
Submitted:
30 May 2024
Posted:
30 May 2024
You are already at the latest version
Abstract
The olive tree (Olea europaea L.) is an evergreen tree that occupies 19% of the woody crop area and is cultivated in 67 countries on five continents. The largest olive production is concentrated in the Mediterranean basin, where the olive tree has had an enormous economic, cultural and environmental impact since the 7th century BC. In the Mediterranean region, salinity stands out as one of the main abiotic stress factors significantly affecting agricultural production. Moreover, climate change is expected to lead to increased salinisation in this region, threatening olive productivity. Salt stress causes combined damage by osmotic stress and ionic toxicity, restricting olive growth and interfering with multiple metabolic processes. A large variability in salinity tolerance among olive cultivars has been described. This paper aims to synthesize information from the published literature on olive adaptations to salt stress and its importance in salinity tolerance. The morphological, physiological, biochemical, and molecular mechanisms of olive tolerance to salt stress are reviewed.
Keywords:
salt stress
; tolerance
; morphological adaptation
; photosynthesis
; antioxidant defense
; osmotic adjustment
; proteins
; molecular mechanisms
1. Introduction
Soil salinity covers approximately 900 million ha of total cultivated lands on a world scale [1] (Table 1). According to Ludwig et al. [2], 30 per cent of arable land is expected to be lost in the next 25 years and 50 per cent by the middle of the 21st century. In saline soils, the accumulation of soluble salts retains water in the soil and accentuates the problem of desertification. As a consequence, the electrical conductivity of the soil increases. A soil is considered saline when it has an EC ≥a 4 dS/m (about 40 mM NaCl) and the percentage of exchangeable sodium is less than 15% [3].
In arid and semi-arid regions such as the Mediterranean basin, characterised by high evaporation and insufficient leaching, salinity is one of the main abiotic stresses that severely affect agricultural production [4,5]. Furthermore, according to the Fifth Report of the United Nations Intergovernmental Panel on Climate Change, precipitation is likely to decrease, increasing the risk of summer drought [6]. This, together with rising temperatures, will lead to increased salinisation in the Mediterranean area. On the other hand, population growth and its need for good quality water restricts the use of freshwater for irrigation. In coastal areas, saline water (CE > 2,0 dS m-1) could be an alternative for crop irrigation [7]. The use of wastewater could also be an interesting practice in sustainable agriculture. However, although wastewater provides nutrients that improve soil fertility, it can also cause serious salinity problems [8,9,10,11].
Salt-tolerant crop cultivars have been developed using transgenic technologies and conventional breeding approaches. However, these two approaches are insufficient, labour- and time-intensive, and alternative technologies need to be used simultaneously to promote sustainable agriculture. One of them is the use of plant growth-promoting bacteria (PGPB) to mitigate or reduce the impact of salt stress on plants. PGPBs are found in the root zone of plants in saline soils and promote salt stress tolerance through biofilm formation, production of extracellular polymeric substances, production of phytohormones, and nitrogen fixation, as well as, enhancement photosynthesis, photosynthetic pigments, of nutrient uptake and ionic homeostasis in the plant, and increased antioxidant activities under salt stress conditions [12,13,14]. PGPBs promote plant growth through production of hormones such as auxin, cytokinin and gibberellin and reduction of ethylene by 1-aminociclopropano-1-carboxilato (ACC) deaminase [12,15,16]. In this line, it has been reported that exogenous application of gibberellic acid, indole-3-acetic acid, salicylic acid, and brassinosteroids can reduce the effect of salinity in olive tree, improving plant growth, chlorophyll, K+ and proline concentration, and/or decreasing Na+ and Cl- accumulation in the plant [17,18,19,20,21]. Melatonin is an indolamine that has a similar action to indole-3-acetic acid. Melatonin has been reported to improve plant resistance to salt stress by scavenging reactive oxygen species and enhancing antioxidant enzyme activity, photosynthetic efficiency and metabolite content, as well as regulating stress-associated transcription factors [22,23,24,25,26]. Other studies have shown that the application of nanoparticles such as ZnO-NP can reduce the harmful effects of salt stress by interacting with transcription factors, inducing the production of certain phytohormones and osmoprotective solutes [27,28,29,30,31].
Predictions that soil salinity will inevitably increase in the Mediterranean basin, where the olive tree is an important industrial crop, and the serious social and economic consequences that a decrease in olive production would entail, justify the drafting of this review which aims to provide an overview of advances in research on salt stress in the olive tree, promoting the sustainable development of the crop. Considering the variability in salinity tolerance in olive, this review provides an updated view of the effects of salinity on olive growth and development, as well as the role of different physiological, biochemical or molecular mechanisms related to salinity tolerance (Figure 1).
1.1. Olive Tree Cultivation
The cultivated olive tree Olea europaea subsp. europaea var. europaea L. originated from Olea europaea subsp. europaea var. sylvestris (Mill) Lehr by artificial selection from wild populations [32]. This evergreen tree is believed to have been one of the first major domesticated fruit crops in the Old World. In Chalcolithic Levant, evidence for the existence of the olive tree has been found about 6,800-6,300 cal BP [33]. The olive tree originated on the eastern shores of the Mediterranean, in what is now southern Turkey, Syria, Lebanon, Palestine and Israel. From there it spread to populate all the countries bordering the Mediterranean [34]. Since then, it has exerted a notable influence on the Mediterranean economy, history, culture and environment [35,36,37,38], where the olive tree is considered to be the dominant fruit tree in the area [39].
Currently, olive cultivation is present in 67 countries on 5 continents (Figure 2). However, its consumption extends to 179 countries. Worldwide, about 11.594.987 ha of olive tree have been recorded [40]. Of its olives production, 13.39% is used for table olive, while 86.61% is used for olive oil production. Figure 3A–D shows the distribution of olive groves by continent in terms of area, amounts of olives produced, destination of olives (table olive or olive oil), and yield average in year 2021. Data indicate that Europe has the world’s largest area under olive cultivation and productions. However, productivity is highest in Oceania for table olives and olive oil, followed by America for table olives and Europe for olive oil [40].
The olive tree is a species adapted to the semiarid regions of the Mediterranean climate, where it has traditionally been grown in rainfed conditions [41] with a low-density planting system (between 70 and 120 trees/ha) [42]. However, the growing demand for olive products has led to an intensive (200-500 trees/ha) or super-intensive (2500 trees/ha) cultivation model and the introduction of irrigation. In 2021, 60% of olive growing was rainfed, while 40% was irrigated (Figure 3E). In addition, 68.5% of the crop was of low density, having been reduced by 5% compared to recent years and by 1.5% in one year (Figure 3F) [40].
The largest olive production is concentrated in the Mediterranean region, where 97% of the world’s olive oil is produced [43]. Among the top 10 producers of olive oil and table olives, Spain stands out for its high production, accounting for 43.58% of olive oil production and 28.86% of table olive production (Figure 4). In olive oil production, Italy (9.61%), Tunisia (7.94%), and Greece (6.77%) also stand out, and in table olive production, Turkey (20.32%), Egypt (19.80%), Algeria (11.06%) and Morocco (5.68%) [37,40].
Figure 5 shows the evolution of olive oil production over the last three decades, at world, European and four main producing countries level. As we can observe, world production has doubled. This is due to the balanced composition of the oil and its link with greater longevity and lower cardiovascular disease rate, cancer and age cognitive decline [44,45,46]. The trend in international production is marked by Spanish production, where the main olive-growing regions are Andalusia, Extremadura and Castilla La Mancha [37].
More than 2600 olive cultivars are cultivated throughout the world, many of which are traditional and very old, with little representation in today’s olive growing (Table 2). These cultivars have different genetic characteristics and respond differently to biotic and abiotic stresses [47], which offers a wide field of study in the adaptation of the olive grove to a changing environment.
2. Mechanisms of Olive Tolerance to Salt Stress
Salt stress tolerance is a multigenic trait in which different mechanisms are involved [48,49]. Olive is considered moderately tolerant to salinity [50], although differences in response to salt stress have been observed between cultivars [51,52,53,54,55,56,57].
The mechanisms of salt tolerance in olive are attributed to its ability to limit the flow of salt ions to sensitive shoot organs, which would accumulate in the root, as well as to its ability to maintain an adequate K+/Na+ ratio in actively growing tissues [7,52,53,58,59,60]. In addition, its tolerance to salt stress can be improved through osmotic adjustment and reactive oxygen species (ROS) scavenging at the cellular level [21,48,56,57,61,62,63,64].
Sodium ion enters the root cells passively via non-selective ion channels and K+ transporters [65,66,67]. In plants under salt stress, the accumulation of Na+ in the tissue reduces the membrane potential and consequently facilitates Cl- uptake under a chemical gradient [68]. Excess Na+ is transported from the root to the shoot by transpiration, damaging the growing tissues [69]. For plant survival, it is essential to maintain Na+ homeostasis under salt stress. The ions exclusion mechanism of leaves has been described in other woody plants, and is closely related to the ability to withstand water deficit [70]. The most sensitive olive cultivars to salt stress have less capacity for Na+ exclusion [71,72,73,74,75]. This tolerance mechanism is shown by most olive cultivars at levels of 50 mM NaCl or lower [52,76]. In studies with the cultivar ‘Leccino’ at 60 mM NaCl it was concluded that Na+ uptake by the root starts within a few hours of salt treatment, however, Na+ transport to the shoot requires more time [77,78,79].
The accumulation of Na+ in the vacuole stimulates the synthesis of osmoprotective solutes to balance the osmotic potential of the cytoplasm with that of the vacuole [80,81,82,83]. In olive, the accumulation of osmolytes in the cytoplasm has been shown to be an adaptive mechanism against salt stress. In this way, the osmotic potential is regulated from the cell to the whole plant [20,48,55,84,85,86,87]. Proline accumulation in roots and leaves has been reported as a response to salinity in some olive cultivars [84,85,86,87,88]. Proline has a high hydration capacity, consequently, proteins that bind to proline prevent their dehydration and denaturation under osmotic stress. In addition, proline may be involved in antioxidant defence by detoxification of excess ROS produced under stress. Therefore, its increase in the cell may be a mechanism activated by the plant to better tolerate salt stress. However, the involvement of proline as an osmolyte is not a widespread response in all olive cultivars and at all salinity levels. Some authors reported a decrease in proline content in olive trees grown under salt stress [21,53,54,82,89]. It has been suggested that proline may also act as a metabolic substrate helping in the maintenance of cellular energy and contributing to other metabolic pathways under stress conditions [90,91,92]. Thus, by acting as a reservoir to enhance plant growth and induce a stress response, it would decrease its levels in the cell [93].
Soluble sugars have also been reported to participate in osmoregulation and ROS scavenging to mitigate salt stress in olive trees [21,54,56,82,86,87,94,95]. Olive trees synthesise mannitol and oligosaccharides as an end product of photosynthesis. Furthermore, the active role of mannitol in the adaptation of olive trees to salinity is evidenced by its increased biosynthesis in olive trees growing under salt stress [20,21,55,86,87,94,96,97,98]. In addition, mannitol helps to quench hydroxyl radical [99,100,101].
Salinity causes oxidative stress by generating ROS that can damage proteins, DNA and membrane lipids [102]. Several studies have addressed the study of oxidative stress caused by salinity in olive trees. Catalase, glutathione reductase, glutathione peroxidase, ascorbate peroxidase and superoxide reductase are among the enzymes that increase their activity in olive trees due to salinisation. However, the role of each of them in combating oxidative stress differs between cultivars [20,54,55,103]. Some studies revealed that olive trees increase superoxide dismutase activity, with the most sensitive genotypes showing the highest activity in a short period of time after exposure to salt [55,103].
3. Morphological, Anatomical and Physiological Adaptation of the Olive Tree to Salt Stress
3.1. Morphological and Anatomical Adaptation
Olive trees can undergo morphological and anatomical changes in response to salinity. Plant growth is a morphological indicator often used in the evaluation of salt stress, although the degree of biomass decline depends on the species and cultivar [53,85,104,105]. In olive plants, growth can be decreased with 50 mM NaCl [20,52,59,61,104,105,106,107]. This decrease in growth in saline medium is manifested in all organs [21,54,60,61,105]. According to studies, the effect of salt is more prominent in the aerial part (shoot + leaves) than in the root at NaCl concentrations above 50 mM, resulting in a higher root/shoot ratio with increasing salinity. Senescent leaf drop may contribute to this [52,53,59,61]. In addition, the elongation of root cells and shoots shows a different tolerance to salt stress [80,108]. The accumulation of salt ions in the leaves is manifested by apical burns, necrosis and old leaf drop [20,54,101,110,111,112] (Figure 6). These toxicity symptoms usually appear at an advanced stage of leaf salinity damage [52,113,114].
The exclusion of saline ions from the shoot is related to structural changes in roots. In studies with ‘Barnea’ olive trees, which is considered a moderately salinity tolerant cultivar, an effective exclusion mechanism is the high turnover rate of the Na+ hyperaccumulating fine roots [64]. In higher root orders salt accumulation and mortality is lower, although they decrease in length and increase in diameter [63,64]. The increase in diameter is due to the increase in size of the periderm cells which accumulate Na+ inside them, acting as an apoplastic barrier against the advance of the ion. In this way, the exclusion of salt ions from vascular tissues is enhanced [64,77]. In addition, these roots decrease the stele area with salinity and increase the diameter of the xylem vessels, increasing the hydraulic conductivity of the root [64,115,116,117]. Consequently, olive tree resistance to salinity has a structural basis.
The higher ion concentration in the outer root layers has also been documented for primary roots of young olive grown under salt stress. In these primary roots, root diameter and cortical cell density decrease, resulting in reduced salt ion uptake. In addition, apoplastic barriers develop in the exodermis and endodermis of sensitive (‘Leccino’) and tolerant (‘Frantoio’) cultivars by accumulation of suberin lamellae [77,118,119]. Rossi et al. [77] suggest that the speed in the development of the apoplastic barrier could be related to the reduced absorption and translocation of large amounts of Na+ into the shoot. In ‘Leccino’ cultivar, early translocation of Na+ to the aerial part has been observed. This may be due to the decreased expression of ATPase 1, SOS1 (plasma membrane Na+/H+ antiporter), vacuolar V-type ATPase and NHX (vacuolar Na+/ H+ antiporter) genes in the early phase of root salt response [79].
Olive leaves show anatomical modifications in response to salinity. Leaf and upper epidermis thickness may increase under salt stress [105,120,121,122]. As leaf thickness increases under salinity stress, internal surface area per unit leaf area increases, internal resistance decreases, and CO2 absorption and water retention potential remain at higher levels [105]. The number of trichomes, the development of collenchyma and sclereids, and the number of conducting vessels increased with salinisation [123,124]. An increase in leaf palisade parenchyma under salt stress has been described in different olive cultivars (‘Chondrolia Chalkidikis’, ‘Manzanilla de Sevilla’ and ‘Kalamon, Meski’) [120,124]. However, Fayed et al. [123] in studies with ‘Picual’, ‘Kalamata’, ‘Hamed’ and ‘Toffahi’ cultivars reported reduction of palisade parenchyma and midrib thickness. In stems, suber, pericycle fibre and liber development were observed under salt stress [124].
3.2. Physiological Adaptation
The slower growth in glycophytes under salt stress is caused by two successive processes: osmotic stress and ionic toxicity in the plant tissue. Firstly, water uptake by the plant is reduced, and then excess ions from the soil enter the plant and damage the plant cells. Salt ions inhibit photosynthesis, alter ionic homeostasis and cause lipid peroxidation of membranes [48,125,126,127,128].
Olive trees under salinity conditions are subject to important physiological changes, e.g., a marked decrease in the rate of photosynthetic assimilation (A), stomatal conductance (gs), and transpiration (E) [20,52,82,95,122,129,130,131,132,133]. Reduced growth under salinity stress is associated with reduced photosynthesis [52,54,105,132]. Studies with different cultivars have shown that photosynthetic rate decreased with increasing level of salt stress [7,20,52,53,54,57,60,73,122,134,135,136]. The effect of salinity on CO2 assimilation rate varies with the salt concentration, the cultivar, duration of the stress, and plant age [105]. The photosynthetic rate decreased in 4-month-old plantlets of the cultivar ‘Frantoio’ with 12.5 mM NaCl for 35 days [73], however in 1-year-old plants of the cultivars ‘Koroneiki’, ‘Kalamata’, ‘Amphisis’ and ‘Kothreiki’ it did not decrease with 50 mM NaCl for 5 months [52]. Similar results were described for 1-year-old plants of ‘Chemlali’ cultivar grown with 50 mM NaCl for 2 months [122]. In most of the cultivars studied there is a decrease in CO2 assimilation with 100 mM NaCl [20,52,54,73,122,133,137,138,139]. It has been reported that there is greater inhibition of CO2 assimilation rate in olive cultivars with high rates of photosynthesis and stomatal conductance [57,122,139,140]. In several studies it has been observed that photosynthesis inhibition is lower in the more tolerant cultivars compared to the more sensitive ones [57,122]. Furthermore, in the salt tolerant cultivar ‘Chemlali’, a more rapid activation of the stress response was observed than in the moderately tolerant ‘Koroneiki’, which was manifested by a later inhibition of photosynthesis [57].
The reduction of the osmotic potential of the olive leaf under salt stress leads to stomata closure [57,73]. This may be due to the accumulation of Na+ ions in stomatal guard cells and can lead to a decrease in internal CO2 availability [20,105,122]. Consequently, the decline in photosynthesis under salt stress may be caused by a decrease in stomatal conductance (gs) due to hydroactive stomatal closure [20,57,95,133,135]. This limitation of CO2 assimilation by stress can lead to the production of ROs by over-reduction of the photosynthetic electron transport chain [54,132,135,141]. However, when leaf turgor potential affects stomatal conductance and net CO2 assimilation rate to different degrees, the ionic (as distinct from the osmotic) component of salt stress would control gas exchange performance [137]. In the cultivars ‘Arbequina’, ‘Royal de Cazorla’, ‘Koroneiki’, ‘Fadak86’, ‘Frantoio’, and ‘Leccino’, photosynthesis and stomatal conductance decreased, while internal CO2 concentration increased under salt stress [54,73,132,137]. In this case, reduced photosynthesis due to salinity could be due to alterations in photosynthetic metabolism and inhibition of Calvin cycle enzymes [54,127,132,142,143,144].
