
Submitted:
15 July 2024
Posted:
16 July 2024
You are already at the latest version
Abstract
Follicular helper CD4+ T cells (TFH) are a major cellular pool for the maintenance of HIV reservoir. Therefore, the delineation of the follicular (F)/germinal center (GC) immune landscape will significantly advance our understanding of HIV pathogenesis. We have applied multiplex confocal imaging, in combination with relevant computational tools, to investigate, F/GC in situ immune dynamics in viremic (vir-HIV), antiretroviral treated (cART HIV) People Living With HIV (PLWH) and compare them to reactive, non-infected controls. Lymph nodes (LNs) from viremic and cART PLWH could be further grouped based on their TFH cell densities in high-TFH and low-TFH subgroups. These subgroups were also characterized by different in situ distribution of PD1hi TFH cells. The significantly accumulated follicular, compared to extrafollicular, FOXP3hi CD4+ T cells found in the low-TFH cART-HIV group were characterized by a less scattered in situ distribution and strongly correlated with the cell density of CD8+ T cells in this group. An inverse correlation between plasma viral load and LN GrzBhiCD8+ T and CD16hiCD15lo cells was found. Our data reveal the complex GC immune landscaping in HIV infection and suggest that follicular FOXP3hi CD4+ T cells could be negative regulators of TFH cell prevalence in cART-HIV.
Keywords:
HIV
; germinal center
; imaging
; CD4 T cells
1. Introduction
Despite the intense research in HIV pathogenesis and cure approaches over the last 40 years, several aspects related to relevant immune cell and viral dynamics are still not well understood. Combination antiretroviral therapy (cART) has extended the life expectancy and improved the quality of life of People Living With HIV (PLWH). Although cART blocks HIV replication, it cannot eradicate the virus[1,2]. Latent infection of resting CD4+ T cells provides a mechanism for lifelong persistence of HIV[3]. Integrated HIV DNA can be detected in various blood and non-blood tissues, but recent evidence indicates that genetically intact proviruses are mainly detected in lymph nodes (LNs)[4]. Chronic HIV infection is characterized by dramatic changes of LN architecture, stromal cell loss and adaptive immune cell composition[5]. Follicular helper CD4+ T cells (TFH), that play a pivotal role as mediators of functional B cell responses[6,7], represent a major HIV reservoir within central memory CD4+ T cells[8,9]. Non-human primate (NHP) studies have shown accumulation of TFH cells in SIV chronically infected compared to non-infected LNs, a phenotype associated with increased frequency of activated germinal center B cells and SIV-specific antibodies[10]. On the other hand, progression to AIDS (acquired immunodeficiency syndrome, advanced stage of disease) was associated with progressive loss of TFH cells in SIV infected NHP[11]. In addition to TFH and GC B cell altered dynamics, chronic HIV/SIV is also characterized by increased cell density of LN/follicular effector CD8+ T cells[12], deregulated immune-regulatory (TREG) and follicular immune-regulatory (TFR) levels[13,14] ,infiltration of inflammatory cell subsets and excessive fibrosis[5,15]. Foxp3hiCD4+ T cells were reported to impair the proliferation and cytokine-releasing capacity of TFH in ex vivo HIV-focused studies[14]. Additionally, IL-10 and CTLA-4 expression of TFR cells were up-regulated in treatment-naïve PLWH[16]. Innate immune cells, which contribute to the adaptive immune responses either by antigen presentation or by secretion of immunomodulatory cytokines[17], play an important role in HIV pathogenesis[18,19] while their role in cure strategies[20] is a field under development.
Several studies have focused on the characterization of LN-derived T cells, mainly using LN-derived cell suspensions and Flow or Mass cytometry-based assays[21] or single cell RNA analysis[22,23]. However, these experimental approaches lack the information of immune cell spatial organization. In situ characterization of immune cell types using multiplex imaging methodologies and appropriate cohorts of control and disease samples, can provide important insight into the prevalence, phenotype, and spatial organization of relevant immune cells in HIV that could also indicate possible mechanistic interactions between specific cell types.
Herein, we applied multiplex imaging combined with advanced computational tools for the comprehensive characterization of the immune landscaping in viremic and cART HIV LNs compared to reactive control LNs or tonsils from non-HIV infected individuals. Our results indicate that both viremic and cART HIV LNs can be further grouped based on TFH cell density. These groups exhibited distinct profiles of Foxp3hi CD4+ T, Granzyme B (GrzB)hi CD8+ T and innate immune cell subsets. Our work points to a possible role of FOXP3hi CD4+ T cells as regulators of TFH cells in HIV, particularly in cART individuals. Our neighboring analysis revealed a distinct distribution pattern for both TFH and follicular (F-)FOXP3hi CD4+ T between PLWH subgroups as well as compared to non-infected donors.
2. Materials and Methods
2.1. Human Material
The tissue samples used in this study were obtained from i) the Centro de Investigacion en Enfermedades Infecciosas (CIENI), Instituto Nacional de Enfermedades Respiratorias (INER) in Mexico City, Mexico (viremic LNs), ii) the University of Washington, Seattle, USA (cART HIV LNs) and iii) the archives of the Institute of Pathology of Lausanne University Hospital, Switzerland (control LNs). Tonsillar tissues were obtained from anonymized children who underwent routine tonsillectomy at the Hospital de l’Enfance of Lausanne. All procedures were in accordance with the Declaration of Helsinki and approved by the appropriate Institutional Review Board/Ethical Committee: i) all tissue samples from PWH were procured with explicit written informed consent from participants prior to donation, adhering strictly to the principles outlined in the Declaration of Helsinki.
2.2. Tissue Processing
Formalin-fixed, paraffin embedded (FFPE) blocks were sequentially cut into 4 μm sections and prepared on Superfrost glass slides (Thermo Scientific, Waltham, MA, Ref. J1800AMNZ), dried overnight at 37°C and stored at 4°C. Before staining, the slides were heated on a metal hotplate (Stretching Table, Medite, Burgdorf, OTS 40.2025, Ref. 9064740715) at 65°C for 20 min. This melting step ensures proper adherence and deparaffinization of tissue section. Fluorescent multiplex immunohistochemistry (mIHC) staining was performed on the Ventana Discovery Ultra Autostainer from Roche Diagnostics.
2.3. Confocal Imaging Assays
2.3.1. Tissue Staining & Data Acquisition
Tissue sections were sequentially subjected to antibody blocking using the Opal blocking/antibody diluent solution, ARD1001EA) staining with primary antibodies (details on antibodies, clones and dilutions are listed in Suppl. Table 1), incubation with secondary HRP-labeled antibodies for 16 minutes, detection with optimized fluorescent Opal tyramide signal amplification (TSA) dyes (Opal 7-color Auto-mation IHC kit, from Akoya, Ref. NEL821001KT) and repeated antibody denaturation cycles. Samples were then counterstained with Spectral DAPI from Akoya for 4 min, rinsed in soapy water and mounted using DAKO mounting medium (Dako/Agilent, Santa Clara, CA, USA, Ref. S302380-2).
Images were acquired using a Leica Stellaris 8 SP8 confocal system, equipped with the LAS-X software, at 512 x 512-pixel density and 0.75x optical zoom using a 20x objective (NA), unless otherwise stated. Frame averaging or summing was never used while obtaining the images. At least 70% of each section was imaged, to ensure accurate representation and minimize selection bias. Tissues stained with a single antibody-fluorophore combination were used to create a compensation matrix via the Leica LAS-AF Channel Dye Separation module (Leica Microsystems), which was used to correct fluorophore spillover (when present), as per user’s manual.
2.3.2. Quantitative Imaging Analysis (Histocytometry)
Confocal image analysis was performed with Imaris software version 9.9.0 (Bitplane). Quantitative data were generated from the images through histocytometry analysis[24,25], as previously reported. In brief, the Surface Creation module of Imaris was used to generate 3-dimensional segmented surfaces (based on nuclear signal) of spillover corrected images. Dat generated from histocytometry such as average voxel intensities for all channels, in addition to volume and sphericity of the 3-dimensional surfaces, were exported in Microsoft Excel format. The files were converted to comma separated value (.CVS) filed and the data were imported into FlowJo (version 10) to be further analyzed and quantitated. Well-defined areas devoid of background staining were included in the analysis and data were quantified either as relative frequencies or as cell counts normalized to total follicular area screened. Optimal z-stack settings were applied in all collected images. Maximum Intensity Projections (MIP) are presented throughout the manuscript.
2.3.3. Data analysis-Neighboring Analysis
The distance between relevant cell subsets (CD20hi, PD1hiCD57hi/lo e.t.c) was calculated with python 3.10.9 using the SciPy library[26]. The matrix interaction was created using X and Y coordinates from each cell phenotype and the median distance was extracted. Furthermore, to characterize the probability to observe different patterns of cellular distribution across Regions Of Interest (ROIs) and patients, we study the curves generated from the Ripley’s G function and the theoretical Poisson curve using pointpats 2.3.0 (https://doi.org/10.5281/zenodo.7706219). The area between the empirical and theoretical Poisson curve was extracted using the NumPy library[27]. ROIs with at least 20 positive cells for each cell subset under investigation were analyzed. The data were presented as i) bar graphs, showing the range of all various distances measured (X axis) and the frequency or count of B cells that fell within each distance range and ii) dot plots, where each dot represents the mean value of the minimum distances between two cell populations for each follicular area.
2.3.4. Viral Load Measurement
The m2000 system (Abbott, Abbot Park, IL, USA) with a detection limit of 40 HIV RNA copies/mL was used to perform automated real-time polymerase chain reaction (PCR) for the determination of HIV plasma viral load (pVL), with a detection limit of 40 HIV RNA copies/mL. Flow cytometry with the AQUIOS Tetra-1 Panel in AQUIOS CL (Beckman Coulter Life Sciences, Indianapolis, IN, USA) was used to determine CD4+ T cell counts. All PWLH involved in the current study were infected with clade B HIV.
2.4. Statistical Analysis
For imaging data analysis, the Mann-Whitney test and simple linear regression analysis were used. The p-values Mann-Whitney test were corrected using the False Discovery Rate (FDR) correction test[28] with q=0.05, for multiple comparisons (both uncorrected and corrected p values for each figure are shown in Suppl. Table 2). Analysis and graphs were generated using the GraphPad Prism 8.3.0 software. For statistical significance, p value <0.05 was considered.
3. Results
3.1. Similar Profiles of Follicular Helper CD4+ T Cell Densities in Viremic and cART HIV LNs
Given the importance of TFH cells for the HIV reservoir maintenance[29] as well as the development of neutralizing antibodies[30],we sought to investigate their in-situ cell density in HIV infected compared to non-infected tissues. We used tonsils and non-infected, cancer free, reactive LNs characterized by follicular hyperplasia as strict control groups (Figure S1, S6). We started our analysis by applying a multiplex imaging assay allowing for the simultaneous detection of main GC B and T cell subsets (Figure 1A). The gating scheme used for the identification and quantitative analysis of corresponding cell subsets by Histocytometry[24,25] is shown (Figure 1B). The in-situ density of CD20hi/dim cells was used for the identification of individual follicular areas (highly enriched in CD20hi/dim cells) as well as the ‘total follicular’ area (Figure 1B). As expected [5,31] , PWLH tissues harbor both ‘preserved’ as well as ‘irregular’ follicular structures (Figure 1A and S1B). CD4+ T cell subsets within the ‘total follicular’ area were analyzed based on their expression of PD1 and CD57 (Figure 1B). Preliminary analysis revealed low expression levels of CD4 in certain TFH cells, particularly the ones expressing CD57 (Figure S2A). Several studies have shown that TFH cells express a unique PD1hi phenotype[9,10,32] compared to other CD4+ T cells while PD1 expression per cell (judged by Mean Fluorescence Intensity) of follicular CD8+ T cells is 4-5 times lower than that of TFH[24]. Therefore, the expression level of PD1 can serve as an in situ TFH identifier. To avoid inconsistencies and misinterpretation of our data, especially for the CD57hi TFH cell subset, we chose to directly analyze the PD1hiCD57hi/low cells in GCs (Figure 1B). The gating for setting the threshold of these biomarkers is shown in Figure S2A. Their expression level in extrafollicular areas as well as the manual inspection of their fluorescence intensities in the raw images, was used as a reference to set the cut-off for ‘high’ values. Back gating of PD1hi and CD57hi cells identified by Histocytometry (shown as spheres) to the original image showed a high concordance between the digitally identified cells and their original counterparts (Figure 1C).
