Submitted:
03 July 2024
Posted:
04 July 2024
You are already at the latest version
Abstract
Extracellular vesicles represent a large heterogeneous class of near and long-distance intercellular communication mediators, released by both prokaryotic and eukaryotic cells. Specifically, the scientific community has shown a growing interest in exosomes, which are nano-sized vesicles with an endosomal origin. Not so long ago, the physiological goal of exosome generation was largely unknown and required more investigation; at first, it was hypothesized that exosomes are able to remove excess, rejects and unnecessary constituents from cells to preserve cellular homeostasis. However, thanks to recent studies, the central role of exosomes in regulating cellular dialogue has emerged. Exosomes act as vectors on cell-cell signaling by their cargo, proteins, lipids, and nucleic acids, and influence physiological and pathological processes. The findings on exosomes are widespread in a large spectrum of biomedical applications from diagnosis and prognosis to thera-pies. In this review, we describe exosome biogenesis and the current methods for their isolation and characterization, emphasizing the role of their cargo in female reproductive processes, from gametogenesis to implantation, and the potential involvement in human female disorders.
Keywords:
extracellular vesicles
; exosomes
; implantation
; embryo
; gametogenesis
; germ cells
; microRNAs
; biomarkers
; blastocoel fluid
1. Introduction
Intercellular communication is mediated by an extensive variety of signaling processes and it is indispensable for the maintenance of homeostasis and the safety of the whole organism [1,2]. Traditionally, cell-to-cell communication involved direct interactions (gap junctions), autocrine, paracrine and endocrine mechanisms as well signaling molecules targeting cellular receptors. Over the last few years, new mediators of intercellular communication in both Prokaryotic and Eukaryotic domains [3] have emerged: the extracellular vesicles (EVs). EVs have become a central hub in cellular biology acting as mediators among cells not only in physiological but also in pathological conditions [4]. EVs include a heterogeneous class of vesicles delimited by a phospholipid bilayer [5]. They are constitutively released by numerous cell types in response to specific stimuli or cellular stress factors [4,6] and transfer their contents to the target cells. Currently, several subgroups of EVs have been defined, such as small EVs, large EVs, ectosomes, extracellular particles, and exosome-like vesicles [3]. However, due to the difficulties in isolation methods and the different classification criteria, three major groups of vesicles are essentially considered. Based on their size, density, morphology, biogenesis and surface markers, they can be classified in microvesicles (MVs), apoptotic bodies and exosomes. MVs, also known as microparticles, are released by budding or shedding of the plasma membrane [7] and show a diameter > 100 nm. The apoptotic bodies, with the broadest range of diameters (50–5000 nm), are produced by cells undergoing apoptosis [8]. Exosomes, nano-sized vesicles (from 30 to 100 nm in diameter) show a characteristic cup-shaped morphology bounded by a bilayer membrane and are secreted by the most of cells both in physiological and pathological conditions [9]. The biogenesis of exosomes mainly goes across three different stages. First the cytoplasmic membrane invaginates to form endosomes, which in turn fuse to form early endosomes (ESEs). Then, ESEs invaginate again to encapsulate the intracellular material, forming multiple small intraluminal vesicles that are further transformed into late endosomes (LSE) which later become multivesicular bodies (MVBs). Finally, MVBs fuse with the cytoplasmic membrane, releasing intraluminal vesicles (ILVs) into the extracellular environment [10,11]. Nowadays, accumulating evidence suggest that these secretory vesicles are released by all types of eukaryotic cells and are widely available in almost all bodily fluids, including urine, blood, cerebrospinal fluid, saliva, breast milk, seminal fluid and follicular fluid [12,13]. Precisely, they can function as intercellular transmitters to deliver their cargo molecules necessary for cell-to-cell communication via interaction on the cells surface, taking part in many biological processes [2]. Their intercellular trafficking includes proteins (such as annexins, tetraspanin, integrin family, proteoglycans, lectins, proteins involved in apoptosis) [14], lipids (such as cholesterol, diglycerides, sphingolipids, phospholipids, prostaglandins, leukotrienes) [15] and nucleic acids (DNA, mRNAs and non-codingRNAs) [16].The term exosome was coined for the first time by Johnstone et al. in 1987 [17], representing the smallest unit of EVs which are delivered to the extracellular space [18]. Exosomes are usually hemispherical, circular, and elliptical cups with a concave side [19]. Despite notable progress, the characterization and quantification of exosomes remain considerable due to the complexity and lack of standard isolation methods [20,21]. Today, a set of standardized methods for exosome isolation and characterization have been widely accepted. In order to ensure reproducibility, comparability across the different studies and define stringent classification parameters, the International Society for Extracellular Vesicles (ISEV) proposed the Minimal Information for Studies of Extracellular Vesicles (MISEV) guidelines for the field in 2018. The MISEV 2018 guidelines include tables and outlines of suggested protocols and steps to follow to document specific EV-associated functional activities [22,23]. The most commonly used methods in exosome isolation are ultrafast-centrifugation, density gradients, filtration (by using filters with pore diameters of 0.8, 0.45, 0.22, and 0.1 μm) [24] and exosome extraction kits based on polyethylene glycol (PEG) precipitation [24]. While scanning and trasmission electron microscopy (SEM and TEM) have been used to observe cup shape, confocal microscopy is employed for live imaging of exosomes uptake and their intracellular mobility [25]. Nanoparticle tracking analysis (NTA) and dynamic light scattering (DLS) are also used to measure particle distribution, concentration and sizes in the range of 30–150 nm [26]. Western blot is commonly applied to detect the presence of molecular markers inside and outside exosomes. Nowadays, the international standard is the detection of three positive proteins (such as transmembrane Tetraspanins CD63, CD9 and/ or CD81, soluble protein TSG101 and cytoplasmic protein Alix) and one negative protein (endoplasmic reticulum protein Calnexin, nuclear proteins histone and Golgi protein GM130) [23,27,28]. In 2019, Zhang and Lyden optimized asymmetric-flowfield-flow fractionation (AF4) technology for separating and characterizing extracellular nanoparticles, particularly exosomes [29]. However, one of the main limitations of AF4 is that it fractionates samples based on their size, thus particles with the same hydrodynamic size but with different morphologies, surface molecules and other biophysical properties can not be separated from each other via AF4 alone [29]. In 2021, a paper published in International Journal of Nanomedicine [21], proposed the silicon nanowires (Si NW) label-free luminescent sensing system. It is functionalized with a specific antibody able to selectively bind the vesicles with CD81+ antigen. Tested with follicular fluid and blastocoel samples, Si NW label-free luminescent sensors showed high selectivity and sensitivity, outperforming standard mass approaches like ELISA. The functionalization processes used for these sensors make them suitable for commercial applications [21]. Initially, exosomes were underestimated as vesicles for cellular waste disposal; however, they are now recognized as crucial contributors in intercellular communication [30,31]. Exosomes are able to release their contents into the cytosol of acceptor cells by using their surface molecules’ receptor-ligand interaction, endocytosis, or direct fusion with the targeting cell membrane [32]. Their bioactive cargoes, uptaken by recipient cells through different mechanisms, can influence molecular pathways and cellular function [33,34,35]. In, 2007 Valadi et al., demonstrated for the first time that, in addition of DNA, mRNAs, proteins and lipids, exosomes can transport non-coding RNAs (ncRNAs): in particular, the authors revealed the presence of microRNAs (miRNAs) in exosomes cargo, which could be delivered to other cells, influencing their functions and gene expression. This represents a novel mechanism of genetic exchange between cells, that they proposed to call “exosomal shuttle RNA” (esRNA) [36]. Research is ongoing to better understand the sorting mechanisms and practical impacts of ncRNAs in exosomes: technological advancements in high-throughput sequencing and bioinformatic analyses are aiding in the detailed characterization of exosomal ncRNAs, emphasizing the complexity of intercellular communication and their potential for innovative diagnostic and therapeutic strategies [37]. These discoveries suggest that the investigation on circulating exosomes and their cargo could provide new opportunities for liquid biopsy in precision medicine, highlighting the potential role of exosomes as biomarkers for cancer diagnosis and prognosis prediction [38,39,37]. Exosomes showed greater advantages in liquid biopsy: in most cases, their abundance in biofluids could contribute to easier isolation of vesicles [35]; exosomes are secreted by living cells and contain several biological information from their origin cells [33,35]; they are innately stable thanks to their lipid bilayers, and thus stably circulate in both physiological conditions and in the harsh tumoral microenvironment [38]. However, one of the most significant limits for the application of exosomes in liquid biopsy is isolation with high efficiency and purity, due to their nanoscale size and intrinsic heterogeneity [40,41,42]. Ultrasensitive and precise detection is necessary for the development of exosome-based cancer diagnostics, since they represent only a small part of all exosomes in bodily fluids [38]. The great predictive potential of exosomal cargo finds application not only in cancer but also in reproductive medicine, pointing out the possibility of using the molecules transported inside exosomes as possible markers of gametes, embryo and implantation quality [43].
