
Submitted:
29 May 2024
Posted:
30 May 2024
You are already at the latest version
Abstract
The cDNA sequences of the common gonadotrophic hormone α-subunit (ag-GTHα) and 13 of the specific follicle-stimulating hormone β-subunit (ag-FSHβ) of the giant Amazonian fish Ara-14 paima gigas, have been previously isolated by our research group. A synthesis of ag-FSH in HEK293 15 cells and its purification and preliminary characterization were also carried out. In the present work, 16 ag-FSH was obtained and purified from the same host cells and an extensive physical chemical 17 characterization was performed via RP-HPLC, HPSEC and MALDI-TOF-MS. Ag-FSH, when com-18 pared to human FSH (hFSH), showed a higher hydrophobicity by RP-HPLC and a higher molecular 19 mass (MM) via HPSEC. The same higher MM was also confirmed via MALDI-TOF-MS: 35,353 Da 20 for ag-FSH against 31,969 Da for hFSH. Its immunological activity was also confirmed via an hFSH 21 ELISA, in comparison with the highly purified pituitary preparation of hFSH-AFP7298A, from the 22 National Hormone and Pituitary Program (NHPP-USA). Finally, an in-vitro bioassay, based on the 23 stimulated release of 11-ketotestosterone (11-KT) from immature A. gigas testis, quantified its bio-24 logical activity in comparison with human chorionic gonadotropin (hCG) and with the WHO Inter-25 national Standard of recombinant hFSH (NIBSC-08/282) This suggests that injections of ag-FSH in 26 A. gigas and ag-FSH cDNA Gene Therapy applications should improve the reproductive functions 27 of this threatened species.
Keywords:
Arapaima gigas
; follicle-stimulating hormone
; in-vitro bioassay
; immature testis
; 11-keto-29 testosterone.
1. Introduction
Follicle-stimulating hormone (FSH) and luteinizing hormone (LH) are two pituitary gonadotrophic hormones that regulate reproductive processes such as gametogenesis and follicular growth in all vertebrates, including fish [1,2,3,4]. These gonadotropins can, therefore, be employed to improve breeding in reproductive centers that could be established for endangered and overexploited species of fish, particularly important for alimentary and commercial purposes, like the giant Amazonian fish Arapaima gigas, called pirarucu in Brazil [5,6,7,8].
A. gigas belongs to the family of Arapaimidae and to the order of Osteoglossiformes; it is native to the Amazon river basin, present in Brazil, Ecuador, Colombia, Peru and Bolivia, and can reach 3 meters in length and weigh up to 250 kg [9]. This species is very important for human nutrition and extractivism in the region, but unfortunately it is in danger of disappearing because of human and industrial exploitation [5,10,11,12] and because of its limited reproductive capacity in captivity [6,7,13,14,15,16].
Given the extensive experience of our research group with human pituitary gonadotrophins, in previous work we cloned and characterized the cDNA of the α-subunit (GTHα) of A. gigas, which is common to the two gonadotropins [17] and both hormone-specific β-subunits (ag-FSHβ and ag-LHβ) [18]. These subunits form, together with the α-subunits, the two heterodimeric, non-covalently bound, glycoprotein hormones. In the latter work, ag-FSH was also synthesized in human embryonic kidney 293 (HEK293) cells and preliminarily purified and characterized. In the present work, purified ag-FSH was extensively characterized via reversed-phase high-performance liquid chromatography (RP-HPLC), high-performance size-exclusion chromatography (HPSEC), matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF-MS), immunoassay (ELISA) and, in particular, by an in-vitro bioassay. In this bioassay, the biological activity of the ag-FSH preparation was determined via release of 11-KT from immature A. gigas testis in comparison with hCG [19] and with the WHO International Standard of recombinant hFSH, NIBSC 08/282.
A schematic figure (Figure 1) representing the main signaling pathways in the control of A. gigas reproduction involving A. gigas growth hormone (ag-GH), ag-FSH and ag-LH, was reported in our previous work: Figure 1 from Lima et al., [20], adapted from Migaud et al., [21].
Purified ag-GH, ag-FSH and ag-LH cDNAs, can be used for preliminary Gene Therapy applications, directed to fertility studies and to improving reproduction in captivity of this important species of fish. Ag-GH cDNA will be particularly useful, considering its action on somatic growth and muscle mass increase.
2. Materials and Methods
2.1. Qualitative and Quantitative Analysis of Purified ag-FSH
Purification of ag-FSH, derived from conditioned medium of transfected HEK293 cells, was performed by two chromatographic steps, RP-HPLC and HPSEC as previously described, obtaining a total of 400 µg of purified material (~80 mg/liter) [18]. A sample of purified material (50 µl) was then analyzed via RP-HPLC on a Vydac C4 column (300 Å pore size, 5 µm particle size, 25 cm x 4.6 mm I.D.) from Grace-Vydac (Fisher Scientific, USA). The elution and gradient conditions were specifically set up for maintaining the FSH heterodimer undissociated. The same quantity of ag-FSH solution (50 µl) was applied to a HPSEC G200 column (125 Ǻ pore size, 10 µm particle size, 60 cm x 7.5 mm I.D.) from Tosoh Bioscience (Tokyo, Japan). The mobile phase was 0.15 M NaCl, sodium phosphate buffer (pH 7.0; 0.02 M), with a flow rate of 1.0 ml/min. In both analyses, a reference preparation of human pituitary FSH-AFP7298A from the National Hormone & Peptide Program (NHPP, Torrance, CA, USA) was used.
2.2. MALDI-TOF-MS
The MM of ag-FSH preparation was determined (using approximately 40 µg of glycoprotein) by MALDI-TOF-MS on an UltrafleXtreme equipment (Bruker Daltonics, Bremen, Germany), operated in the linear positive ion mode. A diluted glycoprotein sample (1:5; 1:10 and 1:20 from a 1 mg/ml solution) was mixed 1:1 with MALDI matrix solution (sinapinic acid 7 mg/mL in 0.1% trifluoroacetic acid and 50% acetonitrile) and spotted directly onto the MALDI plate (1 μL). The analysis was performed in linear positive mode in the range of 5,000–40,000 Da.
