Submitted:
24 June 2024
Posted:
26 June 2024
You are already at the latest version
Abstract
Carbohydrates and lipids integrate into a complex metabolic network that is essential to maintain homeostasis. In insects, as in most metazoans, dietary carbohydrates are taken up as monosaccharides whose excess is toxic, even at relatively low concentrations. To cope with this toxicity, monosaccharides are stored either as glycogen or neutral lipids, the latter constituting a quasi-unlimited energy store. Breakdown of these stores in response to energy demand depends on insect species and on several physiological parameters. In this chapter, we review the multiple metabolic pathways and strategies linking carbohydrates and lipids that insects utilize to respond to nutrient availability, food scarcity or physiological activities.
Keywords:
Monosaccharides
; Glycolysis
; Lipogenesis
; Triacylglycerol
; Midgut
; Fat body
; Oenocytes
; Muscles
; Haemolymph
; Homeostasis
9.1. Introduction
Insects colonize a huge variety of ecological niches on Earth. Despite a great diversity of diets, their food always comprises carbohydrate biomolecules. Carbohydrates are organic compounds containing carbon, oxygen and hydrogen atoms, most frequently with a 2:1 hydrogen-oxygen ratio. They are divided into three main classes ranging from monosaccharide and disaccharide simple sugars (e.g. glucose, maltose) to complex polymers (e.g. glycogen, starch) and glycoconjugates (e.g. glycolipids) (Chandel, 2022). The proportion of each class of dietary carbohydrates depends on the insect feeding source. Since carbohydrates are commonly taken up by enterocytes as monosaccharides, dietary oligosaccharides, polysaccharides and glycoconjugates, they require enzymatic hydrolysis in the midgut (Terra and Ferreira, 2012; Chapman et al., 2013; Sahaka et al., 2020). In insects, the major monosaccharides resulting from enzymatic digestion in the gut lumen are glucose, fructose and galactose; whereas trehalose is the most abundant circulating sugar in the haemolymph (Wyatt and Kale, 1957). Trehalose is a nonreductive disaccharide linking two glucose units in a 1,1-glycosidic bound, which impedes the reductive toxicity of glucose (Mattila and Hietakangas, 2017).
Monosaccharides and their metabolic byproducts are toxic compounds, even at low concentrations (Rabbani and Thornalley, 2013). To prevent this toxicity once taken up into animal cells, glucose is catabolized through glycolysis to produce energy or stored as glycogen granules. However, the capacity for glycogen storage is limited, so that additional storage strategies have been selected throughout evolution to provide energy resources in case of food scarcity (Steele, 1982). Lipogenesis leading to the production of triacylglycerols (TAGs) is directly connected to carbohydrate catabolism through a metabolic axis comprising successively glycolysis, tricarboxylic acid (TCA) cycle and fatty acid (FA) synthesis. TAG storage in lipid droplets is, to a certain extent, unlimited, making it the essential strategy for energy storage (Brookheart et al., 2009; Patel and Abate, 2013). In contrast, glycolipids do not constitute an energy storage form; they are mainly found at the extracellular membrane layer, playing critical roles in numerous cellular functions (Nishihara, 2020). Cholesterol is critical for membrane fluidity and steroid hormone synthesis. Cholesterol biogenesis requires the condensation of two farnesylpyrophosphate units. Insects lack the enzymes for farnesylpyrophosphate condensation to sterol synthesis (Zhang et al., 2019). Therefore, cholesterol is not connected to carbohydrate metabolism and must be dietary provided (Clark and Block, 1959).
Lipid catabolism is not expected to reverse to carbohydrate biogenesis; thus, the carbohydrate to lipid metabolic axis is considered unidirectional. The fate of glucose to energy production or to glycogen/TAG storage is tightly controlled, involving the Mondo/Mlx transcription factor that responds to glucose metabolites, thereby regulating several metabolic routes, including FA synthesis (Mattila and Hietakangas, 2017; Richards et al., 2017; Havula and Hietakangas, 2018). In this chapter, we will focus on the physiological links that connect the carbohydrate to lipid metabolism at the whole insect body level. To this end, we will first summarize the metabolic routes from dietary carbohydrates to lipid synthesis and storage. Next, we will describe the roles of the various organs in these processes. Finally, we will discuss the relevance of this vectorial metabolic axis in maintaining energy homeostasis.
9.2. The Metabolic Network Linking Carbohydrates to Lipids
9.2.1. Monosaccharide Fates
Monosaccharides do not accumulate in the intracellular compartment. They are either stored as glycogen or metabolized through catabolic pathways, including glycolysis or the pentose-phosphate pathway (Figure 1).
Glycogen is a polymer of glucose units linked by α(1→4) bonds, forming chains branched by α(1→6) bonds (Adeva-Andany et al., 2016a; Chandel, 2022). Glycogen synthesis comprises the intermediates glucose-1-phosphate (glucose-1P) and UDP-glucose, and sequentially requires phosphoglucomutase, UDP-glucose phosphorylase, glycogen synthase and a branching enzyme (Figure 1). Breakdown of glycogen is catalyzed in the cytoplasm by glycogen phosphorylase and a debranching enzyme, and in lysosome by an acid α-glucosidase (Roach et al., 2012; Adeva-Andany et al., 2016a; Chandel, 2022).
Glycolysis first proceeds in a preparatory phase that consumes two ATP units, leading to the formation of the trioses glyceraldehyde-3phosphate (GA3P) and dihydroxyacetone-phosphate (DHAP) (Figure 1). In the glycolysis payoff phase, each GA3P unit is oxidized to pyruvate, yielding one NADH and two ATP molecules.
The pentose-phosphate pathway (PPP) (Figure 1) comprises a first oxidative phase, where glucose-6P is oxidized to produce NADPH and ribose-5P and a second non-oxidative phase that regenerates a pool of glucose-6P (Horecker, 2002).
Fructose can be converted to glucose through the intermediate sorbitol (Figure 1) (Krause and Wegner, 2020). A few studies reveal that sorbitol plays a role in immune response, in learning and in the seasonal physiological turnover of overwintering insects (Yang et al., 2019; Sano et al., 2022; Weiglein et al., 2019; Mustard et al., 2018; Drahun et al., 2023). Nonetheless, the relevance of sorbitol in the fructose to glucose conversion to feed glycolysis has been sparsely evaluated in insects. To enter glycolysis (Figure 1), fructose is phosphorylated either by a hexokinase to fructose-6P or by a ketohexokinase to fructose-1P (Johnson et al., 2020). The former enters glycolysis, whereas the latter is cleaved in DAHP and glyceraldehyde (GA), which is phosphorylated to GA3P by a triose kinase (Figure 1). Phosphorylation of fructose-6P to fructose-1,6-bisphosphate by PFK1 is the rate-limiting step of glucose consumption through glycolysis (Mor et al., 2011). The fructose-1P pathway bypasses this key regulatory step and thus enhances lipogenesis, thereby, favoring obesity and metabolic syndrome in human patients (Krause and Wegner, 2020).
The Leloir pathway is the main galactose metabolic pathway leading to the formation of UDP-galactose and glucose-1P, thereby i) contributing to glycogen synthesis, ii) entering glycolysis or iii) initiating the hexosamine pathway required for glycoproteins and glycolipids biosynthesis (Adeva-Andany et al., 2016a; Daenzer and Fridovich-Keil, 2017) (Figure 1). Nonetheless, the glycolytic metabolism of galactose is slower compared to that of glucose and fructose (Chandel, 2022).
The hexosamine pathway (Figure 1) starts with the rate-limiting step catalyzed by glutamine fructose 6-P amidotransferase (GFAT) to finally results in the production of UDP-N-acetylglucosamine (Chandel, 2022). The UDP-galactose epimerase (GALE) catalyzes the reversible interconversion of UDP-galactose to UDP-glucose, but also of UDP-N-acetylglucosamine to N-acetylgalactosamine, all these sugar derivatives being potential substrates for lipid glycosylation (Daenzer and Fridovich-Keil, 2017).
Trehalose synthesis results from the condensation of glucose-6-phosphate and UDP-glucose to trehalose-6-phosphate catalyzed by trehalose phosphate synthase and its dephosphorylation by trehalose phosphatase (Thompson, 2003).
9.2.2. Glycolytic Products and Byproducts
The payoff phase of glycolysis results in the production of one NADH and two ATP molecules through the conversion of each GA3P unit into pyruvate (Figure 2). In the cytoplasm, the reversible conversion of one pyruvate to one lactate molecule is catalyzed by the lactate dehydrogenase (LDH), which concurrently oxidizes one NADH coenzyme to NAD+. In the mitochondria, the oxidation of pyruvate to acetyl-CoA by the pyruvate dehydrogenase enzymatic complex is coupled to the reduction of NAD+ to NADH. Acetyl-CoA reacts with oxaloacetate to form citrate, thereby initiating the TCA cycle. Citrate oxidation to oxaloacetate proceeds through seven enzymatic reactions coupled to the formation of one FAD and three NADH units. Regeneration of these coenzymes by the respiratory chains at the mitochondrial inner membrane yields high amounts of ATP (Towarnicki and Ballard, 2020).
The lactate resulting from anaerobic fermentation in muscles is secreted and can be taken up by other tissues. For instance, in the mammalian liver, lactate from muscles is converted by LDH to pyruvate that is carboxylated to oxaloacetate by pyruvate carboxylase to fuel gluconeogenesis (Adeva-Andany et al., 2016b). To initiate gluconeogenesis, the conversion of oxaloacetate to phosphoenolpyruvate by the phosphoenolpyruvate carboxykinase (PEPCK) bypasses the irreversible phosphoenolpyruvate to pyruvate final reaction of glycolysis. When initiated from TCA cycle intermediates, gluconeogenesis depletes oxaloacetate, which must be compensated by anaplerotic reactions. Anaplerosis is commonly considered to depend on amino acid metabolism, in particular via transaminase-catalyzed reactions that convert aspartate to oxaloacetate and glutamate to α-ketoglutarate (Brooks, 1987; Inigo et al., 2021).
The trioses DHAP and GA3P can be interconverted by the triose phosphate isomerase (Figure 2). GA3P will fuel glycolysis, whereas DHAP can be reduced to G3P (glycerol 3-phosphate) that is used for TAG biogenesis through the Kennedy pathway (Coleman and Mashek, 2011; Ko et al., 2020). In addition, DHAP and GA3P may spontaneously be oxidized to methylglyoxal (MG), which is an α-oxoaldehyde that strongly reacts with the amine groups of proteins, DNA or nucleotides to form advanced-glycation-end-products (AGEs). Several studies support the notion that AGEs ─in particular, glycated proteins that become resistant to proteasome degradation─ are responsible for the toxic effects of sugars (Annandale et al., 2021). Nonetheless, to prevent this toxicity, MG can be metabolized to lactate by the glyoxalase system composed of the glyoxalase 1 (Glo1) and glyoxalase 2 enzymes (Thornalley, 1993) (Figure 2).
9.2.3. Lipid Metabolism
Excess of citrate from the TCA cycle is transferred back to the cytoplasm, where it is converted to oxaloacetate and acetyl-CoA; the latter is used for FA synthesis that proceeds in two main enzymatic steps (Figure 2). First, Acetyl-CoA-Carboxylase (ACC) catalyzes the carboxylation of acetyl-CoA into malonyl-CoA that is the rate-limiting step for FA synthesis (Barber et al., 2005). Next, fatty acid synthase (FASN) sequentially incorporates malonyl-CoA units to one acetyl-CoA primer to produce long chain fatty acids (LCFAs) (Smith et al., 2003; Maier et al., 2010). The biochemical reactions catalyzed by FASN require two NADPH coenzymes for incorporation of each malonyl-CoA unit (Figure 2). The NADPH may be synthesized through i) the pentose phosphate pathway, ii) the reduction of malate or iii) the one-carbon metabolism (Fan et al., 2014).
Newly synthesized LCFA are used for the synthesis of phospholipids or stored as TAGs in lipid droplets (Coleman and Mashek, 2011; Ko et al., 2020) (Figure 3). Synthesis of TAGs that may proceed through three distinct pathways, as well as phospholipid biogenesis are described in chapter 6. In the context of energy demand, the breakdown of TAGs releases LCFAs used for mitochondrial β-oxidation. The resulting acetyl-CoA either feeds the TCA cycle or is converted to ketone bodies that efflux into the circulatory system or the insect haemolymph to sustain the energy demand of other organs (Bailey and Horne, 1972; Huang et al., 2019). Nonetheless, this catabolic route does not happen in the context of FA synthesis, since malonyl-CoA produced by ACC inhibits the CPT-1 mediated transfer of LCFAs into the mitochondria (Saggerson, 2008). Importantly, β-oxidation of even-chain LCFA cannot fuel gluconeogenesis but rather fosters anaplerotic reactions to increase the oxaloacetate pool required to metabolize the resulting high amounts of acetyl-CoA. Even in the case of β-oxidation of odd-chain LCFA, whose final product propionyl-CoA can be converted into succinyl-CoA (Inigo et al., 2021; Fogle et al., 2019), the need in oxaloacetate substrates to metabolize acetyl-CoA impedes gluconeogenesis. Therefore, the metabolic vectorial axis from carbohydrate catabolism to lipid synthesis via the propionyl-CoA resulting from odd-chain LCFA oxidation is unlikely to reverse to carbohydrate biosynthesis.
Glycolipids are cell membrane components acting in signaling. Their synthesis is catalyzed by glycosyltransferases that covalently attach sugar moieties to the hydroxyl group of either a glycerolipid or a sphingolipid backbone. Glycolipid synthesis takes place at the Golgi apparatus, so that glycolipids are presented at the cell surface exposing their sugar moiety in the extracellular milieu (Nishihara, 2020).