The maximum efficiency of photosystem II photochemistry (Fv/Fm) can decrease up to 0.5 in olive plants under salt stress [122,140]. In the early phase of salt stress with 60 mM NaCl (7 days of treatment), no decrease in Fv/Fm of the olive cultivar ‘Leccino’ has been observed [79]. In the cultivars ‘Gemlik’ and ‘Kilis’, Fv/Fm was not affected after 15 days of growth with 100 mM NaCl, but decreased by about 6% with 200 mM NaCl. In contrast, in the ‘Ayvalik’ cultivar it decreased by 7% after 15 days with 100 mM NaCl [55] and by 17% in ‘Chétoui’ after 21 days with 200 mM NaCl [82]. Similar results were described for ‘Frantoio’ (35 days at 200 mM NaCl [137]. Under long-term salt stress conditions, the Fv/Fm ratio decreased only at high salt levels (200 mM NaCl) in the cultivars ‘Arbosana’, ‘Arbequina’, ‘Chétoui’ and ‘Chemlali’ after 5 months of cultivation [122,137] and in ‘Ntopia’ after 6 months [95]. In contrast, no decrease of Fv/Fm has been detected in several cultivars grown for 6 months with high NaCl [95]. The decrease of Fv/Fm under salt stress indicates photoinhibitory damage that could be due to a direct effect of salt ions on leaf tissue, and in particular on photosystem II [145,146]. Then, slight decreases in the Fv/Fm ratio allow to affirm that PSII is not very susceptible to NaCl stress and that leaves possess effective photoprotection mechanisms to cope with salinity-induced stress [122,147].
The effect of salt stress on olive trees is also manifested by changes in the photosynthetic pigment content [21,53,82,122]. A decrease in leaf chlorophyll content due to salt stress has been documented in different olive cultivars [20,21,53,54,122,132,148]. The decrease in chlorophyll content due to salinity is a typical response associated with increased oxidative stress [100,149,150,151,152]. In fact, in studies with different olive cultivars, the decrease in chlorophyll content and photosynthetic rate under salt stress correlated with increased catalytic activity of catalase and glutathione reductase in leaves [54,150,153,154].
In cultivar ‘Chétoui’ and ‘Chemlali’, increased Car content and Car/Chl ratio were observed by salinity, which can be considered as a response of the olive tree to protect the photosynthetic apparatus from photooxidation [82,122]. Similar results were observed for plants pretreated with indole-3-acetic acid, salicylic acid or quinetin of the cultivar ‘Picual’ and grown with 100 and 200 mM NaCl [21]. Carotenoids can act by quenching Chl fluorescence directly by singlet energy transfer from Chl to Car or indirectly mediated by trans-thylakoid membrane-mediated and ΔpH [155,156,157]. In olive cultivars ‘Arbequina’, ‘Koroneiki’, ‘Chemlali’, the increase in thermal energy dissipation and non-photochemical quenching increased with salinity was associated with an increase in the lutein/Chl ratio [122].
3.3. Water Relations
The photosynthetic response, adjusted by stomatal conductance, results in a change in the water use efficiency (WUE) of plants exposed to osmotic stress [55,57,82]. The effect of salinity on WUE varies with the intensity and duration of the stress [57,95]. In saline environments, differences in WUE between cultivars have been observed [55,57,95]. Under salt stress, an increase in foliar WUE may be an indicator of salinity tolerance [57,160]. WUE increases when the decrease in stomatal conductance is greater than the decrease in photosynthesis. This is may be a consequence of an increase in osmotic adjustment and hydraulic resistance, and a decrease in photosynthesis through stomatal limitation, rather than a decrease in photosynthetic capacity [73,161]. In addition, the closure of stomata under salt stress reduces water loss by transpiration [57,73,79,82,133].
Leaf water potential decreased in different olive cultivars grown at different NaCl levels [55]. Olive is a drought tolerant species and can therefore withstand quite low leaf water potentials without losing its turgor [96,160]. Olive can continue to transpire and photosynthesise at water potentials of -6 and -8 MPa [161,162], and can rehydrate in a short time after having lost 40% of the water in the tissues [163]. This is important for salt stress tolerance, as the first phase of salt stress is osmotic stress associated with cellular dehydration due to decreased water uptake and WUE [48,80]. Therios and Misopolinos [109] reported a decrease of water uptake in salinized olive trees, mainly caused by the decrease of osmotic in the external solution. Therefore, reduction of relative leaf water content (RWC) by salinity has been described in different olive cultivars [54,55,82,132,133,164]. Differences in RWC have been described as a function of cultivar, NaCl level and duration of salt stress [54,55]. Higher values of RGR and leaf water status under salt stress are related to a higher salt ion exclusion capacity of the shoot [54,55]. Furthermore, differences in the decrease of RWC of old and young leaves of the cultivar ‘Chondrolia Chalkidikis’ have been reported, where the decrease of RWC was about 6 times higher in old leaves than in young leaves [133].
4. Biochemical Mechanisms in Olive for Adaptation to Salt Stress
Bulleted lists look like this: Olive trees, renowned for their resilience in diverse environments, particularly saline stress, use not only physiological and morphological strategies but also bio-chemical mechanisms to thrive and persist in severe conditions.
4.1. Mineral Nutrients
Several studies have been carried out on the relationship between salinity and mineral nutrition in olive trees, and have found it to be highly complex. Loupassaki et al. [165] studied the impact of salt stress on the concentrations of nitrogen (N), phosphorus (P), potassium (K+), calcium (Ca2+), magnesium (Mg2+) and sodium (Na+) in the leaves, shoots, and roots of six Greek olive cultivars (‘Koroneiki’, ‘Mastoidis’, ‘Kalamon’, ‘Amphissis’, ‘Kothreiki’ and ‘Megaritiki’). They observed a significant increase in Na+ levels in the various tissues of all cultivars, particularly in the roots, while K+ concentrations dropped in all organs. Similarly, Palm et al. [62] discovered that salinity induced a significant accumulation of Na+ in all tissues of olive cultivars ‘Frantoio’, ‘Leccino’, ‘Lecciana’ and ‘Oliana’, with the highest concentrations in the roots, while K+ content decreased with salt level. They also detected a correlation between leaf Na+ concentration and a reduction in photosynthetic parameters. Bader et al. [60] investigated the salinity effect on three cultivars (‘Picholine’, ‘Meski’, and ‘Ascolana’), and revealed that chloride concentration increased in roots with salinity but showed divergent trends in stems and leaves among cultivars. Potassium concentration decreased in all plant parts, except for the cultivar ‘Picholine’ for which a notable increase in leaves was recorded, leading to significant decreases in K+/Na+ and Ca2+/Na+ ratios under salinity treatment. In a recent study carried out on the ‘Leccino’ cultivar by Sodini et al. [79], a salt treatment resulted in an accumulation of Na+ in the stems, while its concentration remained constant in the leaves. The same treatment induced a variation in Ca2+ levels in roots, while K+ and Mg2+ remained unchanged. They also noted a significant linear relationship between Na+ and Ca2+ in roots, indicating a potential interaction. Different patterns were also noted; for instance, the ‘Leccino’ cultivar accumulated Na+ in roots under saline conditions [75,78]. A lower concentration of Ca2+, K+, Mg2+ induced by salt stress has been observed by many authors [164,165,166].
Potassium deficiency in salinized plants is associated with an increase in Na+ accumulation, indicating the presence of ion competition effects [167]. The reduction in K+ concentration in olive roots, resulting in a low K+/Na+ ratio, may contribute to achieving ionic balance after high uptake of sodium ions [168]. Polyphenols play a key role in reducing osmotic stress by improving the K+/Na+ ratio and water absorption, thus reducing oxidative stress in olive leaves under saline conditions [169,170,171]. Other studies have reported evidence of the influence of salinity-nitrogen interaction on plant growth and metabolism, proposing that N fertilization could mitigate the harmful effects of salinity; however, the precise mechanisms of this interaction, including nitrate reductase (NR) activity, require further investigation [135,172].
Cultivated plants, mainly adapted to low salinity, have difficulties to efficiently absorb and use nutrients in saline conditions due to high concentrations of Na+ and Cl- [173]. In fact, such conditions cause nutrient imbalances in plants as a result of various mechanisms, notably reduced availability, competitive absorption, and physiological inactivation of key nutrients such as potassium [174]. Olive cultivars display reduced growth and leaf damage when foliar Na+ levels exceed 0.4% of dry matter [113,129]. Effective mechanisms to mitigate salinity impacts in olive consist mainly of limiting the uptake and transport of sodium and chloride ions from the roots to the aerial parts, especially evident in salt-tolerant cultivars [7,52]. The ability to exclude sodium at low to moderate salinity levels helps to regulate ion concentration in xylem sap, thus preventing the toxic accumulation of ions in aerial parts; however, high salinity can lead to Na+ transport and accumulation in aerial parts, causing toxic symptoms [7,113]. Salt tolerance in olive cultivars involves efficient mechanisms for excluding and retaining Na+ and Cl- ions in the roots, where calcium plays a crucial role in preserving the integrity of plasma membrane, on the other hand, reduced Ca2+ uptake under salt stress is linked to reduced transpiration rates rather than direct competition with Na+ [52,114]. Tabatabaei [135] stated that nutrient availability and absorption in saline environments are dependent in turn on soil composition, solute concentration, pH, accompanying elements, and environmental factors such as aeration, temperature, and stress.
4.2. Osmotic Adjustment
4.2.1. Proline Accumulation
Osmotic adjustment is an additional adaptive mechanism for olive trees in response to severe conditions. It involves organic compounds such as amino acids or soluble carbohydrates [175,176,177]. The process combines both active synthesis and accumulation of osmolytes in cells [178]. Proline, an essential amino acid, is involved in many different physiological processes in plants, particularly olive trees, significantly influences their ability to tolerate salt exposure. Many studies have investigated the response of olive trees to saline conditions, revealing mainly the accumulation of proline as a result of its role in osmoregulation, although its exact function is not yet fully understood [55,84,179]
Recently, Trabelsi et al. [179] reported that full irrigation of a 26-year-old olive orchard (cultivar ‘Chemlali’) with saline water (EC=7.5 dS m-1) increased the proline content in various organs compared to another fully irrigated with tap water (EC=2.46 dS m-1).
This corroborates numerous previous findings; Ben Rouina et al. [84] observed a significant proline accumulation significantly under salt stress, with higher levels in leaves compared to roots, peaking during the summer season coinciding with low values of relative water content and water potential. Proline concentrations in olive leaves (cultivar ‘Sigoise’) were found to be positively correlated with soil salinity, suggesting a potential involvement of proline in osmotic regulation of cytoplasmic pH or N storage for post-stress periods [180). Ayaz et al. [55] published a significant increase in proline levels in the leaves of three Turkish olive cultivars (‘Gemlik’, ‘Kilis’, and ‘Ayvalık’) subjected to different salt stress treatments. The proline content in olive leaves (cultivar ‘Gemlik’) exhibited a significant increase with worsening salt stress severity, followed by a decline, which indicated that olive trees use proline synthesis as a tool for mitigating salinity-induced osmotic stress [181]. All of this reinforces the idea that proline accumulation correlates with the efficiency of salinity tolerance mechanisms in olive trees, suggesting that the time and magnitude of synthesis are probably linked to the level of salinity tolerance and biochemical strategies of cultivars [182].
On the contrary, in another investigation conducted by Regni et al. [54] on the salinity tolerance of the Croatian wild olive genotype and two well-known cultivars ‘Leccino’ and ‘Koroneiki’, proline levels appeared unchanged or even decreased compared to the control. These authors supposed in this case that osmotic adjustment in olive leaves under hyperosmotic conditions could be achieved by the accumulation of K+ and other organic solutes rather than by the accumulation of proline. In a similar vein, Ben Abdallah et al. [82] recorded an 18% reduction in proline content in the leaves of salt-stressed plants (cultivar ‘Chétoui’) irrigated every other day with 200 mM NaCl in 100% Hoagland’s solution, as compared with control conditions. Proline concentration was measured as a potential marker of cellular hydro-saline imbalance in leaves and showed a decrease during salt stress applied to ‘Royal’ and ‘Arbequina’ olive cultivars [183].
Despite its proven involvement in stress response, the precise function of proline is still widely debated but little clarified [81,138,184,185]. Various possible reasons have been suggested to explain the accumulation of proline in stressed plants. Ramanjulu and Sudhakar [186] attribute this accumulation to plant adaptation to salinity, while other studies propose that proline accumulation may result from salt stress-induced damage [187] or serve as an indicator of salt sensitivity [188], calling into question its role as a marker of plant salt tolerance.
Proline could protect the photosynthetic activity of salt-stressed olive trees by regulating hydration and osmotic adjustment, thus promoting growth even under stressful conditions [84]. Salt-stressed olive trees displayed an increasing proline concentration in their cytoplasm in order to enhance water absorption by tissues during active growth and to maintain ionic balance in vacuoles, via osmotic adjustment effects [189]. Proline in the olive tree acts as a protective osmolyte in the face of environmental stresses, particularly saline conditions, both by reducing toxicity and by promoting osmotic regulation [190,191].
Proline can serve as a source of nitrogen in cells under stress conditions, where its accumulation could be utilized as a form of stored N, considering that proline contributed 12.16% of leaf nitrogen [192,193]. In situations of nitrogen limitation, proline functions as an alternative metabolic substrate under conditions of stress, therefore helping the maintenance of cellular energy and the balance between NADP+ and NADPH. This multifaceted role may be extended to its contribution to many pathways, including the tricarboxylic acid cycle and glutathione biosynthesis, which further emphasizes its dynamic nature as an organic reserve capable of supporting plant growth and meeting stress [90–93,194). Ben Ahmed et al. [138,195] documented that proline supply improved photosynthetic activity and antioxidant defense enzyme activities in stressed plants, and its application, both in the presence and absence of salinity, significantly influenced salt ion distribution in the leaves and roots of ‘Chemlali’ olive trees.
4.2.2. Sugar Accumulation
The accumulation of osmolytes, notably soluble sugars, represents a key survival mechanism for plants under salt stress, allowing them to prevent dehydration and protect cells from oxidative damage [88,196]. This adaptation varies across species and depends on the dose and duration of stress exposure [48].
Trabelsi et al. [179] recorded a significant increase in soluble sugars in olive leaf and root tissues during salt and water stress, mainly synthesized in the leaves. Furthermore, these same authors in an earlier study detailed a potential osmo-protective role and stress tolerance for these accumulated soluble sugars, as it could also be linked to reduced vegetative growth shown during these stress periods [197]. Salt stress significantly induced a 51% increase in total soluble sugars in the leaves of young olive seedlings (cultivar ‘Chetoui’) transplanted into 10 L pots [82]. Meanwhile, Tester and Davemport [80] stated that salt accumulation in the vacuole stimulates the synthesis of organic solutes, thus balancing the osmotic potential of the cytoplasm with that of the vacuole. The increase in soluble sugars in olive leaves and roots associated with a reduction in starch, has been observed concurrently with the counteraction of their biosynthetic enzymes, offering a crucial mechanism for plant tolerance to various abiotic stresses, including salinity [198,199].
Olive leaves contain a wide variety of carbohydrates, including glucose, mannitol, fructose, arabinose, myo-inositol, xylotol, xylose, galactinol, galactose, sucrose, raffinose, stachyose and many others. Among them, glucose and mannitol are the most abundant [200, 201). Moula et al. [20] found, in their investigation of the quantification of soluble sugars in young olive plants (cultivars ‘Chemlali’ and ‘Koroneiki’) submitted to salt stress, a significant increase in the concentration of mannitol, which was the most abundant sugar, while glucose and fructose showed no variation under this stress. This confirms the proposal by Tattini et al. [94] that mannitol actively contributes to the osmotic adaptation process in olive trees. According to the same authors, the rise in mannitol concentration is not simply a result of salinity but rather functions as a responsive mechanism, while glucose is the main sugar involved in metabolic functions and storage in olive leaves. Gucci and Tottini [51] noticed an accumulation of soluble carbohydrates, especially mannitol in salt-stressed ‘Frantoio’ and ‘Leccino’ olive cultivars, indicating their contribution to osmotic adjustment.
Ayaz et al. [55] assessed mannitol levels and mannitol dehydrogenase (MDH) activity in three olive cultivars (‘Gemlik’, ‘Ayvalık’, and ‘Kilis’) under varying degrees of salt stress, to examine their contribution to osmotic protection. Their results showed a significant increase in mannitol content, together with changes in mannitol dehydrogenase activities in all cultivars as salt stress was raised, clearly revealing a correlation between mannitol levels and enzymatic activities. Similarly, Conde et al. [202] reported that olive trees (cultivar ‘Galega Vulgar’) subjected to salt and drought stress coordinated mannitol levels and mannitol dehydrogenase activity in source tissues. Salinity greatly elevates mannitol and glucose concentrations in olive leaves, while other soluble carbohydrates remain unaffected [94]. Mannitol accumulation occurs earlier than glucose [94]. 14C-labeling pulsechase experiments supported a preferential allocation of carbon to mannitol synthesis in salt-stressed olive leaves, indicating a central role for mannitol in improving salt tolerance [51].
4.3. Cell Wall Modification
Lignin is a biopolymer that has been identified in the xylem and schlerenchyma cell walls of vascular plants, particularly olive [203]. Its main functions are to provide rigidity to cell walls and ensure the impermeability of the tracheary elements, which facilitate the transport of water and solutes via the vascular system [204]. Many authors have evidenced lignin accumulation during saline stress in several species [205,206,207]. Lignification of vascular constituents reduces water permeability in the apoplasm [208]. Sánchez-Aguayo et al. [207] highlighted a decisive role of lignin accumulation in the process of plant adaptation to salt stress.
Concerning olive trees, Ben Abdallah et al. [82] observed a notable increment in leaf lignin content of the ‘Chétoui’ cultivar under water and salt stress conditions, compared to the control. Sofo et al. [88] asserted that the increase in lignification during stress conditions corresponds to a tolerance strategy adopted by olive plants to mechanically strengthen cell walls. They assume that this reinforcement can reduce water loss and cell dehydration by displacing the space occupied by mesophyll water, which, unlike lignified tissue, exchanges readily with the transpiration stream.
In salt stress conditions, olive leaves exhibit overexpression of two protein isoforms (Fra e 12.10) in olive leaves, which show strong sequence similarity to the main allergens of Fraxinus excelsior (European ash) [209]. Ben Abdallah et al. [82] suppose that this overexpression is related to lignin/lignan metabolism, and suggest that lignification may be a tolerance strategy adopted in olive trees to strengthen cell walls and reduce water loss.
4.4. Antioxidant Defense Activity
Salt stress has a significant impact on plants by inducing oxidative stress through the excessive production of reactive oxygen species (ROS), which can damage vital cellular constituents. The olive tree has various mechanisms to prevent these damaging effects, thanks to its antioxidant defense system. This system consists of enzymes such as superoxide dismutase, catalase, ascorbate peroxidase, and guaiacol peroxidase [88], while the non-enzymatic system includes substances with antioxidant properties such as ascorbate, tocopherol, carotenoids and various phenolic compounds [203].