The Open Source Mass Spectrometry tool data processing software was used for increased resolution analysis in the range 17,000–19,000 Da.
2.3. Immunoenzymatic Assay Based on hFSH ELISA
The immunological activity determination of purified ag-FSH was carried out utilizing a human-FSH ELISA kit (FSH SYM, Vyttra Diagnostics, São Paulo, Brazil) following the manufacturer’s instructions. The kit standard curve was based on six doses determined in duplicate, and the absorbance reading was carried out at 450 nm in a 96-well plate reader (Multiskan EX, Thermo Electron Corp., Finland). The doses provided were: 0; 5.0; 10.0; 25.0; 50.0; 100.0 IU/L, while the calculated standard curve equation was:
Yabs = 0.029 XIU/L + 0.557 (n=6; r=0.924; P<0.01)
Based on WHO protocols defining the potency of highly purified hFSH as 16,000 IU/mg, 1 ng of ultrapure hFSH was found to correspond to 0.016 IU. This relation was, therefore, taken into account when carrying out immunoassay curves based on the commercial standard of the kit or on the pituitary hFSH reference preparation from NHPP. The negative control corresponded to the conditioned medium that was submitted to same transfection procedure with the expression vectors not containing the ag-FSH α- and β-subunit cDNAs and used at different dilutions (1:100, 1:500 and 1:1000).
2.4. In-vitro Bioassay Based on the Release of 11-Ketotestosterone (11-KT) from Immature A. gigas Testes
Testes from three A. gigas (two year-old) were dissected and the immature stage confirmed according to Nuñez and Duponchelle [22]. The dissected testes were ground using a scalpel, uniformly homogenized and kept frozen at -80°C until assays were performed. For assays, 20 mg aliquots of testes were pre-incubated for 1 hour at 20°C with 300 µl of solution containing 0.88 mg/ml of Dulbecco’s Modified Eagle Medium (Gibco®, Life TechnologiesTM, Grand Island, USA), 0.5 mg/ml of glucose (Equiplex®, Aparecida de Goiânia, Brazil) and 1.0% w/v Antibiotics with antimycotics (Gibco®, Life TechnologiesTM). After pre-incubation, this medium was pipetted off and changed with 300 µl of fresh medium containing various concentrations of the hormones under analysis: ag-FSH (in house produced), pituitary hFSH (WHO/NIBSC 08/282, South Mimms, UK) and hCG (Sigma Aldrich CG5-1VL, Saint Louis, USA).These incubations continued for 24 hours at 20°C. Negative controls, were performed with medium alone or with different concentrations of Bovine Serum Albumin-BSA (Sigma Aldrich 5470-1G, Saint Louis, USA). After incubation, the medium was collected and stored at -80°C until ELISA analysis. For this analysis 50 µl medium were used in order to determine the 11-KT concentration according to the ELISA kit protocol (Cayman Chemical Inc., Michigan, USA), reading the microplates at 405 nm using an ELX808 reader (Biotek, Swindon, UK). The 11-KT ELISA assay was previously validated for A. gigas by Torati et al. [16]. Results were back calculated in pg per 100 mg testis tissue. Dose-response curves, relating the logarithm of the applied hormonal dose (ln X, ng/ml) versus the amount of released 11-KT (Y, pg of 11-KT/100 mg testis tissue) were thus obtained.
3. Results
3.1. Qualitative and Quantitative Analysis of ag-FSH by RP-HPLC and HPSEC
Figure 2 shows the qualitative and quantitative analysis of purified ag-FSH. The asymmetric peak of ag-FSH obtained by RP-HPLC (Figure 2B) and the higher amount of material when compared to the HPSEC peak (Figure 2D), may indicate the presence of different glycosylated isoforms, recalling that this purified ag-FSH was obtained from HEK293 cells and does not necessarily have a glycosylation pattern similar to pituitary hFSH. The same chromatograms also indicate a higher hydrophobicity of ag-FSH compared to hFSH. HPSEC exhibited two perfectly symmetric peaks of either hFSH or ag-FSH; considering its retention time, the latter should have a larger MM.
3.2. MALDI-TOF-MS of ag-FSH, in Comparison with the hFSH Standard
Figure 3 compares a MALDI-TOF-MS analysis for the pituitary hFSH-NHPP standard to our preparation of ag-FSH. In Figure 3A the two α- and β-subunits of hFSH are well separated, providing a heterodimeric form of 31,969 Da, while the MM of a similar pituitary hFSH preparation, determined in previous work, was 30,976 Da. In Figure 3B the same analysis for ag-FSH shows that the two α- and β-subunits exhibit very similar MM (17,609 Da and 17,743 Da). The MM of the heterodimeric form is therefore 35,352 Da. Again, this significant difference is surely due to different glycosylation pattern, since the MM of the protein backbones of hFSH and of ag-FSH are 22,689 Da and 22,322 Da, respectively. Considering, moreover, that the identities of the amino acid compositions between hFSH and the ag-FSH α- and β-subunits have been found to be 60.0% and 45.1%, respectively [18], we hypothesized that an anti-hFSH polyclonal antibody acting in a traditional immunoassay could also have some affinity for ag-FSH.
3.3. Immunological Activity Determination of ag-FSH
Indeed, it was possible to demonstrate the immunological activity of ag-FSH against an anti-hFSH polyclonal antibody (Figure 4). The standard kit preparation, as expected, was much less active than the highly purified NHPP reference preparation of hFSH. A negative control curve, carried out with the material obtained from HEK293 cells transfected with the vector not containing ag-FSH cDNA, indicated the presence of a product that was clearly inactive. The hFSH reference preparation from NHPP presented the highest immunological potency (slope of the curve=0.277), while ag-FSH, though still active, had a substantially lower potency when compared to hFSH-NHPP.
3.4. In-Vitro Bioassay of ag-FSH, Based on the Stimulated Release of 11-KT from Immature A. gigas Testes
Figure 5 presents the curves obtained in the in-vitro bioassay, based on the stimulated release of 11-KT from immature A. gigas testes, induced by different hormones. Considering the slopes of the regression lines, ag-FSH exhibits a slightly higher logarithmic dose-biological response profile than hFSH and hCG (1.51-fold and 1.39-fold greater slope, respectively). However, hFSH NIBSC-08/282 is biologically active at substantially lower hormone concentrations, indicating that is probably the purest product.