9.3. Major Organs Implicated
9.3.1. Digestive Tract
The type of dietary carbohydrates directly depends on the insect feeding source (Table 1) (Patrick et al., 2013). Given that monosaccharides are the main form of sugar absorbed by the enterocytes, dietary oligosaccharides, polysaccharides and glycoconjugates require enzymatic hydrolysis of glycosylic bonds in the midgut lumen prior to intestinal uptake. A number of carbohydrase activities have been identified in the salivary glands and in the midgut of several phytophagous bugs, suggesting that carbohydrate hydrolysis starts upon food ingestion (Hori, 1975) or prior to it, as reported for external digestion of starch in Drosophila melanogaster (Haj-Ahmad and Hickey, 1982; Boer and Hickey, 1986; Szyszka and Galizia, 2018). Insect enterocytes synthesize amylases that hydrolyze starch and glycogen, and maltases that hydrolyze maltose (Benkel and Hickey, 1986; Terra and Ferreira, 1994; Zinke et al., 2002; Mattila et al., 2015; Pimentel et al., 2018). Conversely, cellulose and pectin digestion relies in most species on enzymes synthesized by enteric microorganisms (Watanabe and Tokuda, 2010; Ransom-Jones et al., 2012; Calderon-Cortes et al., 2012). Release of sugar moieties from glycolipids involves specific hydrolases secreted by the enterocytes, as reported for the folivorous lepidopteran Epiphyas postvittana and Helicoverpa armigera (Christeller et al., 2011; Sahaka et al., 2020). The origin of a given carbohydrate hydrolase cannot, however, be formally assigned to the genome of either the enterocyte or the microbiota, but varies depending on the insect species. For instance, cellulases and/or pectinases have been reported as encoded by the genome of the termite Reticulitermes speratus, the hemipteran Lygus lineolaris and the coleopteran Phaedon cochleariae, Tribolium castaneum, and Sphenophorus levis (Watanabe et al., 1998; Girard and Jouanin, 1999; Allen and Mertens, 2008; Willis et al., 2011; Evangelista et al., 2015).
The physiology of aphids soaking up an unbalanced diet from plant phloem, containing high amounts of carbohydrates ─mostly sucrose─ with low amino acid and lipid levels, illustrates the tight metabolic link between carbohydrates and lipids through a unidirectional axis. It has been shown in the sap exclusive feeder Acyrthosiphon pisum that most of the dietary sucrose is used for lipid synthesis, mostly neutral lipids (Febvay et al., 1992). Consequently, TAGs are in high levels in the fat body but also in cornicle secretion that contains as well high amounts of sugars (Rahbe et al., 1994). Their midgut symbionts are essential for their growth and their reproductive functions. However, antibiotic-induced depletion of their intestinal symbionts affects amino acid and glycogen synthesis but conversely increases lipid levels (Febvay et al., 1999; Lv et al., 2018), suggesting that these symbionts are required for amino acid but not lipid metabolism. The respective contributions of midgut and fat body cells in this lipogenic process in aphids have not been addressed to date.
Sugar transporters are members of the solute carrier proteins, including the major facilitators GLUT/SLC2, the sodium-driven glucose symporters SGLTs/SLC5 and the plant-specific SWEET/SLC50 (Deng and Yan, 2016). In mammalian enterocytes, glucose uptake proceeds through SGLT1 located at the brush border, whereas passive reversible export at the basal membrane mainly depends on GLUT2 for glucose and galactose and on GLUT5 for fructose (Wright et al., 2007; Gonzalez and Betts, 2019; Taskinen et al., 2019). In insects, the midgut is the main organ where monosaccharide absorption takes place, although this may also occur at the epidermis of endophagous parasitoids as reported for the larvae of the wasp Aphidius ervi (Caccia et al., 2005). Genomic analyses led to the identification of sugar transporters in several insect species (Kikuta et al., 2015; Moreira et al., 2017; Govindaraj et al., 2016; Pimentel et al., 2018; Wang and Wang, 2020). Utilization of chemical inhibitors revealed a powerful approach to confirm sugar transporter specificity. In this way, expression of a potential hexose transporter (NIHT1) has been reported in the midgut of the hemipteran Nilaparvata lugens that feeds on sucrose-enriched phloem sap (Price et al., 2007). NIHT1 expression in vesicles mediates glucose but not fructose transport. This transport is inhibited by cytochalasin B, suggesting that NIHT1 is a functional GLUT2 homologue. Homologues of GLUTs and SGLTs have been found in the anterior midgut of Dysdercus peruvianus (Bifano et al., 2010). In this hemipteran, glucose transport is activated by K2SO4 and inhibited by the GLUT inhibitor phloretin and by the SGLT inhibitor phlorizin, supporting that GLUT- and SGLT-like functions are conserved in insects. In D. melanogaster, the soluble carrier dSLC5A5 has been shown to be the bona fide SGLT1 homologue required for apical endocytosis into the enterocytes (Li et al., 2021). Expression of a trehalose transporter (TreT) has been found in the midgut of Anopheles gambiae and Musca domestica (Liu et al., 2013; Pimentel et al., 2018). However, given the high luminal trehalase levels (Becker et al., 1996) TreT is unlikely fulfilling the uptake of dietary trehalose, but rather buffering haemolymph trehalose levels to maintain homeostasis.
In mammals, the classical view is that nutrients taken up from the intestinal lumen are directly routed to the liver, which is the major site of intermediary metabolism. In insect midgut cells, monosaccharides resulting from carbohydrate hydrolysis can either efflux into the haemolymph or be converted to FAs via neolipogenesis as reported in Pieris brassicae and D. melanogaster (Turunen, 1993; Kokki et al., 2021). In the enterocytes, FAs are transiently esterified to TAGs and stored as lipid droplets, from where they are progressively exported in the form of diacylglycerol (DAG) loaded lipoproteins through the haemolymph to the requiring organs (Ford and Van Heusden, 1994; Pennington and Wells, 2002; Grillo et al., 2003; Palm et al., 2012; Heier and Kuhnlein, 2018).
9.3.2. Fat Body and Oenocytes
Fat body cells are in tight contact with oenocytes in a number of insect species. The fat body is an insect organ that combines hepatic and adipose functions related to energy homeostasis, nutrient storage and detoxification (Arrese and Soulages, 2010; Li et al., 2019). In contrast, the oenocytes fulfil catabolic hepatic-like functions related to starvation (Gutierrez et al., 2007; Moraes and Montagne, 2021; Huang et al., 2022). So far, investigations of the lipid anabolic pathways in the oenocytes of D. melanogaster did not reveal hepatic-like functions; instead, oenocytes produce FAs required to sustain the production of cuticular hydrocarbons (Wicker-Thomas et al., 2015; Storelli et al., 2019; Montagne and Wicker-Thomas, 2020; Blomquist and Ginzel, 2021; Huang et al., 2022) and to maintain tracheal watertightness (Parvy et al., 2012).
The fat body accumulates high amounts of TAGs in the form of lipid droplets, which may involve import of dietary lipids or neolipogenesis. Whether the fat body is solely a storage organ or also contributes to intermediary metabolism has been addressed in a number of studies. Incorporation of dietary radiolabeled lipids in the fat fraction of the fat body of the lepidopteran P. brassicae and of the cockroach Periplaneta americana has been reported (Turunen, 1975; Turunen and Kastari, 1979; Chino and Downer, 1979). Conversely, the fat body of the locusts Schistocerca gregaria and Locusta migratoria has been shown to perform intermediary metabolisms, including TCA cycle, respiration and fatty acid synthesis and to incorporate radiolabeled elements from glucose and amino acids into TAG-containing lipid droplets (Clements, 1959; Tietz, 1961; Hines and Smith, 1963). These studies indicate that acyl groups of the TAG stores may have different origins (Coleman, 2019; Olzmann and Carvalho, 2019; Heier et al., 2021). TAG synthesis through the Kennedy pathway begins with the acylation of a glycerol-3-phosphate backbone in mitochondria or endoplasmic reticulum (ER), whereas the dihydroxyacetone-phosphate (DHAP) pathway begins with DHAP acylation in the peroxisome. Both pathways result in the production of lysophosphatidic acid, which after two acylation reactions in the ER leads successively to phosphatidic acid, DAG and TAG. Acylation of monoacylglycerol (MAG) in the ER is a third pathway for TAG biogenesis. A fourth pathway, which also takes place in the ER, is the transacylation of two DAG units that results in the formation of one TAG and one MAG molecules; the latter may follow the MAG pathway to TAG. In sum, any of these pathways requires additional FA units (Figure 3). The TAG biogenesis pathway that takes place in the fat body has been sparsely addressed and likely depends on the insect species and/or its specific diet type (Kastari and Turunen, 1977). For instance, in Rhodnius prolixus, the Kennedy pathway has been shown to be the only pathway for TAG synthesis (Alves-Bezerra and Gondim, 2012). Nonetheless, given that in most species DAGs but not TAGs are transferred from the midgut to the fat body through the haemolymph, FA synthesis must operate in the fat body to sustain TAG biogenesis. Consistently, in D. melanogaster, fat body knockdown of ACC or FASN results in an almost complete depletion of overall TAGs and of fat body lipid droplets (Parvy et al., 2012; Garrido et al., 2015) and incorporation of high amounts of radiolabeled dietary glucose has been reported in total lipids (Musselman et al., 2013).
Glycogen has been reported to accumulate in the fat body of several insect species, including S. gregaria, Bombyx mori, P.american, Culex pipiens and D. melanogaster (Clements, 1959; Inagaki and Yamashita, 1986; Veenstra, 1989; King et al., 2020; Garrido et al., 2015; Yamada et al., 2018; Yamada et al., 2019). The fat body also synthesizes and accumulates trehalose and is considered to be the only organ where this synthesis takes place (Thompson, 2003; Mattila and Hietakangas, 2017). High levels of trehalose synthesis and the expression of trehalose-6-phosphate synthase have been reported in the fat body of B. mori, L. migratoria and Mythimna separata (Murphy and Wyatt, 1965; Agbanyo and Taylor, 1986; Yang et al., 2023). In D. melanogaster, starvation-induced glycogen breakdown of fat body stores is crucial to maintain homeostasis of circulating trehalose (Yamada et al., 2018).
In the context of starvation, TAGs are hydrolyzed in glycerol and FAs to feed β–oxidation, which ends up with the formation of acetyl-CoA units. Acetyl-CoA may feed the TCA cycle or generate ketone bodies, which are transferred to the requiring organs (Bailey and Horne, 1972). Studies in D. melanogaster suggest that the oenocytes perform β–oxidation upon fasting, as do mammalian hepatocytes that accumulate lipid droplets after the remobilization of adipocyte lipid stores. Likewise, fasting or genetically-provoked breakdown of fat body TAGs induces lipid droplet accumulation in the oenocytes (Gutierrez et al., 2007). Congruently, enzymes responsible for ketone body formation are highly expressed in the oenocytes of D. melanogaster (Huang et al., 2019). Oenocyte-ablated D. melanogaster larvae are almost unable to burn their TAG stores in the context of fasting, which might suggest that the oenocyte are the only organs to perform FA β–oxidation (Gutierrez et al., 2007). Nonetheless, since the oenocytes produce a FA required for the watertightness of the tracheal system, oenocyte-ablated D. melanogaster larvae are in anoxic condition, and thereby cannot perform FA β–oxidation (Parvy et al., 2012). Of note, FA consumption through β–oxidation has been reported in the flight muscles of a number of lepidopteran and orthopteran species (see below). Therefore, it is unlikely that the oenocytes are the only organs able to perform FA β–oxidation in the context of starvation and the respective contributions of the fat body and of the muscles remain to be precisely established.
9.3.3. Muscles
Muscles are necessary for locomotion dispersion in the environment (skeletal muscles) or for internal physiological functions (peristalsis of intestinal visceral muscles; heart activity). The metabolic pathway to provide energy depends on the species and on the muscle type. Metabolism of visceral muscles has been sparsely studied in insects. In D. melanogaster a study focusing on the energy sensor AMP-activated protein kinase (AMPK) revealed that it is essential for intestinal peristalsis and that its mutation impaired nutrient absorption and glucose incorporation in glycogen and neutral lipids (Bland et al., 2010). Conversely, muscle metabolism has been extensively studied in insect skeletal flight muscles, which are the most energy demanding per tissue weight (Wegener, 1996). The immediate on/off state transition at flight onset requires a rapid and efficient production of energy, which is considered to rely on carbohydrate consumption through glycolysis (Auerswald and G, 1995; Canavoso et al., 2003; Suarez et al., 2005; Zhang et al., 2011; Gaviraghi et al., 2019). However, proline oxidation by the proline dehydrogenase at the mitochondrial respiratory chains is proposed to be a sparker metabolite at flight onset in several insect species (Bursell, 1981; Auerswald and G, 1995; Gade and Auerswald, 2002; Soares et al., 2015; Gaviraghi et al., 2019; Stec et al., 2021).
Glucose may originate from muscle glycogen stores, whose breakdown is catalyzed by the debranching enzyme AGL and by the glycogen phosphorylase that releases glucose-1P units (Childress and Sacktor, 1970; Wegener, 1996; Yamada et al., 2018). Glycogen storage and glycogen phosphorylase have been described in the muscles of numerous insect species (Childress et al., 1970; Downer and Matthews, 1976; Turturro and Shafiq, 1979; Steele, 1982; Lorenz, 2007; Zhang et al., 2011; Garrido et al., 2015; Yamada et al., 2019). Remobilization of glycogen from the fat body resulting in the synthesis of trehalose, which is transported through the haemolymph is the other major source of glycolytic substrate in muscles after trehalose uptake and Trehalase hydrolysis to glucose (Kanamori et al., 2010; Thompson, 2003; Murphy and Wyatt, 1965; Agbanyo and Taylor, 1986; Shukla et al., 2015; Mattila and Hietakangas, 2017; Yang et al., 2023). Consistently, glycogen phosphorylase has been reported in the fat body of several insect species including D. melanogaster and the cockroaches Nauphoeta cinerea and P.americana (Steele, 1982; Gade, 1991; Yamada et al., 2018; Yamada et al., 2019).
Glucose consumption through glycolysis produces at the payoff phase a pool of reduced NADH that can be regenerated to NAD+ in the anaerobic reduction of pyruvate to lactate (Figure 4). Slower working skeletal muscles(e.g. leg muscles) may oxidize NADH to NAD+ by pyruvate to lactate, as described in the jumping muscles of Schistocerca americana (Kirkton and Tyler, 2021). Conversely, the high glycolytic rate of flight muscles would produce dramatic amounts of lactate that might be toxic. To cope with this deleterious effect, flight muscles consume pyruvate through the TCA cycle, resulting in the lack of NAD+ to sustain the payoff phase of glycolysis (Figure 4). The fastest metabolic pathway to regenerate the pool of NAD+ proceeds through the G3P shuttle (Mracek et al., 2013; Gaviraghi et al., 2019). G3P is produced from DHAP reduction oxidizing NADH to NAD+. While G3P is a precursor for TAG synthesis through the Kennedy pathway in fat body cells, in muscles, it is regenerated at the mitochondrial inner membrane to DHAP by reducing the FAD coenzyme, which subsequently results in ATP production by the respiratory chains. Therefore, the G3P shuttle in flight muscle cells reduces the cytosolic pool of G3P, which would have been used for TAG or phospholipid synthesis in other tissues. Interestingly, a study on the bumblebee Bombus terrestris, which can fly at low ambient temperatures, suggests that the G3P shuttle plays a thermogenic role in pre-warming flight muscles (Masson et al., 2017).