Calculation of PD1hi cell densities (normalized cell counts per mm2) allowed for further grouping of HIV viremic and cART tissues into two subgroups characterized by significantly different cell densities of PD1hi TFH cells; one with high PD1hi TFH cell densities (hereafter ‘high-TFH’) and one with low PD1hi TFH cell densities (hereafter ‘low-TFH’) (Figure 1D, left panel). In order to assess the heterogeneous (or not) prevalence of PD1hi TFH cells across an individual tissue, the cell densities of PD1hi TFH per follicle for every tissue were analyzed. A great variability was observed in all tissues, particularly in tonsils and control LNs (Figure 1D, right panel).
CD57, a carbohydrate epitope, marks a TFH cell subset with distinct positioning and function in human LNs[33,34]. As expected [34], the PD1hiCD57hi group represents a subset of TFH cells (Figure 1E). PD1hiCD57hi TFH cells exhibited a similar cell density profile with PD1hi cells (Figure 1E) and heterogenous prevalence across the tissue (Figure S2B). A strong correlation between PD1hi and PD1hiCD57hi TFH cell densities was found for tonsil, control and cART LNs (Figure 1F). This association was less significant in the high-TFH PLWH viremic subgroup (Figure 1F). Although not statistically significant, a negative association was observed between the PD1hi TFH cells and pVL in viremic PLWH, as well as a trend for higher pVL in the low-TFH compared to high-TFH viremic PWLH subgroup (Figure 1G, lower and upper panel respectively). Therefore, in agreement with our previous data for viremic SIV infection [10], two subgroups defined by significantly different cell densities of TFH cells were identified in viremic as well as cART HIV LNs.
3.2. A Distinct Positioning Profile of TFH cells in HIV-Infected Compared to Non-Infected Tissues
Next, the spatial positioning of GC B cells and TFH cell subsets was investigated in our tissue cohort. To this end, follicles from all groups, harboring at least 20 cells for each corresponding cell population, were used. The X, Y coordinates of relevant cells were extracted (Figure 2A, upper panel) and digitalized representation of their distribution was generated (Figure 2A, lower panel). Then, the ‘G function’ parameter, as a surrogate for the dispersion/scattering of a distribution, of a given cell type as well as the mean values of the minimum distances between relevant cell types in individual follicular areas were calculated[35]. An example of follicles with low and high mean distance between CD20hi and PD1hi cells and their associated G parameters is shown (Figure 2B). A similar distribution profile for CD20hi cells, among the tissue subgroups analyzed was detected (Figure 2C). Contrary to B cells, PD1hi cells expressed a significantly higher dispersion in the vir-HIV subgroups compared to control LNs (Figure 2D). Regarding the PD1hi cells distribution of cART tissues, a significant difference was found only between control and low-TFH cART LNs (Figure 2D). Furthermore, a significantly higher dispersion was found in low-TFH compared to high-TFH HIV tissues, in both viremic and cART tissues from PLWH (Figure 2D). A significantly higher mean of minimum distance between CD20hi and PD1hi cells was found in the vir- and cART HIV compared to control tissues (Figure 2E), while no difference was observed among the PLWH subgroups (Figure 2E).
Then, the aforementioned parameters were calculated for the PD1hiCD57lo and PD1hiCD57hi cells in all tissue groups. Given the low abundancy of CD57hi TFH cells, a significantly lower number of follicular areas, especially in the low-TFH HIV subgroups, was analyzed (Figure 2F and G). Again, a comparable CD20 G-function profile was found among the tissue groups, in these follicular areas (Figure S3A). Our data showed a similar G function and distance profile between tonsils and reactive control LNs (Figure 2F and G). In general, PD1hiCD57lo and PD1hiCD57hi cells express a significantly higher degree of dispersion in HIV infected compared to tonsils and control LNs (Figure 2F). Interestingly, the cART low-TFH subgroup was the one with the highest dispersion of PD1hiCD57lo cells among the PLWH subgroups (Figure 2F). With respect to the mean minimum distance, a trend, which was statistically significant for many of the comparisons, for longer distance between CD20hi and PD1hiCD57lo OR hi cells was measured in HIV infected compared to tonsils and control LNs (Figure 2G). Our data suggest a distinct profile of the in situ TFH cell distribution and neighboring with B cells in PLWH compared to tonsils and control LNs.
3.3. Significant Accumulation of Follicular compared to Extrafollicular FOXP3hi CD4+ T Cells in cART low-TFH LNs
We used the expression of FOXP3 as a surrogate for potential immune-regulatory CD4+ T cells (Figure 3A). The Histocytometry gating scheme for the identification and calculation of FOXP3hi CD4+ T cells in extrafollicular and intrafollicular areas (highly enriched in CD20hi/dim cells) is shown (Figure 3B). The concordance between digitally/Histocytometry identified FOXP3hi CD4hi T cells (shown as spheres) and their original counterparts is demonstrated in Figure 3C. The lowest cell density of extrafollicular (EF) and follicular (F)FOXP3hi CD4+ T cells was found in tonsils and the highest in reactive control LNs (Figure 3D and E). Comparable EF-FOXP3hiCD4hi T cell densities among the PLWH subgroups were found (Figure 3D). Although not statistically significant, a trend for higher cell densities of F-FOXP3hi CD4+ T cells in cART- compared to vir-HIV LNs was measured (Figure 2E). A broad range of F-FOXP3hi CD4+ T cell densities was observed, particularly in the cART low-TFH subgroup (Figure 2E and S3B).
Next, the EF- and F-FOXP3hi CD4+ T cell densities were compared in each LN. Similar EF- and F-FOXP3hi CD4+ T cell densities were found in control LNs while fewer F-FOXP3hi CD4+ compared to EF-FOXP3hi CD4+ T cells were found for almost all viremic PLWH tissues tested (Figure 3F). However, the opposite profile was observed for the cART tissues (Figure 3F). A consistent, significantly higher cell density of F- FOXP3hi CD4+ compared to EF-FOXP3hi CD4+ T cells was measured, especially in the cART low-TFH subgroup (Figure 3F). The distribution profile (G function) of FOXP3hi CD4+ T cells, across the whole imaged area for each tissue, was also investigated. No significant differences were found among the groups analyzed (Figure S3C). Then, we focused our analysis on F-FOXP3hi CD4+ T cells. An example of identification and corresponding digital representation of their distribution in cART tissues is shown (Figure 3G). A significantly lower dispersion was measured in cART low-TFH compared to cART high-TFH subgroup, when the G factor was calculated for F-FOXP3hi CD4+ T cells (Figure 3H). Our data show a preferential accumulation of follicular immune regulatory CD4 T cells in cART compared to viremic PLWH LNs.
3.4. LN GrzBhiCD8+ T Cells Are Negatively Associated with Blood Viral Load
The in situ profile of bulk and effector (GrzBhi) CD8+ T cells (Figure 4A) was investigated using a multiplex imaging assay and Histocytometry gating scheme is shown in Figure 4B. The applied antibody panel (Supp Table 2, panel II) does not include a follicular/GC biomarker, therefore the cell density of CD8+ T cell subsets was analyzed for the whole imaged area (Figure 4A and B). The concordance between digitally/Histocytometry identified GrzBhi CD4hi T cells (shown as spheres) and their original counterparts is demonstrated in Figure 4C. In line with our previous data[15,24], an accumulation of bulk and GrzBhi CD8+ T cells was measured in viremic donors, particularly the high-TFH tissues, compared to tonsils and control LNs (Figure 4D and S4A). Although not significant, a reduction of GrzBhi CD8+ T cells was observed between the viremic and cART tissues, which was more evident between the high-TFH subgroups (Figure 4D). A positive association between circulating and LN bulk and GrzBhi CD8+ T cells was found in HIV viremic samples (Figure S4B). Contrary to circulating CD8+ T cells (Figure S4C), a significant negative correlation was observed between the viral load and the LN GrzBhi CD8+ T cells (Figure 4E).
Next, the correlation between bulk and GrzBhi CD8+ as well as FOXP3hi CD4+ T cell densities was investigated. A significant association was observed between the two CD8+ T cell populations in viremic LNs (Figure S4D) as well as the cART low-TFH subgroup (Figure 4F). However, this was not the case for the cART high-TFH subgroup (Figure 4F). Among the groups analyzed, a positive correlation was found between bulk CD8+ T - and EF-FOXP3hi CD4+ T cell densities in the cART low-TFH subgroup (Figure 4G, upper panel). This correlation was statistically significant between bulk CD8+ T cells and F-FOXP3hi CD4+ T cells in the same subgroup (Figure 4G, lower panel). The distance profiling revealed a similar, dispersion of GrzBhi CD8+ T cells in LNs from cART compared to those from viremic PLWH (Figure 4H). Conclusively, our data revealed an accumulation of GrzBhi CD8+ T cells in PWLH LNs compared to non-infected tissues, which was inversely correlated with the viral load in viremic PLWH.
3.5. Differential Modulation of Innate Immune Cell Subsets by cART
HIV is a chronic disease characterized by immune activation and inflammation[5,36,37]. Given the role of the innate immunity in HIV pathogenesis, we sought to investigate the tissue dynamics of several innate immune cell subsets using relevant biomarkers (CD163 and CD68, markers for monocytes/macrophages[38], CD15, a surrogate for myeloid cells/granulocytes, CD16 a surrogate for activated myeloid cells, neutrophils and NK cells [39,40] (Figure 5A). The Histocytometry gating scheme for the identification and quantification of these cell subsets is shown in Figure 5B. In general, a reduction was measured for all innate immune cell subsets analyzed in low- compared to high-TFH viremic subgroups (Figure 5C), that was significant for the CD15hiCD16lo cells (Figure 5C). With respect to cART, lower cell densities of CD163hiCD68lo, CD15hiCD16lo, CD16hiCD15lo and CD16hiCD15hi cells were found in cART high-TFH compared to viremic high-TFH tissues (Figure 5C). Comparable cell densities of CD68hiCD163lo cells were counted among all tissue groups analyzed (Figure 5C, upper panel).
Then, the relationship between CD8+ T cells and innate immunity cell types was investigated. A positive association between CD68hi or CD163hi cell subsets and bulk CD8+ T cells in viremic LNs was found (Figure S5A). A significant positive correlation between CD16hiCD15lo and bulk or GrzbhiCD8+ T cells was found in the vir-HIV high-TFH group (Figure 5D). No such correlations were observed in cART LNs. Similar to GrzbhiCD8+ T cells, a significant inverse correlation was also observed between CD16hiCD15lo cells and peripheral blood viral load in viremic PLWH (Figure S5B). Analysis of the in-situ distribution profiling showed a higher dispersion in cART compared to viremic HIV infected tissues for CD163 hi cells while comparable profiles were found for CD68hi and CD16hi cells (Figure S5C). Furthermore, a lower minimum mean distance between GrzBhi CD8+ and CD16hi cells, but not CD68hi or CD163hi, in viremic compared to cART HIV tissues was found (Figure 5E and S5D). Altogether, our data suggest that cART has a differential impact on the cell density of individual innate immunity cell subsets.