4. Discussion
An extensive characterization of purified ag-FSH obtained in HEK293 cells has been carried out via physical chemical, immunological and biological methodologies.
Previous work [18] demonstrated that the α-subunit (ag-GTHα) protein backbone has a MM of 10,489 Da, while that of the ag-FSH β-subunit is of 11,833 Da. For heterodimeric ag-FSH, this corresponds to a total protein backbone of 22,322 Da. Considering that ag-FSH was synthesized in human cells (HEK293), its glycosylation is undoubtedly qualitatively and quantitatively different from that of natural pituitary A. gigas-FSH.
The characterization via RP-HPLC and HPSEC showed a heterodimeric protein that was different from the human hormone (hFSH) in terms of hydrophobicity, presenting a much higher retention time and therefore much higher hydrophobicity via RP-HPLC and a higher molecular mass via HPSEC. Both methodologies were perfectly suitable for a preliminary identity test and for a qualitative and quantitative analysis of ag-FSH.
Accurate MM determination by MALDI-TOF-MS provided a value of 31,969 Da for pituitary hFSH from NHPP, which is only 3.2 % higher than the value of 30,976 Da determined by us in previous work for a similar pituitary-extracted hormone. The MM of ag-FSH, derived from the summation of the MM of the α- and β- subunits, was 35,352 Da, corresponding to a difference of 10.6% compared to hFSH-NHPP. Taking into consideration the protein backbone (22,322 Da), our synthesized ag-FSH should have a carbohydrate moiety of 58.4%, while for hFSH-NHPP (protein backbone=22,689 Da) the carbohydrate moiety accounts for 40.9%.
As already noted, the second characterization was based on the hypothesis that ag-FSH should be recognized by anti-hFSH polyclonal antibodies. This was proven in the experiment shown in Figure 4, where the purest preparation of hFSH (hFSH-NHPP) exhibited an immunoactivity 9.5-fold higher in comparison with the kit standard preparation of hFSH, while in comparison with purified ag-FSH (still clearly immunoreactive against anti-hFSH antibodies), its immunopotency was ~1565-fold higher. We consider this to be an additional, more specific identity test.
Of course, when one considers a hormone and its possible applications, an in-vivo or in-vitro bioassay is, in our opinion, the strongest proof for its identity and relative potency. The results of this bioassay, presented in Figure 5, are based on the release of 11-KT by immature A. gigas testes, stimulated by human-CG, by human FSH or by recombinant ag-FSH.
Androgen release from immature testicular interstitial cells, and consequent spermatogenesis initiation, serves as the basis for different homologous or heterologous in-vivo or in-vitro bioassays for measuring the activity of fish gonadotrophins [19,23,24,25,26,27,28]. In the present work, for the purpose of ag-FSH biological characterization, we developed a homologous in-vitro bioassay based on A. gigas immature testis tissue and compared the data obtained with those of another homologous bioassay that measured aj-FSH activity based on Anguilla japonica immature testis tissue. In this context, it has been reported that eels do not mature spontaneously in captivity and require exogenous gonadotrophin administration for the induction of gonadal maturation [19]; this seems not to be the case of A. gigas, which reaches final oocyte maturation but fails to ovulate in the absence of optimal conditions in captivity [15].
Converting the hCG doses (IU) on the abscissa of Figure 6 of Kamei et al. [19] to nanograms derived from a known highly purified preparation (1 IU = 50 ng) [29] indicated that ~150 ng/ml of pure hCG (~3IU/ml) should stimulate the release of ~3,500 pg 11-KT/100 mg of A. japonica testis tissue. For the corresponding A. gigas testis bioassay, ~3,700 pg 11-KT/100 mg of A. gigas testis tissue, should be released by ~150 ng/ml of hCG. This indicates that both A. japonica and A. gigas testis tissue present approximately the same bioactive response upon hCG stimulation. However, they would probably not present the same response upon ag-FSH or aj-FSH stimulation.
The inter-assay and inter-laboratory precision and accuracy of the proposed bioassay requires further confirmation in order to ensure reliable comparisons between different recombinant ag-FSH productions or different A. gigas pituitary extracts. The recombinant ag-FSH cDNA will, however, be an important tool for A. gigas gene therapy applications directed towards improving the reproduction of this important species of fish in captivity.
Author Contributions
Conceptualization, L.S.T, P.B. and C.N.P; methodology, E.R.L., T.C., J.E.O, T.S. and M.F.S.; software, E.R.L. and M.F.S.; validation, E.R.L. and J.E.O.; formal analysis, E.R.L, T.S. and L.C.T.; investigation, E.R.L., J.E.O. and M.F.S.; resources, L.S.T. and P.B..; data curation, L.S.T., P.B. and C.N.P..; writing—original draft preparation, L.S.T., P.B. and C.N.P..; writing—review and editing, L.S.T., P.B. and C.N.P..; visualization, E.R.L. and J.E.O.; supervision, P.B.; project administration, P.B.; funding acquisition, L.S.T. and P.B.. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the São Paulo State Research Foundation-FAPESP, São Paulo, Brazil, research project No. 2017/26369-1, and No. 2020/16549-5; by CNPq fellowships No. 169008/2018-5, No. 134109/2018-0 and No. 142319/2015-5. Lucas S.Torati, in particular, thanks the Tocantins State Research Foundation-FAPT (Palmas, TO, Brazil) for its support.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest. Testis used in this trial were frozen from a study approved by the Ethics Committee for the Use of Animals–CEUA of the National Research Center on Fisheries, Aquaculture and Agricultural Systems–CNPASA (specific protocol no. 09).
