Submitted:
05 July 2024
Posted:
08 July 2024
You are already at the latest version
Abstract
Cardiomyopathy syndrome (CMS) caused by piscine myocarditis virus (PMCV) has emerged in aquaculture of Atlantic salmon (Salmo salar). The lack of cell culture has hampered studies of the infection. In this study samples from naturally PMCV infected Atlantic salmon from commercial farms were collected and used.
In situ hybridization revealed intense staining for PMCV RNA in myocard in the spongiform layer of the heart, but almost no staining in the compact layer. In kidneys only sporadic staining was seen. Viral RNA was present in all organs, with the highest loads in the heart, kidney, and spleen.
The high viral PMCV RNA loads in the heart were caused by extensive viral mRNA transcription. The high ratio of viral mRNA to viral genomic dsRNA, indicated active transcription but limited production of new viral particles. This suggests that the histopathological changes of the heart are caused by viral mRNA and corresponding viral proteins.
The production of full-length transcripts is regulated, with a reduction in the relative amount of ORF3-containing transcripts at high transcription rates. Efforts to identify alternative hosts, such as fungi, were inconclusive, as fungal sequences were found inconsistently in salmon tissue samples. The results reinforce the need for further research to fully understand PMCV’s life cycle and potential alternative hosts and its whereabouts when it is not infecting the heart of the At-lantic salmon.
Keywords:
piscine myocarditis virus (PMCV)
; Salmo salar
; Totivirus
1. Introduction
The development of large scale, intensive marine aquaculture of Atlantic salmon (Salmo salar) has generated a melting pot for emerging infectious diseases. One example of a disease that has emerged in this industry is cardiomyopathy syndrome (CMS), a disease which predominantly appears in the later part of the salmon’s production cycle and has caused significant fish welfare and economic implications. CMS is commonly observed as a sudden increase in mortality in a net population, with or without observations of preceding clinical signs [1]. The gross pathological findings of diseased fish are circulatory disturbances due to acute or chronic heart failure and the histopathology is characterized by subendocardial inflammation and myocarditis, degeneration, necrosis, and progressive fibrosis of the atrium and the spongious myocardium [2]. The histological changes of CMS was described already in the 1980-ies [3] but it took about twenty years before CMS was demonstrated to be a transmissible disease [4] and then subsequently found to be associated with infection of Piscine myocarditis virus (PMCV) [5,6]. It has not been possible to cultivate PMCV in fish cell lines, and experimental infection is commonly done by injection of each individual fish due to slow horizontal transmission rate [7].
The physical structure of the PMCV particles resembles that of Totiviridae [5], with non-enveloped, single-layered, icosahedral particles composed of one major polypeptide. The genome of classical totiviruses consists of a single molecule linear double-stranded RNA (dsRNA) with two open reading frames (ORFs), where ORF 1 encodes for the major capsid protein (MCP) and ORF2 for the RNA-dependent-RNA-polymerase (RdRp). Due to ribosomal -1 frameshifting ORF2 can be translated as a fusion with ORF1 approximately once every 100 translation events causing some copies of the capsid proteins to be fusion proteins of MCP-RdRp [8]. Some totiviruses has an extra, small toxin encoding dsRNA in a satellite particle [9]. PMCV have been found as distinct from the classical totiviruses by having an extra ORF3 in the 3’- end of the genome, encoding a protein of unknown function [5].
Findings from both field and later experimental studies indicate that horizontal transmission of PMCV from fish to fish may be very slow. For example, transmission of the infection from a heavily infected fish population in one net to another within a farm was observed to take several months, if at all [2]. Similarly, it was observed that PMCV infected fish used in a cohabitation challenge had transmitted the infection only to 54% of the naive fish in a tank after 24 weeks of cohabitation [7], however in earlier experiments it was found that naive co-habitant fish in a tank got infected with a 4-week delay [5].
The totiviruses were originally associated with latent infections in unicellular organisms such as yeast, protozoa, and fungi [10]. Viruses causing latent infections in unicellular organisms may never leave their host cell and do not need the capsid structures that recognize host cell receptors for the entry of cells from the extracellular environment, and the capsids therefore primary shield the genome from cellular antiviral response and facilitate the synthesis of viral RNA. Transmission of virus from one latent infected cell to another is dominantly vertical and occurs at cell division and cell-cell fusion [11]. In contrast, viruses with a capsid structure and a dsRNA genome, which enter cells from the external environment, must possess a mechanism for cell entry. For viruses tentatively listed as totiviruses infecting arthropods, such as the shrimp virus infectious myonecrosis virus (IMNV), it has been shown that the virus particles have fiber protrusions at the five-fold axes that are part of the machinery for the extracellular transmission [12]. The totivirus Omono River virus (OmRV), that infects mosquitos, has an extra crown protein structure on the capsid that may facilitate horizontal cell-to-cell transmission [13]. The ORF-1 encoded proteins of the arthropod infecting totiviruses contain 2A-like motifs, which is an oligopeptide sequence mediating a ribosome skipping effect, producing two polyproteins without the need of an exogenous proteinase [14]. In IMNV 2A motifs gives at least two polypeptides, in addition to the MCP, that are assumed to be part of the fiber protrusions and to counteract the host cell’s antiviral response [15]. It can therefore be argued that the evolution of arthropod-infecting totiviruses have led to the acquisition of fiber-coding sequences to allow extracellular transmission [12]. These viruses can be pathogenic for their hosts, and it has been shown that purified virions can reproduce the disease in pathogen-free animals [16]. Giardia lamblia virus (GLV), a totivirus able to enter its unicellular host from the extracellular environment, has surface loops, and similar are also present in mosquito infecting OmRV, and thought to be of importance for extracellular transmission [17].
Interestingly, the PMCV virion is in many ways similar to classical totiviruses by having overlapping ORF1 and ORF2; the size of the PMCV genome, 6.7 kbp, is within the range of the classical totiviruses, while the genome of the arthropod-infecting totiviruses is bigger (7.5 kbp) [15]; there are no 2A-like motifs present in the ORF1 sequence of PMCV or in classical totiviruses, as it is in arthropod totiviruses [5]; and there are no protrusions visible on EM of the PMCV particle [5]. However, the presence of ORF3 of the PMCV differentiates the PMCV from the classical totiviruses.
In this study we addressed the question whether Altantic salmon is the natural host of PMCV. We characterized the infection dynamics and tissue tropism to determine if they represent a natural infection and searched for indications of co-infection with other host organisms. Additionally, we investigated the genome organization of PMCV to see if it aligned with classical totiviruses.
2. Materials and Methods
2.1. Samples
All samples originated from commercial marine aquaculture sites for Atlantic salmon, where PMCV had earlier been detected in the fish populations. The production sites were located along the Norwegian coast. Samplings were conducted by local fish health veterinarians that regularly monitored the health status of the fish populations at the production sites. Ten fish were usually included in each sampling and the procedures followed the national regulations called “The aquaculture act” for animal welfare (“Regulation on the operation of aquaculture facilities §34. Killing of fish.”).
At sampling the fish population was clinically characterized by the local fish health veterinarians as apparently healthy or with clinical signs of CMS. The fish were euthanized with a lethal dose of anesthetic. Dissections were conducted aseptically, and the organs collected included heart, head kidney, spleen, liver, gills, skin (base of pectoral fin and scrapes of skin), muscle, pyloric appendices, mid- and hindgut, and put in RNAlater® (LifeTechnologies, Carlsbad, CA, USA), L15 (Gibco, NY, USA) or in 96 % ethanol or in PBS and shipped cool overnight. Upon arrival, samples intendent for RNA isolation were stored at − 20 °C. Heart and kidney samples for in-situ hybridizations were fixed in 10% neutral buffered formalin and after 24 – 36 hrs the formalin was replaced with ethanol and samples stored at 4 °C.
Blood samples were centrifuged, serum collected and stored at -20°C.
Additionally, heart and head kidney samples from clinically healthy, PMCV negative, fish were sampled at Centre for fish trials, Norwegian University of Life Sciences, Ås, Norway.
2.2. RNA Extraction
Total RNA was isolated using RNeasy mini QIAcube kit (Qiagen, Hilden, Germany). Briefly, a 2 mm3 cube, sliced into small pieces was placed in a safe lock tube with 500 µl buffer RLT (Qiagen) with 40 mM DTT and a steel bead (5 mm, Qiagen). The tissue was disrupted and homogenised in a TissueLyser II (Qiagen) for 5 minutes at 25 Hz, total RNA was isolated following the instructions of the manufacturer and eluted in 30 µl RNase-free water.
Viral RNA was isolated using QIAamp Viral RNA Mini (Qiagen). Briefly, 140 µl tissue homogenate was mixed with 560 µl buffer AVL (Qiagen) containing carrier RNA, and viral RNA was isolated following the instructions of the manufacturer and eluted in 50 µl buffer AVE (RNase-free water with 0.04% sodium azide).
2.3. Extraction of Virion dsRNA
A 1 mm3 cube was placed in a safe lock tube with 250 µl buffer (10 mM Tris, 1 mM EDTA, pH 8.0) and a steel bead (5 mm, Qiagen). The tissue was disrupted and homogenised in a TissueLyser II (Qiagen) for 10 minutes at 25 Hz, then 4 U PureLinkTM RNase A, (LifeTechnologies) was added and incubated at 37 °C for 30 min. Thereafter 4 U DNase, (ThermoFisher, Lithuania) was added and further incubated at 37 °C for 30 min. ViralRNA was isolated as described above.
2.4. RT-qPCR
The Brilliant III Ultra-Fast QRT-PCR Master Mix, (Agilent Technologies, Santa Clara, CA, USA) was used for detection and estimation of amount of viral RNA. The RT-qPCR was set up using the following conditions: 1 x QRT-PCR master mix, 30 nM ROX reference dye, 400 nM primers (Table 1), 300 nM probe (Table 1), 1 mM DTT, 0,75 µl RT/RNase block and 100 ng total RNA and tested in a total volume of 15 µl.