4. Discussion
Here, we have investigated the immune landscaping in reactive LNs from PLWH and compared it non-infected, control LNs and tonsils (Figure S6). We chose to compare the in situ immune dynamics in HIV LNs to non-infected, reactive LNs characterized by active follicles (follicular hyperplasia), as a reference for highly active F/GCs. Given the abundance in GC B and TFH cells as well as the preservation of the follicular and sub-follicular structures tonsils are considered as a ‘prototype’ lymphoid organ for the investigation of F/GC immune cell types. We should emphasize that none of the viremic PLWH had active opportunistic infections or AIDS-defining pathologies at the time of the sampling. The time since diagnosis was also similar among the individuals. Our analysis revealed two subgroups of HIV infected LNs with respect to the cell density of TFH cells, in both viremic and cART PLWH. This profile, at least for viremic individuals, is in line with the TFH cell dynamics in SIV non-human primate model[10,11].
No association of TFH cell densities with gender, age, CD4+ and CD8+ counts was found in both viremic and cART PLWH. HIV infection of LN CD4+ T cells per se, intrinsic factors and/or the interaction of TFH cells with the GC microenvironment represent potential mechanisms that could contribute to TFH cell in-situ dynamics by altering the differentiation and/or turnover rate of total (or specific subsets) of TFH cells in PLWH. Regarding the viremic PLWH, we observed a negative association between pVL and GC/TFH cells that could reflect a preferential infection/loss of TFH cells or a generalized loss of TFH cells in highly viremic PLWH. The relatively low numbers (~5% of TFH cells) of HIV DNA+ TFH cells [9] are challenging the impact of the productive HIV infection per se. Similar infection rates for CD57hi and CD57lo TFH cells have been reported previously[41] challenging the preferential infection/loss of CD57hi TFH cells and the observed weaker association between these two TFH cell subsets in viremic compared to cART PLWH. Abortive HIV infection represents an alternative mechanism for the loss of tonsillar CD4 T cells at least in vitro [42]. The role of such mechanism for the regulation of vir-HIV TFH cells needs to be investigated. Alternatively, progressive fibrosis and damage of vital LN structural elements (e.g. the Fibroblastic Reticular Cell network[43] and/or loss of GC survival signals (e.g. due to damage of the Follicular Dendritic Cell network[44] for TFH cells in highy viremics could affect the differentiation and maintenance or turnover of TFH cells. The observed heterogeneity of TFH cell densities in different follicles across an individual tissue indicates that the locality of such mechanisms is possibly an important factor to consider in future studies. Our data urge for further investigation of LN structure elements in conjunction with the in-situ dynamics of immune cell types. The development of appropriate imaging tools will greatly facilitate such efforts.
Two groups (LoViReT and HiViReT) of treated PLWH, harboring a relatively wide range of very low viral reservoir, was recently described [45]. Whether the cART TFH subgroups correspond to a status is not known and merits further investigation. Despite the viral control, cART is not able to fully restore the LN/follicular damage in PLWH back to normal. FRC reconstitution is one of the factors that could affect the reconstitution of LN CD4+ T cell pool and presumably the TFH cell prevalence[46]. No association of TFH cell densities with treatment duration was found. However, the capacity of individual PLWH to differentially respond to cART and restore relevant LN elements could contribute to the observed high- and low-TFH subgroups in the cART HIV group. Our distribution analysis showed an overall higher dispersion of the PD1hi TFH cells in HIV infected compared to non-infected tissues, that is also associated with a longer mean distance between TFH and B cells in the infected donors. A mutual regulation between TFH and GC B cells has been reported previously[47]. We hypothesize that the described spatial distribution profile may reflect a lower probability for these two cell subsets to interact in the infected LNs leading to the subsequent loss of vital signals for TFH cells. The aforementioned profile was more evident in the low-TFH HIV subgroups, further supporting our hypothesis.
Follicular immune-regulatory CD4+ T cells (TFR) represent an important ‘microenvironment cell factor’ for the development of TFH cells[14,48,49]. Tonsils harbor the lowest number of FOXP3hi CD4+ T cells, in line with previous reports[48,50]. Our data revealed a contrasting profile regarding the cell density between EF- and F-FOXP3hi CD4+ T cells in control and viremic compared to cART-HIV LNs. Within the cART HIV group, we measured a significant increase of F-FOXP3hi CD4+ T specifically in the cART-low TFH subgroup, suggesting a negative role for the TFH cell development in these individuals. Supplementary to this is the significantly less scattered distribution of the FOXP3hi CD4+ T cells within the follicles of low- compared to high-TFH cART tissues. Whether the FOXP3hi CD4+ T cells represent bona fide TFR cells or cells originated from TFH cells[51] is not known and needs further investigation. Our data urge for a comprehensive in situ phenotypic and functional characterization of FOXP3hi CD4+ T cells, especially in cART-HIV PLWH.
In contrast to PLWH, the majority of follicular CD8+ T cells in tonsils and control LNs express a GrzBlo phenotype, in line with our previous observation[24]. The positive association between the numbers of circulating and LN CD8+ T cell density suggests that increased trafficking of bulk and presumably effector CD8+ T cells may support, at least in part, their increased cell density in viremic LNs. A CXCR3/CXCR3L mediated mechanism, that has been previously proposed HIV PLWH[52] and SIV infected non-human primates[15], could contribute to the CD8+ T cell density profile we observed for viremic PLWH. Several studies have shown the role of CD8+ T cells in controlling HIV and SIV[53,54]. In line with these studies, we found a negative correlation between LN CD8+ and GrzBhi CD8+ with pVL that indicates a potential role of CD8+ T cells for viral control, at least in part, in our cohort. The reduction of GrzBhi CD8+ T cells in cART-HIV LNs, particularly in the high-TFH subgroup, could reflect a lower immune activation due to therapy, in line with the overall lower cell density of several innate immunity cell subsets observed in cART-HIV. Further comprehensive in situ phenotypic (e.g. expression of homing receptors), functional (e.g. ‘regulatory’ function [55]) and topological characterization of CD8+ T cells, especially in cART-HIV PLWH, is of great interest given their potential role for immunotherapies aiming to eliminate the virus. We found a positive correlation between CD8+ and F-FOXP3hi CD4+ T cells specifically in the low-TFH cART-HIV subgroup. Whether and how these two cell types could affect the cell density of TFH cells in this group remains to be elucidated.
Overall, we observed a diverse profile of innate immunity cell types among the groups. Individual cell types were differentially modulated by cART while no consistent association with the low- or high-TFH status was observed. We measured different cell densities for the CD68hiCD163hi and CD68loCD163hi cell subsets. CD163 is a receptor that can be cleaved and therefore the cell density of CD163hi macrophages/monocytes can be underestimated. Our results, however, indicate that possible cleavage/loss of CD163 is not responsible for the observed in-situ dynamics [44]. Our data point to a diverse innate immunity LN microenvironment that could affect the host-virus interplay in these LNs [18]. We found a strong correlation between LN CD8+ T cells and CD16hiCD15lo cells in high-TFH viremic tissues. This profile was also associated with i) a less dispersed CD8+ T cells distribution, ii) a shorter distance between CD8+ and CD16hi cells and iii) a negative association between pVL and LN CD8+ or CD16hiCD15lo cells. These findings urge for further investigation of neutrophils/granulocytes, in addition to macrophages, as possible key determinants for the innate immunity / CD8+ T cell cross talk and virus dynamics[56]. Whether the same or different innate immune cell subsets mediate the host-virus interaction in viremic and cART PLWH is not known and remains to be elucidated.