References
- Huang, H.; Zhang, Y.; Huang, W.; Li, S.; Zhu, P.; Liu, Y.; Yin, S.; Liu, X.; Lin, H. Molecular characterization of marbled eel (Anguilla marmorata) gonadotropin subunits and their mRNA expression profiles during artificially induced gonadal development. Gen. Comp. Endocrinol. 2009, 162, 192–202. [CrossRef]
- Levavi-Sivan, B.; Bogerd, J.; Mañanós, E.L.; Gómez, A.; Lareyre, J.J. Perspectives on fish gonadotropins and their receptors. Gen.Comp. Endocrinol. 2010, 165, 412–437. [CrossRef]
- Schulz, R.W.; de França, L.R.; Larteyre, J.J.; Le Gac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen.Comp. Endocrinol. 2010, 165, 390–411. [CrossRef]
- Senthilkumaran, B. Recent advances in meiotic maturation and ovulation: Comparing mammals and pisces. Front. Biosci. 2011,16, 1898–1914. [CrossRef]
- Borella, M.I.; Venturieri, R.; Mancera, J.M. Immunocytochemical identification of adenohypophyseal cells in the pirarucu (Arapaima gigas), an Amazonian basal teleost. Fish. Physiol. Biochem. 2009, 35, 3–16. [CrossRef]
- Mylonas, C.C.; Fostier, A.; Zanuy, S. Broodstock management and hormonal manipulations of fish reproduction. Gen. Comp. Endocrinol. 2010, 165, 516–534. [CrossRef]
- Castello, L.; Stewart, D.J.; Arantes, C.C. Modeling population dynamics and conservation of arapaima in the Amazon. Rev. Fish Biol. Fish. 2011, 21, 623–640. [CrossRef]
- Zohar, Y.; Zmora, N; Trudeau, V.L.; Munoz-Cueto, J.A.; Golan, M.A. A half century of fish gonadotropin-releasing hormones: Breaking paradigms. J. Neuroendocrinol. 2022, 34, e13069. [CrossRef]
- Nelson, J.S.; Grande T.C.; Wilson, M.V.H. Fishes of the world, 5th ed.; Wiley: New Jersey, 2016.
- Cavole, L.M.; Arantes, C.C.; Castello, L. How illegal are tropical small-scale fisheries? An estimate for Arapaima in the Amazon. Fish. Res. 2015, 168, 1–5. [CrossRef]
- Watson, L.C.; Stewart, D.J.; Kretzer, A.M. Genetic diversity and population structure of the threatened Giant Arapaima in southwestern Guyana: Implications for their conservation. Copeia 2016, 104, 864–872. [CrossRef]
- Castello, L.; Stewart, D.J. Assessing CITES non-detriment findings procedures for Arapaima in Brazil. J. Appl. Ichthyol. 2010, 26, 49-56. [CrossRef]
- Chu-Koo, F.; Dugue, R.; Aguilar, M.A.; Daza, A.C.; Bocanegra, F.A.; Veintemilla, C.C.; Duponchelle, F.; Renno, J.F.; Tello, S.; Nunes, J. Gender determination in the Paiche or Pirarucu using plasma vitellogenin. 17 beta-estradiol and 11-ketotestosterone levels. Fish. Physiol. Biochem. 2009, 35, 125–136.
- Garcia, A.; Tello, S.; Duponchelle, F. Patterns of commercial fish landings in the Loreto region (Peruvian Amazon) between 1984 and 2006. Fish. Physiol. Biochem. 2009, 35, 53–67. [CrossRef]
- Torati, L.S.; Lima, A.F.; Kirschnik, L.N.G.; Migaud, H. Endoscopy and cannulation as non-invasive tools to identify sex and monitor reproductive development in Arapaima gigas. Copeia 2019, 107, 287–296. [CrossRef]
- Torati, L.S.; Taylor, J; Mesquita P.E.C.; Migaud, H. ‘GnRHa implants and size pairing effects on plasma and cephalic secretion sex steroids in Arapaima gigas, Gen. Comp. Endocrinol. 2020, 113614. [CrossRef]
- Faria, M.T.; Carvalho, R.F.; Sevilhano, T.C.A.; Oliveira, N.A.J.; Silva, C.F.P.; Oliveira, J.E.; Soares, C.R.J.; Garcez, R.; Santo, P.R.; Bartolini, P. Isolation of the pituitary gonadotrophic α-subunit hormone of the giant Amazonian fish: Pirarucu (Arapaima gigas). Fish. Phys. Biochem. 2013, 39, 683–693. [CrossRef]
- Sevilhano, T.; Carvalho, R.F.; Oliveira, N.A.; Oliveira, J.E.; Maltarollo, V.G.; Trossini, G.; Garcez, R.; Bartolini, P. Molecular cloning and characterization of pirarucu (Arapaima gigas) follicle-stimulating hormone and luteinizing hormone β-subunit cDNAs. PLoS ONE 2017, 12, e0183545. [CrossRef]
