Submitted:
05 June 2024
Posted:
06 June 2024
You are already at the latest version
Abstract
Porins are crucial proteins located in the outer membrane that directly influence antimicrobial resistance mechanisms and virulence in bacteria. In this study, a porin gene (Vp-porin) was cloned in V. parahaemolyticus, and the function of Vp-porin on biological characteristics and virulence was investigated. The results of sequence analysis showed that Vp-porin is highly conserved in Vibrio spp. and the predicted 3D structure showed it could form a transmembrane channel with 20 β barrel. Membrane permeabilization provides evidence that membrane integrity of ∆Vp-porin was damaged and the sensitivity to tetracycline, polymyxin B, rifampicin and cephalothin of ∆Vp-porin obvious increased. In addition, loss of Vp-porin damaged motility due to down-regulated flagellar synthesis. Besides, ∆Vp-porin exhibited attenuated cytotoxicity to Tetra-hymena. The relative survival rate of Tetrahymena infection with ∆Vp-porin was 86%, which is much high than that with WT (49%). Take together, Vp-porin in V. parahaemolyticus plays various roles in biological characteristics in membrane integrity, antimicrobial resistance and motility, and contributes to virulence.
Keywords:
Vibrio parahaemolyticus
; Vp-porin
; antimicrobial resistance
; motility
; virulence
1. Introduction
Vibrio parahaemolyticus is a Gram-negative bacterium commonly associated with gastroenteritis resulting from the consumption of contaminated seafood [1]. Sepsis resulting from gastroenteritis and wound infections can result in death, especially in individuals with underlying liver conditions. Antibiotics have traditionally served as the primary treatment for V. parahaemolyticus infection [2]. Studies have indicated that the widespread multi-drug resistance observed in V. parahaemolyticus strains isolated from both environmental and clinical sources is largely attributed to the inappropriate use of antibiotics in aquaculture production [3,4,5,6].
The emergence of antibiotic resistance in V. parahaemolyticus presents a considerable public health challenge, underscoring the need for a comprehensive comprehension of the molecular mechanisms underpinning bacterial drug resistance. V. parahaemolyticus demonstrates notable environmental adaptability, closely linked to its effective and precise regulatory system [7,8]. The evolution of antibiotic resistance in bacteria is a multifaceted process encompassing diverse mechanisms, including porin-mediated efflux pumps, target modification, enzymatic degradation or alteration, cell wall and membrane adjustments, and horizontal gene transfer [9,10,11]. Among these mechanisms, the interaction between porins and antimicrobial compounds plays a significant role in the development of resistance [7]. However, there is limited research on the molecular mechanism of antibiotic resistance in V. parahaemolyticus, understanding these mechanisms is essential for effective management and control of infections.
Porins are outer membrane proteins in Gram-negative bacteria that play a crucial role in modulating cellular permeability and antibiotic resistance. They are transmembrane pore-forming proteins with a β-barrel structure, forming water-filled channels for the passive transport of hydrophilic compound. Porins can be classified into non-specific or specific types based on their activity and as monomeric, dimeric, or trimeric based on their structural arrangement [9,12,13,14]. The function of porins in antibiotic resistance among Gram-negative bacteria lies in their role in mediating the entry of antibiotics across the outer membrane. Mutants lacking specific porins, such as OmpF, have been associated with antibiotic resistance in pathogens like Escherichia coli, Serratia marcescens, V. parahaemolyticus, and Enterobacter aerogenes [15,16,17,18]. Conversely, the deletion of porins like OmpU has been shown to increase susceptibility to antibiotics, as observed in V. cholerae [19]. Despite the known impact of porins on antibiotic resistance in various bacteria, the understanding of their role in V. parahaemolyticus remains limited. Specifically, the contribution of specific porins to antibiotic resistance in this bacterium is not well elucidated. Additionally, porins located on the bacterial outer membrane can have implications beyond antibiotic resistance, influencing factors like flagellum function, virulence, protein export, and adhesion, thus affecting the bacterium's adaptability to external conditions [19,20,21,22].
In this study, we aimed to identify a novel porin gene in V. parahaemolyticus and explore its role in antibiotic resistance and other characteristics of the bacterium. By constructing a Vp-porin-deletion mutant using overlapping PCR and two-step homologous recombination, we found that the deletion mutant displayed altered susceptibility to different antibiotics. Moreover, the mutants showed regulatory effects on flagellar synthesis and virulence in V. parahaemolyticus. These findings contribute to a deeper understanding of the role of porins in V. parahaemolyticus and their impact on bacterial adaptation and antibiotic response.
2. Materials and Methods
2.1. Strains, Media and Experimental Animals
The strains and plasmids utilized in this investigation are detailed in Table 1. For the construction of deletion mutants and subsequent functional analyses, we employed the Vibrio parahaemolyticus strain (ATCC® 17802™). V. parahaemolyticus and its derived mutants were cultured in Luria-Bertani (LB) medium supplemented with 3% NaCl, incubated at 37 °C with continuous shaking at 150 rpm. Tetrahymena thermophila, provided by Prof. Shan Gao from the Ocean University of China, was used for comparative studies. Tetrahymena was axenically cultured in SPP medium at a temperature of 30 °C.
2.2. Protein Domain and Structure Analysis
The amino acids of Vp-porin were achieved from the genome of ATCC (ID:17802), and validated by PCR and sequencing. The ID of Vp-porin protein is AMG08901.1. To investigate the conservation of protein sequences across different genera, we employed Clustal Omega X2.1 and ESPript 3.0 for generating a multiple sequence alignment of deduced protein sequences of Vp-porin, which were retrieved from the NCBI database for various Vibrio spp. The secondary structure of Vp-porin was predicted using the SOPMA tool (available at http://npsa.lyon.inserm.fr/cgi-bin/secpred_sopma.pl). For the construction of the 3D structure of Vp-porin, homology modeling was carried out using the Swiss Model server (http://swissmodel.expasy.org), with the protein 3D model template obtained from the Protein Data Bank (PDB) server (http://pdbbeta.rcsb.org/pdb/Welcome.do).
2.3. Construction of ∆Vp-Porin Deletion Mutant and Phenotype Characterization
The Vp-porin gene sequence were obtained from genome from ATCC17802 strain and validated by PCR and sequencing (the sequence was listed in supplemental materials). The gene ID of Vp-porin is NP_800037.1. Then the Vp-porin gene was deleted using an allelic replacement strategy. The primers designed for constructing the Vp-porin mutant strains are listed in Table 2. PCR amplification was performed to obtain upstream and downstream flanking fragments of the target gene, generating 772 bp upstream and 917 bp downstream overlap fragments through overlap PCR. These overlapped DNA products from Vp-porin were digested with Sac I and Spe I enzymes and subsequently inserted into the pSR47s plasmid at the Sac I and Spe I restriction sites. The resulting recombinant plasmid, pSR47S-∆Vp-porin, was transformed into CC118 λpir strain and validated through gene sequencing. Following validation, the pSR47S-∆Vp-porin recombinant plasmid was introduced into the wild-type (WT) strain via conjugation and selected on LB agar plates containing kanamycin (Kan) and ampicillin (Amp). The second cross-over recombination event was then selected for on LB agar containing 10% sucrose to isolate the ∆Vp-porin mutant through sucrose resistance screening. The mutation in the ∆Vp-porin strain was confirmed by PCR using primers T1/T2 and subsequent sequencing. To assess the growth kinetics of the different strains, overnight bacterial cultures were diluted at a 1:100 ratio into 15 ml of fresh LB medium supplemented with 3% NaCl. The cultures were incubated at 37 °C with shaking at 150 rpm until they reached an OD600 value of 1.0. Subsequently, these cultures were further diluted 1:100 into 100 ml of the same medium and grown in a temperature-controlled incubator. Samples were collected every hour to monitor growth.
2.4. Proteolysis Activity Assay
The assessment of protease production and enzymatic activity was performed using LB agar plates supplemented with 2% skim milk. Overnight cultures of both the wild-type (WT) and the ∆Vp-porin mutant strains were adjusted to an optical density at 600 nm (OD600) of 1.0. Following this adjustment, 5 μL aliquots of these diluted cultures were spotted onto separate agar plates and incubated at 37ºC for a duration of 24 hours. The presence of proteolytic activity was determined by observing clear zones around the bacterial colonies. The diameters of these transparent zones were quantitatively measured using ImageJ software.
2.5. Outer Membrane Permeabilization Assay
The outer membrane permeability of bacteria was assessed using the N-phenyl-1-napthylamine (NPN) uptake assay, following established procedures [23]. Initially, cells cultured overnight were washed and suspended in buffer (5 mM HEPES, 5 mM glucose, pH 7.4). Subsequently, NPN was introduced to 2 ml of cells in a quartz cuvette to achieve a final concentration of 10 mM, and the background fluorescence was recorded (excitation wavelength = 350 nm, emission wavelength = 420 nm). Changes in fluorescence were monitored using BioTek Synergy HTX (Agilent, Uk).
