Submitted:
22 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
The objective of this paper was to study the expression pattern of the GLOD4 gene in the testis and its regulatory effect on testicular cells. Currently, the regulated mechanism through are unclear. Here, we explored how GLOD4 functions in goat leydig cells to enhance our understanding of spermatogenesis and improve reproduction in breeding rams. In this study, we demonstrated that the localization of GLOD4 in testicular cells using immunohistochemistry and subcellular localization analyses. Subsequently, we analyzed the GLOD4 expression pattern in 4 age-based groups (0, 6, 12, and 18 months old) using real-time quantitative polymerase chain reaction (qRT‒PCR) and protein blotting. Finally, we performed GLOD4 gene silencing and overexpression studies in testicular cells from goats and explored the effects of GLOD4 on cell proliferation, the cell cycle, steroid hormone secretion and the expression of candidate testosterone hormone-regulated genes. GLOD4 was mainly expressed in testicular leydig cells, and the subcellular localization results showed that the GLOD4 protein was mainly localized in the cytoplasm and nucleus. Silencing of GLOD4 significantly suppressed the mRNA expression levels of the testosterone secretion-related genes CYP11A1, 3β-HSD and CYP17A1 and the mRNA expression levels of cell-cycle related genes CDK6, PCNA, and Cyclin E. Moreover, the cell cycle was blocked at the G2/M phase after GLOD4 silencing, which significantly suppressed testosterone secretion. In contrast, GLOD4 overexpression significantly increased the mRNA expression levels of the testosterone secretion-related genes CYP11A1, 3β-HSD, and CYP17A1 and increased the expression of the cell-cycle related genes CDK6, PCNA, and Cyclin E. Moreover, GLOD4 overexpression promoted the cell cycle from G0/G1 phases to enter the S phase and G2/M phases,promoted the secretion of testosterone. Taken together, our experimental results indicate that GLOD4 may affect the development of cells in Qianbei Ma goats of different ages by influencing the cell cycle, Proliferation and testosterone hormone synthesis. These findings enhance our understanding of the functions of GLOD4 in the cells of goats.
Keywords:
Qianbei Ma goat
; Testosterone
; Proliferation
; Leydig cell
; GLOD4
1. Introduction
In the testis, spermatogenesis begins with the mitotic division of spermatogonia in the seminiferous tubules. Spermatogonia undergoes two meiotic divisions to form spermatocytes, which subsequently metamorphose, continue to develop and mature in the seminiferous tubules, and ultimately undergo a complex series of changes to produce mature spermatozoa. In addition, it is regulated by many highly complex genes[1,2,3,4,5]. Spermatogenesis is a highly complex and organized process that occurs in the testis. Another important biological function of the testis, which is an important male reproductive organ, is androgen secretion. For example, testosterone increases the libido and sexual behavior, improves spermatogenesis and sperm quality in male animals, increases the success rate of fertilization[6], and can induce the testicular tubules to produce more sperm and support sperm maturation and motility[4,7]. In addition, estrogen and other hormones such as luteinizing hormone (LH) and follicle stimulating hormone (FSH) are indispensable throughout spermatogenesis[8,9].
The glyoxalase gene family is an important regulatory family consisting of six structurally and functionally distinct enzymes, including glyoxalase domain-containing protein 4 (GLOD4). GLOD4, also known as HC6-type or HC71-type, was originally cloned as the C17orf25 cDNA from human hepatocellular carcinoma[10,11]. GLO1 is an ancient analog of GLOD4 located at Chr 17p13.3 and consists of 13 exons[12]. The human GLOD4 gene region is approximately 23 kb long and contains 10 exons and 9 introns[13]. The full-length cDNA sequence of human GLOD4 is 1814 bp long and encodes an open reading frame (ORF) consisting of 313 amino acidsTreatment of mouse embryonic stem cells with cyclopiroxamine (CPX) results in the altered expression of multiple proteins, whereas downregulation of GLOD4 protein expression may inhibit cell proliferation and block the cell cycle[14]. In a canine model of retinal degeneration caused by mutations in the RPGRORF15 gene, researchers analyzed the expression profiles of mutant and normal retinas. In conjunction with other genes, GLOD4 was found to have an effect on cell-cycle progression and to potentially influence processes related to mitochondrial function and cellular energy metabolism[15]. In Blmh (Bleomycin Hydrolase) -/-mice, GLOD4 isoform 3 mRNA expression was decreased in the kidney but no effect on isoform 1 mRNA expression was observed. This suggests that GLOD4 protein expression may be subjected to Blmh+/+-mediated posttranscriptional regulation, which, in turn, has a detoxification effect[16]. Using the cryptic nematode Caenorhabditis elegans (C. elegans) as a model to explore the role of the TRPA1-Nrf signaling pathway in methylglyoxal detoxification revealed that it regulates the expression of GLOD4. This study also emphasized the potential role of GLOD4 in human diseases, especially in pathologies associated with α-diketone bodies (α-DCs) accumulation, such as diabetes and neurodegenerative diseases[17]. However, current GLOD4 research still focuses mainly on humans and rodents, with few reports on goats.
Testicular leydig cells are in the interstitial tissue of the testes and play an important role in the male reproductive system. One of the main functions of leydig cells is to secrete testosterone, a hormone that stimulates sperm production and development within the testes[18,19]. Since the peak of testosterone secretion mainly occurs during the reproductive and youthful stages of the animal, testosterone secretion also has an indispensable role in muscle protein synthesis, calcium uptake and bone density maintenance during these developmental stages. In addition to testosterone secretion, leydig cells also regulate the proper testis temperature and the Proliferation and function of different cell types within the testis, thus ensuring normal spermatogenesis[20].
As a domesticated animal with important economic value, the economic impact of goats can be increased by improving the fertility of breeding rams and the efficiency of goat breeding[21]. Recent studies have revealed that the GLOD4 gene is differentially expressed in leydig cells of Tibetan sheep at different developmental stages[22]. However, the mechanism by which GLOD4 gene expression affects leydig cells in goats has rarely been reported, especially within the context of testicular development. As a goat breed that is well adapted to the highland mountain environment, Qianbei Ma goats have a long history of breeding in Guizhouwith a medium body size, rough feeding resistance, strong disease resistance, good fur quality and strong fertility. Thus, Qianbei Ma goats are a good model animal for exploring the reproductive traits of plateau goats[23]. Therefore, the aim of this study was to investigate the profile of GLOD4 gene expression in the testes of goats at different ages and to analyze the regulatory effect of the GLOD4 gene on testicular leydig cells. We first silenced and overexpressed the GLOD4 gene in the testicular leydig cells of goats and investigated the effects of the GLOD4 gene on the cells including on the cell cycle and testosterone hormone secretion. By analyzing the effects of the GLOD4 gene expression on testicular leydig cells, we established a foundation for understanding the potential regulatory mechanism through which the GLOD4 gene functions in spermatogenesis and provided a theoretical basis for improving the reproductive traits of breeding rams.
2. Materials and Methods
The experimental protocol used in this study has been approved by the Animal Protection and Use Committee of Guizhou University, Guiyang, China (Approval number: EAE-GZU-2021-E021). The animal handling procedures were in line with the Chinese Animal Welfare Guidelines and were approved by the Animal Protection and Use Committee of Guizhou University, Guiyang, China (Approval number: EAE-GZU-2021-E021).