- Kamei, H.; Ohira, T.; Yoshiura, Y.; Uchida, N.; Nagasawa, H.; Aida, K. Expression of a biologically active recombinant follicle stimulating hormone of Japanese Eel Anguilla japonica using methylotropic yeast, Pichia pastoris. Gen. Comp. Endocrinol. 2003, 134, 244-254. [CrossRef]
- Lima, E.R.; Passos Freire, R.; Suzuki, M.F.; Oliveira, J.E.; Yosidaki, V.L.; Peroni, C.N.; Sevilhano, T.; Zorzeto, M.; Torati, L.S.; Soares, C.R.J.; Lima, I.D.M.; Kronenberger, T.; Maltarollo, V.G.; Bartolini, P. Isolation and characterization of the Arapaima gigas Growth Hormone (ag-GH) cDNA and three-dimensional modeling of this hormone in comparison with the human hormone (hGH). Biomolecules 2023, 13, 158. [CrossRef]
- Migaud, H.; Davie, A.; Taylor, J.F. Current knowledge on the photoneuroendocrine regulation of reproduction in temperature fishspecies. J. Fish Biol. 2010, 76, 27–68. [CrossRef]
- Núñez, J.; Duponchelle, F. Towards a universal scale to assess sexual maturation and related life history traits in oviparous teleost fishes’, Fish Physiol. and Biochem. 2009, 35: 167-80. [CrossRef]
- Miura, C.; Miura. T.; Yamashita, M.; Yamauchi, K.; Nagahama, Y. Hormonal induction of all stages of spermatogenesis in germ-somatic cell coculture from immature Japanese eel testis. Develop. Growth Differ. 1996, 38, 257-262. [CrossRef]
- Kamei, H.; Kawazoe, I.; Kaneko, T.; Aida, K. Purification of follicle-stimulating hormone from immature Japanese eel, Anguilla japonica, and its biochemical properties and steroidogenic activities. Gen. Comp. Endocrinol. 2005, 143, 257-266. [CrossRef]
- Kamei, H.; Kaneko, T.; Aida, K. In vivo gonadotropic effects of recombinant Japanese eel follicle-stimulating hormone. Aquaculture, 2006, 262 (2), 771-775. [CrossRef]
- Ko, H.; Park, W.D.; Kim, D.J.; Kobayashi, M.; Sohn, Y.C. Biological activities of recombinant Manchurian trout FSH and LH: their receptor specificity, steroidogenic and vitellogenic potencies. J. Mol. Endocrinol. 2007, 38, 99-111. [CrossRef]
- Kazeto, Y.; Kohara, M.; Miura, T.; Miura, C.; Yamaguchi, S.; Trant, J.M.; Adachi, S.; Yamaguchi, K. Japanese eel follicle-stimulating hormone (Fsh) and luteinizing hormone (Lh): production of biologically active recombinant Fsh and Lh by Drosophila S2 cells and their differential actions on the reproductive biology. Biol. Reprod. 2008, 79, 938-946. [CrossRef]
- Hayakawa, Y.; Nagaya, H.; Kaki, H.; Hotta, K.; Kobayashi, M. Induction of spermatogenesis in Japanese eel by recombinant goldfish gonadotropins. Fish.Sci. 2009, 75, 137-144. [CrossRef]
- Almeida, B.E.; Oliveira, J.E.; Carvalho, C.M.; Dalmora, S.L.; Bartolini, P.; Ribela, M.T.C.P. Analysis of human luteinizing hormone and human chorionic gonadotropin of different origins by reversed-phase high-performance liquid chromatography. J. Pharmaceut. Biomed. 2010; 53: 90-97. [CrossRef]
Figure 1.
Schematic figure representing the main signaling pathways in the control of A. gigas reproduction.
Figure 1.
Schematic figure representing the main signaling pathways in the control of A. gigas reproduction.
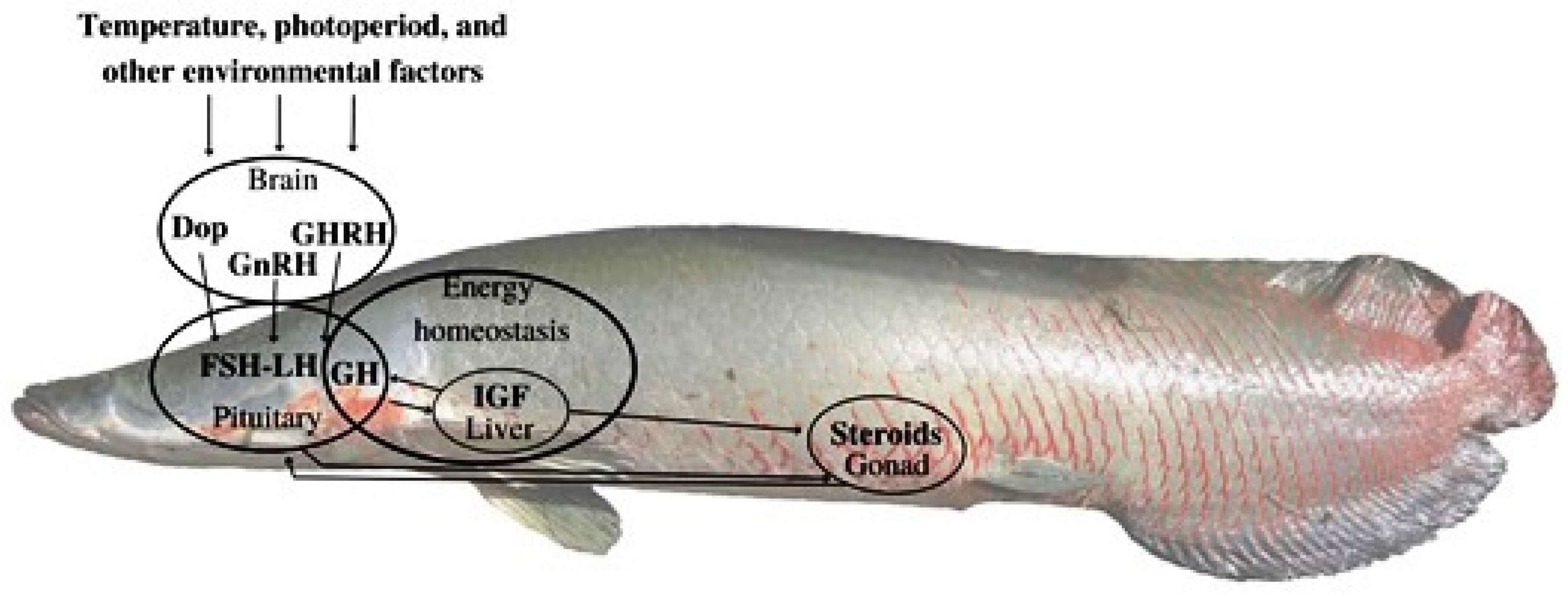
Figure 2.
Qualitative and quantitative analysis of the final product (ag-FSH) after two purification steps. (A) Reference preparation of pituitary hFSH from NHPP, 5 µg/5 µL, via RP-HPLC. (B) Analysis of the final product, ag-FSH (4.8 µg/50 µl), via RP-HPLC. (C) Reference preparation of pituitary hFSH from NHPP, 5 µg/5 µL, via HPSEC. (D) Analysis of the final product of ag-FSH (4 µg/50µl), via HPSEC.
Figure 2.