Altogether, we have analyzed several immunological cell types in a well characterized cohort of LNs and provide evidence i) for the subgrouping of HIV-infected LNs (both from viremic and cART PLWH) based on their TFH cell density, ii) of a distinct profile of potential immunosuppressive FOXP3hi CD4+ T cells in cART LNs with respect to their cell densities, distribution between extrafollicular and follicular areas, and spatial distribution within the follicular compartment and iii) the effect of cART on the cell density of CD8+ T and innate immune cells (Table 2). Altered TFH cell density in HIV subgroups is associated with different CD8+ and F-FOXP3hi CD4+ T cell density and distribution profiles too. The data suggest that further investigation of CD8 and immune-regulatory CD4 T cells could provide insights for the TFH cell prevalence in HIV and particularly in cART PLWH. Understanding the follicular/GC microenvironment in HIV infection could further inform for the role of TFH cells in HIV pathogenesis and for possible combinatorial interventions aiming to manipulate a major HIV tissue reservoir.

Table 1.
Demographic and clinical information of study PLWH.
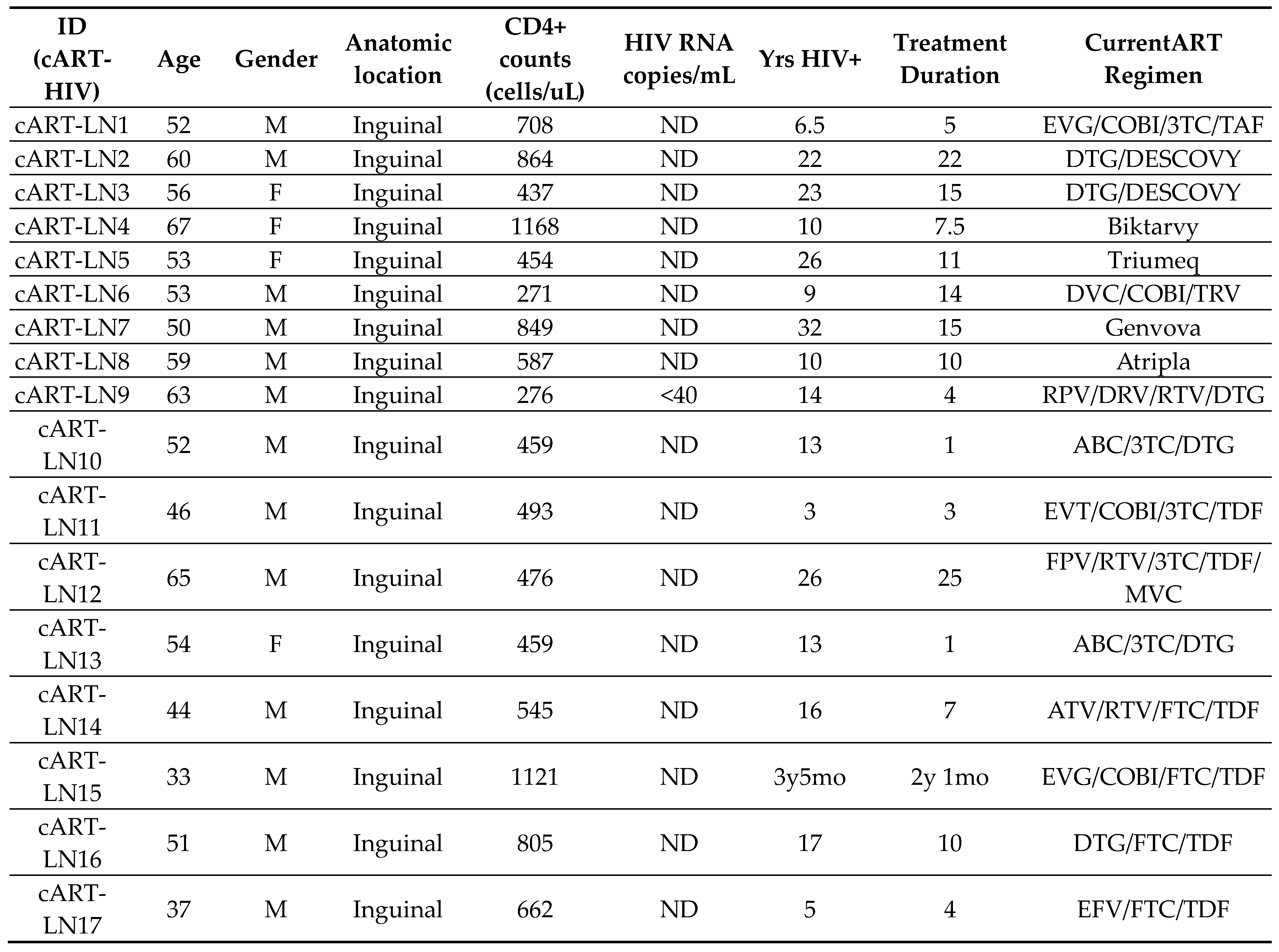
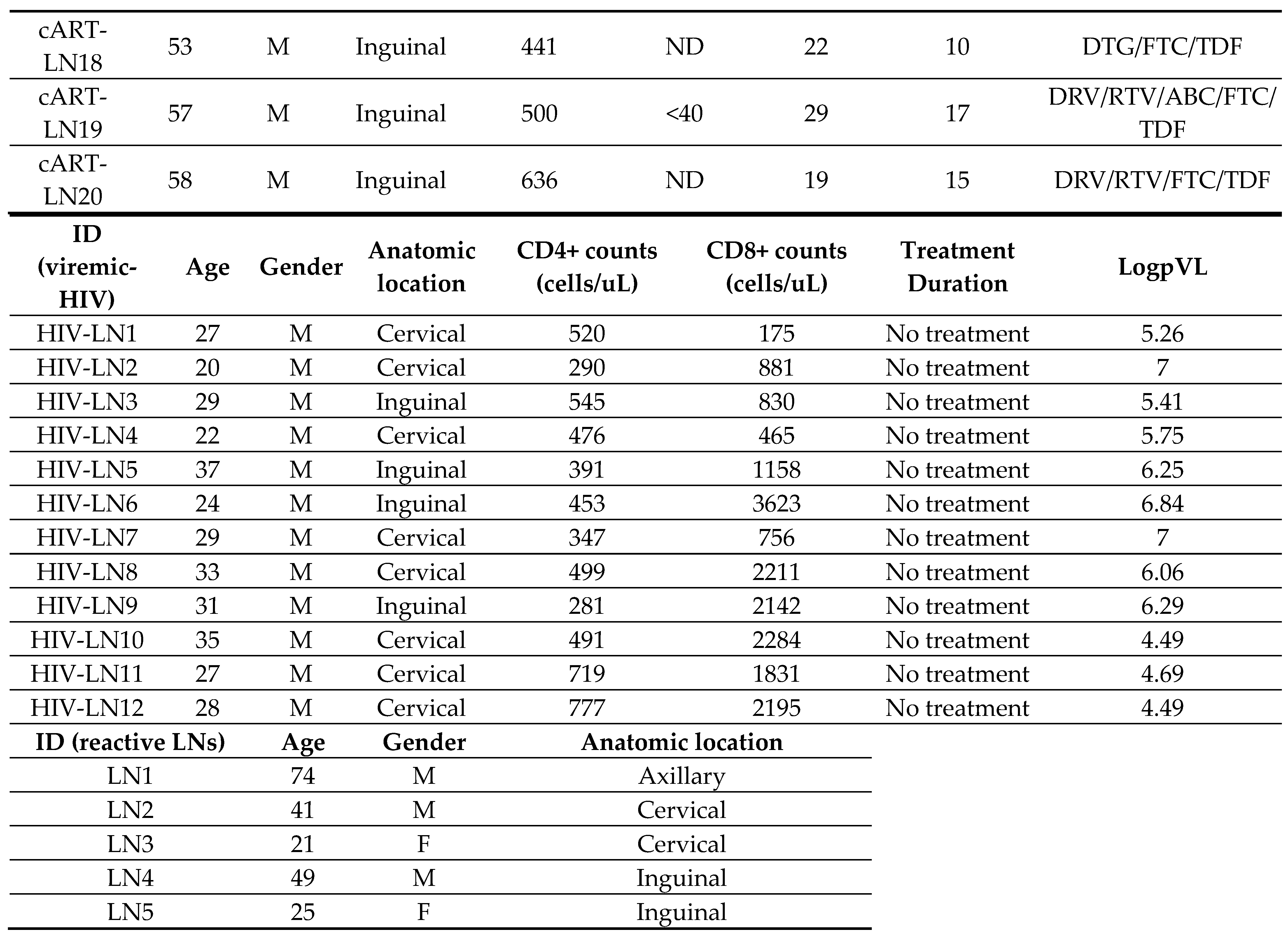
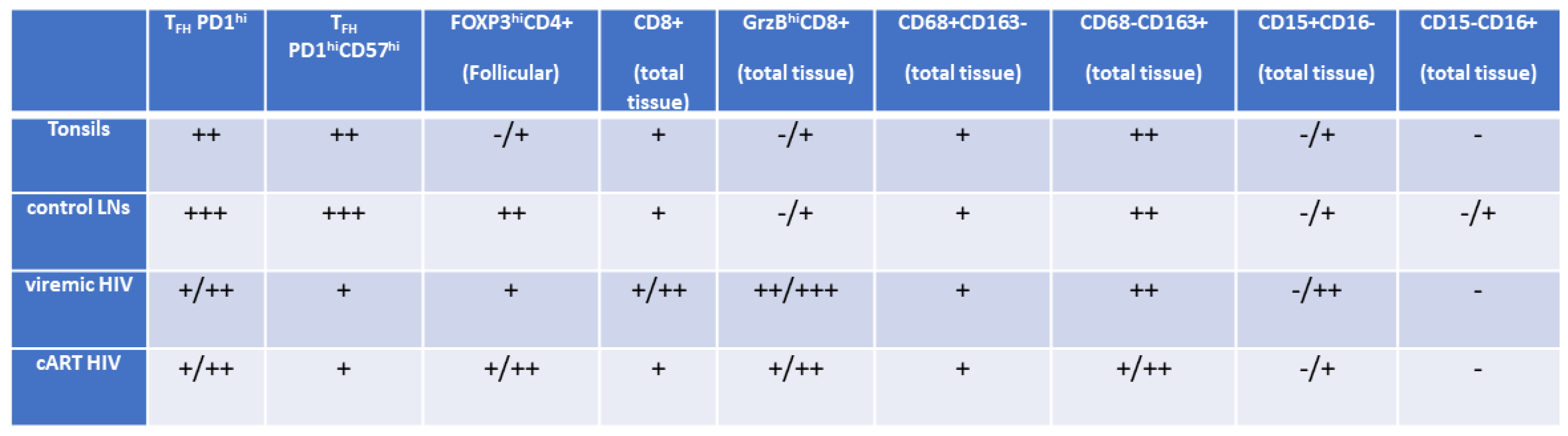
Table 2.
A table summarizing the main comparisons for immune cell subsets among the tissue groups.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Acknowledgments
The authors would like to thank Pr Laurence de Leval for assisting with control lymph nodes and Dr Natalie Piazzon (operational director of the Tissue Biobank), Damien Maison and Emilie Lingre, Institute of Pathology, CHUV, for their help with the tissue processing.
Funding
These studies were supported by grants from the Swiss National Science Foundation (SNF, 310030_204226 to C.P.) and in part by National Institute of Health (USA 7UM AI164561 to C.P.) and the Intramural Research Program of NIAID.
Institutional Review Board Statement
All procedures were in accordance with the Declaration of Helsinki and approved by the appropriate Institutional Review Board/Ethical Committee. The utilization of these samples was formally sanctioned by both the Research Committee and the Ethics in Research Committee of the National Institute of Respiratory Diseases "Ismael Cosío Villegas," Mexico City as part of the "C71-18" protocol, ii) University of Washington (study ID: STUDY00001091, CR ID:CR00004042) and iii) the Canton de Vaud-CER-VD, Switzerland for LNs (2021-01161) and tonsils (PB_2016-02436 (201/11)). Written informed consent was obtained from all participants.
Authorship Contributions
S.G. and M.O. performed experiments, analyzed and interpreted data and drafted the manuscript; C.B. and S.B. assisted with distance analysis; O.Y.C. supervised the statistical analysis, P.D.R.E., S.A.R., F.T.R., G.P. and R.A.K. provided tissue material, R.A.K. provided input for the data interpretation, CP conceived, designed and supervised the study, interpreted data, and revised the manuscript. All authors have read and approved the final version for submission.
Disclosure of Conflicts of Interest
The authors declare no competing conflicts of interest.
Data Sharing Statement
The authors agree to share all publication-related data. For further information, please contact the corresponding author at Konstantinos.Petrovas@chuv.ch.
References
- Dey, B.; Berger, E.A. Towards an HIV cure based on targeted killing of infected cells: different approaches against acute versus chronic infection. Curr Opin HIV AIDS 2015, 10, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Maldarelli, F. The role of HIV integration in viral persistence: no more whistling past the proviral graveyard. J Clin Invest 2016, 126, 438–447. [Google Scholar] [CrossRef] [PubMed]
- De Flora, S.; Serra, D.; Basso, C.; Zanacchi, P. Mechanistic aspects of chromium carcinogenicity. Arch Toxicol Suppl 1989, 13, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Dufour, C.; Ruiz, M.J.; Pagliuzza, A.; Richard, C.; Shahid, A.; Fromentin, R.; Ponte, R.; Cattin, A.; Wiche Salinas, T.R.; Salahuddin, S.; et al. Near full-length HIV sequencing in multiple tissues collected postmortem reveals shared clonal expansions across distinct reservoirs during ART. Cell Rep 2023, 42, 113053. [Google Scholar] [CrossRef] [PubMed]
- Estes, J.D. Pathobiology of HIV/SIV-associated changes in secondary lymphoid tissues. Immunol Rev 2013, 254, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Mudd, P.A.; Minervina, A.A.; Pogorelyy, M.V.; Turner, J.S.; Kim, W.; Kalaidina, E.; Petersen, J.; Schmitz, A.J.; Lei, T.; Haile, A.; et al. SARS-CoV-2 mRNA vaccination elicits a robust and persistent T follicular helper cell response in humans. Cell 2022, 185, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Noto, A.; Suffiotti, M.; Joo, V.; Mancarella, A.; Procopio, F.A.; Cavet, G.; Leung, Y.; Corpataux, J.M.; Cavassini, M.; Riva, A.; et al. The deficiency in Th2-like Tfh cells affects the maturation and quality of HIV-specific B cell response in viremic infection. Front Immunol 2022, 13, 960120. [Google Scholar] [CrossRef] [PubMed]
- Aid, M.; Dupuy, F.P.; Moysi, E.; Moir, S.; Haddad, E.K.; Estes, J.D.; Sekaly, R.P.; Petrovas, C.; Ribeiro, S.P. Follicular CD4 T Helper Cells As a Major HIV Reservoir Compartment: A Molecular Perspective. Front Immunol 2018, 9, 895. [Google Scholar] [CrossRef] [PubMed]
- Perreau, M.; Savoye, A.L.; De Crignis, E.; Corpataux, J.M.; Cubas, R.; Haddad, E.K.; De Leval, L.; Graziosi, C.; Pantaleo, G. Follicular helper T cells serve as the major CD4 T cell compartment for HIV-1 infection, replication, and production. J Exp Med 2013, 210, 143–156. [Google Scholar] [CrossRef]
- Petrovas, C.; Yamamoto, T.; Gerner, M.Y.; Boswell, K.L.; Wloka, K.; Smith, E.C.; Ambrozak, D.R.; Sandler, N.G.; Timmer, K.J.; Sun, X.; et al. CD4 T follicular helper cell dynamics during SIV infection. J Clin Invest 2012, 122, 3281–3294. [Google Scholar] [CrossRef]
- Xu, H.; Wang, X.; Malam, N.; Lackner, A.A.; Veazey, R.S. Persistent Simian Immunodeficiency Virus Infection Causes Ultimate Depletion of Follicular Th Cells in AIDS. J Immunol 2015, 195, 4351–4357. [Google Scholar] [CrossRef] [PubMed]
- Chirila, T.V.; Russo, A.V.; Constable, I.J. Chemical investigations of ultraviolet-absorbing hydrogel material for soft intraocular lenses. J Cataract Refract Surg 1989, 15, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Graff-Dubois, S.; Rouers, A.; Moris, A. Impact of Chronic HIV/SIV Infection on T Follicular Helper Cell Subsets and Germinal Center Homeostasis. Front Immunol 2016, 7, 501. [Google Scholar] [CrossRef] [PubMed]
- Miles, B.; Miller, S.M.; Folkvord, J.M.; Kimball, A.; Chamanian, M.; Meditz, A.L.; Arends, T.; McCarter, M.D.; Levy, D.N.; Rakasz, E.G.; et al. Follicular regulatory T cells impair follicular T helper cells in HIV and SIV infection. Nat Commun 2015, 6, 8608. [Google Scholar] [CrossRef] [PubMed]
- Ferrando-Martinez, S.; Moysi, E.; Pegu, A.; Andrews, S.; Nganou Makamdop, K.; Ambrozak, D.; McDermott, A.B.; Palesch, D.; Paiardini, M.; Pavlakis, G.N.; et al. Accumulation of follicular CD8+ T cells in pathogenic SIV infection. J Clin Invest 2018, 128, 2089–2103. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Xu, W.; Tu, B.; Hong, W.G.; Zhang, Z.; Chen, W.W.; Zhao, M. Alterations of the frequency and functions of follicular regulatory T cells and related mechanisms in HIV infection. J Infect 2020, 81, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Luster, A.D. The role of chemokines in linking innate and adaptive immunity. Curr Opin Immunol 2002, 14, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Chan, C.N.; Rovira-Clave, X.; Chen, H.; Bai, Y.; Zhu, B.; McCaffrey, E.; Greenwald, N.F.; Liu, C.; Barlow, G.L.; et al. Combined protein and nucleic acid imaging reveals virus-dependent B cell and macrophage immunosuppression of tissue microenvironments. Immunity 2022, 55, 1118–1134. [Google Scholar] [CrossRef] [PubMed]
- Banga, R.; Procopio, F.A.; Lana, E.; Gladkov, G.T.; Roseto, I.; Parsons, E.M.; Lian, X.; Armani-Tourret, M.; Bellefroid, M.; Gao, C.; et al. Lymph node dendritic cells harbor inducible replication-competent HIV despite years of suppressive ART. Cell Host Microbe 2023, 31, 1714–1731. [Google Scholar] [CrossRef]
- Board, N.L.; Moskovljevic, M.; Wu, F.; Siliciano, R.F.; Siliciano, J.D. Engaging innate immunity in HIV-1 cure strategies. Nat Rev Immunol 2022, 22, 499–512. [Google Scholar] [CrossRef]
- Neidleman, J.; Luo, X.; Frouard, J.; Xie, G.; Hsiao, F.; Ma, T.; Morcilla, V.; Lee, A.; Telwatte, S.; Thomas, R.; et al. Phenotypic analysis of the unstimulated in vivo HIV CD4 T cell reservoir. Elife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M. Keeping the ill at ease. Postgrad Med 1990, 88, 44–45. [Google Scholar] [CrossRef] [PubMed]