2.6. Antimicrobial Susceptibility Testing and Survival Assay
The antimicrobial resistance of WT and mutant to (chloramphenicol, tetracycline, streptomycin, kanamycin, polymyxin B, rifampicin, cephalothin, levofloxacin, ampicillin and amoxicillin were determined by Antimicrobial Susceptibility Testing (AST). The experiment was carried out using Kirby–Bauer Disk Diffusion Susceptibility Test method [24]. Two strains were cultured overnight and adjusted OD600 nm to 1. Then the bacteria strains were spread on the LB agar (Merck, Germany) using sterile cotton swab. Sterile antimicrobial susceptibility disks were then placed on the agar plate and incubated at 37 °C for 24 h. The zone of inhibition was observed after 24 h of incubation. All assays were performed in triplicate. For survival assays, overnight cultures were adjusted to 0.5 (OD600nm) in LB and diluted in 1:100. Aliquots of 1mL of culture were added to different antibiotics. Titrations of tetracycline were performed using concentrations ranging from 0.15-1.25 μg/mL and 0.3-2.5 μg/mL for rifamycin. Cultures were incubated overnight at 37 °C in a rotary shaker after which the colony forming units were calculated by plating serial dilutions on LB agar. The percentage of bacterial survival was calculated by comparing the CFU/mL of the treated versus the untreated.
2.7. Motility Assay
Overnight cultures of the wild-type (WT) and the ∆Vp-porin mutant strain were first adjusted to an optical density at 600 nm (OD600) of 1.0. Subsequently, 2 μL aliquots of these diluted cultures were spotted onto separate plates for motility assays. For assessing swimming motility, LB medium containing 0.3% agar was used, while LB medium with 1.5% agar was employed for evaluating swarming motility. The plates were then incubated at 37°C for 24 hours. Following the incubation period, the swarming capabilities of the strains were observed and recorded. To ensure the accuracy and reproducibility of the results, each experiment was conducted six times.
2.8. qRT-PCR Analysis
Total RNA extraction was performed utilizing the RNeasy Plus Mini Kit (Qiagen), and genomic DNA contamination was eliminated using RNase-free DNase I. For cDNA synthesis, equal quantities of RNA (1 μg) from each sample were reverse transcribed using the PrimeScript RT Reagent Kit with gDNA Eraser (Takara, Tokyo, Japan). The specific primers employed for the qRT-PCR are detailed in Table 3. The qRT-PCR reactions were executed using the SLAN 96S Real-Time PCR System (Xiamen Zeesan Biotech, China) and FastStart Universal SYBR Green Master (Yeasen, Shanghai, China). To quantify the relative expression levels of the target genes, the transcript levels were normalized to those of the 16S rRNA gene, applying the 2^−ΔΔCt method for calculation. To ensure the robustness and reproducibility of the results, three independent biological replicates were performed for each experiment, with each individual replicate run in triplicate.
2.9. Assessment of Strains Virulence using Tetrahymena thermophila
The virulence of ∆Vp-porin mutant was assessed via Tetrahymena infection model by the relative survival of bacteria and Tetrahymena after co-culturing as previously described [25]. Briefly, Tetrahymena was cultured in sterile SPP medium at 30 °C for 48 h using an initial inoculum of 10^3 cells/mL. Cells in this culture were harvested by centrifugation at 2,000 g for 10 min at 10°C, washed twice with sterile SPP medium and adjusted to 1x10^5 cells/mL. Each strain after being overnight cultured was harvested, washed twice in SPP medium and adjusted to 3x10^9 CFU/mL. 5,000:1 co-cultures of V. parahaemolyticus (3x10^9 CFU/mL) and Tetrahymena (1x10^5 cells/mL) was mixed and cultured 6 h at 30 °C. Bacterial growth in these mixed cell suspensions was determined every 1 h by measuring absorbance at 450 nm every 1 h. Controls contained the bacterial strains mixed with an equal volume of SPP medium. Sterile SPP medium was used as the blank well. The relative survival of bacteria (%) was counted as the number of bacteria remaining in culture relative to the number of bacterial cells grown alone. The relative survival of Tetrahymena (%) was calculated as the number of Tetrahymena cells remaining in culture relative to the number of cells cultured alone. The cellular morphology of Tetrahymena cells incubated for 6 h was examined under a light microscope (Nikon 80i) after they had been fixed with 4% PFA. Each experiment was repeated at least three times.
2.10. Statistical Analysis
Statistical analysis was performed by GraphPad Prism 9 (Graph Pad Software, Inc, San Diego, CA, USA). Experimental data were expressed as the mean ± SD. Two-group comparison was analyzed using Student’s t-test. ns, p > 0.05; *p < 0.05; **p < 0.01; ***p < 0.001.
3. Results
3.1. Vp-Porin Sequence and Structural Analysis
The Vp-porin gene from V. parahaemolyticus encompasses an open reading frame (ORF) spanning 1026 bp, encoding a protein of 342 amino acids with an estimated molecular weight of 37 kDa. A comprehensive multiple sequence alignment of the amino acid sequences revealed significant homology between Vp-porin and porins from other Vibrio species, including V. antiquarius (82.95%), V. harveyi (86.30%), V. owensii (88.89%), V. jasicida (87.83%), V. vulnificus (64.53%), V. campbellii (85.47%), and V. alginolyticus (82.95%) (Figure 1A). The secondary structure prediction for Vp-porin indicates a composition of 18.13% α-helix, 26.90% extended strand, and 54.97% random coil (Figure 1B). The secondary structural elements of Vp-porin, aligned above the sequence comparison in Figure 1A, suggest a relatively simple fold comprising 20 antiparallel β-strands forming a hollow β-barrel. The three-dimensional (3D) structure of Vp-porin (Figure 1C), reveals a channel formed by its transmembrane segment. The depiction of the monomer emphasizes Vp-porin's role as a transmembrane channel, underscoring its significance in facilitating the transport of molecules across the bacterial membrane. In a word, Vp-porin is a highly conserved protein that is speculated to function as an outer membrane porin, playing an important role in the regulation of cellular permeability and potentially impacting antibiotic resistance mechanisms.
3.2. Construction and Characterization of the Deletion Mutant of Vp-Porin Gene in V. parahaemolyticus
To investigate the significance of the Vp-porin gene for V. parahaemolyticus, a deletion mutant of Vp-porin was generated using an allelic replacement strategy. The primers utilized for the construction of the Vp-porin mutant are detailed in Table 2. Following the successful deletion of Vp-porin, the nucleotide fragment size of Vp-porin was reduced from its original 1029 bp to 78 bp (Figure 2A). PCR verification confirmed the successful creation of the mutant strain ∆Vp-porin (Figure 2B). Initially, we evaluated the growth of the strain in LB media supplemented with 0.3% NaCl. No discernible differences in colony morphology were observed on agar media between the WT and ∆Vp-porin strains (Figure 2C). Nonetheless, the growth curve analysis revealed that the mutant exhibited a marginally slower growth rate compared to the WT strain, with OD600 values of 0.931 and 0.874 for the WT and ∆Vp-porin strains, respectively, after 12 hours of cultivation at 37°C (Figure 2D). Further analysis focused on the protease activity of ∆Vp-porin in the presence of 2% skim milk. The ∆Vp-porin strain displayed a smaller clear zone compared to the WT (Figure 2E), with the relative diameter of the transparent zone for the WT being 1.64 mm, surpassing that of ∆Vp-porin at 1.46 mm. This indicates a proteolytic deficiency in the ∆Vp-porin strain on skim milk plates (Figure 2F). These findings underscore the pivotal role of Vp-porin in facilitating essential physiological functions within V. parahaemolyticus, particularly with regard to protease activity.
3.3. Permeabilization of Outer Membranes
The outer membrane permeabilization of WT and mutant was determined by using the NPN uptake assay. NPN is a nonpolar hydrophobic fluorescent probe, which is typically blocked by the outer membrane. However, when the integrity of the cell membrane is disrupted, it can enter the outer membrane and exhibit higher fluorescence intensity. As is shown in Figure 3, an increased outer membrane permeability was observed in ∆Vp-porin.
3.4. Comparison of Antimicrobial Susceptibility between WT and ∆Vp-Porin Strain
To study whether Vp-porin we identified play a role in antimicrobial resistance, we performed antibacterial activity measurement. The sensitivity of the WT and ∆Vp-porin to ten antimicrobials (chloramphenicol, tetracycline, streptomycin, kanamycin, polymyxin B, rifampicin, cephalothin, levofloxacin, ampicillin and amoxicillin) were tested. As shown in Figure 4A, ∆Vp-porin was obvious increased sensitivity to tetracycline, polymyxin B, rifampicin and cephalothin, and ∆Vp-porin was slightly increased sensitivity to chloramphenicol. Whereas ∆Vp-porin displays similar sensitivity as the WT to other antibiotics. In detail, for polymyxin B, ∆Vp-porin show a drastic increase in inhibition zone diameter (12.81 mm) compared to WT 8.19 mm (Figure 4B). ∆Vp-porin also display an increasing sensitivity to tetracycline (the inhibition zone diameter: 18.70 mm), comparable to WT (the inhibition zone diameter: 15.74 mm). In the presence of rifampicin, the inhibition zone diameter of WT (17.63 mm) is smaller than ∆Vp-porin (23.0 mm). There was barely visible zone of inhibition both in WT and ∆Vp-porin mutant, indicating both of them exhibited resistance to ampicillin and amoxicillin. Survival curve of WT and ∆Vp-porin in the presence of varying concentrations of rifampicin and tetracycline was tested (Figure4C). After exposure to 1.25 μg /mL rifampicin, WT has a 55 % survival rate whereas the ∆Vp-porin mutant exhibits a survival rate of 17 %. ∆Vp-porin mutant display lower survival rate than the WT when exposing to rifampicin. After exposure to 0.625 μg/mL tetracycline, WT has a 35 % survival rate whereas the ∆Vp-porin mutant exhibits a low survival rate of 7 %. Results indicates ∆Vp-porin mutant displays increased susceptibility to rifampicin and tetracycline. Overall, our results uncover Vp-porin contributes to antimicrobial resistance.