2.1. Tissue Collection
Experiment animals and sampling A total of 20 healthywere randomly selected from Xishui County, Guizhou Province, China, Fuxing Dairy Company Limited, and divided into four age groups (n = 5 for each age group): 0Y, 0 month old (before sexual maturity, body weight: 1.66 kg ± 0.5 kg); 6M, 6 months (after sexual maturity, body weight: 30.30 kg ± 1.0 kg); 12M, 12 months old (after physical maturity, body weight: 43.32 kg ± 1.5 kg); 18M, 18 months (sell on the market,body weight:47.53 kg±2.0 kg). The left testicular tissue was excised immediately after anesthesia, collected with sterile scissors, trimmed to approximately 5 mm testicular tissue, and washed in phosphate-buffered saline solution within 20 min, with three biological replicates per age. After clipping the testicles of the goats, the wounds were sterilized, and the goats were continued with subsequent feeding. After the experiment, the animals survived normally. One part of the sample was fixed in 4% paraformaldehyde solution for H&E staining, and the remainder was rapidly frozen in liquid nitrogen and subsequently transferred to 80C for storage in lab for other analyses such as extraction of total RNA and total protein.
2.2. Cell Culture and Transfection
Fine leydig cells was obtained from 12-month-old goat testes and cultured as previously described[24]. Briefly, testicular tissue from goats was immediately transferred to Dulbecco’s phosphate-buffered saline (DPBS) supplemented with 5% penicillin/streptomycin (Gibco, Grand Island, NY, USA) and incubated at 4°C. The white membrane on the surface of the testis was removed with eye clippers and then digested with 0.5 mg/mL collagenase IV (Sigma, St. Louis, MO, USA) in DMEM/F12 (Gibco, Grand Island, NY, USA) at 37°C for 20 min. The dispersed cells were then filtered using a 100 mm cell strainer (Biosharp, Shanghai, China), and the filtrate was centrifuged at 1200 × g for 5 min. The supernatant was discarded, and the cells were washed three times in DPBS and centrifuged at 1200 × g for 5 min and resuspended. The resuspended cells were purified by the differential applanation method. The harvested leydig cells (LCs) were cultured in DMEM/F12 containing 10% fetal bovine serum (Gibco, China, Beijing, China) and 1% penicillin/streptomycin and incubated in a cell culture incubator (Thermo Fisher Scientific, Waltham, MA, USA) at 37°C with 5% CO2. Once the confluence reached 75-90%, the cells were evenly divided between 6 empty plates (NEST Biotechnology Co., Ltd., Wuxi, China). These cells were then transfected with GLOD4 overexpression and silencing plasmids using a Lipofectamine 2000 kit (Thermo Fisher Scientific Waltham, MA, USA). After 48 h, RNA luminescence was assessed as good and the cells were harvested to conduct all other experiments.
2.3. Pasmid Construction
After PCR amplification, amplicons were subjected to gel purification to obtain the desired target sequences, which were then ligated to pcDNA3.1 using T4 DNA ligase. Plasmids were amplified in transformed DH5α competent cells. Then, the desired plasmids were extracted from the cells by oscillation. The PCR reaction (25 µL) consisted of 1 µL of DNA template, 1 µL of each of the upstream and downstream primers (10 pmol/µL), 12.5 µL of Green Mix, and 9.5 µL of ddH2O. shRNA-GLOD4 interfering sequences were designed and synthesized by Shanghai Gemma Pharmaceuticals & Biologicals (see Table 1 for details).
2.4. Total RNA Was Extracted and Reverse-Transcribed
Total RNA was extracted from goat testicular tissue and testicular leydig cells using TRIzol reagent (Solarbio, Beijing, China). Subsequently, first-strand cDNA was performed using the StarScript II First-Strand cDNA Kit (GenStar, Beijing, China) according to the manufacturer’s instructions, and the reaction products were stored at -20°C.
2.5. Real-Time Polymerase Chain Reaction
The expression levels of the GLOD4 gene, and that of other genes, was measured in goat leydig cells on a CFX 9600 real-time PCR instrument (Bio-Rad, Hercules, CA, USA). The details of the real-time PCR primers used are summarized in Table 2. A 10 µL real-time PCR mixture (containing 5 µL of 2× Es-Taq Master mix, 3.2 µL of RNase-free ddH2O, 0.5 µL of cDNA, and 0.4 µL of forward and reverse primers (10 pmol/µL)) was prepared according to the manufacturer’s instructions and subjected to a three-step PCR program (95°C for 2 min, 95°C for 15 s, annealing temperature (see Table 2 for details) for 30 s, and 72°C for 30 s; the melting curve was generated automatically by the machine). The PCR assay was repeated three times for each test sample. The expression data were normalized to the expression of β-actin, which we used as an internal reference. The relative change in gene expression was then calculated using the relative quantification method 2-∆∆CT. β-actin primers were synthesized by Bioengineering (Shanghai) Co. (see Table 2 for details).
2.6. Western-Blot
Testicular tissues from different developmental stages were homogenized and lysed using a radioimmunoprecipitation assay (RIPA) protein extraction kit (Solarbio, Beijing, China) with the addition of phenylmethylsulfonyl fluoride (PMSF) (Solarbio, Beijing, China). Protein concentrations were quantified using a commercial bicinchoninic acid (BCA) protein assay (Solarbio, Beijing, China). Protein samples were separated on a 10% SDS‒PAGE gel and then transferred to a polyvinylidene fluoride (PVDF) membrane by wet transfer. After being blocked in 5% skim milk for 2 h, the membranes were incubated overnight with rabbit anti-GLOD4 polyclonal antibody (1:1000 dilution, Bioss, Beijing, China) at 4°C. The membranes were then washed three times in Tris-buffered saline containing Tween-20 (TBST), incubated with an HRP-conjugated secondary antibody (1:10,000 dilution) for 1 hr in TBST, washed again, and then incubated with an HRP-conjugated secondary antibody (1:10000 dilution, Bioss, Beijing, China) and visualized using an NcmECL Ultra kit (P10200, NCM, Suzhou, China) on a Universal Hood II instrument (Bio-Rad, CA, USA). The grayscale value of each band was analyzed using ImageJ software.
2.7. Immunohistochemistry
For immunohistochemistry, after the prepared paraffin sections of testis were dewaxed in water, they were boiled in 0.1 M citrate buffer (pH = 6.0) for 15 min in a microwave oven and then cooled to 37°C for antigen retrieval. The sections were subsequently washed with phosphate-buffered saline (PBS) and incubated in 3% hydrogen peroxide for 15 min to block endogenous peroxidase activity. The sections were further incubated in blocking buffer (5% bovine serum albumin (BSA) in PBS) for 15 min at 37°C to block nonspecific binding. The membranes were then incubated with rabbit anti-GLOD4 antibody (1:1000; Bioss Biotechnology Co., Ltd., Beijing, China) in PBS overnight, washed with PBS, incubated with the biotinylated secondary antibody for 2 h at 37°C and with horseradish peroxidase (HRP)-streptavidin for 15 min, and then visualized using diaminobenzidine. Nonspecific rabbit immunoglobulin G (IgG) was used instead of the primary antibody as a negative control.