- Szabo, P.A.; Levitin, H.M.; Miron, M.; Snyder, M.E.; Senda, T.; Yuan, J.; Cheng, Y.L.; Bush, E.C.; Dogra, P.; Thapa, P.; et al. Single-cell transcriptomics of human T cells reveals tissue and activation signatures in health and disease. Nat Commun 2019, 10, 4706. [Google Scholar] [CrossRef] [PubMed]
- Petrovas, C.; Ferrando-Martinez, S.; Gerner, M.Y.; Casazza, J.P.; Pegu, A.; Deleage, C.; Cooper, A.; Hataye, J.; Andrews, S.; Ambrozak, D.; et al. Follicular CD8 T cells accumulate in HIV infection and can kill infected cells in vitro via bispecific antibodies. Sci Transl Med 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Moysi, E.; Del Rio Estrada, P.M.; Torres-Ruiz, F.; Reyes-Teran, G.; Koup, R.A.; Petrovas, C. In Situ Characterization of Human Lymphoid Tissue Immune Cells by Multispectral Confocal Imaging and Quantitative Image Analysis; Implications for HIV Reservoir Characterization. Front Immunol 2021, 12, 683396. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: fundamental algorithms for scientific computing in Python. Nat Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Lovell, D.P. Biological importance and statistical significance. J Agric Food Chem 2013, 61, 8340–8348. [Google Scholar] [CrossRef] [PubMed]
- Miles, B.; Connick, E. TFH in HIV Latency and as Sources of Replication-Competent Virus. Trends Microbiol 2016, 24, 338–344. [Google Scholar] [CrossRef]
- Havenar-Daughton, C.; Lee, J.H.; Crotty, S. Tfh cells and HIV bnAbs, an immunodominance model of the HIV neutralizing antibody generation problem. Immunol Rev 2017, 275, 49–61. [Google Scholar] [CrossRef]
- Biberfeld, P.; Ost, A.; Porwit, A.; Sandstedt, B.; Pallesen, G.; Bottiger, B.; Morfelt-Mansson, L.; Biberfeld, G. Histopathology and immunohistology of HTLV-III/LAV related lymphadenopathy and AIDS. Acta Pathol Microbiol Immunol Scand A 1987, 95, 47–65. [Google Scholar] [CrossRef]
- Crotty, S. Follicular helper CD4 T cells (TFH). Annu Rev Immunol 2011, 29, 621–663. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Rott, L.S.; Clark-Lewis, I.; Campbell, D.J.; Wu, L.; Butcher, E.C. Subspecialization of CXCR5+ T cells: B helper activity is focused in a germinal center-localized subset of CXCR5+ T cells. J Exp Med 2001, 193, 1373–1381. [Google Scholar] [CrossRef]
- Padhan, K.; Moysi, E.; Noto, A.; Chassiakos, A.; Ghneim, K.; Perra, M.M.; Shah, S.; Papaioannou, V.; Fabozzi, G.; Ambrozak, D.R.; et al. Acquisition of optimal TFH cell function is defined by specific molecular, positional, and TCR dynamic signatures. Proc Natl Acad Sci U S A 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Parra, E.R. Methods to Determine and Analyze the Cellular Spatial Distribution Extracted From Multiplex Immunofluorescence Data to Understand the Tumor Microenvironment. Front Mol Biosci 2021, 8, 668340. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med 2006, 12, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.W.; Lee, S.A.; Siedner, M.J. Immunologic Biomarkers, Morbidity, and Mortality in Treated HIV Infection. J Infect Dis 2016, 214 Suppl 2, S44–50. [Google Scholar] [CrossRef]
- Austermann, J.; Roth, J.; Barczyk-Kahlert, K. The Good and the Bad: Monocytes' and Macrophages' Diverse Functions in Inflammation. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Yeap, W.H.; Wong, K.L.; Shimasaki, N.; Teo, E.C.; Quek, J.K.; Yong, H.X.; Diong, C.P.; Bertoletti, A.; Linn, Y.C.; Wong, S.C. CD16 is indispensable for antibody-dependent cellular cytotoxicity by human monocytes. Sci Rep 2016, 6, 34310. [Google Scholar] [CrossRef]
- McKenna, E.; Mhaonaigh, A.U.; Wubben, R.; Dwivedi, A.; Hurley, T.; Kelly, L.A.; Stevenson, N.J.; Little, M.A.; Molloy, E.J. Neutrophils: Need for Standardized Nomenclature. Front Immunol 2021, 12, 602963. [Google Scholar] [CrossRef]
- Boritz, E.A.; Darko, S.; Swaszek, L.; Wolf, G.; Wells, D.; Wu, X.; Henry, A.R.; Laboune, F.; Hu, J.; Ambrozak, D.; et al. Multiple Origins of Virus Persistence during Natural Control of HIV Infection. Cell 2016, 166, 1004–1015. [Google Scholar] [CrossRef] [PubMed]
- Doitsh, G.; Cavrois, M.; Lassen, K.G.; Zepeda, O.; Yang, Z.; Santiago, M.L.; Hebbeler, A.M.; Greene, W.C. Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue. Cell 2010, 143, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Smith, A.J.; Wietgrefe, S.W.; Southern, P.J.; Schacker, T.W.; Reilly, C.S.; Estes, J.D.; Burton, G.F.; Silvestri, G.; Lifson, J.D.; et al. Cumulative mechanisms of lymphoid tissue fibrosis and T cell depletion in HIV-1 and SIV infections. J Clin Invest 2011, 121, 998–1008. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Q.; Schuler, T.; Cavert, W.; Notermans, D.W.; Gebhard, K.; Henry, K.; Havlir, D.V.; Gunthard, H.F.; Wong, J.K.; Little, S.; et al. Reversibility of the pathological changes in the follicular dendritic cell network with treatment of HIV-1 infection. Proc Natl Acad Sci U S A 1999, 96, 5169–5172. [Google Scholar] [CrossRef] [PubMed]
- Galvez, C.; Urrea, V.; Dalmau, J.; Jimenez, M.; Clotet, B.; Monceaux, V.; Huot, N.; Leal, L.; Gonzalez-Soler, V.; Gonzalez-Cao, M.; et al. Extremely low viral reservoir in treated chronically HIV-1-infected individuals. EBioMedicine 2020, 57, 102830. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.; Southern, P.J.; Reilly, C.S.; Beilman, G.J.; Chipman, J.G.; Schacker, T.W.; Haase, A.T. Lymphoid tissue damage in HIV-1 infection depletes naive T cells and limits T cell reconstitution after antiretroviral therapy. PLoS Pathog 2012, 8, e1002437. [Google Scholar] [CrossRef] [PubMed]
- Baumjohann, D.; Preite, S.; Reboldi, A.; Ronchi, F.; Ansel, K.M.; Lanzavecchia, A.; Sallusto, F. Persistent antigen and germinal center B cells sustain T follicular helper cell responses and phenotype. Immunity 2013, 38, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Sayin, I.; Radtke, A.J.; Vella, L.A.; Jin, W.; Wherry, E.J.; Buggert, M.; Betts, M.R.; Herati, R.S.; Germain, R.N.; Canaday, D.H. Spatial distribution and function of T follicular regulatory cells in human lymph nodes. J Exp Med 2018, 215, 1531–1542. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, A.; Del Rio Estrada, P.M.; Tharp, G.K.; Trible, R.P.; Amara, R.R.; Chahroudi, A.; Reyes-Teran, G.; Bosinger, S.E.; Silvestri, G. Decreased T Follicular Regulatory Cell/T Follicular Helper Cell (TFH) in Simian Immunodeficiency Virus-Infected Rhesus Macaques May Contribute to Accumulation of TFH in Chronic Infection. J Immunol 2015, 195, 3237–3247. [Google Scholar] [CrossRef]
- Amodio, D.; Cotugno, N.; Macchiarulo, G.; Rocca, S.; Dimopoulos, Y.; Castrucci, M.R.; De Vito, R.; Tucci, F.M.; McDermott, A.B.; Narpala, S.; et al. Quantitative Multiplexed Imaging Analysis Reveals a Strong Association between Immunogen-Specific B Cell Responses and Tonsillar Germinal Center Immune Dynamics in Children after Influenza Vaccination. J Immunol 2018, 200, 538–550. [Google Scholar] [CrossRef]
- Jacobsen, J.T.; Hu, W.; TB, R.C.; Solem, S.; Galante, A.; Lin, Z.; Allon, S.J.; Mesin, L.; Bilate, A.M.; Schiepers, A.; et al. Expression of Foxp3 by T follicular helper cells in end-stage germinal centers. Science 2021, 373. [Google Scholar] [CrossRef] [PubMed]
- Brainard, D.M.; Tager, A.M.; Misdraji, J.; Frahm, N.; Lichterfeld, M.; Draenert, R.; Brander, C.; Walker, B.D.; Luster, A.D. Decreased CXCR3+ CD8 T cells in advanced human immunodeficiency virus infection suggest that a homing defect contributes to cytotoxic T-lymphocyte dysfunction. J Virol 2007, 81, 8439–8450. [Google Scholar] [CrossRef] [PubMed]
- Collins, D.R.; Gaiha, G.D.; Walker, B.D. CD8(+) T cells in HIV control, cure and prevention. Nat Rev Immunol 2020, 20, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Policicchio, B.B.; Cardozo-Ojeda, E.F.; Xu, C.; Ma, D.; He, T.; Raehtz, K.D.; Sivanandham, R.; Kleinman, A.J.; Perelson, A.S.; Apetrei, C.; et al. CD8(+) T cells control SIV infection using both cytolytic effects and non-cytolytic suppression of virus production. Nat Commun 2023, 14, 6657. [Google Scholar] [CrossRef]
- Miles, B.; Miller, S.M.; Folkvord, J.M.; Levy, D.N.; Rakasz, E.G.; Skinner, P.J.; Connick, E. Follicular Regulatory CD8 T Cells Impair the Germinal Center Response in SIV and Ex Vivo HIV Infection. PLoS Pathog 2016, 12, e1005924. [Google Scholar] [CrossRef]
- Di Pilato, M.; Palomino-Segura, M.; Mejias-Perez, E.; Gomez, C.E.; Rubio-Ponce, A.; D'Antuono, R.; Pizzagalli, D.U.; Perez, P.; Kfuri-Rubens, R.; Benguria, A.; et al. Neutrophil subtypes shape HIV-specific CD8 T-cell responses after vaccinia virus infection. NPJ Vaccines 2021, 6, 52. [Google Scholar] [CrossRef]
Figure 1.