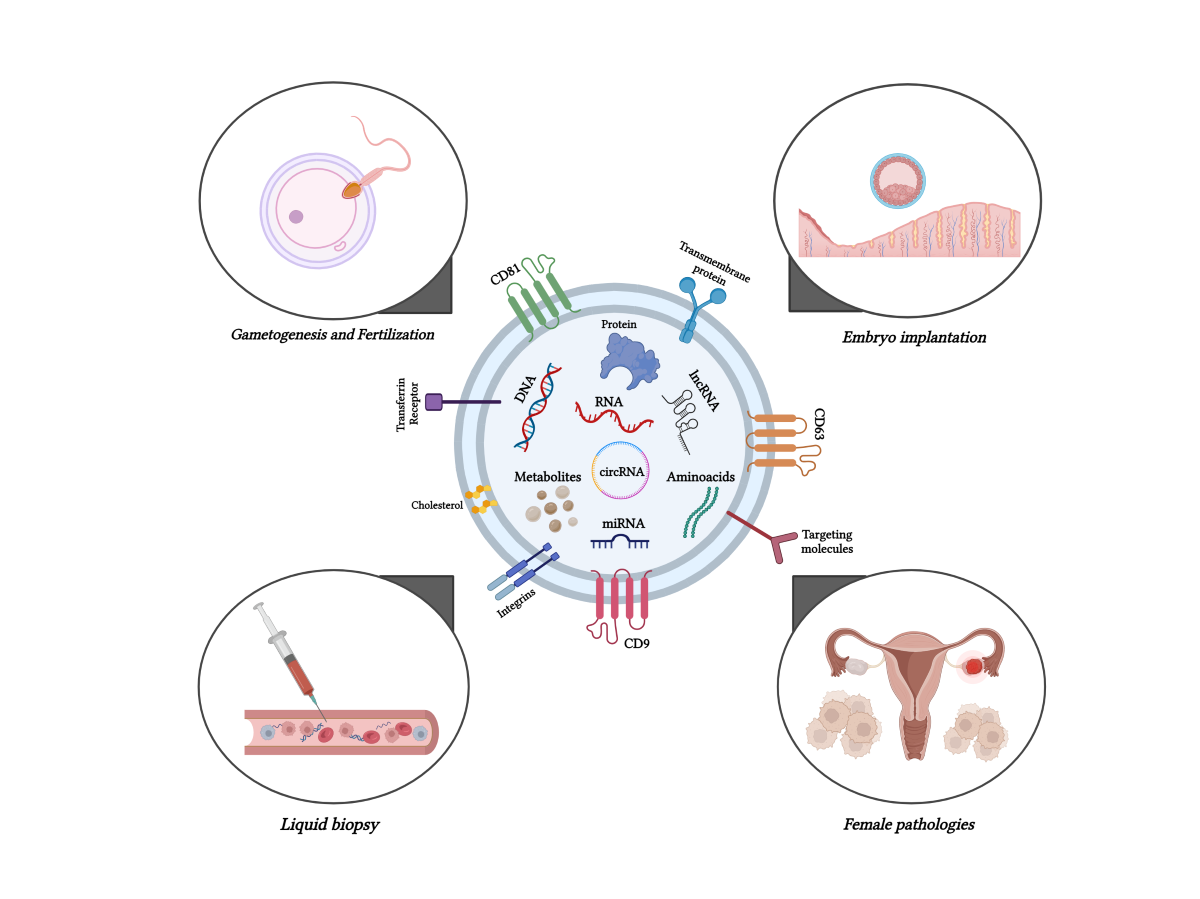
Figure 1.
Graphical abstract edited by Biorender https://www.biorender.com/ (accessed on 30 June 2024).
Figure 1.
Graphical abstract edited by Biorender https://www.biorender.com/ (accessed on 30 June 2024).
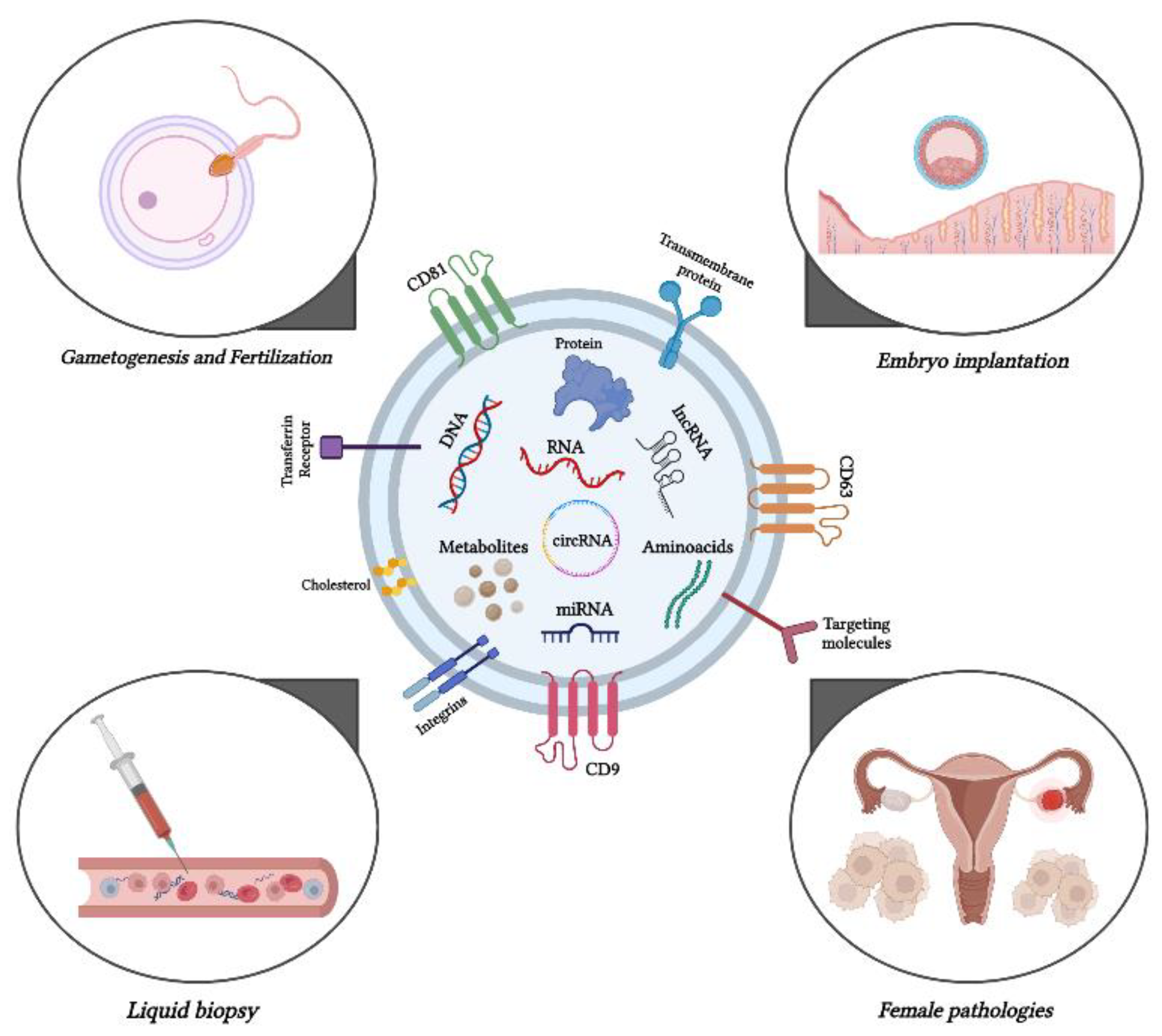
2. Exploring the activity of exosomes in gametogenesis and fertilization
In sexual reproduction, sperms and oocytes represent the fundamental protagonists generating a new organism. The maturation and the functionalities of germinal cells are critical points for reproductive success [44]. In the last years, several scientific evidence suggest that exosomes play significant roles in gametogenesis, facilitating communication between germ cells and the somatic cells and influencing proliferation, differentiation, and maturation of the gametes [44,45,46,47,48]. In spermatogenesis, the sperm maturation process and the acquisition of fertilization capability involve a series of intricate interactions along the male reproductive tract [49,50]. The Sertoli cells support and coordinate germ cell development and spermatogenesis by offering nutrients and participating in the formation of the blood-testis barrier [51]. A few studies, reveal that Sertoli cells are able to release exosomes in the testis and allow sperm differentiation into the spermatogenic tubules [44], [52,53]. The epididymis, comprising the caput, corpus, and cauda, facilitates sperm maturation, transport, and storage: each segment offers a unique habitat optimized for different stages of sperm development [54,55]. Sperms undergo crucial changes in morphology and function during their transit through the epididymis, where they interact with different microenvironments. Throughout this transit, sperm modifies membrane composition and surface proteins, influenced by interactions with the surrounding intraluminal fluid. Numerous studies have demonstrated EVs secretion by epididymal epithelial cells (epididymosomes) and by prostate (prostasomes) [56,57,58,59,60]. These vesicles are able to orchestrate sperm pathways – such as capacitation, acrosome reaction, protection against oxidative stress, stability and fluidity – by transporting bioactive molecules such as sperm adhesion molecule 1 (SPAM1), glioma pathogenesis-related 1-like protein 1 (GliPr1L1), metalloprotease, proto-oncogene tyrosine-protein kinase Src (cSrc), macrophage migration inhibitory factor (MIF), dicarbonyl/L-xylulose reductase (DCXR), Liprin α3,and also miRNAs [45,61,62,63]. In fact, it seems that exosome-derived miRNAs are a new component of cell-cell communication: a recent study reveal that Sertoli cell exosome-derived miR-486-5p is able to down-regulate PTEN and up-regulate the expression of Stra8, promoting the differentiation of spermatogonial stem cells (SSCs) into spermatogonia in mice [19,64]. A study conducted in 2022., [65] investigated and analyzed the protein and miRNA components of human testicular endothelial cell-derived exosomes (HTEC-Exos) revealing that the deregulation of the expression of miRNAs (hsa-miR-511, hsa-miR-222-3p, hsa-miR-320a) in sperm cells, epididymis, seminal plasma, and EVs (i.e., exosomes and microvesicles) may lead to the alterations of spermatogenesis and various forms of infertility [66]. In mammalian female gametogenesis, mature ovarian follicle, comprising oocyte, somatic cells (cumulus granulosa, mural granulosa and theca cells), and follicular fluid (FF), represent a critical dynamic microenvironment rich in metabolites and hormones to ensure follicular development and oocyte maturation [67]. Bidirectional communication between gametes and granulosa cells in the follicle occurs either directly by a network of gap junctions or through paracrine, autocrine and endocrine signalling factors in the FF [68,69]. These interactions guide to the modulation of oocyte transcriptional activity, in order to produce a competent oocyte able to support fertilization and preimplantation embryo development [70,71,72,73]. Emerging evidence highlight that exosomes contribute to this intricate interplay by transporting miRNAs that regulate several pathways involved in folliculogenesis [74], including WNT, TGFβ, MAPK, ErbB, and ubiquitin-mediated signalling pathways [45,75,76]. Many studies about the involvement of EVs from FF in follicular maturation use animal models. Hung et al.[77] have demonstrated that bovine follicular EVs are up-taken by bovine or mouse cumulus cell, consequently inducing the expansion of cumulus-oocyte complex (COC) thanks to the modulation of Prostaglandin-endoperoxide synthase 2 (Ptgs2) and Pentraxin 3 (Ptx3) expression, genes known to be involved in this process [77]. EVs originating in early, mid-estrus and pre-ovulation stages of oocyte maturation in mares, present dissimilar concentrations and different miRNA contents depending on the stage. In particular, miR-125 and miR-199, probably secreted by granulosa cells and packaged in EVs, were particularly expressed in the pre-ovulatory stages, being involved in oocyte maturation and cumulus proliferation. MiR-21, miR-132, and miR-212, whose expression seems to be hCG/LH-related [10] regulate gene expression inducing granulosa cell proliferation and the maturation of the cumulus-oocyte complex [78]. In 2014, Diez-Fraile et al. [79] identified four different expressed miRNAs in EVs from FF comparing younger with older women: specifically, they found miR-99b, miR-134 and miR-190b upregulated and miR-21-5p downregulated in older women [10]. These miRNAs regulate genes involved in heparan-sulphate proteoglycan expression, carbohydrate digestion and absorption, and apoptosis and their altered regulation could affect follicle development and oocyte maturation [10,45,79]. Finally, it seems that exosomes may take part also in the fertilization process, which represents a complex regulatory mechanism involving sequential events essential for the successful merge of sperm and oocyte [45]. When sperm passes through the uterus, the uterine cell-derived exosomes (uterosomes) carry transmembrane proteins and glycosyl phosphatidylinositol junction protein (SPAM1), which are essential for sperm fertilization and enhance the ability to cross the cumulus cell [80]. Similarly, when sperm passes through the oviduct, oocyte-derived exosomes (Oc-Exo) deliver specific sugar protein membrane Ca2+-ATpase4 (PMCA4) to the sperm surface improving the resistance to zona pellucida hydrolysis and reducing multiple sperm fertilization. Besides, Oc-Exo promote spermatozoa motility and their capacitation, thanks to the induction of tyrosine phosphorylation and consequently activating the acrosome reaction [81]: in particular, Oc-Exo carry two tetraspanins - CD81 and CD9 - [44] involved in sperm-egg fusion, acting independently of each other [44,82]. Other molecules participate in the process of sperm-egg fusion, such as glutathione peroxidase-5 (GPX5), SPAM1, prostate-specific antigen (PSA), kinesin family member 5B (KIF5B), annexin A2 (ANXA2) and kallikrein 2 (KLK2) which are delivered by human semen-derived exosomes [83]. Thanks to this finely regulated communication among gametes, orchestrated by a variety of factors and signalling molecules, the development and the implantation of embryo will be able to proceed.
3. Exosomes and embryo implantation
