
Submitted:
31 May 2024
Posted:
06 June 2024
You are already at the latest version
Abstract
Fluorinated cytokinins have emerged as promising alternatives to traditional cytokinins in plant tissue culture, offering enhanced stability and bioactivity. However, their metabolic fate and impact on endogenous cytokinin profiles remain largely unexplored. This study investigated the cytokinin metabolism of Phalaenopsis orchids under varying exogenous cytokinin treatments (BA, FmT, FmTR) and culture conditions (open vs. closed containers). The results reveal distinct metabolic profiles associated with each treatment, highlighting the complex interplay between exogenous and endogenous cytokinin levels. Additionally, we proposed an alternative inactivation pathway involving the conversion of FmTR and BA to pT and pTR. These findings provide valuable insights into the intricate relationship between cytokinin metabolism and plant growth under in vitro conditions, with implications for optimizing micropropagation protocols
Keywords:
Phalaenopsis
; orchid
; cytokinin metabolism
1. Introduction
Our recent research has demonstrated that fluorinated cytokinins significantly enhance shoot proliferation in orchids grown in vitro, particularly in hermetically sealed containers [1]. However, the mechanisms behind this phenomenon remain elusive. To explore this subject, we performed a study focusing on the metabolites of the traditionally used BA-type cytokinin and “second” generation topolins 6-(3-fluorobenzylamino)purine (FmT) and 6-(3-fluorobenzylamino)purine 9-riboside (FmTR) with supposed improved stability and bioactivity [2]. Understanding the metabolic response of orchids to fluorinated cytokinins is crucial for developing optimized micropropagation protocols, not only for orchids but potentially for other recalcitrant plant species as well.
Cytokinins, a class of essential plant hormones, play a pivotal role in in vitro plant culture. These phytohormones regulate numerous developmental processes, including meristem maintenance, cell division, shoot formation, and leaf senescence [3]. During in vitro micropropagation, the exogenous application of cytokinins is crucial for achieving optimal plant multiplication and quality [4,5]. Plants have evolved complex mechanisms to maintain homeostasis between the exogenous supply and endogenous levels of cytokinins through intricate metabolic pathways, essential for overall plant growth and development [6]. Despite their widespread use in micropropagation, the comprehensive metabolic profiles of cytokinins have been elucidated only for a limited number of plant species. Moreover, the relative abundance of various cytokinin metabolites is influenced by the specific type of cytokinin applied and exhibits interspecies variability [7,8].
Upon absorption from the culture medium, cytokinins are subjected to extensive metabolic transformations within plant tissues, resulting in a dynamic array of molecular forms with varying biological activities. The metabolic profile encompasses active forms, predominantly free bases like BA, mT, 2iP, and Z, which directly influence plant growth and development. These active forms are often converted into less active transport forms, including ribosides and ribotides, facilitating their movement within the plant. Additionally, cytokinins can be reversibly stored as O-glucosides, inactive conjugates that can be readily hydrolyzed back to their active free base forms when required. Conversely, N-glucosides are generally considered to be irreversible and completely inactive storage forms [9,10]. This intricate metabolic network allows plants to tightly regulate cytokinin levels in response to developmental and environmental cues, a critical factor in maintaining plant homeostasis. In the context of in vitro culture, where exogenous cytokinin application and the absence of natural environmental cues can disrupt this delicate balance, understanding and manipulating cytokinin metabolism is paramount for achieving optimal conditions for de novo organogenesis and successful micropropagation [11,12,13,14]. We investigated the metabolic profile of cytokinins under two factors affecting the micropropagation of Phalaenopsis: (1) cytokinin type and (2) headspace aeration [1]. We demonstrate the balance between endogenous and exogenous cytokinins and interaction with the culture conditions.
2. Materials and Methods
2.1. Plant Material and Growth Conditions
An unnamed cultivar of Phalaenopsis amabilis served as the model plant for this study. The initial plant material was propagated on 5/8 MS (Murashige and Skoog, 1962) basal medium supplemented with 10 µM FmTR. Following a two-month growth period, these shoots were transferred to fresh medium of the same composition, but supplemented with one of three different cytokinins: 6-(3-fluorobenzylamino)purine (FmT), 6-(3-fluorobenzylamino)purine 9-riboside (FmTR) (Olchemim, Czech Republic), or N6-benzyladenine (BA), each at a fixed concentration of 10 µM.