Ayaz et al. [55] reported that salt stress caused increased levels of reactive oxygen species, which require an active antioxidant defense system for tolerance, thus suggesting that antioxidant enzyme activity could serve as a marker of salinity tolerance in olive cultivars. They analyzed the activities of superoxide dismutase (SOD), guaiacol peroxidase (APX), glutathione reductase (GR), peroxidase (POX), and catalase (CAT), in response to higher salinity levels, and observed a notable increase in three olive cultivars ‘Gemlik’, ‘Ayvalık’, and ‘Kilis’. Regni et al. [54] documented a systemic increase of GSH and CAT activities in the leaves of salt-treated olive plants (cultivars ‘Fadak 86’, ‘Royal de Cazorla’, ‘Koroneiki’ and ‘Arbequina’). Similarly, Sofo et al. [88] found higher activities of antioxidant enzymes, namely ascorbate peroxidase (APX) and catalase (CAT), as well as superoxide dismutase (SOD) and peroxidase (POD), lipoxygenase (LOX) activity and malonaldehyde (MDA) concentrations, indicating membrane lipid oxidation, which is linked to photosynthetic apparatus damage. The same authors also detected a lower polyphenol oxidase (PPO) activity to protect against phenol oxidation. These enzymes are widely documented to be activated in response to abiotic oxidative stress by plants [131,184,210,211,212]. They play a central role in protecting chloroplasts from oxidative damage [141). Plants have been shown a tolerance to saline conditions and succeed in thriving even under these severe environments thanks to a significant increase in the production of antioxidant enzymes [182,213,214].
Salt-tolerant species induce the production of antioxidant enzymes, thus improving their ability to eliminate reactive oxygen species. In contrast, salt-sensitive species exhibit a reduction in antioxidant activity [215]. This decline in enzymatic activity caused an accumulation of hydrogen peroxide and malondialdehyde. Ertani et al. [216] explained the accumulation of hydrogen peroxide as the result of reduced APX when exposed to salt stress. Del Buono et al. [132] observed that salt exposure of olive plants (cultivar ‘Arbequina’) caused significant reductions in the activities of antioxidant enzymes, especially ascorbate peroxidase (APX) to a large extent, and others such as superoxide dismutase (SOD), guaiacol peroxidase (GPX) and catalase (CAT). In another similar study, Bano et al. [217] found that salinity strongly reduced SOD, CAT, and peroxidase activities, as well as this effect was cultivar-dependent.
The total antioxidant capacity of olive leaves (cultivar ‘Changlot Real’, ‘Picual’, and ‘Arbequina’) was significantly reduced under salinity stress [218]. The antioxidant activity, in particular that of superoxide dismutase (SOD), increased in response to salt stress, thus reducing oxidative stress in olive trees, notably via non-enzymatic antioxidants such as polyphenols [219]. Total phenols and phenolic compounds are involved in the antioxidant mechanisms developed by the olive tree in response to oxidative stress induced by salt stress [182,195].
In some olive cultivars (‘Gemlik’, ‘Nizip Yaglık’, and ‘Kilis Yaglık’), salt treatment showed no significant impact on polyphenol and protein content, but it reduced total flavonoid content, indicating a consistent response to salt-induced stress [220]. Phenolic compounds play a crucial role in combating abiotic and biotic stress drivers, as both their biosynthesis and accumulation are activated by these factors, although some studies indicate a decrease in their content as stressors increase [221,222]. Flavonoids have been reported to improve the ability of trees to tolerate stress through their influence on physiological performance leading to reduced lipid peroxidation [223]. Demiral et al. [181] noted a change in the leaf antioxidant activity of the ‘Gemlik’ olive cultivar during salt stress, linked to proline and total phenol levels. Proline enhances photosynthetic activity and antioxidant defense enzymes more effectively, as well as acting on the distribution of salt ions in olive trees [195].
4.5. Lipid Peroxidation
Salt stress can also alter the structural properties and integrity of membranes, leading to their deterioration. Many researchers have used lipid peroxidation analysis to assess the damage inflicted on olive trees by salt stress.
Ben Abdallah et al. [82] conducted a study on the effect of salt stress on oxidative damage in Chétoui variety leaves and revealed a 50% rise in lipid peroxidation, as measured using (malondialdehyde) MDA content, compared with the control. They speculated that salt-stressed olive trees might accumulate both reactive oxygen species and electrolytes, leading to heightened oxidative damage. Ayaz et al. [55] have determined MDA levels in the leaves of olive cultivars ‘Gemlik’, ‘Kilis’, and ‘Ayvalık’, and also revealed some spectacular increases in MDA content, with differences observed among varieties. Several other research reports have cited similar trends of significant rises in MDA under salt stress [131,132,179].
Recently, Lima-Cabello et al. [218] have analyzed lipid peroxidation as an indicator of salinity resistance, by measuring the combined levels of Malondialdehyde (MDA) and 4-hydroxynonenal (HNE) [MDA + HNE] in commercial olive trees subjected to saline treatments. The results showed higher levels of MDA and HNE in the ‘Arbequina’, ‘Cobrançosa’, ‘Pico-Limón’ and ‘Cornezuelo’ cultivars compared with controls, while no significant difference was observed for the ‘Changlot Real’ and ‘Picual’ cultivars, renowned for their resistance to salinity. Similarly, Azariadis et al. [224] found that MDA levels increased significantly in salt-sensitive varieties (‘Koroneiki’ and ‘Arvanitoli’), but did not occur in tolerant varieties (‘Lefkolia’ and ‘Gaidourelia’), when all were exposed to saline conditions. Meanwhile, in another study, MDA levels in two olive cultivars (‘Canino’ and ‘Sirole’) under different salt concentrations remained unchanged [225]. Non-significant MDA variations were found in the roots of olive cultivar ‘Leccino’ plants under salt stress [79].
4.6. Adaptations at Protein Level
Different defense mechanisms are triggered when plants are subjected to salt stress, mobilizing a complex set of proteins to mitigate the adverse effects of salinity on growth and development. For olive trees, the crucial role of proteins and their involvement in salt stress tolerance are the subject of intense investigations.
Bashir et al. [225] reported a close relationship between salt stress and protein concentration in ‘Canino’ and ‘Sirole’ olive cultivars. Demir and Cetinkaya [220] stated a slight change in protein content when olive cultivars ‘Gemlik’, ‘Kilis Yaglık’, and ‘Nizip Yaglık’ were exposed to high salinity levels. Valderrama et al. [226] conducted a research study on olive response to salt stress and found a 30-50% increase in protein content. Likewise, Parida et al. [227] affirmed that salinity rise can induce changes in free amino acid levels and reduce total protein content, thereby boosting the activity of acid and alkaline proteases, thus offering resistance to stress conditions. The accumulation of these proteins constitutes a source of nitrogen for osmotic adjustment [228]. Salt stress-induced proteins can be classified into two main groups: salt-stress proteins, which accumulate specifically in response to salt stress, and stress-related proteins, which accumulate in response to various abiotic stresses [229].
Arabinogalactan proteins (AGPs) are key glycoproteins in plant cell walls, with a significant role in growth, development, and responses to environmental stresses [230]. A related claim was reported by Ouyang et al. [231] who consider wall-associated kinases and AGPs to be potential candidates for detecting stress-induced changes in plant physiology. The same authors noted that transcriptional analyses have indicated a repression of cell wall-related gene expression including AGPs and xyloglucan, as a result of salt stress. Similarly, Azariadis et al. [224] reported that immunodetection revealed a weak AGP signal in the ‘Koroneiki’ cultivar, which was associated with irregular cells and intercellular spaces, resulting in the formation of aerenchyma after 45 days of salt treatment. Hydroxyproline-rich cell wall glycoproteins (HRGP), such as arabinogalactans proteins (AGP), are involved in salinity stress [232,233].
Ben Abdallah et al. [82] achieved reproducible 2-DE proteomic maps of ‘Chetoui’ olive leaves for the first time in an original investigation. The analysis of these maps identified 26 differentially expressed protein spots, with the majority corresponding to proteins involved in photosynthesis. Other identified proteins were associated with nitrogen metabolism, protein storage, energy, and other functional categories. Profiles of identified proteins in control and salt samples are provided, revealing significant differences in expression patterns. Under salt stress conditions, several photosynthesis-related proteins showed reduced expression levels compared to the control. These included ribulose-1,5- bisphosphate carboxylase/oxygenase (RuBisCO) large subunit isoforms, oxygen-evolving enhancer protein 1 (OEE1) isoforms, oxygen-evolving enhancer protein 2 (OEE2) isoforms, ferredoxin-NADP reductase (FNR), BSP, glutamine synthetase cytosolic isozymes (GSc), salicylic acid-binding protein (SABP), and carbonic anhydrase isozyme (CA), while others like ATP synthase subunit β, GS nodule isoenzyme (GSn) and Fra e 12.10 allergen isoforms were over-expressed. The down-regulation of RuBisCO isoforms and some other proteins in saline stress conditions indicates a limitation of photosynthetic efficiency due to factors linked to low intercellular CO2 and reduced electron transport activity [234,235]. Moreover, the down-regulation of enzymes involved in CO2 maintenance and salicylic acid signaling, such as CA and SABP respectively, suggests possible mechanisms for reduced stress tolerance [236,237,238,239]. In addition, the overexpression of specific proteins as Fra e 12.10 may reflect an adaptive response, including lignification, to mitigate stress effects [240,241,242]. Alterations in cytoplasmic proteins affect cell viscosity under saline conditions [243].
5. Molecular Mechanisms in Olive for Adaptation to Salt Stress
Plant adaptation or tolerance to salt stress involves a complex process of physiological and biochemical traits and metabolic pathways controlled by a network of genes [244,245,246]. This process begins with detecting stress signals, followed by signal transduction that activates stress-responsive genes, ultimately leading to metabolic adjustments able to counter salinity-induced osmotic stress and other various stressors [247,248].
From recent literature on olive tree responses to salinity, we can note a serious gap concerning the molecular network that governs its adaptation to such severe conditions, compared to the physiological, biochemical, and metabolic mechanisms that have been widely investigated. The lack of molecular research into olive resistance to salt stress has recently been filled by some studies seeking both to identify the genes induced by stress and to analyze their functions.
Bazakos et al. [110] conducted a comparative transcriptomic analysis of two olive cultivars under salt stress conditions, which is widely acknowledged as a seminal study in understanding the molecular response of olive trees to salt stress. They found that in cultivar. Kalamon, 159 transcripts consistently showed up-regulation during stress, with 50 others exhibiting delayed up-regulation, but all 209 were down-regulated post-stress. In contrast, cultivar ‘Chondrolia Chalkidikis’ displayed limited transcriptional activation (20 transcripts), leading the authors to suggest that this reduced transcriptional response might be partly responsible for the sensitivity of cultivar ‘Chondrolia Chalkidikis’ compared to cultivar ‘Kalamon’ in terms of gene expression. Seven transcripts in cultivar ‘Kalamon’ were recognized by these authors as salt-specific, namely putative small glutamine-rich tetratricopeptide repeat-containing protein, 40S ribosomal protein, NAD+ ADP-ribosyltransferase, annexin A4, xyloglucan endotransglycosylase, UDP-galactose epimerase, and stress-induced protein. JEREF and bZIP transcription factors have been identified as key regulators of the response of olive trees to salt stress, with differences in expression between the two cultivars indicating their importance in salt tolerance. Zhang et al. [249] reported the involvement of JERF in the response to abiotic stress, including salinity, by regulating the expression of stress-responsive and ABA biosynthesis-related genes. Similarly, Yoshida et al. [250] showed that AREB1, a group A bZIP, is crucial in ABA signaling in response to drought stress.
In further research, Bazakos et al. [251] used next-generation sequencing to explore the molecular basis of the salt stress response in the olive cultivar ‘Kalamon’, focusing on leaves and roots. Transcriptomic analysis disclosed 24 genes differentially expressed in roots (9 down-regulated and 15 up-regulated) and 70 in leaves (14 down- and 56 up-regulated), giving tissue-specific responses to salt stress. The differentially expressed transcripts are consistent with findings in chickpeas and maize, suggesting common stress response pathways to plant species [252,253]. In addition, the authors stated that BlastX analysis identified 14 transcripts associated with cell wall hydrolases (8 down-regulated in roots and only 1 in leaves), which could have an impact on wall elasticity under salt stress. A reduction in hydrolase activity may decrease wall elasticity, making plants more rigid and tolerant to salt stress [252]. In addition, Bazakos et al. [251] noted that 19 clusters encode cytochrome P450s showing significant differential expression in leaves and roots. This gene family is involved in lignin biosynthesis, known for its role in the response to abiotic stress [254].
Rossi et al. [78] assessed the expression levels of genes implicated in the phenylpropanoid metabolic pathway, compounds known to be specialized in responses to environmental stimuli, in ‘Leccino’ (salt-sensitive) and ‘Frantoio’ (salt-tolerant) cultivar olive plants grown in a phytotron chamber and treated with NaCl. They highlighted in this study the salt stress impact on gene expression along phenolic pathways in olive trees, revealing a variable response depending on the organ, genotype, and specific enzyme involved. Phenylalanine-Ammonia-Lyase (PAL), the primary enzyme converting phenylalanine into cinnamic acid, is up-regulated in response to salinity, varying by organ and genotype. Expression of Cinnamate-4-Hydroxylase (C4H), involved in the conversion of cinnamic acid to p-coumaric acid, remained stable in all treatments at most organs, except for significant up-regulation in ‘Leccino’ roots under moderate salt stress. 4-Coumarate: CoA ligase (4CL), responsible for 4-coumarate activation, showed higher expression in ‘Leccino’ aerial organs, with up-regulation in leaves. Chalcone Synthase (CHS) and Chalcone Isomerase (CHI), key enzymes in flavonoid biosynthesis, exhibited low expression levels but were up-regulated in new leaves of both cultivars under NaCl stress. The same authors observed that expression of the Na+/H+ Exchanger (NHX) gene, a recognized molecular marker of salt stress in other plant species [255], was significantly increased in all ‘Leccino’ organs in response to salt treatment, while it remained insignificant in ‘Frantoio’, indicating a differential perception of salt stress between the cultivars.
Said [246] focused his investigation more specifically on the change in expression of the AtTPS1 gene, which codes for trehalose-6-phosphate synthase (TPS), in salt-stressed plants. This study was conducted on two olive cultivars, ‘Pecaul’ (salt-tolerant) and ‘Aggizi Shame’ (moderately salt-tolerant), which were treated with validamycin A and grown under saline conditions. Results revealed successful amplification of AtTPS1 in both cultivars, with an additional fragment at 210 bp observed particularly in the salt-tolerant cultivar (‘Pecual’). Trehalose synthesis relies on the enzymes trehalose-6-phosphate synthase (TPS) and trehalose-6-phosphate phosphatase (TPP). The increase in cytoplasmic trehalose induced by validamycin A inhibits TPP activity, increasing trehalose-6-phosphate (T6P) levels [256]. Accordingly, the results of Said [246] suggest the involvement of olive TPS in salt stress response with validamycin A in both cultivars. AtTPS1 and AtTPP genes have previously been discovered in Arabidopsis thaliana, emphasizing their relevance to plant stress responses [257,258,259]. Penna [260] asserted the potential strategy of modulating trehalose levels, a key player in the stress response, either through inhibition of trehalase or enhancement of its biosynthesis.
Mousavi et al. [261] analyzed the differential regulation of candidate genes putatively for the salt stress response across four olive cultivars ‘(Koroneiki’, ‘Picual’, ‘Royal de Cazorla’, and ‘Fadak86’). They revealed that only OeNHX7, OeP5CS, OeRD19A, and OePetD exhibited upregulation in tolerant cultivars (‘Royal’ and ‘Fadak86’), indicating their key role in the activation of a salt tolerance mechanism. Up-regulation of NHX, as observed with OeNHX7, was related to an enhanced accumulation of Na+ in the vacuole, hence contributing to mitigate salt stress [128,255]. P5CS gene expression has been shown to improve salt tolerance through proline accumulation, which regulates Na+ accumulation in leaves [262,263,264]. The cysteine protease genes RD21A and RD19A are involved in the programmed cell death pathway during stress [265]. The up-regulation of OeRD19A supports its implication in improving salt tolerance in olive trees [261]. The chloroplast gene PetD (CytB6), a component of the plastoquinone-plastocyanin reductase, plays a critical role in electron transport and ATP production, thus offering a defense against salinity-induced oxidative damage [261,266,267].
Recently, Sodini et al. [79] delved into the molecular mechanisms underlying the response of ‘Leccino’ olive tree (salt-sensitive) roots to NaCl stress, focusing on the expression of genes associated with proton pumps and Na+/H+ exchangers. They specifically examined genes such as PATPase, V-type ATPase sub E, SOS1, and NHX, designing primers based on the Olea europaea genome and genes from other tree species. They revealed that salt treatment resulted in a decrease in the expression of P-ATPase 1 and SOS1 after 24 hours, followed by NHX and V-ATPase sub E after 48 hours, and ultimately P-ATPase 8 within 7 days. Membrane H+-ATPase genes are typically under-expressed in response to salinity [268]. P-type H+-ATPase has housekeeping functions, aiding turgor pressure and cell wall extension [269]. SOS1 gene in roots functions in Na+ exclusion and loading into the xylem, potentially leading to sodium accumulation in leaves [80,128,270]. V-ATPase subunits and NHX genes are involved in the mechanisms of root Na+ storage, and their downregulation aligns with the observed lack of sodium accumulation in the roots [271].
6. Factors Affecting Salinity Tolerance
The olive tree is widely recognized for its resilience and ability to tolerate severe environments [82,272,273], including those with high levels of salinity, thanks to a series of adaptive mechanisms [84,86,217,219]. Nevertheless, the extent of its tolerance to saline stress is also conditioned by many other factors. Studies have revealed significant variation in the response of olive trees to stress depending on the cultivar, as some cultivars such as ‘Leccino’, ‘Arvanitoli’, ‘Arbequina’ displayed salt sensitivity, while others like ‘Frantoio’, ‘Lefkolia’, ‘Gaidourelia’, ‘Manzanillo’, and ‘Hojiblanc’a showed more resilience [78,224]. The specific genetic characteristics of each cultivar define its salt tolerance, thereby affecting ion absorption/exclusion regulation, osmolyte accumulation, resistance to oxidative stress, and the activation of metabolic pathways [110,246,261]. Furthermore, prior investigations have demonstrated that both young olive trees and organs are more susceptible to salt stress compared to their older counterparts [135,274,275]. The effects of salt stress on olive trees are not limited to cultivar tolerance. The physiological, biochemical, and molecular responses of the olive tree are in part dependent on the concentration and nature of salts and nutrients available in the soil and irrigation water. In fact, high salt concentrations exacerbate the damaging actions of salt stress on olive trees [62,225].