Embryo implantation is a critical step and requires a close dialogue between the embryo and maternal tissues coordinated by molecular and physiological signalling. The embryo adhesion to receptive endometrium needs the protection of embryo from the maternal immune system before implantation, but also the maternal endometrial inflammatory response after implantation promoting embryo invasion [84,85,86]. During the embryo attachment, embryonic EVs seem to transport molecules potentially able to modulate the host endometrial immune system [87,88]. An increasing repertoire of uterine CD56, CD16, natural killer (uNK) cells, T lymphocytes, B cells, and dendritic cells are recruited at implantation site during embryo implant due to their ability to secrete immunosuppressive IL-10 [89,90] to protect the embryo from maternal immune attacks. Moreover, this modulation seems to depend on miRNA and protein EV cargo [91] able to influence the establishment of pregnancy, fetal development and survival during pregnancy [92,93]. Moreover, there is increasing evidence that Uterine Fluid EVs (UF-EVs) have potent effects on the maternal immune system during implantation. Nakamura et al. in two papers published in 2019 and in 2021 [94,95] reported that in cattle, during receptivity, UF-derived EVs carry miRNAs, bta-miR-98 [94] and bta-miR-26b [95], which target and negatively regulate immunoregulatory genes in endometrial recipient cells (including CTSC, IL6, CASP4, and IKBKE, and PSMC6, CD40, and IER3, respectively) [96]. Gene set enrichment analyses of downstream target genes highlighted the involvement of neutrophil activation in immune response and neutrophil-mediated immunity as potential immunomodulatory functions. Embryos are able to release some immunosuppressive molecules (as hCG, HLAG) inside EVs, inducing the production of immunosuppressive factors (e.g., IL-10 [97]) to protect themselves from maternal immune responses during the implantation process. For instance, earlier evidence suggests that mouse embryonic EVs containing progesterone-induced-blocking factor 1 (PIBF), could interact with CD4+ and CD8+ peripheral T-cells, causing IL-10 production and immunosuppression [97]. Similarly, in a study published in 2019 [98] the authors suggested that HSPE1-associated trophoblast cell line BeWo can release EVs able to suppress regulatory T-cell (Treg) signalling by modulation of miRNAs (like hsa-miR-23b, hsamiR-146a, hsa-miR-155, hsa-miR-22, and hsa-miR-221) and protein (HSPE1) expression on the T-cell surface and lumen highlighting their potential to regulate immune cell function during embryo implantation [96]. Another study conducted by Rai et al. in 2021 [99] highlights that EVs present in human UF contain antioxidative regulators (SOD1, GSTO1, MPO, CAT) specifically during the secretory phase [99]: they are able to reduce ROS levels facilitating the embryo apposition and his attachment to the endometrium [96].The control and regulation of the communication between the mother and developing embryo require also the contribution of the placenta. It is an essential organ performing vital functions (ensuring gas exchange, nutrient and waste transfer, immunoglobulin transport, and hormone secretion [18] for the fetus to support its growth and survival and maintain the pregnancy. The crosstalk between fetus and mother can involve the simple diffusion of molecules through tissue layers but also the extracellular vesicles, especially exosomes [93]. During pregnancy, various types of placental cells are able to secrete exosomes - placenta-derived exosomes - which concentration continuously increases in maternal circulation over the first trimester of pregnancy [44]. They may be differentiated from other exosomes by the presence of placenta-specific growth factors, DNA fragments, mRNAs and miRNAs, which are involved in regulating the physiological function of the maternal uterus and fetal development [44]. Among the most interesting miRNAs, we can find several miRNAs located in chromosome 19 - miRNA cluster C19MC - which is the largest cluster of miRNAs in the human genome [100]. MiRNAs within the human C19MC include 46 miRNAs which are expressed during pregnancy only in the placenta, the so-called placenta-associated miRNAs [101,102], secreted into the maternal circulation by exosomes where they function in placental-maternal signalling [103,104]. For example, exosomal miR-517b increased the expression of TNFα and/or other death ligands [93], instead miR-516b-5p, miR-517-5p, and miR-518a-3p are shown to inlfuence the PI3K-Akt and therefore the insulin signalling pathways, and their expression levels seems to be regulated by various stimuli, including oxidative stress and glucose levels [18,105]. Furthermore, the exosomal transfer of placenta-specific miR-571a-3p into NK cells repressed cGMP-dependent protein kinase 1, a crucial mediator of nitric oxide signalling [93,106]. Different papers underline the EVs cargo as able to modulate the embryo-mother bidirectional cross-talk [107,108]. In vivo, EVs have been found in uterine fluid and, in vitro, in the culture medium, released from endometrial cancer cell lines (ECC1) [109]. Even human embryos, obtained from In Vitro Fertilization cycles (IVF), can secrete exosomes and microvesicles in the culture medium and it has been demonstrated that embryo EVs are received by primary endometrial cells [110]. Therefore, accurate characterization of their cargo could provide useful information about the quality of the embryo and endometrial receptivity in assisted reproductive medicine; actually, one of the most fascinating challenges in reproductive medicine is the identification of new biomarkers able to indicate the best quality embryos to implant [10]. Currently, Preimplantation Genetic Screening (PGS) is often associated with the morphological evaluation of embryo quality, to detect euploid embryos to implant. Even though the potential of PGS, the invasive removal of cells from the trophectoderm is still considered a critical procedure [111]. For these reasons, the spent culture medium analysis could represent an innovative method to assess embryo competence during implantation [10]. Giacomini at al. in 2017 demonstrated for the first time that human preimplantation embryos at different developmental stages (at day 3 and day 5after fertilization) can release EVs during their in vitro culture for ART (Assisted Reproductive Technologies) [112]. The authors isolated and characterized the exosomes from embryo-derived conditioned media, according to the guidelines of the MISEV; moreover, the embryo vesicles can be internalized by cultured endometrial cells, suggesting their ability to communicate with the maternal side [112]. Furthermore, co-culture techniques using both embryonic and endometrial cells have been introduced in a paper conducted by Bhadarka et al. [113], with intent to mimic the uterine microenvironment and improve the quality of the embryo; the authors obtained better quality blastocyst with a higher implantation rate, by culturing human embryos coming from women who had undergone Intra Cytoplasmic Sperm Injection (ICSI) with cumulus cells [10,113]. Investigating the nucleic acid constituent of the EVs released by the embryo in the culture media, in 2017 Pallinger et al. demonstrated, by using flow cytometry, that only the embryo releasing a low number of vesicles were competent, probably because a higher presence of nucleic acid could be related to cell injury and, consequently, to embryo damage [114]. Other studies have also shown the presence of miRNAs in the embryo culture medium, correlated with in vitro fertilization methods, embryo aneuploidy and pregnancy outcome [115,116]; Abu-Halima et al. by analyzing extracellular vesicle secretion and miRNA expression in the spent culture media after embryo transfer, observed a reduced miRNA amount, related to decreased EVs secretion from an embryo successfully implanted in comparison with an embryo with a negative outcome [10,117]. Another approach to evaluate embryo-quality is revealed in a study published in 2019 by Battaglia et al., [118]: for the first time, the authors demonstrated the human embryo’s ability to secrete exosomes enriched with miRNAs inside the human blastocoel fluid (BF). The bioinformatic and comparative analyses identified the biological function of these miRNAs in critical signalling pathways controlling embryo development, such as pluripotency, cell reprogramming, epigenetic modifications, intercellular communication, cell adhesion and cell fate. MiRNAs of BF reflect the miRNome of embryonic cells and their presence in exosomes, strongly suggests their important role in mediating cellular signalling among blastocyst cells. Their characterization is important to study the earliest stages of embryogenesis and the complex paths regulating pluripotency. Above all, miRNA expression profiles in BF, could be used as possible minimally invasive biomarkers of embryos, predicting implantation rate in IVF cycles [10,118].
