Submitted:
28 May 2024
Posted:
29 May 2024
You are already at the latest version
Abstract
The most recent National Research Council Nutrient Requirements for Sheep and Goats was published in 2007, one of the most consequential nutrient requirement recommendations for sheep and goats in the world. It is pertinent to continue the refinement and discussion of nutrient utilization and requirements for the purposes of enhancing the efficiency of production systems, maximizing resource economy, and minimizing the carbon footprint, among others. Progress has been made in the estimation of energy and protein requirements in sheep and goats, mainly utilizing empirical feeding experimentation, comparative slaughter technique and minimum endogenous loss method. In sheep, newer estimates of energy and protein requirements for maintenance and growth and partial efficiencies had been reported since 2007. There were suggestions that energy and protein requirements could have been affected by breed, wool growth, gender and size; with these reported values being similar or lower than the recommended values in international feeding systems such as NRC, ARC, INRA and AFRC. In goats, energy and protein requirements for growing goats were reported to be either higher or lower than the established recommendations, depending upon meat or dairy breeds. Effect of gender on energy requirement appeared to be related to the stage of growth or degree of maturity. Newer data also suggested that the guidelines of the major feeding systems for the entire growth phase may not be adequate for non-pregnant and non-lactating pubertal females. In multiparous pregnant goats, energy and protein requirements for maintenance did not appear to be affected by days of pregnancy, but efficiencies of metabolizable energy and metabolizable protein utilization for pregnancy were. There were suggestions that that metabolizable protein synthesis in the rumen can be predicted from energy intake using combined equations that encompass both sheep and goats, but more data on goats were called for to account for specific differences in nutrition. In addition to sulfur, there has been progress made on the estimation of maintenance and growth requirements of calcium, phosphorus, potassium and magnesium in goats, with suggestions on the consideration of gender and breed differences. Species, level of Intake, genotype, mature size, age, gender, body composition, use of tissue energy, activity, season, and acclimatization continue to be important considerations for the estimation of nutrient requirements in sheep and goats, with emerging factors such as climate change, heat stress, parasitism and secondary plant compounds. Model equations and partial efficiencies used by NRC to predict energy and protein requirements for maintenance, growth, lactation, and fiber have been highlighted and discussed for the purpose of a more focus discussion and refinement for the future. Potential and limitation of both traditional and emerging methodologies in determining the nutrient requirements in sheep and goats were discussed. To justify the research investment, emerging methodologies such as nutrigenomics will have to be linked more directly to the improvement of production efficiency via more precise prediction of nutrient requirement.
Keywords:
Sheep
; goat
; nutrient requirement
; nutrient utilization
; nutrigenomics
1. Introduction
The economic and social impacts of sheep and goat production in the improvement of living standards and alleviation of poverty in rural communities are recognized throughout the world [1]. Understanding nutrient utilization by sheep and goats enhances the efficiency of production systems, maximizes resource economy, and minimizes the carbon footprint. Nutrient requirements can be one of the most important considerations for the precision nutrition [2] in sheep and goat production. Requirements and utilization of structural and nonstructural carbohydrates, degradable and bypass proteins, lipids, minerals, and vitamins are essential parts of this important scientific understanding. Nutrient requirements are affected by multiple factors such as individuality, genetics, climates, diet, age, and physiological stages [3], and a continuous update of knowledge is essential. Relatively recent advancements in nutrigenomics [4,5,6,7] have led to improved knowledge on the molecular interaction between nutrients and other dietary bioactive components with respect to the genome, effect of food constituents on gene expression, and influence of genetic variation on nutrition; therefore, contributing to a better understanding of nutrient utilization and requirements. Such understanding allows optimizing and customizing nutrition with respect to a subject’s genotype and offers great potential for a more precise determination of nutrient requirements in sheep and goats. The most recent National Research Council (NRC) Nutrient Requirements for Sheep and Goats was published in 2007 [3]. It is pertinent to continue the refinement and discussion of nutrient utilization and requirements in sheep and goats from the perspectives of traditional nutrition and nutrigenomics. As an introductory article for the special issue on “Nutrient utilization, requirements, and nutrigenomics in sheep and goats”, the objective of this paper is to provide an overview of the progress in establishing nutritional standards through traditional research in nutrient utilization and nutrient requirements, but also considering new approaches such as precision nutrition and nutrigenomics.
2. National Research Council and History of Nutrient Requirements in Sheep and Goats
The National Research Council was organized in 1916 under a congressional charter requested by then President Wilson of the United States [8]. It is the operating arm of National Academy of Sciences, National Academy of Engineering, and Institute of Medicine; with the purposes of improving government decision and public policy, increasing public understanding, and disseminating knowledge in science, engineering, technology and health. The National Research Council supports studies in six divisions including Earth and Live Sciences. The Division of Earth and Live Sciences conducted studies and publishes “Nutrient Requirements of Animals”. Under the Division of Earth and Live Sciences, Board on Agriculture and Natural Resources approves nutrient requirements of various animal species upon the recommendation of Committee on Nutrition and Subcommittee of Nutrition of various animal species.
