Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
The type of fertilization used influences plant compounds and pest infestation. We measured, through chemical stimuli, the preference of Melanaphis sacchari for sorghum plants fertilized by means of conventional fertilization (CF) or organic fertilization (OF). Leaves were collected from sorghum plants fertilized with 200 kg N ha-1 using ammonium sulfate and poultry manure. Extracts were obtained using Soxhlet extraction, and the compounds were identified using a gas chromatograph coupled with mass spectrometry (GC/MS). Sorghum extracts were individually tested through bioassays to determine M. sacchari preference. The abundance and number of compounds in sorghum differed depending on the type of fertilization used. M. sacchari showed a preference for the extract from CF sorghum plants (76.66%) over the extract from OF plants (23.34%). Therefore, the type of fertilization can be used as a tactic to prevent higher infestations of M. sacchari. The biological activity of the compounds identified here with M. sacchari should be determined for future pest management strategies using allelochemicals, given that the sugarcane aphid uses chemical signals to locate its host plant.
Keywords:
Alellochemicals
; bioassays
; chemical stimuli
; fertilization
; plant extracts
; sorghum
; yellow aphid
1. Introduction
Sorghum bicolor (L.) Moench, commonly called sorghum, belongs to the family Poaceae. Mexico contributes 10.6% of the world’s production and imports 5.01 million tons of this grain utilized mainly as feed for cattle, pigs, and poultry [1]. The sugarcane aphid (Melanaphis sacchari) poses a persistent challenge in sorghum cultivation; it is distributed globally and, specifically in Mexico, is found in 27 states [2]. This insect negatively affects sorghum and other important crops. Sugarcane aphids are economically important; they feed on plant sap, leading to reduced yields and, in some cases, total sorghum losses [2,3]. Pest management strategies proposed for M. sacchari include biological control, determination of optimal planting dates, elimination of alternative hosts, and the use of pesticides, with the latter being the primary control measure for this insect [4]. Pesticides can potentially enter the environment, contaminating both the environment and food, thereby affecting biodiversity and human health [5]. Therefore, it is crucial to implement new strategies, such as utilizing insect behavior to compounds emitted by plants or also called allelochemicals. [6]. These compounds serve as chemical signals for insects to identify their host plants, yet their potential benefits in M. sacchari management remain poorly explored [7]. Nitrogen (N) fertilization, mainly synthetic or conventional N, is essential for obtaining high crop yields. However, this practice causes contamination, similar to the use of pesticides, and has been associated with a higher incidence of pest and aphid species, such as Brachycaudus cardui, which has shown increased attraction to fertilized host plants [8,9]. In addition, there is evidence that insects have a lower preference for organically fertilized (OF) plants. Organic fertilization has recently gained importance because it shows less nutrient loss and is more environmentally friendly than conventional fertilization (CF) [10]. In line with the above, it has been reported that the type of fertilization used affects plant chemical compounds [11,12].
The aim of the present study was to determine whether the type of fertilization affects the compounds found in sorghum plants and the preference of the sugarcane aphid. To do this, compounds from S. bicolor were identified and bioassays were conducted to assess the behavior of M. sacchari in response to chemical stimuli, which could serve as another important factor in the management of this pest.
2. Materials and Methods
2.1. Plant Material
S. bicolor seeds were sown in 4 L polyethylene bags in a greenhouse of the Colegio de Postgraduados (COLPOS), Campus Puebla (19°04′26.5´´N; 98°15′41.3´´W). Plants were grown for 60 days at 20-25 °C and 60 ± 10% humidity, and watered every 3 days. The experiment followed a completely randomized design consisting of 3 treatments, with 3 replicates (Table 1).
2.2. Insect Breeding
Sugarcane aphid nymphs were obtained from sorghum plots in the municipality of Izúcar de Matamoros, Puebla (18°36′10´´N; 98°27′5´´W). The nymphs were transported to the greenhouse and maintained on healthy sorghum plants (under the above-mentioned conditions) for reproduction until their use in bioassays using extracts.
2.3. Extraction and Identification of Compounds
The third and fourth alternate leaves of sorghum plants (300 g) were ground using a mill, wrapped in filter paper, and then placed in a Soxhlet extractor mounted atop a distillation flask containing 150 mL of 90% ethanol (Sigma Aldrich). The extraction process was carried out for 3 h. Then, 1 µL of the extract was injected into an Agilent Technologies 7890A gas chromatograph coupled with an Agilent Technologies 5975C mass spectrometer (Santa Clara, CA, USA). The system was equipped with a 30 m x 0.25 mm HP-5MS column with a film thickness of 0.50 µm (Agilent J&W, Santa Clara, CA, USA). The GC-MS parameters were as follows: helium as the carrier gas, injector temperature of 250 °C in splitless mode, initial oven temperature of 36 °C for 1 min and then increased by 10 °C per min until reaching 250 °C, which was maintained for 3 min. Compounds were identified through comparison with mass spectra from the National Institute of Standards and Technology library (NIST 8 and NIST 11).
2.4. Bioassays Using Extracts from S. bicolor
Healthy-looking 1.2 mm-long, 13-day-old females of M. sacchari were fasted for 1 h prior to individual bioassays. A glass Petri dish (10 cm in diameter) was used as the behavioral arena (Figure 1). Bioassays were carried out under laboratory conditions (22 ± 3 °C and 60 ± 10% RH). Briefly, 10 µL of extract were placed on 1 cm x 1 cm filter paper pieces (Whatman No. 1) and allowed to evaporate for 30 s. For the control (C), an equivalent volume of solvent (ethanol) was used, following the same procedure. Each piece of filter paper was placed end to end of the Petri dish randomly in each bioassay. A 5 min response time was given, or until it moved towards the extract (past the response line). A total of 30 individual bioassays were conducted. After 3 bioassays, the Petri dish used was replaced by a clean one. The extracts tested were C, F0 (unfertilized plant), CF, and OF. Six combinations of extracts were made, as described in Table 2.