4. Exosomes involved in female pathologies
Several studies proposed exosomes as some of the protagonists involved in a high number of female pathologies - premature ovarian failure (POF), recurrent implantation failure (RIF), preeclampsia, endometriosis, cervical and endometrial cancer, polycystic ovarian syndrome (PCOS) – delivering different cargoes, especially miRNAs which could be regarded as diagnostic biomarkers [43,119,120,121,122,123]. The investigation of regulatory disorders caused by the altered EVs secretion, modifying the regulation of genes and protein expression and several signalling pathways such as wingless signalling pathway (WNT), transforming growth factor beta (TGF-β), neurotrophins, insulin signalling pathways, mitogen-activated protein kinase (MAPK), epidermal growth factor receptor (ErbB) pathways and pathways associated with ubiquitin, could offer insights into implantation failure in women with reproductive diseases [124,125]. For instance, a recent study, conducted by Zhou et al. illustrates for the first time the differential expression patterns of exosomal miRNAs from endometrial stroma cells of women with endometriosis-associated infertility [120]. Hsa-miR-494-3p, hsa-miR-10b-3p, hsa-125b-2-3p, and hsa-miR-1343-3pdisplay higher expression levels and some of their predicted target genes are related to endometrial receptivity: homeobox A10 (HOXA10) and leukemia inhibitory factor (LIF). HOXA10 is involved in morphological arrangements of the uterus and in endometrial regeneration during menstrual cycle [126,127]; instead, LIF is one of the most important cytokines essential for regular implantation [128]. Their expression is significantly decreased in patients with endometriosis, affecting endometrial receptivity and implantation [120]. Another study revealed that circulating exosomes in PCOS follicular fluid had differential miRNAs expression compared to healthy female controls: miR-146a-5p and miR-126-3p expression levels were increased, instead miR-20b-5p, miR-106a-5p, and miR-18a-3p showed a decreasing trend in PCOS patients. The computational analysis highlighted the involvement of miRNAs in MAPK signalling pathway, axon guidance, circadian rhythms, endocytosis, and tumorigenesis circuits suggesting that they may confer a risk of PCOS [129]. Li, H., et al. in a study conducted in 2020, isolated exosomes from women with preeclampsia, reporting that the concentration and the mean diameter of plasma exosomes were greater than healthy controls (women with uncomplicated term pregnancies) [130]. The authors found the up-regulation of miR-153-3p and miR-325-3p into exosomes from preeclampsia [130], which are known to be able to inhibit cell proliferation and promote apoptosis [131]. Recently, researchers have started to investigate the relationship between exosomal miRNAs and their influence on ovarian cancer [123]. Previous studies have revealed that exosomes could influence the chemo-susceptibility in recipient cells by regulating different biological pathways, including cell cycle and apoptosis: for instance, miR-106a, miR-130a, miR-221, miR-222, miR-433, and miR-591 are introduced as modulators of drug resistance in ovarian cancer [132,133]. Additionally, a newer analysis indicated that macrophage-derived exosomes transfer miR-223 to epithelial ovarian cancer cells to promote drug resistance through the PI3K/AKT signalling pathway [123,134]. Besides, Li et al. [135] showed that cancer-associated fibroblasts (CAFs)-derived exosomes induce endometrial cancer progression partially due to the loss of miR-148b in the exosomes, which represent an important tumoral suppressor. MiR-148b targets DNA (cytosine-5) methyltransferase1 (DNMT1) that suppresses endometrial cancer metastasis by increasing epithelial-mesenchymal transition (EMT) [135]. Another study observed that exosomal miR-320a, derived from CAFs, has a lower expression in endometrial cancer cells and tissues: it targets hypoxia-inducible factor 1 subunit alpha (HIF1α) which reduce the vascular endothelial growth factor a (VEGFA) expression inhibiting cell proliferation [136]. More investigations on the central role of exosomes and their miRNAs cargo in the pathophysiology of reproduction are required to describe the effect of exosomes on the activity of cancer. A better approaches to learn their activities in the female reproductive system secretome could promote the development of innovative diagnostic and therapeutic tools [18,123].
5. Conclusion and Future Perspectives
This review summarizes the principal implications of exosomal vesicles in a wide range of biological processes related to reproduction, acting as essential mediators of intercellular communication and major regulators of cell functions. The great versatility of exosomes is evident in their involvement in a large number of processes such as germinal cell development and regulation of the reproductive system, tumor growth, tissue homeostasis, immune regulation, disease progression. However, despite the notable progress during the recent years, several challenges remain to be addressed. These include the standardization of isolation and characterization techniques, investigation of specific cargo-loading mechanisms, and optimization of exosome-based therapeutic strategies for clinical translation medicine. Overall, ongoing and future studies on exosomes will be able to underline their full potential in biomedical applications and personalized medicine, to improve diagnosis, treatment and management of diseases, as well as increasing positive outcomes in assisted reproduction techniques.
Author Contributions
Conceptualization A.F., A.C., C.F., M.P., C.D.P., R.B.; writing—review and editing A.F., A.C., C.F., M.P., C.D.P., R.B; visualization and supervision A.F., A.C., C.F., M.P., C.D.P., R.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sung, B.H.; Parent, C.A.; Weaver, A.M. Extracellular vesicles: Critical players during cell migration. Dev. Cell 2021, 56, 1861–1874. [Google Scholar] [CrossRef] [PubMed]
- Ginini, L.; Billan, S.; Fridman, E.; Gil, Z. Insight into Extracellular Vesicle-Cell Communication: From Cell Recognition to Intracellular Fate. Cells 2022, 11, 1375. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Bedina Zavec, A.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, M. A. , Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology (Basel) 2022, 11, (6). [Google Scholar] [CrossRef] [PubMed]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [PubMed]
- Abramowicz, A.; Widłak, P.; Pietrowska, M. Different Types of Cellular Stress Affect the Proteome Composition of Small Extracellular Vesicles: A Mini Review. Proteomes 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed]
- Menck, K.; Bleckmann, A.; Schulz, M.; Ries, L.; Binder, C. , Isolation and Characterization of Microvesicles from Peripheral Blood. J Vis Exp 2017, (119).