Several other environmental factors are critical in regulating the response of olive trees to saline conditions. Soil composition and pH, as well as solute and nutrient content, all influence ion exchange [135]. In addition, water availability has proven to be essential for the olive tree in managing salt in the soil; unbalanced conditions, particularly a water shortage, may exacerbate water stress and increase salt concentration [82], which further emphasizes the importance of finding the right balance to support the resistance of the olive tree to salt stress. High temperatures can also aggravate the effects of salt stress by accelerating transpiration [276]. Hence, we also hypothesize that cultivation and management practices, like irrigation methods and fertilization, may influence the salinity tolerance of the olive tree.
7. Conclusions
Olive cultivation has experienced rapid expansion worldwide, although its primary production remains concentrated in the regions of the Mediterranean and the Middle East. However, these regions are facing major challenges associated with demographic pressure and climate change, significantly impacting water availability and quality. This situation is giving rise to the growing risk of soil and water salinization, particularly threatening agricultural production, including that of olives and olive oil.
The literature on the response of the olive tree (Olea europaea) to saline conditions has revealed multiple tolerance mechanisms to cope with these stressful environments. The olive tree may undergo structural changes in the root to favour ion exclusion from the shoot. It can remove senescent leaves where there is accumulation of toxic ions and adjust its morphology to limit water loss caused by salt stress, in particular by reducing growth, especially of the aerial part, and modifying leaf anatomy. In addition, stomatal conductance is increased by salinity, leading to a decrease in transpiration and photosynthesis and an increase in water use efficiency. In fact, the decrease in photosynthesis is mainly due to stomatal closure, since most studies revealed that PSII is not very susceptible to saline ions. Moreover, to resist and adapt to salinity, the olive tree relies on biochemical mechanisms that include the accumulation of osmolytes like sugars and amino acids, as well as the regulation of ionic flux, to maintain osmotic and ionic balance. Additionally, research has unveiled the synthesis of specific metabolites and enzymes crucial for protecting plant cells against oxidative damage induced by salt stress. While Molecular exploration of the response of olive trees to salt stress has lagged behind the other aspects, however, recently considerable attention has been devoted to understanding the expression and functions of involved genes, especially ion transporters and signaling proteins that regulate olive responses to saline conditions.
Profiling genes contributing to physiological and biochemical processes related to salt tolerance in olive trees could enable the identification of specific markers. Understanding all these mechanisms is essential for the development of selection and breeding strategies to improve the ability of olive trees to grow in saline environments, thus ensuring their sustainability.
Author Contributions
Conceptualization, M.E.Y. and M.d.P.C.; resources, M.d.P.C. and M.E.Y.; data curation, M.E.Y. and M.d.P.C.; writing—original draft preparation, M.d.P.C. and M.E.Y.; writing—review and editing, M.E.Y. and M.d.P.C.; visualization, M.d.P.C. and M.E.Y.; supervision M.E.Y. and M.d.P.C.; project administration, M.E.Y.; funding acquisition, M.d.P.C.; All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the R&D project of Viveros Jarico S.L. funding by the IDEA agency of the Junta de Andalucía. grant number 805 (University of Jaén) Improvement of salinity tolerance in olive by pretreatment with growth regulators.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mukhopadhyay, R.; Sarkar, B.; Jat, H.S.; Sharma, P.C.; Bolan, N.S. Soil salinity under climate change: Challenges for sustainable agriculture and food security. J. Environ. Manage. 2021, 280, 111736. [CrossRef]
- Ludwig, M.; Wilmes, P.; Schrader, S. Measuring soil sustainability via soil resilience. Sci. Total Environ. 2018, 626, 1484–1493. [CrossRef]
- Yuvaraj, M.; Bose, K.S.C.; Elavarasi, P.; Tawfik, E. Soil salinity and its management. In Soil Moisture Importance. Meena, R.S., Datta, R., Eds. IntechOpen, 2021; pp. 109–119. https://www.intechopen.com/chapters/72995.
- El hasini, S.; Halima, I.O.; Azzouzi, M.E.; Douaik, A.; Azim, K.; Zouahri, A. Organic and inorganic remediation of soils affected by salinity in the sebkha of sed el mesjoune – marrakech (Morocco). Soil Tillage Res. 2019, 193, 153–160. [CrossRef]
- Mousavi, S.; Regni, L.; Bocchini, M.; Mariotti, R.; Cultrera, N.G.M.; Mancuso, S.; Googlani, J.; Chakerolhosseini, M.R.; Guerrero, C.; Albertini, E.; Baldoni, L.; Proietti, P. Physiological, epigenetic and genetic regulation in some olive cultivars under salt stress. Sci. Rep. 2019, 9, 1093. [CrossRef]
- Field, C.B.; Barros, V.R.; Mach, K.J.; Mastrandrea, M.D. Technical summary. In In: Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., Girma, B., Kissel, E.S., Levy, A.N., MacCracken, S., Mastrandrea, P.R., White, L.L., Eds. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, pp. 35-94.
- Chartzoulakis, K. Salinity and olive: Growth, salt tolerance, photosynthesis and yield. Agric. Water Manag. 2005, 78, 108–121. [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Álvarez, S.; Hernández, J.A.; Sánchez-Blanco, M.J. The use of reclaimed water is a viable and safe strategy for the irrigation of myrtle plants in a scenario of climate change. Water Supply. 2019, 19, 1741-1747. [CrossRef]
- Meena, M.D.; Yadav, R.K.; Narjary, B.; Gajender, Y.; Jat, H.S.; Sheoran, P.; Meena, M.K.; Antil, R.S.; Meena, B.L.; Singh, H.V.; Meena, V.S.; Rai, P.K.; Ghosh, A.; Moharana, P.C. Municipal solid waste (MSW): Strategies to improve salt affected soil sustainability: A review. Waste Manage. 2019. 84, 38-53. [CrossRef]
- Romero-Trigueros, C.; Vivaldi, G.A.; Nicolás, E.N.; Paduano, A.; Salcedo, F.P.; Camposeo, S. Ripening Indices, Olive Yield and Oil Quality in Response to Irrigation with Saline Reclaimed Water and Deficit Strategies. Front. Plant Sci. 2019, 10, 1243. [CrossRef]
- Vivaldi, G.A.; Camposeo, S.; Lopriore, G.; Romero-Trigueros, C.; Salcedo, F.P. Using saline reclaimed water on almond grown in Mediterranean conditions: Deficit irrigation strategies and salinity effects. Water Supply. 2019, 19, 1413–142. [CrossRef]
- Ansari, F.A.; Ahmad, I.; Pichtel, J. Growth stimulation and alleviation of salinity stress to wheat by the biofilm forming Bacillus pumilus strain FAB10. Appl. Soil Ecol. 2019, 143, 45–54. [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A. K.; Srivastava, S.; Verma, J. P. Plant growth-promoting bacteria: Biological tools for the mitigation of salinity stress in plants. Front. Microbiol. 2020, 11, 1216. [CrossRef]
- Galicia-Campos, E.; Garcia-Villaraco, A.; Montero-Plamero, M.B.; Gutierrez-Mañero, F.J.; Ramos-Solano, B. Modulation of photosynthesis and ROS scavenging response by benficial bacteria in Olea europaea Plantlets under Salt Stress Conditions. Plants. 2022, 11, 2748. [CrossRef]
- Bhise, K.K.;Bhagwat, P.K.; Dandge, P.B. Synergistic effect of Chryseobacterium gleum sp. SUK with ACC deaminase activity to alleviate salt stress and promote plant growth in Triticum aestivum L. 3 Biotech. 2017, 7, 105. [CrossRef]
- Zamanzadeh-Nasrabadi, S.M.; Mohammadiapanah, F.; Sarikhan, S.; Shariati, V.; Saghafi, K.; Hosseini-Mazinani, M. Comprehensive genome analysis of Pseudomonas sp. SWRIQ11, a new plant growth-promoting bacterium that alleviates salinity stress in olive. 3 Biotech. 2023, 13, 347. [CrossRef]
- Shekafandeh, A.; Sirooeenejad, S.; Alsmoushtaghi, E. Influence of gibberellin on increasing of sodium chloride tolerance via some morpho-physiological changes in two olive cultivars. Agric. Conspec. Sci. 2017, 82, 367-373. https://acs.agr.hr/acs/index.php/acs/article/view/1225.
- Metheni, K.; Abdallah, M.B.; Nouairi, I; Smaoui, A.; Ammar, B.; Zarrouk, M.; Youssef, N.B. Salicylic acid and calcium pretreatments alleviate the toxic effect of salinity in the Oueslati olive variety. Sci. Hortic. 2018, 233, 349-358. [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [CrossRef]
- Moula, I.; Boussadia, O.; Koubouris, G.; Hassine, MB; Boussetta, W.; van Labeke, MC; Braham, M. Ecophysiological and biochemical aspects of olive (Olea europaea L.) in response to salt stress and gibberellic acid-induced relief. S. Africa. J.Bot. 2020, 132, 38–44. [CrossRef]
- Cordovilla, M.P.; Aparicio, C.; Melendo, M.; Bueno, M. Exogenous application of indol-3-acetic and salicylic acid improves tolerance to salt stress in olive plantlets (Olea europaea L. cultivar Picual) in growth chamber environments. Agronomy, 2023, 13, 647. [CrossRef]
- Li, J.; Liu, J.; Zhu, T., Zhao, C.; Li, L.; Chen, M. The Role of Melatonin in Salt Stress Responses. Int. J. Mol. Sci. 2019, 20, 1735. [CrossRef]
- Zhan, H.; Nie, X.; Zhang, T.; Li, S.; Wang, X.; Du, X.; Tong, W.; Song W. Melatonin: A small molecule but important for salt stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 709. [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Hoveizeh N.F.; Gholami, R.; Abdelrahman, M.; Phan Tran, L.S. Exogenous melatonin mitigates salinity-induced damage in olive seedlings by modulating ion homeostasis, antioxidant defense, and phytohormone balance. Physiol. Plant. 2021, 173, 1682-1694. [CrossRef]
- Khan, T.A., Saleem, M., Fariduddin, Q. Recent advances and mechanistic insights on Melatonin-mediated salt stress signaling in plants. Plant Physiol. Biochem. 2022, 188, 97-107. [CrossRef]
- Fan, Z.; He, F.; Wu, T.; Li, B.; Xiao, J.; Wang, Z. Breaking the salty spell: Understanding action mechanism of melatonin and beneficial microbes as nature’s solution for mitigating salt stress in soybean. S. Afr. J. Bot. 2023, 161, 555-567. [CrossRef]
- Ahmed, M.; Decsi, K.; Tóth, Z. Different tactics of synthesized zinc oxide nanoparticles, homeostasis ions, and phytohormones as regulators and adaptatively parameters to alleviate the adverse effects of salinity stress on plants. Life 2022, 13, 73. [CrossRef]
- Sarraf, M.; Vishwakarma, K.; Kumar, V.; Arif, N.; Das, S.; Johnson, R. Metal/Metalloid-Based Nanomaterials for Plant Abiotic Stress Tolerance: An Overview of the Mechanisms. Plants 2022, 11, 316. [CrossRef]
- Ahmed, M.; Tóth, Z.; Decsi, K. The Impact of Salinity on Crop Yields and the Confrontational Behavior of Transcriptional Regulators, Nanoparticles, and Antioxidant Defensive Mechanisms under Stressful Conditions: A Review. Int. J. Mol. Sci. 2024, 25, 2654. [CrossRef]
- Al-Saif, A.M.; Sas-Paszt, L.; Mosa, W.F.A. Olive performance under the soil application of humic acid and the spraying of titanium and zinc nanoparticles under soil salinity stress. Horticulturae 2024, 10, 295. [CrossRef]
- Kumar, D.; Singh, R.; Upadhyay, S.K.; Verma, K.K.; Tripathi, R.M.; Liu, H.; Dhankher, O.P.; Tripathi, R.D.; Sahi, S.V.; Seth, C.S. Review on interactions between nanomaterials and phytohormones: Novel perspectives and opportunities for mitigating environmental challenges. Plant Sci. 2024, 340, 111964. [CrossRef]
- Bernard, G.; Khadari, B.; Naavascues, M.; Fernandez-Mazuecos, M.; El Bakkali, A.: Arrigo, N.; Baali-Cherif, D.; Caraffa, B.B.; Santoni, S.; Vargas, P.; Savolainen, V. The complex history of the olive tree: From Late Quaternary diversification of Mediterranean lineages to primary domestication in the northern Levant. Proc. Biol. Sci. 2013, 280, 1756. [CrossRef]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th ed.; Oxford University Pres: Oxford, UK, 2012, pp. 193. [CrossRef]
- Vossen, P. Olive oil: History, production, and characteristics of the world’s classic oils. Hortscience 2007, 42, 1093–1100. [CrossRef]
- Schäfer-Schuchardt, H. Expansión cultural y artística. In VVAA, Enciclopedia Mundial del Olivo; Plaza and Jané: Barcelona, Spain, 1996, pp. 21-26.
- Besnard, G.; Terral, J.F.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ana. Bot. 2018, 121, 385-403. [CrossRef]
- IOC. International Olive Council. 2018. Available online: https://www.internationaloliveoil.org (accesed on 11 April 2024).
- Vilar, J.; Pereira, J.E.; Urieta, D.; Menor, A.; Caño, S., Barreal, J.; Velasco, M.M.; Puentes, R. La olivicultura Internacional. Difusión histórica, análisis estratégico y visión descriptiva. Fundación Caja Rural: Jaén, Spain, 2018; pp. 153.
- IOC. International Olive Council. 2003. Available online: https://www.internationaloliveoil.org (accesed on 11 April 2024).
- IOC. International Olive Council. 2022. Available online: https://www.internationaloliveoil.org (accesed on 11 April 2024).
- Issaoui, M.; Flamini, G.; Brahmi, F.; Dabbou, S.; Ben Hassine, K.; Taamali, A.; Chehab, H.; Ellouz, M.; Zarrouk, M.; Hammami, M. Effect of the growing area conditions on differentiation between Chemlali and Chétoui olive oils. Food Chem. 2010, 119, 220-225. [CrossRef]
- Tous, J., Romero, A., Hermoso, J.F. New trends in olive orchard design for continuous mechanical harvesting. Adv. Hortic. Sci. 2010, 24, 43-52.
- Loumou, A.; Giourga, C. Olive groves: The life and identity of the Mediterranean. Agric. Human Values 2003, 20, 87-95.
- Pessoa, H.R.; Zago, L.; Chaves C.; Ferraz da Costa, D.C. Modulation of biomarkers associated with risk of cancer in humans by olive oil intake: A systematic review. J. Funct. Food. 2022, 98, 105275. [CrossRef]
- Fazlollahi, A.; Motlagh Asghari, K.; Aslan, C.; Noori, M.; Nejadghaderi, S.A.; Araj-Khodaei, M.; Sullman, M.J.M.; Karamzad, N., Kolahi; A.A.; Safiri S. Front. Nutr. 2023, 10, 1218538. [CrossRef]
- Chrysant, S.G.; Chrysant, G.S. Olive Oil Consumption and Cardiovascular Protection: Mechanism of Action. Cardiol. Rev. 2024, 32, 57-61. [CrossRef]
- Therios, I.N. Olives; CABI: Oxfordshire, UK, 2009. Pp. 409.
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651-681. [CrossRef]
- Shabala S.; Wu, H.; Bose, J. Salt stress sensing and early signalling events in plant roots: Current knowlegde and hypothesis. Plant Sci. 2015, 241, 109–199. [CrossRef]
- Rugini, E.; Fedeli, E. Olive as an oilseed crop. In: Legumes and Oilseed Crops; Bajaj YPS ed.; Sorubger-Verlag: Berlín, Germany, 1990; pp 593-641.
- Gucci, R.; Tattini, M. Salinity tolerance in olive. Hortic. Rev. 1997, 21, 177-213.
- Chartzoulakis, K.; Loupassaki, M.; Bertaki, M.; Androulakis, I. Effects of NaCl salinity on growth, ion content and CO2 assimilation rate of six olive cultivars. Sci. Hortic. 2002, 96, 235–247. [CrossRef]
- Aparicio, C.; Urrestarazu, M.; Cordovilla, M.P. Comparative physiological analysis of salinity effects in six olive genotypes. HortScience 2014, 49, 901-904. [CrossRef]
- Regni, L.; Del Pino, AM; Mousavi, S.; Palmerini, California; Baldoni, L.; Mariotti, R.; Mairech, H.; Gardi, T.; D’Amato, R.; Proietti, P. Behaviour of four olive cultivars during salt stress. Front. Plant Sci. 2019, 10, 867. [CrossRef]
- Ayaz, M.; Varol, N.; Yolcu, S.; Pelvan, A.; Kaya,Ü.; Aydogdu, E.; Bor, M.; Ördemir, F.; Türkan, İ. Three (Turkish) olive cultivars display contrasting salt stress-coping mechanisms under high salinity. Trees. 2021. 35, 1283-1298. [CrossRef]
- Dbara, S.; Abboud, S.; Bchir, A. Foliar potassium nitrate spray induces changes in potassium-sodium balance and biochemical mechanisms in olive (Olea europaea L. cv Chemlali) plants submitted to salt stress. J. Hortic. Postharvest Res. 2022, 5, 309-322. [CrossRef]
- Boussadia, O.; Zgallai, H.; Mzid, N.; Zaabar, R.; Braham, M.; Doupis, G.; Koubouris, G. Physiological responses of two olive cultivars to salt stress. Plants 2023, 12, 1926. [CrossRef]
- Tattini, M. Ionic relations of aeroponically-grown olive plants during salt stress. Plant Soil. 1994. 161, 251-256. https://www.webofscience.com/wos/alldb/full-record/WOS:A1994NY60100011.