3.5. The Vp-Porin Mutant Exhibits Lower Motility and Decreases Transcription of Polar Flagellar Genes and Lateral Flagellar Genes in V. parahaemolyticus
To evaluate the swimming and swarming capabilities, we compared the wild-type and ∆Vp-porin strains. The wild-type strain displayed initial inoculum spreading throughout the plate in an asymmetrical pattern, characteristic of swimming (Figure 4A). Conversely, the ∆Vp-porin strain exhibited minimal movement beyond the initial inoculum site, indicating a defect in swimming in this medium. Similarly, when colonies of the wild-type and ∆Vp-porin strains were inoculated onto 1.5% agar plates and incubated, the wild-type strain demonstrated colony growth expanding in a symmetrical pattern, indicating swarming behavior. In contrast, the colonies of the ∆Vp-porin strain exhibited limited movement on the plate, suggesting a slight defect in swarming (Figure 4A). Statistical analysis of the zone of swimming and swarming further confirmed that the ability of ∆Vp-porin to swim and swarm was significantly lower than that of the WT (Figure 4B, C). V. parahaemolyticus harbors both polar flagella and lateral flagella. To investigate the impact of Vp-porin on flagellar gene expression, qRT-PCR analysis was conducted. Notably, in the ∆Vp-porin strain, all polar flagellar cluster I genes (flgB, flgM, flgK, flgC) exhibited significant down-regulation compared with the WT strain, while polar flagellar cluster II genes fliE and fliK showed no significant difference between WT and ∆Vp-porin (Figure 4D). Furthermore, qRT-PCR was employed to assess the regulation of Vp-porin on lateral flagellar gene clusters. The results demonstrated significant down-regulation of flgA, flgB, flgG, lafA, motY, and fliE genes in the ∆Vp-porin strain compared to the WT (Figure 4E). Collectively, these findings suggest that Vp-porin acts as a sigma factor, regulating the expression of polar flagellar genes to modulate swimming motility in V. parahaemolyticus. Additionally, qRT-PCR analysis was also used to examine genes from the T6SS2, which plays a crucial role in virulence and adhesion. The mRNA expression level of all genes (VPA1043, VPA1044, VPA1045) was markedly down-regulated in the ∆Vp-porin strain compared to the WT (Figure 4F). These results consolidate the role of Vp-porin in regulating the expression of T6SS2 genes. Thus, the deficiency of ∆Vp-porin in motility is a result of the downregulated expression level of polar and lateral flagellar synthesis gene in V. parahaemolyticus.
3.6. Assessment of Virulence of ∆Vp-Porin using Tetrahymena
To investigate the virulence of the ∆Vp-porin strain, a co-culture experiment was conducted using Tetrahymena thermophila. In the control group without bacteria, Tetrahymena cells exhibited an elliptical or pear-shaped morphology, with a large number of cells displaying rapid activity (Figure 3A). However, when co-cultured with bacteria, the morphology of Tetrahymena cells underwent changes. The cells became shrunken and round, displaying more vacuoles in the cytoplasm compared to the control group. Some cells even showed signs of cell death. Notably, co-culturing Tetrahymena with the wild-type (WT) strain resulted in severe cell shrinkage, deformation, and evident inhibition of cell growth. In contrast, when co-cultured with the ∆Vp-porin strain, Tetrahymena exhibited relatively improved growth, with decreased abnormal cell morphology and increased cell numbers observed compared to the WT group (Figure 3A). These observations suggest that the ∆Vp-porin strain possesses reduced virulence compared to the WT strain, as evidenced by the ameliorated effects on Tetrahymena growth and morphology. During infection, both the WT and the ∆Vp-porin mutant cultivated alone exhibited slow growth in sterile SPP medium. However, the biomass of both the WT and the ∆Vp-porin mutant decreased continuously when co-cultured with Tetrahymena, suggesting that a large number of bacteria were preyed upon by Tetrahymena (Figure 5B, C). Notably, the biomass of the ∆Vp-porin strain declined more rapidly than that of the WT strain when co-cultivated with the same number of Tetrahymena (Figure 5C), indicating that the ∆Vp-porin strain was less resistant to predation by Tetrahymena compared to the WT strain. As depicted in Figure 5D, the relative survival of the ∆Vp-porin strain significantly decreased in the co-culture model, highlighting its reduced resistance to predation by Tetrahymena in comparison to the WT strain. The growth dynamics of Tetrahymena were also investigated in the co-culture model (Figure 5E). When cultured alone, the biomass of Tetrahymena increased during incubation, reaching a maximum concentration of 3.96 x 10^5 cells/mL at 6 hours. However, the growth of Tetrahymena was inhibited when co-cultivated with both the WT and ∆Vp-porin strains (Figure 5E). The number of Tetrahymena cells grown in the presence of WT initially increased before declining, with a relative survival rate of 49% at 6 hours. In contrast, the number of Tetrahymena cells co-cultured with the ∆Vp-porin strain continued to increase throughout the incubation period. Co-culture with the ∆Vp-porin strain had a lesser impact on Tetrahymena viability, as evidenced by a relative survival rate of 86% (Figure 3F). In total, these data indicated the virulence of ∆Vp-porin was greatly attenuated.
4. Discussion
In this study, we aimed to explore the influence of porin on Vibrio parahaemolyticus by generating the ∆Vp-porin mutant. Our results uncovered that the absence of Vp-porin led to increased susceptibility to partially antibiotics and hindered both swimming and swarming capabilities. Additionally, utilizing a Tetrahymena infection model, we demonstrated that the deletion of Vp-porin significantly diminishes the virulence of V. parahaemolyticus.
The Vp-porin gene in V. parahaemolyticus displays significant sequence homology to porins found in various Vibrio species, suggesting a conserved structural and functional role for Vp-porin across these organisms. Its predicted secondary structure and 3D configuration, characterized by 20 antiparallel strands forming a hollow β barrel with distinct periplasmic and extracellular features, indicate potential channel-forming characteristics. The increasing prevalence of antimicrobial resistance (AMR) among bacterial pathogens, especially Gram-negative bacteria, emphasizes the necessity for a comprehensive understanding of the mechanisms contributing to this phenomenon [18,21,26]. Porins represent major proteins present in the outer membrane and play a direct role in antimicrobial resistance mechanisms [9]. It has been confirmed distinct role of outer membrane porins in antibiotic resistance and membrane integrity in Escherichia coli. Choi et al. put forward that porins can be classified into three groups according to their roles in antibiotic transport and membrane integrity: antibiotic transport-related specific porin, membrane integrity-related non-specific porin, and non-specific porins involved in both antibiotic transport and membrane integrity [14]. In this study, the sensitivity of the ∆Vp-porin strain to tetracycline, polymyxin B, rifampicin and cephalothin was significantly increased, revealing that Vp-porin is involve in regulating antimicrobial resistance in V. parahaemolyticus, which is contrary to mutants of OmpF porins that the increased resistance impact to several antibiotics [14]. Antimicrobial can penetrate the outer membrane by two different pathways, through the lipid bilayer or through porins.
Research showed mutant of OmpU porin and OmpA contributes to increased susceptibility to some antibiotics due to impaired membrane integrity, which is similar to Vp-porin in V. parahaemolyticus [14,19]. In membrane integrity-related non-specific porin, the impaired membrane integrity can increase the intracellular diffusion of antibiotics [23]. An increase in the NPN fluorescence of ∆Vp-porin indicated the membrane integrity maybe disrupted. Thus Vp-porin is likey to be a membrane integrity-related non-specific porin. Interestingly, no difference in sensitivity to kanamycin, ampicillin, and amoxicillin was observed between the WT and ∆Vp-porin strains, indicating a specific role of Vp-porin in modulating resistance to certain classes of antibiotics. Generally, chemicals with a molecular weight of more than 600 Da could not penetrate the envelope of the Gram-negative bacteria [14]. But the molecular weight of kanamycin, ampicillin, and amoxicillin are 484, 349 and 365 Da respectively, which is much less than 600 Da. We speculate there are other resistance mechanism existed in V. parahaemolyticus, such as the presence pf drug resistance genes.