Qualitative and quantitative analysis of the final product (ag-FSH) after two purification steps. (A) Reference preparation of pituitary hFSH from NHPP, 5 µg/5 µL, via RP-HPLC. (B) Analysis of the final product, ag-FSH (4.8 µg/50 µl), via RP-HPLC. (C) Reference preparation of pituitary hFSH from NHPP, 5 µg/5 µL, via HPSEC. (D) Analysis of the final product of ag-FSH (4 µg/50µl), via HPSEC.
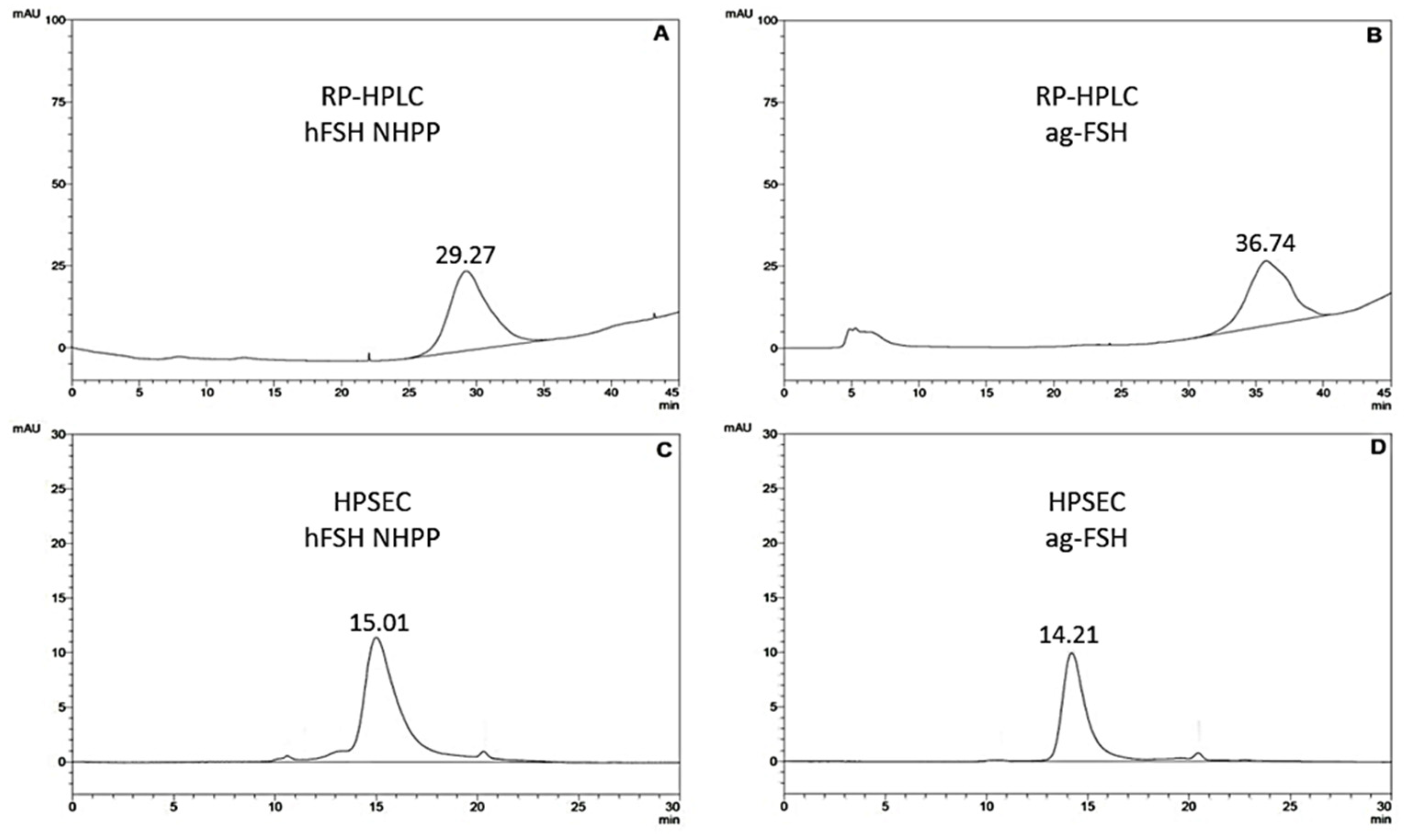
Figure 3.
(A) MALDI-TOF-MS analysis of pituitary hFSH reference preparation (FSH-SIAFP-B-3 from the National Hormone & Peptide Program, NHPP, Torrance, CA, USA): hFSH, MM = 31,969 Da and (B) ag-FSH, MM= 35,352 Da. In B the peaks of interest are also shown at a higher resolution.
Figure 3.
(A) MALDI-TOF-MS analysis of pituitary hFSH reference preparation (FSH-SIAFP-B-3 from the National Hormone & Peptide Program, NHPP, Torrance, CA, USA): hFSH, MM = 31,969 Da and (B) ag-FSH, MM= 35,352 Da. In B the peaks of interest are also shown at a higher resolution.