Prior to RT-qPCR, the RNA was denatured at 95°C for 5 min and a reverse transcription phase for 10 minutes at 50°C followed by a hot start phase at 95°C for 3 min and 40 cycles of 95°C for 5 s and 60°C for 10 s. The samples were run in duplicate on AriaMx (Agilent Technologies, Penang, Malaysia), and a sample was defined as positive if both parallel samples had a Cq <40.
To test the efficiency and reproducibility of PMCV primer sets, cDNA from total RNA of heart samples from two fish were serial 1:10 diluted and used as templates.
Detection of Serna and dsRNA. RACE
Total RNA (1 µg) isolated from heart and head kidney samples with a Cq value for PMCV 24 or lower, was incubated with 1,12 U PureLink™ RNaseA at low salt concentration or high salt concentration (0.5 M NaCl). At 0.5 M NaCl, dsRNA but not ssRNA is selectively protected from RNaseA degradation [20]. Following incubation with RNaseA at 25 °C for 45 min, RNA was recovered using RNeasy MinElute Cleanup Kit (Qiagen) as per manufacturer’s instructions.
We also adapted the method established by Polinski et al. used for quantifying reoviral ssRNA and dsRNA [21]. The premise is that dsRNA segments will not be transcribed to cDNA unless they are first denatured into single strands by heating (95°C for 3 min).
To try to characterize 5' ends from eventually low-copy viral transcripts not corresponding to full-length copies of the genome a 5´ RACE System for Rapid Amplification of cDNA Ends was used following the instructions of the manufacturer (Thermo Fisher Scientific, USA).
2.5. Statistical Analysis
Means of PMCV dsRNA and ssRNA found by RT-qPCR in different organs were compared using the Student’s t-test.
In-situ hybridization (ISH)
Chromogenic detection: RNAscope® 2.5 HD Detection Kit RED, (Advanced Cell Diagnostic (ACD), Newark, CA, USA) was used for chromogenic detection of PMCV in tissue samples. Following the manufacturer’s instructions, paraffin embedded tissue sections (5 µm) of heart and kidney from PMCV infected fish and control fish were dewaxed at 60 °C for 90 min in ACD HybEZ™ II oven, followed by hydrogen peroxide treatment for 10 min at room temperature. Samples were boiled in RNAscope target antigen retrieval reagent for 15 min and then incubated with RNAscope protease plus at 40 °C for 15 min in a HybEZ™ oven. Each section was hybridized by a RNAscope probe (Table 2) designed against the positive strand (mRNA) of PMCV-ORF1 or PMCV-ORF2 or the negative strand (genomic) of PMCV-ORF2 (ACD catalogue numbers, 812021-C2, 555231, 1219761-C3) for 2 hrs at room temperature. Probe targeting Peptidylpropyl Isomerase B (PPIB) in Atlantic salmon (ACD, catalogue number-494421) was used as reference target gene to test for RNA integrity in the samples. As negative control, probe-DapB (ACD catalogue number-310043) was used to evaluate cross reactivity. Fast Red chromogenic substrate was used for detection of signals amplified following manufacturer’s instructions. Counterstaining was done with 50% Gill’s hematoxylin solution and the samples mounted with EcoMount (BioCare Medical, Pacheco, CA, USA).
Imaging was performed by bright field microscopy, DM6B-Z (Leica Microsystems, Wetzlar, Germany).
Fluorescent In-situ hybridization (FISH): RNAscope® Multiplex Fluorescent Reagent Kit v2, (Advanced Cell Diagnostic) was used for RNA-FISH. Samples were treated the same way as described for RuneScape Chromogenic. The probes were assigned a fluorophore, Opal 520, with emission/excitation wavelength 494/525. Each section was counter stained by adding DAPI (fluorescent DNA stain) for 30 sec at room temperature. Mounting was performed by adding 1-2 drops of Prolong Gold antifade mounting reagent (Thermo Fisher Scientific).
Fluorescent images were obtained using a DMi8-CS Confocal Microscope (Leica Microsystems, Wetzlar, Germany) or DM6 B Thunder Microscope (Leica Microsystems, Wetzlar, Germany). Lasers with wavelength of 405 (DAPI) and 488 (FITC) nm were used.
3. Results
3.1. In Situ Localization of PMCV RNA In Heart Ventricle And Kidney
To determine the localization of PMCV RNA in the heart ventricle, ISH protocols targeting the PMCV-ORF2, the RNA-dependent RNA polymerase gene, were established. The specificity of the detection was shown by positive and negative ISH controls and that probes against PMCV ORF1 and ORF2 recognized the same cells (Figure A1).
Detection by chromogenic labeled probes in heart with low Cq values, i.e. high viral RNA loads showed an intense and confluent staining of myocardial cells in stratum spongiosum with a sharp demarcation to the stratum compactum (Figure 1A). A few positive cells were detected in the stratum compactum. The intense, and confined staining of the spongious layer with a sharp demarcation to the compact layer were confirmed by fluorescent ISH probes where a cross section of the heart ventricle showed strong staining in the spongiform trabeculae and lack of emission from the compact and epicardial layers (Figure 1B). The strong, demarcated staining in spongiform layer of heart seen by ISH was in line with earlier described histopathological changes largely restricted to the spongy portion of the heart [22]. Counter staining for nuclear DNA revealed that PMCV infection could not be observed in the blood cells filling the space between spongiform trabecula (Figure 1C1, C2) and PMCV RNA could only be detected in parts of the spongiosum trabeculae.
The ISH staining for PMCV in kidney, on the other hand, was sporadic and punctate (Figure 1D). There was no indication of staining of the kidney melano-macrophages, positive signals for PMCV were also seen in tubuli structures (Figure 1D, arrow) possibly indicating shedding or infection of kidney epithelial cells. The scattered presence of PMCV RNA in the kidney was in sharp contrast to the massive staining of the heart in individuals with high viral RNA load.
3.2. PMCV RNA Load in Different Organs
Ten fish from farm A were sampled two months after PMCV had been detected at the farm. At the time of sampling the fish did not show clinical signs of disease and there was no increased mortality. Six fish were RT-qPCR positive for PMCV in all organs tested and in two of these the viral load in the heart were assessed as high, i.e., Cq 16.4 and 16.9. In fish that were RT-qPCR positive for PMCV the largest viral RNA loads were present in the heart, kidney, and spleen and not significantly different, with average Cqs of RT-qPCR 21.9, 23.1 and 23.2, respectively. Significantly less viral RNA was present in gills, liver, muscle, skin, pyloric ceca, mid- and hindgut with average Cqs from 26.2 to 29.1 in positive fish (Table 3). One fish was positive only in heart, kidney and in skin scrape, and interestingly one fish in skin scrape only, while two fish were negative for PMCV by RTqPCR in all samples tested (Table 3). As for field samples in general, we do not know when the population had originally been exposed to PMCV. The efficiency and reproducibility of the primer set used, and probe have been determined earlier [6].
The further studies of viral RNA loads focused on heart and kidney representing two organs with high virus load. Ten fish from each of farm B and C, where similarly to farm A, the fish did not show clinically signs of disease at sampling had average Cqs in heart and kidney, not significantly different, of 19.5, 22.6 (Farm B) and 21.0, 20.3 (Farm C), respectively.
In Farm D on the other hand, where the fish had tentative clinical signs of CMS at time of sampling, there was significantly higher (p<0.01) amount of RNA in heart samples compared to kidney samples, with average Cqs of 14.6 ±1.8 and 20.4 ±0.94, respectively.
3.3. PMCV RNA form In Various Organs At Low And High Viral Load
The findings of ISH and RT-qPCR indicated that PMCV in some individual fish replicates extensively in the spongiform part of the heart ventricle, but less so in the other organs tested like the kidneys. To determine if the massive replication in the heart is causing a high production of new viral particles, indicating that fish with clinical symptoms are particularly important in the spread of the disease, we set out to examine what form viral RNA has in different organs.
To determine if PMCV RNA is present as dsRNA genomic RNA (gRNA) or viral mRNA in infected cells we postulated that PMCV gRNA would not be detected by RTqPCR in infected cells unless it was denatured into single strands by heating prior to reverse transcription of cDNA. This hypothesis was based on an established method designed to distinguish between the genomic dsRNA and ssRNA transcripts of the reovirus piscine orthoreovirus [21].
To test the validity of the method, we used total RNA from two heart samples with no difference in the Cq values between heating of the extracted RNA at 95°C for 5 min or no heating prior to cDNA synthesis. For such samples a major part of RNA would be expected to be ssRNA. Digestion with RNase A in the presence of 0.5 M NaCl, which selectively degrades ssRNA, versus no RNaseA digestion, gave a difference in Cq values (ΔCq) for the two test samples of 15.4 and 16.8, a reduction in the size of 1: 65 000. RNase digestion in low salt conditions completely degraded the RNA. This demonstrated that most of the RNA detected by RT-qPCR without denaturing prior to cDNA synthesis is ssRNA.
To determine if there was a difference of the relative proportion of PMCV gRNA versus mRNA between organs and between fish with or without clinical symptoms, samples from heart and kidney were tested. In the heart, PMCV ssRNA made up close to 100% of the detectable total RNA for 9 out of the 10 fish selected for a low Cq in the heart, as seen in Table 4, with an average ΔCq between heating and nonm-heating of 0.02 for those 9 fish.