Furthermore, we also found that the ∆Vp-porin mutant exhibited defective motility in swimming and swarming, suggesting a potential role of Vp-porin in flagellar synthesis and motility regulation. This phenomenon was also reported in ompX and ompA that is critical for flagellar assembly and swimming ability in Stenotrophomonas maltophilia and E. coli [19,21,22]. V. parahaemolyticus possesses dual flagellar system, polar flagellum for swimming in liquids and the peritrichous lateral flagella for swarming over surfaces or in viscous liquids [27]. The significant down-regulation of polar flagellar genes in cluster I and lateral flagellar genes in the ∆Vp-porin strain underscores Vp-porin is a sigma factor that could regulate the expression of polar flagellar genes to mediate swimming motility and the expression of lateral flagellar genes to mediate swarming motility in V. parahaemolyticus. Interestingly, no significant differences in the mRNA expression levels of fliE and fliK from cluster II suggests a more nuanced regulatory mechanism involving Vp-porin in the control of specific subsets of polar flagellar genes. The T6SS2 as one major virulence determinant in V. parahaemolyticus plays a role in bacterial invasion into host cells [28,29,30]. Our qRT-PCR analysis exhibited that the expression of T6SS2 genes were down-regulated in the ∆Vp-porin strain compared to the WT strain, suggesting Vp-porin could positively regulate the expression of T6SS2. Above all, our results further suggest a multifaceted role of Vp-porin in modulating gene expression profiles crucial for flagellar assembly, motility, and virulence in V. parahaemolyticus.
Tetrahymena, a single-celled ciliate, has emerged as a valuable model organism for studying host-pathogen interactions. Recent research has utilized Tetrahymena to investigate bacterial infection mechanisms and host defense responses, shedding light on fundamental aspects of microbial pathogenesis [31]. In this study, we aimed to evaluate the virulence of ∆Vp-porin using a Tetrahymena infection model. Similar to Tetrahymena-Aeromonas co-culture models [25], significant differences were observed in the relative survival of Tetrahymena co-cultured with the WT strain compared to ∆Vp-porin. Compared with the WT group, which exhibited shrinkage, deformation, and a lower cell count, Tetrahymena co-cultured with ∆Vp-porin showed slightly improved growth and decreased abnormal cell morphology. This phenomenon is reminiscent of experiments involving Listeria monocytogenes, where hemolytic L. monocytogenes induces lysis of Tetrahymena pyriformis, while only a few protozoa undergo lysis in the presence of nonhemolytic Listeria innocua [32]. Previous molecular studies and animal infection experiments have indicated the critical role of certain porins, such as OmpX and OmpA from E. coli, and Acinetobacter baumannii, in bacterial pathogenicity by influencing bacterial adhesion and virulence factors [21,22,33]. Furthermore, it has been observed that OmpU from toxigenic strains evolves in the environment and serves as preadaptations to virulence in the context of the human host [34]. Similarly, we found the ∆Vp-porin strain exhibits attenuated virulence compared to the WT strain, indicating Vp-porin plays important role in virulence of V. parahaemolyticus. V. parahaemolyticus has many virulence factors, T6SS2 and adhesion factors are crucial factors by controlling the virulence. T6SS2 gene transcription but also a mediator of V. parahaemolyticus adhesion to host cells [29]. Besides, the flagellum also affects bacterial virulence by promoting early biofilm formation and promoting adherence and invasion in V. parahaemolyticus [2,35]. Our results confirm that Vp-porin can down-regulate the expression level of T6SS2 genes and flagellar synthesis genes. Thus, the virulence of ∆Vp-porin strain is high likely associated to the down-regulated of T6SS2 and blocked flagellar synthesis.
5. Conclusions
In conclusion, we constructed the Vp-porin deletion mutant and preliminary investigated the effects of Vp-porin gene on resistance antimicrobials and virulence-associated properties in V. parahaemolyticus. The present results suggest that Vp-porin modulates antimicrobial resistance and positively regulate flagellar synthesis in V. parahaemolyticus. Further analysis using a Vibrio-Tetrahymena co-culture model demonstrated that Vp-porin could contribute to the virulence of V. parahaemolyticus. These findings not only are helpful for better understanding the function of Vp-porin, but also provide further potential evidence supporting the feasibility of engineering strategies aimed at mitigating antimicrobial resistance of V. parahaemolyticus.
Author Contributions
Data curation, Qitong Fang; Formal analysis, Binghong Liu; Funding acquisition, Xiu Fang and Baolong Bao; Investigation, Jinyuan Che, Qitong Fang and Shaojie Hu; Methodology, Jinyuan Che and Qitong Fang; Software, Jinyuan Che and Shaojie Hu; Validation, Jinyuan Che and Binghong Liu; Visualization, Lekang Li; Writing – original draft, Jinyuan Che; Writing – review & editing, Lei Wang, Lekang Li, Tuyan Luo and Baolong Bao.
Funding
This work was supported by grants from the National Key R&D Program of China (2022YFD2400103), the Regional Development Project of Fujian (2021N3016), and Fisheries High Quality Development Project of Fujian (FJHYF-L-2023-16).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ghenem, L.; Elhadi, N.; Alzahrani, F.; Nishibuchi, M. Vibrio Parahaemolyticus: A Review on Distribution, Pathogenesis, Virulence Determinants and Epidemiology. Saudi J Med Med Sci 2017, 5, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Meng, H.; Gu, D.; Li, Y.; Jia, M. Molecular mechanisms of Vibrio parahaemolyticus pathogenesis. Microbiol Res 2019, 222, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Cheng, J.; Wu, Q.; Zhang, J.; Xie, T. Prevalence, characterization, and antibiotic susceptibility of Vibrio parahaemolyticus isolated from retail aquatic products in North China. BMC Microbiol 2016, 16, 32. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Sun, J.; Qiu, Y.; Zhang, M.; Xue, X.; Li, X.; Yang, W.; Zhou, D.; Hu, L.; Zhang, Y. The quorum sensing regulator OpaR is a repressor of polar flagellum genes in Vibrio parahaemolyticus. J Microbiol 2021, 59, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Haifa-Haryani, W.O.; Amatul-Samahah, M.A.; Azzam-Sayuti, M.; Chin, Y.K.; Zamri-Saad, M.; Natrah, I.; Amal, M.N.A.; Satyantini, W.H.; Ina-Salwany, M.Y. Prevalence, Antibiotics Resistance and Plasmid Profiling of Vibrio spp. Isolated from Cultured Shrimp in Peninsular Malaysia. Microorganisms 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Wang, Y.; Xu, H.; Jin, X.; Yan, B.; Zhang, W. Antibiotic Resistance and Epidemiology of Vibrio parahaemolyticus from Clinical Samples in Nantong, China, 2018-2021. Infect Drug Resist 2023, 16, 7413–7425. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.; Hancock, R.E. Adaptive and mutational resistance: role of porins and efflux pumps in drug resistance. Clin Microbiol Rev 2012, 25, 661–681. [Google Scholar] [CrossRef] [PubMed]