2.2. Cytokinin Analyses
Endogenous plant hormones level was measured in in vitro shoots collected from two types of containers, 1) hermetically closed polypropylene container and 2) container with ‘Large’ filter (‘L’ = #10filter (Microbox™, Belgium). Shoots were grown for 10 weeks before analysis, and leaf from ex vitro grown Phalaenopsis plant was collected and used as a control/reference sample.
Five random plants of each treatment per condition were selected. shoot parts were removed weighed and lyophilized. All samples were stored at -80 ͦC before CK analysis. Cytokinin metabolites were quantified using a modified protocol [15]. In brief, lyophilized samples were finely ground in liquid nitrogen and 5 mg of dry weight was aliquoted for analysis. Subsequently, 1 µl of cold extraction solution (Modified Bieleski buffer MeOH/H2O/HCOOH (15/4/1), Bieleski, 1964) and 23 deuterium-labelled cytokinin (CK) internal standards (Olchemim Ltd., Czech Republic) were added to each sample. The internal standards, applied at concentrations of 1 pmol (free bases) and 2 pmol (O-glucoside, nucleotides) per sample, served to assess the recovery of target cytokinins during purification and validate their quantification. Following homogenization in a vibration mill (27 Hz, 3 min), sonication (4°C, 5 min), and agitation (4°C, 30 min), samples were centrifuged (20,000 rpm, 10 min, 4°C) and the resulting pellets were re-extracted using the same procedure. The collected supernatants were then subjected to a multi-step purification process involving Octadecyl C18 (100 mg/1 ml) speed SPE cartridges, Oasis MCX (30 mg/1 ml) columns, and immunoaffinity chromatography (IAC). Blank samples were prepared in parallel using an identical procedure.
Detailed purification and quantification procedures were previously described [16]. Briefly, samples were analyzed using an Acquity UPLC™ system (Waters) coupled to a Xevo™ TQ MS™ (Waters) triple quadrupole mass spectrometer equipped with an electrospray ionization (ESI) interface. CK quantification was achieved via multiple reaction monitoring (MRM) of [M + H]+ and the appropriate product ions. Optimal MRM conditions (dwell time, cone voltage, collision energy) were determined for each CK. The ratio of CKs to their corresponding labelled standards was used to quantify CK levels based on the known internal standard concentrations [15]. All CK quantifications were performed in triplicate.
2.3. Statistical Analysis
The experiment followed a completely randomized design. Three repetitions for each sample were carried out to assess CKs. The data was analyzed using SPSS software, version 25.
3. Results
3.1. Effect of Exogenously Administered Cytokinins (CKs) on Total Quantified CK Levels and Their Metabolic Profile
The total content of cytokinins (CKs) pmol / 1g FW was determined in eight-week-old in vitro shoots of Phalaenopsis grown in the presence of 10 μM BA, 10 μM FmT, and 10 μM FmTR, and compared between plants grown in filtered or hermetically sealed containers. A total of 48 different cytokinins and their metabolites were detected in varying concentrations and is listed in Table 1. The concentrations of different cytokinin types vary depending on the treatment and container type, both aromatic cytokinins such as BA; ortho-topolin (oT), meta-topolin (mT), para-topolin (pT) and their derivatives, and isoprenoid cytokinins such as trans -Zeatin (tZ), cis-Zeatin (cZ), dihydrozeatin (DHZ), isopentenyladenine (iP) and its derivatives were detected, with diverse abundance across treatments and container types.
The presence and abundance of specific cytokinin metabolites varied significantly under different treatment and culture conditions. For instance, tZ7G, present in trace amounts under BA treatment, was completely absent in plants grown in closed containers and supplemented with FmT or FmTR. Additionally, the abundance of cZ and its riboside form, cZR, exhibited distinct patterns depending on treatment and container type. While cZ remained detectable at minimal concentrations in BA-treated plants, it fell below the limit of detection (LOD) in FmT or FmTR-treated plants. Intriguingly, cZR was consistently abundant across all treated plants. Although cis-Zeatin was previously considered biologically inactive, potentially as a byproduct of tRNA degradation, recent studies challenge this notion and suggest its involvement in various stages of plant development [17]. Notably, neither cZ nor its metabolites were detected in control leaves from ex vitro plants.
Endogenous isoprenoid metabolites, specifically the reversible storage form DHZROG and the transport form IPR, exhibited higher levels in filter pots compared to closed containers, contrary to expectations. As anticipated, BA treatment generally led to elevated concentrations of BA, BAR, and their derivatives, while FmT and FmTR treatments resulted in the accumulation of fluorinated cytokinins. Notably, less glucosylation of BA occurred in closed containers, with higher levels of free BA observed.