2.5. Statistical Analysis
The frequencies of behavioral responses were analyzed through an exact binomial test using the R Studio software.
3. Results
3.1. Extraction and Identification of Compounds
The number of compounds identified in sorghum leaf extracts were 12 in F0, 34 in CF, and 16 in OF. Butanedioic acid, 1-tetradecene, (Z)-7-hexadecene, and phenol were found in leaves from all three treatments. Butanedioic acid and 1-tetradecene were most abundant in F0, followed by CF and OF. In contrast, phenol was most abundant in CF (23.35%), followed by F0 (22.64%) and OF (14.01%). The abundances of (E)-5-octadecene and benzophenone were higher in CF than in OF. On the other hand, 1,2-benzenedicarboxylic acid was more abundant in OF (1.62%) than in CF (0.77%). The most abundant compound in F0 and OF was (4-methoxy-phenyl)-(5-p-tolyl-furan-2-ylmethylene)-amine (46.89% and 68.02%, respectively). In CF, 5-[[[3,4,5-trimethoxyphenyl]imino]methyl]-2,4-pyrimidinediamine was the most abundant compound (37.73%). The least abundant compounds were (2,2-dichlorocyclopropyl) methanol in F0 (0.06%), 5,6-dihydro-2-(4-nitrophenyl)-4H-1,3-oxazin-5-one in CF (0.11%), and 3-methyl-1-(4-toluidino)pyrido [1,2 -a]benzimidazole-4-carbonitrile in OF (0.2%). A total of 28 compounds were found exclusively in CF sorghum plants.
3.2. Bioassays Using Extracts from S. bicolor
M. sacchari females showed a preference for S. bicolor extracts compared to the control (C), with higher response in F0 (93.34%), CF (86.66%), and OF (76.66%). However, bioassays comparing F0 (20%) with CF (80%) and F0 (26.67%) with OF (73.33%), exhibited preference for extracts from fertilized S. bicolor (CF and OF). Finally, when comparing CF (76.66%) with OF (23.34%) extracts, CF was preferred (Figure 2).
4. Discussion
The type of fertilization affected the abundance and number of compounds found in sorghum plants. This is in agreement with observations in Rubus idaeus plants, where the type of fertilization had both qualitative and quantitative effects on the compounds emitted [13]. In other plants such as willow, strawberry, and tomato, the compounds were affected by CF with N [12,13,14]. In addition, OF has been shown to modify the abundance of compounds compared to unfertilized tomato plants [14]. In line with previous reports for other species, we found that the number of compounds in S. bicolor was higher in CF, followed by OF and F0. These results are attributed to the availability of N, which leads to increased protein synthesis and, therefore, increased synthesis of secondary metabolites that affect plant defense [15]. CF has been associated with a higher abundance of compounds [12,16], as evinced by the results here obtained for (E)-5-octadecene, benzophenone, and phenol. However, said trend was not observed when comparing F0 with CF. Butanedioic acid, 1-tetradecene, and (Z)-7-hexadecene were more abundant in F0. In this regard, it has been reported that compounds exhibit individual responses to fertilization, which could be attributed to different biosynthetic pathways and environmental factors [12]. Eleven of the compounds exclusively identified in CF were mostly aromatic compounds, which act as chemical signals involved in the attraction of insects, such as Drosophila melanogaster, Eupeodes corolla, and Sitona humeralis [17,18,19]. These compounds, which were not common to other treatments, could be a key factor in the attraction of M. sacchari to extracts from CF sorghum plants.
Some of the compounds identified in this study have been reported to have biological activity in certain insects, including some found exclusively in CF plants, such as acetic acid. Acetic acid has been related to high-dose N fertilization in Brassica napus plants and serves as a potential attractant of Meligethes aeneus and D. melanogaster [11,17]. Butyric acid is a key attractant of pests such as Holotrichia paralela and Bubas bison [20,21]. A compound induced by herbivory, 2-octanol, has been reported as an attractant of Spilosoma obliqua [22]. In Camellia sinensis plants, 1,3-benzenedicarboxylic acid has been related to N fertilization doses and infestation of the aphid Toxoptera aurantii [23]. The most abundant compound in CF was phenol, which acts as an attractant of beetles of the species B. bison [21].
In OF, we found butanedioic acid, a component of the silkworm cocoon, which may be involved in its protection [24]. Likewise, higher levels of butanedioic acid were found in chickpea plants and were linked to increased resistance to the leaf miner Liriomyza cicerina [25]. Another compound that could have repellent activity against M. sacchari is 1,2-benzenedicarboxylic acid, which has potential as a natural insecticide [26]. In Capsicum spp. plants infested with Aphis gossypii aphids, 1-undecanol has been identified as a compound involved in plant’s defense [27]. In the present study, 1-undecanol was exclusively found in F0 and OF sorghum plants, suggesting its potential role as a repellent against M. sacchari. Finally, 1-tetradecene exhibits repellent activity against the aphids Acyrthosiphon pisum and Myzus persicae [28], while showing attractant activity towards Apolygus lucorum, Adelphocoris suturalis, and Megalurothrips sjostedti [29,30]. Although previous reports on the compounds identified in this study may provide an indication of their activity against or towards M. sacchari, further research is needed to confirm their biological activity.