- Yu, L.; Zhu, G.; Zhang, Z.; Yu, Y.; Zeng, L.; Xu, Z.; Weng, J.; Xia, J.; Li, J.; Pathak, J.L. Apoptotic bodies: bioactive treasure left behind by the dying cells with robust diagnostic and therapeutic application potentials. J. Nanobiotechnology 2023, 21, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Rajput, A.; Varshney, A.; Bajaj, R.; Pokharkar, V. Exosomes as New Generation Vehicles for Drug Delivery: Biomedical Applications and Future Perspectives. Molecules 2022, 27, 7289. [Google Scholar] [CrossRef]
- Andronico, F.; Battaglia, R.; Ragusa, M.; Barbagallo, D.; Purrello, M.; Di Pietro, C. Extracellular Vesicles in Human Oogenesis and Implantation. Int. J. Mol. Sci. 2019, 20, 2162. [Google Scholar] [CrossRef]
- Xu, M.; Ji, J.; Jin, D.; Wu, Y.; Wu, T.; Lin, R.; Zhu, S.; Jiang, F.; Ji, Y.; Bao, B.; et al. The biogenesis and secretion of exosomes and multivesicular bodies (MVBs): Intercellular shuttles and implications in human diseases. Genes Dis. 2023, 10, 1894–1907. [Google Scholar] [CrossRef]
- Yim, K.H.W.; Krzyzaniak, O.; Al Hrout, A.; Peacock, B.; Chahwan, R. Assessing Extracellular Vesicles in Human Biofluids Using Flow-Based Analyzers. Adv. Heal. Mater. 2023, 12, e2301706. [Google Scholar] [CrossRef] [PubMed]
- Calvani, R.; Picca, A.; Guerra, F.; Coelho-Junior, H.J.; Bucci, C.; Marzetti, E. Circulating extracellular vesicles: friends and foes in neurodegeneration. Neural Regen. Res. 2022, 17, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Hánělová, K.; Raudenská, M.; Masařík, M.; Balvan, J. Protein cargo in extracellular vesicles as the key mediator in the progression of cancer. Cell Commun. Signal. 2024, 22, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Donoso-Quezada, J.; Ayala-Mar, S.; González-Valdez, J. The role of lipids in exosome biology and intercellular communication: Function, analytics and applications. Traffic 2021, 22, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Jang, H.; Cho, H.; Choi, J.; Hwang, K.Y.; Choi, Y.; Kim, S.H.; Yang, Y. Recent Advances in Exosome-Based Drug Delivery for Cancer Therapy. Cancers 2021, 13, 4435. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.M.; Adam, M.; Hammond, J.R.; Orr, L.; Turbide, C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 1987, 262, 9412–9420. [Google Scholar] [CrossRef] [PubMed]
- Ghafourian, M.; Mahdavi, R.; Jonoush, Z.A.; Sadeghi, M.; Ghadiri, N.; Farzaneh, M.; Salehi, A.M. The implications of exosomes in pregnancy: emerging as new diagnostic markers and therapeutics targets. Cell Commun. Signal. 2022, 20, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-Y.; Liu, S.-P.; Dai, X.-F.; Lan, D.-F.; Song, T.; Wang, X.-Y.; Kong, Q.-H.; Tan, J.; Zhang, J.-D. The emerging role of exosomes in the development of testicular. Asian J. Androl. 2023, 25, 547–555. [Google Scholar] [CrossRef]
- Russell, A.E.; Sneider, A.; Witwer, K.W.; Bergese, P.; Bhattacharyya, S.N.; Cocks, A.; Cocucci, E.; Erdbrügger, U.; Falcon-Perez, J.M.; Freeman, D.W.; et al. Biological membranes in EV biogenesis, stability, uptake, and cargo transfer: an ISEV position paper arising from the ISEV membranes and EVs workshop. J. Extracell. Vesicles 2019, 8, 1684862. [Google Scholar] [CrossRef]
- Leonardi, A.A.; Battaglia, R.; Morganti, D.; Faro, M.J.L.; Fazio, B.; De Pascali, C.; Francioso, L.; Palazzo, G.; Mallardi, A.; Purrello, M.; et al. A Novel Silicon Platform for Selective Isolation, Quantification, and Molecular Analysis of Small Extracellular Vesicles. Int. J. Nanomed. 2021, ume 16, 5153–5165. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Welsh, J. A.; Goberdhan, D. C. I.; O'Driscoll, L.; Buzas, E. I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T. A. P.; Erdbrugger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J Extracell Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef]
- Konoshenko, M.Y.; Lekchnov, E.A.; Vlassov, A.V.; Laktionov, P.P. Isolation of Extracellular Vesicles: General Methodologies and Latest Trends. BioMed Res. Int. 2018, 2018, 8545347. [Google Scholar] [CrossRef]
- Reclusa, P.; Verstraelen, P.; Taverna, S.; Gunasekaran, M.; Pucci, M.; Pintelon, I.; Claes, N.; de Miguel-Pérez, D.; Alessandro, R.; Bals, S.; et al. Improving extracellular vesicles visualization: From static to motion. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Szatanek, R.; Baj-Krzyworzeka, M.; Zimoch, J.; Lekka, M.; Siedlar, M.; Baran, J. The Methods of Choice for Extracellular Vesicles (EVs) Characterization. Int. J. Mol. Sci. 2017, 18, 1153. [Google Scholar] [CrossRef]
- Schageman, J.; Zeringer, E.; Li, M.; Barta, T.; Lea, K.; Gu, J.; Magdaleno, S.; Setterquist, R.; Vlassov, A.V. The Complete Exosome Workflow Solution: From Isolation to Characterization of RNA Cargo. BioMed Res. Int. 2013, 2013, 1–15. [Google Scholar] [CrossRef]
- Jankovičová, J.; Sečová, P.; Michalková, K.; Antalíková, J. Tetraspanins, More than Markers of Extracellular Vesicles in Reproduction. Int. J. Mol. Sci. 2020, 21, 7568. [Google Scholar] [CrossRef]
- Zhang, H.; Lyden, D. Asymmetric-flow field-flow fractionation technology for exomere and small extracellular vesicle separation and characterization. Nat. Protoc. 2019, 14, 1027–1053. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Shao, H.; Im, H.; Castro, C.M.; Breakefield, X.; Weissleder, R.; Lee, H. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018, 118, 1917–1950. [Google Scholar] [CrossRef]
- He, J.; Ren, W.; Wang, W.; Han, W.; Jiang, L.; Zhang, D.; Guo, M. Exosomal targeting and its potential clinical application. Drug Deliv. Transl. Res. 2021, 12, 2385–2402. [Google Scholar] [CrossRef]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Mark, M.T.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef]
- Kawamura, S.; Iinuma, H.; Wada, K.; Takahashi, K.; Minezaki, S.; Kainuma, M.; Shibuya, M.; Miura, F.; Sano, K. , Exosome-encapsulated microRNA-4525, microRNA-451a and microRNA-21 in portal vein blood is a high-sensitive liquid biomarker for the selection of high-risk pancreatic ductal adenocarcinoma patients. J Hepatobiliary Pancreat Sci 2019, 26, (2), 63–72. [Google Scholar] [CrossRef]
- Cai, X.; Janku, F.; Zhan, Q.; Fan, J.-B. Accessing Genetic Information with Liquid Biopsies. Trends Genet. 2015, 31, 564–575. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Qiu, Y.; Li, P.; Zhang, Z.; Wu, M. , Insights Into Exosomal Non-Coding RNAs Sorting Mechanism and Clinical Application. Front Oncol 2021, 11, 664904. [Google Scholar] [CrossRef]
- Yu, D.; Li, Y.; Wang, M.; Gu, J.; Xu, W.; Cai, H.; Fang, X.; Zhang, X. Exosomes as a new frontier of cancer liquid biopsy. Mol. Cancer 2022, 21, 1–33. [Google Scholar] [CrossRef]
- Irmer, B.; Chandrabalan, S.; Maas, L.; Bleckmann, A.; Menck, K. Extracellular Vesicles in Liquid Biopsies as Biomarkers for Solid Tumors. Cancers 2023, 15, 1307. [Google Scholar] [CrossRef]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in Exosome Isolation Techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef]
- He, C.; Zheng, S.; Luo, Y.; Wang, B. Exosome Theranostics: Biology and Translational Medicine. Theranostics 2018, 8, 237–255. [Google Scholar] [CrossRef]