Interestingly, limited amounts of FmT were presumably de-fluorinated to yield BA in both open and closed containers, a phenomenon not observed with FmTR. Another intriguing observation regarding potential regulatory mechanisms of cytokinin metabolism in Phalaenopsis was the presence of pT and pTR, proposed deactivation products of mF and mFR treatment, suggesting that mFR may be degraded to BA and BAR, which are subsequently converted to pT and pTR. Because para-topolin and its derivatives exhibit very low biological activity [21], this pathway can serve as an alternative form of inactivation, because it is well known that aromatic cytokinins are less susceptible to degradation by CKX enzyme [22]. However, identity of products has to be confirmed by high resolution mass spectrometry. Notably, putative pT and pTR levels significantly declined in closed containers. The presence of FmT and FmTR in BA-treated plants is not unexpected, as their previous subculture involved FmTR, and its stability allows for carryover to subsequent subcultures.
To further investigate the role of treatment and culture condition on in vitro Phalaenopsis development, we next, compared absolute concentration of cytokinin between containers. The highest level of total cytokinin was observed in FmTR-treated plants in closed containers. It appears that added FmTR did not degraded thus represented the largest fraction of the total CK pool across all treatment. Figure 1.a. Notably, culture condition not only affected the total CK pool but also the ration between IsCKs and ArCKs levels as shown in Figure 1b. analysis result shows that, levels of IsCKs and ArCKs vary significantly across the treatments, for instance, the highest concentration of IsCKs is observed in BA (F) treated shoots, representing 38.97% proportion of total CK content, whereases the lowest concentration is founded in the FmTR (Cl) treatment, representing approximately 2.65% of total CKs.
Data analysis revealed a correlation between the levels of isoprenoid cytokinins (IsCKs) and the observed number of de novo shoots in Phalaenopsis plants, consistent with previous findings [1]. Additionally, within the same treatment group, a higher concentration of aromatic cytokinins (ArCKs) was associated with an increased number of shoots, particularly evident in plants grown in closed containers (Cl). This observation suggests that aromatic cytokinins may play a beneficial role in Phalaenopsis.
Figure 2 illustrates the distribution of cytokinin (CK) metabolites in Phalaenopsis under different treatments and container types. In filter containers (FC), ribosides constituted the majority of the total CK content across all treatments, consistent with their role as long-distance transport forms capable of interconversion into active forms [3,18]. Free bases represented the second largest proportion, with varying percentages depending on the applied cytokinin (BA: 26%; FmT: 42%; FmTR: 7%). In closed containers (CC), free bases predominated, except in the case of FmTR treatment.
O-glucosides, detectable only in FmT and BA-treated plants (FC), accounted for a small proportion of the total CK pool (3% with BA, 6% with FmT). These reversible storage forms can contribute to the active CK pool upon hydrolysis [18,19]. In CC, only negligible amounts of cZROG were found. N-glucosides were largely below the detection limit, except in BA-treated plants where tZ7G and BAP9G were present. The predominance of BAP9G suggests that 9-glycosylation is a primary BA deactivation pathway in Phalaenopsis. Notably, FmT-treated plants exhibited extremely low levels of N-glucosides.
Interestingly, the presence of BA, pT, and pTR as metabolites of mF and mFR treatment suggests a potential alternative regulatory pathway in Phalaenopsis, whereby mFR is degraded to BA and BAR, subsequently converted to the inactive pT and pTR. In future experiments, we will make an effort to confirm existence of this novel pathway as well as the identity of formed catabolites by high resolution mass spectrometry. complete lack of pT and pTR in closed containers highlights the influence of container type on cytokinin metabolism (Table 2).
4. Discussion
Our study reveals a complex interplay between exogenous cytokinin application, headspace aeration, and the resulting endogenous cytokinin profiles in Phalaenopsis. The differential accumulation of specific metabolites under varying treatments and container types underscores the dynamic nature of cytokinin metabolism in this orchid species. Notably, the absence of tZ7G in plants treated with FmT or FmTR, particularly in closed containers, suggests a potential inhibitory effect of these fluorinated cytokinins on the N7-glucosylation pathway. Furthermore, the unexpected abundance of cZR across all treatments, despite the low or undetectable levels of its precursor cZ, raises questions about the biological significance of these compounds in Phalaenopsis. While traditionally considered inactive, recent evidence suggests a potential role for cZ in plant development [17], and the absence of cZ and its metabolites in ex vitro leaves further supports the notion that this cytokinin may be specifically involved in responses to in vitro conditions.