The production of energy for flight muscles relies on carbohydrates for short-term and medium-term flights in several insect species (Beenakkers, 1969), but may also depend on lipid consumption (Li et al., 2023), as reported for Manduca sexta, L. migratoria and Gryllus bimaculatus (Ziegler and Schulz, 1986; van der Horst et al., 1993; Lorenz, 2007) (Table 2). Given that neutral lipids represent an enormous energy store, consumption of remobilized lipids represents an advantage for the long-term flight of non-feeding adult insects (Anand and Lorenz, 2008). In L. migratoria, carbohydrate is the predominant energy source during the first 30 minutes of flight, while lipid remobilization from fat body stores in the form of circulating DAGs progressively increases to become the major energy source for the following 30 minutes of flight (Jutsum and Goldsworthy, 1976). In Panstrongylus megistus, haemolymph carbohydrates and muscle glycogen progressively drop during 45 minutes flight, while fat body glycogen slightly decreases, suggesting that the latter do not sustain energy flight. In contrast, between 15 and 45 minutes of flight, lipid stores from muscles and fat body strongly decrease, while those in the haemolymph increase, indicating that lipids become the predominant energy source during extended flight duration (Canavoso et al., 2003).
Studies to identify the mechanism of store remobilization to provide energy to flight muscles revealed the key role of neuropeptides (Li et al., 2023), including the functional glucagon homologue adipokinetic hormone (AKH). AKHs are small peptides synthesized in the corpora cardiaca and transferred through the haemolymph to the fat body, where they bind their cognate G-protein-coupled receptors (Gade and Auerswald, 2003; Gade, 2009). AKH-dependent remobilization is not store specific. In L. migratoria, AKH controls the remobilization of lipid and carbohydrate stores from the fat body to feed flight muscle activity (Robinson and Goldsworthy, 1977). In M. sexta, AKH controls lipid mobilization to sustain adult flight, but glycogen phosphorylase in starving larvae (Ziegler et al., 1990). In D. melanogaster, AKH deficiency results in lipid consumption defects and obese flies, which surprisingly are not flight deficient, although this study assayed only the immediate flying performance, but not long-term flying ability (Galikova et al., 2015). In A. gambia injection of a synthetic AKH increases flight performances and haemolymph carbohydrate but not lipids levels (Kaufmann and Brown, 2008). In several beetle species that use proline to pre-warm flight muscles but also for flight, AKH controls haemolymph proline levels (Gade and Auerswald, 2002). Likely because of its role in remobilizing fat body stores, a number of studies focused on the fat body AKH receptor, although it has been detected in a number of distinct cell types (Galikova and Klepsatel, 2023). In R. prolixus, the AKH receptor is highly expressed in fat body and flight muscles, and its knockdown results in lipid accumulation in both organs (Alves-Bezerra et al., 2016). In sum, the insulin/glucagon balance to control homeostatic storage and remobilization of carbohydrates and lipids is tightly conserved in insects (Chowanski et al., 2021) (see also chapter 6).
9.4. Integration of Carbohydrate/Lipid Metabolism at the Body Level
9.4.1. Carbohydrate and Lipid Homeostasis
Systemic regulators of metabolic homeostasis conserved throughout evolution have been identified in insects. In feeding state, dietary sugar and amino acids act through the fat body to trigger insulin-like peptide (ILP) secretion from neurosecretory cells, which in turn mediate several aspects of body growth and metabolic homeostatic response, in particular sugar storage in the form of glycogen and TAGs (Wu and Brown, 2006; Toprak, 2020; Chowanski et al., 2021). Conversely, in case of high energy demand, neuropeptides such as AKH, AKH/Corazonin-related peptide and octopamine stimulate store breakdown to maintain homeostasis (Oguri and Steele, 2003; Roeder, 2005; Michitsch and Steele, 2008; Zhou et al., 2018; Toprak, 2020; Galikova and Klepsatel, 2023). Neutral lipids are always the major components of fat body stores. However, the TAG/glycogen storage ratio as well as their consumption depends on the insect species and on several parameters, including behavior, physiological status, aging and energy level requirement (Lorenz and Anand, 2004; Anand and Lorenz, 2008; Wang et al., 2016). This section focuses on the relationships between carbohydrate and lipid metabolism per se, whereas the mechanisms of endocrine regulation of lipid metabolism are extensively documented in other chapters.
A number of insect-based studies emphasize the metabolic links between carbohydrates and lipids. As mentioned above, this metabolic link is nicely illustrated by the phloem exclusive feeder A. pisum, producing lipids in excess that accumulate in fat body but also in cornicle secretions (Febvay et al., 1992; Rahbe et al., 1994). In R. prolixus, it has been shown that deficiency in insulin receptor signaling strongly drops TAG synthesis in the fat body after blood meal (Silva-Oliveira et al., 2021). In Blattella germanica, high sucrose diet increases the expression of ACC ─the rate-limiting enzyme of fatty acid synthesis─, while ACC-knockdown decreases TAG and cuticular hydrocarbons levels, and increases circulating sugar and glycogen storage (Pei et al., 2023). It has been shown in P. americana that FAs stimulate trehalose synthesis in isolated FB cells and that trehalose efflux into the haemolymph depends on arachidonic acid-derived metabolites (Ali et al., 1998; Ali and Steele, 1997). Furthermore, a co-dependence in lipid and glycogen synthesis has been reported in C. pipiens where inhibition of either results in decreased biogenesis of both glycogen and TAGs (Olademehin et al., 2020). In Aedes aegypti females, where lipid stores are mobilized for egg production, a constant carbohydrate dietary source is required to sustain the accomplishment of the gonotrophic cycles (Briegel et al., 2002; Zhou et al., 2004). In Grapholita molesta, while flight muscle energy relies on carbohydrate consumption, lipid stores constitute the main energy source in starvation condition(s) (Su et al., 2022). The flying morph of Gryllus firmus female consumes carbohydrates for flight, while TAG consumption is likely related to reproductive behavior in the non-flying reproductive morph (Zhang et al., 2011). A metabolic-based conflict between flight ability and egg production has been reported in several insect species. For instance, in G. bimaculatus adult females, carbohydrate and lipid stores are directed to egg production, while flight muscles are concurrently hydrolyzed (Lorenz, 2007). A screening-based study in D. melanogaster identified sirtuin 1 as a gene whose mutation results in fat accumulation but improved survival on a feeding media depleted in amino acid but supplemented with sugar (Reis et al., 2010). Another study investigating the relationship between dietary sugar and fat storage revealed that fructose was a more potent obesity-inducer than glucose (Rovenko et al., 2015). This effect likely results from the fructose-1P glycolytic route (§9.2.1), which bypasses the PFK1 rate-limiting reaction of glycolysis (Mor et al., 2011; Krause and Wegner, 2020). Dietary sugar excesses are harmful in humans but also in insects. In D. melanogaster, high sugar diet (HSD) has been shown to provoke fat accumulation in the cardiac tube and heart dysfunction, which in part depends on hexosamine metabolism (Na et al., 2013). These studies strengthen the notion that carbohydrate and lipid metabolisms are inseparable processes to maintain homeostasis and might suggest that high amounts of dietary glucose are directly routed to TAG storage through the glycolytic/TCA/Lipogenic axis. However, a D. melanogaster study indicates that under chronic HSD, mitochondrial acetyl-coA could be recycled through an anaplerotic/gluconeogenic pathway before being re-metabolized and incorporated into TAGs (Musselman et al., 2013). Further, a study focusing on muscle ACC in D. melanogaster supports that life-span extension upon diet restriction depends on a recycling loop of fat synthesis/oxidation (Katewa et al., 2012). In sum, the carbohydrate/lipid connection does not obligatory follow a unique direct axis but likely integrates within a complex metabolic network.
9.4.2. Sugar Diet and Type 2 Diabetic-Like Syndrome in D. melanogaster
The D. melanogaster model has been of powerful help in investigating the harmful effect of HSD. The Target of rapamycin (TOR) signaling network plays a critical role in metabolic-related diseases (Saxton and Sabatini, 2017; Moraes and Montagne, 2021). TOR is present in two distinct complexes, TORC1 and TORC2, which operate via two parallel branches in D. melanogaster (Radimerski et al., 2002). TORC1 integrates nutrient availability through the tumor suppressor TSC and the small GTPase Rheb, while TORC2 resides in the insulin signaling branch that includes the tumor suppressor PTEN and the kinase AKT. The metabolism of D. melanogaster larvae over-activating either the TORC1 or the insulin response branch exhibits a significant decrease in overall glycogen and trehalose levels (Devilliers et al., 2021). Nonetheless, insulin- but not TORC1-induced cell-autonomous overgrowth is significantly decreased by glycolysis or FA synthesis defect, suggesting that alternative metabolic strategies operate for TORC1- but not insulin-dependent growth (Devilliers et al., 2021). In mammals, mTOR-dependent metabolic regulations are in part mediated by the NAD+-dependent deacetylases Sirtuins, as shown for SIRT4 (Csibi et al., 2013). In D. melanogaster, mutation of Sirt4 results in shorter lifespan and increased TAG stores; these mutants are sensitive to starvation and weakly consume their carbohydrate and lipid upon fating (Wood et al., 2018). In D. melanogaster, while amino acid controls the production of ilp2 (Colombani et al., 2003), circulating sugar promotes selective release of Dilp3 from insulin producing cells (Kim and Neufeld, 2015).
To characterize the genomic response to dietary sugar, a microarray analysis identified the sucrose-induced transcription factor encoded by sugarbabe and showed that it can repress genes required for dietary lipid absorption and fat breakdown (Zinke et al., 2002). Further studies revealed that HSD fed larvae decrease food intake, were hyperglycemic and developmentally delayed compared to larvae fed control diet. HSD-fed larvae accumulate more TAGs, express higher ILPs both at mRNA and protein levels and exhibit decreased insulin response (Musselman et al., 2011; Pasco and Leopold, 2012; Musselman et al., 2013). In addition, larvae fed chronic HSD up-regulate genes of glycolysis, lipogenesis and trehalose synthesis, but also of gluconeogenesis and FA β-oxidation. Interestingly, obesity, ketogenesis, insulin resistance and increased gluconeogenesis are typical hallmarks of type 2 diabetes (Hatting et al., 2018). Congruent with a type 2 diabetic-like syndrome (Back and Kaufman, 2012), genes involved in endoplasmic reticulum and oxidative stresses are also misregulated in larvae fed chronic HSD (Musselman et al., 2013).
The E-box transcription factor Mondo ─homologue to the mammalian carbohydrate responsive element binding protein (ChREBP)─ is up-regulated in HSD fed larvae (Musselman et al., 2011). In mammals, ChREBP activity is stimulated by glycolytic metabolites to direct glucose usage (Richards et al., 2017; Havula and Hietakangas, 2018). In D. melanogaster, Mondo knockdown as well as mutation of its partner encoded by Mlx, result in high sugar sensibility and in the down-regulation of glycolytic and lipogenic genes in the fat body (Havula et al., 2013; Musselman et al., 2013). Consequently, Mondo and Mlx deficient larvae exhibited an increase in haemolymph sugar and a decrease in TAG and phospholipid levels. Finally, it has been shown that in response to dietary sugar, the Mondo/Mlx complex stimulates glycolysis and lipogenesis but also the pentose phosphate pathway amino acid biosynthesis, while repressing glucose intake and gluconeogenesis (Bartok et al., 2015; Mattila et al., 2015). These regulatory processes in part depend on Sugarbabe and on the TGF-β/Activin ligand Dawdle, either of them being metabolic regulators. On one hand, Dawdle has been shown to control the expression of carbohydrases and lipases in the enterocytes (Chng et al., 2014) and Dawdle mutant larvae exhibit elevated haemolymph sugars, TAG and glycogen stores (Ghosh and O'Connor, 2014). On the other hand, over-expressing Sugarbabe in insulin producing cells represses ilp3 expression, which in turn results in overall elevated TAG levels (Varghese et al., 2010). Finally, Sugarbabe has been shown to reside within an ERK7/TOR/PWP1 signaling axis to control fat body adiposity, where ERK7 is an anti-anabolic atypical MAP kinase and PWP1 a chromatin binding protein (Hasygar et al., 2021). Taken together these studies demonstrate that Mondo and TOR signaling integrate in a complex network to regulate lipid metabolism in response to carbohydrate load.
9.4.3. Sugar Toxicity and Lipogenesis
The metabolic links between obesity and carbohydrate support the notion that lipogenesis protects against excess dietary sugar. This lipogenic-dependent protection is strongly supported by the phenotype induced by the mutation in the FASN1 gene that encodes the ubiquitous FASN enzyme in D. melanogaster (Garrido et al., 2015). These mutants are early larval lethal but can be rescued by an appropriate mixture of dietary lipids. These rescued mutants decrease their food intake and TAG stores and increase their glycogen stores and haemolymph trehalose, but fail to survive on sucrose-supplemented media. Surprisingly, this sensitivity to dietary sugar also operates at the cell-autonomous level, since FASN1 mutant clones in the fat body are strongly reduced in size in HSD- but not standard- fed larvae (Garrido et al., 2015; Devilliers et al., 2021). An increase in AGEs (§ 9.2.2) is directly associated with high levels of circulating sugar in type 2 diabetic patients and is responsible for the harmful effects (Liu et al., 2023). Methylglyoxal that formed spontaneously from the glycolytic trioses phosphate GA3P and DHAP (Figure 2) is a very active glycation reagent, which is detoxified by the glyoxalase complex (Rabbani and Thornalley, 2012). Genetic interaction between FASN1 mutation and glyoxalase1 knockdown in D. melanogaster revealed that the toxic effect of sugar also depends on α-oxoaldehyde reagents, both at the organismal and cell-autonomous levels (Garrido et al., 2015). In addition, mutants for glyoxalase 1 ─encoding the first enzyme of the glyoxalase complex─ are hyperglycemic, insulin resistant and exhibit elevated methylglyoxal levels and FASN1 activity (Moraru et al., 2018).