The findings in the kidney were remarkably different, with an average ΔCq of 3.4. The ssRNA in the kidney made up only 9.4% of the total viral RNA, and the dominating form of PMCV RNA was genomic dsRNA which made up more than 90% of the viral RNA. Even for those individuals where approximately 100% of the detectable viral RNA in the heart was ssRNA, the genomic dsRNA was the major form in kidney, (Table 4). This indicates that the dominating viral process in the heart in the acute phase of infection, i.e. high viral RNA load, is viral mRNA transcription, while new synthesis of dsRNA, that would indicate particle formation was not detectable with the method we used. In kidney, the dsRNA was the dominating form and the massive mRNA production found in heart was not observed.
3.4. Genomic Organization of PMCV.
The detection of viral RNA by RT-qPCR using extracted, heat-denatured total RNA does not differentiate between viral genomic (gRNA) and transcribed ssRNAs. Genomic viral RNA within intact viral particles is protected against nucleases. Thus, to exclusively detect viral gRNA, cDNA synthesis should be preceded by RNase treatment, which degrades viral mRNA outside viral particles but not the RNA genomes within intact viral particles, a method used to estimate molecularly between infectious and non-infectious viral particles, assuming that intact particles are infectious [23]. Consequently, for the study of viral genomic versus mRNA we used samples that had been sampled and stored in PBS, rather than in a RNase-inhibiting-fixative. These samples were treated with RNaseA prior to RNA extraction. The RNaseA treatment degrades extra-particle viral RNA while leaving the genomic RNA within intact virus particles undamaged. The extracted viral genomic RNA was then used in RT-qPCR with primers targeting ORF2 or ORF3. The results of preceding RNase treatment before RNA extraction and RT-qPCR are shown in Figure 5. Positive control was total RNA extracted from sample without preceding RNase treatment. RT-qPCR with primers from either ORF2 or ORF3 both gave a correct sized product (264 bp and 266 bp) demonstrating that preceding RNase treatment had not destroyed the genomic RNA. When combining primers from ORF2 and ORF3, i.e., primers shown as red and blue arrows, no amplification were seen, apart from one sample (#5). This could indicate a discontinuous genome as seen in toxin producing totiviruses [9]. However, without preceding RNase treatment, i.e. containing full-length mRNA transcripts a product was amplified using the ORF2 + ORF3 primers, as seen in lane P in Figure 5C, indicating a continuous genome. A rapid amplification of cDNA ends (RACE) was run but it did not demonstrate evidence for discontinuity. In Figure 5D-F primer pairs combinations from various regions of ORF2 and ORF3 using viral genomic RNA as target demonstrate that amplification of parts of ORF3 is only partly successful (Figure 5c and f), but this is not the case when total RNA, i.e. mRNA is included as a target.
Totiviruses transcribe a full-length copy of the genome which is extruded from the cytoplasmic particle [24]. The difficulty to amplify parts of the ORF3 from genomic RNA could indicate structural problems for the reverse transcriptase used in the cDNA synthesis of the RT-qPCR, but it could also indicate that there is difference in number mRNA transcripts containing ORF3 versus the transcripts containing ORF1/2. First the efficiency and reproducibility, R2, of different sets of primers were determined, which ideally should be between 90 – 110% and higher that 0.980, respectively. The primer sets of ORF2, SP3-orf2Fw – orf2Rw, and of ORF3, orf3Fw – SP3-orf3Rw (Table 1), had estimated efficiency of 100% and 93%, respectively, and both had a reproducibility R2 of 0.99, and the sizes of these amplicons were 264 bp and 266 bp, respectively. Total RNA from heart and kidney samples from four fish was used, where there had been no preceding RNase treatment nor heat denaturing prior to the cDNA synthesis, thus the RT-qPCR mainly detected ssRNA. The relative number of ORF2 containing PMCV transcripts in heart relative to that of the kidney of the four fish were approximately 1000:1 as the ΔCq value between heart and kidney were -10 and 210 = 1024 (Table 5). The relative amounts of amplicons from ORF3 ssRNA versus ORF2 ssRNA were from 3-10% in the heart samples and from 19-40% in the kidney samples (Table 5). The difference between measured amounts of ORF2 and ORF3 could be due to difference in sensitivity between these primer sets, but this could not explain the difference between the organs. The difference in the relative amounts of transcripts with ORF3 versus ORF2 between heart and kidney was statistically significant, (p = 0.0020). The ssRNA containing ORF2 in the heart was thus, on average, 16 times more prevalent than that of the ORF3 for the samples tested, and approximately three times more prevalent in the kidney.
To test if this was reproduced with different primer/probe sets the PMCV-F/R/probe (ORF 2) and ORF3 F/R/Probe (ORF3) (Table 1) were used on 15 different heart samples, with an oobtained average Cq for ORF2 of 19.6 and for ORF3 24.3. The ΔCq of 4.7 between ORF2 and ORF3, indicated 26 times more ORF2 than ORF3 transcripts.
Together this indicates that the relative number of transcripts containing the ORF3 is lower than the ORF2 containing transcripts, especially in the heart at high transcription activity.
3.5. Are There Alternative Host Organisms for PMCV?
The indication that expression of viral transcripts containing ORF 3 is regulated might suggest that the ORF 3 could be an analogue to the toxin gene found in totiviruses of unicellular organisms where the toxin gene is found in a satellite particle. We did not find evidence of a discontinuous genome for the PMCV, but the changes of the atrium and spongious myocardium characteristic for CMS could be indications of toxic effect. Furthermore, several findings have found that PMCV does not spread efficiently from fish to fish [2,7], indicating that the capsid stability can be a limiting factor for virion survival in the external environment. This could justify a search for other host organisms than salmon. The approach we used was PCR with primers developed for amplification and subsequent sequencing and barcoding of conserved parts of the genome of various genera of organisms to assess their eventual presence. Primers are listed in Table A1. The DNA was extracted from heart and kidney samples of salmon being RT-qPCR positive for PMCV. The results were negative for all primer pairs listed in Table A1, except for the LROR/LR5 pair, which detected the large subunit rRNA of fungus in some individual fish (Figure 6). The sequence of the amplicon of approximately 1.2 kb size, seen in almost all lanes in Figure 6, matched that of Salmo salar ribosomal DNA, while the sequence of the amplicon of approximately 1.0 kb size, seen in lanes 9, 16, 18, matched that of Malassezia restricta and Debaryomyces sp. with 99.9% identity. Malassezia is a known host for totivirus [25] and Debaryomyces is a genus of yeasts in the family Saccharomycetaceae, also a known host of totiviruses [26]. This could indicate that Malassezia or Debaryomyces is present in some of the samples being positive for PMCV. This raises the question if this is an accidental finding or a contamination from the sampling process. Furthermore, one would expect that all PMCV positive samples were PCR positive for these agents if they are a mandatory host for PMCV. To study if the finding was accidental two approaches were followed. The first was PCR with primers specific for Malassezia and Debaryomyces (Table A1). This gave only negative results, indicating that the above finding was incidental. The second approach was NGS datasets obtained from tissue material collected in the present study and from CMS studies from 2010 (Data from National Center for Biotechnology Information (NCBI) Sequence Read Archive (SRA) under accession SRP002117) [6] and from NGS data from a non-CMS related salmon study. Sequences from Malassezia restricta were detected in both CMS materials collected 10 years apart, and not in the non-CMS material.
4. Discussion
The strong correlations found between viral RNA loads, location of viral RNA and severity of histopathological changes are robust indicators of PMCV's role in the etiology of CMS [5]. However, the difficulties to propagate the PMCV in cell lines has made controlled experimental infections challenging, and thus no studies have been published to date that reproduce CMS in fish using purified virus particles. Replication of PMCV has earlier been indicated in both CHH-1 and GF-1 cells [5], but these observations have not been easily reproduced. Experimental infection protocols for PMCV therefore typically use injection of material from fish that have tested positive for PMCV by RT-qPCR [7,27], i.e. experimentally PMCV infected fish are not infected through its natural route. In the current study, samples were collected in commercial farms from naturally PMCV infected fish. The nature of field sampling implies uncertainty about the time of introduction of the virus infection to the fish population, and furthermore, as horizontal transmission PMCV is observed as a slow process [2,7], one would expect large individual variation in the progress of the infection in a naturally infected fish population.
We tested fish from farms where the populations had earlier detection of PMCV by RT-qPCR. At time of sampling the fish were predominantly without clinical or pathological signs of circulatory disturbance. In most fish, PMCV RNA was found in all organs tested. For PMCV infected fish without clinical symptoms the highest viral RNA loads were found in heart, kidney, and spleen with a relatively even load of viral RNA between these organs, as indicated by Cq. However, some individual fish without clinical signs had higher viral RNA loads in heart compared to kidney/spleen with Cqs of approximately 16 versus 22-23.
When we tested fish that were assessed as having clinical signs in line with CMS at time of sampling and therefore assumed to represent the clinical phase of the disease, a significantly higher amount (p<0.01) of viral RNA was detected in heart samples compared to kidney samples, which corresponds well with the heart being the prime target organ for the virus in the acute phase, as described earlier [5]. Furthermore, the RT-qPCR results were in line with the ISH findings and also with earlier findings where PMCV was almost exclusively present in the lesions when micro-dissecting inflamed ventricular tissue [28]. It is interesting that PMCV was present in skin scrape, i.e. mucus, in some individuals that had very low level or non-detectable PMCV in heart and kidney, that could perhaps indicate transmission through water.
A natural follow up of this was to study the viral transcription activity. During the process where naked, icosahedral viruses with dsRNA genomes enter cells from the exterior, i.e. crossing the cellular membrane, they lose parts of their capsid structure and reorganize to form transcriptionally active core particles that are released into the cytoplasm. These core particles synthesize full-length, positive-sense, non-polyadenylated single-stranded mRNAs (ssRNAs) [26,29]. The viral mRNAs are identical to the positive strand of the dsRNA genome and are either translated or packaged into new particles before the synthesis of the complementary strand. Therefore, two types of viral RNA exist within the cell: dsRNA, which represents viral genomic RNA (gRNA), and ssRNA, which is synthesized and either capped to act as mRNA for viral protein translation, or incorporated into new, assembling particles, acting as a template for the viral polymerase in the minus-strand synthesis. The relative amount of viral single-stranded RNA transcripts is thus an indicator of active transcription, while double-stranded genomic RNA is an indicator of old or newly made viral particles. We validated that absence of denaturing RNA prior to cDNA synthesis used to distinguish between totiviral double stranded, genomic RNA and single stranded RNA, [21], by comparing it to an enzymatic based method, RNaseA treatment in high or low salt concentrations.