Interestingly, isoprenoid metabolites DHZROG and IPR were found at higher levels in filter pots compared to closed containers, contrary to the observation that closed containers typically yield more shoots [1]. This finding suggests that the fluorinated cytokinins FmT and FmTR, as well as headspace aeration, may modulate isoprenoid cytokinin biosynthesis or metabolism. Additionally, the de-fluorination of FmT to BA was proposed, putatively observed in both open and closed containers but not measured with FmTR, highlights the potential for metabolic interconversions between different cytokinin types and underscores the importance of considering the stability and metabolic fate of synthetic cytokinins in plant tissue culture.
The abundance of total cytokinin content in closed containers suggests that a hermetically sealed environment might prevent the degradation of certain cytokinins, potentially explaining the earlier reported increased shoot proliferation under closed containers [1]. However, the exact mechanism behind this action remains unclear.
Notably, hydroxylated metabolites of BA, such as mT, oT, and pT, were observed in our study, despite not being detected in greenhouse-grown Phalaenopsis. Previous research has documented the presence of oT and pT, along with their derivatives, in other plant species under in vitro culture conditions, suggesting a potential role of hydroxylation in regulating the activity of aromatic cytokinins [20,21,23].
The putative presence of pT and pTR as metabolites of mF and mFR treatment suggests a novel metabolic pathway in Phalaenopsis, representing a potential mechanism for regulating cytokinin activity in this orchid. However, this has to be confirmed by future HR-MS identification experiments. The decline of pT and pTR levels in closed containers may be linked to altered metabolic flux under these conditions.
In conclusion, our study provides new insights into the intricate relationship between exogenous cytokinin application, headspace aeration, and endogenous cytokinin metabolism in Phalaenopsis. These findings have important implications for optimizing micropropagation protocols and understanding the complex regulatory networks governing plant growth and development. Our findings challenge future research on the possible mechanisms underlying cytokinin metabolism and ethylene crosstalk in plant in vitro culture.
5. Conclusions
In conclusion, our investigation into the cytokinin metabolism of Phalaenopsis under varying exogenous cytokinin treatments and culture conditions has revealed a complex and dynamic system. The differential accumulation of specific metabolites highlights the intricate interplay between exogenous and endogenous cytokinin levels, influenced by both the chemical structure of the applied cytokinin and the culture environment.
The observed metabolic alterations, such as the unexpected abundance of cZR, and proposed de-fluorination of FmT, underscore the need for a deeper understanding of cytokinin metabolism in orchids. The identification of a potential novel metabolic pathway involving pT and pTR further emphasizes the complexity of this system.
These findings have significant implications for optimizing Phalaenopsis micropropagation protocols. By tailoring cytokinin treatments and culture conditions to specific metabolic profiles, we can potentially enhance shoot proliferation, improve plant quality, and overcome recalcitrance in orchid tissue culture. Moreover, our results contribute to the broader understanding of cytokinin metabolism in plants and its role in regulating growth and development. Further research is warranted to elucidate the precise mechanisms underlying these metabolic processes and to explore their potential applications in plant biotechnology.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, NM and SPOW; methodology, NM validation, SPOW and KD; formal analysis, NM an KD.; investigation, SPOW; resources, SPOW; ; writing—original draft preparation, NM; writing—review and editing, NM and SPOW; visualization, SPOW.; supervision, SPOW;. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Data Availability Statement
all data are included in the article
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Murvanidze N, Doležal K, Werbrouck SP. Fluorine containing topolin cytokinins for Phalaenopsis Amabilis (L.) blume micropropagation. Propag Ornam Plants. 2019, 19, 48–51.
- Plíhalová L. Synthesis and chemistry of meta-Topolin and related compounds. Meta-topolin: A Growth Regulator for Plant Biotechnology and Agriculture. 2021, 11–22.
- Kieber JJ, Schaller GE. Cytokinin signaling in plant development. Development 2018, 145(4), dev149344. [CrossRef] [PubMed]
- Singh J, Sengar RS, Prasad M, Kumar A. Effect of Different Plant Growth Regulators on In-vitro Micropropagation of Banana Cultivar Grand Naine (Musa spp.). J Adv Biol Biotechnol. 2024, 27(3), 90–98.