2.8. Subcellular Localization
For transfection, cell lysates were added before the cells were inoculated into six-well plates, and after 36 h of transfection, the cells were washed 3 times with PBS and fixed at room temperature for 20 min by the addition of precooled 4% paraformaldehyde (Solarbio, Shanghai, China); the cells were washed 3 times with PBS and fixed at room temperature for 5 min by the addition of 0.25% Triton X-100 (Biosharp Anhui, China); to conduct all other experiments, and the nuclei were stained with DAPI (Solarbio, Shanghai, China) to stain the cells; and the cells were washed 3 times with PBS. The fluorescence images were merged using Photoshop, and the subcellular localization of the fusion protein was assessed based on the overlap between the fluorescence of the fusion protein and the fluorescence of the cell nucleus.
2.9. Cell Proliferation Analysis
A CCK-8 assay was used to detect the effect of GLOD4 gene overexpression and silencing on the proliferative activity of leydig cells. Plasmids from the negative control and experimental groups were incubated at 37°C and 5% CO2 for 6, 12, 24, 48 and 72 h after transfection, as described in the instructions of the CCK-8 assay kit (Monmouth Junction, NJ, USA). The absorbance (OD) at 450 nm was subsequently measured using enzyme markers (Thermo Fisher Scientific, Waltham, MA, USA).
2.10. Steroid Assay
The culture medium was collected to measure the testosterone concentration using an ELISA kit (Kamels, Shanghai, China) according to the manufacturer’s instructions[25]; the absorbance of the samples was measured at 450 nm, and the concentration of testosterone was determined using a standard curve.
2.11. Flow Cytometry Analysis
Harvested cells were fixed in 70% ice-cold ethanol overnight at 20°C. After washing with PBS, the cells were incubated in 0.5 mg/mL RNase for 30 min at 35°C and stained with 0.025 mg/mL propidium iodide for 10 min. Finally, cells at different stages of the cell cycle were identified by flow cytometry analysis (BD Biosciences, Canto II plus, USA). The data from at least 20,000 cells were collected per sample.
2.12. Statistics
Statistical analyses were performed using Graphpad prism10. All trials were repeated at least three times, and the data were presented as means ± SD. Comparisons between two independent groups were determined by t-test; meanwhile, the differences among multiple groups were analyzed by one-way ANOVA or Two-way ANOVA. (significant, P < 0.05; highly significant, P < 0.01).
3. Results
3.1. Localisation of GLOD4 in the Testis of Qianbei Ma Goats
We detected GLOD4-positive protein signals by immunohistochemistry in testicular tissue sections from 12-month-old rams and found that the immunopositivity was confined to testicular leydig cells (Figure 1A). After transfection of the pcDNA3.1-NC vector containing GFP fluorescence, the subcellular localization results showed that GLOD4 was highly expressed in the nucleus and cytoplasm of leydig cells. (Figure 1B).
3.2. Expression Patterns of the GLOD4 Transcript at Different Developmental Stages of Qianbei Ma Goats Testes
Both the Western blot and qRT‒PCR showed that GLOD4 was expressed in the testes of Qianbei Ma goats at 0, 6, 12 and 18 months of age, and GLOD4 expression increased with age. GLOD4 protein expression was significantly higher at 18 months of age than at 0 months of age (p<0.01) and that at both 6 and 12 months of age (p<0.05). The GLOD4 mRNA expression was significantly higher at 6, 12 and 18 months of age than at 0 months of age (p<0.01; Figure 2A,B).
3.3. GLOD4 Promotes Qianbei Ma Goat LCs Proliferation In Vitro
To investigate how GLOD4 affects leydig cells, we transfected cells with GLOD4 silencing and overexpression vectors and then conducted mRNA and protein assays. After GLOD4 silencing and, both the mRNA and protein levels of GLOD4 were significantly downregulated and upregulated (p<0.01 each), respectively, compared to those in the control group (sh-NC, pcDNA3.1-NC) (Figure 2C–F). Furthermore, we performed a CCK-8 assay at 6, 12, 24, 48 and 72 h after transfection, which revealed that silencing of GLOD4 inhibited the proliferation of leydig cells, whereas the GLOD4 overexpression significantly promotes cell proliferation. (Figure 2H,I). In addition, we examined the expression of proliferating cell nuclear antigen (PCNA) and found that GLOD4 interference and overexpression resulted in the downregulation and upregulation of PCNA, respectively. (Figure 2G).
3.4. GLOD4 Promotes Cell Cycle Progress of Qianbei Ma Goat LCs In Vitro
After exploring the effect of the GLOD4 gene on the cells, we wanted to determine how GLOD4 regulates the cell cycle in leydig cells. We found that the percentage of cells in the G2/M phase was (p<0.05) lower than that in the control group after GLOD4 silencing and expression of the cell cycle-regulated protein kinase (CDK6) and Cyclin E (Cyclin E) was significantly reduced in these cells (p<0.01; Figure 3A,B). In contrast, overexpression of the GLOD4 gene significantly reduced the percentage of cells arrested in the G0/G1 phase and increased the percentage of cells in the G2/M phase compared to those in the control group, and highly significantly (p<0.01) upregulated the expression levels of CDK6 and Cyclin E (Figure 3B,C).
3.5. Promotion of Testosterone Hormone Secretion by GLOD4
After GLOD4 silencing, leydig cells secreted less testosterone (p<0.01) than that in the controls at 48 and 72 h, and the expression of 3β-HSD (p<0.05), CYP11A1 (p<0.01), and CYP17A1 (p<0.01) significantly decreased (Figure 4A,C). In addition, after overexpression of GLOD4, leydig cells secreted(p<0.01) more testosterone hormone than did the control group at 48 h and 72 h, significantly increased expression of 3β-HSD (p<0.01) and CYP17A1 (p<0.05) and had no effect on CYP11A1 expression (Figure 4B,D).
4. Discussion
Previous reports on GLOD4 have focused on its metabolic function and detoxification properties[17,26,27], although some have reported that GLOD4 may have certain effects on the cell cycle and other cellular functions[14,28]. However, few studies have explored the role of GLOD4 in reproduction. Therefore, by exploring GLOD4 expression in the testes of Qianbei Ma goats and its effect on leydig cells, we provided an in depth analysis of the function of GLOD4 and provided a theoretical basis for improving ram fertility.
Wang et al. noted that GLOD4 expression was significantly upregulated in the testis of Tibetan sheep wwith age, and GLOD4 was only expressed in leydig cells and that GLOD4 was mainly observed in the cytoplasm[23], our immunohistochemistry revealed that GLOD4 was expressed only in leydig cells, which is consistent with the findings of Wang et al. However, our subcellular localization analysis revealed that GLOD4 was widely expressed in both the nucleus and the cytoplasm of leydig cells. Interestingly, in a recent study, fluorescently tagged GLOD4 protein was widely distributed in the cytoplasm and nucleus of human retinal pigment epithelial cells (hTERT-RPE1)[28], which may result from differential gene expression in different species[29,30,31].