In conclusion, a critical function of lipogenesis is to protect against excesses of dietary carbohydrates. At first glance it may be surprising that the basic enzymatic pathway of FA synthesis is conserved throughout evolution suggesting that TAG synthesis is more important than the essential-FA metabolites for animal life. Therefore, it is tempting to surmise that the protection against dietary carbohydrates is more critical than the essential FA requirement. Evolution is canalized by environmental constraints. To survive to alternation of food availability and scarcity episodes, metabolic pathways have been selected in a way that carbohydrates, which are abundant but extremely toxic, can be stored in high amounts in the form of neutral lipids. Excess of food may induce metabolic syndrome and insulin resistance, in part because of lipotoxicity. However, the strong gene conservation of the glycolytic/TCA/lipogenic axis suggests that lipotoxicity is not as harmful as glucotoxicity.
References
- Adeva-Andany, M.M.; González-Lucán, M.; Donapetry-García, C.; Fernández-Fernández, C.; Ameneiros-Rodríguez, E. Glycogen metabolism in humans. BBA Clin. 2016, 5, 85–100. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Pérez-Felpete, N.; Fernández-Fernández, C.; Donapetry-García, C.; Pazos-García, C. Liver glucose metabolism in humans. Biosci. Rep. 2016, 36, e00416. [Google Scholar] [CrossRef]
- Agbanyo, M.; Taylor, N.F. Incorporation of 3-deoxy-3-fluoro-D-glucose into glycogen and trehalose in fat body and flight muscle in Locusta migratoria. Biosci. Rep. 1986, 6, 309–316. [Google Scholar] [CrossRef]
- Ali, I.; Finley, C.; Steele, J. Evidence for the participation of arachidonic acid metabolites in trehalose efflux from the hormone activated fat body of the cockroach (Periplaneta americana). J. Insect Physiol. 1998, 44, 1119–1126. [Google Scholar] [CrossRef]
- Ali, I.; Steele, J. Fatty Acids Stimulate Trehalose Synthesis in Trophocytes of the Cockroach (Periplaneta americana) Fat Body. Gen. Comp. Endocrinol. 1997, 108, 290–297. [Google Scholar] [CrossRef]
- Allen, M.L.; Mertens, J.A. Molecular Cloning and Expression of Three Polygalacturonase cDNAs from the Tarnished Plant Bug,Lygus lineolaris. J. Insect Sci. 2008, 8, 1–14. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; De Paula, I.F.; Medina, J.M.; Silva-Oliveira, G.; Medeiros, J.S.; Gäde, G.; Gondim, K.C. Adipokinetic hormone receptor gene identification and its role in triacylglycerol metabolism in the blood-sucking insect Rhodnius prolixus. Insect Biochem. Mol. Biol. 2016, 69, 51–60. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Gondim, K.C. Triacylglycerol biosynthesis occurs via the glycerol-3-phosphate pathway in the insect Rhodnius prolixus. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2012, 1821, 1462–1471. [Google Scholar] [CrossRef]
- Anand, A.N.; Lorenz, M.W. Age-dependent changes of fat body stores and the regulation of fat body lipid synthesis and mobilisation by adipokinetic hormone in the last larval instar of the cricket, Gryllus bimaculatus. J. Insect Physiol. 2008, 54, 1404–1412. [Google Scholar] [CrossRef]
- Annandale, M.; Daniels, L.J.; Li, X.; Neale, J.P.H.; Chau, A.H.L.; Ambalawanar, H.A.; James, S.L.; Koutsifeli, P.; Delbridge, L.M.D.; Mellor, K.M. Fructose Metabolism and Cardiac Metabolic Stress. Front. Pharmacol. 2021, 12, 695486. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect Fat Body: Energy, Metabolism, and Regulation. Annu. Rev. Èntomol. 2010, 55, 207–225. [Google Scholar] [CrossRef]
- Auerswald, L. & G, G. A. 1995. Energy substrates for flight in the blister beetle Decapotoma lunata (Meloidae). J Exp Biol, 198(Pt 6), pp 1423-31.
- Auerswald, L.; Schneider, P.; Gäde, G. Utilisation of Substrates During Tethered Flight with and without Lift Generation in the African Fruit Beetle Pachnoda Sinuata (Cetoniinae). J. Exp. Biol. 1998, 201, 2333–2342. [Google Scholar] [CrossRef]
- Back, S.H.; Kaufman, R.J. Endoplasmic Reticulum Stress and Type 2 Diabetes. Annu. Rev. Biochem. 2012, 81, 767–793. [Google Scholar] [CrossRef]
- Bailey, E.; A Horne, J.; Izarr, M.E.G.; Hill, L. The role of ketone bodies in the metabolism of the adult desert locust. Biochem. J. 1972, 128, 79P–79P. [Google Scholar] [CrossRef]
- Barber, M. C. , Price, N. T. & Travers, M. T. 2005. Structure and regulation of acetyl-CoA carboxylase genes of metazoa. Biochim Biophys Acta, 1733(1), pp 1-28.
- Bartok, O.; Teesalu, M.; Ashwall-Fluss, R.; Pandey, V.; Hanan, M.; Rovenko, B.M.; Poukkula, M.; Havula, E.; Moussaieff, A.; Vodala, S.; et al. The transcription factor Cabut coordinates energy metabolism and the circadian clock in response to sugar sensing. EMBO J. 2015, 34, 1538–1553. [Google Scholar] [CrossRef]
- Becker, A.; Schlöder, P.; Steele, J.E.; Wegener, G. The regulation of trehalose metabolism in insects. Experientia 1996, 52, 433–439. [Google Scholar] [CrossRef]
- Beenakkers, A. Carbohydrate and fat as a fuel for insect flight. A comparative study. J. Insect Physiol. 1969, 15, 353–361. [Google Scholar] [CrossRef]
- Benkel, B.F.; A Hickey, D. GLUCOSE REPRESSION OF AMYLASE GENE EXPRESSION IN DROSOPHILA MELANOGASTER. Genetics 1986, 114, 137–144. [Google Scholar] [CrossRef]
- Bifano, T.D.; Alegria, T.G.; Terra, W.R. Transporters involved in glucose and water absorption in the Dysdercus peruvianus (Hemiptera: Pyrrhocoridae) anterior midgut. Comp. Biochem. Physiol. Part B: Biochem. Mol. Biol. 2010, 157, 1–9. [Google Scholar] [CrossRef]
- Bland, M.L.; Lee, R.J.; Magallanes, J.M.; Foskett, J.K.; Birnbaum, M.J. AMPK supports growth in Drosophila by regulating muscle activity and nutrient uptake in the gut. Dev. Biol. 2010, 344, 293–303. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Ginzel, M.D. Chemical Ecology, Biochemistry, and Molecular Biology of Insect Hydrocarbons. Annu. Rev. Èntomol. 2021, 66, 45–60. [Google Scholar] [CrossRef]
- Boer, P. H. & Hickey, D. A. 1986. The alpha-amylase gene in Drosophila melanogaster: nucleotide sequence, gene structure and expression motifs. Nucleic Acids Res, 14(21), pp 8399-411.
- Braz, V.; Selim, L.; Gomes, G.; Costa, M.L.; Mermelstein, C.; Gondim, K.C. Blood meal digestion and changes in lipid reserves are associated with the post-ecdysis development of the flight muscle and ovary in young adults of Rhodnius prolixus. J. Insect Physiol. 2023, 146, 104492. [Google Scholar] [CrossRef]
- Briegel, H.; Hefti, M.; DiMarco, E. Lipid metabolism during sequential gonotrophic cycles in large and small female Aedes aegypti. J. Insect Physiol. 2002, 48, 547–554. [Google Scholar] [CrossRef]
- Brookheart, R. T. , Michel, C. I. & Schaffer, J. E. 2009. As a matter of fat. Cell Metab, 10(1), pp 9-12.
- Brooks, G.A. Amino acid and protein metabolism during exercise and recovery. Med. Sci. Sports Exerc. 1987, 19, S150–6. [Google Scholar] [CrossRef]
- Bursell, E. 1981. The Role of Proline in Energy Metabolism. In: Downer, R. G. H. (ed.) Energy Metabolism in Insects. Plenum Press, New York.
- Caccia, S.; Leonardi, M.; Casartelli, M.; Grimaldi, A.; de Eguileor, M.; Pennacchio, F.; Giordana, B. Nutrient absorption by Aphidius ervi larvae. J. Insect Physiol. 2005, 51, 1183–1192. [Google Scholar] [CrossRef]
- Calderón-Cortés, N.; Quesada, M.; Watanabe, H.; Cano-Camacho, H.; Oyama, K. Endogenous Plant Cell Wall Digestion: A Key Mechanism in Insect Evolution. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 45–71. [Google Scholar] [CrossRef]
- E Canavoso, L.; Stariolo, R.; Rubiolo, E.R. Flight metabolism in Panstrongylus megistus (Hemiptera: Reduviidae): the role of carbohydrates and lipids. Mem. Do Inst. Oswaldo Cruz 2003, 98, 909–914. [Google Scholar] [CrossRef]
- Chandel, N. S. 2022. Carbohydrate Metabolism. Cold Spring Harb Perspect Biol, 13(a040568.
- Chapman, R. F. , Simpson, S. J. & Douglas, A. E. 2013. The Insects. Structure and Function., Cambridge University Press, New York.
- Childress, C.C.; Sacktor, B. Regulation of glycogen metabolism in insect flight muscle. Purification and properties of phosphorylases in vitro and in vivo.. 1970, 245, 2927–36. [Google Scholar] [CrossRef]
- Childress, C. C. , Sacktor, B., Grossman, I. W. & Bueding, E. 1970. Isolation, ultrastructure, and biochemal characterization of glycogen in insect flight muscle. J Cell Biol, 45(1), pp 83-90.
- Chino, H. & Downer, R. G. H. 1979. The role of diacylglycerol in absorption of dietary glyceride in the american cockroach Periplanata americana L.. Insect Biochem., 9(379-382.
- Chng, W. A. , Sleiman, M. S. B., Schupfer, F. & Lemaitre, B. 2014. Transforming growth factor beta/activin signaling functions as a sugar-sensing feedback loop to regulate digestive enzyme expression. Cell Rep, 9(1), pp 336-348.
- Chowański, S.; Walkowiak-Nowicka, K.; Winkiel, M.; Marciniak, P.; Urbański, A.; Pacholska-Bogalska, J. Insulin-Like Peptides and Cross-Talk With Other Factors in the Regulation of Insect Metabolism. Front. Physiol. 2021, 12, 701203. [Google Scholar] [CrossRef]
- Christeller, J.T.; Amara, S.; Carrière, F. Galactolipase, phospholipase and triacylglycerol lipase activities in the midgut of six species of lepidopteran larvae feeding on different lipid diets. J. Insect Physiol. 2011, 57, 1232–1239. [Google Scholar] [CrossRef]
- Clark, A.J.; Block, K. The absence of sterol synthesis in insects. . 1959, 234, 2578–82. [Google Scholar] [CrossRef]
- Clements, A. N. 1959. Studies on the metabolism of locust fat body J. Exptl. Biol., 36(665-.
- Coleman, R.A. It takes a village: channeling fatty acid metabolism and triacylglycerol formation via protein interactomes. J. Lipid Res. 2019, 60, 490–497. [Google Scholar] [CrossRef]
- Coleman, R.A.; Mashek, D.G. Mammalian Triacylglycerol Metabolism: Synthesis, Lipolysis, and Signaling. Chem. Rev. 2011, 111, 6359–6386. [Google Scholar] [CrossRef]
- Colombani, J.; Raisin, S.; Pantalacci, S.; Radimerski, T.; Montagne, J.; Léopold, P. A Nutrient Sensor Mechanism Controls Drosophila Growth. Cell 2003, 114, 739–749. [Google Scholar] [CrossRef]
- Csibi, A.; Fendt, S.-M.; Li, C.; Poulogiannis, G.; Choo, A.Y.; Chapski, D.J.; Jeong, S.M.; Dempsey, J.M.; Parkhitko, A.; Morrison, T.; et al. TThe mTORC1 pathway stimulates glutamine metabolism and cell proliferation by repressing SIRT4. Cell 2013, 153, 840–854. [Google Scholar] [CrossRef]
- Daenzer, J. M. & Fridovich-Keil, J. L. 2017. Drosophila melanogaster Models of Galactosemia. Curr Top Dev Biol, 121(377-395.
- Deng, D.; Yan, N. GLUT, SGLT, and SWEET: Structural and mechanistic investigations of the glucose transporters. Protein Sci. 2015, 25, 546–558. [Google Scholar] [CrossRef]
- Devilliers, M. , Garrido, D., Poidevin, M., Rubin, T., Le Rouzic, A. & Montagne, J. 2021. Differential Metabolic Sensitivity of Insulin-like-response- and mTORC1-Dependent Overgrowth in Drosophila Fat Cells. Genetics, 217(1), pp 1-12.
- Downer, R.; Matthews, J. Glycogen depletion of thoracic musculature during flight in the American cockroach, Periplaneta americana L. Comp. Biochem. Physiol. Part B: Comp. Biochem. 1976, 55, 501–502. [Google Scholar] [CrossRef]
- Drahun, I.; Poole, E.A.; Hunt, K.A.; van Herk, W.G.; LeMoine, C.M.; Cassone, B.J. Seasonal turnover and insights into the overwintering biology of wireworms (Coleoptera: Elateridae) in the Canadian Prairies. Pest Manag. Sci. 2023, 79, 526–536. [Google Scholar] [CrossRef]
- Eanes, W.F.; Merritt, T.J.S.; Flowers, J.M.; Kumagai, S.; Sezgin, E.; Zhu, C.-T. Flux control and excess capacity in the enzymes of glycolysis and their relationship to flight metabolism inDrosophila melanogaster. Proc. Natl. Acad. Sci. 2006, 103, 19413–19418. [Google Scholar] [CrossRef]
- Evangelista, D.E.; de Paula, F.F.P.; Rodrigues, A.; Henrique-Silva, F. Pectinases From Sphenophorus levis Vaurie, 1978 (Coleoptera: Curculionidae): Putative Accessory Digestive Enzymes. J. Insect Sci. 2015, 15, 5–5. [Google Scholar] [CrossRef]
- Fan, J.; Ye, J.; Kamphorst, J.J.; Shlomi, T.; Thompson, C.B.; Rabinowitz, J.D. Quantitative flux analysis reveals folate-dependent NADPH production. Nature 2014, 510, 298–302. [Google Scholar] [CrossRef]
- Faust, J.E.; Manisundaram, A.; Ivanova, P.T.; Milne, S.B.; Summerville, J.B.; Brown, H.A.; Wangler, M.; Stern, M.; McNew, J.A. Peroxisomes Are Required for Lipid Metabolism and Muscle Function in Drosophila melanogaster. PLOS ONE 2014, 9, e100213. [Google Scholar] [CrossRef]
- Febvay, G. , Pageaux, J. & G., B. 1992. Lipid Composition of the Pea Aphid, Acyrthosiphon pisum (Harris) (Hornoptera: Aphididae), Reared on Host Plant and on Artificial Media. Archives of Insect Biochemistry and Physiology, 21(103-118.