It is natural that the ratio of PMCV gRNA versus mRNA in infected cells varies through the infection process. A very high ratio of viral mRNA compared to gRNA, was observed in hearts with high viral RNA load, and in some fish the only viral RNA detected in the heart was mRNA, i.e. the gRNA was below the detection limit of the method used. In fish with low viral RNA level in heart the ratio between viral mRNA and gRNA was much lower. The relative amount of viral mRNA versus gRNA in individuals regarded as clinically affected with CMS, indicated a massive transcription activity. In an earlier study Cq values for PMCV in heart were found to be as low as 7 [2]. However, we did not find indications that the massive viral transcription in heart was reflected in production of new viral particles in the heart as the amount of dsRNA was low, which indicate lack of packing of the positive strand transcripts or lack of the subsequent synthesis of the negative strand. Both the chromogenic and fluorescent ISH clearly demonstrated that the presence of large amount of viral RNA was restricted to the spongiform part of the heart ventricle. Only very little staining was found in the stratum compactum. There is no connective tissue layer between these two muscle compartments of the heart ventricle [30]. Together this indicates that in fish with high loads of viral RNA in the heart, typically having clinical signs of CMS, the high viral RNA loads are due to viral transcription that are restricted to specific parts of the heart. We did not observe spread of virus from the spongiform layer to nearby heart compartments. The massive viral transcription in the heart was not present in the kidney and in line with this there was only punctual ISH staining found in this organ.
The pathological effects observed in the spongiform part of the heart are due to presence of viral RNA and proteins and not production of new virus particles. Degeneration and necrosis of the inner, spongious myocardium of the ventricle are characteristic histological findings, but also infiltration with inflammatory cells, and proliferation of the endocardium are observed in CMS, that indicate immune response and recovery phase [22]. The ORF 3 protein is highly cytotoxic and harmful to cells and assessed as important for the pathogenesis of CMS [31]. Toxin production is well described for totiviruses of yeast [32] where the toxin is encoded by a dsRNA segment that is encapsidated in a satellite capsid. The segmentation of the genome is increasingly found in totiviruses of various hosts such as fungi [25]. This encouraged us to study the organization of genomic RNA of PMCV. The genome of naked, icosahedral virus particles is protected against RNase activity in intact viral particles, while viral RNA outside the virion is not similarly protected [23]. Therefore, we collected field samples in PBS, i.e. without any RNase inhibitor in the sample collection medium, then treated the samples with RNase and extracted RNA. RT-qPCR was run separately for ORF2 and ORF3 and we obtained the expected amplicons, which demonstrated the presence of viral genomic RNA after the RNase treatment. When we run RT-PCR of this material with primers from ORF2 together with ORF3 primers, we did net get amplification for most samples, which could indicate that there a discontinuation of the genomic dsRNA. However, when the preceding RNase treatment was omitted, and thus the full-length transcripts were present, a product of expected size was amplified using the ORF2 + ORF3 primers. This finding did not indicate a discontinuation of the genome. Previous sequencing of PMCV and other totiviruses found in fish have not indicated segmented genomes [5,33]. Our RT-qPCR results using primer pairs combinations from various regions of ORF2 and ORF3 and viral gRNA as target could reflect that the viral RNA polymerase is able to make a continuous transcript while the similar cDNA synthesis in the RT-qPCR process is difficult.
We subsequently evaluated the relative amounts of mRNA transcripts containing ORF3 versus ORF2 in fish where the predominant form of viral RNA in the heart was identified as mRNA. Our findings revealed that transcripts containing ORF3 versus ORF2 were present in significantly lower relative quantities in the heart compared to the kidney. The higher transcription activity but less efficient transcription of ORF3 could indicate a premature termination rather than synthesis a full-length mRNA. Cytoplasmic core particles of dsRNA viruses typically synthesize full-length copies of the genome that would have led to equal amounts of ORF3 and ORF2 containing transcripts. However, our results indicate a transcript termination before the end of ORF3. This finding could represent a regulatory mechanism employed by the virus to modulate the relative amounts of viral proteins or to generate various truncated forms of the ORF3 protein. Such a mechanism might enhance the virus protein variability.
Additionally, previous unpublished work referenced in an earlier publication from expression studies of PMCV ORF3 in cell culture indicates the presence of truncated ORF3 forms [33].
We also pursued and searched for other possible natural hosts of PMCV than salmon. We used PCR and DNA extracted from organs of PMCV positive fish as target, using primers from conserved parts of the genome for various groups of organisms. These primers were originally developed for barcoding, i.e. taxonomic placement. Earlier phylogenetic studies have indicated that PMCV are most genetically related, among the classical totivirus genera, to the genus Giardiavirus [5,33], but also possibly to the totiviruses of arthropods [34]. Several of the totiviruses associated to fish are clustered to a separate genus tentatively called Pistolvirus. We tested quite broadly, with an emphasis on unicellular organisms. Using this approach, the only positive amplifications came using primers from fungi and sequencing of the amplicon showed a very high identity to the fungi Malssezia spp. and Debaryomyces spp. However, relatively few PMCV positive samples were positive by PCR for these fungi, and furthermore these fungi are almost ubiquitous even in the marine environment [35]. Using the approach described previously by our group [36], novel NGS data generated in the context of this project as well as NGS data from CMS infected fish were downloaded from public databases and mined for the presence of fungal sequences. Though sequences from Malssezia spp./Malassezia restricta could be identified even in CMS samples collected a decade apart, contamination could not be ruled out as a source.
5. Conclusions
High viral PMCV RNA loads predominantly found in the spongiform layer of the heart are attributed to extensive viral mRNA transcription. There is a correlation between PMCV viral mRNA loads, and the severity of histopathological changes observed using ISH techniques. This correlation suggests that the observed changes result from extensive viral transcription and corresponding products, rather than cellular damage caused by production of new viral particles. The production of full-length transcripts is regulated, with a reduction in the relative amount of ORF3-containing transcripts at high transcription rates. The low or non-detectable production of new viral particles in the heart together with extensive transcription activity and corresponding pathological consequences, indicates that the heart is partly a dead-end for virus production. Attempts to identify alternative hosts for PMCV, such as fungi, were inconclusive. These finding underscore the need for further research to fully understand PMCV’s life cycle and potential alternative hosts and its whereabouts when it is not infecting the heart of the Atlantic salmon.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure A1.; Table A1.
Author Contributions
ER launched the project idea and participated in the overall design and coordination of the study and interpretation of data, drafted the manuscript. IBN performed sample preparation, RT-qPCR, mRNA analysis and in situ hybridization. ØW participated in the overall design and coordination of the study and interpretation of data. HB and MA performed morphological analyses of the in vivo expression. IBN, TT, MA and ØW participated in the coordination of the study, interpretation of data, and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The Norwegian Seafood Research Fund (FHF), Grant #901671, supported this study financially.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Figure A1.
In situ hybridization of heart ventricle with fluorescent probes detecting PMCV ORF1 and ORF2. Same section. The probes detect the same cells.
Figure A1.
In situ hybridization of heart ventricle with fluorescent probes detecting PMCV ORF1 and ORF2. Same section. The probes detect the same cells.
Appendix B

Table A1.
Primers used for different classes of organisms.