Based on the differential expression of GLOD4 at different testicular development stages and GLOD4 expression being confined to leydig cells, we hypothesized that the observed increase in the GLOD4 expression might relate to the continuous development of the testis. To date this, we isolated and cultured cells extracted from testes to further explore GLOD4 function. Previously, using a yeast two-hybrid method, Zhang et al. screened for NUDT9 gene interactors and reported that GLOD4 may inhibit cell growth by interacting with NUDT9[32]. In a recent report on GLOD4, GLOD4-deficient Hidradenitis elegans nematode worms were shown to exhibit increased α-DCs, as hortened lifespan, increased neuronal damage, and tactile sensitization, indicating that GLOD4 expression has a protective effect on the cells and ensures cellular proliferation[27]. After successfully constructing GLOD4 interference and overexpression models, and considering that GLOD4 may act with other genes to influence cell Proliferation[14,15], we performed a CCK-8 assay in leydig cells. We found that GLOD4 gene silencing significantly inhibited cell proliferation at two time points and downregulated the mRNA expression of the cellular nuclear antigen (PCNA), which is a proliferation marker. PCNA is widely distributed in a variety of cells, reflecting not only their proliferation rate but also affecting cellular Proliferation as a key gene[12,33]. In contrast, overexpression of GLOD4 had a significant effect on almost every time and significantly upregulated the expression of PCNA. Therefore, we speculate that GLOD4 may play an important regulatory role in cell function.
The cell cycle, which is regulated by the cell cycle proteins CDK and CKI, controls cell growth and plays a key role in cell proliferation[34,35,36]. Our results showed that both GLOD4 silencing and overexpression significantly inhibited and promoted the progression of the cell cycle from the S phase to the G2/M phase, respectively. These results side-step the assertion of Dihazi et al., in which they assert that downregulation of GLOD4 protein expression may inhibit cellular proliferation and block the cell cycle in mouse embryonic stem cells[14]. In a genome-wide study of Chlamydomonas reinhardtii cilia, Albee Alison et al. identified GLOD4 as one of the genes whose expression is upregulated during ciliogenesis. Moreover, knockdown of GLOD4 in human retinal pigment epithelial cells (hTERT-RPE1) results in shorter cellular cilia and cell cycle progression defects[28]. Cyclin-dependent kinase 6 (CDK6), a key gene in the CDK family, regulates cell cycle progression from G1 to S phase by phosphorylating retinoblastoma (RB) proteins when it is complexed with cyclin D[37]. In addition, Cyclin E performs a similar function via regulating the cellular transition from the G0/G1 phase to the S phase to promote cell cycle progression[38]. Thus, examining CDK6 and Cyclin E gene expression and found that silencing and overexpression GLOD4 significantly downregulated and upregulated both CDK6 and Cyclin E, respectively could be reasonable. In addition, overexpression of GLOD4 accelerated the progression from G0/G1, and we speculated that CDK6 and Cyclin E might play a role in this. Therefore, we speculate that GLOD4 may regulate cell proliferation by affecting the cell cycle.
Taken together, GLOD4 may be crucial for leydig cell proliferaiton, but how GLOD4 function in the regulation of testosterone secretion remains unknown. To address this, we examined testosterone secretion in cells cultured for different durations and the expression of genes involved in testosterone secretion. It is well known that in mammalian testes, the testicular cellular receptor (SCARB1) binds specifically to the raw material for androgen synthesis[39], and a series of cascade reactions stimulate the expression and phosphorylation of steroidogenic acute regulatory protein (STAR)[40]. In turn, STAR catalyzes the substrate of the cytochrome P450 cholesterol side chain cleavage enzyme (CYP11A1) reaction and converts it to pregnenolone, which is then catalyzed to progesterone by the micronutrient enzyme 3B-hydroxysteroid dehydrogenase 1 (3β-HSD1) in the smooth endoplasmic reticulum[41]. Progesterone is ultimately converted to testosterone by a series of steroid synthases[42,43,44]. These functional genes play a critical role in regulating testosterone secretion. 3β-HSD and CYP17A1 were significantly downregulated and upregulated, respectively, by GLOD4 suppression and overexpression. In addition, the suppression and overexpression of GLOD4 decreased and increased testosterone secretion in leydig cells at 48 and 72 h, respectively. These results suggest that GLOD4 influences testosterone secretion by affecting the expression of a range of genes involved in steroid synthesis.
5. Conclusions
In summary, GLOD4 was differentially expressed in the testes of goats at different ages. In addition, after the silencing and overexpression of GLOD4 in goats, the proliferation of leydig cells was decreased and increased, respectively; similar effects were observed on the cell cycle; and testosterone secretion was significantly decreased and increased respectively. Therefore, we speculate that GLOD4 may regulate testicular development and spermatogenesis by increasing the number of leydig cells and regulation of testosterone secretion, indicating that GLOD4 plays a key role in spermatogenesis by affecting the fate of LCs in goats. Although our study confirms the value of exploring the role of GLOD4 in reproduction, there remains a large gap in studies exploring the role of this gene in reproduction, and larger studies with longer follow-up periods are needed to explore the mechanism by which GLOD4 regulates testicular development in vitro and in vivo.
Author Contributions
Jinqian Wang and Xiang Chen conceived and designed the experiments; Jinqian Wang and Jiajing Chen conducted the experiments; Jinqian Wang and Xiang Chen analyzed the data; Jinqian Wang wrote the paper; Jinqian Wang and Wei Sun revised the paper.
Funding
This research was funded by the National Natural Science Foundation of China (32260835), the Science and Technology Project of Guizhou Province (Qian Kehe Foundation—ZK [(2021)] General 151), and the Guizhou High-Level Innovative Talents Project (Qian Kehe Platform Talents [(2022)] 021-1).
Institutional Review Board Statement
Institutional Review Board Statement: The animal handling procedures were in line with the Chinese Animal Welfare Guidelines and were approved by the Animal Protection and Use Committee of Guizhou University, Guiyang, China (Approval number: EAE-GZU-2021-E021).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank X. Chen for their help and support, and Guizhou University for providing the experimental platform. This work was supported by the National Natural Science Foundation of China (32260835), Guizhou Science and Technology Program (Qiankehe Fund-ZK [(2021)] General 151), and Guizhou High-level Innovative Talent Program (Qiankehe Platform Talent [(2022)]021-1).
Conflicts of Interest
There is no conflicts of interest among all authors.
Appendix B
Here are some files related to the raw data of the experiment:Western-blot images(PPTX) 、CCK-8(XLSX)、Relative gene expression QRT-PCR(XLSX)、protein gray value(XLSX)、Plasmid transfection(PPTX)、Immunohistochemistry images(PPTX)、flow cytometry datas(XLS)、subcellular localization(PPTX)、flow cytometry-sh-NC(PPTX)、flow cytometry-sh-GLOD4(PPTX)、flow cytometry-pcNDNA3.1-NC(PPTX)、flow cytometry-pcNDNA3.1-GLOD4(PPTX)、Animal welfare protocol1(PDF)、Application form for Animal Ethical Review(PDF)、Cover letter(PDF)、Declaration of Interest Statement(PDF).