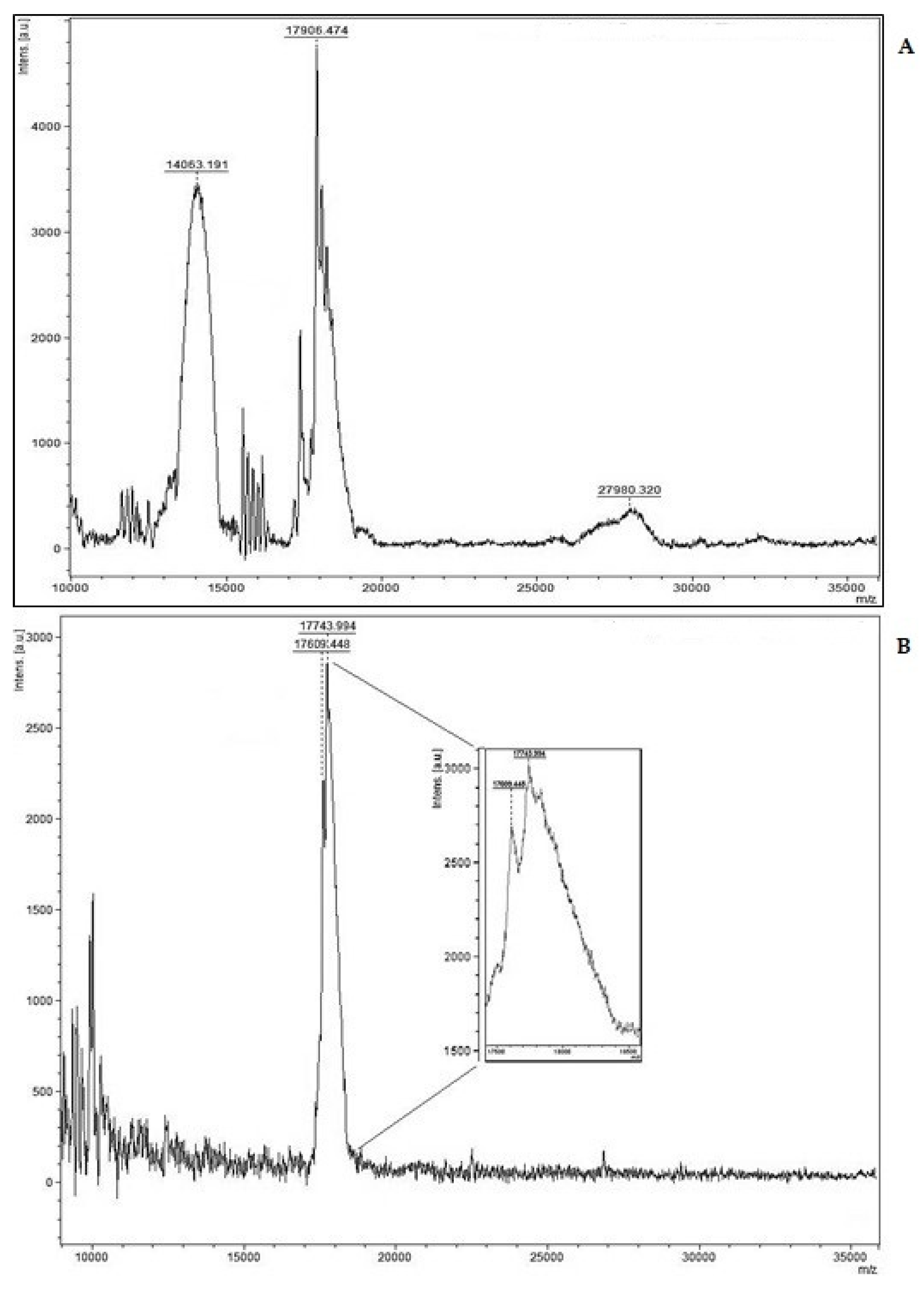
Figure 4.
Dose-response curves based on the human-FSH ELISA kit applied to immunological activity determination of purified ag-FSH. (A) Standard curve based on ELISA kit reagents: YAbs=0.029XIU/ml + 0.557 (n=6; r=0.924; P<0.01) i.e., 9.5-fold less active than hFSH-NHPP (B) Negative control at different dilutions. (C) hFSH reference preparation from NHPP, used also as quantitative standard: YAbs=0.277Xng/ml + 0.501 (n=5; r=0.971; P<0.01) (D) purified ag-FSH: YAbs=0.177.10-3 Xng/ml + 0.004 (n=5; r=0.999; P<0.001), i.e., 1,565-fold less active than hFSH-NHPP.
Figure 4.
Dose-response curves based on the human-FSH ELISA kit applied to immunological activity determination of purified ag-FSH. (A) Standard curve based on ELISA kit reagents: YAbs=0.029XIU/ml + 0.557 (n=6; r=0.924; P<0.01) i.e., 9.5-fold less active than hFSH-NHPP (B) Negative control at different dilutions. (C) hFSH reference preparation from NHPP, used also as quantitative standard: YAbs=0.277Xng/ml + 0.501 (n=5; r=0.971; P<0.01) (D) purified ag-FSH: YAbs=0.177.10-3 Xng/ml + 0.004 (n=5; r=0.999; P<0.001), i.e., 1,565-fold less active than hFSH-NHPP.
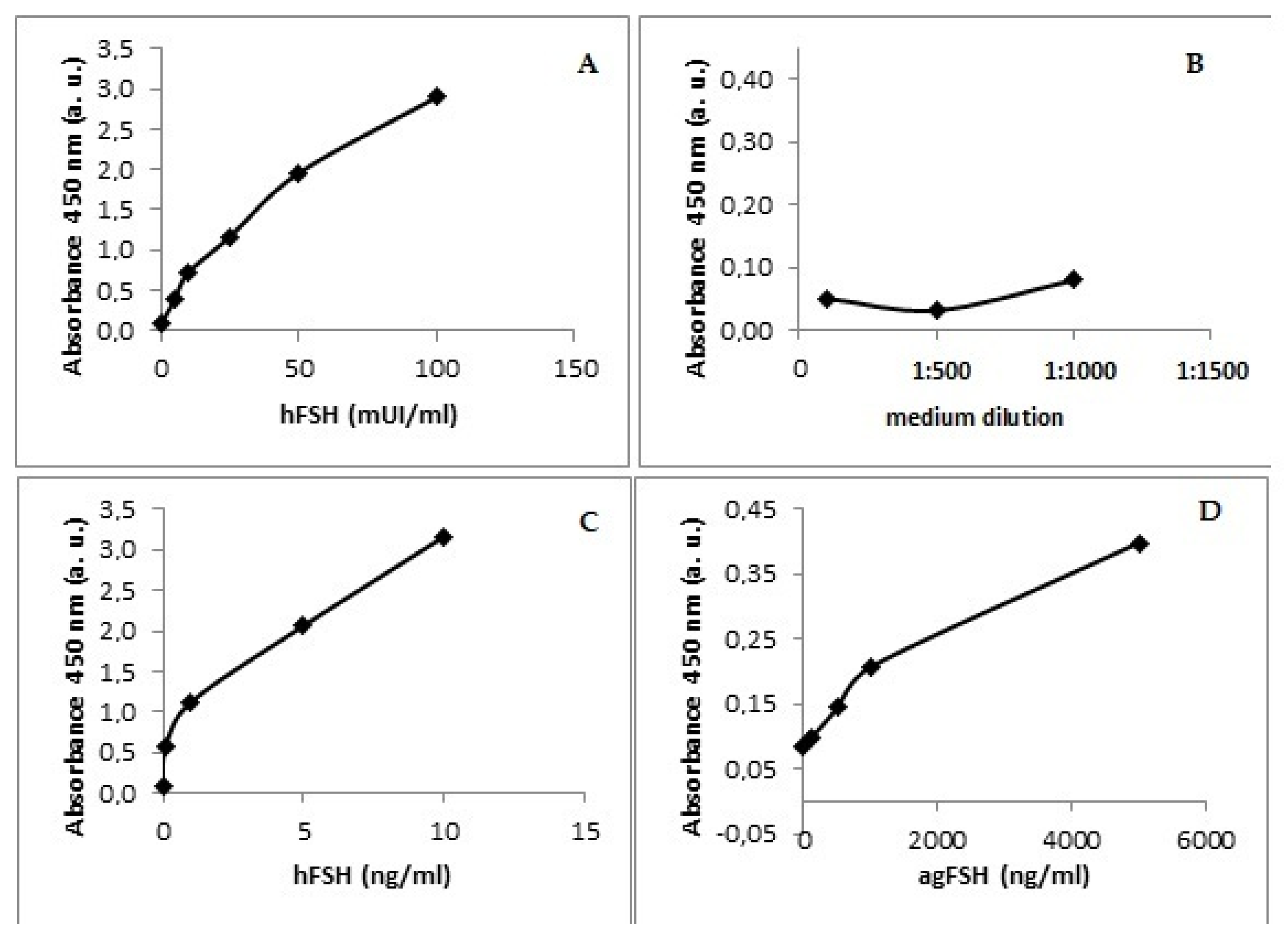
Figure 5.