| Target | Primer name | Sequence 5’- 3’ | Reference |
|---|---|---|---|
| Fungi | ITS1ITS4 (ytre) | TCCGTAGGTGAACCTGCGG TCCTCCGCTTATTGATATGC |
[37] |
| Fungi | ITS2ITS3 | GCTGCGTTCTTCCATCGATGC GCATCGATGAAGAACGCAGC |
[37] |
| Fungi | NL-1NL-4 | GCATATCAATAAGCGGAGGAAAAG GGTCCGTGTTTCAAGACGG |
[37] |
| Fungi | EF1-1018F EF1-1620R | GAYTTCATCAAGAACATGAT GACGTTGAADCCRACRTTGTC |
[37] |
| Fungi | ITS5ITS4 | GGAAGTAAAAGTCGTAACAAGG TCCTCCGCTTATTGATATGC |
[38] |
| Trypanozoma | Try-FwTry-Rw | CCAWACAACAAACATATGATGCTGC TCCHGATATGGTWTTKCCYCG |
[39] |
| Apicomplexa | Api-FwApi-Rw | GAAACTGCGAATGGCTCATT CTTGCGCCTACTAGGCATTC |
[40] |
| Mikrosporidia | CM-V5FCM-V5R | GATTAGANACCNNNGTAGTTC TAANCAGCACAMTCCACTC |
[41] |
| Mikrosporidia | bcdF01bcdR06 | CATTTTCHACTAAYCATAARGATATTGG GGDGGRTAHACAGTYCAHCCNGT |
[41] |
| Myxozoa | Myxo_617FMyxo_2313R_all | CGCGCAAATTACCCAMTCCA CGTTACCGGAATRRCCTGACAG |
[42] |
| Ichtyophonus | NS1-FNS8 deg-R | GTAGTCATATGCTTGTCTC TCCGCAGGTTCACCWACGGA |
[43] |
| Ichtyophonus | vc7Fvc5R | GTCTGTACTGGTACGGCAGTTTC TCCCGAACTCAGTAGACACTCAA |
[43] |
| Ichthyophonus | Out ITS1-FOut-ITS2-R | GCGGAAGGATCATTACCAAATAACG GCCTGAGTTGAGGTCAAATTT |
[43] |
| Ichthyophonus | Ich7fIch6r | GCTCTTAATTGAGTGTCTAC CATAAGGTGCTAATGGTGTC |
[43] |
| Malassezia | LRORLR5 | ACCCGCTGAACTTAAGC TCCTGAGGGAAACTTCG |
[44] |
| Malassezia | Mal63 Fw Mal487 Rw | TTGGCTACAGCGGCGACGACCTG CATCGCCTTGCCGACCGTCG |
[44] |
| Malassezia | LR5 - 2LROR-2 | ATCCTGAGGGAAACTTC GTACCCGCTGAACTTAAGC |
[44] |
| Malassezia | Mala_28S_F2 Mala_28S_R2 | CGCGTTGTAATCTCGAGACG CCACCCAAAAACTCGCACA |
[44] |
| Debaryomyces | Deb FwDeb Rw | TCACCATCTTTCGGGTCCCAACAGC CCCGTGCGATGAGATGCCCAATTC |
[45] |
| Oomycetea | Cox2-FCox2-R | GGCAAATGGGTTTTCAAGATCC CCATGATTAATACCACAAATTTCACTAC |
[46] |
| Crusteacea | LCO1490HCO2198 | GGTCAACAAATCATAAAGATATTGG TAAACTTCAGGGTGACCAAAAAATCA |
[47] |
References
- Brun, E.; Poppe, T.; Skrudland, A.; Jarp, J. Cardiomyopathy syndrome in farmed atlantic salmon salmo salar: Occurrence and direct financial losses for norwegian aquaculture. Dis Aquat Organ 2003, 56, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Fritsvold, C.; Mikalsen, A.B.; Poppe, T.T.; Taksdal, T.; Sindre, H. Characterization of an outbreak of cardiomyopathy syndrome (cms) in young atlantic salmon, salmo salar l. J Fish Dis 2021, 44, 2067–2082. [Google Scholar] [CrossRef]
- Amin, A.; Trasti, J. Endomyocarditis in atlantic salmon in norwegian seafarms. Bull Eur Assoc Fish Pathol 1988, 8, 70–71. [Google Scholar]
- Fritsvold, C.; Kongtorp, R.T.; Taksdal, T.; Orpetveit, I.; Heum, M.; Poppe, T.T. Experimental transmission of cardiomyopathy syndrome (cms) in atlantic salmon salmo salar. Dis Aquat Organ 2009, 87, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Haugland, O.; Mikalsen, A.B.; Nilsen, P.; Lindmo, K.; Thu, B.J.; Eliassen, T.M.; Roos, N.; Rode, M.; Evensen, O. Cardiomyopathy syndrome of atlantic salmon (salmo salar l.) is caused by a double-stranded rna virus of the totiviridae family. Journal of Virology 2011, 85, 5275–5286. [Google Scholar] [CrossRef]
- Lovoll, M.; Wiik-Nielsen, J.; Grove, S.; Wiik-Nielsen, C.R.; Kristoffersen, A.B.; Faller, R.; Poppe, T.; Jung, J.; Pedamallu, C.S.; Nederbragt, A.J. , et al. A novel totivirus and piscine reovirus (prv) in atlantic salmon (salmo salar) with cardiomyopathy syndrome (cms). Virol J 2010, 7, 309. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; van Eerde, A.; Steen, H.S.; Heldal, I.; Haugslien, S.; Ørpetveit, I.; Wüstner, S.C.; Inami, M.; Løvoll, M.; Rimstad, E. Establishment of a piscine myocarditis virus (pmcv) challenge model and testing of a plant-produced subunit vaccine candidate against cardiomyopathy syndrome (cms) in atlantic salmon salmo salar. Aquaculture 2021, 541, 736806. [Google Scholar] [CrossRef]
- Dinman, J.D.; Icho, T.; Wickner, R.B. A -1 ribosomal frameshift in a double-stranded rna virus of yeast forms a gag-pol fusion protein. Proc Natl Acad Sci U S A 1991, 88, 174–178. [Google Scholar] [CrossRef]
- Rodríguez-Cousiño, N.; Esteban, R. Relationships and evolution of double-stranded rna totiviruses of yeasts inferred from analysis of la-2 and l-bc variants in wine yeast strain populations. Applied and Environmental Microbiology 2017, 83, e02991–02916. [Google Scholar] [CrossRef]
- Fermin, G.; Mazumdar-Leighton, S.; Tennant, P. Viruses of prokaryotes, protozoa, fungi, and chromista. Viruses, Academic Press, London 2018, 217-244.
- Wickner, R.B.; Ribas, J.C. Totivirus. In The springer index of viruses, 2011.
- Tang, J.; Ochoa, W.F.; Sinkovits, R.S.; Poulos, B.T.; Ghabrial, S.A.; Lightner, D.V.; Baker, T.S.; Nibert, M.L. Infectious myonecrosis virus has a totivirus-like, 120-subunit capsid, but with fiber complexes at the fivefold axes. Proceedings of the National Academy of Sciences 2008, 105, 17526–17531. [Google Scholar] [CrossRef]
- Shao, Q.; Jia, X.; Gao, Y.; Liu, Z.; Zhang, H.; Tan, Q.; Zhang, X.; Zhou, H.; Li, Y.; Wu, D. Cryo-em reveals a previously unrecognized structural protein of a dsrna virus implicated in its extracellular transmission. PLoS pathogens 2021, 17, e1009396. [Google Scholar] [CrossRef] [PubMed]
- de Lima, J.G.; Lanza, D.C. 2a and 2a-like sequences: Distribution in different virus species and applications in biotechnology. Viruses 2021, 13, 2160. [Google Scholar] [CrossRef] [PubMed]
- Dantas, M.D.A.; Cavalcante, G.H.O.; Oliveira, R.A.; Lanza, D.C. New insights about orf1 coding regions support the proposition of a new genus comprising arthropod viruses in the family totiviridae. Virus research 2016, 211, 159–164. [Google Scholar] [CrossRef]
- Poulos, B.T.; Tang, K.F.; Pantoja, C.R.; Bonami, J.R.; Lightner, D.V. Purification and characterization of infectious myonecrosis virus of penaeid shrimp. Journal of General Virology 2006, 87, 987–996. [Google Scholar] [CrossRef]
- Wang, H.; Marucci, G.; Munke, A.; Hassan, M.M.; Lalle, M.; Okamoto, K. High-resolution comparative atomic structures of two giardiavirus prototypes infecting g. Duodenalis parasite. PLoS Pathog 2024, 20, e1012140. [Google Scholar] [CrossRef]
- Wessel, O.; Olsen, C.M.; Rimstad, E.; Dahle, M.K. Piscine orthoreovirus (prv) replicates in atlantic salmon (salmo salar l.) erythrocytes ex vivo. Vet Res 2015, 46, 26. [Google Scholar] [CrossRef] [PubMed]
- Hodneland, K.; Endresen, C. Sensitive and specific detection of salmonid alphavirus using real-time pcr (taqman). J Virol Methods 2006, 131, 184–192. [Google Scholar] [CrossRef]
- Park, C.; Raines, R.T. Origin of the 'inactivation' of ribonuclease a at low salt concentration. FEBS Lett 2000, 468, 199–202. [Google Scholar] [CrossRef]
- Polinski, M.P.; Marty, G.D.; Snyman, H.N.; Garver, K.A. Piscine orthoreovirus demonstrates high infectivity but low virulence in atlantic salmon of pacific canada. Sci Rep 2019, 9, 3297. [Google Scholar] [CrossRef]
- Ferguson, H.; Poppe, T.; Speare, D.J. Cardiomyopathy in farmed norwegian salmon. Diseases of Aquatic Organisms 1990, 8, 225–231. [Google Scholar] [CrossRef]
- Marti, E.; Ferrary-Américo, M.; Barardi, C.R.M. Detection of potential infectious enteric viruses in fresh produce by (rt)-qpcr preceded by nuclease treatment. Food Environ Virol 2017, 9, 444–452. [Google Scholar] [CrossRef]
- Wickner, R.B. Double-stranded rna viruses of saccharomyces cerevisiae. Microbiological reviews 1996, 60, 250–265. [Google Scholar] [CrossRef]
- Applen Clancey, S.; Ruchti, F.; LeibundGut-Landmann, S.; Heitman, J.; Ianiri, G. A novel mycovirus evokes transcriptional rewiring in the fungus malassezia and stimulates beta interferon production in macrophages. mBio 2020, 11. [Google Scholar] [CrossRef]
- Wickner, R.B.; Tang, J.; Gardner, N.A.; Johnson, J.E.; Patton, J. The yeast dsrna virus la resembles mammalian dsrna virus cores. Caister Academic Press: 2008.