- Mazri MA. Role of cytokinins and physical state of the culture medium to improve in vitro shoot multiplication, rooting and acclimatization of date palm (Phoenix dactylifera L.) cv. Boufeggous. J Plant Biochem Biotechnol. 2015, 24, 268–275. [CrossRef]
- Zürcher E, Müller B. Cytokinin synthesis, signaling, and function—advances and new insights. Int Rev Cell Mol Biol. 2016, 324, 1–38.
- Aremu AO, Bairu MW, Szüčová L, Doležal K, Finnie JF, Van Staden J. Genetic fidelity in tissue-cultured ‘Williams’ bananas–The effect of high concentration of topolins and benzyladenine. Sci Hortic. 2013, 161, 324–327. [CrossRef]
- Ahmad MZ, Hussain I, Roomi S, Zia MA, Zaman MS, Abbas Z, Shah SH. In vitro response of cytokinin and auxin to multiple shoot regeneration in Solanum tuberosum L. Am Eurasian J Agric Environ Sci. 2012, 12(11), 1522–1526.
- Sakakibara H. Cytokinins: activity, biosynthesis, and translocation. Annu Rev Plant Biol. 2006; 57:431-449.
- Mok DWS, Mok MC. Cytokinin metabolism and action. Annu Rev Plant Biol. 2001, 52, 89–118. [CrossRef] [PubMed]
- Klemš M, Plačková L, Doležal K, Bettaieb T, Werbrouck SP. Changes in cytokinin levels and metabolism in tobacco (Nicotiana tabacum L.) explants during in vitro shoot organogenesis induced by trans-zeatin and dihydrozeatin. Plant Growth Regul. 2011, 65, 427–437. [CrossRef]
- Abdouli D, Plačková L, Doležal K, Bettaieb T, Werbrouck SP. Topolin cytokinins enhanced shoot proliferation, reduced hyperhydricity and altered cytokinin metabolism in Pistacia vera L. seedling explants. Plant Sci. 2022, 322, 111360. [CrossRef] [PubMed]
- Abdouli M, Plačková L, Doležal K, Bettaieb T, Werbrouck SP. Cytokinin regulation in in vitro plant propagation. J Plant Growth Regul. 2022.
- Smykalova I, Plačková L, Doležal K, Bettaieb T, Werbrouck SP. The role of cytokinin regulation in micropropagation. Plant Cell Rep. 2019, 38, 123–132.
- Novák O, Hauserová E, Amakorová P, Doležal K, Strnad M. Cytokinin profiling in plant tissues using ultra-performance liquid chromatography–electrospray tandem mass spectrometry. Phytochemistry. 2008, 69(11), 2214–2224.
- Plačková L, Hrdlička J, Smýkalová I, Cvečková M, Novák O, Griga M, Doležal K. Cytokinin profiling of long-term in vitro pea (Pisum sativum L.) shoot cultures. Plant Growth Regul. 2015, 77, 125–132. [CrossRef]
- Schäfer M, Plačková L, Doležal K, Bettaieb T, Werbrouck SP. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J Exp Bot. 2015, 66(16), 4873–4884.
- Kamada-Nobusada T, Sakakibara H. Molecular basis for cytokinin biosynthesis. Phytochemistry 2009, 70(4), 444–449. [CrossRef] [PubMed]
- Kieber JJ, Schaller GE. Cytokinins. The Arabidopsis Book/American Society of Plant Biologists. 2014;12.
- Werbrouck SP, Doležal K, Strnad M, Van Staden J. Meta-topolin, an alternative to benzyladenine in tissue culture? Physiol Plant. 1996, 98(2), 291–297. [CrossRef]
- Van Staden J, Drewes FE. The stability and metabolism of benzyladenine glucosides in soybean callus. J Plant Physiol. 1992, 140(1), 92–95. [CrossRef]
- Galuszka P, Popelková H, Werner T, Frébortová J, Pospíšilová H, Mik V, Köllmer I, Schmülling T, Frébort I. Biochemical characterization of cytokinin oxidases/dehydrogenases from Arabidopsis thaliana expressed in Nicotiana tabacum L. Journal of Plant Growth Regulation. 2007 Sep; 26:255-67.
- Holub J, Plačková L, Doležal K, Strnad M. Biological activity of cytokinins derived from ortho-and meta-hydroxybenzyladenine. Plant Growth Regul. 1998, 26, 109–115. [CrossRef]
Figure 1.