- Kanwar, S.S.; Dunlay, C.J.; Simeone, D.M.; Nagrath, S. Microfluidic device (ExoChip) for on-chip isolation, quantification and characterization of circulating exosomes. Lab a Chip 2014, 14, 1891–1900. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Wrzecińska, M.; Czerniawska-Piątkowska, E.; Kupczyński, R. Exosomes – Spectacular role in reproduction. Biomed. Pharmacother. 2022, 148, 112752. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Z.; Gu, X.; Sheng, X.; Xiao, L.; Wang, X. Exosomes: New regulators of reproductive development. Mater. Today Bio 2023, 19, 100608. [Google Scholar] [CrossRef]
- Machtinger, R.; Laurent, L.C.; Baccarelli, A.A. Extracellular vesicles: roles in gamete maturation, fertilization and embryo implantation. Hum. Reprod. Update 2016, 22, 182–193. [Google Scholar] [CrossRef]
- Qamar, A.Y.; Mahiddine, F.Y.; Bang, S.; Fang, X.; Shin, S.T.; Kim, M.J.; Cho, J. Extracellular Vesicle Mediated Crosstalk Between the Gametes, Conceptus, and Female Reproductive Tract. Front. Veter- Sci. 2020, 7, 589117. [Google Scholar] [CrossRef]
- Capra, E.; Lange-Consiglio, A. , The Biological Function of Extracellular Vesicles during Fertilization, Early Embryo-Maternal Crosstalk and Their Involvement in Reproduction: Review and Overview. Biomolecules 2020, 10. [Google Scholar] [CrossRef]
- Aleksejeva, E.; Zarovni, N.; Dissanayake, K.; Godakumara, K.; Vigano, P.; Fazeli, A.; Jaakma, U.; Salumets, A. , Extracellular vesicle research in reproductive science: Paving the way for clinical achievementsdagger. Biol Reprod 2022, 106, 408–424. [Google Scholar] [CrossRef]
- Gervasi, M.G.; Visconti, P.E. Molecular changes and signaling events occurring in spermatozoa during epididymal maturation. Andrology 2017, 5, 204–218. [Google Scholar] [CrossRef]
- Sullivan, R.; Saez, F.; Girouard, J.; Frenette, G. Role of exosomes in sperm maturation during the transit along the male reproductive tract. Blood Cells, Mol. Dis. 2005, 35, 1–10. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Mruk, D.D. The Blood-Testis Barrier and Its Implications for Male Contraception. Pharmacol. Rev. 2011, 64, 16–64. [Google Scholar] [CrossRef]
- Rimmer, M.P.; Gregory, C.D.; Mitchell, R.T. The transformative impact of extracellular vesicles on developing sperm. Reprod. Fertil. 2021, 2, R51–R66. [Google Scholar] [CrossRef]
- Amiri, N.; Mohammadi, P.; Allahgholi, A.; Salek, F.; Amini, E. The potential of sertoli cells (SCs) derived exosomes and its therapeutic efficacy in male reproductive disorders. Life Sci. 2023, 312, 121251. [Google Scholar] [CrossRef]
- Dacheux, J.-L.; Belleannée, C.; Guyonnet, B.; Labas, V.; Teixeira-Gomes, A.-P.; Ecroyd, H.; Druart, X.; Gatti, J.-L.; Dacheux, F. The contribution of proteomics to understanding epididymal maturation of mammalian spermatozoa. Syst. Biol. Reprod. Med. 2012, 58, 197–210. [Google Scholar] [CrossRef]
- Belleannée, C. ; Calvo, ; Caballero, J.; Sullivan, R. Epididymosomes Convey Different Repertoires of MicroRNAs Throughout the Bovine EpididymisBiol. Reprod. 2013, 89, 30. [Google Scholar] [CrossRef]
- Ronquist, G.; Brody, I. The prostasome: its secretion and function in man. Biochim. et Biophys. Acta (BBA) - Rev. Biomembr. 1985, 822, 203–218. [Google Scholar] [CrossRef]
- Ronquist, G.; Brody, I.; Gottfries, A.; Stegmayr, B. , An Mg2+ and Ca2+-stimulated adenosine triphosphatase in human prostatic fluid: part I. Andrologia 1978, 10, (4), 261–72. [Google Scholar]
- Ronquist, K.G.; Ronquist, G.; Carlsson, L.; Larsson, A. Human prostasomes contain chromosomal DNA. Prostate 2009, 69, 737–743. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Kamiguchi, Y.; Mikamo, K.; Suzuki, F.; Yanagimachi, H. Maturation of spermatozoa in the epididymis of the Chinese hamster. Am. J. Anat. 1985, 172, 317–330. [Google Scholar] [CrossRef]
- Sullivan, R.; Saez, F. Epididymosomes, prostasomes, and liposomes: their roles in mammalian male reproductive physiology. Reproduction 2013, 146, R21–R35. [Google Scholar] [CrossRef]
- Vojtech, L.; Woo, S.; Hughes, S.; Levy, C.; Ballweber, L.; Sauteraud, R.P.; Strobl, J.; Westerberg, K.; Gottardo, R.; Tewari, M.; et al. Exosomes in human semen carry a distinctive repertoire of small non-coding RNAs with potential regulatory functions. Nucleic Acids Res. 2014, 42, 7290–7304. [Google Scholar] [CrossRef]
- Arienti, G.; Carlini, E.; Palmerini, C. Fusion of Human Sperm to Prostasomes at Acidic pH. J. Membr. Biol. 1997, 155, 89–94. [Google Scholar] [CrossRef]
- Arienti, G.; Carlini, E.; Nicolucci, A.; Cosmi, E.V.; Santi, F.; Palmerini, C.A. The motility of human spermatozoa as influenced by prostasomes at various pH levels. Biol. Cell 1999, 91, 51–54. [Google Scholar] [CrossRef]
- Li, Q.; Li, H.; Liang, J.; Mei, J.; Cao, Z.; Zhang, L.; Luo, J.; Tang, Y.; Huang, R.; Xia, H.; et al. Sertoli cell-derived exosomal MicroRNA-486-5p regulates differentiation of spermatogonial stem cell through PTEN in mice. J. Cell. Mol. Med. 2021, 25, 3950–3962. [Google Scholar] [CrossRef]
- Guan, R.-L.; Song, W.-P.; Gu, S.-J.; Tan, X.-H.; Gu, Y.-Y.; Song, W.-D.; Zeng, J.-Y.; Xin, Z.-C. Proteomic analysis and miRNA profiling of human testicular endothelial cell-derived exosomes: the potential effects on spermatogenesis. Asian J. Androl. 2021, 24, 478. [Google Scholar] [CrossRef]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Carrell, D.T.; Jenkins, T.G.; Yeste, M. The role of miRNAs in male human reproduction: a systematic review. Andrology 2019, 8, 7–26. [Google Scholar] [CrossRef]
- Da Broi, M.G.; Giorgi, V.S.I.; Wang, F.; Keefe, D.L.; Albertini, D.; Navarro, P.A. Influence of follicular fluid and cumulus cells on oocyte quality: clinical implications. J. Assist. Reprod. Genet. 2018, 35, 735–751. [Google Scholar] [CrossRef]
- Eppig, J.J.; Chesnel, F.; Hirao, Y.; O'Brien, M.J.; Pendola, F.L.; Watanabe, S.; Wigglesworth, K. Oocyte control of granulosa cell development: how and why. . 1997, 12, 127–32. [Google Scholar]
- Matzuk, M.M.; Burns, K.H.; Viveiros, M.M.; Eppig, J.J. Intercellular Communication in the Mammalian Ovary: Oocytes Carry the Conversation. Science 2002, 296, 2178–2180. [Google Scholar] [CrossRef]
- Buccione, R.; Schroeder, A.C.; Eppig, J.J. Interactions between Somatic Cells and Germ Cells Throughout Mammalian OogenesisBiol. Reprod. 1990, 43, 543–547. [Google Scholar] [CrossRef]
- Adashi, E. Y. Endocrinology of the ovary. Hum Reprod 1994, 9, 815–27. [Google Scholar] [CrossRef]
- Senbon, S.; Hirao, Y.; Miyano, T. Interactions between the Oocyte and Surrounding Somatic Cells in Follicular Development: Lessons from In Vitro Culture. J. Reprod. Dev. 2003, 49, 259–269. [Google Scholar] [CrossRef]
- Hamel, M.; Dufort, I.; Robert, C.; Gravel, C.; Leveille, M.-C.; Leader, A.; Sirard, M.-A. Identification of differentially expressed markers in human follicular cells associated with competent oocytes. Hum. Reprod. 2008, 23, 1118–1127. [Google Scholar] [CrossRef]
- Di Pietro, C. Exosome-mediated communication in the ovarian follicle. J. Assist. Reprod. Genet. 2016, 33, 303–311. [Google Scholar] [CrossRef]
- da Silveira, J.C.; Veeramachaneni, D.R.; Winger, Q.A.; Carnevale, E.M.; Bouma, G.J. Cell-Secreted Vesicles in Equine Ovarian Follicular Fluid Contain miRNAs and Proteins: A Possible New Form of Cell Communication Within the Ovarian Follicle1. Biol. Reprod. 2012, 86, 71. [Google Scholar] [CrossRef]
- Santonocito, M.; Vento, M.; Guglielmino, M.R.; Battaglia, R.; Wahlgren, J.; Ragusa, M.; Barbagallo, D.; Borzì, P.; Rizzari, S.; Maugeri, M.; et al. Molecular characterization of exosomes and their microRNA cargo in human follicular fluid: bioinformatic analysis reveals that exosomal microRNAs control pathways involved in follicular maturation. Fertil. Steril. 2014, 102, 1751–1761. [Google Scholar] [CrossRef]