- Perica, S.; Goreta, S.; Selak, G.V. Growth, biomass allocation and leaf ion concentration of seven olive (Olea europaea L) cultivars under increased salinity. Sci. Hortic. 2008, 117, 123-129. [CrossRef]
- Bader, B.; Aissaoui, F.; Kmicha, I.; Ben Salem, A.; Chehab, H.; Gargouri, K.; Boujnah, D.; Chaieb, M. Effects of salinity stress water desalination, olive tree (Olea europaea L. cvs ’Picholine’, ’Meski’ and ’Ascolana’) growth and ion accumulation. Desalination. 2015, 364, 46-52. [CrossRef]
- Kchaou, H.; Larbi, A.; Gargouri, K.; Chaieb, M.; Morales, F.; Msallem, M. Assessment of tolerance to NaCl salinity of five olive cultivars, based on growth characteristics and Na+ and Cl- exclusion mechanisms. Sci. Hortic. 2010, 124, 306-315. [CrossRef]
- Palm, E.R.; Salzano, A.M.; Vergine, M., Negro, W.G.N.; Sabbatini, L.; Balestrini, R.; De Pinto, M.C.; Fortunato, S.; Gohari, G.; Mancuso, S.; Luvisi, A.; De Bellis, L.; Scaloni, A.; Vita, F. Response to salinity stress in four Olea europaea L. genotypes: A multidisciplinary approach. Environ. Exp. Bot. 2024, 218, 105586. [CrossRef]
- Soda, N.; Ephrath, J.E.; Dag, A.; Beiersdorf, I.; Presnov, E.; Yermiyahu, U.; Ben-Gal, A. Root growth dynamics of olive (Olea europaea L.) affected by irrigation induced salinity. Plant Soil 2017, 411, 305–318. [CrossRef]
- Tan, J.; Ben-Gal, A.; Shtein, I.; Bustan, A.; Dag, A.; Erel, R. Root structural plasticity enhances salt tolerance in mature olives. Environ. Exp. Bot. 2020. 179, 104224. [CrossRef]
- Horie, T.; Costa, A.; Kim, T.H.; Han, M.J.; Horie, R.; Leung, H.Y.; Miyao, A.; Hirochika, H.; An, G.; Schroeder, J.I. Rice OsHKT2;1 transporter mediates large Na+ influx component into K+-starved roots for growth. EMBO J. 2007, 26, 3003–3014. [CrossRef]
- Almeida, P.; Katschnig, D.; de Boer, A.H. HKT transporters—State of the art. Int. J. Mol. Sci. 2013, 14, 20359–20385. [CrossRef]
- Hamamoto, S.; Horie, T.; Hauser, U.; Deinlein, U.; Schroeder, J.I.; Uozumi, N. HKT transporters mediate salt stress resistance in plants: From structure and function to the field. Curr. Opin. Biotechnol. 2015, 32, 113–120. [CrossRef]
- Hao, H.; Wrang, Y.; Yan, V.; Liu, Y.; Wang, J.; Chen, S. A Review on Plant Responses to Salt Stress and Their Mechanisms of Salt Resistance. Horticulturae 2021, 7, 132. [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo,W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371-379. [CrossRef]
- Connor, J.D.; Fereres, E. The physiology of adaptation and yield in olive. Hort. Rev. 2005, 31, 155–229.
- Chartzoulaki, K.S. Salinity and olive trees: Growth, salt tolerance, photosynthesis and yield. Agric. Water Manage. 2005, 78, 108–121. [CrossRef]
- Møller, I.S.; Gilliham, M.; Jha, D.; Mayo, G.M.; Roy, S.J.; Coates, J.C.; Haseloff, J.; Tester, M. Shoot Na+ exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na+ transport in Arabidopsis. Plant Cell. 2009, 21, 63–78. [CrossRef]
- Zhang, J.L.; Flowers, T.J.; Wang, S.M. Mechanisms of sodium uptake by roots of higher plants. Plant Soil 2010, 326, 45–60. [CrossRef]
- Fernández, J. Understanding olive adaptation to abiotic stresses as a tool to increase crop performance. Environ. Exp. Bot. 2014, 103, 158-179. [CrossRef]
- Rossi, L.; Francini, A.; Minnocci, A.; Sebastiani, L. Salt stress modifies apoplastic barriers in olive (Olea europaea L.): A comparison between a salt-tolerant and a saltsensitive cultivar. Sci. Hortic. 2015, 192, 38–46. [CrossRef]
- Tattini, M.; Lombardi, L.; Gucci, R. The effect of NaCl stress and relief on gas exchange properties of two olive cultivars differing in tolerance to salinity. Plant Soil 1997, 197, 87-93. [CrossRef]
- Rossi, L.; Francini, A.; Minnocci, A.; Sebastini, L. Salt stress modifies apoplastic barriers in olive (Olea europaea L.): A comparison between a salt-tolerant and a salt-sensitive cultivar. Sci. Hortic. 2015, 192, 38-46. [CrossRef]
- Rossi, L.; Borghi, M.; Francini, A.; Lin, X.; Xie, D.Y.; Sebastiani, L. Salt stress induces differential regulation of the phenylpropanoid pathway in Olea europaea cultivars Frantoio (salt-tolerant) and Leccino (salt-sensitive). J. Plant Physiol. 2016, 204, 8–15. [CrossRef]
- Sodini, M.; Francini, A.; Sebastini, L. Early salinity response in root of salt sensitive Olea europaea L. cv Leccino. Plant Stress 2023, 10, 100264. [CrossRef]
- Tester, M.; Davemport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503-527. [CrossRef]
- Hayat, S.; Hayat, Q._; Alyemeni, M. N.; Wani, A. S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [CrossRef]
- Ben Abdallah, M.; Trupiano, D.; Polzella, A.; De Zio, E.; Sassi, M.; Scaloni, A.; Zarrouk, M.; Youssef, N.B., Scippa, G.S. Unraveling physiological, biochemical and molecular mechanisms involved in olive (Olea europaea L. cv. Chétoui) tolerance to drought and salt stresses. J. Plant Physiol. 2018, 220, 83-95. [CrossRef]
- Munns, R.; Passioura, J.B.; Colmer, T.D.; Byrt, C.S. Osmotic adjustment and energy limitations to plant growth in saline soil. New Phytol. 2020, 225, 1091–1096. [CrossRef]
- Ben Rouina, B., Ben Ahmed, C., Athar, H.R., Boukhris, M. Water relations, proline accumulation and photosynthetic activity in olive tree (Olea europaea L. cv. Chemlali) in response to salt stress. Pak. J. Bot. 2006, 38, 1397–1406. https://www.webofscience.com/wos/alldb/full-record/WOS:000203707400007.
- Ben Ahmed, C.; Ben Rouina, B.; Boukhris, M. Comparative responses of three Tunisian olive cultivars to salt stress: From physiology to management practice. In: Agricultural Irrigation Research Progress; Alonso, D., Iglesias, H.J. Eds.; Nova Science Publishers: New York, USA; 2008; pp. 109–129.
- Ben Ahmed, C.; Ben Rouina, B.; Boukhris, M. Changes in water relations, photosynthetic activity and proline accumulation in one-year-old olive trees (Olea europaea L. cv. Chemlali) in response to NaCl salinity. Acta Physiol. Plant. 2008, 154, 553–560. [CrossRef]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2009, 67, 345–352. [CrossRef]
- Sofo, A.; Dichio, B.; Xiloyannis, C.; Masia, A. Lipoxygenase activity and proline accumulation in leaves and roots of olive tree in response to drought stress. Physiol. Plant. 2004, 121, 58–65. [CrossRef]
- Ayala-Astorga, G.I.; Alcaraz-Meléndez, L. Salinity effects on protein content, lipid peroxidation, pigments, and proline in Paulownia imperialis (Siebold & Zuccarini) and Paulownia fortunei (Seemann & Hemsley) grown in vitro. Electron. J. Biotechnol. 2010, 13,13-14. [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Redox Signal. 2013, 19, 998-1011. [CrossRef]
- Goel, P.; Singh, A.K. Abiotic stresses downregulate key genes involved in nitrogen uptake and assimilation in Brassica juncea L. PLoS ONE 2015. 10, e0143645. [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Nitrogen modifies NaCl toxicity in eggplant seedlings: Assessment of chlorophyll a fluorescence, antioxidative response and proline metabolism. Biocatal. Agric. Biotechnol. 2016, 7, 76-86. [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89-97. [CrossRef]
- Tattini, M.; Gucci, R.; Romani, A.; Baldi, A.; Everard, J.D. Changes in non-structural carbohydrates in olive (Olea europaea L.) leaves during root zone salinity stress. Physiol. Plant. 1996, 98, 117–124. [CrossRef]
- Roussos, P.A.; Assimakopoulou, A.; Nikoloudi, A.; Salmas, I.; Nifakos, K.; Kalogeropoulos, P.; Kostelenos, G. Intra- and inter-cultivar impacts of salinity stress on leaf photosynthetic performance, carbohydrates and nutrient content of nine indigenous Greek olive cultivars. Acta Physiol. Plant. 2017, 39, 136. [CrossRef]
- Gucci, R.; Lombardini, L.; Tattini, M. Analysis of leaf water relations in leaves of two olive cultivars (Olea europaea) differing in tolerance to salinity. Tree Physiol. 1997, 17, 13-21. [CrossRef]
- Chartzoulakis, K.; Psarras, G.; Vemmos, S.; Loupassaki, M.; Bertaki, M. Response of two olive cultivars to salt stress and potassium supplement. J. Plant Nutr. 2006, 29, 2063-2078. [CrossRef]
- Roussos, P.A.; Gasparatos, D.; Kechrologou, K.; Katsenos, P.; Bouchagier, P. Impact of organic fertilization on soil properties, plant physiology and yield in two newly planted olive (Olea europaea L.) cultivars under Mediterranean conditions. Sci. Hortic. 2017, 220, 11-19. [CrossRef]
- Gill, S.S.; Tuteja, N. Polyamines and abiotic stress tolerance in plants. Plant Signal. Behav. 2010, 5, 26–33. [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and anti-oxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [CrossRef]
- Kaya, C.; Sonmez, O.; Aydemir, S.; Ashraf, M.; Dikilitas, M. Exogenous application of mannitol and thiourea regulates plant growth and oxidative stress responses in salt-stressed maize (Zea mays L.). J. Plant Interact. 2013, 8, 234–241. [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response antioxidant as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [CrossRef]
- Goreta, S.; Bučević-Popović, V.; Pavela-Vrančić, M.; Perica, S. Salinity-induced changes in growth, superoxide dismutase activity, and ion content of two olive cultivars. J. Plant Nutr. 2007, 170, 398-403. [CrossRef]
- Li, N.; Wang, X.; Ma, B.; Du, C.; Zheng, L.; Wang, Y. Expression of a Na+/H+ antiporter Rtnhx1 from a recreto-halophyte Reaumuria Trigyna improved salt tolerance of transgenic Arabidopsis Thaliana. J. Plant Physiol. 2017, 218, 109–120. [CrossRef]
- Azimi, M.; Khoshzaman, T.; Taheri, M.; Dadras, A. Evaluation of salinity tolerance of three olive (Olea europaea L.) cultivars. J. Cent. Eur. Agric. 2021, 22, 571–581. [CrossRef]
- Aparicio, C. The olive tree (Olea europaea L.) and salt stress. Importance of growth regulators. University of Jaén (Spain), 2015. https://www.educacion.gob.es/teseo/mostrarRef.do?ref=1273335.
- Gholami, R.; Hashempour, A.; Hoveized, N.F.; Zahedi, S.M. Evaluation of salinity tolerance between six young potted elite olive cultivars using morphological and physiological characteristics. Appl. Fruit Sci. 2024, 66, 163-171. [CrossRef]
- Navarro, J.M.; Pérez-Tornero, O.; Morte, A. Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the rootstock salt tolerance. J. Plant Physiol. 2014. 171, 76–85. [CrossRef]
- Therios, I.N.; Misopolinos, N.D. Genotypic response to sodium chloride salinity of four major olive cultivars (Olea europea L.). Plant Soil 1988. 106, 105–111. [CrossRef]
- Bazakos, C.; Manioudaki, M.E.; Therios, I.; Voyiatzis, D.; Kafetzopoulos, D.; Awada, Kalaitzis, P. Comparative transcriptome analysis of two olive cultivars in response to NaCl-stress. PLoS ONE 2012, 7, e42931. [CrossRef]
- Karimi, H.R.; Hasanpour, Z. Effects of salinity and waters on growth and macro nutrients concentration of pomegranate (Punica granatum L.). J. Plant Nutr. 2012, 37, 1937-1951. [CrossRef]
- Poury, N.; Seifi, E.; Alizadeh, M. The effect of proline and salt stress on growth characteristics of three olive cultivars at three different stages of the growing season. J.Chem. Health Risks. 2019, 9, 133-147. [CrossRef]
- Tattini, M.; Bertoni, P.; Caselli, S. Genotypic responses of olive plants to sodium chloride. J. Plant Nutr. 1992. 15, 1467–1485. [CrossRef]
- Klein, I.; Ben-Tal, Y.; Lavee, S.; de Malach, Y.; David, I. Saline irrigation of cv. Manzanillo and Uovo di Piccione trees. Acta Hort. 1994, 356, 176-180.
- Rewald, B.; Leuschner, C.; Wiesman, Z.; Ephrath, J. Influence of salinity on root hydraulic properties of three olive varieties. Plant Biosyst. 2011, 145, 12–22. [CrossRef]
- Rewald, B.; Rachmilevitch, S.; Ephrath, J. Salt stress effects on root systems of two mature olive cultivars. Acta Hortic. 2011, 888, 109–118. [CrossRef]
- Rewald, B.; Shelef, O.; Ephrath, J.E.; Rachmilevitch, S. Adaptive plasticity of salts-tressed root systems. In Ecophysiology and Responses of Plants Under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V. Springer: Berlin, Germany, 2013; pp. 169–201.
- Schreiber, L.; Hartmann, K.; Skrabs, M.; Zeier, J. Apoplastic barriers in roots: Chemical composition of endodermal and hypodermal cell walls. J. Exp. Bot. 1999, 50, 1267–1280. [CrossRef]
- Stoláriková, M.; Vaculík, M.; Lux, A.; Di Baccio D.; Minnocci, A.; Andreucci, A.; Sebastiani, L. Anatomical differences of poplar (Populus × euramericana clone I-214) roots exposed to zinc excess. Biologia 2012, 67, 483–489. [CrossRef]
- Vigo, C.; Therios, I.N.; Bosabalidis, A.M. Plant growth, nutrient concentration, and leaf anatomy of olive plants irrigated with diluted seawater. J. Plant Nutr. 2005, 28, 1001–11021. [CrossRef]
- Melgar, J.C.; Syvertsen, J.P.; Martínez, V.; García-Sánchez, F. Leaf gas exchange, water relations, nutrient content and growth in citrus and olive seedlings under salinity. Biol. Plant. 2008, 52, 385–390. [CrossRef]
- Kchaou, H.; Larbi, A.; Chaieb, M.; Sagardoy, R.; Msallem, M.; Morales, F. Genotypic differentiation in the stomatal response to salinity and contrasting photosynthetic and photoprotection responses in five olive (Olea europaea L.) cultivars. Sci. Hortic. 2013, 160, 129-138. [CrossRef]
- Fayek, M.A.; Fayed, T.A.; Emtithal, H.; El-Sayed, E.; El-Hameed, A. Comparative impacts of salt stress on survival and leaf anatomy traits in olive genotypes. Biosci. Res. 2018, 15, 565-574. https://www.webofscience.com/wos/alldb/full-record/WOS:000441155500002.
- Saidana Naija, D.; Ben Mansour Gueddes, S.; Braham, M. Not. Bot. Horti Agrobot. Cluj-Na. 2021, 49, 12157. [CrossRef]
- Barba-Espín, G.; Clemente-Moreno, M.J.; Álvarez, S.; García-Legaz, M.F.; Hernandez, J.A.; Diaz-Vivancos, P. Salicylic acid negatively affects the response to salt stress in pea plants. Plant Biol. 2011, 13, 909–917. [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Díaz-Vivancos, P.; Sánchez-Blanco, M.J.; Hernández, J.A. Plant responses to salt stress: Adaptative mechanisms. Agronomy 2017, 7, 18. [CrossRef]
- Wu, H. Plant salt tolerance and Na+ sensing and transport. Crop J. 2018, 6, 215–225. [CrossRef]
- Tattini, M.; Gucci, R.; Coradeschi, M.A.; Ponzio, C.; Everarard, J.D. Growth, gas exchange and ion content in Olea europaea plants during salinity and subsequent relief. Physiol. Plant. 1995, 95, 203–210. [CrossRef]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; Lopes, J.I.; Gonçalvez, C.; Ferreira, T.C.; Corrira, C.M. Physiological behaviour, oxidative damage and antioxidative protection of olive trees grown under different irrigation regimes. Plant Soil 2007, 292, 1-12. [CrossRef]
- Proietti, P.;Nasini, L.; Del Buono, D.; D’Amato, R.; Tedeschini, E.; Businelli, D. Selenium protects olive (Olea europaea L.) from drought stress. Sci. Hortic. 2013, 164, 165–171. [CrossRef]
- Del Buono, D.; Regni, L.; Del Pino, A.M.; Bartucca, M.L.; Palmerini, C.A.; Proietti, P. Effects of megafol on the olive cultivar ‘Arbequina’ grown under serve saline stress in terms of physiological traits, oxidative stress, antioxidant defenses, and cytosolic Ca2+. Front. Plant Sci. 2021, 11, 603576. [CrossRef]
- Skodra, C.; Michailidis, M.; Dasenaki, M.; Ganopoulos, I.; Thomaidis, N.S.; Tanou, G.; Molassiotis, A. Unraveling salt-responsive tissue-specific metabolic pathways in olive tree. Physiol. Plant. 2021, 173, 13565. [CrossRef]
- Centritto, M.; Loreto, F.; Chartzoulakis, K. The use of low [CO2] to estimate diffusional and non-diffusional limitations of photosynthetic capacity of salt-stressed olive saplings. Plant Cell Environ. 2003, 26, 585-594. [CrossRef]
- Tabatabaei, S.J. Salinity Stress and Olive: An Overview. Plant Stress 2007, 1, 105–112.
- Iqbal, M.; Ashraf, M. Gibberellic acid mediated induction of salt tolerance in wheat plants: Growth, ionic partitioning, photosynthesis, yield and hormonal homeostasis. Environ. Exp. Bot. 2013, 86, 76-85. [CrossRef]
- Tattini, M.; Traversi, M.L. On the mechanism of salt tolerance in olive (Olea europaea L.) under low- or high-Ca2+ supply. Environ. Exp. Bot. 2009, 65, 72-81. [CrossRef]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhriss, M.; Ben Abdullah, F. Exogenous proline effects on Photosynthetic performance and antioxidant defense system of young olive tree. J. Agric. Food Chem. 2010, 58, 4216-4222. [CrossRef]
- Bongi, G.; Loreto, F. Gas-exchange properties of salt stressed olive (Olea europea L.) leaves. Plant Physiol. 1989. 90, 1408–1416. [CrossRef]
- Loreto, F.; Centritto, M.; Chartzoulakis, K. Photosynthetic limitations in olive cultivars with different sensitivity to salt stress. Plant Cell Environ. 2003, 26, 595–601. [CrossRef]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286-291. [CrossRef]
- Singh, S. K.; and Reddy, K. R. Regulation of photosynthesis, fluorescence, stomatal conductance and water-use efficiency of cowpea (Vigna unguiculata [L.] Walp.) under drought. J. Photochem. Photobiol. B 2011, 105, 40–50. [CrossRef]
- Proietti, P.; Nasini, L.; Ilarioni, L. Photosynthetic behavior of Spanish arbequina and Italian Maurino olive (Olea europaea L.) cultivars under superintensive grove conditions. Photosynthetica 2012, 50, 239–246. [CrossRef]
- Yamane, K.; Mitsuya, S.; Taniguchi, M.; Miyake, H. Salt-induced chloroplast protrusion is the process of exclusion of ribulose-1,5-bisphosphate carboxylase/ oxygenase from chloroplasts into cytoplasm in leaves of rice. Plant Cell Environ. 2012, 35, 1663-1671. [CrossRef]
- Munns, R.; James, R.A.; Lauchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [CrossRef]
- Kaouther, Z.; Mariem, B.F.; Fardaous, M.; Cherif, H. Impact of salt stress (NaCl) on growth, chlorophyll content and fluorescence of Tunisian cultivars of chili pepper (Capsicum frutescens L.). J. Stress Physiol. Biochem. 2012, 8, 236–252.