References
- Hess, R.A.; Renato, D.F.L. Spermatogenesis and cycle of the seminiferous epithelium. Asian J. Androl. 2008, 636, 1–15. [Google Scholar] [CrossRef]
- Staub, C.; Johnson, L. Review: Spermatogenesis in the bull. Animal 2018, 12, s27–s35. [Google Scholar] [CrossRef]
- Chalmel, F.; Rolland, A.D. Linking transcriptomics and proteomics in spermatogenesis. Reproduction 2015, 150, R149–R157. [Google Scholar] [CrossRef] [PubMed]
- Mruk, D.D.; Cheng, C.Y. The Mammalian Blood-Testis Barrier: Its Biology and Regulation. Endocr. Rev. 2015, 36, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Chen, W.; Cheng, Z.; Wu, S.; He, J.; Han, L.; He, Z.; Qin, W. Novel Gene Regulation in Normal and Abnormal Spermatogenesis. Cells 2021, 10, 666. [Google Scholar] [CrossRef]
- Grande, G.; Barrachina, F.; Soler-Ventura, A.; Jodar, M.; Mancini, F.; Marana, R.; Chiloiro, S.; Pontecorvi, A.; Oliva, R.; Milardi, D. The Role of Testosterone in Spermatogenesis: Lessons From Proteome Profiling of Human Spermatozoa in Testosterone Deficiency. Front. Endocrinol. 2022, 13, 852661. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Chen, S.; Chen, Y.; Yuan, X.; Dong, H. Testicular fat deposition attenuates reproductive performance via decreased follicle-stimulating hormone level and sperm meiosis and testosterone synthesis in mouse. Anim. Biosci. 2024, 37, 50–60. [Google Scholar] [CrossRef]
- Schulster, M.; Bernie, A.M.; Ramasamy, R. The role of estradiol in male reproductive function. Asian J. Androl. 2016, 18, 435–440. [Google Scholar] [CrossRef]
- Rosati, L.; Di Fiore, M.M.; Prisco, M.; Di Giacomo, R.F.; Venditti, M.; Andreuccetti, P.; Chieffi, B.G.; Santillo, A. Seasonal expression and cellular distribution of star and steroidogenic enzymes in quail testis. Journal of Experimental Zoology. Part B, Molecular and Developmental Evolution 2019, 332, 198–209. [Google Scholar] [CrossRef]
- Qin, W.X.; Wan, F.; Sun, F.Y.; Zhang, P.P.; Han, L.W.; Huang, Y.; Jiang, H.Q.; Zhao, X.T.; He, M.; Ye, Y.; et al. Cloning and characterization of a novel gene (C17orf25) from the deletion region on chromosome 17p13.3 in hepatocelular carcinoma. Cell Res. 2001, 11, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Farrera, D.O.; Galligan, J.J. The Human Glyoxalase Gene Family in Health and Disease. Chem. Res. Toxicol. 2022, 35, 1766–1776. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.Y.; Xu, J.; Mao, Q.; Fu, L.L.; Gu, J.R.; De Zhu, J. The promoter analysis of the human C17orf25 gene, a novel chromosome 17p13.3 gene. Cell Res. 2002, 12, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic. Acids. Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef] [PubMed]
- Dihazi, G.H.; Bibi, A.; Jahn, O.; Nolte, J.; Mueller, G.A.; Engel, W.; Dihazi, H. Impact of the antiproliferative agent ciclopirox olamine treatment on stem cells proteome. World J. Stem Cells 2013, 5, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Genini, S.; Zangerl, B.; Slavik, J.; Acland, G.M.; Beltran, W.A.; Aguirre, G.D. Transcriptional profile analysis of RPGRORF15 frameshift mutation identifies novel genes associated with retinal degeneration. Invest. Ophthalmol. Vis. Sci. 2010, 51, 6038–6050. [Google Scholar] [CrossRef] [PubMed]
- Suszynska-Zajczyk, J.; Wroblewski, J.; Utyro, O.; Luczak, M.; Marczak, L.; Jakubowski, H. Bleomycin hydrolase and hyperhomocysteinemia modulate the expression of mouse proteins involved in liver homeostasis. Amino Acids 2014, 46, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, J.; Bose, N.; Gong, J.; Hall, D.; Rifkind, A.; Bhaumik, D.; Peiris, T.H.; Chamoli, M.; Le, CH.; Liu, J.; et al. A Caenorhabditis elegans Model Elucidates a Conserved Role for TRPA1-Nrf Signaling in Reactive alpha-Dicarbonyl Detoxification. Curr. Biol. 2016, 26, 3014–3025. [Google Scholar] [CrossRef]
- Heinrich, A.; Defalco, T. Essential roles of interstitial cells in testicular development and function. Andrology 2020, 8, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Bittman, E.L. Timing in the Testis. J. Biol. Rhythms 2016, 31, 12–36. [Google Scholar] [CrossRef]
- Mendis-Handagama, S.M.; Ariyaratne, H.B. Effects of thyroid hormones on Leydig cells in the postnatal testis. Histol. Histopathol. 2004, 19, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Mueller, J.P.; Getachew, T.; Rekik, M.; Rischkowsky, B.; Abate, Z.; Wondim, B.; Haile, A. Converting multi-trait breeding objectives into operative selection indexes to ensure genetic gains in low-input sheep and goat breeding programmes. Animal 2021, 15, 100198. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, T.; Liu, N.; Zhang, H.; Zhao, X.; Ma, Y. Characterization of GLOD4 in Leydig Cells of Tibetan Sheep During Different Stages of Maturity. Genes 2019, 10, 796. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, X.; Zhou, Z.; Tian, X.; Yang, P.; Fu, K. CYP19A1 May Influence Lambing Traits in Goats by Regulating the Biological Function of Granulosa Cells. Animals 2022, 12, 1911. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhong, L.; Zhu, Y.; Liu, G. Research on the isolation of mouse Leydig cells using differential digestion with a low concentration of collagenase. J. Reprod. Dev. 2011, 57, 433–436. [Google Scholar] [CrossRef]
- Duan, P.; Ha, M.; Huang, X.; Zhang, P.; Liu, C. Intronic miR-140-5p contributes to beta-cypermethrin-mediated testosterone decline. The Science of the Total Environment 2022, 806, 150517. [Google Scholar] [CrossRef]