- Timmerhaus, G.; Krasnov, A.; Nilsen, P.; Alarcon, M.; Afanasyev, S.; Rode, M.; Takle, H.; Jørgensen, S.M. Transcriptome profiling of immune responses to cardiomyopathy syndrome (cms) in atlantic salmon. BMC Genomics 2011, 12, 459. [Google Scholar] [CrossRef]
- Wiik-Nielsen, J.; Løvoll, M.; Fritsvold, C.; Kristoffersen, A.B.; Haugland, Ø.; Hordvik, I.; Aamelfot, M.; Jirillo, E.; Koppang, E.O.; Grove, S. Characterization of myocardial lesions associated with cardiomyopathy syndrome in a tlantic salmon, s almo salar l., using laser capture microdissection. Journal of Fish Diseases 2012, 35, 907–916. [Google Scholar] [CrossRef]
- Tenorio, R.; Fernández de Castro, I.; Knowlton, J.J.; Zamora, P.F.; Sutherland, D.M.; Risco, C.; Dermody, T.S. Function, architecture, and biogenesis of reovirus replication neoorganelles. Viruses 2019, 11. [Google Scholar] [CrossRef]
- Pieperhoff, S.; Bennett, W.; Farrell, A.P. The intercellular organization of the two muscular systems in the adult salmonid heart, the compact and the spongy myocardium. J Anat 2009, 215, 536–547. [Google Scholar] [CrossRef]
- Garseth, A.H.; Fritsvold, C.; Svendsen, J.C.; Jensen, B.B.; Mikalsen, A.B. Cardiomyopathy syndrome in atlantic salmon salmo salar l.: A review of the current state of knowledge. Journal of Fish Diseases 2018, 41, 11–26. [Google Scholar] [CrossRef]
- Bevan, E.; Herring, A.; Mitchell, D.J. Preliminary characterization of two species of dsrna in yeast and their relationship to the “killer” character. Nature 1973, 245, 81–86. [Google Scholar] [CrossRef]
- Sandlund, L.; Mor, S.K.; Singh, V.K.; Padhi, S.K.; Phelps, N.B.D.; Nylund, S.; Mikalsen, A.B. Comparative molecular characterization of novel and known piscine toti-like viruses. Viruses 2021, 13. [Google Scholar] [CrossRef]
- Tighe, A.J.; Ruane, N.M.; Carlsson, J. Potential origins of fish toti-like viruses in invertebrates. Journal of General Virology 2022, 103, 001775. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, R.M.; El Baidouri, F.; Mitchison-Field, L.M.; Lim, F.Y.; Ekena, J.; Vogt, E.J.; Gladfelter, A.; Theberge, A.B.; Amend, A.S. Malassezia is widespread and has undescribed diversity in the marine environment. Fungal Ecology 2023, 65, 101273. [Google Scholar] [CrossRef]
- Tengs, T.; Rimstad, E. Emerging pathogens in the fish farming industry and sequencing-based pathogen discovery. Dev Comp Immunol 2017, 75, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Uyaguari-Diaz, M.I.; Chan, M.; Chaban, B.L.; Croxen, M.A.; Finke, J.F.; Hill, J.E.; Peabody, M.A.; Van Rossum, T.; Suttle, C.A.; Brinkman, F.S. A comprehensive method for amplicon-based and metagenomic characterization of viruses, bacteria, and eukaryotes in freshwater samples. Microbiome 2016, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Bal, J.; Yun, S.-H.; Yeo, S.-H.; Kim, J.-M.; Kim, D.-H. Metagenomic analysis of fungal diversity in korean traditional wheat-based fermentation starter nuruk. Food Microbiology 2016, 60, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.S.; Morelli, K.A.; Jansen, A.M. Cytochrome c oxidase subunit 1 gene as a DNA barcode for discriminating trypanosoma cruzi dtus and closely related species. Parasites & vectors 2017, 10, 1–18. [Google Scholar]
- Renoux, L.P.; Dolan, M.C.; Cook, C.A.; Smit, N.J.; Sikkel, P.C. Developing an apicomplexan DNA barcoding system to detect blood parasites of small coral reef fishes. Journal of Parasitology 2017, 103, 366–376. [Google Scholar] [CrossRef]
- Trzebny, A.; Slodkowicz-Kowalska, A.; Becnel, J.J.; Sanscrainte, N.; Dabert, M. A new method of metabarcoding microsporidia and their hosts reveals high levels of microsporidian infections in mosquitoes (culicidae). Molecular Ecology Resources 2020, 20, 1486–1504. [Google Scholar] [CrossRef]
- Hartikainen, H.; Bass, D.; Briscoe, A.G.; Knipe, H.; Green, A.J.; Okamura, B. Assessing myxozoan presence and diversity using environmental DNA. International Journal for Parasitology 2016, 46, 781–792. [Google Scholar] [CrossRef]
- LaDouceur, E.E.; Leger, J.S.; Mena, A.; Mackenzie, A.; Gregg, J.; Purcell, M.; Batts, W.; Hershberger, P. Ichthyophonus sp. Infection in opaleye (girella nigricans). Veterinary pathology 2020, 57, 316–320. [Google Scholar] [CrossRef]
- Clancey, S.A.; Ruchti, F.; LeibundGut-Landmann, S.; Heitman, J.; Ianiri, G. A novel mycovirus evokes transcriptional rewiring in malassezia and provokes host inflammation and an immunological response. bioRxiv [Preprint].
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; De Vries, M.; Verkleij, G.; Crous, P.; Boekhout, T. DNA barcoding analysis of more than 9 000 yeast isolates contributes to quantitative thresholds for yeast species and genera delimitation. Studies in mycology 2016, 85, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Robideau, G.P.; De Cock, A.W.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Desaulniers, N.; Eggertson, Q.A.; Gachon, C.M. DNA barcoding of oomycetes with cytochrome c oxidase subunit i and internal transcribed spacer. Molecular ecology resources 2011, 11, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Raupach, M.J.; Barco, A.; Steinke, D.; Beermann, J.; Laakmann, S.; Mohrbeck, I.; Neumann, H.; Kihara, T.C.; Pointner, K.; Radulovici, A. The application of DNA barcodes for the identification of marine crustaceans from the north sea and adjacent regions. PloS one 2015, 10, e0139421. [Google Scholar] [CrossRef] [PubMed]
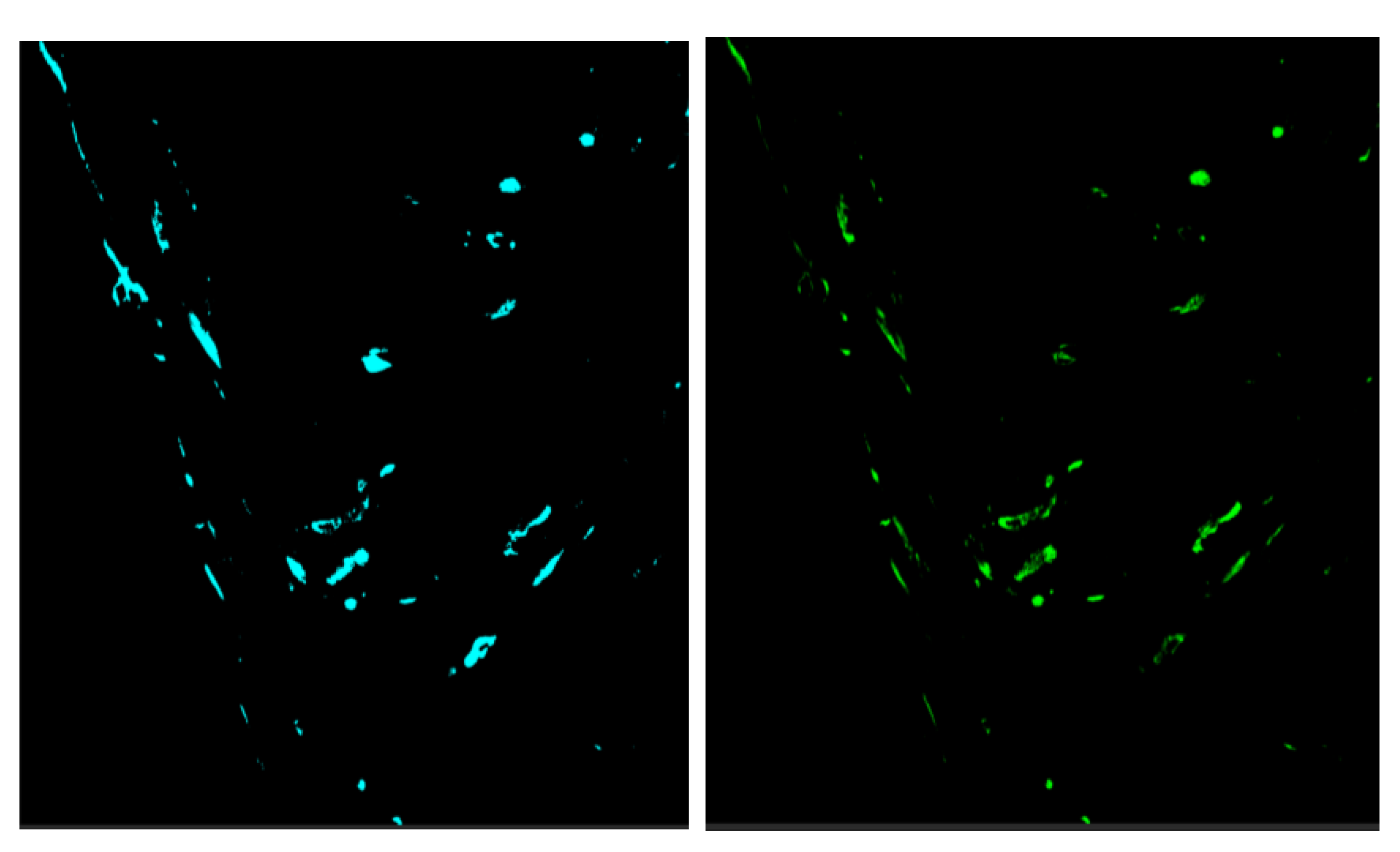
Figure 1.
In situ hybridization of heart for PMCV with A) chromogenic (red) and B-D) fluorescent staining (green). A) The chromogenic staining is restricted to the stratum spongiosum with only a few stained spots (arrow) in compactum. The sample used had a Cq= 13, i.e indicating a high viral load. 10 X objective. B) Fluorescent in situ hybridization. Transverse section of the ventricle. Staining is restricted to stratum spongiosum. 10 X objective. C) Staining with DAPI (C1) where blue dots represent nucleated hematocytes including red blood cells and fluorescent in situ hybridization of stratum spongiosum against PMCV (C2). The same section is shown in C1 and C2, and no PMCV staining of heamatocytes is seen. 20 X objective. D) Kidney, scattered staining for PMCV. Counterstained with DAPI. Black spots represent melano-macrophages. Arrow points to staining in tubular structure. S = spongiosum, C = Compactum.