- Morales, F.; Abadía, A.; Abadía, J. 2006. Photoinhibition and photoprotection under nutrient deficiencies, drought and salinity. In: Photoprotection. Photoinhibition, Gene Regulation,and Environment. Demmig-Adams, B., Adams III, W.W., Mattoo, A.K., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 65-85.
- Shaheen, M.A.; Hegazi, A.A.; Hmmam, I.S.A. Effect of salinity treatments on vegetative characteristics and leaves chemical content of transplants of five olive cultivars. J. Horticult. Sci. Ornament. Plants. 2011, 3, 143–151.
- Yasar, F.; Ellialtioglu, S.; Yildiz, K. Effect of salt stress on antioxidant defense systems, lipid peroxidation, and chlorophyll content in green bean. Russ. J. Plant Physiol. 2008, 55, 782–786. [CrossRef]
- Saha, P.;Chatterjee, P.; Biswas, A. K. NaCl pretreatment alleviates salt stress by enhancement of antioxidant defense system and osmolyte accumulation in mungbean (Vigna radiata L. Wilczek). Indian J. Exp. Biol. 2010, 48, 593–600. https://www.webofscience.com/wos/alldb/full-record/WOS:000278586800011.
- Din, J.; Khan, S.U.; Ali, I.; Gurmani, A. R. Physiological and agronomic response of canola varieties to drought stress. J. Anim. Plant Sci. 2011, 21, 78–82. https://www.webofscience.com/wos/alldb/full-record/WOS:000288250800016.
- Arjenaki, F. G.; Jabbari, R.; Morshedi, A. Evaluation of drought stress on relative water content, chlorophyll content and mineral elements of wheat (Triticum aestivum L.) varieties. Int. J. Agric. Crop Sci. 2012, 4, 726–729.
- Sevengor, S.; Yasar, F.; Kusvuran, S.; Ellialtioglu, S. The effect of salt stress on growth, chlorophyll content, lipid peroxidation and antioxidative enzymes of pumpkin seedling. Afr. J. Agric. Res. 2011, 6, 4920–4924. [CrossRef]
- Keshavkant, S.; Padhan, J.; Parkhey, S.; Naithani, S. C. Physiological and antioxidant responses of germinating Cicer arietinum seeds to salt stress. Russ. J. Plant Physiol. 2012, 59, 206–211. [CrossRef]
- Demming-Adams, B.; Adams III, W.W. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 599–626. [CrossRef]
- Takahashi, S.; Badger, M.R. Photo protection in plants: A new light on photosystem II damage. Trends Plant Sci. 2011, 16, 53–60. [CrossRef]
- García-Plazaola, J.I.; Esteban, R., Fernández-Marín, B.; Kranner, I.; Porcar-Castell, A. Thermal energy dissipation and xanthophyll cycles beyond the Arabidopsis model. Photosynth. Res. 2012, 113, 89–103. [CrossRef]
- Syvertsen, J.P.; Melgar, J.C.; García-Sánchez, F. Salinity Tolerance and Leaf Water Use Efficiency in Citrus. J. Am. Soc. Hortic. Sci. 2010, 135, 33–39. [CrossRef]
- Li, H.; Hou, X.; Bertin, N.; Ding, R.; Du, T. Quantitative Responses of Tomato Yield, Fruit Quality and Water Use Efficiency to Soil Salinity under Different Water Regimes in Northwest China. Agric. Water Manag. 2023, 277, 108134. [CrossRef]
- Larsen, F.S.; Higgins Wir, A. Diurnal water relations of apple, apricot, grape, olive and peach in an arid environment. Sci. Hortic. 1989, 39, 211–222. [CrossRef]
- Rhızopoulou, S.; Meletiou-Christou, M.S.; Diamandoglou, S. Water relations for sun and shade leaves of four Mediterranean evergreen sclerophytes. J. Exp. Bot. 1991, 42, 627–635. [CrossRef]
- Dichio, B.; Xiloyannis, C.; Angelopoulos, K.; Nuzzo, V.; Bufo, S.A.; Celano, G. Drought-induced variations of water relations parameters in Olea europea. Plant Soil 2003, 257, 381–389. [CrossRef]
- Vitagliano, C.; Sebastiani, L. Physiological and biochemical remarks on environmental stress in olive (Olea europaea L.). Acta Hortic. 2002, 586, 435–440. [CrossRef]
- Larbi, A.; Kchaou, H.; Gaaliche, B.; Gargouri, K.; Boulal, H.K.; Morales, F. Supplementary potassium and calcium improves salt tolerance in olive plants. Sci. Hortic.2020, 260, 108912. [CrossRef]
- Loupassaki, M.H.; Chartzoulakis, K.S.; Digalaki, N.B.; Androulakis, I.I. Effects of salt stress on concentration of nitrogen, phosphorus, potassium, calcium, magnesium, and sodium in leaves, shoots, and roots of six olive cultivars. J. Plant Nutr. 2002, 25, 2457-2482. [CrossRef]
- Tattini, M.; Traversi, M.L. Responses to changes in Ca2+ supply in two Mediterranean evergreens, Phillyrea latifolia and Pistacia lentiscus, during salinity stress and subsequent relief. Ann. Bot. 2008, 102, 609-622. [CrossRef]
- Rus, A.; Yokoi, S.; Sharkhuu, A.; Reddy, M.; Lee, B.H.; Matsumoto, T.K et al. AtHKT1 is a salt tolerance determinant that controls Na+ entry into plant roots. Proceedings of the national academy of sciences, 2001, 98, 14150-14155. [CrossRef]
- Slama, F. Intervention des racines dans le sensibilite ou tolerance a NaCl des plantes cultivees. Agronomie 1986, 6, 651–658.
- Ismail, H.; Maksimović, J.D.; Maksimović, V.; Shabala, L.; Živanović, B.D.; Tian, Y. et al. Rutin, a flavonoid with antioxidant activity, improves plant salinity tolerance by regulating K+ retention and Na+ exclusion from leaf mesophyll in quinoa and broad beans. Functional Plant Biology, 2015, 43, 75-86. [CrossRef]
- El-Shafey, N.M.; AbdElgawad, H. Luteolin, a bioactive flavone compound extracted from Cichorium endivia L. subsp. divaricatum alleviates the harmful effect of salinity on maize. Acta Physiol. Plant, 2012, 34, 2165-2177. [CrossRef]
- Parvin, K.; Hasanuzzaman, M.; Bhuyan, M.B.; Mohsin, S.M.; Fujita, M. Quercetin mediated salt tolerance in tomato through the enhancement of plant antioxidant defense and glyoxalase systems. Plants, 2019, 8, 247. [CrossRef]
- Shenker, M.; Ben-Gal, A.; Shani, U. Sweet corn response to combined nitrogen and salinity environmental stresses. Plant soil, 2003, 256, 139-147. [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol, 1980, 31, 149-190. [CrossRef]
- Grattan, S.R.; Grieve, C.M.. Salinity–mineral nutrient relations in horticultural crops. Sci Hortic, 1998, 78, 127-157.
- Giorio, P.; Sorrentino, G.; d’Andria, R. Stomatal behaviour, leaf water status and photosynthetic response in field-grown olive trees under water deficit. Environ. Exp. Bot. 1999, 42, 95-104. [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551-560. [CrossRef]
- Boughalleb, F.; Mhamdi, M. Possible involvement of proline and the antioxidant defense systems in the drought tolerance of three olive cultivars grown under increasing water deficit regimes. Agric. J. 2011, 6, 378-391.
- Pierantozzi, P.; Torres, M.; Bodoira, R.; Maestri, D. Water relations, biochemical–physiological and yield responses of olive trees (Olea europaea L. cvs. Arbequina and Manzanilla) under drought stress during the pre-flowering and flowering period. Agric. Water Manag. 2013, 125, 13-25. [CrossRef]
- Trabelsi, L.; Gargouri, K.; Ghrab, M.; Mbadra, C.; Hassena, A.B.; Ncube, B. et al. (2024). The effect of drought and saline water on the nutritional behaviour of the olive tree (Olea europaea L.) in an arid climate. S. Afr. J. Bo., 2024, 165, 126-135. [CrossRef]
- Boualem, S.; Boutaleb, F.; Ababou, A.; Gacem, F. Effect of salinity on the physiological behavior of the olive tree (variety sigoise). J. Fundam. Appl. Sci. 2019, 11, 525-538. [CrossRef]
- Demiral, M.A.; Uygun, D.A.; Uygun, M.; Kasirğa, E.; Karagözler, A.A. Biochemical response of Olea europaea cv. Gemlik to short-term salt stress. Turk. J. Biol. 2011, 35, 433-442. [CrossRef]
- Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S., Boukhriss, M.; Abdullah, F.B. Saline water irrigation effects on antioxidant defense system and proline accumulation in leaves and roots of field-grown olive. J. Agric. Food Chem. 2009, 57, 11484-11490. [CrossRef]
- Regni, L.; Del Pino, A.M.; Mousavi, S.; Palmerini, C.A.; Baldoni, L.; Mariotti, R.; et al. Physiological and biochemical traits of different olive tree cultivars during salt stress. In IX International Symposium on Irrigation of Horticultural Crops 2019, 1335, 171-178. [CrossRef]
- Ikbal, F.E.; Hernández, J.A.; Barba-Espín, G.; Koussa, T.; Aziz, A.; Faize, M.; Diaz-Vivancos, P. (2014). Enhanced salt-induced antioxidative responses involve a contribution of polyamine biosynthesis in grapevine plants. J. Plant Physiol. 2014, 171, 779-788. [CrossRef]
- Cardi, M.; Castiglia, D.; Ferrara, M.; Guerriero, G.; Chiurazzi, M.; Esposito, S. The effects of salt stress cause a diversion of basal metabolism in barley roots: Possible different roles for glucose-6-phosphate dehydrogenase isoforms. Plant Physiol. Biochem. 2015, 86, 44–54. [CrossRef]
- Ramanjulu, S.; Sudhakar, C. Proline metabolism during dehydration in two mulberry genotypes with contrasting drought tolerance. J. Plant Physiol. 2000, 157, 81-85. [CrossRef]
- De Lacerda, C.F.; Cambraia, J.; Oliva, M.A.; et al. Solute accumulation and distribution during shoot and leaf development in two sorghum genotypes under salt stress. Environ. Exp. Bot. 2003, 49, 107-120. [CrossRef]
- Chen, Z.; Cuin, T.A.; Zhou, M.; Twomey, A.; Naidu, B.P.; Shabala, S. Compatible solute accumulation and stress-mitigating effects in barley genotypes contrasting in their salt tolerance. J. Exp. Bot. 2007, 58, 4245-4255. [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463-499. [CrossRef]
- Khatkar, D.; Kuhad, M.S. Short-term salinity induced changes in two wheat cultivars at different growth stages. Biol. Plant. 2000, 43, 629-632. [CrossRef]
- Ashraf, M.; Foolad, M.R. Improving plant abiotic-stress resistance by exogenous application of osmoprotectants glycine betaine and proline. Environ. Exp. Bot. 2007, 59, 206-216. [CrossRef]
- Gholami, R.; Fahadi Hoveizeh, N.; Zahedi, S.M.; Gholami, H.; Carillo, P. Effect of three water-regimes on morpho-physiological, biochemical and yield responses of local and foreign olive cultivars under field conditions. BMC Plant Biol. 2022, 22, 477. [CrossRef]
- Zahedi, S.M.; Hosseini, M.S.; Meybodi, N.D.H.; Abadía, J.; Germ, M.; Gholami, R.; Abdelrahman, M. Evaluation of drought tolerance in three commercial pomegranate cultivars using photosynthetic pigments, yield parameters and biochemical traits as biomarkers. Agric. Water Manag. 2022, 261, 107357. [CrossRef]
- Ben Hassine, A.; Ghanem, M.E.; Bouzid, S.; Lutts, S. An inland and a coastal population of the Mediterranean xero-halophyte species Atriplex halimus L. differ in their ability to accumulate proline and glycinebetaine in response to salinity and water stress. J. Exp. Bot. 2008, 59, 1315-1326. [CrossRef]
- Ben Ahmed, C.; Magdich, S.; Rouina, B.B.; Boukhris, M.; Abdullah, F.B. Saline water irrigation effects on soil salinity distribution and some physiological responses of field grown Chemlali olive. J. Environ. Manage. 2012, 113, 538-544. [CrossRef]
- Van den Ende, W.; Valluru, R. Sucrose, sucrosyl oligosaccharides, and oxidative stress: Scavenging and salvaging?. J. Exp. Bot. 2009, 60, 9-18. [CrossRef]
- Trabelsi, L.; Gargouri, K.; Hassena, A.B.; Mbadra, C.; Ghrab, M.; Ncube, B.; et al.. Impact of drought and salinity on olive water status and physiological performance in an arid climate. Agric. Water Manag. 2019; 213, 749-759. [CrossRef]
- Gargouri, K.; Influence of Soil Fertility on Olive (Olea europaea L.) Tree Nutritional Status and Production Under Arid Climates. Doctoral Thesis in Arboriculture, Universita Degli Studi Di Palermo, Italy. 2007.
- Ben Hassena, A.; Zouari, M.; Trabelsi, L.; Decou, R.; Amar, F.B.; Chaari, A.; et al. Potential effects of arbuscular mycorrhizal fungi in mitigating the salinity of treated wastewater in young olive plants (Olea europaea L. cv. Chetoui). Agric. Water Manage. 2021, 245, 106635. [CrossRef]
- Annabi, K.; Laaribi, I.; Gouta, H.; Laabidi, F.; Mechri, B.; Ajmi, L.; et al. Protein content, antioxidant activity, carbohydrates and photosynthesis in leaves of girdled stems of four olive cultivars. Sci. Hortic. 2019, 256, 108551. [CrossRef]
- Lama-Muñoz, A.; del Mar Contreras, M.; Espínola, F.; Moya, M.; Romero, I.; Castro, E. Characterization of the lignocellulosic and sugars composition of different olive leaves cultivars. Food Chem. 2000, 329, 127153. [CrossRef]
- Conde, A.; Silva, P.; Agasse, A.; Conde, C.; Gerós, H. Mannitol transport and mannitol dehydrogenase activities are coordinated in Olea europaea under salt and osmotic stresses. Plant Cell Physiol., 2011, 52, 1766-1775. [CrossRef]
- Žuna Pfeiffer, T.; Štolfa, I.; Hoško, M.; Žanić, M.; Pavičić, N.; Cesar, V.; Lepeduš, H. Comparative study of leaf anatomy and certain biochemical traits in two olive cultivars. Agric. Conspec. Sci. 2020, 75, 91-97.
- Barceló, A.R. Lignification in plant cell walls. Int. Rev. Cytol. 1997, 176, 87-132. [CrossRef]
- Espartero, J.; Pintor-Toro, J.A.; Pardo, J.M. Differential accumulation of S-adenosylmethionine synthetase transcripts in response to salt stress. Plant Mol. Biol. 1994, 25, 217-227. [CrossRef]
- Quiroga, M.; Guerrero, C.; Botella, M.A.; Barceló, A.; Amaya, I.; Medina, M.I.; et al. A tomato peroxidase involved in the synthesis of lignin and suberin. Plant Physiol. 2000, 122, 1119-1128. [CrossRef]
- Sánchez-Aguayo, I.; Rodríguez-Galán, J.M.; García, R.; Torreblanca, J.; Pardo, J.M. Salt stress enhances xylem development and expression of S-adenosyl-L-methionine synthase in lignifying tissues of tomato plants. Planta. 2004, 220, 278-285. [CrossRef]
- Ben Nja, R. Effet d’un stress salin sur la teneur en polymères pariétaux dans les feuilles de luzerne (Medicago sativacv Gabès) et sur la distribution dans les cellules de transfert des fines nervures. Doctoral dissertation, Faculté des Sciences et Techniques, Université de Limoges, France, 2014.
- Niederberger, V.; Purohit, A.; Oster, J.P.; Spitzauer, S.; Valenta, R.; Pauli, G. The allergen profile of ash (Fraxinus excelsior) pollen: Cross-reactivity with allergens from various plant species. Clin. Exp. Allergy. 2002, 32, 933-941. [CrossRef]
- Lee, M.H.; Cho, E.J.; Wi, S.G.; Bae, H.; Kim, J.E.; Cho, J.Y.; et al. Divergences in morphological changes and antioxidant responses in salt-tolerant and salt-sensitive rice seedlings after salt stress. Plant Physiol. Biochem. 2013, 70, 325-335. [CrossRef]
- Mimmo, T.; Bartucca, M.L.; Del Buono, D.; Cesco, S. Italian ryegrass for the phytoremediation of solutions polluted with terbuthylazine. Chemosphere, 2015, 119, 31-36. [CrossRef]
- Del Buono, D.; Pannacci, E.; Bartucca, M.L.; Nasini, L.; Proietti, P.; Tei, F. Use of two grasses for the phytoremediation of aqueous solutions polluted with terbuthylazine. Int. J. Phytoremediation. 2016; 18, 885-891. [CrossRef]
- Bhaduri, A.M.; Fulekar, M.H. Antioxidant enzyme responses of plants to heavy metal stress. Rev. Environ. Sci. Biotechnol. 2012, 11, 55-69. [CrossRef]
- Keunen, E.L.S.; Peshev, D.; Vangronsveld, J.; Van Den Ende, W.I.M; Cuypers, A.N.N. Plant sugars are crucial players in the oxidative challenge during abiotic stress: Extending the traditional concept. Plant Cell Environ. 2013, 36, 1242-1255. [CrossRef]
- López-Gómez, E.; San Juan, M.A.; Diaz-Vivancos, P.; Beneyto, J.M.; García-Legaz, M.F.; Hernández, J.A.. Effect of rootstocks grafting and boron on the antioxidant systems and salinity tolerance of loquat plants (Eriobotrya japonica Lindl.). Environ. Exp. Bot. 2007,60, 151-158. [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil, 2013, 364, 145-158. [CrossRef]
- Bano, S.; Muhammad Ashraf, M.A.; Akram, N.A.; Al-Qurainy, F. Regulation in some vital physiological attributes and antioxidative defense system in carrot (Daucus carota L.) under saline stress. J. Appl. Bot. Food Qual. Angew. Bol. 2012, 85, 105–115.