Absolute cytokinin content, and Isoprenoid and Aromatic Cytokinin Ratio (%) in Axillary Shoots of Phalaenopsis Grown on Media Supplemented with Different Types of Cytokinins: BA (N6-Benzyladenine), 6-(3-fluorobenzylamino)purine (FmT), and 6-(3-fluorobenzylamino)purine 9-riboside (FmTR), in Two Types of Cultured Vessels: Closed Container (CC) and Filtered Container (FC).
Figure 1.
Absolute cytokinin content, and Isoprenoid and Aromatic Cytokinin Ratio (%) in Axillary Shoots of Phalaenopsis Grown on Media Supplemented with Different Types of Cytokinins: BA (N6-Benzyladenine), 6-(3-fluorobenzylamino)purine (FmT), and 6-(3-fluorobenzylamino)purine 9-riboside (FmTR), in Two Types of Cultured Vessels: Closed Container (CC) and Filtered Container (FC).
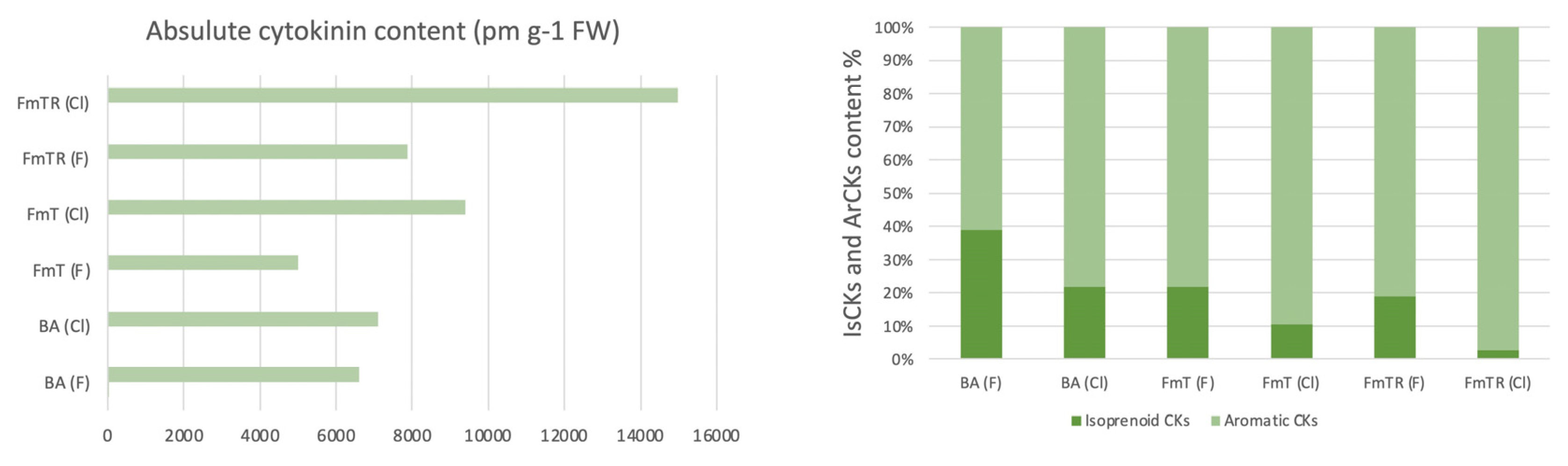
Figure 2.
The chart displays the relative distribution of various cytokinin metabolites, categorized as free bases (cZ, IP, BAP, mT, oT, pT, FmT), ribosides (FmTR, pTR, oTR, mTR, BAR, iPR, cZR, DHZR), O-glucosides ribosides and N-glucosides (DHZROG, tZ7G, DHZ7G, cZ9G BAP9G, mT9G), among different treatments and container types.
Figure 2.
The chart displays the relative distribution of various cytokinin metabolites, categorized as free bases (cZ, IP, BAP, mT, oT, pT, FmT), ribosides (FmTR, pTR, oTR, mTR, BAR, iPR, cZR, DHZR), O-glucosides ribosides and N-glucosides (DHZROG, tZ7G, DHZ7G, cZ9G BAP9G, mT9G), among different treatments and container types.