Figure 1.
In situ hybridization of heart for PMCV with A) chromogenic (red) and B-D) fluorescent staining (green). A) The chromogenic staining is restricted to the stratum spongiosum with only a few stained spots (arrow) in compactum. The sample used had a Cq= 13, i.e indicating a high viral load. 10 X objective. B) Fluorescent in situ hybridization. Transverse section of the ventricle. Staining is restricted to stratum spongiosum. 10 X objective. C) Staining with DAPI (C1) where blue dots represent nucleated hematocytes including red blood cells and fluorescent in situ hybridization of stratum spongiosum against PMCV (C2). The same section is shown in C1 and C2, and no PMCV staining of heamatocytes is seen. 20 X objective. D) Kidney, scattered staining for PMCV. Counterstained with DAPI. Black spots represent melano-macrophages. Arrow points to staining in tubular structure. S = spongiosum, C = Compactum.
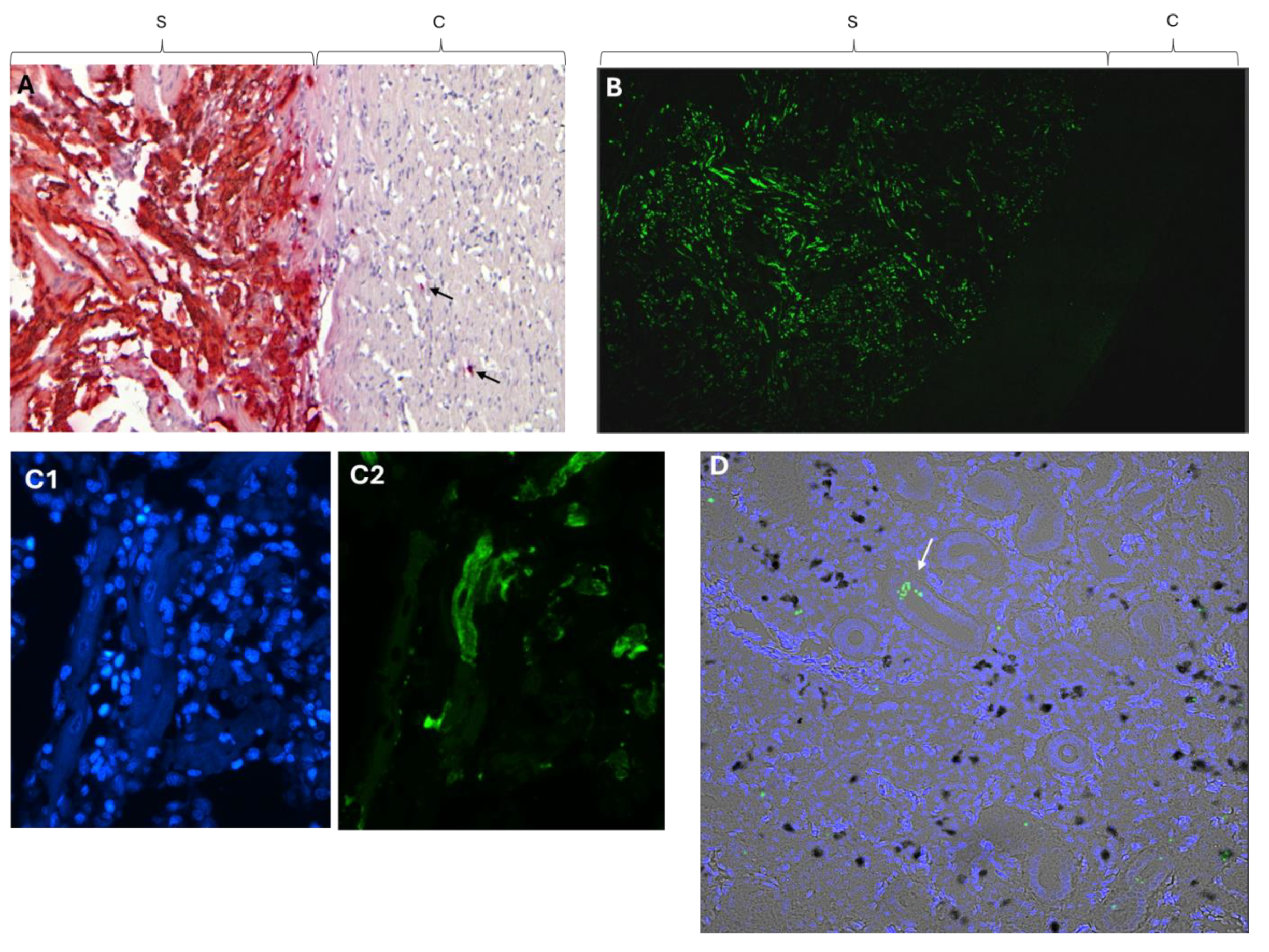
Figure 5.
Top: Location of primers used on extracted viral genomic RNA from intact virus particles.RT-qPCR of genomic RNA of viral particles, i.e from samples RNase treated prior to RNA extraction. Primer sequences came from ORF2, ORF3 and color-coded arrows indicate positions and combinations. M = molecular marker, P = positive control, i.e. total RNA extracted from non-RNase treated material. N = negative control.
Figure 5.
Top: Location of primers used on extracted viral genomic RNA from intact virus particles.RT-qPCR of genomic RNA of viral particles, i.e from samples RNase treated prior to RNA extraction. Primer sequences came from ORF2, ORF3 and color-coded arrows indicate positions and combinations. M = molecular marker, P = positive control, i.e. total RNA extracted from non-RNase treated material. N = negative control.
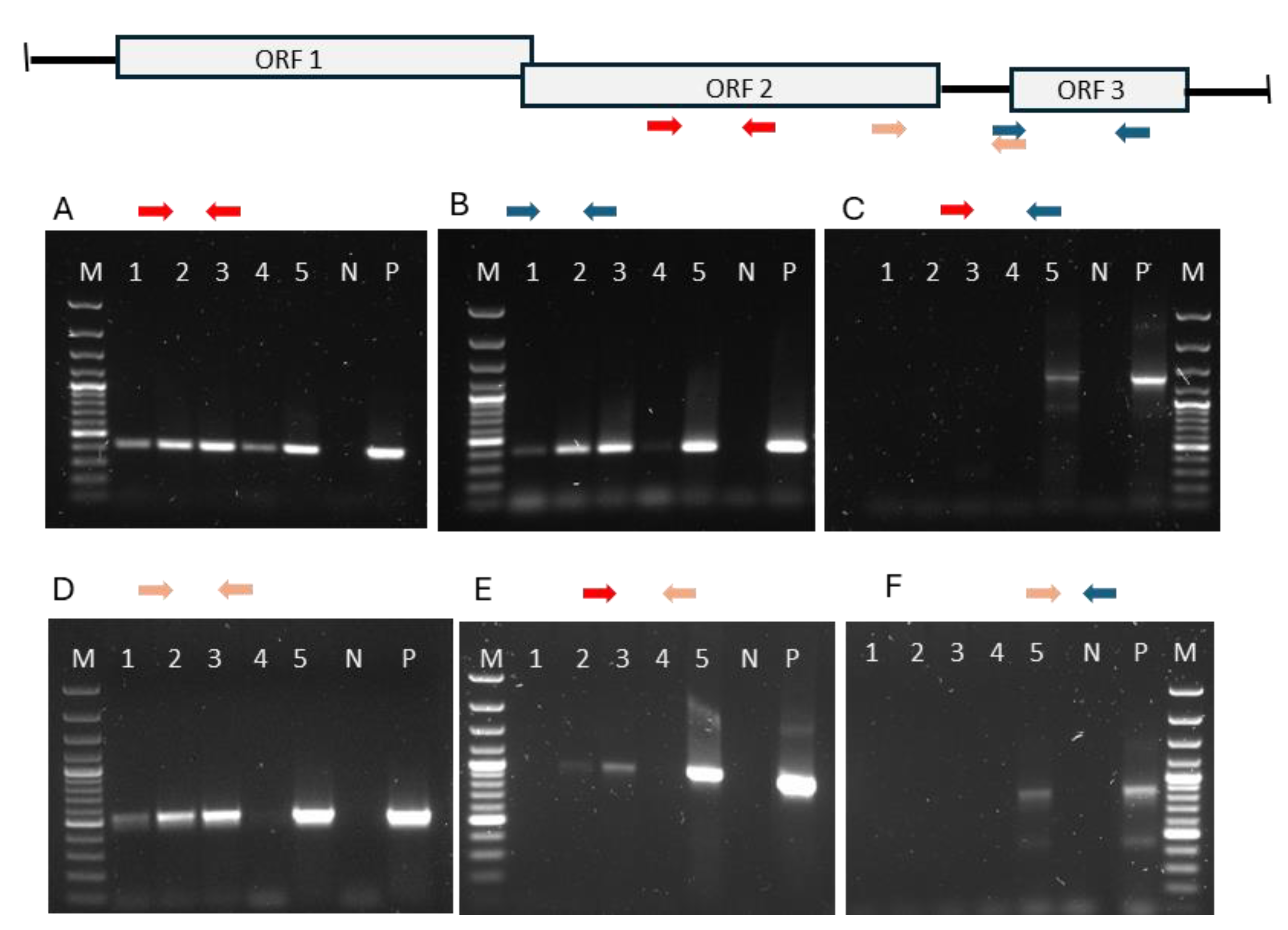
Figure 7.
PCR amplification using primers developed for detection of conserved part of ribosomal DNA of the fungus Malassezia. The sequence of the larger corresponds to ribosomal DNA of Atlantic salmon, while the sequence of the bands of lanes 9, 16 and 18 matched Malassezia sp.
Figure 7.