- Lima Cabello, E.; de Torres, S.; Torrellas López, E.; Medina-Correa, C.; León, I.; Bejarano, P.; et al. Resilience of olive tree cultivars to intensive salt stress. High School Students for Agricultural Science Research 2023, 12, 66-76.
- Tadić, J.; Dumičić, G.; Veršić Bratinčević, M.; Vitko, S.; Radić Brkanac, S. Physiological and biochemical response of wild olive (Olea europaea Subsp. europaea var. sylvestris) to salinity. Front. Plant Sci. 2021, 12, 712005. [CrossRef]
- Demir, S.; Cetinkaya, H. Effects of saline conditions on polyphenol and protein content and photosynthetic response of different olive (Olea europaea L.) cultivars. Appl. Ecol. Environ. Res. 2020, 18. 2599-2610. [CrossRef]
- Naczk, M.; Shahidi, F. Extraction and analysis of phenolics in food. J. Chromatogr. A 2004, 1054, 95-111. [CrossRef]
- Ranalli, A.; Contento, S.; Lucera, L.; Di Febo, M.; Marchegiani, D.; Di Fonzo, V. Factors affecting the contents of iridoid oleuropein in olive leaves (Olea europaea L.). J. Agric. Food Chem. 2006, 54, 434-440. [CrossRef]
- Chutipaijit, S.; Cha-Um, S.; Sompornpailin, K. Differential accumulations of proline and flavonoids in indica rice varieties against salinity. Pak. J. Bot. 2009, 41, 2497-2506.
- Azariadis, A.; Vouligeas, F.; Salame, E.; Kouhen, M.; Rizou, M.; Blazakis, K.; et al. Response of Prolyl 4 Hydroxylases, Arabinogalactan Proteins and Homogalacturonans in Four Olive Cultivars under Long-Term Salinity Stress in Relation to Physiological and Morphological Changes. Cells 2023, 12, 1466. [CrossRef]
- Bashir, M.A.; Silvestri, C.; Coppa, E.; Brunori, E.; Cristofori, V.; Rugini, E. ; et al.. Response of olive shoots to salinity stress suggests the involvement of sulfur metabolism. Plants 2021, 10, 350. [CrossRef]
- Valderrama, R.; Corpas, F. J.; Carreras, A.; Fernández-Ocaña, A.; Chaki, M.; Luque, F.; et al. Nitrosative stress in plants. Febs Lett. 2007, 581, 453-461. [CrossRef]
- Parida, A.K., Das, A.B.; Mittra, B.; Mohanty, P. Salt-stress induced alterations in protein profile and protease activity in the mangrove Bruguiera parviflora. Z. Naturforsch. C. 2004, 59, 408-414. [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324-349. [CrossRef]
- Mansour, M.M.F. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant. 2000, 43, 491-500. [CrossRef]
- Kouhen, M. Involvment of the cell wall arabinogalactan proteins in the salinity tolerance mechanism of four greek olive cultivars. Degree Master of Science, Department of Horticultural Genetics and Biotechnology CIHEAM-Mediterranean Agronomic Institute of Chania, Greece, 2019.
- Ouyang, B.; Yang, T.; Li, H.; Zhang, L.; Zhang, Y.; Zhang, J.; et al. Identification of early salt stress response genes in tomato root by suppression subtractive hybridization and microarray analysis. J. Exp. Bot. 2007, 58, 507-520. [CrossRef]
- Olmos, E.; García De La Garma, J.; Gomez-Jimenez, M. C.; Fernandez-Garcia, N. Arabinogalactan proteins are involved in salt-adaptation and vesicle trafficking in tobacco by-2 cell cultures. Front. Plant Sci. 2017, 8, 274585. [CrossRef]
- Zhao, C.; Zayed, O.; Zeng, F.; Liu, C.; Zhang, L.; Zhu, P.; et al. Arabinose biosynthesis is critical for salt stress tolerance in Arabidopsis. New Phytol. 2019, 224, 274-290. [CrossRef]
- Meyer, S.; Genty, B. Mapping intercellular CO2 mole fraction (C i) in Rosa rubiginosa leaves fed with abscisic acid by using chlorophyll fluorescence imaging: Significance of C i estimated from leaf gas exchange. Plant Physiol., 116, 1998, 947-957. [CrossRef]
- Seeber, F.; Aliverti, A.; Zanetti, G. The plant-type ferredoxin-NADP+ reductase/ferredoxin redox system as a possible drug target against apicomplexan human parasites. Curr. Pharm. Des. 2005, 11, 3159-3172. [CrossRef]
- Badger, M.R.; Price, G.D. The role of carbonic anhydrase in photosynthesis. Annu. Rev. Plant Biol. 1994, 45, 369-392. [CrossRef]
- Park, S. W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl salicylate is a critical mobile signal for plant systemic acquired resistance. Science, 2007, 318, 113-116. [CrossRef]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot., 2011, 62, 3321-3338. [CrossRef]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N. A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front Plant Sci. 2015, 6, 135066. [CrossRef]
- Cabane, M.; Afif, D.; Hawkins, S. Lignins and abiotic stresses. In Advances in botanical research. Academic Press. 2012, 61, 219-262. [CrossRef]
- Jimenez-Lopez, J.C.; Kotchoni, S.O.; Hernandez-Soriano, M.C.; Gachomo, E. W.; Alché, J.D. Structural functionality, catalytic mechanism modeling and molecular allergenicity of phenylcoumaran benzylic ether reductase, an olive pollen (Ole e 12) allergen. J. Comput. Aided Mol. Des. 2013, 27, 873-895. [CrossRef]
- Castro, L.; Crespo, J.F.; Rodríguez, J.; Rodríguez, R.; Villalba, M. Immunoproteomic tools are used to identify masked allergens: Ole e 12, an allergenic isoflavone reductase from olive (Olea europaea) pollen. Biochim. Biophys. Acta, Proteins Proteomics. 2015, 1854, 1871-1880. [CrossRef]
- Mansour, M.M.F.; Salama, K.H. Cellular basis of salinity tolerance in plants. Environ. Exp. Bot. 2004, 52, 113-122. [CrossRef]
- Ashraf, M.P.J.C.; Harris, P.J. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3-16. [CrossRef]
- Almeida, A.M.; Cardoso, L.A.; Santos, D.M.; Torné, J.M.; Fevereiro, P.S. Trehalose and its applications in plant biotechnology. In Vitro Cell Dev. Biol. Plant. 2007, 43, 167-177. [CrossRef]
- Said, E.M. Expression analysis of AtTPS1 gene coding for trehalose-6-phosphate synthase in salt stressed olive (Olea eurropaea L.). Biosci. Res. 2017, 14, 809-816. https://www.webofscience.com/wos/alldb/summary/3f514112-e14b-4a82-909a-1737254acf2b-e9cc0e08/relevance/1.
- Bray, E.A. Plant responses to water deficit. Trends Plant Sci. 1997, 2, 48-54. [CrossRef]
- Seki, M.; Kamei, A.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Molecular responses to drought, salinity and frost: Common and different paths for plant protection. Curr. Opin. Biotechnol. 2003, 14, 194-199. [CrossRef]
- Zhang, H.; Huang, Z.; Xie, B.; Chen, Q.; Tian, X.; Zhang, X.; et al. The ethylene-, jasmonate-, abscisic acid-and NaCl-responsive tomato transcription factor JERF1 modulates expression of GCC box-containing genes and salt tolerance in tobacco. Planta 2004, 220, 262-270. [CrossRef]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; et al. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672-685. [CrossRef]
- Bazakos, C.; Manioudaki, M.E.; Sarropoulou, E.; Spano, T.; Kalaitzis, P. 454 pyrosequencing of olive (Olea europaea L.) transcriptome in response to salinity. PLoS ONE 2015, 10, e0143000. [CrossRef]
- Mantri, N. L., Ford, R., Coram, T. E., & Pang, E. C. (2007). Transcriptional profiling of chickpea genes differentially regulated in response to high-salinity, cold and drought. BMC genomics 2007, 8, 1-14. [CrossRef]
- Qing, D.J.; Lu, H.F.; Li, N.; Dong, H.T.; Dong, D. F.;Li, Y.Z. Comparative profiles of gene expression in leaves and roots of maize seedlings under conditions of salt stress and the removal of salt stress. Plant Cell Physiol. 2009, 50, 889-903. [CrossRef]
- Ehlting, J.; Provart, N.J.; Werck-Reichhart, D. Functional annotation of the Arabidopsis P450 superfamily based on large-scale co-expression analysis. Biochem. Soc. Trans. 2006, 34, 1192-1198. [CrossRef]
- Gálvez, F.J.; Baghour, M.; Hao, G.; Cagnac, O.; Rodríguez-Rosales, M.P.; Venema, K. Expression of LeNHX isoforms in response to salt stress in salt sensitive and salt tolerant tomato species. Plant Physiol. Biochem. 2012, 51, 109-115. [CrossRef]
- Ahmed, H.E.; Youssef, E.A.; Kord, M.A.; Qaid, E.A. Trehalose accumulation in wheat plant promotes sucrose and starch biosynthesis. Jordan J. Biol. Sci. 2013, 6, 143-149. [CrossRef]
- Blázquez, M.A.; Santos, E.; Flores, C.L.; Martínez-Zapater, J.M.; Salinas, J.; Gancedo, C. Isolation and molecular characterization of the Arabidopsis TPS1 gene, encoding trehalose-6-phosphate synthase. Plant J. 1998, 13, 685-689. [CrossRef]
- Vogel, G.; Aeschbacher, R.A.; Müller, J.; Boller, T.; Wiemken, A. Trehalose-6-phosphate phosphatases from Arabidopsis thaliana: Identification by functional complementation of the yeast tps2 mutant. Plant J. 1998, 13, 673-683. [CrossRef]
- Leyman, B.; Van Dijck, P.;Thevelein, J.M. An unexpected plethora of trehalose biosynthesis genes in Arabidopsis thaliana. Trends Plant Sci. 2001, 6, 510-513. [CrossRef]
- Penna, S. Building stress tolerance through over-producing trehalose in transgenic plants. Trends Plant Sci. 2003, 8, 355-357. [CrossRef]
- Mousavi, S.; Mariotti, R.; Valeri, M.C.; Regni, L.; Lilli, E.; Albertini, E.; et al. Characterization of differentially expressed genes under salt stress in olive. Int. J. Mol. Sci. 2021, 23, 154. [CrossRef]
- Reddy, P.S.; Jogeswar, G.; Rasineni, G.K.; Maheswari, M.; Reddy, A.R.; Varshney, R.K.; Kishor, P.K. Proline over-accumulation alleviates salt stress and protects photosynthetic and antioxidant enzyme activities in transgenic sorghum [Sorghum bicolor (L.) Moench]. Plant Physiol. Biochem. 2015, 94, 104-113. [CrossRef]
- Guan, C.; Cui, X.; Liu, H.Y.; Li, X.; Li, M.Q.; Zhang, Y.W. Proline biosynthesis enzyme genes confer salt tolerance to switchgrass (Panicum virgatum L.) in cooperation with polyamines metabolism. Front. Plant Sci. 2020, 11, 46. [CrossRef]
- Pakzad, R.; Goharrizi, K.J.; Riahi-Madvar, A.; Amirmahani, F.; Mortazavi, M.; Esmaeeli, L. Identification of Lepidium draba Δ 1-pyrroline-5-carboxylate synthetase (P5CS) and assessment of its expression under NaCl stress: P5CS identification in L. draba plant. Proceedings of the National Academy of Sciences. India Sect. B: Biol. Sci. 2021, 91, 195-203. [CrossRef]
- Liu, H.; Hu, M.; Wang, Q.; Cheng, L.; Zhang, Z. Role of papain-like cysteine proteases in plant development. Front. Plant Sci. 2018, 9, 1717. [CrossRef]
- Joaquín-Ramos, A.; Huerta-Ocampo, J.Á.; Barrera-Pacheco, A.; De León-Rodríguez, A.; Baginsky, S.; de la Rosa, A.P.B. Comparative proteomic analysis of amaranth mesophyll and bundle sheath chloroplasts and their adaptation to salt stress. J. Plant Physiol. 2014, 171, 1423-1435. [CrossRef]
- Wang, M. J.; Ding, L.; Liu, X.H.; Liu, J.X. Two B-box domain proteins, BBX28 and BBX29, regulate flowering time at low ambient temperature in Arabidopsis. Plant Mol. Biol. 2021; 106, 21-32. [CrossRef]
- Zhang, M.; Chen, Q.; Zhou, P.; Zhang, Q.; Fang, Y. NaCl-induced changes in vacuolar H+-ATPase expression and vacuolar membrane lipid composition of two shrub willow clones differing in their response to salinity. Plant Growth Regul. 2018, 86, 445-453. [CrossRef]
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ response mechanisms to salinity stress. Plants 2023, 12, 2253. [CrossRef]
- Shabala, S.; Hariadi, Y.; Jacobsen, S.E. Genotypic difference in salinity tolerance in quinoa is determined by differential control of xylem Na+ loading and stomatal density. J. Plant Physiol. 2013, 170, 906-914. [CrossRef]
- Seidel, T. The Plant V-ATPase. Front. Plant Sci. 2022, 13, 931777. [CrossRef]
- El Yamani. M.; Sakar, E.H.; Boussakouran, A.; Rharrabti, Y. Physiological and biochemical responses of young olive trees (Olea europaea L.) to water stress during flowering. Arch. Biol. Sci. 2019, 71, 123-132. [CrossRef]
- El Yamani, M.; Sakar, E.H., Boussakouran, A.; Rharrabti, Y. Leaf water status, physiological behavior and biochemical mechanism involved in young olive plants under water deficit. Sci. Hortic. 2020, 261, 108906. [CrossRef]
- Abd El Rahman, A.; Sharkawi, M.H. Effect of salinity and water supply on olive. Plant Soil 1968, 280-290. [CrossRef]
- Gucci, R.; Tattini, M.; Lombardini, L. Meccanismi di resistenza allo stress osmosalino in olivo. In Olivicoltura mediterranea: Stato e rpospettive della coltura e della ricerca. La Grafica Commerciale. 1996, pp. 305-312.
- Sadok, W.; Lopez, J.R.; Smith, K.P. Transpiration increases under high-temperature stress: Potential mechanisms, trade-offs and prospects for crop resilience in a warming world. Plant Cell Environ. 2021, 44, 2102-2116. [CrossRef]
Figure 1.
General outline of the aspects to be reviewed in this paper.
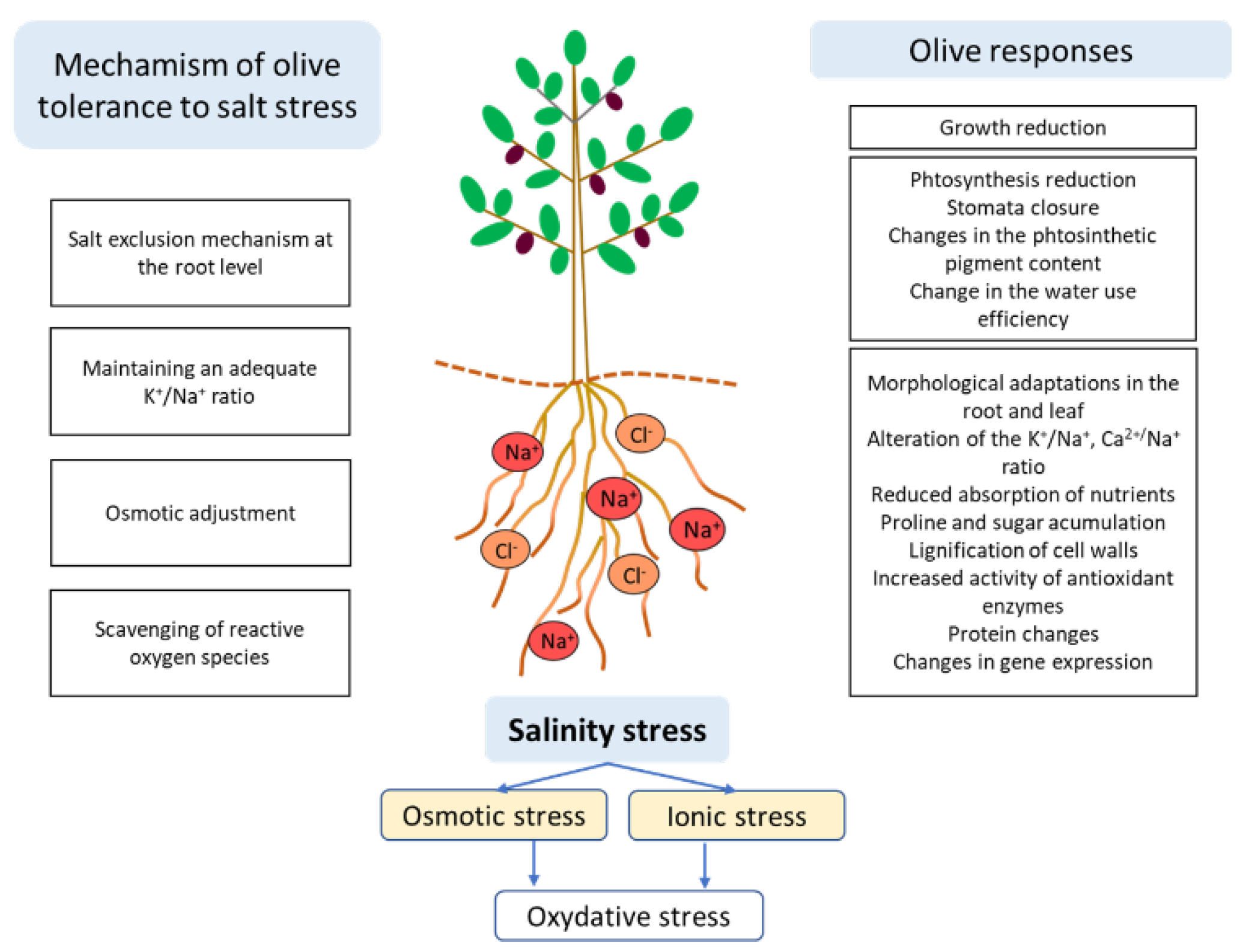
Figure 2.
Distribution of the olive groves in the world [38].
Figure 2.
Distribution of the olive groves in the world [38].