Based on our results, M. sacchari has a preference for CF and OF treatments. In this regard, it has already been reported that there is a preferential relationship of M. sacchari for plants fertilized with higher levels of N [4]. Here, we compared the preference for CF or OF plants. The release of nutrients in organic fertilization is slower compared to chemical fertilizers. This differential rate impacts sap-sucking insects, as leaf sap composition is affected by fertilizer sources and dosages [31,32]. To date, only M. persicae has been tested using chemical stimuli. Olfactometer tests revealed that this species was more attracted to volatile compounds from cabbage plants with higher N doses [33]. A higher incidence and preference for fertilized plants has also been observed in other aphids, such as M. persicae, which preferred capsicum plants with higher N doses [34]. Similarly, the aphids T. aurantii, Lipaphis erysimi, Bemisia tabaci, Rhopalosiphum padi, and Sitobion avenae were more attracted to host plants fertilized with high N doses compared to unfertilized plants. This preference affected their fecundity and longevity [23,35,36,37].
Insect preference for fertilized plants is attributed to physiological changes in the plants, which cause changes in their metabolites and, consequently, in the chemical signals they emit [37]. In addition, M. persicae showed a preference for white clover CF plants over plants fertilized with poultry manure [38]. It has also been reported that maize plants fertilized with synthetic fertilizers had a higher percentage of infestation by R. maidis aphid than plants fertilized with animal manure [39]. In this regard, chemical nitrogen fertilizers enhance the vigorous qualities of plants, making them attractive to insects [40,41]. On the other hand, organic fertilization, characterized by the slow release of nutrients and consequently a lower N dose, may affect the production of toxic compounds, helping to maintain pest populations at low levels and to enhance plant resistance [42,43,44].
There are no reports indicating the existence of a chemical signal involved in the interaction between M. sacchari and S. bicolor. Moreover, it has been claimed that this insect relies primarily on visual signals. However, aphids have large antennae, suggesting that olfactory signals must be important factors in locating host plants [45,46]. Our findings confirm the above, as chemical signals were used to evaluate the preference of M. sacchari for S. bicolor extracts.
5. Conclusions
The type of fertilization used affected the compounds extracted from S. bicolor plants, which is related to the attractant activity shown by the extracts from fertilized sorghum plants. The sugarcane aphid showed a preference for extracts from CF plants, confirming that M. sacchari uses chemical stimuli to locate its host plant, and that CF makes sorghum plants susceptible to attack by this aphid. Therefore, OF represents a key method for pest prevention.
The type of fertilization and its relationship with the chemical compounds of sorghum plants should be taken into account when devising management strategies for M. sacchari. It is evident that sugarcane aphids rely on allelochemicals to locate the sorghum plant. We recommend further studies to determine the biological activity of the compounds identified here against M. sacchari.
Author Contributions
Conceptualization, E.N.S. and E.S.C.; methodology, E.N.S. and M.D.C.A.; software, E.N.S.; validation, E.S.C., M.D.C.A., A.D.A., A.H.d.l.P. and I.O.F.; formal analysis, E.N.S.; investigation, E.N.S.; resources, E.N.S. and M.D.C.A; data curation, E.N.S.; writing—original draft preparation, E.N.S.; writing—review and editing E.N.S., M.D.C.A., A.D.A., A.H.d.l.P. and I.O.F.; visualization, E.N.S. and E.S.C.; supervision, E.N.S. and M.D.C.A.; project administration, E.N.S., E.S.C. and M.D.C.A.; funding acquisition, E.N.S., M.D.C.A. and E.S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study is a product of the doctoral dissertation of the first author, who is grateful to the Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCyT) for grant number 814396 in support of her doctoral studies.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Peña-Martínez, R.; Brujanos-Muñiz, R.; Muñoz-Viveros, A.; Vanegas-Rico, J.; Salas, R.; Hernández-Torres, O.; Marín-Jarillo, A.; Ibarra, J.; Lomeli-Flores, R. Pulgón amarillo del sorgo, (PAS), Melanaphis sacchari (Zehntner, 1897), interrogantes biológicas y tablas de vida, 1st ed.; Fundación Produce Guanajuato A. C., Celaya Gto., México, 2018; pp 1-47.