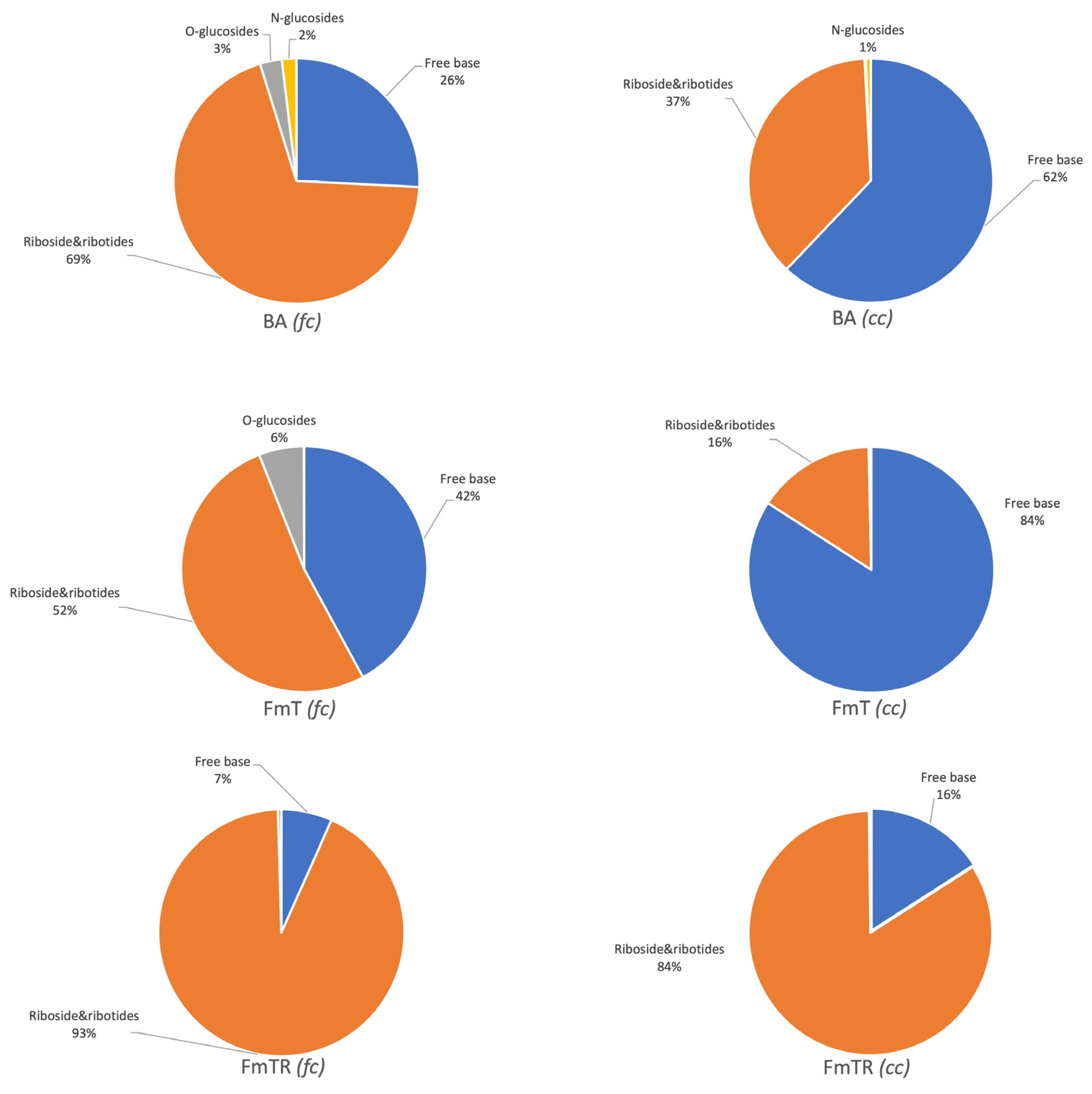

Table 1.
Concentrations of cytokinins (pmol/g FW) and its metabolites in Phalaenopsis in vitro culture grown in filtered and closed Containers.
Table 1.