- Febvay, G.; Rahbé, Y.; Rynkiewicz, M.; Guillaud, J.; Bonnot, G. Fate of dietary sucrose and neosynthesis of amino acids in the pea aphid,Acyrthosiphon pisum, reared on different diets. J. Exp. Biol. 1999, 202, 2639–2652. [Google Scholar] [CrossRef]
- Fogle, K.J.; Smith, A.R.; Satterfield, S.L.; Gutierrez, A.C.; Hertzler, J.I.; McCardell, C.S.; Shon, J.H.; Barile, Z.J.; Novak, M.O.; Palladino, M.J. Ketogenic and anaplerotic dietary modifications ameliorate seizure activity in Drosophila models of mitochondrial encephalomyopathy and glycolytic enzymopathy. Mol. Genet. Metab. 2019, 126, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Ford, P. S. & Van Heusden, M. C. 1994. Triglyceride-rich lipophorin in Aedes aegypti (Diptera: Culicidae). J Med Entomol, 31(3), pp 435-41.
- Gäde, G. Glycogen Phosphorylase in the Fat Body of Two Cockroach Species, Periplaneta americana and Nauphoeta cinerea: Isolation, Partial Characterization of Three Forms and Activation by Hypertrehalosaemic Hormones. Z. Fur Naturforschung Sect. C-A J. Biosci. 1991, 46, 149–162. [Google Scholar] [CrossRef]
- Gade, G. 2009. Peptides of the adipokinetic hormone/red pigment-concentrating hormone family: a new take on biodiversity. Ann N Y Acad Sci, 1163(125-36.
- Gade, G. & Auerswald, L. 2002. Beetles' choice--proline for energy output: control by AKHs. Comp Biochem Physiol B Biochem Mol Biol, 132(1), pp 117-29.
- Gäde, G.; Auerswald, L. Mode of action of neuropeptides from the adipokinetic hormone family. Gen. Comp. Endocrinol. 2003, 132, 10–20. [Google Scholar] [CrossRef]
- Gáliková, M.; Diesner, M.; Klepsatel, P.; Hehlert, P.; Xu, Y.; Bickmeyer, I.; Predel, R.; Kühnlein, R.P. Energy Homeostasis Control in Drosophila Adipokinetic Hormone Mutants. Genetics 2015, 201, 665–683. [Google Scholar] [CrossRef]
- Gáliková, M.; Klepsatel, P. Endocrine control of glycogen and triacylglycerol breakdown in the fly model. Semin. Cell Dev. Biol. 2023, 138, 104–116. [Google Scholar] [CrossRef]
- Garrido, D.; Rubin, T.; Poidevin, M.; Maroni, B.; Le Rouzic, A.; Parvy, J.-P.; Montagne, J. Fatty Acid Synthase Cooperates with Glyoxalase 1 to Protect against Sugar Toxicity. PLOS Genet. 2015, 11, e1004995–e1004995. [Google Scholar] [CrossRef] [PubMed]
- Gaviraghi, A.; Soares, J.B.C.; Mignaco, J.A.; Fontes, C.F.L.; Oliveira, M.F. Mitochondrial glycerol phosphate oxidation is modulated by adenylates through allosteric regulation of cytochrome c oxidase activity in mosquito flight muscle. Insect Biochem. Mol. Biol. 2019, 114, 103226. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.C.; O’connor, M.B. Systemic Activin signaling independently regulates sugar homeostasis, cellular metabolism, and pH balance in Drosophila melanogaster. Proc. Natl. Acad. Sci. 2014, 111, 5729–5734. [Google Scholar] [CrossRef] [PubMed]
- Girard, C.; Jouanin, L. Molecular cloning of cDNAs encoding a range of digestive enzymes from a phytophagous beetle, Phaedon cochleariae. Insect Biochem. Mol. Biol. 1999, 29, 1129–1142. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, J.T.; Betts, J.A. Dietary sugars, exercise and hepatic carbohydrate metabolism. Proc. Nutr. Soc. 2018, 78, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Govindaraj, L.; Gupta, T.; Esvaran, V.G.; Awasthi, A.K.; Ponnuvel, K.M. Genome-wide identification, characterization of sugar transporter genes in the silkworm Bombyx mori and role in Bombyx mori nucleopolyhedrovirus (BmNPV) infection. Gene 2016, 579, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Grillo, L.; Pontes, E.; Gondim, K. Lipophorin interaction with the midgut of Rhodnius prolixus: characterization and changes in binding capacity. Insect Biochem. Mol. Biol. 2003, 33, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, E.; Wiggins, D.; Fielding, B.; Gould, A.P. Specialized hepatocyte-like cells regulate Drosophila lipid metabolism. Nature 2006, 445, 275–280. [Google Scholar] [CrossRef]
- Haj-Ahmad, Y.; Hickey, D.A. A molecular explanation of frequency-dependent selection in Drosophila. Nature 1982, 299, 350–352. [Google Scholar] [CrossRef]
- Hasygar, K.; Deniz, O.; Liu, Y.; Gullmets, J.; Hynynen, R.; Ruhanen, H.; Kokki, K.; Käkelä, R.; Hietakangas, V. Coordinated control of adiposity and growth by anti-anabolic kinase ERK7. Embo Rep. 2020, 22, e49602. [Google Scholar] [CrossRef]
- Hatting, M.; Tavares, C.D.; Sharabi, K.; Rines, A.K.; Puigserver, P. Insulin regulation of gluconeogenesis. Ann. New York Acad. Sci. 2017, 1411, 21–35. [Google Scholar] [CrossRef]
- Havula, E. & Hietakangas, V. 2018. Sugar sensing by ChREBP/Mondo-Mlx-new insight into downstream regulatory networks and integration of nutrient-derived signals. Curr Opin Cell Biol, 51(89-96.
- Havula, E. , Teesalu, M., Hyotylainen, T., Seppala, H., Hasygar, K., Auvinen, P., Oresic, M., Sandmann, T. & Hietakangas, V. 2013. Mondo/ChREBP-Mlx-regulated transcriptional network is essential for dietary sugar tolerance in Drosophila. PLoS Genet, 9(4), pp e1003438.
- Heier, C.; Klishch, S.; Stilbytska, O.; Semaniuk, U.; Lushchak, O. The Drosophila model to interrogate triacylglycerol biology. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2021, 1866, 158924. [Google Scholar] [CrossRef]
- Heier, C.; Kühnlein, R.P. Triacylglycerol Metabolism in Drosophila melanogaster. Genetics 2018, 210, 1163–1184. [Google Scholar] [CrossRef] [PubMed]
- Hines, W.; Smith, M. Some aspects of intermediary metabolism in the desert locust (Schistocerca gregaria Forskål). J. Insect Physiol. 1963, 9, 463–468. [Google Scholar] [CrossRef]
- Horecker, B. L. 2002. The pentose phosphate pathway. J Biol Chem, 277(50), pp 47965-71.
- Hori, K. Digestive carbohydrases in the salivary gland and midgut of several phytophagous bugs. Comp. Biochem. Physiol. Part B: Comp. Biochem. 1975, 50, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Chen, W.; Zhu, F.; Li, P.W.-L.; Kapahi, P.; Bai, H. RiboTag translatomic profiling of Drosophila oenocytes under aging and induced oxidative stress. BMC Genom. 2019, 20, 50. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Liu, Y.; Perrimon, N. Roles of Insect Oenocytes in Physiology and Their Relevance to Human Metabolic Diseases. Front. Insect Sci. 2022, 2. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, S.; Yamashita, O. Metabolic shift from lipogenesis to glycogenesis in the last instar larval fat body of the silkworm, Bombyx mori. Insect Biochem. 1986, 16, 327–331. [Google Scholar] [CrossRef]
- Inigo, M.; Deja, S.; Burgess, S.C. Ins and Outs of the TCA Cycle: The Central Role of Anaplerosis. Annu. Rev. Nutr. 2021, 41, 19–47. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Stenvinkel, P.; Andrews, P.; Sánchez-Lozada, L.G.; Nakagawa, T.; Gaucher, E.; Andres-Hernando, A.; Rodriguez-Iturbe, B.; Jimenez, C.R.; Garcia, G.; et al. Fructose metabolism as a common evolutionary pathway of survival associated with climate change, food shortage and droughts. J. Intern. Med. 2019, 287, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Jutsum, A.; Goldsworthy, G. Fuels for flight in Locusta. J. Insect Physiol. 1976, 22, 243–249. [Google Scholar] [CrossRef]
- Kanamori, Y.; Saito, A.; Hagiwara-Komoda, Y.; Tanaka, D.; Mitsumasu, K.; Kikuta, S.; Watanabe, M.; Cornette, R.; Kikawada, T.; Okuda, T. The trehalose transporter 1 gene sequence is conserved in insects and encodes proteins with different kinetic properties involved in trehalose import into peripheral tissues. Insect Biochem. Mol. Biol. 2010, 40, 30–37. [Google Scholar] [CrossRef]
- Kastari, T. & Turunen, S. 1977. Lipid utilization in Pieris brassicae reared on meridic and natural diest: implication for dietary improvement. Ent. exp. & appl., 22(71-80.
- Katewa, S.D.; Demontis, F.; Kolipinski, M.; Hubbard, A.; Gill, M.S.; Perrimon, N.; Melov, S.; Kapahi, P. Intramyocellular Fatty-Acid Metabolism Plays a Critical Role in Mediating Responses to Dietary Restriction in Drosophila melanogaster. Cell Metab. 2012, 16, 97–103. [Google Scholar] [CrossRef]
- Kaufmann, C.; Brown, M.R. Regulation of carbohydrate metabolism and flight performance by a hypertrehalosaemic hormone in the mosquito Anopheles gambiae. J. Insect Physiol. 2007, 54, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Kikuta, S.; Nakamura, Y.; Hattori, M.; Sato, R.; Kikawada, T.; Noda, H. Herbivory-induced glucose transporter gene expression in the brown planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2015, 64, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Neufeld, T.P. Dietary sugar promotes systemic TOR activation in Drosophila through AKH-dependent selective secretion of Dilp3. Nat. Commun. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- King, B.; Li, S.; Liu, C.; Kim, S.J.; Sim, C. Suppression of glycogen synthase expression reduces glycogen and lipid storage during mosquito overwintering diapause. J. Insect Physiol. 2019, 120, 103971–103971. [Google Scholar] [CrossRef] [PubMed]
- Kirkton, S. D. & Tyler, S. K. 2021. American locust (Schistocerca americana) post-exercise lactate fate dataset. Data Brief, 37(107263.
- Ko, C.-W.; Qu, J.; Black, D.D.; Tso, P. Regulation of intestinal lipid metabolism: current concepts and relevance to disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Kokki, K.; Lamichane, N.; Nieminen, A.I.; Ruhanen, H.; Morikka, J.; Robciuc, M.; Rovenko, B.M.; Havula, E.; Käkelä, R.; Hietakangas, V. Metabolic gene regulation by Drosophila GATA transcription factor Grain. PLOS Genet. 2021, 17, e1009855. [Google Scholar] [CrossRef] [PubMed]
- Krause, N.; Wegner, A. Fructose Metabolism in Cancer. Cells 2020, 9, 2635. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yu, X.; Feng, Q. Fat Body Biology in the Last Decade. Annu. Rev. Èntomol. 2019, 64, 315–333. [Google Scholar] [CrossRef]
- Li, X.; Zhou, Y.; Wu, K. Biological Characteristics and Energy Metabolism of Migrating Insects. Metabolites 2023, 13, 439. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Lim, H.-Y. Drosophila Solute Carrier 5A5 Regulates Systemic Glucose Homeostasis by Mediating Glucose Absorption in the Midgut. Int. J. Mol. Sci. 2021, 22, 12424. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Pan, S.; Wang, X.; Liu, Z.; Zhang, Y. Role of advanced glycation end products in diabetic vascular injury: molecular mechanisms and therapeutic perspectives. Eur. J. Med Res. 2023, 28, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liu, K. , Dong, Y., Huang, Y., Rasgon, J. L. & Agre, P. 2013. Impact of trehalose transporter knockdown on Anopheles gambiae stress adaptation and susceptibility to Plasmodium falciparum infection. Proc Natl Acad Sci U S A, 110(43), pp 17504-9.
- Lorenz, M.W. Oogenesis-flight syndrome in crickets: Age-dependent egg production, flight performance, and biochemical composition of the flight muscles in adult female Gryllus bimaculatus. J. Insect Physiol. 2007, 53, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.W.; Anand, A.N. Changes in the biochemical composition of fat body stores during adult development of female crickets, Gryllus bimaculatus. Arch. Insect Biochem. Physiol. 2004, 56, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Lv, N.; Wang, L.; Sang, W.; Liu, C.-Z.; Qiu, B.-L. Effects of Endosymbiont Disruption on the Nutritional Dynamics of the Pea Aphid Acyrthosiphon pisum. Insects 2018, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Maier, T.; Leibundgut, M.; Boehringer, D.; Ban, N. Structure and function of eukaryotic fatty acid synthases. Q. Rev. Biophys. 2010, 43, 373–422. [Google Scholar] [CrossRef] [PubMed]
- Masson, S.W.C.; Hedges, C.P.; Devaux, J.B.L.; James, C.S.; Hickey, A.J.R. Mitochondrial glycerol 3-phosphate facilitates bumblebee pre-flight thermogenesis. Sci. Rep. 2017, 7, 13107. [Google Scholar] [CrossRef]
- Mattila, J.; Havula, E.; Suominen, E.; Teesalu, M.; Surakka, I.; Hynynen, R.; Kilpinen, H.; Väänänen, J.; Hovatta, I.; Käkelä, R.; et al. Mondo-Mlx Mediates Organismal Sugar Sensing through the Gli-Similar Transcription Factor Sugarbabe. Cell Rep. 2015, 13, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Mattila, J.; Hietakangas, V. Regulation of Carbohydrate Energy Metabolism in Drosophila melanogaster. . 2017, 207, 1231–1253. [Google Scholar] [CrossRef]
- Michitsch, J.; Steele, J.E. Carbohydrate and lipid metabolism in cockroach (Periplaneta americana) fat body are both activated by low and similar concentrations of Peram-AKH II. Peptides 2008, 29, 226–234. [Google Scholar] [CrossRef]
- Montagne, J. & Wicker-Thomas, C. 2020. Drosophila pheromone production. In: Blomquist G. & Vogt R. G. (eds.) Insect Pheromone Biochemistry and Molecular Biology. Elsevier Academic Press, London.