- Garay, I.; Díaz, A.; Herrera, J.; Rubio, A.; Vázquez, C. Evaluación de cuatro híbridos de sorgo (Sorghum bicolor L), abonados con vermicompost en Úrsulo Galván, Ver. Braz. J. Anim. Environ. Res. 2021, 4, 1910–1919. [Google Scholar] [CrossRef]
- Xoconostle-Cázares, B.; Ramírez-Pool, J.A.; Núñes-Muñoz, L.A.; Calderón-Pérez, B.; Vargas-Hernández, B.Y.; Bujanos-Muñiz, R.; Ruiz-Medrano, R. The Characterization of Melanaphis sacchari Microbiota and antibiotic treatment effect on insects. Insects 2023, 14, 807. [Google Scholar] [CrossRef] [PubMed]
- Schlickmann-Tank, J.; Morales-Galván, O.; Pineda-Pineda, J.; Espinosa-Vásquez, G.; Colinas-León, M.; Vargas-Hernández, M. Relationship between chemical fertilization in sorghum and Melanaphis sacchari/sorghi (Hemiptera: Aphididae) populations. Agronomía Colombiana 2020, 38, 357–366. [Google Scholar] [CrossRef]
- Tudi, M.; Ruan, H.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D. Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 2021, 18, 1–23. [Google Scholar]
- Arora, S.; Husain, T.; Prasad, S. Allelochemicals as biocontrol agents: Promising aspects, challenges and oppotunities. S. Afr. J. Bot. 2024, 166, 503–511. [Google Scholar] [CrossRef]
- Caballero-Vidal, G.; Bouysset, C.; Gévar, J.; Mbouzid, H.; Nara, C.; Delaroche, J.; Golebiowski, J.; Montagné, N.; Fiorucci, S.; Jacquin-Joly, E. Reverse chemical ecology in a moth: machine learning on odorant receptors identifies new behaviorally active agonists. Cell. Mol. Life Sci. 2021, 78, 6593–6603. [Google Scholar] [CrossRef]
- Yang, Q.; Ma, J.; Yang, F.; Zheng, H.; Lu, Z.; Qiao, F.; Zhang, K.; Gong, H.; Men, X.; Li, J.; Ouyang, F.; Ge, F. The hidden indirect environmental effect undercuts the contribution of crop nitrogen fertilizer application to the net ecosystem economic benefit. J. Clean. Prod. 2023, 426, 139204. [Google Scholar] [CrossRef]
- Lescano, M.; Quintero, C.; Farji, A.; Balseiro, E. Excessive nutrient input an ecological cost for aphids by modifying their attractiveness toward mutualist ants. Funct. Ecol. 2022, 36, 2661–2672. [Google Scholar] [CrossRef]
- Fabroni, S.; Bontempo, L; Campanelli, G. ; Canali, S.; Montmurro, F. Innovative tools for the nitrogen fertilization traceability of organic farming products. Horticulturae 2023, 9, 723. [Google Scholar] [CrossRef]
- Veromann, E.; Toome, M.; Kännaste, A.; Kaasik, R.; Copolovici, L.; Flink, J.; Kovács, G.; Narits, L.; Luik, A.; Niinemets, Ü. Effects of nitrogen fertilization on insect pests, their parasitoids, plant diseases and volatiles organic compounds in Brassica napus. Crop Prot. 2013, 43, 79–88. [Google Scholar] [CrossRef]
- Hu, B.; Jarosch, A.; Gauder, M.; Graeff-Hönninger, S.; Schnitzler, J.; Grote, R.; Rennenberg, H.; Kreuzwieser, J. VOC emissions and carbon balance of two bioenergy plantations in response to nitrogen fertilization: A comparison of Miscanthus and Salix. Environ. Pollut. 2018, 237, 205–217. [Google Scholar] [CrossRef]
- Estrada, A.; Salas, N.; Parra, R.; González, A.; Soto, M.; Rodriguez, M.; Flores, M.; Chavez, A. Effect of conventional and organic fertilizers on volatile compounds of raspberry fruit. Not. bot. Horti Agrobot. Cluj-Napoca 2020, 48, 862–870. [Google Scholar] [CrossRef]
- Paskovic, I.; Soldo, B.; Ban, S.; Rdic, T.; Lukic, M.; Urlic, B.; Mimica, M.; Bubola, K.; Colla, G.; Rouphael, Y.; Major, N.; Simpraga, M.; Ban, D.; Palcic, I.; Franic, M.; Grozic, K.; Lukic, I. Fruit quality and volatile compound composition of processing tomato as affected by fertilisation practices and arbuscular mycorrhizal fungi application. Food Chem. 2021, 359, 129961. [Google Scholar] [CrossRef]
- Ibrahim, M.; Jaafar, H.; Rahmat, A.; Rahman, Z. Effects of nitrogen fertilization on synthesis of primary and secondary metabolites in three varieties of Kacip Fatimah (Labisia Pumila Blume). Int. J. Mol. Sci. 2011, 12, 5238–5254. [Google Scholar] [CrossRef]