Similar in situ cell density of TFH cells in viremic and cART PLWH LNs. (A)Representative examples of CD20 (red), CD4 (green), DAPI (blue), CD57 (grey) and PD1 (cyan) staining pattern from a tonsil and control, viremic and cART HIV LNs (scale bar: 30 μm). (B) Histocytometry gating strategy used for the sequential identification of TFH cells based on PD1 and CD57 expression after nuclear staining-based cell segmentation on Imaris and FlowJo analysis. F and EF areas were defined based on the density for CD20 signal and manually gated on FlowJo to extract cell counts for specific tissue localities. (C) Histocytometry identified PD1hiCD57hi TFH cells were backgated to the original image using Imaris software. Each sphere represents one cell. (D) Bar graph (left) demonstrating the cell density (normalised per mm2 counts) of follicular PD1hi TFH cells in tonsils (N=5), control LNs (N=5), viremic HIV (N=12) and cART HIV LNs (N=20). Viremic and cART were further subdivided based on their TFH counts (HIV vir-high TFH(N=6), HIV vir-low-TFH (N=6), HIV cART-high-TFH (N=6) and HIV cART-low-TFH (N=14)). Each symbol represents one donor. The p values were calculated using the Mann-Whitney test and were corrected using FDR correction with q=0.05 (Suppl.Table 2). Dot graph (right) shows the distribution of PD1hi cell densities among samples. Each symbol represents a follicle. (E) Bar graph demonstrating the normalized per mm2 numbers of follicular PD1hiCD57hi TFH cells in the same groups/subgroups of samples. (F) Linear regression analysis to address the correlation between PD1hi and PD1hiCD57hi absolute follicular counts among different subgroups. Each symbol represents a follicle. R-squared and p-values are displayed on graphs. (G) Linear regression analysis (lower) to address the correlation between normalized follicular PD1hi counts and blood viral load-pVL. Dot graph (upper) showing the pVL differences (as log scale) between the viremic HIV subgroups.
Figure 1.
Similar in situ cell density of TFH cells in viremic and cART PLWH LNs. (A)Representative examples of CD20 (red), CD4 (green), DAPI (blue), CD57 (grey) and PD1 (cyan) staining pattern from a tonsil and control, viremic and cART HIV LNs (scale bar: 30 μm). (B) Histocytometry gating strategy used for the sequential identification of TFH cells based on PD1 and CD57 expression after nuclear staining-based cell segmentation on Imaris and FlowJo analysis. F and EF areas were defined based on the density for CD20 signal and manually gated on FlowJo to extract cell counts for specific tissue localities. (C) Histocytometry identified PD1hiCD57hi TFH cells were backgated to the original image using Imaris software. Each sphere represents one cell. (D) Bar graph (left) demonstrating the cell density (normalised per mm2 counts) of follicular PD1hi TFH cells in tonsils (N=5), control LNs (N=5), viremic HIV (N=12) and cART HIV LNs (N=20). Viremic and cART were further subdivided based on their TFH counts (HIV vir-high TFH(N=6), HIV vir-low-TFH (N=6), HIV cART-high-TFH (N=6) and HIV cART-low-TFH (N=14)). Each symbol represents one donor. The p values were calculated using the Mann-Whitney test and were corrected using FDR correction with q=0.05 (Suppl.Table 2). Dot graph (right) shows the distribution of PD1hi cell densities among samples. Each symbol represents a follicle. (E) Bar graph demonstrating the normalized per mm2 numbers of follicular PD1hiCD57hi TFH cells in the same groups/subgroups of samples. (F) Linear regression analysis to address the correlation between PD1hi and PD1hiCD57hi absolute follicular counts among different subgroups. Each symbol represents a follicle. R-squared and p-values are displayed on graphs. (G) Linear regression analysis (lower) to address the correlation between normalized follicular PD1hi counts and blood viral load-pVL. Dot graph (upper) showing the pVL differences (as log scale) between the viremic HIV subgroups.
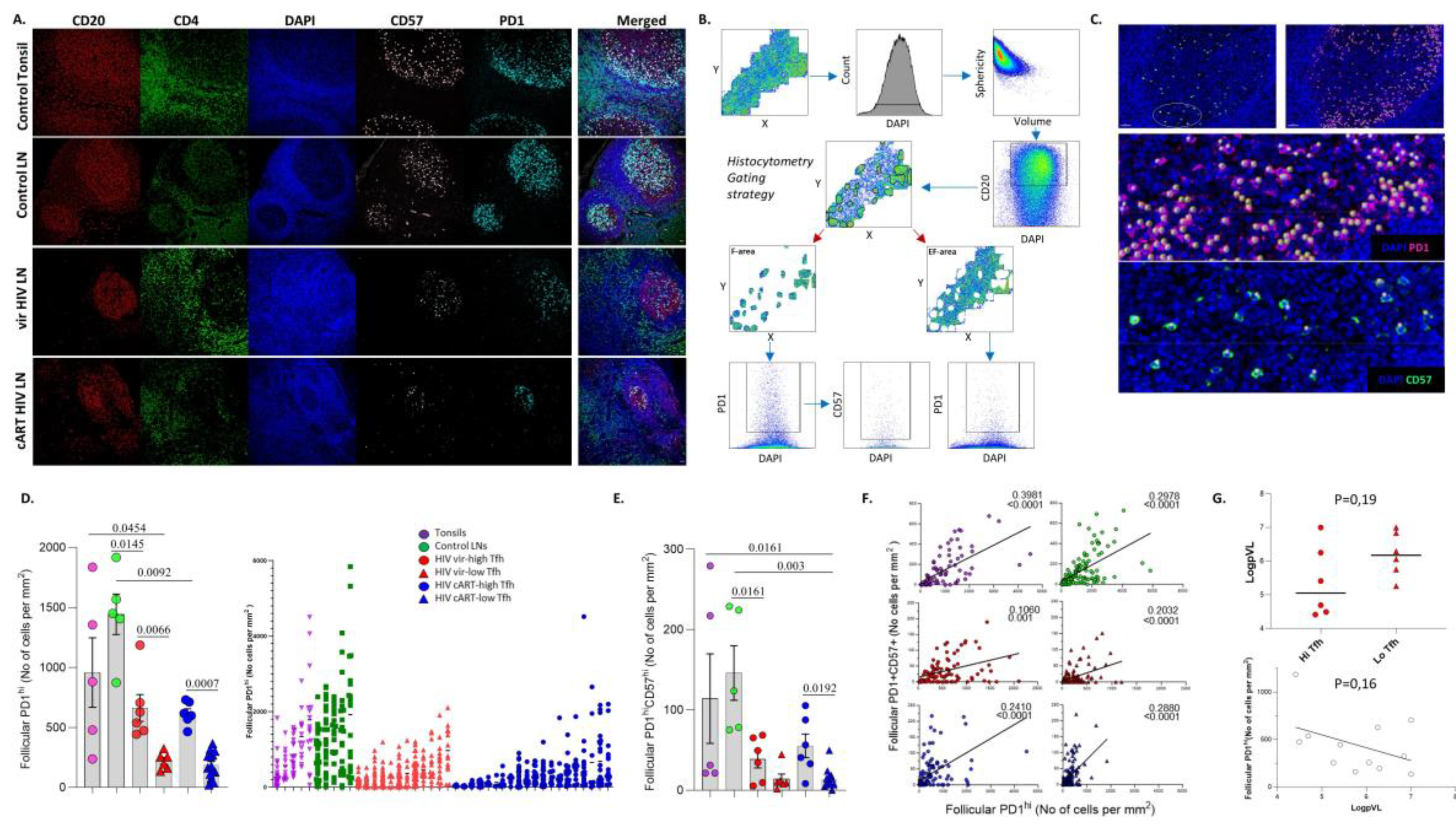
Figure 2.
A highly scattered distribution of TFH cells in HIV infected compared to non-infected lymphoid tissues. (A) Representative immunofluorescence images showing the distribution of CD20hi (blue) and PD1hi (red) cells in follicles from one control, one vir-HIV high-TFH and one cART HIV high-TFH tissue. The corresponding digitalized (generated by the Python distance analysis script) images are shown too. (B) The distribution bar graphs for the minimum distance between CD20 and PD1hi cells in two follicular areas (control-left, cART-right) (upper panel). Diagrams showing the theoretical (blue) and experimental (red) Poisson curves for the distribution of PD1hi cells in two follicular areas (control-left, cART-right) (lower panel). Bar graphs showing the G function analysis for total CD20hi (C) and PD1hi TFH cells ((control LNs (N=54), vir-HIV high-TFH (N=42), vir-HIV low-TFH (N=51), cART HIV high-TFH (N=31) and cART HIV low-TFH (N=55). (D) in individual follicles from control and infected LNs. (E) The mean of minimum distance values between CD20hi and PD1hi cells in individual follicles from control and infected LNs is shown ((control LNs (N=54), vir- HIV high-TFH (N=42), vir-HIV low-TFH (N=51), cART HIV high-TFH (N=31) and cART HIV low-TFH (N=55)). (F) The G function analysis for PD1hiCD57hi (left) and PD1hiCD57lo (right) TFH cells in tonsils, control and infected LNs is shown (( tonsils (N=29), (control LNs (N=29), vir-HIV high-TFH (N=13), vir-HIV low-TFH(N=5), cART HIV high-TFH (N=11) and cART HIV low-TFH (N=7)). (G) Bar graphs showing the mean of minimum distances between CD20hi and PD1hiCD57lo (left) or PD1hiCD57hi (right) TFH cells in tonsils, control and infected LNs ( tonsils (N=29), control LNs (N=29), vir-HIV high-TFH (N=13), vir-HIV low-TFH (N=5), cART HIV high-TFH (N=11) and cART HIV low-TFH (N=7)). Each dot represents one follicle in all presented graphs. The p values were calculated using the Mann-Whitney test and were corrected using FDR correction with q=0.05 (Suppl.Table 2).
Figure 2.