- Mor, I.; Cheung, E.C.; Vousden, K.H. Control of Glycolysis through Regulation of PFK1: Old Friends and Recent Additions. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Moraes, K.C.M.; Montagne, J. Drosophila melanogaster: A Powerful Tiny Animal Model for the Study of Metabolic Hepatic Diseases. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Moraru, A.; Wiederstein, J.; Pfaff, D.; Fleming, T.; Miller, A.K.; Nawroth, P.; Teleman, A.A. Elevated Levels of the Reactive Metabolite Methylglyoxal Recapitulate Progression of Type 2 Diabetes. Cell Metab. 2018, 27, 926–934. [Google Scholar] [CrossRef] [PubMed]
- Moreira, N.R.; Cardoso, C.; Dias, R.O.; Ferreira, C.; Terra, W.R. A physiologically-oriented transcriptomic analysis of the midgut of Tenebrio molitor. J. Insect Physiol. 2017, 99, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Mráček, T.; Drahota, Z.; Houštěk, J. The function and the role of the mitochondrial glycerol-3-phosphate dehydrogenase in mammalian tissues. Biochim. et Biophys. Acta (BBA) - Bioenerg. 2013, 1827, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.A.; Wyatt, G. The Enzymes of Glycogen and Trehalose Synthesis in Silk Moth Fat Body. J. Biol. Chem. 1965, 240, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.P.; Fink, J.L.; Narzinski, K.; Ramachandran, P.V.; Hathiramani, S.S.; Cagan, R.L.; Baranski, T.J. A high-sugar diet produces obesity and insulin resistance in wild-type Drosophila. Dis. Models Mech. 2011, 4, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Musselman, L.P.; Fink, J.L.; Ramachandran, P.V.; Patterson, B.W.; Okunade, A.L.; Maier, E.; Brent, M.R.; Turk, J.; Baranski, T.J. Role of Fat Body Lipogenesis in Protection against the Effects of Caloric Overload in Drosophila. J. Biol. Chem. 2013, 288, 8028–8042. [Google Scholar] [CrossRef] [PubMed]
- Mustard, J.A.; Alvarez, V.; Barocio, S.; Mathews, J.; Stoker, A.; Malik, K. Nutritional value and taste play different roles in learning and memory in the honey bee (Apis mellifera). J. Insect Physiol. 2018, 107, 250–256. [Google Scholar] [CrossRef]
- Na, J.; Musselman, L.P.; Pendse, J.; Baranski, T.J.; Bodmer, R.; Ocorr, K.; Cagan, R. A Drosophila Model of High Sugar Diet-Induced Cardiomyopathy. PLOS Genet. 2013, 9, e1003175. [Google Scholar] [CrossRef]
- Nishihara, S. Functional analysis of glycosylation using Drosophila melanogaster. Glycoconj. J. 2019, 37, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Oguri, E.; Steele, J.E. A novel function of cockroach (Periplaneta americana) hypertrehalosemic hormone: translocation of lipid from hemolymph to fat body. Gen. Comp. Endocrinol. 2003, 132, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Olademehin, O.P.; Liu, C.; Rimal, B.; Adegboyega, N.F.; Chen, F.; Sim, C.; Kim, S.J. Dsi-RNA knockdown of genes regulated by Foxo reduces glycogen and lipid accumulations in diapausing Culex pipiens. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Olzmann, J.A.; Carvalho, P. Dynamics and functions of lipid droplets. Nat. Rev. Mol. Cell Biol. 2019, 20, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Palm, W. , Sampaio, J. L., Brankatschk, M., Carvalho, M., Mahmoud, A., Shevchenko, A. & Eaton, S. 2012. Lipoproteins in Drosophila melanogaster--assembly, function, and influence on tissue lipid composition. PLoS Genet, 8(7), pp e1002828.
- Parvy, J. P. , Napal, L., Rubin, T., Poidevin, M., Perrin, L., Wicker-Thomas, C. & Montagne, J. 2012. Drosophila melanogaster Acetyl-CoA-carboxylase sustains a fatty acid-dependent remote signal to waterproof the respiratory system. PLoS Genet, 8(8), pp e1002925.
- Pasco, M.Y.; Leopold, P. High sugar-induced insulin resistance in Drosophila relies on the lipocalin Neural Lazarillo. PLoS ONE 2012, 7, e36583. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Abate, N. Role of Subcutaneous Adipose Tissue in the Pathogenesis of Insulin Resistance. J. Obes. 2013, 2013, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Patrick, J.W.; Botha, F.C.; Birch, R.G. Metabolic engineering of sugars and simple sugar derivatives in plants. Plant Biotechnol. J. 2012, 11, 142–156. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Bai, T.; Luo, Y.; Zhang, Z.; Li, S.; Fan, Y.; Liu, T. Acetyl coenzyme A carboxylase modulates lipogenesis and sugar homeostasis in Blattella germanica. Insect Sci. 2023. [Google Scholar] [CrossRef]
- Pennington, J. E. & Wells, M. A. 2002. Triacylglycerol-rich lipophorins are found in the dipteran infraorder Culicomorpha, not just in mosquitoes. J Insect Sci, 2(15.
- Pimentel, A.C.; Barroso, I.G.; Ferreira, J.M.; Dias, R.O.; Ferreira, C.; Terra, W.R. Molecular machinery of starch digestion and glucose absorption along the midgut of Musca domestica. J. Insect Physiol. 2018, 109, 11–20. [Google Scholar] [CrossRef]
- Price, D.R.; Wilkinson, H.S.; Gatehouse, J.A. Functional expression and characterisation of a gut facilitative glucose transporter, NlHT1, from the phloem-feeding insect Nilaparvata lugens (rice brown planthopper). Insect Biochem. Mol. Biol. 2007, 37, 1138–1148. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Methylglyoxal, glyoxalase 1 and the dicarbonyl proteome. Amino Acids 2010, 42, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N. & Thornalley, P. J. 2013. Glyoxalase in diabetes, obesity and related disorders. Semin Cell Dev Biol, 22(3), pp 309-17.
- Radimerski, T.; Montagne, J.; Hemmings-Mieszczak, M.; Thomas, G. Lethality of Drosophila lacking TSC tumor suppressor function rescued by reducing dS6K signaling. Genes Dev. 2002, 16, 2627–2632. [Google Scholar] [CrossRef] [PubMed]
- Rahbé, Y.; Delobel, B.; Febvay, G.; Chantegrel, B. Aphid-specific triglycerides in symbiotic and aposymbiotic Acyrthosiphon pisum. Insect Biochem. Mol. Biol. 1994, 24, 95–101. [Google Scholar] [CrossRef]
- Rajapakse, S.; Qu, D.; Ahmed, A.S.; Rickers-Haunerland, J.; Haunerland, N.H. Effects of FABP knockdown on flight performance of the desert locust, Schistocerca gregaria. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef] [PubMed]
- Ransom-Jones, E.; Jones, D.L.; McCarthy, A.J.; McDonald, J.E. The Fibrobacteres: an Important Phylum of Cellulose-Degrading Bacteria. Microb. Ecol. 2012, 63, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Reis, T.; Van Gilst, M.R.; Hariharan, I.K. A Buoyancy-Based Screen of Drosophila Larvae for Fat-Storage Mutants Reveals a Role for Sir2 in Coupling Fat Storage to Nutrient Availability. PLOS Genet. 2010, 6, e1001206. [Google Scholar] [CrossRef]
- Richards, P.; Ourabah, S.; Montagne, J.; Burnol, A.-F.; Postic, C.; Guilmeau, S. MondoA/ChREBP: The usual suspects of transcriptional glucose sensing; Implication in pathophysiology. Metab. Clin. Exp. 2017, 70, 133–151. [Google Scholar] [CrossRef] [PubMed]
- Roach, P.J.; Depaoli-Roach, A.A.; Hurley, T.D.; Tagliabracci, V.S. Glycogen and its metabolism: some new developments and old themes. Biochem. J. 2012, 441, 763–787. [Google Scholar] [CrossRef]
- Robinson, N.; Goldsworthy, G. Adipokinetic hormone and the regulation of carbohydrate and lipid metabolism in a working flight muscle preparation. J. Insect Physiol. 1977, 23, 9–16. [Google Scholar] [CrossRef]
- Roeder, T. TYRAMINE AND OCTOPAMINE: Ruling Behavior and Metabolism. Annu. Rev. Èntomol. 2005, 50, 447–477. [Google Scholar] [CrossRef]
- Rovenko, B.M.; Perkhulyn, N.V.; Gospodaryov, D.V.; Sanz, A.; Lushchak, O.V.; Lushchak, V.I. High consumption of fructose rather than glucose promotes a diet-induced obese phenotype in Drosophila melanogaster. Comp. Biochem. Physiol. Part A: Mol. Integr. Physiol. 2015, 180, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Saggerson, D. Malonyl-CoA, a Key Signaling Molecule in Mammalian Cells. Annu. Rev. Nutr. 2008, 28, 253–272. [Google Scholar] [CrossRef] [PubMed]
- Sahaka, M.; Amara, S.; Wattanakul, J.; Gedi, M.A.; Aldai, N.; Parsiegla, G.; Lecomte, J.; Christeller, J.T.; Gray, D.; Gontero, B.; et al. The digestion of galactolipids and its ubiquitous function in Nature for the uptake of the essential α-linolenic acid. Food Funct. 2020, 11, 6710–6744. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Nakamura, A.; Yamane, M.; Niwa, H.; Nishimura, T.; Araki, K.; Takemoto, K.; Ishiguro, K.-I.; Aoki, H.; Kato, Y.; et al. The polyol pathway is an evolutionarily conserved system for sensing glucose uptake. PLOS Biol. 2022, 20, e3001678. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Shukla, E.; Thorat, L.J.; Nath, B.B.; Gaikwad, S.M. Insect trehalase: Physiological significance and potential applications. Glycobiology 2014, 25, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Silva-Oliveira, G.; De Paula, I.F.; Medina, J.M.; Alves-Bezerra, M.; Gondim, K.C. Insulin receptor deficiency reduces lipid synthesis and reproductive function in the insect Rhodnius prolixus. Biochim. et Biophys. Acta (BBA) - Mol. Cell Biol. Lipids 2021, 1866, 158851. [Google Scholar] [CrossRef]
- Smith, S.; Witkowski, A.; Joshi, A.K. Structural and functional organization of the animal fatty acid synthase. Prog. Lipid Res. 2003, 42, 289–317. [Google Scholar] [CrossRef]
- Soares, J.B.R.C.; Gaviraghi, A.; Oliveira, M.F. Mitochondrial Physiology in the Major Arbovirus Vector Aedes aegypti: Substrate Preferences and Sexual Differences Define Respiratory Capacity and Superoxide Production. PLOS ONE 2015, 10, e0120600–e0120600. [Google Scholar] [CrossRef]
- Stec, N.; Saleem, A.; Darveau, C.-A. Proline as a Sparker Metabolite of Oxidative Metabolism during the Flight of the Bumblebee, Bombus impatiens. Metabolites 2021, 11, 511. [Google Scholar] [CrossRef] [PubMed]
- Steele, J. Glycogen phosphorylase in insects. Insect Biochem. 1982, 12, 131–147. [Google Scholar] [CrossRef]
- Storelli, G.; Nam, H.-J.; Simcox, J.; Villanueva, C.J.; Thummel, C.S. Drosophila HNF4 Directs a Switch in Lipid Metabolism that Supports the Transition to Adulthood. Dev. Cell 2018, 48, 200–214. [Google Scholar] [CrossRef]
- Su, S. , Zhang, X., Jian, C., Huang, B., Peng, X., Vreysen, M. J. B. & Chen, M. 2022. Effects of Adult Feeding Treatments on Longevity, Fecundity, Flight Ability, and Energy Metabolism Enzymes of Grapholita molesta Moths. Insects, 13(8), pp.
- Suarez, R.K.; Darveau, C.-A.; Welch, K.C.; O'Brien, D.M.; Roubik, D.W.; Hochachka, P.W. Energy metabolism in orchid bee flight muscles: carbohydrate fuels all. J. Exp. Biol. 2005, 208, 3573–3579. [Google Scholar] [CrossRef]
- Szyszka, P.; Galizia, C.G. The Role of the Sucrose-Responsive IR60b Neuron for Drosophila melanogaster: A Hypothesis. Chem. Senses 2018, 43, 311–312. [Google Scholar] [CrossRef]
- Taskinen, M.-R.; Packard, C.J.; Borén, J. Dietary Fructose and the Metabolic Syndrome. Nutrients 2019, 11, 1987. [Google Scholar] [CrossRef]
- Terra, W.R.; Ferreira, C. Insect digestive enzymes: properties, compartmentalization and function. Comp. Biochem. Physiol. Part B: Comp. Biochem. 1994, 109, 1–62. [Google Scholar] [CrossRef]
- Terra, W. R. & Ferreira, C. 2012. Biochemistry and molecular biology of digestion. In: Gilbert, L. I. (ed.) Insect Molecular Biology and Biochemistry.. Academic Press/Elsevier, London.
- Thompson, S. N. 2003. Trehalose – The Insect ‘Blood’ Sugar. Advances in insect physiology, 31(.
- Thornalley, P.J. The glyoxalase system in health and disease. Mol. Asp. Med. 1993, 14, 287–371. [Google Scholar] [CrossRef]
- Tietz, A. Fat synthesis in cell-free preparations of the locust fat-body. J. Lipid Res. 1961, 2, 182–187. [Google Scholar] [CrossRef]
- Toprak, U. The Role of Peptide Hormones in Insect Lipid Metabolism. Front. Physiol. 2020, 11, 434. [Google Scholar] [CrossRef] [PubMed]
- Towarnicki, S.G.; Ballard, J.W.O. Towards understanding the evolutionary dynamics of mtDNA. Mitochondrial DNA Part A 2020, 31, 355–364. [Google Scholar] [CrossRef]
- Turturro, A.; Shafiq, S.A. Quantitative Morphological Analysis of Age-Related Changes in Flight Muscle of Musca Domestica L. J. Gerontol. 1979, 34, 823–833. [Google Scholar] [CrossRef]
- Turunen, S. Absorption and transport of dietary lipid in Pieris brassicae. J. Insect Physiol. 1975, 21, 1521–1529. [Google Scholar] [CrossRef]
- Turunen, S. Metabolic pathways in the midgut epithelium of Pieris brassicae during carbohydrate and lipid assimilation. Insect Biochem. Mol. Biol. 1993, 23, 681–689. [Google Scholar] [CrossRef]
- Turunen, S.; Kastari, T. Digestion and absorption of lecithin in larvae of the cabbage butterfly, Pieris brassicae. Comp. Biochem. Physiol. Part A: Physiol. 1979, 62, 933–937. [Google Scholar] [CrossRef]
- van der Horst, D. J. , van Doorn, J. M., Passier, P. C., Vork, M. M. & Glatz, J. F. 1993. Role of fatty acid-binding protein in lipid metabolism of insect flight muscle. Mol Cell Biochem, 123(1-2), pp 145-52.