- Campos, F.; Ribeiro, M.; Esteves, A.; delaCruz-Chacón, I.; Mayo, M.; Ferreira, G.; Fernandes, B. Nitrogen in the defense system of Annona emarginata (Schltdl.) H. Rainer. PLos ONE, 2019, 14, e0217930. [Google Scholar] [CrossRef]
- Becher, P.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpedowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piskur, J. Chemical signaling and insect attraction is a conserved trait in yeast. Ecol. Evol. 2018, 8, 2962–2974. [Google Scholar] [CrossRef]
- Li, H.; Liu, W.; Yang, L.; Cao, H.; Pelosi, P.; Wang, G.; Wang. B. Aromatic volatiles and odorant receptor 25 mediate attraction of Eupeodes corollae to flowers. J. Agric. Food Chem. 2020, 68, 12212–12220. [Google Scholar] [CrossRef]
- Lohonyai, Z.; Vuts, J.; Kárpáti, Z.; Koczor, S.; Domingue, M.; Fail, J.; Birkett, M.; Tóth, M.; Imrei, Z. Benzaldehyde: an alfalfa related compound for the spring attraction os pest weevil Sitona humeralis (Coleoptera: Curculionidae). Pest. Manag. Sci. 2019, 75, 3153–3159. [Google Scholar] [CrossRef]
- Nyamwasa, I.; Li, K.; Zhang, S.; Yin, J.; Li, X.; Liu, J.; Li, E.; Sun, X. Overlooked side effects of organic farming inputs attract soil insect crop pests. Ecol. Appl. 2020, 30, e02084. [Google Scholar] [CrossRef]
- Perera, N.; Weston, P.; Barrow, R.; Weston, L.; Gurr, G. Contrasting volatilomes of livestock dung drive preference of the dung beetle Bubas bison (Coleoptera: Scarabaeidae). Molecules 2022, 27, 4152. [Google Scholar] [CrossRef]
- Mobarak, S.; Koner, A.; Debnath, R.; Barik, A. The role of gram plant volatile blends in the behavior of arctiid moth, Spilosoma obliqua. J. Chem. Ecol. 2022, 48, 802–816. [Google Scholar] [CrossRef]
- Pokharel, S.; Zhong, Y.; Changning, L.; Shen, F.; Likun, L.; Parajulee, M.; Fang, W.; Chen, F. Influence of reduced N-fertilizer application on foliar chemicals and functional qualities of tea plants under Toxoptera aurantii infestation. BMC Plant Biol. 2022, 22, 166. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, D.; Meng, Z.; Dong, Z.; Lin, Y.; Chen, S.; Xia, Q.; Zhao, P. Wild silkworm cocoon contains more metabolites than domestic silkworm cocoon to improve its protection. J. Insect Sci. 2017, 17, 105. [Google Scholar] [CrossRef]
- Chrigui, N.; Sari, D.; Sari, H.; Eker, T.; Cengiz, M.; Ikten, C.; Toker, C. Introgression of resistance to Leafminer (Liriomyza cicerina Rondani) from Cicer reticulatum Ladiz. to C. arietinum L. and relationships between potential biochemical selection criteria. Agronomy 2021, 11, 57. [Google Scholar] [CrossRef]
- Chidambaram, K.; Alqahtani, T.; Alghazwani, Y.; Aldahish, A.; Annadurai, S.; Venkatesan, K.; Dhandapani, K.; Thilagam, E.; Venkatesan, K.; Paulsamy, P.; Vasudevan, R.; G. Kandasamy. Medicinal plants of Solanum species: the promising sources of phyto-insecticidal compounds. J. Trop. Med. 2022, 2022, 1–22. [Google Scholar]
- Gomes, J.; Vieira, E.; Riffel, A.; Birkett, M.; Bleicher, E.; Goulart, A. Differential preference of Capsicum spp. Cultivars by Aphis gossypii is conferren by variation in volatile semiochemistry. Euphytica 2010, 177, 299–307. [Google Scholar] [CrossRef]
- Dancewicz, K.; Kordan, B.; Gabrys, B.; Szumny, A.; Wawrzenczyk, C. Feeding deterrent activity of α-methylenelactones to pea aphid Acyrthosiphon pisum (Harris) and green peach aphid Myzus persicae (Sulzer). Pol. J. Nat. Sci. 2006, 20, 23–31. [Google Scholar]
- Yin, H.; Li, W.; Xu, M.; Xu, D.; Wan, P. The role of tetradecane in the identification of host plants by the mirid bugs Apolygus lucorum and Adelphocoris suturalis and potential application in pest management. Front. physiol. 2022, 13, 1061817. [Google Scholar]
- Alabi, O.; Onochie, I. Repellence and attraction properties of selected synthesized volatile organic compounds to flower bud thrips (Megalurothrips sjostedti Trybom). Adeleke University J. Eng. Technol. 2023, 6, 142–150. [Google Scholar]