A highly scattered distribution of TFH cells in HIV infected compared to non-infected lymphoid tissues. (A) Representative immunofluorescence images showing the distribution of CD20hi (blue) and PD1hi (red) cells in follicles from one control, one vir-HIV high-TFH and one cART HIV high-TFH tissue. The corresponding digitalized (generated by the Python distance analysis script) images are shown too. (B) The distribution bar graphs for the minimum distance between CD20 and PD1hi cells in two follicular areas (control-left, cART-right) (upper panel). Diagrams showing the theoretical (blue) and experimental (red) Poisson curves for the distribution of PD1hi cells in two follicular areas (control-left, cART-right) (lower panel). Bar graphs showing the G function analysis for total CD20hi (C) and PD1hi TFH cells ((control LNs (N=54), vir-HIV high-TFH (N=42), vir-HIV low-TFH (N=51), cART HIV high-TFH (N=31) and cART HIV low-TFH (N=55). (D) in individual follicles from control and infected LNs. (E) The mean of minimum distance values between CD20hi and PD1hi cells in individual follicles from control and infected LNs is shown ((control LNs (N=54), vir- HIV high-TFH (N=42), vir-HIV low-TFH (N=51), cART HIV high-TFH (N=31) and cART HIV low-TFH (N=55)). (F) The G function analysis for PD1hiCD57hi (left) and PD1hiCD57lo (right) TFH cells in tonsils, control and infected LNs is shown (( tonsils (N=29), (control LNs (N=29), vir-HIV high-TFH (N=13), vir-HIV low-TFH(N=5), cART HIV high-TFH (N=11) and cART HIV low-TFH (N=7)). (G) Bar graphs showing the mean of minimum distances between CD20hi and PD1hiCD57lo (left) or PD1hiCD57hi (right) TFH cells in tonsils, control and infected LNs ( tonsils (N=29), control LNs (N=29), vir-HIV high-TFH (N=13), vir-HIV low-TFH (N=5), cART HIV high-TFH (N=11) and cART HIV low-TFH (N=7)). Each dot represents one follicle in all presented graphs. The p values were calculated using the Mann-Whitney test and were corrected using FDR correction with q=0.05 (Suppl.Table 2).
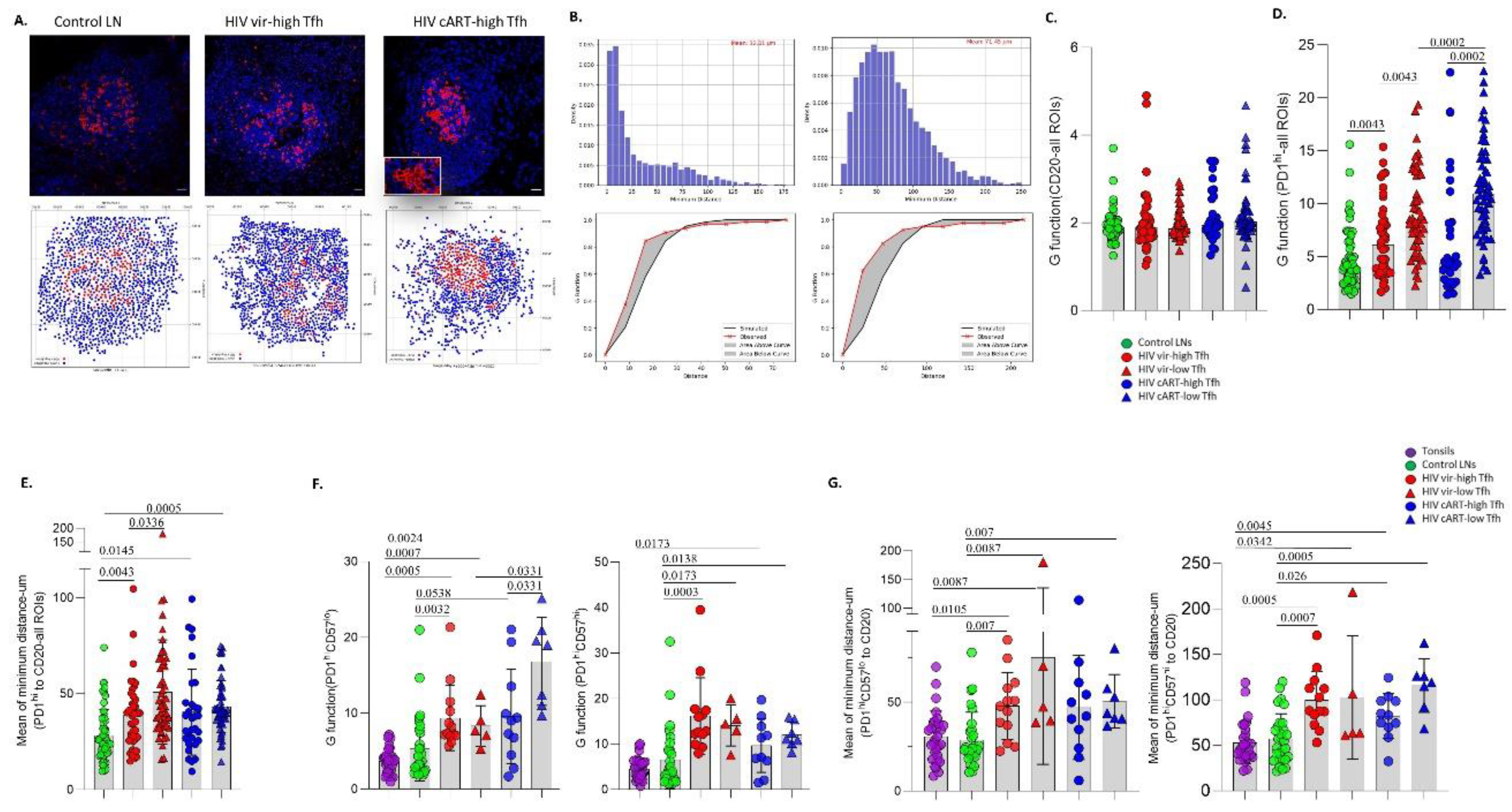
Figure 3.
Preferential accumulation of FOXP3hi CD4+ T cells in cART HIV follicular areas. (A) Representative examples of CD20(red), CD4 (green), DAPI (blue) and FOXP3 (yellow) staining patternfrom a tonsil and control, viremic and cART HIV LNs (scale bar: 30 μm). (B) Histocytometry immunophenotyping gating strategy used for the sequential identification of FOXP3hi CD4+ T cells in follicular and extrafollicular areas in a control LN (C) Representative back gating of Histocytometry identified FOXP3hi CD4+ T cells, using Imaris software. Each sphere represents one cell. (D) Bar graph (left) demonstrating the normalised per mm2counts of extrafollicular FOXP3hi CD4+ T cells in tonsils (N=5), control LNs (N=5), HIV vir-high-TFH (N=6), HIV vir-low-TFH(N=6), HIV cART-high-TFH (N=6) and HIV cART-low-TFH (N=14). (E) Bar graph showing the normalised per mm2 counts of follicular FOXP3hi CD4+ T cells in the same tissue samples. Each symbol represents one donor. (F) Bar graph with connecting lines demonstrating the normalised per mm2 counts of FOXP3hi CD4+ T cells in follicular and extrafollicular regions in each tissue analysed. (G) Representative immunofluorescence images showing the distribution of FOXP3hi (green) and CD20hi (red) cells in follicles from one cART-high-TFH and one cART-low-TFH tissue. The corresponding digitalized (generated by the Python distance analysis script) images are shown too. (H) Bar graph showing the calculated G function values for follicular FOXP3hi CD4+ T cells in cART-high TFH (blue circles) and cART-low TFH (blue triangles) (tissues HIV cART-high TFH (N=22) and HIV cART-low TFH (N=64)). The p values were calculated using the Mann-Whitney test and were corrected using FDR correction with q=0.05(Suppl.Table 2).
Figure 3.
Preferential accumulation of FOXP3hi CD4+ T cells in cART HIV follicular areas. (A) Representative examples of CD20(red), CD4 (green), DAPI (blue) and FOXP3 (yellow) staining patternfrom a tonsil and control, viremic and cART HIV LNs (scale bar: 30 μm). (B) Histocytometry immunophenotyping gating strategy used for the sequential identification of FOXP3hi CD4+ T cells in follicular and extrafollicular areas in a control LN (C) Representative back gating of Histocytometry identified FOXP3hi CD4+ T cells, using Imaris software. Each sphere represents one cell. (D) Bar graph (left) demonstrating the normalised per mm2counts of extrafollicular FOXP3hi CD4+ T cells in tonsils (N=5), control LNs (N=5), HIV vir-high-TFH (N=6), HIV vir-low-TFH(N=6), HIV cART-high-TFH (N=6) and HIV cART-low-TFH (N=14). (E) Bar graph showing the normalised per mm2 counts of follicular FOXP3hi CD4+ T cells in the same tissue samples. Each symbol represents one donor. (F) Bar graph with connecting lines demonstrating the normalised per mm2 counts of FOXP3hi CD4+ T cells in follicular and extrafollicular regions in each tissue analysed. (G) Representative immunofluorescence images showing the distribution of FOXP3hi (green) and CD20hi (red) cells in follicles from one cART-high-TFH and one cART-low-TFH tissue. The corresponding digitalized (generated by the Python distance analysis script) images are shown too. (H) Bar graph showing the calculated G function values for follicular FOXP3hi CD4+ T cells in cART-high TFH (blue circles) and cART-low TFH (blue triangles) (tissues HIV cART-high TFH (N=22) and HIV cART-low TFH (N=64)). The p values were calculated using the Mann-Whitney test and were corrected using FDR correction with q=0.05(Suppl.Table 2).
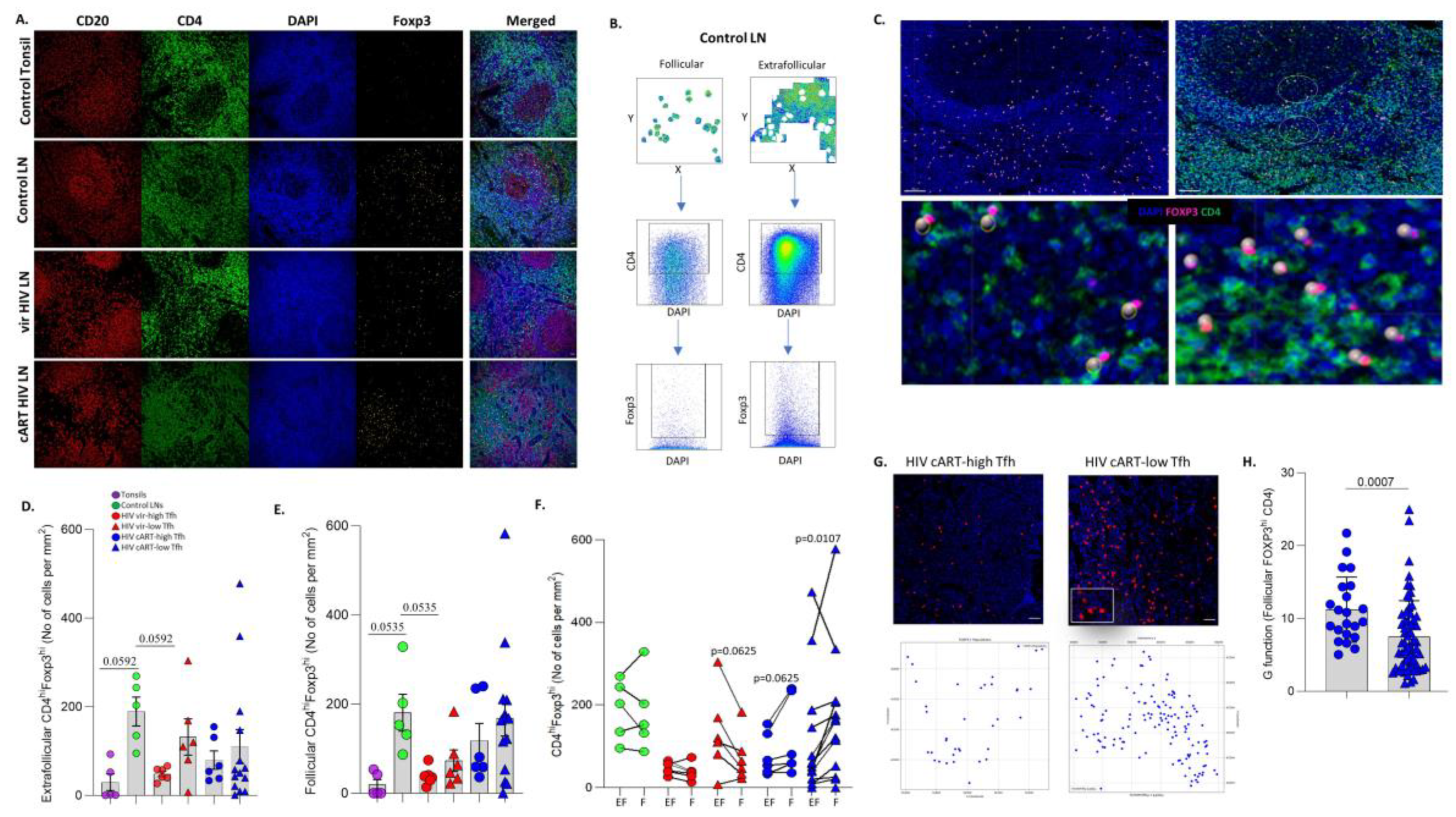
Figure 4.