- Hung, W.-T.; Hong, X.; Christenson, L.K.; McGinnis, L.K. Extracellular Vesicles from Bovine Follicular Fluid Support Cumulus Expansion1. Biol. Reprod. 2015, 93, 117. [Google Scholar] [CrossRef]
- da Silveira, J.C.; de Ávila, A.C.F.C.M.; Garrett, H.L.; E Bruemmer, J.; A Winger, Q.; Bouma, G.J. Cell-secreted vesicles containing microRNAs as regulators of gamete maturation. J. Endocrinol. 2018, 236, R15–R27. [Google Scholar] [CrossRef]
- Diez-Fraile, A.; Lammens, T.; Tilleman, K.; Witkowski, W.; Verhasselt, B.; De Sutter, P.; Benoit, Y.; Espeel, M.; D'Herde, K. , Age-associated differential microRNA levels in human follicular fluid reveal pathways potentially determining fertility and success of in vitro fertilization. Hum Fertil (Camb) 2014, 17, (2), 90–8. [Google Scholar] [CrossRef]
- Griffiths, G.S.; Galileo, D.S.; Aravindan, R.G.; Martin-DeLeon, P.A. Clusterin Facilitates Exchange of Glycosyl Phosphatidylinositol-Linked SPAM1 Between Reproductive Luminal Fluids and Mouse and Human Sperm Membranes1. Biol. Reprod. 2009, 81, 562–570. [Google Scholar] [CrossRef]
- Murdica, V.; Giacomini, E.; Alteri, A.; Bartolacci, A.; Cermisoni, G.C.; Zarovni, N.; Papaleo, E.; Montorsi, F.; Salonia, A.; Viganò, P.; et al. Seminal plasma of men with severe asthenozoospermia contain exosomes that affect spermatozoa motility and capacitation. Fertil. Steril. 2019, 111, 897–908. [Google Scholar] [CrossRef]
- Kharazi, U.; Badalzadeh, R. A review on the stem cell therapy and an introduction to exosomes as a new tool in reproductive medicine. Reprod. Biol. 2020, 20, 447–459. [Google Scholar] [CrossRef]
- Baskaran, S.; Panner Selvam, M. K.; Agarwal, A. , Exosomes of male reproduction. Adv Clin Chem 2020, 95, 149–163. [Google Scholar] [PubMed]
- Franasiak, J.M.; Alecsandru, D.; Forman, E.J.; Gemmell, L.C.; Goldberg, J.M.; Llarena, N.; Margolis, C.; Laven, J.; Schoenmakers, S.; Seli, E. A review of the pathophysiology of recurrent implantation failure. Fertil. Steril. 2021, 116, 1436–1448. [Google Scholar] [CrossRef]
- Sehring, J.; Beltsos, A.; Jeelani, R. Human implantation: The complex interplay between endometrial receptivity, inflammation, and the microbiome. Placenta 2021, 117, 179–186. [Google Scholar] [CrossRef]
- Fan, W.; Qi, Y.; Wang, Y.; Yan, H.; Li, X.; Zhang, Y. Messenger roles of extracellular vesicles during fertilization of gametes, development and implantation: Recent advances. Front. Cell Dev. Biol. 2023, 10, 1079387. [Google Scholar] [CrossRef]
- Giacomini, E.; Alleva, E.; Fornelli, G.; Quartucci, A.; Privitera, L.; Vanni, V.S.; Viganò, P. Embryonic extracellular vesicles as informers to the immune cells at the maternal–fetal interface. Clin. Exp. Immunol. 2019, 198, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Kook, H.; Kang, S.; Lee, J. Study of immune-tolerized cell lines and extracellular vesicles inductive environment promoting continuous expression and secretion of HLA-G from semiallograft immune tolerance during pregnancy. J. Extracell. Vesicles 2020, 9. [Google Scholar] [CrossRef]
- Bulmer, J.N.; Morrison, L.; Longfellow, M.; Ritson, A.; Pace, D. Granulated lymphocytes in human endometrium: histochemical and immunohistochemical studies. Hum. Reprod. 1991, 6, 791–798. [Google Scholar] [CrossRef]
- Quenby, S.; Bates, M.; Doig, T.; Brewster, J.; Lewis-Jones, D.; Johnson, P.; Vince, G. Pre-implantation endometrial leukocytes in women with recurrent miscarriage. Hum. Reprod. 1999, 14, 2386–2391. [Google Scholar] [CrossRef]
- Wu, H.-M.; Chen, L.-H.; Hsu, L.-T.; Lai, C.-H. Immune Tolerance of Embryo Implantation and Pregnancy: The Role of Human Decidual Stromal Cell- and Embryonic-Derived Extracellular Vesicles. Int. J. Mol. Sci. 2022, 23, 13382. [Google Scholar] [CrossRef]
- Kaminski, V.d.L.; Ellwanger, J.H.; Chies, J.A.B. Extracellular vesicles in host-pathogen interactions and immune regulation — exosomes as emerging actors in the immunological theater of pregnancy. Heliyon 2019, 5, e02355. [Google Scholar] [CrossRef]
- Czernek, L.; Düchler, M. Exosomes as Messengers between Mother and Fetus in Pregnancy. Int. J. Mol. Sci. 2020, 21, 4264. [Google Scholar] [CrossRef]
- Nakamura, K.; Kusama, K.; Ideta, A.; Kimura, K.; Hori, M.; Imakawa, K. Effects of miR-98 in intrauterine extracellular vesicles on maternal immune regulation during the peri-implantation period in cattle. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Kusama, K.; Hori, M.; Imakawa, K. , The effect of bta-miR-26b in intrauterine extracellular vesicles on maternal immune system during the implantation period. Biochem Biophys Res Commun 2021, 573, 100–106. [Google Scholar] [CrossRef]
- Poh, Q.H.; Rai, A.; Salamonsen, L.A.; Greening, D.W. Omics insights into extracellular vesicles in embryo implantation and their therapeutic utility. Proteomics 2023, 23, e2200107. [Google Scholar] [CrossRef]
- Pallinger, E.; Bognar, Z.; Bogdan, A.; Csabai, T.; Abraham, H.; Szekeres-Bartho, J. PIBF+ extracellular vesicles from mouse embryos affect IL-10 production by CD8+ cells. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kovács, .F.; Fekete, N.; Turiák, L.; Ács, A.; Kőhidai, L.; Buzás, E.I.; Pállinger. Unravelling the Role of Trophoblastic-Derived Extracellular Vesicles in Regulatory T Cell Differentiation. Int. J. Mol. Sci. 2019, 20, 3457. [Google Scholar] [CrossRef]
- Rai, A.; Poh, Q.H.; Fatmous, M.; Fang, H.; Gurung, S.; Vollenhoven, B.; Salamonsen, L.A.; Greening, D.W. Proteomic profiling of human uterine extracellular vesicles reveal dynamic regulation of key players of embryo implantation and fertility during menstrual cycle. Proteomics 2021, 21, 2000211. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Mouillet, J.-F.; Coyne, C.; Sadovsky, Y. Review: Placenta-specific microRNAs in exosomes – Good things come in nano-packages. Placenta 2013, 35, S69–S73. [Google Scholar] [CrossRef]
- Noguer-Dance, M.; Abu-Amero, S.; Al-Khtib, M.; Lefèvre, A.; Coullin, P.; Moore, G.E.; Cavaillé, J. The primate-specific microRNA gene cluster (C19MC) is imprinted in the placenta. Hum. Mol. Genet. 2010, 19, 3566–3582. [Google Scholar] [CrossRef]
- Yuan, Z.; Sun, X.; Jiang, D.; Ding, Y.; Lu, Z.; Gong, L.; Liu, H.; Xie, J. Origin and evolution of a placental-specific microRNA family in the human genome. BMC Evol. Biol. 2010, 10, 346–346. [Google Scholar] [CrossRef] [PubMed]
- Donker, R.B.; Mouillet, J.F.; Chu, T.; Hubel, C.A.; Stolz, D.B.; Morelli, A.E.; Sadovsky, Y. The expression profile of C19MC microRNAs in primary human trophoblast cells and exosomes. Mol. Hum. Reprod. 2012, 18, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.-S.; Ishibashi, O.; Ishikawa, G.; Ishikawa, T.; Katayama, A.; Mishima, T.; Takizawa, T.; Shigihara, T.; Goto, T.; Izumi, A.; et al. Human Villous Trophoblasts Express and Secrete Placenta-Specific MicroRNAs into Maternal Circulation via Exosomes. Biol. Reprod. 2009, 81, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Song, G.; Lim, W. Effects of extracellular vesicles on placentation and pregnancy disorders. Reproduction 2019, 158, R189–R196. [Google Scholar] [CrossRef] [PubMed]
- Kambe, S.; Yoshitake, H.; Yuge, K.; Ishida, Y.; Ali, M.; Takizawa, T.; Kuwata, T.; Ohkuchi, A.; Matsubara, S.; Suzuki, M.; et al. Human Exosomal Placenta-Associated miR-517a-3p Modulates the Expression of PRKG1 mRNA in Jurkat Cells1. Biol. Reprod. 2014, 91, 129. [Google Scholar] [CrossRef] [PubMed]