- Barrios, M.; García, J.; Basso, C. Efecto de la fertilización nitrogenada sobre el contenido de nitrato y amonio en el suelo y la planta de maíz. Bioagro 2012, 24, 213–220. [Google Scholar]
- Hsu, C; Lai, H. Comprehensive assessment of the influence of applying two kinds of chicken-manure-proccessed organic fertilizers on soil properties, mineralization of nitrogen, and yields of three crops. Agronomy 2022, 12, 2355. [Google Scholar] [CrossRef]
- Ahmed, N.; Darshanee, H.; Khan, I.; Zhang, Z.; Liu, T. Host selection behavior of the green peach aphid, Myzus persicae, in response to volatile organic compounds and nitrogen contents of cabbage cultivars. Front. Plant Sci, 2019, 10, 79. [Google Scholar] [CrossRef]
- Kamnqa, U.; Etsassala, N.; Akinpelu, E.; Nchu, F. Effects of varying nitrogen fertilization on growth, yield and flowering of Capsicum annuum (California wonder). 18th SOUTH AFRICA Int’l Conference on Agricultural, Chemical, Biological and Environmental Sciences (ACBES-20), Johannesburg, 16-17 nov, 2020. [CrossRef]
- Aqueel, M.; Leather, S. Effect of nitrogen fertilizer on the growth and survival of Rhopalosiphum padi (L.) and Sitobion avenae (F.) (Homoptera: Aphididae) on different wheat cultivars. Crop Protec. 2011, 30, 216–221. [Google Scholar] [CrossRef]
- Zhao, Z.; Hui, C.; He, D.; Li, B. Effects of agricultural intensification on ability of natural enemies to control aphids. Sci. Rep. 2015, 5, 8024. [Google Scholar] [CrossRef]
- Fallahpour, F.; Ghorbani, R.; Nassiri, M.; Hosseini, M. Plant fertilization helps plants to compensate for aphid damage, positively affects predator efficiency and improves canola yield. J. Pest Sci. 2019, 93, 251–260. [Google Scholar] [CrossRef]
- Staley, J.; Stewar, A.; Pope, T.; Wright, D.; Leather, S.; Hadley, P.; Rossiter, J.; Emden, H.; Poppy, G. Varying responses of insect herbivores to altered plant chemistry under organic and conventional treatments. Proc. R. Soc. 2010, 277, 779–786. [Google Scholar] [CrossRef]
- Naroz, M.; Mahmoud, H.; El-Rahman, S. Influence of fertilization and plant density on population of some maize insects pests yield. J. Plant Prot. Pathol. 2021, 12, 403–411. [Google Scholar] [CrossRef]
- Figueroa, R.; Villa, P.; López, J.; Huerta, A.; Pacheco, J.; Ramos, M. Nitrogen fertilization sources and insecticidal activity of aqueous seeds extract of Carica papaya against Spodoptera frugiperda in maize. Cienc. Invest. Agrar. 2013, 40, 567–577. [Google Scholar] [CrossRef]
- Blundell, R.; Schmidt, J.; Igwe, A.; Cheung, A.; Vannette, R.; Gaudin, C.; Casteel, C. Organic mangement promotes natural pest control through altered plant resistance to insects. Nat. Plants 2020, 6, 483–489. [Google Scholar] [CrossRef]
- Rowen, E.; Tooker, J.; Blubaugh, C. Managing fertility with animal waste to promote arthropod pest suppression. Biol. Control 2019, 134, 130–140. [Google Scholar] [CrossRef]
- Parizad, S.; Bera, S. The effect of organic farming on water reusability, sustainable ecosystem, and food toxicity. Environ. Sci. Pollut. Res. 2021, 30, 1–12. [Google Scholar] [CrossRef]
- Gao, H.; Xu, R.; Du, Z.; Ye, H.; Tian, J.; Huang, W.; Xu, S.; Xu, F.; Hou, M.; Zhong, F. Molecular regulation of volatile organic compounds accumulation in tomato leaf by different nitrogen treatments. Plant Growth Regul. 2023. [Google Scholar] [CrossRef]
- Holopainen, J. Importance of olfactory and visual signals of autumn leaves in the coevolution of aphids and trees. Bioessays 2008, 30, 889–896. [Google Scholar] [CrossRef]
- Guerrieri, E.; Digilio, M. Aphid-plant interactions: a review. J. Plant Interact. 2012, 3, 223–232. [Google Scholar] [CrossRef]
Figure 1.
Behavioral arena for bioassays with sorghum extracts. RL: Response line; E: Extract.
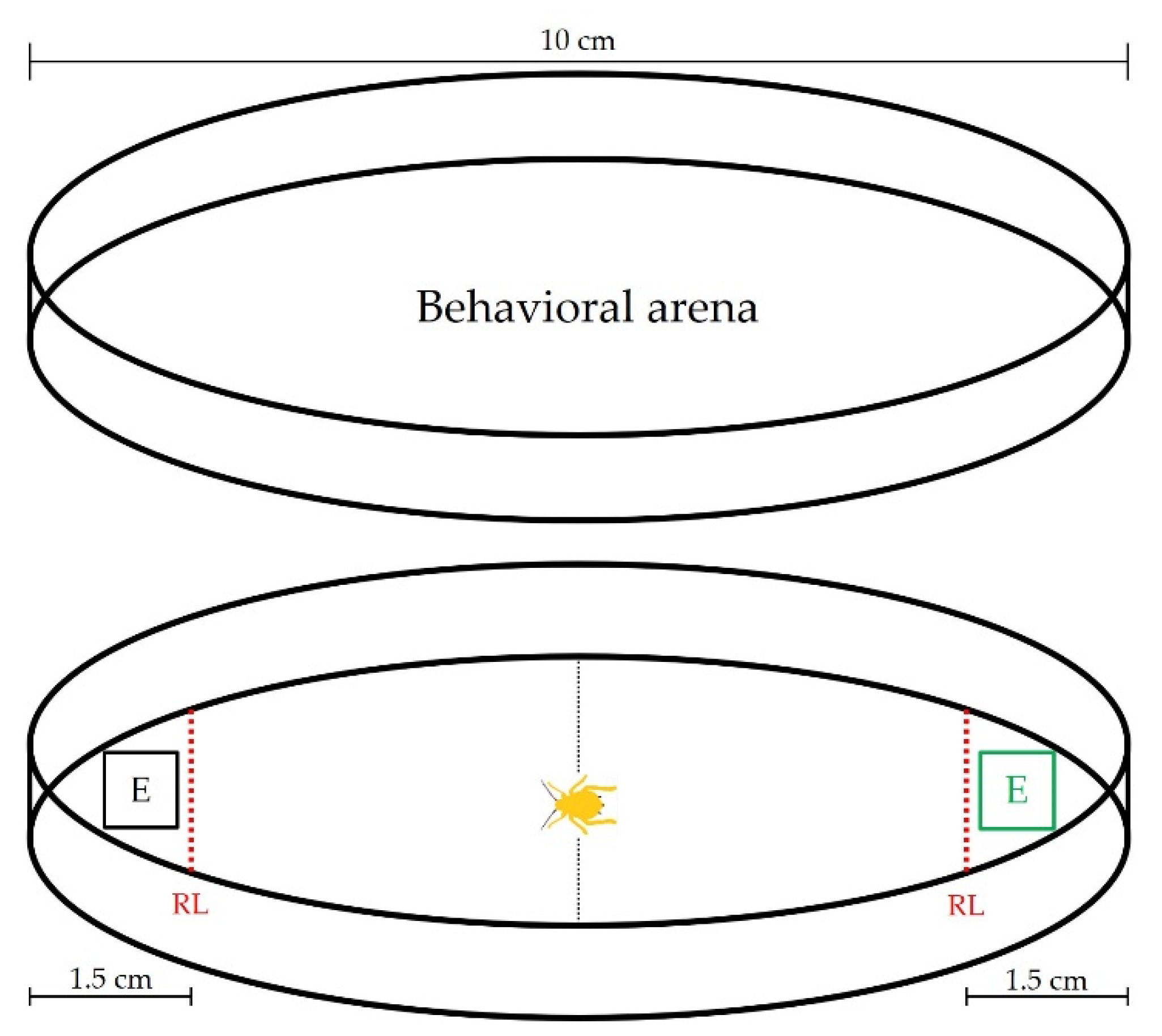
Figure 2.