- Varghese, J. , Lim, S. F. & Cohen, S. M. 2010. Drosophila miR-14 regulates insulin production and metabolism through its target, sugarbabe. Genes Dev, 24(24), pp 2748-53.
- Veenstra, J.A. Simulation of the activation of fat body glycogen phosphorylase and trehalose synthesis by peptide hormones in the American cockroach. Biosystems 1989, 23, 31–40. [Google Scholar] [CrossRef]
- Wang, M.; Wang, J. Glucose transporter GLUT1 influences Plasmodium berghei infection in Anopheles stephensi. Parasites Vectors 2020, 13, 1–11. [Google Scholar] [CrossRef]
- Wang, Y.; Campbell, J.B.; Kaftanoglu, O.; Page, R.E.; Amdam, G.V.; Harrison, J.F. Larval starvation improves metabolic response to adult starvation in honey bees (Apis mellifera L.). J. Exp. Biol. 2016, 219, 960–968. [Google Scholar] [CrossRef]
- Watanabe, H.; Noda, H.; Tokuda, G.; Lo, N. A cellulase gene of termite origin. Nature 1998, 394, 330–331. [Google Scholar] [CrossRef]
- Watanabe, H.; Tokuda, G. Cellulolytic Systems in Insects. Annu. Rev. Èntomol. 2010, 55, 609–632. [Google Scholar] [CrossRef]
- Wegener, G. Flying insects: model systems in exercise physiology. Cell. Mol. Life Sci. 1996, 52, 404–412. [Google Scholar] [CrossRef]
- Weiglein, A.; Gerstner, F.; Mancini, N.; Schleyer, M.; Gerber, B. One-trial learning in larval Drosophila. Learn. Mem. 2019, 26, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Wicker-Thomas, C.; Garrido, D.; Bontonou, G.; Napal, L.; Mazuras, N.; Denis, B.; Rubin, T.; Parvy, J.-P.; Montagne, J. Flexible origin of hydrocarbon/pheromone precursors in Drosophila melanogaster. J. Lipid Res. 2015, 56, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Willis, J.D.; Oppert, B.; Oppert, C.; Klingeman, W.E.; Jurat-Fuentes, J.L. Identification, cloning, and expression of a GHF9 cellulase from Tribolium castaneum (Coleoptera: Tenebrionidae). J. Insect Physiol. 2011, 57, 300–306. [Google Scholar] [CrossRef]
- Wone, B. W. M. , Kinchen, J. M., Kaup, E. R. & Wone, B. 2018. A procession of metabolic alterations accompanying muscle senescence in Manduca sexta. Sci Rep, 8(1), pp 1006.
- Wood, J.G.; Schwer, B.; Wickremesinghe, P.C.; Hartnett, D.A.; Burhenn, L.; Garcia, M.; Li, M.; Verdin, E.; Helfand, S.L. Sirt4 is a mitochondrial regulator of metabolism and lifespan in Drosophila melanogaster. Proc. Natl. Acad. Sci. 2018, 115, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M.; Hirayama, B.A.; Loo, D.F. Active sugar transport in health and disease. J. Intern. Med. 2007, 261, 32–43. [Google Scholar] [CrossRef]
- Wu, Q.; Brown, M.R. SIGNALING AND FUNCTION OF INSULIN-LIKE PEPTIDES IN INSECTS. Annu. Rev. Èntomol. 2006, 51, 1–24. [Google Scholar] [CrossRef]
- Wyatt, G. R. & Kale, G. F. 1957. The chemistry of insect hemolymph. II. Trehalose and other carbohydrates. J Gen Physiol, 40(6), pp 833-47.
- Yamada, T.; Habara, O.; Kubo, H.; Nishimura, T. Fat body glycogen serves as a metabolic safeguard for the maintenance of sugar levels in Drosophila. Development 2018, 145, dev–158865. [Google Scholar] [CrossRef]
- Yamada, T.; Habara, O.; Yoshii, Y.; Matsushita, R.; Kubo, H.; Nojima, Y.; Nishimura, T. Role of glycogen in development and adult fitness in Drosophila. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Yang, H. J. , Cui, M. Y., Zhao, X. H., Zhang, C. Y., Hu, Y. S. & Fan, D. 2023. Trehalose-6-phosphate synthase regulates chitin synthesis in Mythimna separata. Front Physiol, 14(1109661.
- Yang, S.; Zhao, Y.; Yu, J.; Fan, Z.; Gong, S.-T.; Tang, H.; Pan, L. Sugar Alcohols of Polyol Pathway Serve as Alarmins to Mediate Local-Systemic Innate Immune Communication in Drosophila. Cell Host Microbe 2019, 26, 240–251. [Google Scholar] [CrossRef]
- Zhang, B.-C.; Jiang, C.-J.; An, C.-J.; Zhang, Q.-W.; Zhao, Z.-W. Variations in Fuel Use in the Flight Muscles of Wing-Dimorphic Gryllus Firmus and Implications for Morph-Specific Dispersal. Environ. Èntomol. 2011, 40, 1566–1571. [Google Scholar] [CrossRef]
- Zhang, T.; Yuan, D.; Xie, J.; Lei, Y.; Li, J.; Fang, G.; Tian, L.; Liu, J.; Cui, Y.; Zhang, M.; et al. Evolution of the Cholesterol Biosynthesis Pathway in Animals. Mol. Biol. Evol. 2019, 36, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, X.; Shi, X.; Karpac, J. Diet-MEF2 interactions shape lipid droplet diversification in muscle to influence Drosophila lifespan. Aging Cell 2020, 19, e13172. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Pennington, J.E.; Wells, M.A. Utilization of pre-existing energy stores of female Aedes aegypti mosquitoes during the first gonotrophic cycle. Insect Biochem. Mol. Biol. 2004, 34, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Fukumura, K.; Nagata, S. Effects of adipokinetic hormone and its related peptide on maintaining hemolymph carbohydrate and lipid levels in the two-spotted cricket, Gryllus bimaculatus. Biosci. Biotechnol. Biochem. 2018, 82, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, R.; Eckart, K.; Law, J.H. Adipokinetic hormone controls lipid metabolism in adults and carbohydrate metabolism in larvae of Manduca sexta. Peptides 1990, 11, 1037–1040. [Google Scholar] [CrossRef]
- Ziegler, R.; Schulz, M. Regulation of lipid metabolism during flight in Manduca sexta. J. Insect Physiol. 1986, 32, 903–908. [Google Scholar] [CrossRef]
- Zinke, I.; Schütz, C.S.; Katzenberger, J.D.; Bauer, M.; Pankratz, M.J. Nutrient control of gene expression in Drosophila: microarray analysis of starvation and sugar-dependent response. EMBO J. 2002, 21, 6162–6173. [Google Scholar] [CrossRef]
Figure 1.
Monosaccharide fates. Glucose, fructose and galactose are the main monosaccharides taken by enterocytes after hydrolysis of dietary carbohydrates. Glucose is metabolized to glycogen or glycolysis. Fructose can enter glycolysis at various steps. Galactose enters this metabolic network as galactose-1P. Glycolytic intermediates may be used by the pentose phosphate pathway (PPP) or the hexosamine pathway.
Figure 1.
Monosaccharide fates. Glucose, fructose and galactose are the main monosaccharides taken by enterocytes after hydrolysis of dietary carbohydrates. Glucose is metabolized to glycogen or glycolysis. Fructose can enter glycolysis at various steps. Galactose enters this metabolic network as galactose-1P. Glycolytic intermediates may be used by the pentose phosphate pathway (PPP) or the hexosamine pathway.
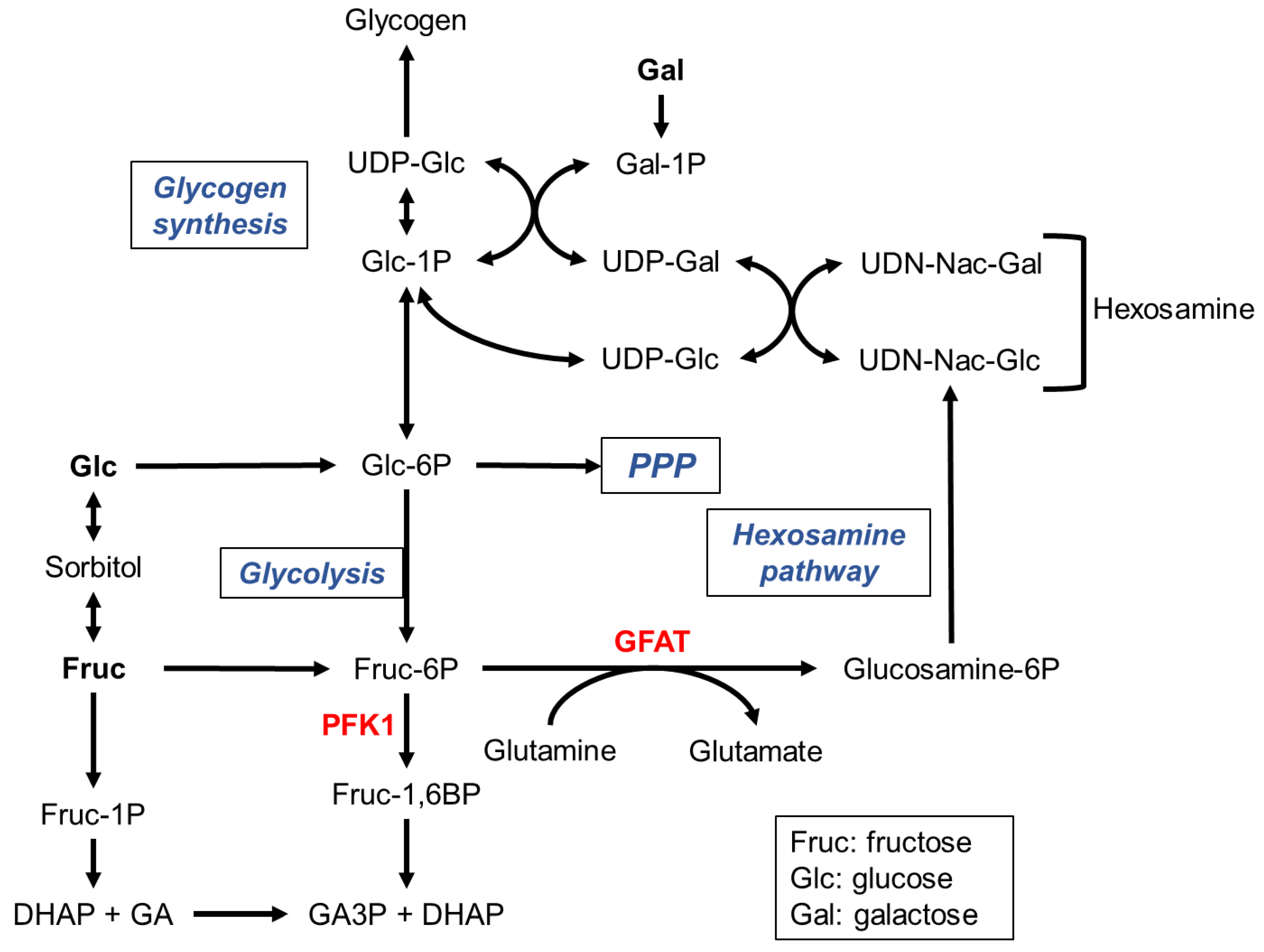
Figure 2.
Connecting glycolysis to lipogenesis. The final glycolytic product pyruvate is either fermented to lactate or oxidized to acetyl-CoA in the mitochondria. Excess of citrate is transferred back to the cytoplasm and used for FA synthesis. The NADPH required for this FA synthesis may be produced by the PPP oxidative phase. Glucose, fructose and trioses phosphate may lead to α-oxoaldehydes that react with amine groups to form AGEs. The toxicity of methylglyoxal (MG) can be neutralized by the glyoxalase complex. G3P is required for TAG synthesis through the Kennedy pathway.
Figure 2.
Connecting glycolysis to lipogenesis. The final glycolytic product pyruvate is either fermented to lactate or oxidized to acetyl-CoA in the mitochondria. Excess of citrate is transferred back to the cytoplasm and used for FA synthesis. The NADPH required for this FA synthesis may be produced by the PPP oxidative phase. Glucose, fructose and trioses phosphate may lead to α-oxoaldehydes that react with amine groups to form AGEs. The toxicity of methylglyoxal (MG) can be neutralized by the glyoxalase complex. G3P is required for TAG synthesis through the Kennedy pathway.