- Kurian, N.K.; Modi, D. Extracellular vesicle mediated embryo-endometrial cross talk during implantation and in pregnancy. J. Assist. Reprod. Genet. 2018, 36, 189–198. [Google Scholar] [CrossRef]
- Saadeldin, I.; Oh, H.J; Lee, B. Embryonic–maternal cross-talk via exosomes: Potential implications. Stem Cells Clon. Adv. Appl. 2015, 8, 103–107. [Google Scholar] [CrossRef]
- Ng, Y.H.; Rome, S.; Jalabert, A.; Forterre, A.; Singh, H.; Hincks, C.L.; Salamonsen, L.A. Endometrial Exosomes/Microvesicles in the Uterine Microenvironment: A New Paradigm for Embryo-Endometrial Cross Talk at Implantation. PLOS ONE 2013, 8, e58502. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Liang, J.; Qin, T.; Zhang, Y.; Chen, X.; Wang, Z. The Role of Extracellular Vesicles in Embryo Implantation. Front. Endocrinol. 2022, 13, 809596. [Google Scholar] [CrossRef]
- Scott, R.T.; Upham, K.M.; Forman, E.J.; Zhao, T.; Treff, N.R. Cleavage-stage biopsy significantly impairs human embryonic implantation potential while blastocyst biopsy does not: a randomized and paired clinical trial. Fertil. Steril. 2013, 100, 624–630. [Google Scholar] [CrossRef]
- Giacomini, E.; Vago, R.; Sanchez, A.M.; Podini, P.; Zarovni, N.; Murdica, V.; Rizzo, R.; Bortolotti, D.; Candiani, M.; Viganò, P. Secretome of in vitro cultured human embryos contains extracellular vesicles that are uptaken by the maternal side. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bhadarka, H.K.; Patel, N.H.; Patel, N.H.; Patel, M.; Patel, K.B.; Sodagar, N.R.; Phatak, A.G.; Patel, J.S. Impact of embryo co-culture with cumulus cells on pregnancy & implantation rate in patients undergoing in vitro fertilization using donor oocyte. 2017; 146, 341–345. [Google Scholar] [CrossRef]
- Pallinger, E.; Bognar, Z.; Bodis, J.; Csabai, T.; Farkas, N.; Godony, K.; Varnagy, A.; Buzas, E.; Szekeres-Bartho, J. A simple and rapid flow cytometry-based assay to identify a competent embryo prior to embryo transfer. Sci. Rep. 2017, 7, 39927. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, A.; Ubaldi, F.M.; Cimadomo, D.; Noli, L.; Khalaf, Y.; Farcomeni, A.; Ilic, D.; Rienzi, L. MicroRNAs in spent blastocyst culture medium are derived from trophectoderm cells and can be explored for human embryo reproductive competence assessment. Fertil. Steril. 2015, 105, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Rosenbluth, E.M.; Shelton, D.N.; Wells, L.M.; Sparks, A.E.; Van Voorhis, B.J. Human embryos secrete microRNAs into culture media—a potential biomarker for implantation. Fertil. Steril. 2014, 101, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Häusler, S.; Backes, C.; Fehlmann, T.; Staib, C.; Nestel, S.; Nazarenko, I.; Meese, E.; Keller, A. Micro-ribonucleic acids and extracellular vesicles repertoire in the spent culture media is altered in women undergoing In Vitro Fertilization. Sci. Rep. 2017, 7, 13525–13525. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, R.; Palini, S.; Vento, M.E.; La Ferlita, A.; Faro, M.J.L.; Caroppo, E.; Borzì, P.; Falzone, L.; Barbagallo, D.; Ragusa, M.; et al. Identification of extracellular vesicles and characterization of miRNA expression profiles in human blastocoel fluid. Sci. Rep. 2019, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- E Schjenken, J.; Panir, K.; A Robertson, S.; Hull, M.L. Exosome-mediated intracellular signalling impacts the development of endometriosis—new avenues for endometriosis research. Mol. Hum. Reprod. 2018, 25, 2–4. [Google Scholar] [CrossRef]
- Zhou, W.; Lian, Y.; Jiang, J.; Wang, L.; Ren, L.; Li, Y.; Yan, X.; Chen, Q. Differential expression of microRNA in exosomes derived from endometrial stromal cells of women with endometriosis-associated infertility. Reprod. Biomed. Online 2020, 41, 170–181. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; Wang, R.; Xu, T.; Wang, J.; Cui, Z.; Cheng, F.; Wang, W.; Yang, X. Endometrial stem cell-derived exosomes repair cisplatin-induced premature ovarian failure via Hippo signaling pathway. Heliyon 2024, 10, e31639. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Hao, J.; Huang, X.; Liu, M.; Lv, M.; Su, C.; Mu, Y. L. , miRNA-122-5p in POI ovarian-derived exosomes promotes granulosa cell apoptosis by regulating BCL9. Cancer Med 2022, 11, (12), 2414–2426. [Google Scholar] [CrossRef]
- Esfandyari, S.; Elkafas, H.; Chugh, R.M.; Park, H.-S.; Navarro, A.; Al-Hendy, A. Exosomes as Biomarkers for Female Reproductive Diseases Diagnosis and Therapy. Int. J. Mol. Sci. 2021, 22, 2165. [Google Scholar] [CrossRef] [PubMed]
- Mobarak, H.; Rahbarghazi, R.; Lolicato, F.; Heidarpour, M.; Pashazadeh, F.; Nouri, M.; Mahdipour, M. Evaluation of the association between exosomal levels and female reproductive system and fertility outcome during aging: a systematic review protocol. Syst. Rev. 2019, 8, 1–6. [Google Scholar] [CrossRef]
- Jiang, N.-X.; Li, X.-L. The Complicated Effects of Extracellular Vesicles and Their Cargos on Embryo Implantation. Front. Endocrinol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake, D. L.; Malopolska, M. M.; Schwarz, T.; Tuz, R.; Bartlewski, P. M. , The roles and expression of HOXA/Hoxa10 gene: A prospective marker of mammalian female fertility? Reprod Biol 2022, 22, 100647. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Taylor, H. S. , The Role of Hox Genes in Female Reproductive Tract Development, Adult Function, and Fertility. Cold Spring Harb Perspect Med 2015, 6, a023002. [Google Scholar] [CrossRef] [PubMed]
- Kimber, S.J. Leukaemia inhibitory factor in implantation and uterine biology. Reproduction 2005, 130, 131–145. [Google Scholar] [CrossRef]
- Jiang, X.; Li, J.; Zhang, B.; Hu, J.; Ma, J.; Cui, L.; Chen, Z.-J. Differential expression profile of plasma exosomal microRNAs in women with polycystic ovary syndrome. Fertil. Steril. 2021, 115, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ouyang, Y.; Sadovsky, E.; Parks, W.T.; Chu, T.; Sadovsky, Y. Unique microRNA Signals in Plasma Exosomes from Pregnancies Complicated by Preeclampsia. Hypertension 2020, 75, 762–771. [Google Scholar] [CrossRef]
- Zeng, H.F.; Yan, S.; Wu, S.F. MicroRNA-153-3p suppress cell proliferation and invasion by targeting SNAI1 in melanoma. Biochem. Biophys. Res. Commun. 2017, 487, 140–145. [Google Scholar] [CrossRef]
- Liang, T.; Guo, Q.; Li, L.; Cheng, Y.; Ren, C.; Zhang, G. MicroRNA-433 inhibits migration and invasion of ovarian cancer cells via targeting Notch1. Neoplasma 2016, 63, 696–704. [Google Scholar] [CrossRef]
- Azmi, A.S.; Bao, B.; Sarkar, F.H. Exosomes in cancer development, metastasis, and drug resistance: a comprehensive review. Cancer Metastasis Rev. 2013, 32, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Shen, H.; Yin, X.; Yang, M.; Wei, H.; Chen, Q.; Feng, F.; Liu, Y.; Xu, W.; Li, Y. Macrophages derived exosomes deliver miR-223 to epithelial ovarian cancer cells to elicit a chemoresistant phenotype. J. Exp. Clin. Cancer Res. 2019, 38, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Lu, W.; Qu, J.; Ye, L.; Du, G.; Wan, X. Loss of exosomal miR-148b from cancer-associated fibroblasts promotes endometrial cancer cell invasion and cancer metastasis. J. Cell. Physiol. 2018, 234, 2943–2953. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, Y.; Liu, H.; Shen, W. , Extracellular vesicle encapsulated microRNA-320a inhibits endometrial cancer by suppression of the HIF1alpha/VEGFA axis. Exp Cell Res 2020, 394, 112113. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