- Pazhani, G.P.; Chowdhury, G.; Ramamurthy, T. Adaptations of Vibrio parahaemolyticus to Stress During Environmental Survival, Host Colonization, and Infection. Front Microbiol 2021, 12, 737299. [Google Scholar] [CrossRef] [PubMed]
- Pages, J.M.; James, C.E.; Winterhalter, M. The porin and the permeating antibiotic: a selective diffusion barrier in Gram-negative bacteria. Nat Rev Microbiol 2008, 6, 893–903. [Google Scholar] [CrossRef]
- MacLean, R.C.; San Millan, A. The evolution of antibiotic resistance. Science 2019, 365, 1082–1083. [Google Scholar] [CrossRef]
- Zhou, G.; Wang, Q.; Wang, Y.; Wen, X.; Peng, H.; Peng, R.; Shi, Q.; Xie, X.; Li, L. Outer Membrane Porins Contribute to Antimicrobial Resistance in Gram-Negative Bacteria. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and function of bacterial outer membrane proteins: barrels in a nutshell. Mol Microbiol 2000, 37, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Schulz, G.E. The structure of bacterial outer membrane proteins. Biochim Biophys Acta 2002, 1565, 308–317. [Google Scholar] [CrossRef]
- Choi, U.; Lee, C.R. Distinct Roles of Outer Membrane Porins in Antibiotic Resistance and Membrane Integrity in Escherichia coli. Front Microbiol 2019, 10, 953. [Google Scholar] [CrossRef]
- Bornet, C.; Davin-Regli, A.; Bosi, C.; Pages, J.M.; Bollet, C. Imipenem resistance of enterobacter aerogenes mediated by outer membrane permeability. J Clin Microbiol 2000, 38, 1048–1052. [Google Scholar] [CrossRef]
- Ziervogel, B.K.; Roux, B. The binding of antibiotics in OmpF porin. Structure 2013, 21, 76–87. [Google Scholar] [CrossRef]
- Moya-Torres, A.; Mulvey, M.R.; Kumar, A.; Oresnik, I.J.; Brassinga, A.K.C. The lack of OmpF, but not OmpC, contributes to increased antibiotic resistance in Serratia marcescens. Microbiology (Reading) 2014, 160, 1882–1892. [Google Scholar] [CrossRef]
- Meng, X.; Huang, D.; Zhou, Q.; Ji, F.; Tan, X.; Wang, J.; Wang, X. The Influence of Outer Membrane Protein on Ampicillin Resistance of Vibrio parahaemolyticus. Can J Infect Dis Med Microbiol 2023, 2023, 8079091. [Google Scholar] [CrossRef]
- Grant, T.A.; Lopez-Perez, M.; Haro-Moreno, J.M.; Almagro-Moreno, S. Allelic diversity uncovers protein domains contributing to the emergence of antimicrobial resistance. PLoS Genet 2023, 19, e1010490. [Google Scholar] [CrossRef]
- Confer, A.W.; Ayalew, S. The OmpA family of proteins: roles in bacterial pathogenesis and immunity. Vet Microbiol 2013, 163, 207–222. [Google Scholar] [CrossRef]
- Hirakawa, H.; Suzue, K.; Takita, A.; Kamitani, W.; Tomita, H. Roles of OmpX, an Outer Membrane Protein, on Virulence and Flagellar Expression in Uropathogenic Escherichia coli. Infect Immun 2021, 89. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.H.; Chang, C.L.; Huang, H.H.; Lin, Y.T.; Li, L.H.; Yang, T.C. Interplay between OmpA and RpoN Regulates Flagellar Synthesis in Stenotrophomonas maltophilia. Microorganisms 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Wang, J.; Gao, H.; Wang, Z.; Dong, N.; Ma, Q.; Shan, A. Antimicrobial properties and membrane-active mechanism of a potential alpha-helical antimicrobial derived from cathelicidin PMAP-36. PLoS One 2014, 9, e86364. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Tech Bull Regist Med Technol 1966, 36, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.; Xie, X.; Dong, Y.; Du, H.; Wang, N.; Lu, C.; Liu, Y. Identification of novel virulence-related genes in Aeromonas hydrophila by screening transposon mutants in a Tetrahymena infection model. Vet Microbiol 2017, 199, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Siu, L.K.; Fung, C.P.; Lin, J.C.; Yeh, K.M.; Chen, T.L.; Tsai, Y.K.; Chang, F.Y. Contribution of outer membrane protein K36 to antimicrobial resistance and virulence in Klebsiella pneumoniae. J Antimicrob Chemother 2010, 65, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B.J.; McCarter, L.L. Lateral flagellar gene system of Vibrio parahaemolyticus. J Bacteriol 2003, 185, 4508–4518. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Osei-Adjei, G.; Zhang, Y.; Gao, H.; Yang, W.; Zhou, D.; Huang, X.; Yang, H.; Zhang, Y. CalR is required for the expression of T6SS2 and the adhesion of Vibrio parahaemolyticus to HeLa cells. Arch Microbiol 2017, 199, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Garcia, F.; Ruiz-Perez, F.; Cataldi, A.; Larzabal, M. Type VI Secretion System in Pathogenic Escherichia coli: Structure, Role in Virulence, and Acquisition. Front Microbiol 2019, 10, 1965. [Google Scholar] [CrossRef]
- Gu, D.; Zhang, Y.; Wang, K.; Li, M.; Jiao, X. Characterization of the RpoN regulon reveals the regulation of motility, T6SS2 and metabolism in Vibrio parahaemolyticus. Front Microbiol 2022, 13, 1025960. [Google Scholar] [CrossRef]
- Vadivelu, J.; Pang, M.-D.; Lin, X.-Q.; Hu, M.; Li, J.; Lu, C.-P.; Liu, Y.-J. Tetrahymena: An Alternative Model Host for Evaluating Virulence of Aeromonas Strains. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Ly, T.M.; Muller, H.E. Ingested Listeria monocytogenes survive and multiply in protozoa. J Med Microbiol 1990, 33, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, B.L.; Leal, B.F.; Leyser, M.; de Barros, M.P.; Trentin, D.S.; Ferreira, C.A.S.; de Oliveira, S.D. Increased ompW and ompA expression and higher virulence of Acinetobacter baumannii persister cells. BMC Microbiol 2023, 23, 157. [Google Scholar] [CrossRef] [PubMed]
- Ganie, H.A.; Choudhary, A.; Baranwal, S. Structure, regulation, and host interaction of outer membrane protein U (OmpU) of Vibrio species. Microb Pathog 2022, 162, 105267. [Google Scholar] [CrossRef]
- Haiko, J.; Westerlund-Wikstrom, B. The role of the bacterial flagellum in adhesion and virulence. Biology (Basel) 2013, 2, 1242–1267. [Google Scholar] [CrossRef]
Figure 1.
Homology modelling of the V. parahaemolyticus Vp-porin structure. (A) Multiple sequence alignment of Vp-porin in different vibrio species. Amino acid conservation is shown in red and secondary structure was based on the V. parahaemolyticus Vp-porin structure. The η symbol refers to a 310-helix. Helices and strands are shown as black helices with squiggles and arrows, respectively. Red box, white character indicates strict identity; (B) Secondary structure of vp-porin using SOPMA; (C) 3D Structure of Vp-porin using Swiss-model.
Figure 1.
Homology modelling of the V. parahaemolyticus Vp-porin structure. (A) Multiple sequence alignment of Vp-porin in different vibrio species. Amino acid conservation is shown in red and secondary structure was based on the V. parahaemolyticus Vp-porin structure. The η symbol refers to a 310-helix. Helices and strands are shown as black helices with squiggles and arrows, respectively. Red box, white character indicates strict identity; (B) Secondary structure of vp-porin using SOPMA; (C) 3D Structure of Vp-porin using Swiss-model.
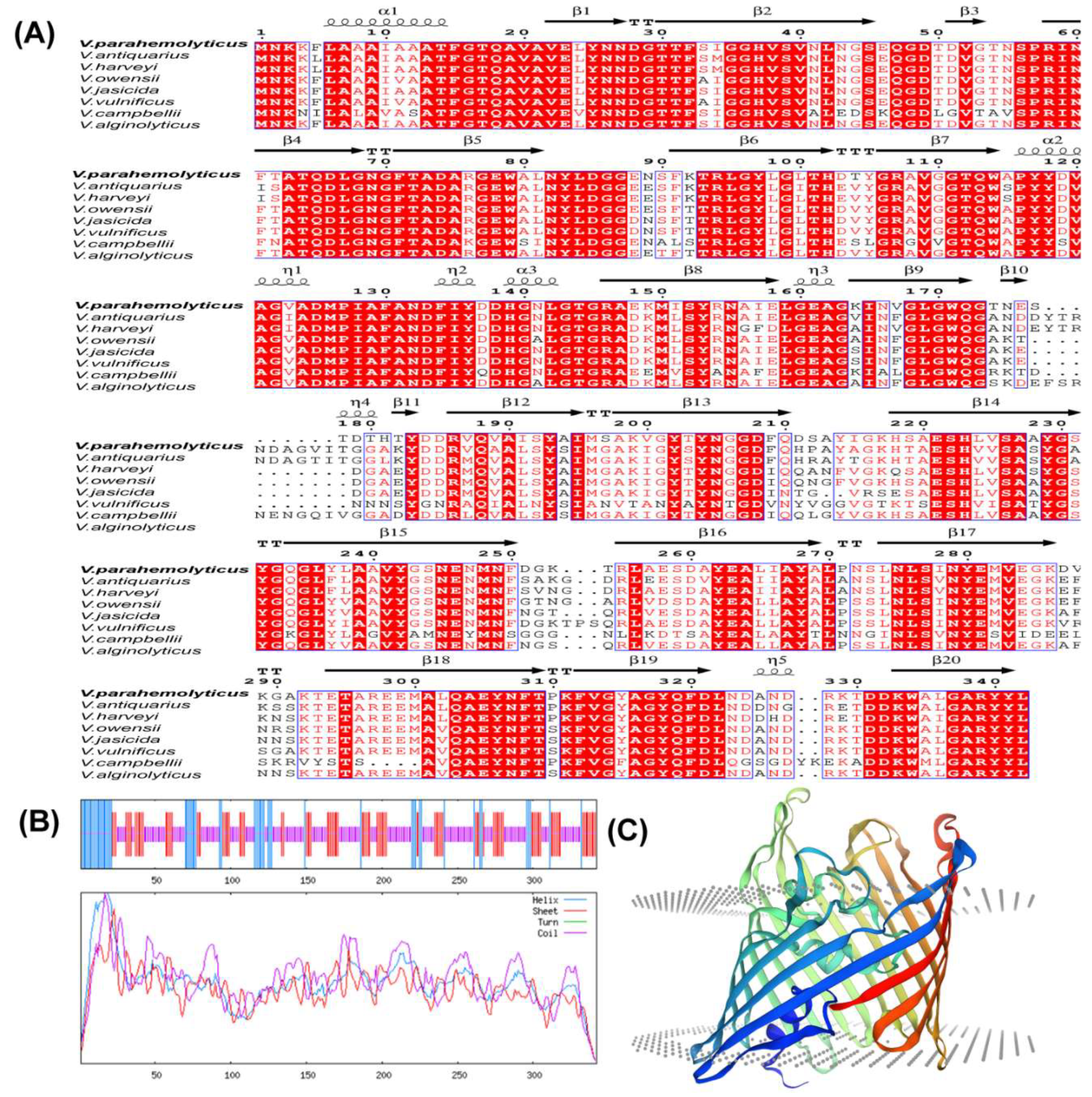
Figure 2.