Accumulated LN GrzBhiCD8+ T cells are negatively associated with PLWH blood viral load. (A) Representative examples of GRZb (red), CD8 (green) and DAPI (blue) staining pattern from a tonsil and control, vir HIV and cART HIV LNs (scale bar: 30 μm). (B) Histocytometry gating strategy used for the sequential identification of bulk CD8+ and GrzBhi CD8+ T cells. (C) Representative back gating of Histocytometry identified GrzBhi CD8+ cells using Imaris software. Each sphere represents one cell. (D) Bar graph (upper) demonstrating the normalized per mm2 counts of bulk CD8+ cells in tonsils (N=5), control LNs (N=5), vir-HIV high-TFH (N=6), vir-HIV low-TFH (N=6), cART HIV high-TFH (N=6) and cART HIV low-TFH (N=14). Bar graph (lower) showing the normalized per mm2 counts of GrzBhi CD8+ T cells in the same tissue samples. Each symbol represents one donor. (E) Linear regression analysis showing the correlation between LN CD8+ (upper panel) and LN GrzBhi CD8+ (lower panel) normalized T cell counts with blood viral load. (F) Linear regression analysis showing the correlation between LN CD8+ and LN GrzBhi CD8+ T cell counts in cART HIV high-TFH (upper panel) and cART HIV low-TFH (lower panel). (G) Linear regression analysis to address the correlation between extrafollicular (upper panel) and follicular (lower panel) FOXP3hi CD4+ and LN CD8+ normalized T cell counts in cART HIV low-TFH subgroup. (H) Bar graph showing the G function values for the LN GrzBhi CD8+ T cells in the HIV subgroups (vir-HIV high-TFH (N=6), vir-HIV low-TFH (N=5), cART HIV high-TFH (N=6) and cART HIV low-TFH (N=10)). The p values were calculated using the Mann-Whitney test and corrected using FDR correction with q=0.05 (Suppl.Table 2).
Figure 4.
Accumulated LN GrzBhiCD8+ T cells are negatively associated with PLWH blood viral load. (A) Representative examples of GRZb (red), CD8 (green) and DAPI (blue) staining pattern from a tonsil and control, vir HIV and cART HIV LNs (scale bar: 30 μm). (B) Histocytometry gating strategy used for the sequential identification of bulk CD8+ and GrzBhi CD8+ T cells. (C) Representative back gating of Histocytometry identified GrzBhi CD8+ cells using Imaris software. Each sphere represents one cell. (D) Bar graph (upper) demonstrating the normalized per mm2 counts of bulk CD8+ cells in tonsils (N=5), control LNs (N=5), vir-HIV high-TFH (N=6), vir-HIV low-TFH (N=6), cART HIV high-TFH (N=6) and cART HIV low-TFH (N=14). Bar graph (lower) showing the normalized per mm2 counts of GrzBhi CD8+ T cells in the same tissue samples. Each symbol represents one donor. (E) Linear regression analysis showing the correlation between LN CD8+ (upper panel) and LN GrzBhi CD8+ (lower panel) normalized T cell counts with blood viral load. (F) Linear regression analysis showing the correlation between LN CD8+ and LN GrzBhi CD8+ T cell counts in cART HIV high-TFH (upper panel) and cART HIV low-TFH (lower panel). (G) Linear regression analysis to address the correlation between extrafollicular (upper panel) and follicular (lower panel) FOXP3hi CD4+ and LN CD8+ normalized T cell counts in cART HIV low-TFH subgroup. (H) Bar graph showing the G function values for the LN GrzBhi CD8+ T cells in the HIV subgroups (vir-HIV high-TFH (N=6), vir-HIV low-TFH (N=5), cART HIV high-TFH (N=6) and cART HIV low-TFH (N=10)). The p values were calculated using the Mann-Whitney test and corrected using FDR correction with q=0.05 (Suppl.Table 2).
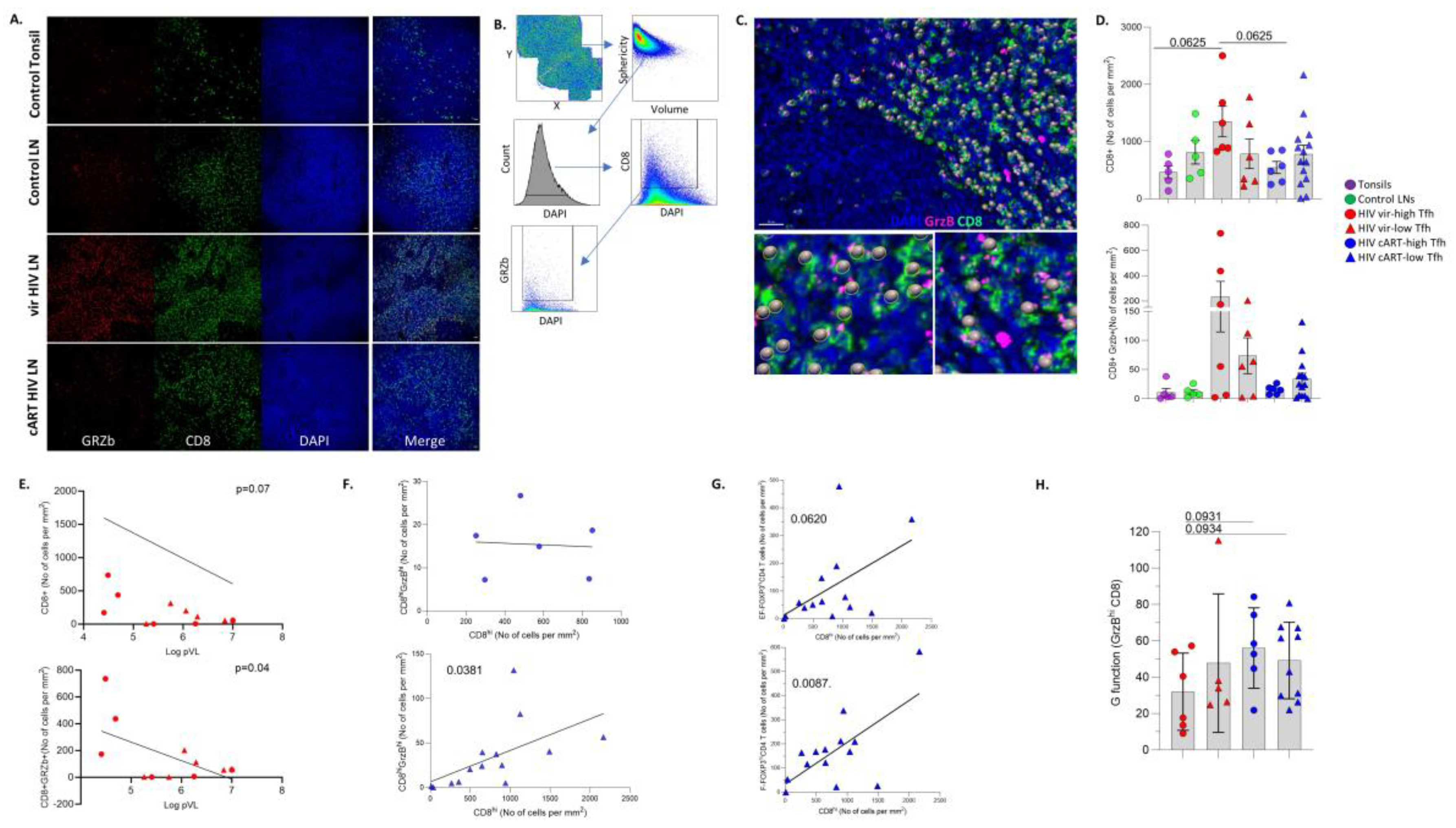
Figure 5.
Altered innate immunity signatures among control and HIV samples. (A) Representative examples of DAPI (blue), CD15 (cyan), CD16 (magenta), CD163 (yellow) and CD68 (red) staining pattern from a tonsil and control, vir-HIV and cART-HIV LNs (scale bar: 30 μm). (B) Histocytometry gating strategy used for the sequential identification of CD15hi, CD16hi, CD163hi and CD68hi innate cells. (C) Bar graphs demonstrating the normalised per mm2 counts of bulk CD68hiCD163lo (upper left), CD68loCD163hi (upper middle), CD68hiCD163hi (upper right), CD15hiCD16lo (lower left), CD15hiCD16lo(lower middle) and CD15hiCD16lo (lower right) in tonsils (N=5), control LNs (N=5), vir-HIV high-TFH (N=6), vir-HIV low-TFH (N=6), cART HIV high-TFH (N=6) and cART HIV low-TFH (N=14)). Each symbol represents one donor. (D) Linear regression analysis to address the correlation between CD8+ (upper panel) or GrzBhi CD8+ (lower panel) T cells and CD16hiCD15lo cells in vir-HIV high-TFH tissues. (E) Bar graph showing the mean values of the minimum distances between GrzBhiCD8+ T and CD16hi cells in tissues from the HIV subgroups vir- HIV high-TFH (N=6), vir-HIV low-TFH (N=3), cART HIV high-TFH (N=4) and cART HIV low-TFH (N=10). Each dot represents a different donor. The p values were calculated using the Mann-Whitney test and p values were corrected using FDR correction with q=0.05 (Suppl.Table 2).
Figure 5.
Altered innate immunity signatures among control and HIV samples. (A) Representative examples of DAPI (blue), CD15 (cyan), CD16 (magenta), CD163 (yellow) and CD68 (red) staining pattern from a tonsil and control, vir-HIV and cART-HIV LNs (scale bar: 30 μm). (B) Histocytometry gating strategy used for the sequential identification of CD15hi, CD16hi, CD163hi and CD68hi innate cells. (C) Bar graphs demonstrating the normalised per mm2 counts of bulk CD68hiCD163lo (upper left), CD68loCD163hi (upper middle), CD68hiCD163hi (upper right), CD15hiCD16lo (lower left), CD15hiCD16lo(lower middle) and CD15hiCD16lo (lower right) in tonsils (N=5), control LNs (N=5), vir-HIV high-TFH (N=6), vir-HIV low-TFH (N=6), cART HIV high-TFH (N=6) and cART HIV low-TFH (N=14)). Each symbol represents one donor. (D) Linear regression analysis to address the correlation between CD8+ (upper panel) or GrzBhi CD8+ (lower panel) T cells and CD16hiCD15lo cells in vir-HIV high-TFH tissues. (E) Bar graph showing the mean values of the minimum distances between GrzBhiCD8+ T and CD16hi cells in tissues from the HIV subgroups vir- HIV high-TFH (N=6), vir-HIV low-TFH (N=3), cART HIV high-TFH (N=4) and cART HIV low-TFH (N=10). Each dot represents a different donor. The p values were calculated using the Mann-Whitney test and p values were corrected using FDR correction with q=0.05 (Suppl.Table 2).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