Concentrations of cytokinins (pmol/g FW) and its metabolites in Phalaenopsis in vitro culture grown in filtered and closed Containers.
| |
Cytokinin treatment | |||||
| BA | FmT | FmTR | ||||
| Cytokinin metabolites |
Container type | |||||
| Filtered | Closed | Filtered | Closed | Filtered | Closed | |
| tZ | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| tZOG | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| tZR | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| tZROG | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| tZ7G | 20.2 ± 2.07 | 4.9±0.2 | 1.6 ± 0.05 | <LOD | <LOD | <LOD |
| tZ9G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| tZR5´MP | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| cZ | 20.8 ± 2.5 | 11.09 ± 1.4 | <LOD | <LOD | <LOD | <LOD |
| cZOG | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| cZR | 2059.5 ± 96.3 | 1296.1 ± 9.8 | 743.5 ± 56.5 | 893.7 ± 77.0 | 1362.8 ± 129.3 | 270.5 ± 21.8 |
| cZROG | 26.1 ± 1.7 | 11.3 ± 0.5 | <LOD | 3.85 ± 0.2 | <LOD | 4.1 ± 0.1 |
| cZ7G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| cZ9G | <LOD | <LOD | <LOD | 8.6 ± 0.2 | <LOD | 6.1 ± 0.4 |
| cZR5´MP | <LOD | 74.6 ± 5.3 | <LOD | 40.7 ± 3.6 | <LOD | 66.1 ± 3.9 |
| DHZ | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| DHZOG | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| DHZR | 5.7 ± 0.8 | 3.8 ± 0.3 | 3.4 ± 0.06 | 1.5 ± 0.2 | <LOD | <LOD |
| DHZROG | 164.1 ± 4.8 | <LOD | 296.6 ± 20.1 | <LOD | 34.9 ± 1.1 | <LOD |
| DHZ7G | <LOD | 3.3 ± 0.07 | <LOD | 2.4 ± 0.2 | <LOD | 3.8 ± 0.1 |
| DHZ9G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| DHZR5´MP | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| iP | 8.6 ± 0.1 | 5.2 ± 0.1 | 1.8 ± 0.1 | 3.1 ± 0.2 | 3.1 ± 0.3 | 2.5 ± 0.2 |
| iPR | 272.9 ± 4.7 | 125.8 ± 5.3 | 53.2 ± 1.1 | 22.1 ± 0.4 | 87.07 ± 3.8 | 23.8 ±1.2 |
| iP7G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| iP9G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| iPR5´MP | <LOD | 21.5 ± 2.03 | <LOD | 14.4 ± 1.3 | <LOD | 20.01 ± 0.8 |
| BA | 1398.3 ± 151.1 | 4159.68 ± 497.2 | 9.03 ± 0.6 | 15.4 ± 3.1 | <LOD | <LOD |
| BAR | 84.04 ± 2.9 | 61.4 ± 1.3 | 0.5 ± 0.09 | <LOD | 1.1 ± 0.08 | 3.1 ± 0.3 |
| BA7G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| BA9G | 106.5 ± 3.4 | 37.3 ± 1.3 | <LOD | <LOD | <LOD | <LOD |
| BAR5´MP | <LOD | 19.1 ± 0.4 | <LOD | <LOD | <LOD | <LOD |
| mT | 30.8 ± 1.9 | 16.1 ± 0.3 | <LOD | 9.1 ± 1,0 | <LOD | 19.4 ± 1.3 |
| mTR | 67.2 ± 6.3 | 17.5 ± 0.9 | 27.1 ± 1.3 | 7.8 ± 0. 2 | 32.2 ± 2.0 | 26.6 ± 1.8 |
| mT7G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| mT9G | <LOD | <LOD | <LOD | 16.8 ± 0,1 | <LOD | 15.1 ± 1.3 |
| oT | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| oTR | 17.1 ± 1.5 | 4.7 ± 0.4 | <LOD | <LOD | <LOD | <LOD |
| oT7G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| oT9G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| pT | 147.6 ± 9.4 | 27.8 ± 2.3 | 140.7 ± 10.6 | <LOD | 196.5 ± 6.1 | <LOD |
| pTR | 1242.2 ± 27.8 | 251.9 ± 17.8 | 1142.1 ± 24.9 | 21.6 ± 0.8 | 1346.1 ± 74.4 | 25.8 ±1.3 |
| pT7G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| pT9G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| K | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| KR | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| K9G | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| FmT | 99.2 ± 11.2 | 140.8 ± 11.4 | 1952.3 ± 220.9 | 7836.4 ± 713.3 | 326.1 ± 25.4 | 2350.7 ± 360.5 |
| FmTR | 844.4 ± 107.2 | 804.2 ± 88.6 | 627.4 ± 44.5 | 490.7 ± 41.03 | 4478.6 ± 150.7 | 12156.5 ± 1712.8 |
| Values are presented as mean ± SD; LOD = below detection limit | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