Construction of the Vp-porin deletion mutant strain of V. parahaemolyticus and phenotype characterization of ∆Vp-porin. (A) Construction of strain of V. parahaemolyticus. (B) The deletion of Vp-porin was confirmed by colony PCR. (C) Growth of three strains on agar media and colony morphology following incubation for 24 h; (D) Growth curves in 3% NaCl LB medium over 12 h period; (E) Protease production on 2% skim milk agar; (F) Zone of proteolysis (mm) surrounding bacterial colonies after 48 h incubation at 37°C. Columns have been marked with an asterisk (*p < 0.05; **p < 0.01; ***p < 0.001).
Figure 2.
Construction of the Vp-porin deletion mutant strain of V. parahaemolyticus and phenotype characterization of ∆Vp-porin. (A) Construction of strain of V. parahaemolyticus. (B) The deletion of Vp-porin was confirmed by colony PCR. (C) Growth of three strains on agar media and colony morphology following incubation for 24 h; (D) Growth curves in 3% NaCl LB medium over 12 h period; (E) Protease production on 2% skim milk agar; (F) Zone of proteolysis (mm) surrounding bacterial colonies after 48 h incubation at 37°C. Columns have been marked with an asterisk (*p < 0.05; **p < 0.01; ***p < 0.001).
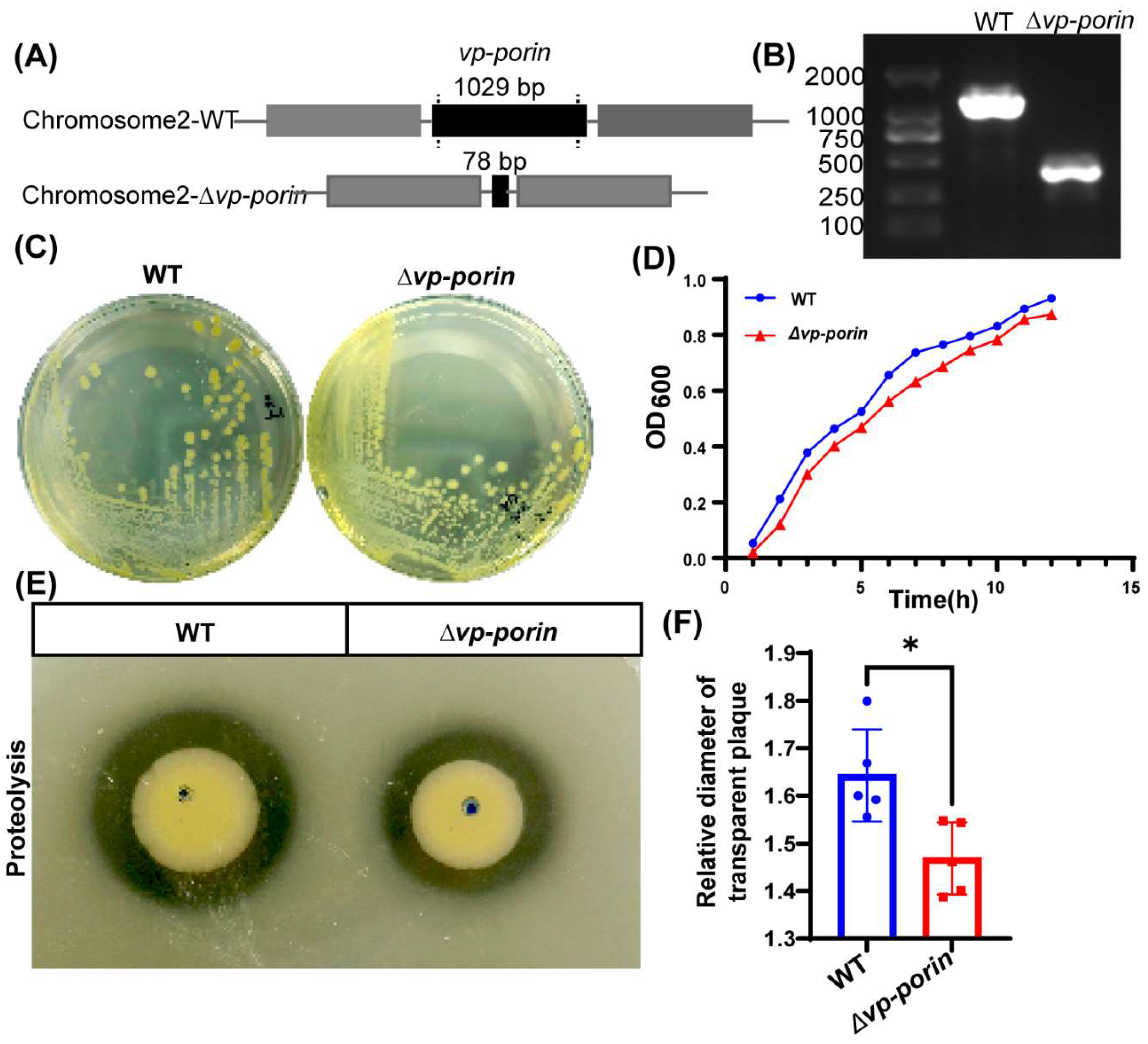
Figure 3.
Comparison of outer membrane permeability of WT and ∆Vp-porin. The fluorescence intensity was determined with excitation at 350 nm and emission at 428 nm. Each bar represents as the mean ± SD. n=10. Columns have been marked with an asterisk (*p < 0.05; **p < 0.01; ***p < 0.001).
Figure 3.
Comparison of outer membrane permeability of WT and ∆Vp-porin. The fluorescence intensity was determined with excitation at 350 nm and emission at 428 nm. Each bar represents as the mean ± SD. n=10. Columns have been marked with an asterisk (*p < 0.05; **p < 0.01; ***p < 0.001).
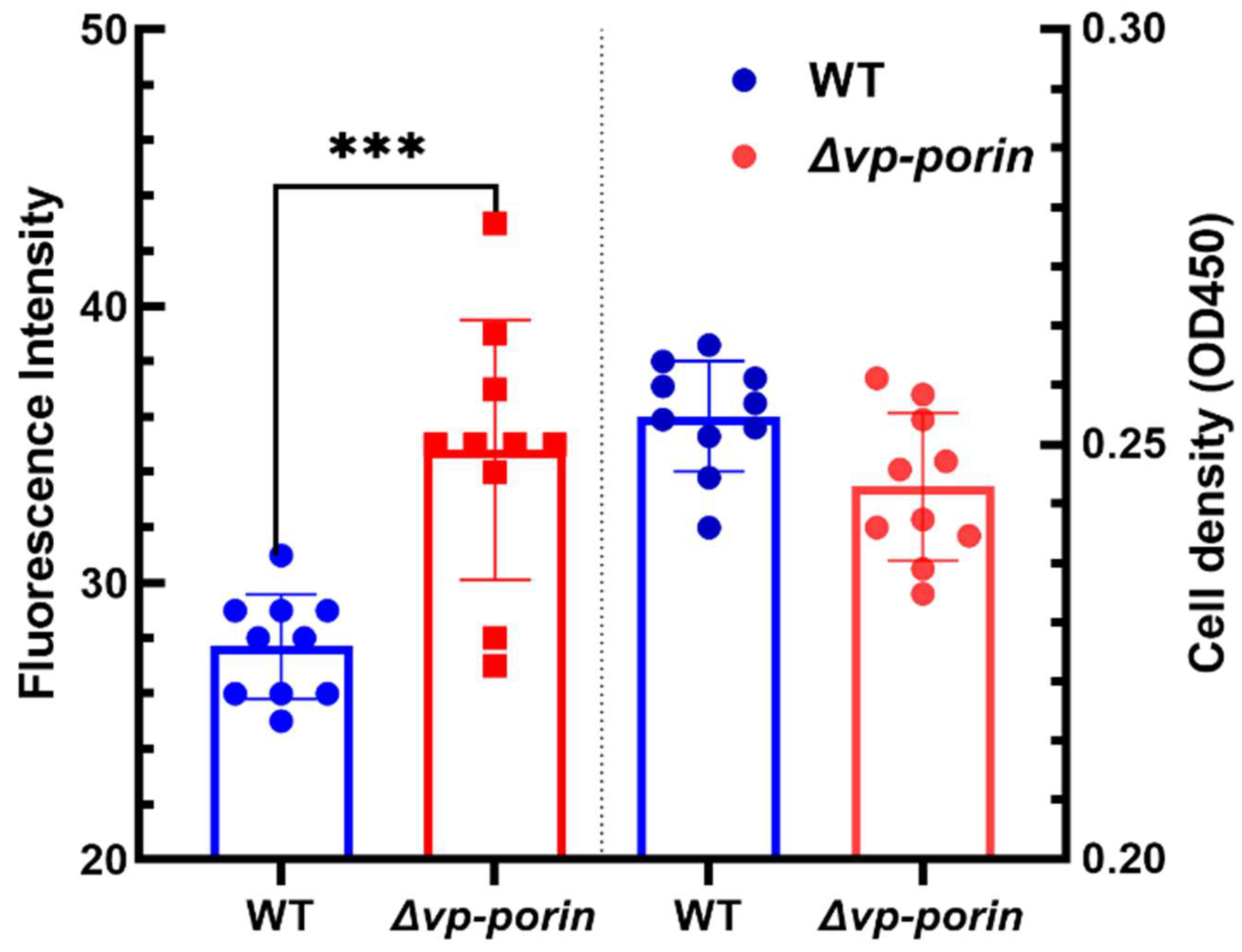
Figure 4.