Behavioral bioassays of M. sacchari in response to S. bicolor extracts. C (control), F0 (unfertilized plant), CF (plant fertilized with ammonium sulfate), and OF (plant fertilized with poultry manure). * denotes significant differences (Binomial test: p < 0.05); ** denotes significant differences (Binomial test: p < 0.0001).
Figure 2.
Behavioral bioassays of M. sacchari in response to S. bicolor extracts. C (control), F0 (unfertilized plant), CF (plant fertilized with ammonium sulfate), and OF (plant fertilized with poultry manure). * denotes significant differences (Binomial test: p < 0.05); ** denotes significant differences (Binomial test: p < 0.0001).
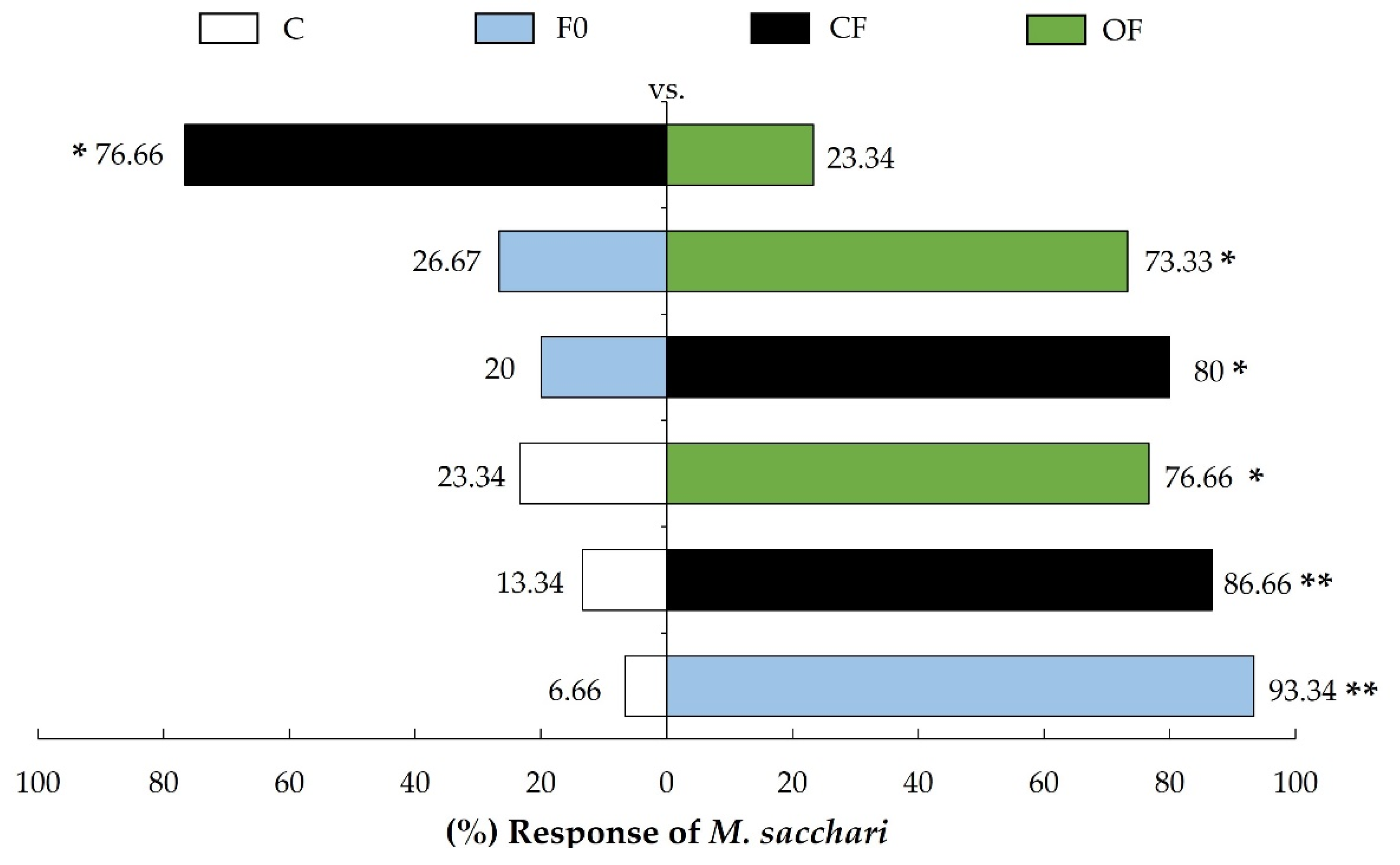

Table 1.
Fertilization treatments in S. bicolor.
| Treatments | Description |
|---|---|
| F0 | Soil (4.4% organic material; nitrogen 100 ppm; phosphorus 0.80 ppm; potassium 5.50 ppm and pH 7.4) |
| CF OF |
200 kg ha-1 N (ammonium sulfate) + soil 200 kg ha-1 N (poultry manure) + soil |
F0: Zero fertilization; CF: Conventional fertilization; OF: Organic fertilization.
Table 2.