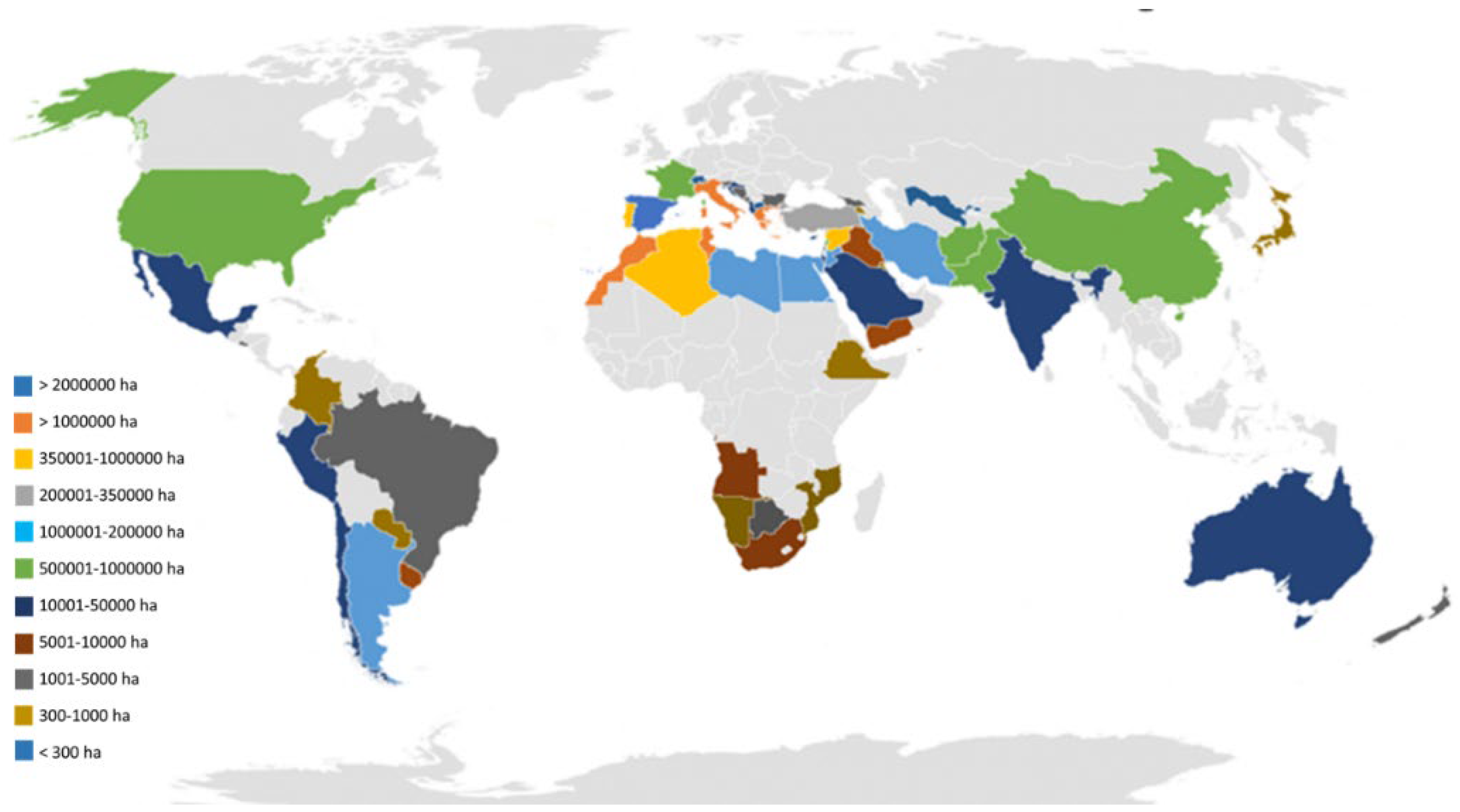
Figure 3.
(A) Area harvested (Mha), (B) production (t), (C) destination of olives (Mha), (D) yield (t/ha), (E) area (Mha) of rainfed, and irrigated olive grove cultivation, and (F) area (%) of low-density, intensive and super-intensive olive grove cultivation in 2021. Prepared by the authors based on data from the IOC [40].
Figure 3.
(A) Area harvested (Mha), (B) production (t), (C) destination of olives (Mha), (D) yield (t/ha), (E) area (Mha) of rainfed, and irrigated olive grove cultivation, and (F) area (%) of low-density, intensive and super-intensive olive grove cultivation in 2021. Prepared by the authors based on data from the IOC [40].
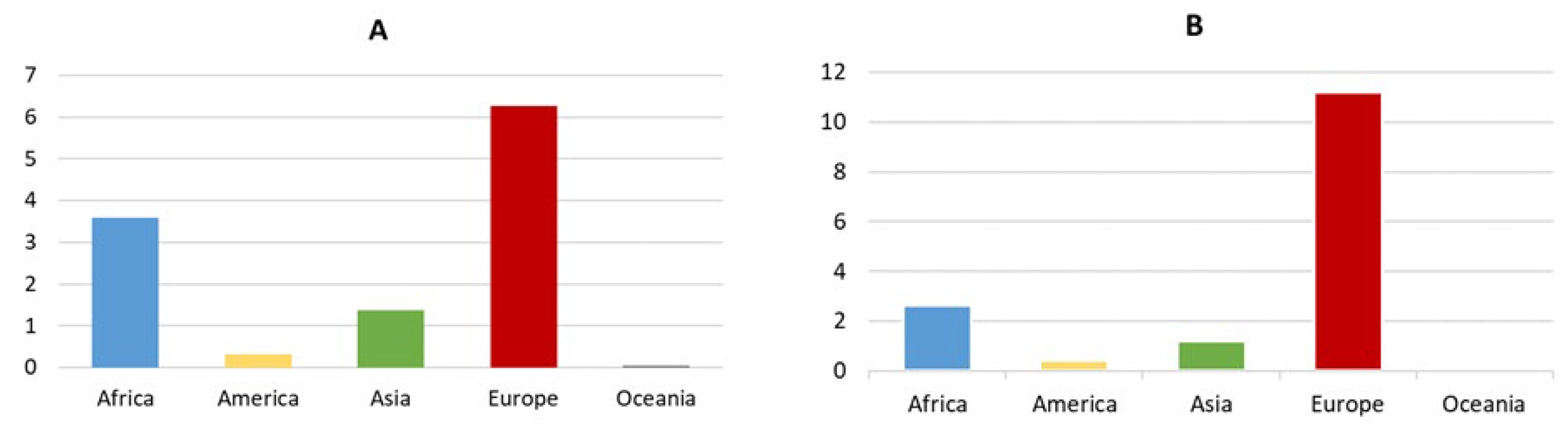
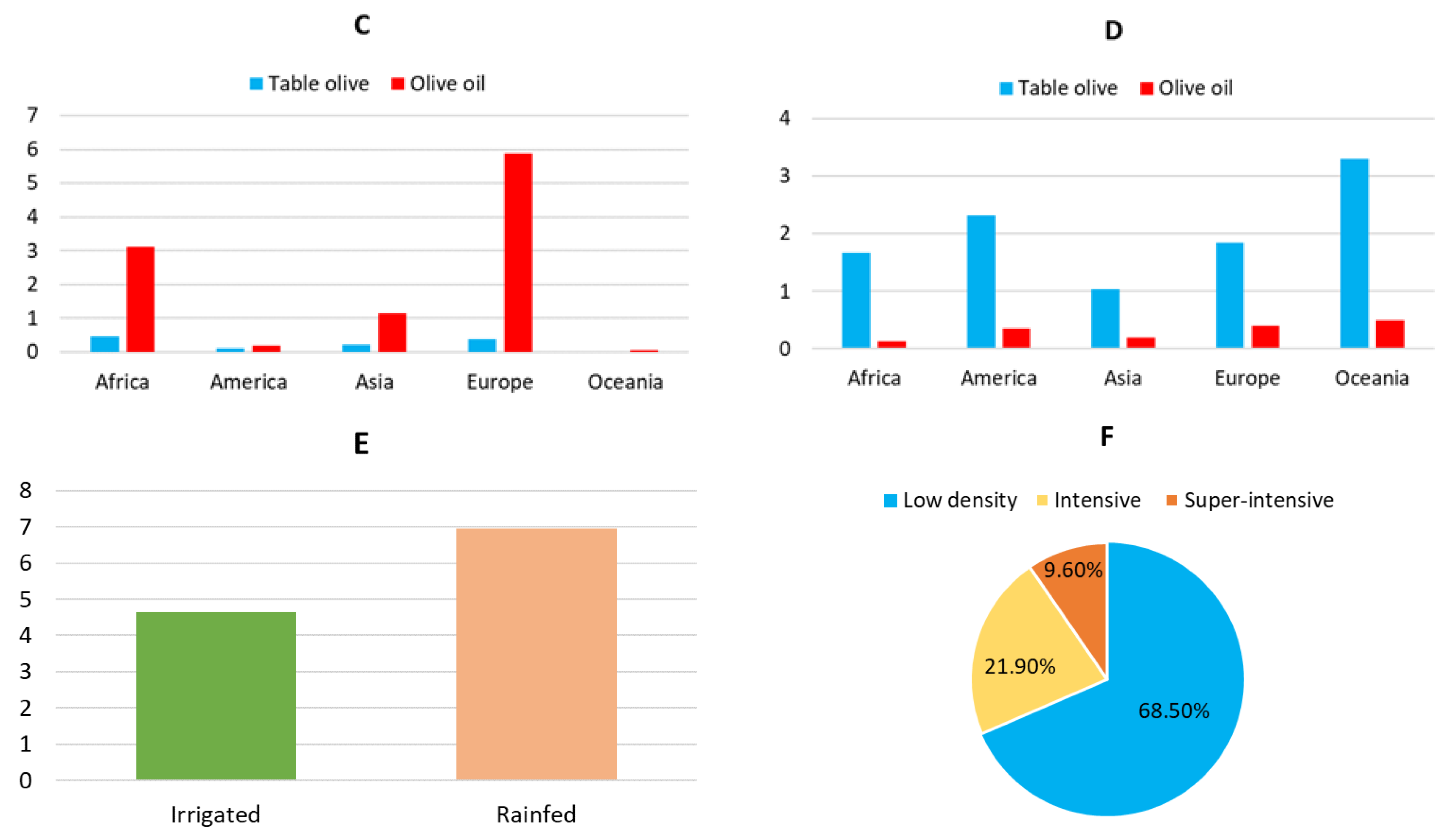
Figure 4.
(Average (A) table olive (x 103 t) and (B) olive oil production (x 103 t) in the main producing countries from 2013 to 2016. Prepared by author based on data from the IOC [37].
Figure 4.
(Average (A) table olive (x 103 t) and (B) olive oil production (x 103 t) in the main producing countries from 2013 to 2016. Prepared by author based on data from the IOC [37].
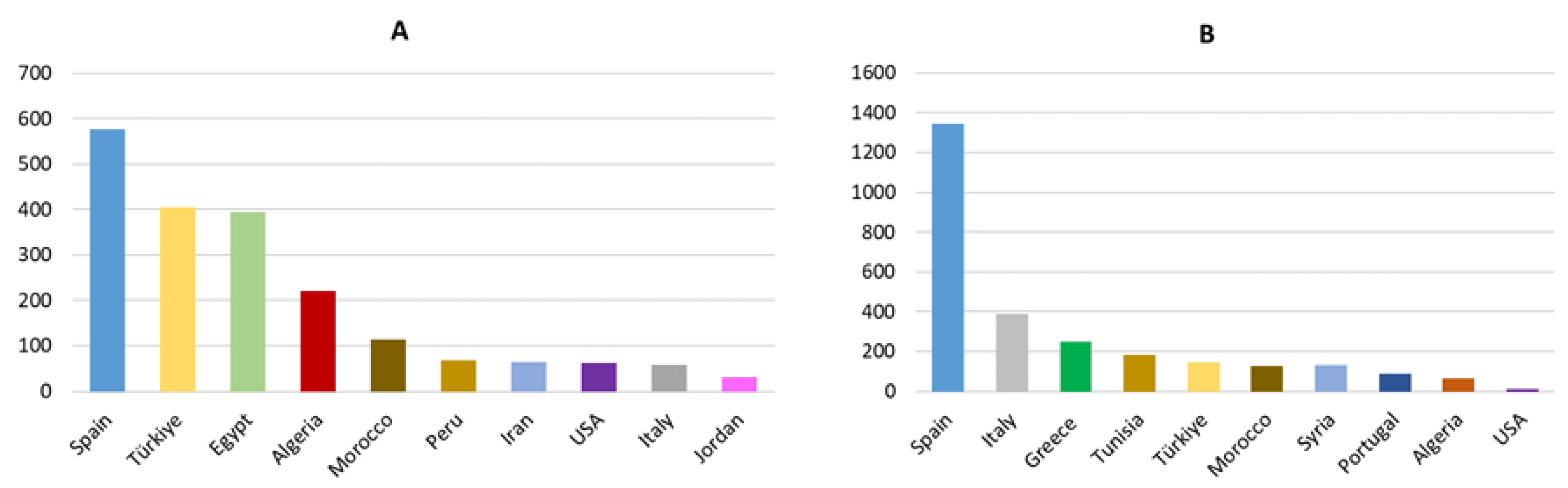
Figure 5.
Olive oil production (x 103 t) from 1990 to 2022. Prepared by author based on data from the IOC [40].
Figure 5.
Olive oil production (x 103 t) from 1990 to 2022. Prepared by author based on data from the IOC [40].
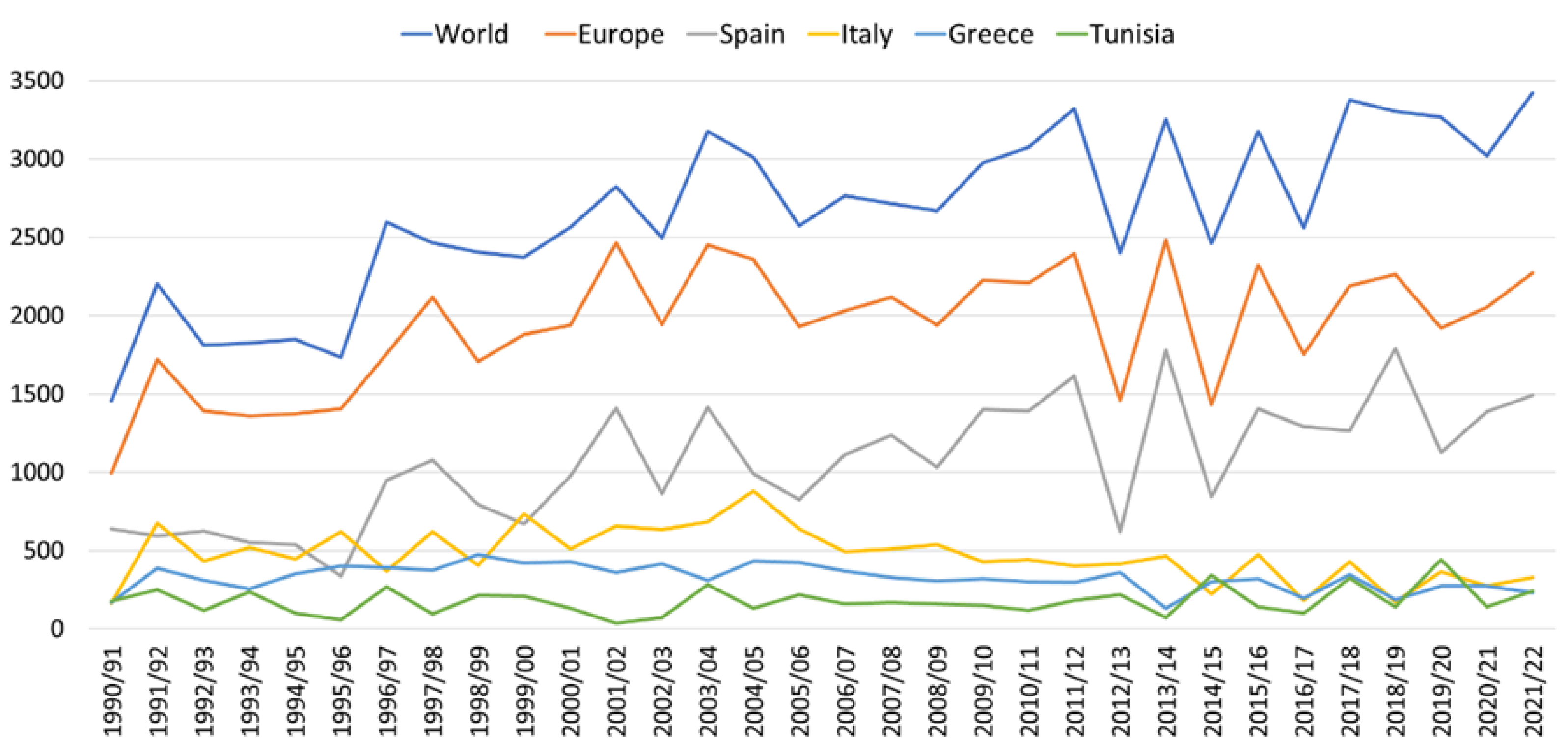
Figure 6.
(A) Control leaf and (B) leaves with ion accumulation. Necrosis starts at the leaf apex and progresses towards the base [106].
Figure 6.
(A) Control leaf and (B) leaves with ion accumulation. Necrosis starts at the leaf apex and progresses towards the base [106].


Table 1.
Continental distribution of the area of saline soils (Mha). Prepared by the authors based on data from Mukhopadhyay et al. [1].
Table 1.
Continental distribution of the area of saline soils (Mha). Prepared by the authors based on data from Mukhopadhyay et al. [1].
| Continents | Area (Mha) |
|---|---|
| Africa | 80.44 |
| America | 144.73 |
| Asia | 316.50 |
| Europe | 30.70 |
| Oceania | 357.33 |
| World | 931.70 |
Table 2.
List of the most important cultivars of each continent [38].
Table 2.
List of the most important cultivars of each continent [38].
| Continents | Main Cultivars |
|---|---|
| Africa | Picholine, Marroqui, Hacuzia, Meslala, Menara, Arbosana, Arbequina, Koroneiki, Woira, Mission, Kalamata |
| America | Arbequina, Azapá, Criolla, Coratina, Barouni, Manzanilla, Ascolana, Mision, Arauco, Picus |
| Asia | Arjosi, Barmagui, Basica, Kikkam, Kasb, Souri, Nabal Baladi, Mehravia, Muhasan, Mastoidis |
| Europe | Arbequina, Picual, Hojiblanca, Koroneiki, Arbosana, Alentajana, Frantoio, Verdial, Picudo, Carbonella |
| Oceania | Hardy’s, Mammoth, Fs17, Dai21, Azapa, Picual, Hojiblanca, Manzanilla, Barnea, Frantoio, Koroneiki |
| World | Arbequina, Arbosana, Koroneiki, Picual, Frantoio, Leccino, Hojiblanca, Verdial, Kalamata, Picholine, Alentajana, Nabal Baladi |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