Antimicrobial activity of WT and ∆Vp-porin. (A) Antimicrobial screening showing zones of inhibition; (B) Inhibition zone was calculated to determine the antimicrobial activity of WT and ∆Vp-porin to various antimicrobials; (C) Survival of WT and ∆Vp-porin in the presence of varying concentrations of rifampicin. Columns have been marked with an asterisk (*p < 0.05; **p < 0.01; ***p < 0.001).
Figure 4.
Antimicrobial activity of WT and ∆Vp-porin. (A) Antimicrobial screening showing zones of inhibition; (B) Inhibition zone was calculated to determine the antimicrobial activity of WT and ∆Vp-porin to various antimicrobials; (C) Survival of WT and ∆Vp-porin in the presence of varying concentrations of rifampicin. Columns have been marked with an asterisk (*p < 0.05; **p < 0.01; ***p < 0.001).
Figure 5.
Vp-porin regulates swimming and swarming motility in V. parahaemolyticus. (A) Swimming motility assay and Swarming motility assay of WT and ∆Vp-porin on LB plates with 0.3% and 1.5% agar at 37 °C; (B, C) Analyze of swimming (B) and swarming (C) of WT, ∆Vp-porin strains in LB medium respectively. The Diameters of swimming zone reflecting bacterial migration on the 0.3% agar. The data are presented as the mean ± SD (n = 3). Columns have been marked with an asterisk (*p < 0.05; **p < 0.01; ***p < 0.001); (D) qRT-PCR analysis of the transcription levels of polar flagellar cluster I genes (flgB, flgM, flgK, flgC) and polar flagellar cluster II genes (fliE, fliK) in ∆Vp-porin compared to WT. The data are presented as the mean ± SD (n = 3). *p < 0.05; **p < 0.01; ***p < 0.001; (E) qRT-PCR analysis of the transcription levels of lateral flagellar cluster I (flgA, flgB, flgG, lafA, motY) and lateral flagellar cluster II (fliE) genes in ∆Vp-porin compared to WT. The data are presented as the mean ± SD (n = 3). ***p < 0.001. Student’s t test analyzes ∆Vp-porin compared to WT; (F) qRT-PCR analysis of the expression levels of genes located in T6SS2 in WT, ∆Vp-porin strains. The data are presented as the mean ± SD (n = 3). Student’s t test analyzes the different mutant strains compared to WT. ***p < 0.001.
Figure 5.
Vp-porin regulates swimming and swarming motility in V. parahaemolyticus. (A) Swimming motility assay and Swarming motility assay of WT and ∆Vp-porin on LB plates with 0.3% and 1.5% agar at 37 °C; (B, C) Analyze of swimming (B) and swarming (C) of WT, ∆Vp-porin strains in LB medium respectively. The Diameters of swimming zone reflecting bacterial migration on the 0.3% agar. The data are presented as the mean ± SD (n = 3). Columns have been marked with an asterisk (*p < 0.05; **p < 0.01; ***p < 0.001); (D) qRT-PCR analysis of the transcription levels of polar flagellar cluster I genes (flgB, flgM, flgK, flgC) and polar flagellar cluster II genes (fliE, fliK) in ∆Vp-porin compared to WT. The data are presented as the mean ± SD (n = 3). *p < 0.05; **p < 0.01; ***p < 0.001; (E) qRT-PCR analysis of the transcription levels of lateral flagellar cluster I (flgA, flgB, flgG, lafA, motY) and lateral flagellar cluster II (fliE) genes in ∆Vp-porin compared to WT. The data are presented as the mean ± SD (n = 3). ***p < 0.001. Student’s t test analyzes ∆Vp-porin compared to WT; (F) qRT-PCR analysis of the expression levels of genes located in T6SS2 in WT, ∆Vp-porin strains. The data are presented as the mean ± SD (n = 3). Student’s t test analyzes the different mutant strains compared to WT. ***p < 0.001.
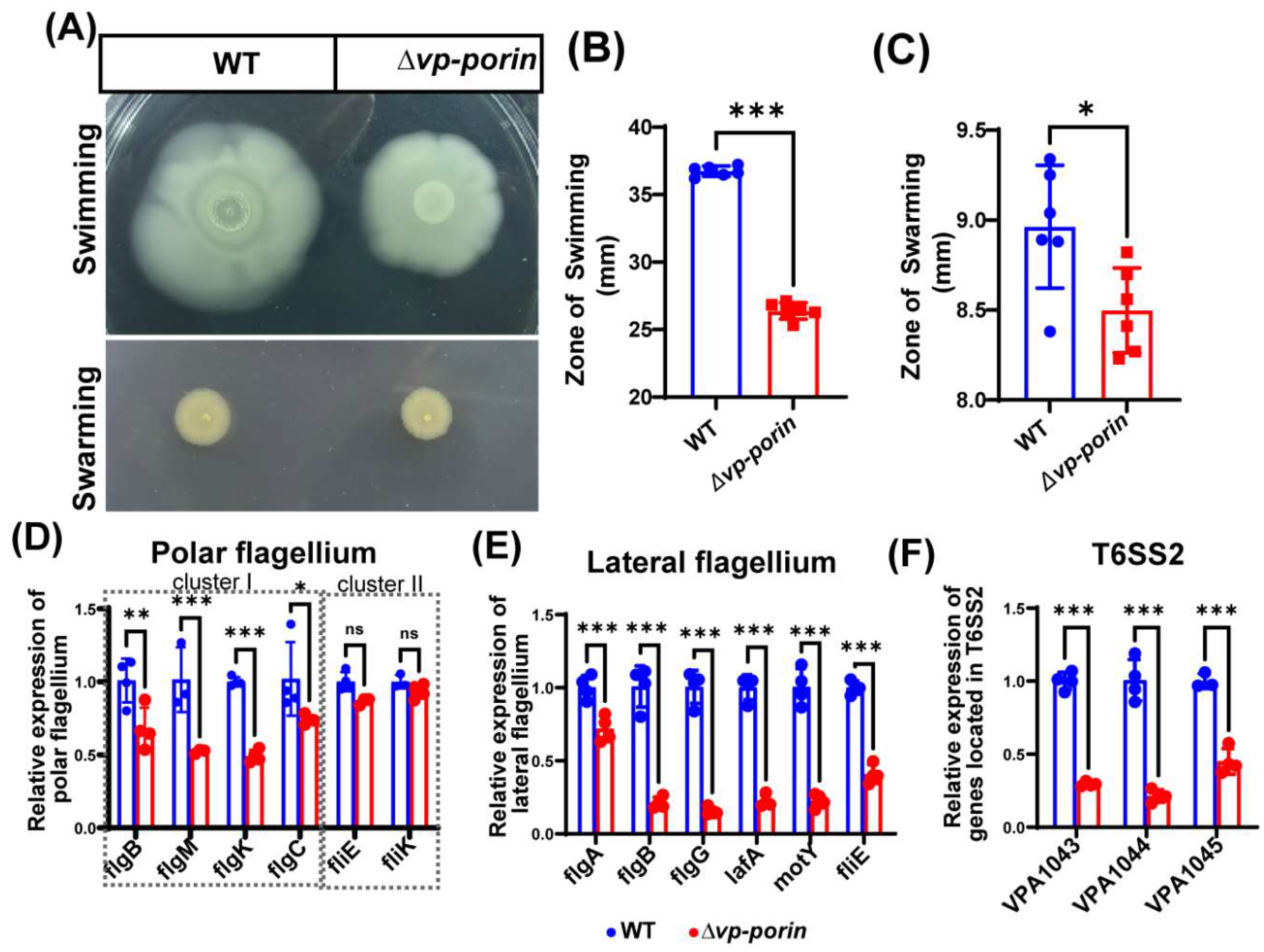
Figure 6.
Assessment virulence of WT, ∆Vp-porin using Tetrahymena. (A) Morphological changes of Tetrahymena cells after co-cultivation with ∆Vp-porin and WT strains. Red arrow represents abnormal cell; Black arrow represent normal cell. Scale bar: 200 μm; (B-C) Growth of ∆Vp-porin and WT co-cultured in the presence or absence of Tetrahymena; (D) Relative survival of ∆Vp-porin and WT strains co-cultured with Tetrahymena. The relative survival of bacteria was expressed as the OD450 value of strains co-cultured with Tetrahymena divided by that of bacteria grown alone at different time. Data are expressed as the mean ± SD of three measurements per time point. *** P < 0.001, **** P < 0.0001; (E) Growth of Tetrahymena co-cultivated with ∆Vp-porin and WT strains. The control group was Tetrahymena grown alone in sterile SPP medium. Data are expressed as the mean ± SD of three measurements per time point; (F) Relative survival of Tetrahymena co-cultivated with ∆Vp-porin and WT strains at 2 h, 4 h and 6 h. Relative survival of Tetrahymena is calculated by the number of Tetrahymena cells in the co-culture with different strains relative to that of Tetrahymena cells cultured alone.
Figure 6.