Combinations for behavioral bioassays.
| Bioassay | Combinations extracts |
|---|---|
| 1 | CF vs. OF |
| 2 | F0 vs. OF |
| 3 | F0 vs. CF |
| 4 | C vs. OF |
| 5 | C vs. CF |
| 6 | C vs. F0 |
C: Control (etanol); F0: Zero fertilization; CF: Conventional fertilization; OF: Organic fertilization.
Table 3.
Area of compounds detected in S. bicolor leaves.
| Number | Compound | Area F0 (%) | Area CF (%) | Area OF (%) |
|---|---|---|---|---|
| 1 | Butyric acid | ND | 0.33 | ND |
| 2 | Tridecyl trifluoroacetate | ND | 0.15 | ND |
| 3 | (R)-2-octanol | ND | 0.21 | ND |
| 4 | Propanoic acid | ND | 0.36 | ND |
| 5 | 1-Methyldecylamine | 1.43 | ND | ND |
| 6 | Ethylamine | ND | 0.28 | ND |
| 7 | 4-methyl-2-Pentanamine | ND | 0.33 | ND |
| 8 | 4-fluorohistamine | ND | 0.61 | ND |
| 9 | 1-undecanol | 7 | ND | 2.69 |
| 10 | 2H-pyran-2-one | ND | 5.48 | ND |
| 11 | 2-heptanol | ND | 0.35 | ND |
| 12 | Acetic acid | ND | 0.63 | ND |
| 13 | (2,3-dimethyloxiranyl) methanol | ND | 0.39 | ND |
| 14 | 2,3-diethoxy-propionic acid, ethyl ester | ND | 0.42 | ND |
| 15 | 2-nonanol | ND | 0.24 | ND |
| 16 | Butanedioic acid | 2.24 | 1.83 | 1.09 |
| 17 | 1-tetradecene | 6.36 | 5.47 | 2.5 |
| 18 | 1-(1-propynyl)-cyclohexene | ND | 0.42 | ND |
| 19 | (Z)-7-hexadecene | 3.4 | 2.85 | 1.46 |
| 20 | Phenol | 22.64 | 23.35 | 14.01 |
| 21 | 4-(2-methylamino)ethyl)pyridine | ND | 0.04 | ND |
| 22 | 2-fluoro-2′,4,5-trihydroxy-N-methyl-benzenethanamine | ND | 0.89 | ND |
| 23 | (E)-5-octadecene | ND | 0.93 | 0.55 |
| 24 | N,N′-dimethyl-2-butene-1,4-diamine | 0.68 | ND | ND |
| 25 | 1-dodecanamine | ND | 1.53 | ND |
| 26 | 4-hydroxy-benzeneacetonitrile | ND | 5.29 | ND |
| 27 | N,N-dimethyl-dimethylphosphoric amide | ND | 0.39 | ND |
| 28 | 2-Octyl benzoate | ND | 0.72 | ND |
| 29 | Benzophenone | ND | 1.06 | 0.69 |
| 30 | N-(3-pyridinylmethylene)benzenamine | 1.09 | ND | ND |
| 31 | 4-amino-2-oxy-furazan-3-carboxylic acid | ND | 0.20 | ND |
| 32 | Piperidin-4-ol, 1,3,3-trimethyl-4phenyl | ND | 0.37 | ND |
| 33 | Benzene, 4-bromo-1,3-dimethoxy-6-(4-acetylphenyliminomethyl) | ND | ND | 0.93 |
| 34 | Phthalic acid, isobutyl 4-isopropylphenyl ester | ND | 0.24 | ND |
| 35 | 9-hydroxy-3,4-dihydro-2H-1,4-ethanoquinoline-9-carboxylic acid | ND | ND | 1.14 |
| 36 | 5,6-dihydro-2-(4-nitrophenyl)-4H-1,3-oxazin-5-one | ND | 0.11 | ND |
| 37 | 1H-pyrrolo [1,2-a]benzimidazolium,2,3-dihydro-4-(1,2,3,4-tetrahydro-6-hydroxy-1,3-dimethyl-2,4-dioxo-5-pyrimidinyl)-, hydroxide | ND | ND | 1.07 |
| 38 | 5-[[[3,4,5-trimethoxyphenyl]imino]methyl]-2,4-pyrimidinediamine | ND | 37.73 | ND |
| 39 | (4-methoxy-phenyl)-(5-p-tolyl-furan-2-ylmethylene)-amine | 46.89 | ND | 68.02 |
| 40 | 1,2-benzenedicarboxylic acid | ND | 0.77 | 1.62 |
| 41 | 2-methyl-benzothiazole | 0.55 | ND | ND |
| 42 | 2-bromo-N-methyl-2-propen-1-amine | ND | 0.35 | ND |
| 43 | 4-methyl-2-pentanamine | 0.83 | ND | ND |
| 44 | (2,2-dichlorocyclopropyl)methanol | 0.06 | ND | ND |
| 45 | 1,3-benzenedicarboxylic acid | ND | 0.34 | ND |
| 46 | 4-phenoxy-2-phenyl-1-naphthalenol | ND | ND | 1.06 |
| 47 | 3-methyl-1-(4-toluidino)pyrido [1,2-a]benzimidazole-4-carbonitrile | ND | ND | 0.2 |
| 48 | 5-(p-aminophenyl)-4-(p-nitrophenyl)-2-thiazolamin | ND | ND | 0.3 |
| 49 | 2-oxo-1,4,5-triphenyl-4-imidazolin | ND | ND | 0.68 |
| Number of compounds detected per treatment | 12 | 34 | 16 |
ND: Not detected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