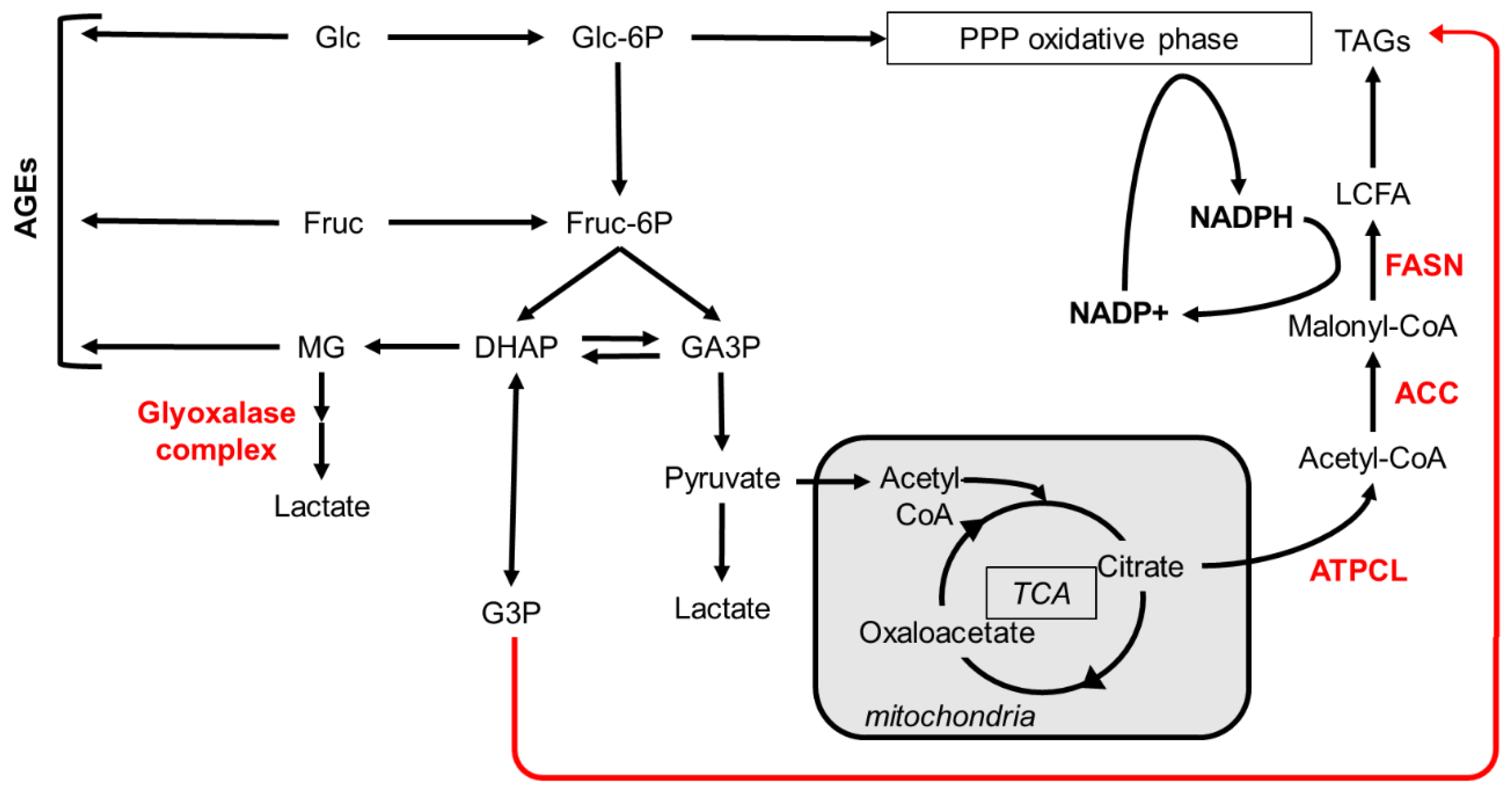
Figure 3.
TAG synthesis pathways. This synthesis may originate from G3P (Kennedy pathway), DHAP, MAG or transacylation of DAGs. Note that newly synthesized FAs are always required.
Figure 3.
TAG synthesis pathways. This synthesis may originate from G3P (Kennedy pathway), DHAP, MAG or transacylation of DAGs. Note that newly synthesized FAs are always required.
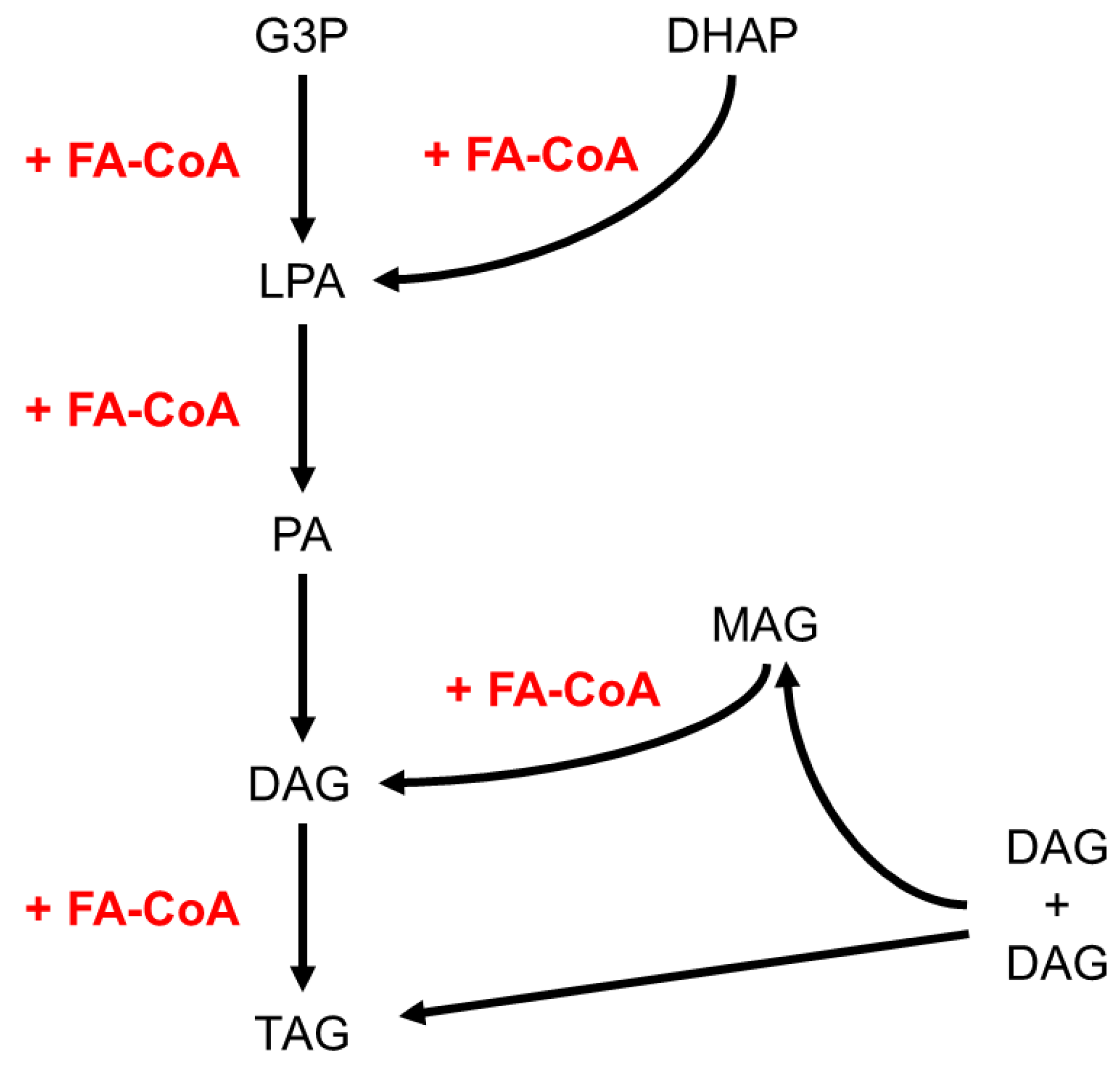
Figure 4.
Metabolism of insect flight muscles. Glucose consumption in flight muscles follows the aerobic glycolytic pathway through mitochondrial respiration; regeneration of the cytoplasmic NAD+ cofactor proceeds through the G3P shuttle. Energy production through anaerobic glycolysis yields high amounts of lactate that is toxic in flight muscles, but is used in locomotion muscles. Long-term flight usually consumes TAG stores through FA β-oxidation.
Figure 4.
Metabolism of insect flight muscles. Glucose consumption in flight muscles follows the aerobic glycolytic pathway through mitochondrial respiration; regeneration of the cytoplasmic NAD+ cofactor proceeds through the G3P shuttle. Energy production through anaerobic glycolysis yields high amounts of lactate that is toxic in flight muscles, but is used in locomotion muscles. Long-term flight usually consumes TAG stores through FA β-oxidation.
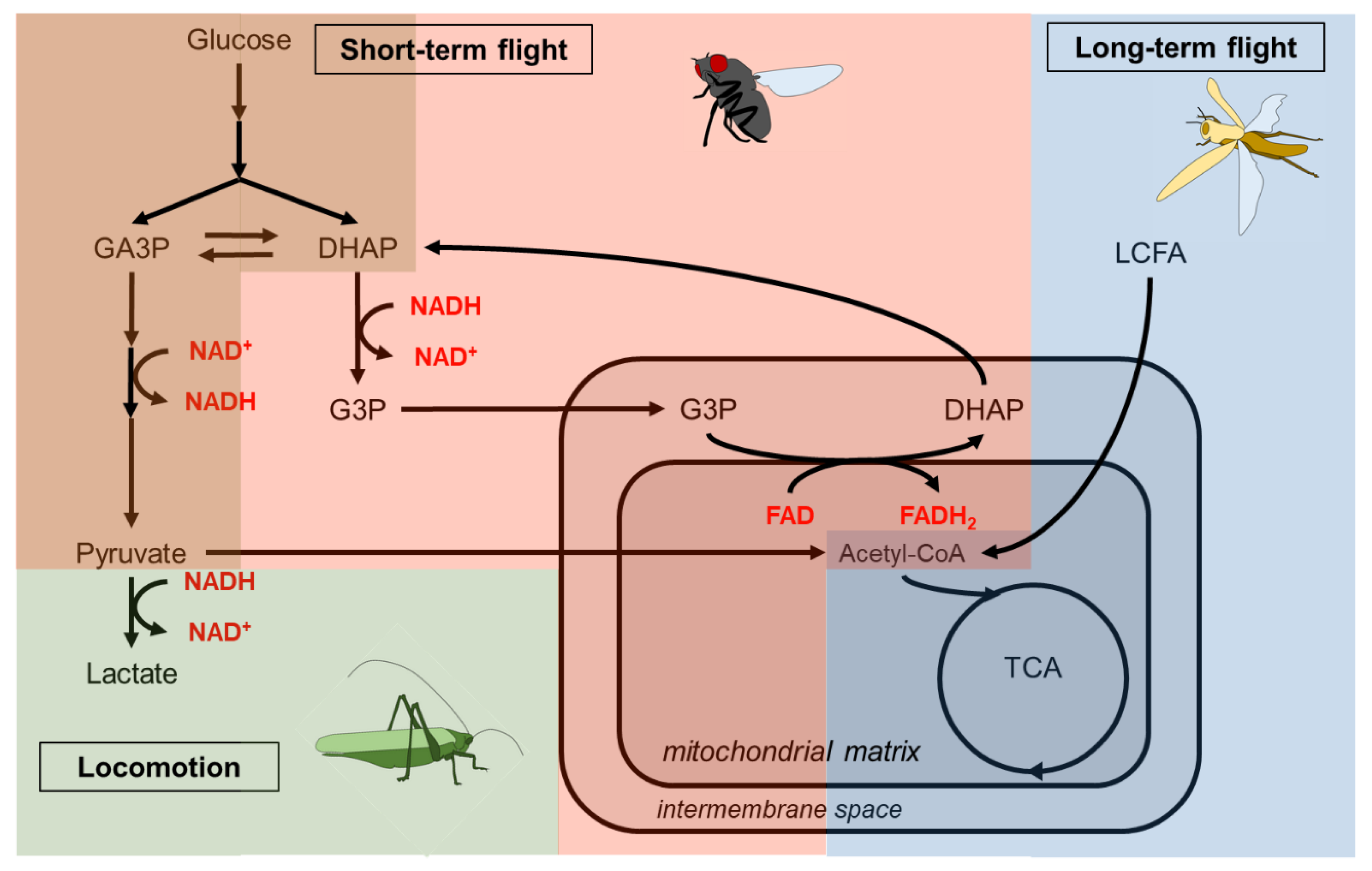

Table 1.
Dietary carbohydrates of insects.
| Carbohydrate | Type | Main dietary source |
|---|---|---|
| Glucose | monosaccharide C6H12O6 | universal |
| Fructose | monosaccharide C6H12O6 | fruits |
| Galactose | monosaccharide C6H12O6 | milk, honey, plant galactolipids |
| Sucrose | 1 glucose + 1 fructose | plant roots, fruits and sap |
| Maltose | 2 glucose | germinating seeds |
| Trehalose | 2 glucose | insect haemolymph |
| Cellobiose | 2 glucose | cellulose digestion |
| Raffinose | 1 galactose + 1 glucose + 1 fructose | plant seeds |
| Melezitose | 2 glucose + 1 fructose | plant sap, honey |
| Glycogen | branched glucose polymer | animal stores |
| Starch | branched glucose polymer | plant stores |
| Inulin | branched fructose polymer | plant stores |
| Cellulose | linear glucose polymer | plant cell walls |
| Hemicellulose | branched monosaccharide polymer | plant cell walls |
| Pectin | galacturonic acid polymer | plant cell walls |
| Glycolipid | monosaccharide + diacylglycerol | cell membranes |
| Galactolipid | galactose + diacylglycerol | thylakoid membranes |
Table 2.
Metabolites for flight muscles in various insect species.
| Insect species | Flight muscle metabolism requirements | Reference |
|---|---|---|
| A. aegypti | proline, pyruvate and G3P | (Gaviraghi et al., 2019) |
| A. gambia | AKH-I dependent haemolymph trehalose | (Kaufmann and Brown, 2008) |
| Bombus impatiens | proline at flight onset shift to carbohydrate | (Stec et al., 2021) |
| B. terrestris | G3P shuttle to pre-warm flight muscle | (Masson et al., 2017) |
| D. melanogaster | Glycolytic metabolites AKH-dependent lipid metabolism Peroxisome-dependent lipid metabolism Carbohydrate |
(Eanes et al., 2006) (Katewa et al., 2012) (Faust et al., 2014) (Zhao et al., 2020) |
| Decapotoma lunata | Haemolymph carbohydrate | (Auerswald and G, 1995) |
| G. molesta | Carbohydrate | (Su et al., 2022) |
| G. firmus | Fat body glycogen, haemolymph trehalose | (Zhang et al., 2011) |
| L. migratoria | Trehalose and lipids Lipids, high FABP levels in flight muscles |
(Robinson and Goldsworthy, 1977) (van der Horst et al., 1993) |
| M. sexta | Haemolymph lipids FA oxidation shifts to glycolysis with aging |
(Ziegler and Schulz, 1986) (Wone et al., 2018) |
| Pachnoda sinuata | Proline and haemolymph carbohydrates | (Auerswald et al., 1998) |
| P. megistus | Carbohydrate first, shift to lipids | (Canavoso et al., 2003) |
| P.americana | Glycogen High expression of glycogen phosphorylase |
(Downer and Matthews, 1976) (Steele, 1982) |
| R. prolixus | Lipids | (Braz et al., 2023) |
| S. gregaria | Carbohydrate first, shift to lipids | (Rajapakse et al., 2019) |
| Orchid bees | Carbohydrate | (Suarez et al., 2005) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