Assessment virulence of WT, ∆Vp-porin using Tetrahymena. (A) Morphological changes of Tetrahymena cells after co-cultivation with ∆Vp-porin and WT strains. Red arrow represents abnormal cell; Black arrow represent normal cell. Scale bar: 200 μm; (B-C) Growth of ∆Vp-porin and WT co-cultured in the presence or absence of Tetrahymena; (D) Relative survival of ∆Vp-porin and WT strains co-cultured with Tetrahymena. The relative survival of bacteria was expressed as the OD450 value of strains co-cultured with Tetrahymena divided by that of bacteria grown alone at different time. Data are expressed as the mean ± SD of three measurements per time point. *** P < 0.001, **** P < 0.0001; (E) Growth of Tetrahymena co-cultivated with ∆Vp-porin and WT strains. The control group was Tetrahymena grown alone in sterile SPP medium. Data are expressed as the mean ± SD of three measurements per time point; (F) Relative survival of Tetrahymena co-cultivated with ∆Vp-porin and WT strains at 2 h, 4 h and 6 h. Relative survival of Tetrahymena is calculated by the number of Tetrahymena cells in the co-culture with different strains relative to that of Tetrahymena cells cultured alone.
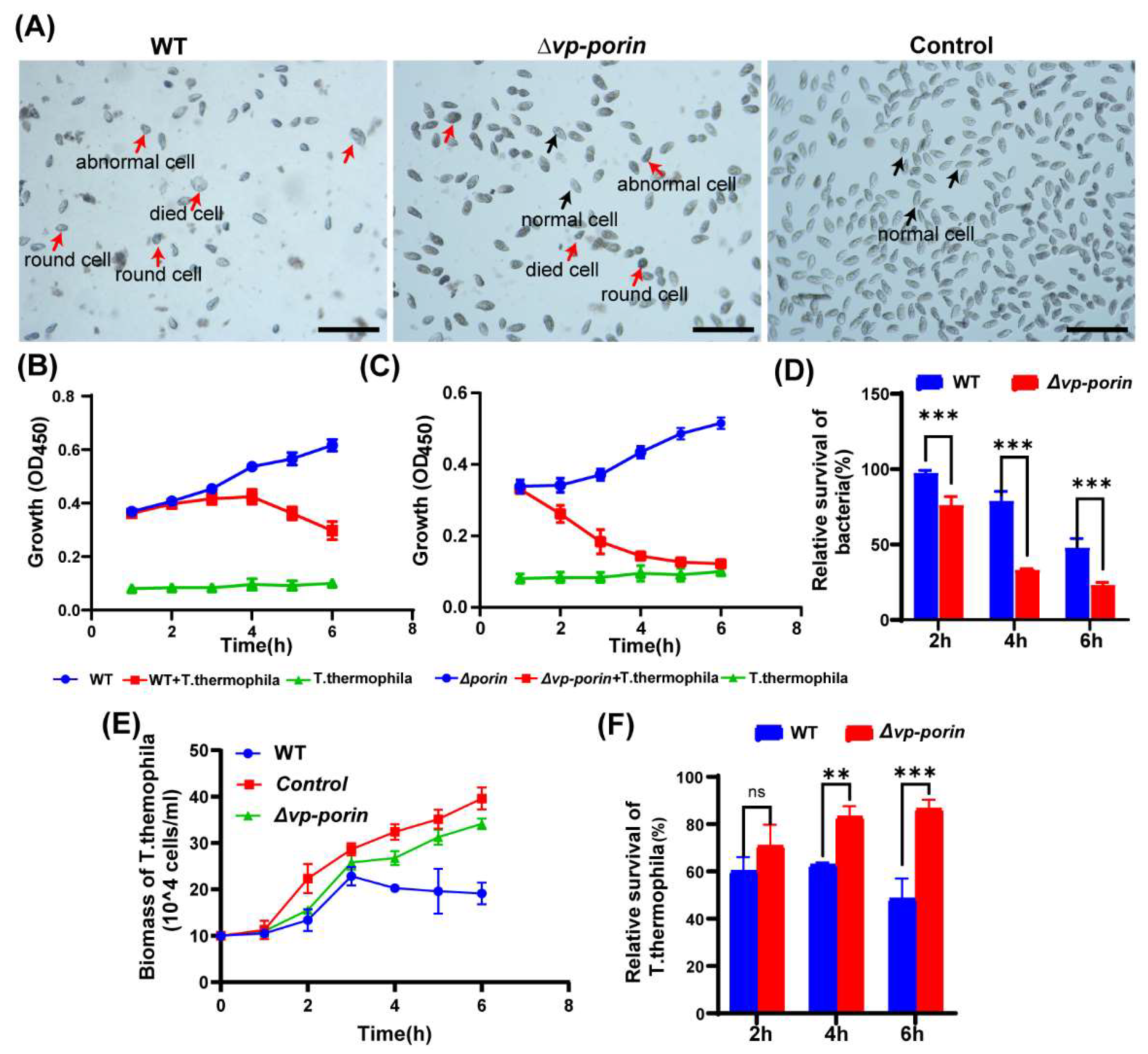

Table 1.
Strains and plasmids.
| Strains | Genotype & Characteristics | Source |
|---|---|---|
| V.parahaemolyticus17802 | Cms, Kms, Ampr, Wild type strain, | ATCC |
| ∆Vp-porin | V.parahaemolyticus strain in-frame deletion in Vp-porin | This study |
| Escherichia coli | ||
| CC118 | λpir lysogen of CC118 (Δ(ara-leu) araD ΔlacX74galEgalKphoA20 thi-1rpsE rpoB argE (Am) recA1 | Our lab |
| CC118/pHelper | CC118 λpir harboring plasmid pHelper | Our lab |
| Plasmids | ||
| pSR47S | Bacterial allelic exchange vector with sacB, KanR | Our lab |
| pSR47S-∆Vp-porin | A 1689 bp fragment containing the upstream and downstream sequences of the ∆Vp-porin gene in pSR47S, KanR | This study |
Table 2.
Sequences of PCR oligonucleotide primers.
| Primer Name | Primer Sequence (5’ to 3’) | Purpose |
|---|---|---|
| UP-F | CGAGCTCCTTGATGGACTTCGCCAAC | Creation of ∆Vp-porin deletion fusion fragment |
| UP-R | CAACATTCGGTACTCAAGCAGCACTTGGTGCACGTTACTAC | |
| DOWN-F | GTAGTAACGTGCACCAAGTGCTGCTTGAGTACCGAATGTTG | |
| DOWN-R | GACTAGTGTACACACCGAATGCAGAC | |
| Vp-porin-T1 | GAACAACACTAGAACGCGC | Comfirmation of ∆Vp-porin deletion |
| Vp-porin-T2 | TCGGTTACCGAAGAGTCTTC | |
| Note: Restriction sites are italic. Complementary sites are underlined. | ||
Table 3.
Primers used for q RT-PCR.
| Primer name | Primer sequence (5' to 3') | Target |
|---|---|---|
| flgB-F | ACAAGGCACTAGGCATCC | polar flagellar cluster I genes |
| flgB-R | GACCATCTGTTCGGCTAAG | |
| flgC-F | GCGTCATGCTGTATTTGGTG | |
| flgC-R | AACCTGCACATTCGTTTGGT | |
| flgM-F | ATTCAAGTGCGACATCAAG | |
| flgM-R | CGGAGAAGCTGCCATATC | |
| flgK-F | GCCGTCAGTCAGTGATTC | |
| flgK-R | GTAGAGGACAGGTTGAGTTC | |
| fliE-F | CACTGTGCCCGTTTGCTTAC | polar flagellar cluster II genes |
| fliE-R | TCCGGCGGATGCTTCTATTC | |
| fliK-F | GTCGAGAAGAATGGCGAGAG | |
| fliK-R | CCAACTGAGCCTCTGACTCC | |
| flgA-F | TACCGACTGGCAAAGGTTGG | |
| flgA-R | TACCGACTGGCAAAGGTTGG | lateral flagellar cluster I genes |
| flgB-F | GCAGGTTCAGGCCCAGTATT | |
| flgB-R | TCATGTTGAGAAACGTCAGGCT | |
| flgG-F | AGATCTAGCGGTAATGGGGC | |
| flgG-R | GAGAAAGAGGTCGCGTTGTC | |
| lafA-F | GCTGGTGGCCTTATCGAAGA | |
| lafA-R | TACTGCGAAGTCTGCATCCAT | |
| motY-F | ATTAGTGAGGGTGCGCCTTT | |
| motY-R | GGTGAAGGGAAGGAATGGCA | |
| fliE-F | CGCTTGAGAAAACGACAGTGG | lateral flagellar cluster I genes |
| fliE-R | CCTACTAATGCGGTCTCGGC | |
| VPA1043-F | TCGAACAGCACGTAGAATCG | T6SS2 genes |
| VPA1043-R | GTGGCACTTCAGTTTCGTGA | |
| VPA1044-F | TCCTCAACCAAATCCTCGAC | |
| VPA1044-R | GCGTAGTTAGGCGTGTAGCC | |
| VPA1045-F | CCGATGCTCAATGGCTTAAT | |
| VPA1045-R | GCTGCTCTTTACCCAACTGC | |
| 16s rRNA-F | TTAAGTAGACCGCCTGGGGA | qPCR of 16s rRNA |
| 16s rRNA-R | GCAGCACCTGTCTCAGAGTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