- Lim, Y.; Gang, D.Y.; Lee, W.Y.; Yun, S.H.; Cho, Y.B.; Huh, J.W.; Park, Y.A.; Kim, H.C. Proteomic identification of arginine-methylated proteins in colon cancer cells and comparison of messenger RNA expression between colorectal cancer and adjacent normal tissues. Ann. Coloproctol. 2022, 38, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Muthaiyan, S.M.; Chaudhuri, J.; Sellegounder, D.; Sahu, A.K.; Guha, S.; Chamoli, M.; Hodge, B.; Bose, N.; Roberts, C.; Farrera, D.O.; et al. Methylglyoxal-derived hydroimidazolone, MG-H1, increases food intake by altering tyramine signaling via the GATA transcription factor ELT-3 in Caenorhabditis elegans. Elife 2023, 12, e82446. [Google Scholar] [CrossRef] [PubMed]
- Albee, A.J.; Kwan, A.L.; Lin, H.; Granas, D.; Stormo, G.D.; Dutcher, S.K. Identification of cilia genes that affect cell-cycle progression using whole-genome transcriptome analysis in Chlamydomonas reinhardtti. G3-Genes Genomes Genet. 2013, 3, 979–991. [Google Scholar] [CrossRef]
- Xie, M.C.; Ai, C.; Jin, X.M.; Liu, S.F.; Tao, S.X.; Li, Z.D.; Wang, Z. Cloning and characterization of chicken SPATA4 gene and analysis of its specific expression. Mol. Cell. Biochem. 2007, 306, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, B.; He, S.; Zhao, Y.; Wang, Z. Cloning and characterization of zebra fish SPATA4 gene and analysis of its gonad specific expression. Biochem.-Moscow 2005, 70, 638–644. [Google Scholar] [CrossRef]
- Liu, B.; Liu, S.; He, S.; Zhao, Y.; Hu, H.; Wang, Z. Cloning and expression analysis of gonadogenesis-associated gene SPATA4 from rainbow trout (Oncorhynchus mykiss). Journal of Biochemistry and Molecular Biology 2005, 38, 206–210. [Google Scholar] [CrossRef]
- Zhang, H.T.; Yan, Z.Q.; Hu, X.B.; Yang, S.L.; Gong, Y. Interaction of C17orf25 with ADP-ribose pyrophosphatase NUDT9 detected via yeast two-hybrid method. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao (Shanghai) 2003, 35, 747–751. [Google Scholar] [PubMed]
- Strzalka, W.; Ziemienowicz, A. Proliferating cell nuclear antigen (PCNA): a key factor in DNA replication and cell cycle regulation. Ann. Bot. 2011, 107, 1127–1140. [Google Scholar] [CrossRef]
- Swaffer, M.P.; Jones, A.W.; Flynn, H.R.; Snijders, A.P.; Nurse, P. CDK Substrate Phosphorylation and Ordering the Cell Cycle. Cell 2016, 167, 1750–1761. [Google Scholar] [CrossRef]
- Price, M.A. CKI, there’s more than one: casein kinase I family members in Wnt and Hedgehog signaling. Genes. Dev. 2006, 20, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Lavi, O.; Ginsberg, D.; Louzoun, Y. Regulation of modular Cyclin and CDK feedback loops by an E2F transcription oscillator in the mammalian cell cycle. Mathematical Biosciences and Engineering : Mbe 2011, 8, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Boudreault, J.; Wang, N.; Poulet, S.; Daliah, G.; Yan, G.; Moamer, A.; Burgos, S.A.; Sabri, S.; Ali, S.; et al. Differential Regulation of Cancer Progression by CDK4/6 Plays a Central Role in DNA Replication and Repair Pathways. Cancer Research (Chicago, Ill.) 2021, 81, 1332–1346. [Google Scholar] [CrossRef] [PubMed]
- Moroy, T.; Geisen, C. Cyclin E. The International Journal of Biochemistry & Cell Biology 2004, 36, 1424–1439. [Google Scholar] [CrossRef]
- Omoboyowa, D.A.; Balogun, T.A.; Saibu, O.A.; Chukwudozie, O.S.; Alausa, A.; Olubode, S.O.; Aborode, A.T.; Batiha, G.E.; Bodun, D.S.; Musa, S.O. Structure-based discovery of selective CYP(17)A(1) inhibitors for Castration-resistant prostate cancer treatment. Biol. Methods Protoc. 2022, 7, bpab26. [Google Scholar] [CrossRef]
- Janer, G.; Leblanc, G.A.; Porte, C. A comparative study on androgen metabolism in three invertebrate species. Gen. Comp. Endocrinol. 2005, 143, 211–221. [Google Scholar] [CrossRef]
- Ohta, Y.; Kawate, N.; Inaba, T.; Morii, H.; Takahashi, K.; Tamada, H. Feeding hydroalcoholic extract powder of Lepidium meyenii (maca) enhances testicular gene expression of 3beta-hydroxysteroid dehydrogenase in rats. Andrologia 2017, 49, e12792. [Google Scholar] [CrossRef] [PubMed]
- Gunther, J.; Schuler, G.; Teppa, E.; Furbass, R. Charged Amino Acids in the Transmembrane Helix Strongly Affect the Enzyme Activity of Aromatase. International Journal of Molecular Sciences 2024, 25, 1440. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. The New England Journal of Medicine 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, J.; Yang, L.; Zhang, H.; Zhang, Y.; Gao, D.; Jiang, H.; Li, Y.; Dong, H.; Ma, T.; et al. Glyphosate exposure attenuates testosterone synthesis via NR1D1 inhibition of StAR expression in mouse Leydig cells. The Science of the Total Environment 2021, 785, 147323. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) Immunohistochemical staining of GLOD4 protein in testes of 12-month Qianbei Ma goat. (a) Immunostaining pattern of GLOD4 protein in 12M testes (100×) of Qianbei Ma goat. (b) Immunostaining pattern of GLOD4 protein in 12M testes (400×) of QianbeiMa goat; (c) Phosphate buffered saline (PBS) instead of the primary antibody was used as negative control (400×). (B) Subcellular localization of GLOD4 on goat leydig cells.GLOD4 was labeled with GFP fluorescence (a1–a3) and nuclei were labeled with DAPI dye (b1–b3) (100×). c (c1–c3) Diagram combining a (a1–a3) + b (b1–b3); (d1–d3) blank control (n = 3).
Figure 1.
(A) Immunohistochemical staining of GLOD4 protein in testes of 12-month Qianbei Ma goat. (a) Immunostaining pattern of GLOD4 protein in 12M testes (100×) of Qianbei Ma goat. (b) Immunostaining pattern of GLOD4 protein in 12M testes (400×) of QianbeiMa goat; (c) Phosphate buffered saline (PBS) instead of the primary antibody was used as negative control (400×). (B) Subcellular localization of GLOD4 on goat leydig cells.GLOD4 was labeled with GFP fluorescence (a1–a3) and nuclei were labeled with DAPI dye (b1–b3) (100×). c (c1–c3) Diagram combining a (a1–a3) + b (b1–b3); (d1–d3) blank control (n = 3).
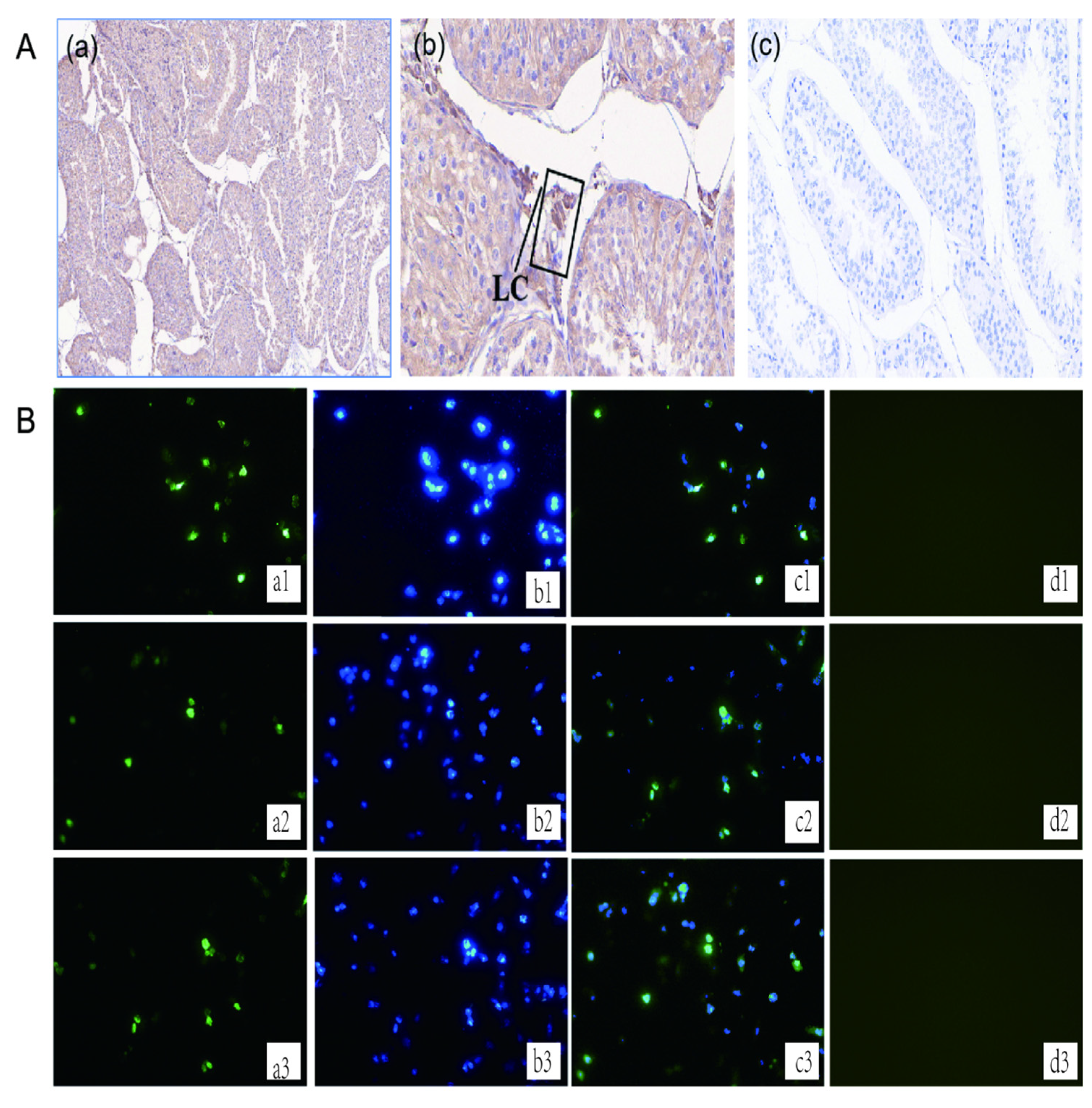
Figure 2.