Dose-dependent 11-KT secreting activity of hFSH  ; ag-FSH
; ag-FSH  and hCG
and hCG  in immature Arapaima gigas testis.
in immature Arapaima gigas testis.
; ag-FSH and hCG in immature Arapaima gigas testis.
Figure 5.
Dose-dependent 11-KT secreting activity of hFSH ; ag-FSH and hCG in immature Arapaima gigas testis.
; ag-FSH and hCG in immature Arapaima gigas testis.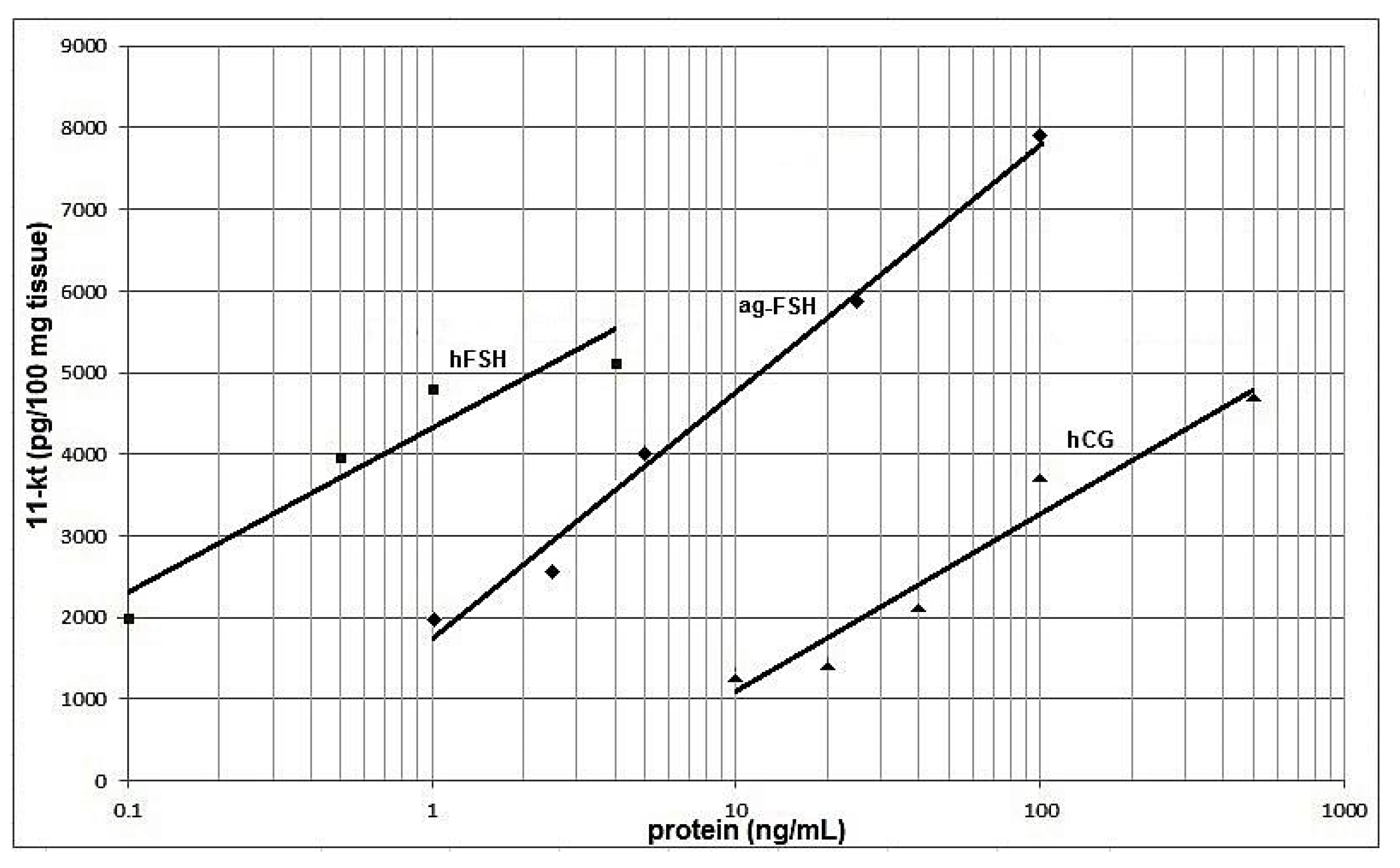
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