PCR amplification using primers developed for detection of conserved part of ribosomal DNA of the fungus Malassezia. The sequence of the larger corresponds to ribosomal DNA of Atlantic salmon, while the sequence of the bands of lanes 9, 16 and 18 matched Malassezia sp.
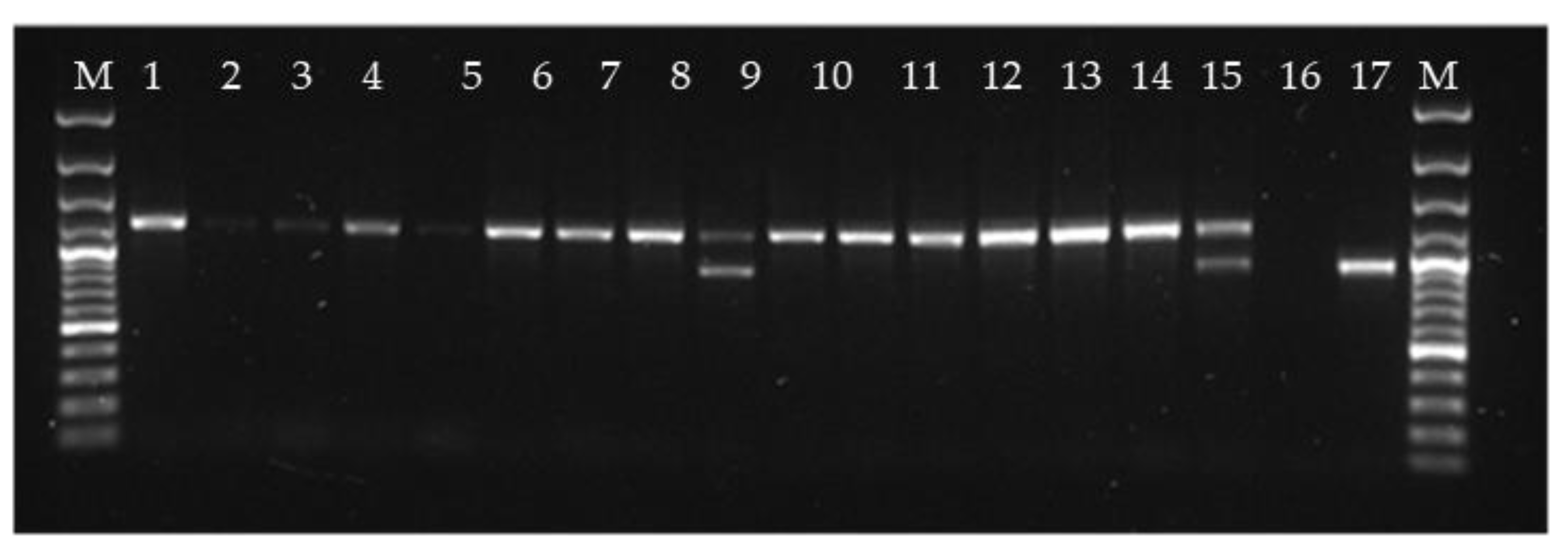
Table 1.
Primers and probes used for detection of viruses.
| Virus | Primer/Probe | Sequence (5’ → 3’) | Amplicon (bp) | Reference |
|---|---|---|---|---|
| PMCV | ||||
| ORF2 | PMCV-F | TTCCAAACAATTCGAGAAGCG | 140 | [6] |
| PMCV-R | ACCTGCCATTTTCCCCTCTT | |||
| PMCV probe | FAM-CCGGGTAAAGTATTTGCGTC- MGBNFQ | |||
| ORF3 | Orf3-3’Fw | TTACAGAGGGCGGGAACCTGTGTGG | ||
| Orf3-3’Rw | TGGCTTCTTGTGAATTGTCAACAC | 114 | ||
| Orf3 probe | FAM -TCTTCGATAATACGCAGTGTA- MGBNFQ | |||
| SYBR primers | SP3-orf2Fw | CTAAGGCCAGTGGCGGAATC | 264 | |
| Orf2 Rw | TGGTGGCATACTTACCCATG | |||
| Orf3 Fw | GGCGAGAATGGTGTTTGTGCACTGC | 266 | ||
| SP3-orf3Rw | GAATGAAGCAAGATGGAACC | |||
| Orf2-3’Fw | TTGGGTTCAAGAGGATAGAG | 122 | ||
| Orf2-3’Rw | GAATTTTGGTACCTGTGATG | |||
| PRV 1 | Sigma3 659 Fw | TGCGTCCTGCGTATGGCACC | [18] | |
| Sigma3 801Rw | GGCTGGCATGCCCGAATAGCA | |||
| Sigma3_693 probe | FAM-ATCACAACGCCTACCT–MGBNFQ | |||
| SAV | QnsP1 17F | CCGGCCCTGAACCAGTT | [19] | |
| QnsP1 122R | GTAGCCAAGTGGGAGAAAGCT | |||
| QnsP1_53probe | FAM-CTGGCCACCACTTCGA-MGBNFQ |
Table 2.
Target and control probes for in situ hybridization.
| Probe | Accession no. | Target region (bp) | Catalogue no. | |
|---|---|---|---|---|
| V-Piscine-myocarditis-ORF1-C2) | JQ728724.1 | 1050-1757 | 812021-C2 | |
| V-PMCV-ORF2 | HQ339954.1 | 3441 - 4500 | 555231 | |
| V-PMCV-ORF2-sense-C3 | HQ339954.1 | 3441 - 4500 | 1219761-C3 | |
| DapB (negative control) | EF191515 | 414 - 862 | 310043 | |
| PPIB (positive control) | NM_001140870 | 20 - 934 | 494421 |
Table 3.
Cq values of RT-qPCR for PMCV of various organs of clinically healthy, PMCV infected population of Atlantic salmon.
Table 3.
Cq values of RT-qPCR for PMCV of various organs of clinically healthy, PMCV infected population of Atlantic salmon.
| Fish no. | Heart | Gills | Kidney | Spleen | Liver | Muscle | Skin scrape | Pyloric ceca | Mid-gut | Hind-gut |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | - | - | - | - | - | - | 35.9 | - | - | - |
| 2 | 24.5 | 26.0 | 22.9 | 21.9 | 29.9 | 27.4 | 26.4 | 27.5 | 25.7 | 27.5 |
| 3 | - | - | - | - | - | - | - | - | - | - |
| 4 | 16.4 | 28.6 | 22.5 | 22.7 | 26.7 | 26.8 | 28.4 | 26.3 | 25.3 | 26.3 |
| 5 | 36.9 | - | 35.0 | - | - | - | 37.4 | - | - | - |
| 6 | - | - | - | - | - | - | - | - | - | - |
| 7 | 24.4 | 26.4 | 22.9 | 22.9 | 29.5 | 28.1 | 29.9 | 27.4 | 22.2 | 27.1 |
| 8 | 22.9 | 28.8 | 21.9 | 21.8 | 27.1 | 27.8 | 29.6 | 29.6 | 29.1 | 30.5 |
| 9 | 16.9 | 28.0 | 24.5 | 24.9 | 30.5 | 28.4 | 28.7 | 28.9 | 28.3 | 27.7 |
| 10 | 26.2 | 26.3 | 23.9 | 25.1 | 30.8 | 28.0 | 27.3 | 29.5 | 26.5 | 27.6 |
| Ave | 21.9 | 27.3 | 23.1 | 23.2 | 29.1 | 27.8 | 28.4 | 28.2 | 26.2 | 27.8 |
Table 4.
Relative amount of viral ssRNA and dsRNA in heart and kidney.
| Heat treated (ssRNA + dsRNA) |
Not heat treated (ssRNA) |
ΔCq for PMCV ssRNA versus total PMCV RNA |
||||
|---|---|---|---|---|---|---|
| Fish# | Heart | Kidney | Heart | Kidney | Heart | Kidney |
| 1 | 17.05 | 21.66 | 17.23 | 25.47 | 0.18 | 3.81 |
| 2 | 13.36 | 20.20 | 13.32 | 23.06 | -0.04 | 2.86 |
| 3 | 14.19 | 20.26 | 14.49 | 24.24 | 0.3 | 3.94 |
| 4 | 12.79 | 21.74 | 12.78 | 25.27 | -0.01 | 3.53 |
| 5 | 17.70 | 19.00 | 18.65 | 23.21 | 0.95 | 4.21 |
| 6 | 21.8 | 23.5 | 21.6 | 24.5 | -0.2 | 1 |
| 7 | 17.7 | 20.6 | 17.6 | 24.2 | -0.1 | 3.6 |
| 8 | 13.6 | 19.3 | 13.7 | 22.9 | 0.1 | 3.6 |
| 9 | 17.4 | 22.7 | 17.3 | 25.9 | -0.1 | 3.2 |
| 10 | 17.5 | 20.4 | 17.6 | 24.5 | 0.1 | 4.1 |
Table 5.
Relative amount of ORF3 ssRNA versus ORF2.
| ORF 2 | ORF 3 | ΔCt | Relative amount of ORF3 ssRNA versus ORF2 | |
|---|---|---|---|---|
| Heart 2 | 12.08 | 15.36 | -3.28 | 10% |
| Heart 3 | 12.97 | 17.94 | -4.97 | 3% |
| Heart 4 | 11.34 | 16.60 | -5.26 | 3% |
| Heart 5 | 18.31 | 21.72 | -3.41 | 9% |
| Average | 13.675 | 17.90 | 6.25% | |
| Kidney 2 | 22.91 | 24.23 | -1.32 | 40% |
| Kidney 3 | 23.84 | 25.43 | -1.59 | 33% |
| Kidney 4 | 24.68 | 27.10 | -2.42 | 19% |
| Kidney 5 | 23.13 | 24.71 | -1.58 | 33% |
| Average | 23.64 | 25.37 | 31.25% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