(A)GLOD4 mRNA expression in testes of goats of different ages. (B)GLOD4 protein expression in testes of goats of different ages.(C,D ) GLOD4 mRNA and protein expression levels in goat LCs transfected with shRNA-GLOD4 and sh-NC.(E,F) GLOD4 mRNA and protein expression levels in goat LCs transfected with pcDNA3.1-GLOD4 and pcDNA3.1-NC. (G) mRNA expression levels of PCNA in goat LC transfected with shRNA-GLOD4、sh-NC、pcDNA3.1-NC and pcDNA3.1-GLOD4. (H, I) Cell proliferation was detected by CCK-8 after silencing and overexpressing GLOD4. Data are expressed as mean ± standard deviation, *P<0.05***P<0.01.
Figure 2.
(A)GLOD4 mRNA expression in testes of goats of different ages. (B)GLOD4 protein expression in testes of goats of different ages.(C,D ) GLOD4 mRNA and protein expression levels in goat LCs transfected with shRNA-GLOD4 and sh-NC.(E,F) GLOD4 mRNA and protein expression levels in goat LCs transfected with pcDNA3.1-GLOD4 and pcDNA3.1-NC. (G) mRNA expression levels of PCNA in goat LC transfected with shRNA-GLOD4、sh-NC、pcDNA3.1-NC and pcDNA3.1-GLOD4. (H, I) Cell proliferation was detected by CCK-8 after silencing and overexpressing GLOD4. Data are expressed as mean ± standard deviation, *P<0.05***P<0.01.
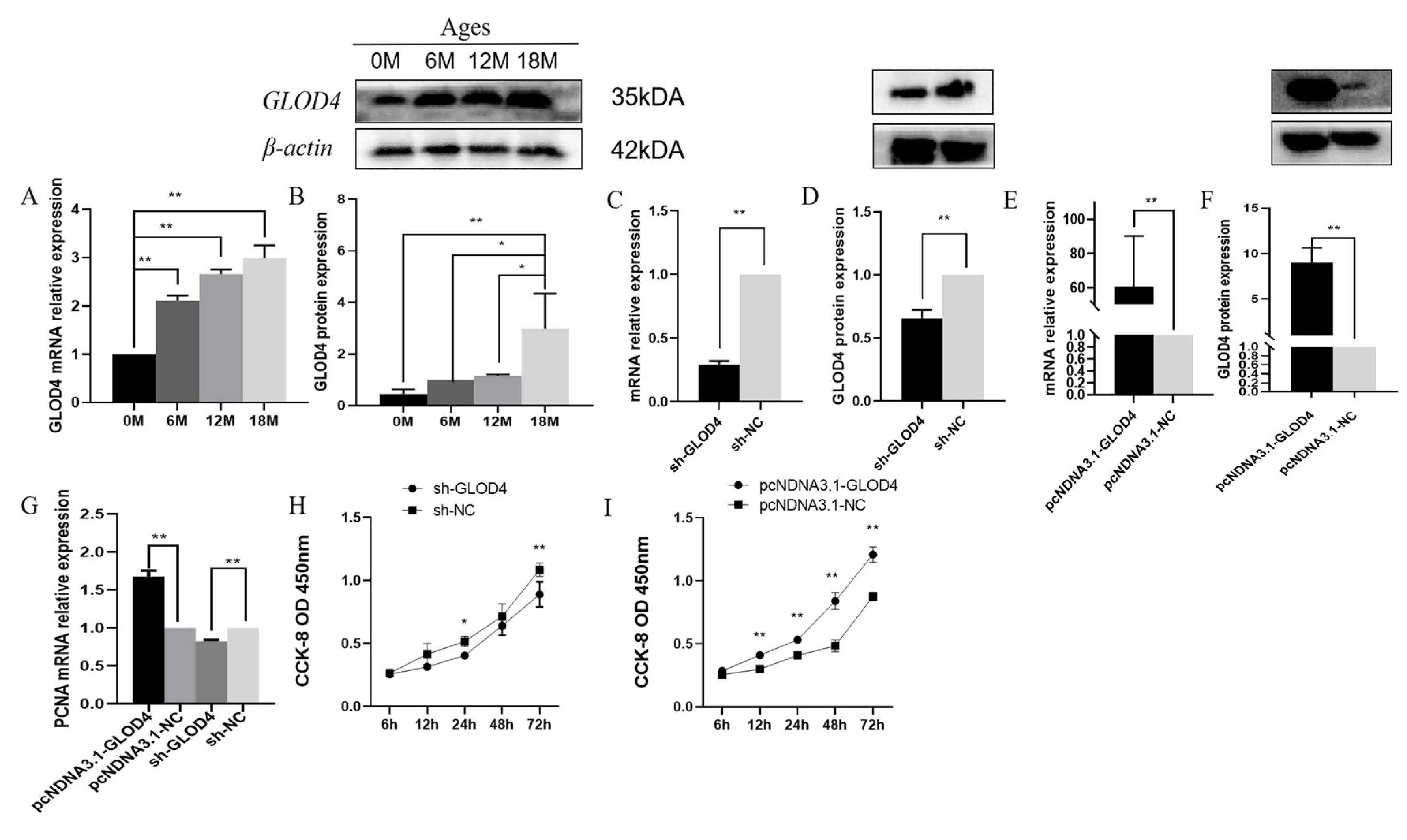
Figure 3.
(A, C) Effects of goat LCs transfected with shRNA-GLOD4 and pcDNA3.1-GLOD4 on cell cycle progression. (B, D) Expression levels of cell cycle-related genes in Qianbei Ma goat LCs after silencing and overexpressing GLOD4. All tests were performed at least three times. Data are expressed as mean ± standard deviation, *P<0.05***P<0.01.
Figure 3.
(A, C) Effects of goat LCs transfected with shRNA-GLOD4 and pcDNA3.1-GLOD4 on cell cycle progression. (B, D) Expression levels of cell cycle-related genes in Qianbei Ma goat LCs after silencing and overexpressing GLOD4. All tests were performed at least three times. Data are expressed as mean ± standard deviation, *P<0.05***P<0.01.
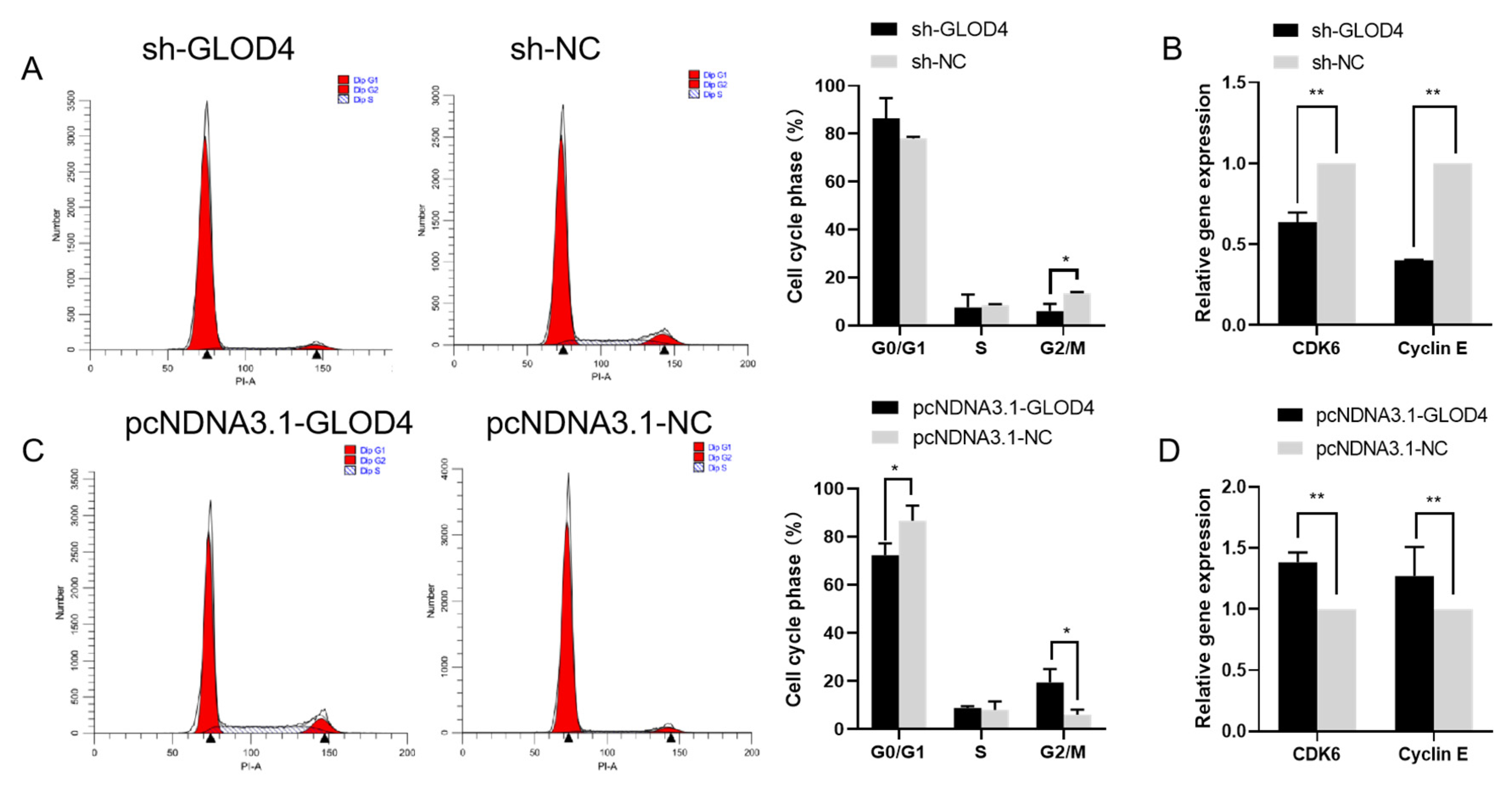
Figure 4.
(A, B) Secretion of testosterone hormone in testicular LCs of Qianbei Ma goat transfected with shRNA-GLOD4 or pcDNA3.1-GLOD4. (C, D) Expression levels of testosterone secretion-related genes in LCs of Qianbei Ma goat after silencing and overexpressing GLOD4. All tests were performed at least three times. Data are expressed as mean ± standard deviation, *P<0.05***P<0.01.
Figure 4.
(A, B) Secretion of testosterone hormone in testicular LCs of Qianbei Ma goat transfected with shRNA-GLOD4 or pcDNA3.1-GLOD4. (C, D) Expression levels of testosterone secretion-related genes in LCs of Qianbei Ma goat after silencing and overexpressing GLOD4. All tests were performed at least three times. Data are expressed as mean ± standard deviation, *P<0.05***P<0.01.


Table 1.
vector construction.
| Gene | Primer sequence (5′→3′) |
|---|---|
| Sh-GLOD4 | F:CACCGATATAAGTTCTATTTGCAGGATTCAAGAGATCCTGCAAATAGAACTTATATTTTTTTG R:GATCCAAAAAAATATAAGTTCTATTTGCAGGATCTCTTGAATCCTGCAAATAGAACTTATATC |
| Sh-NC | F:CACCGTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAATTTTTTG R:GATCCAAAAAATTCTCCGAACGTGTCACGTTCTCTTGAAACGTGACACGTTCGGAGAAC |
Table 2.
Real-time fluorescent primers information.
| Gene | Primer sequence (5′→3′) |
Gen Bank ID | Fragment Size(bP) | Tm/℃ |
|---|---|---|---|---|
| CYP17A1 | F:GCTCACCCTCGCCTETTTETT R:GTCTCCTGACACTGCTCACA |
NM_001314145.1 | 169 | 58 |
| CYP11A1 | F:CTCCAGAGGCAETAAAGAA R:TCAAAGGCAAAGTGAAACA |
NM_001287574.1 | 145 | 60 |
| 3BHSD | F:AGACCAGAAGTTCGGGAGGAA R:TCTCCCTGTAGGAGTTGGGC |
NM_001285716.1 | 292 | 60 |
| GLOD4 | F:AGCTCTGCACTTCGTGTTCA R: GCAATGCGTCCAAAACCTGT |
XM_013971906 |
554 | 60 |
| CDK6 | F:GTGGACCTCTGGAGCGTTGG R:TGCCTTGCTCATCAATGTCTGTTAC |
XM_018047426.1 | 223 | 58 |
| PCNA | F:GTAGCCGTGTCETTGCGACTCC R:GCTCTGTAGGTTCACGCCACTTG |
XM_005688167.3 | 145 | 60 |
| CylinE | F:GETGTCGGCTGCTTAGAET R:GTCTCCTGACACTGCTCACA |
XM_018062248.1 | 104 | 60 |
| β-actin | F:GETGTCGGCTGCTTAGAET R:CCAETCTCETCTCGTTTTCTG |
NM_00297986.1 | 139 | 60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