The 1st Edition of Nutrient Allowances for Sheep was published by NRC in 1945 [9] and was revised in 1949 [10], 1957 [11], 1964 [12], 1968 [13], 1975 [14], with the 6th revised Edition published in 1985 [15]. The 6th revised Edition [15] was the last published NRC recommendations solely focusing on sheep. The 1st Edition of Nutrient Requirements of Goats was published by NRC in 1981 [16]. Since the NRC published its 1st Edition of Nutrient Requirements of Goats, there has been advancement in research pertaining to energy and protein [17,18,19,20,21,22] and sulfur [23,24,25,26,27,28,29] requirements in milk, meat and fiber producing goats that may or may not have been considered fully in the most recent NRC publication on nutrient requirements of goats [3]. A series of articles pertaining to energy and protein requirements in goats [30,31,32,33,34] were published prior to and considered in the 2007 NRC publication [3]. The latest version of nutrient requirements for sheep and goats combinedly appeared in “Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids and New World Camelids” and was published sixteen years ago [3]. Incorporated in the 2007 publication were the concepts of ruminally undegraded protein and microbial crude protein as sources of metabolizable protein (MP). For large ruminants, “Nutrient Requirements of Beef Cattle” and “ Nutrient Requirements of Dairy Cattle have been revised and published more recently [35,36]. “Nutrient Requirements of Animals” is considered one of the most important nutritional standards once published. They are commonly used as a standard for future animal experiments, and NRC recommendations are among the most cited publications in animal nutrition. They can be considered as the culmination of nutritional accomplishment at a particular time period.
In addition to NRC, there are other notable international nutrition standards for sheep and goats that have been recommended by the Agricultural Food and Research Council in the United Kingdom [37,38,39,40], the Instituto National de la Recherche Agronomique in France [41,42,43], and the Commonwealth Scientific and Industrial Research Organization in Australia [44,45]. Most of these recommendation predated the most recent NRC recommendation for sheep and goats [3]. Nutrient requirements of meat-type sheep and goats were revised in China, after a decade-long coordinated national effort utilizing comparative slaughter trial, digestibility trial, and carbon–nitrogen balance technique [46].
3. Energy and Protein Requirements in Sheep
Since 2007, there have been progress made in the estimation of energy and protein requirements for maintenance and growth in sheep. Galvani et al. [47] conducted comparative slaughter experiment using Texel crossbred lambs (n = 30) and reported that net energy (NEm) and metabolizable energy requirements for maintenance (MEm) were 58.6 and 91 kcal/kg 0.75 of shrunk body weight (SBW), respectively. Partial efficiency of energy use for maintenance was calculated to be 0.64. Net energy requirements for growth (NEg) of lambs at 15 and 35 kg of SBW with an average daily gain (ADG) of 250 g were calculated to be 424 and 553 kcal/d, respectively. Partial efficiency of energy use for growth was calculated to be 0.47. To estimate protein requirements by endogenous nitrogen (N) loss in Texel crossbred lambs [48], the growth pattern of the wool was found to influence protein requirements and the estimated growth requirements were lower than those reported by the most nutritional systems. They reported that net protein requirement for maintenance (NPm) was 1.52 ± 0.18 g/kg 0.75 of SBW, metabolizable protein requirement for maintenance (MPm) was 2.31 g/ kg 0.75 of SBW, and the efficiency of MP use for maintenance was 0.66. Net protein requirements for body weight gain (NPg) and wool growth (NPw) of lambs at 15 and 35 kg of SBW, with an ADG of 250 g, were 28.7 and 27.3 g/day and 3.8 and 5.8 g/day, respectively. Estimated efficiencies of MP use for body weight gain (kpg) and wool growth (kpw) were 0.71 and 0.46, respectively. Salah et al. [49] conducted a meta-analysis study of 590 publications and concluded that energy and protein requirements of tropical and warm-area ruminants compared with those proposed in the international feed system standards such as the NRC, ARC, INRA and AFRC tables were higher and little or no differences between species were found.
Comparative slaughter and digestion trials were conducted on Dorper × Hu crossbred lambs to determine energy and protein requirements [50]. Energy and protein requirement for growth in growing lambs were founded to similar to the NRC recommendations for early and later maturating growing sheep. They reported that MEm (400.61 ± 20.31 vs. 427.24 ± 18.70 kJ kg−1 /SBWkg0.75), partial efficiency of ME utilization for maintenance (km, 0.64 ± 0.02 vs. 0.65 ± 0.03), partial efficiency of ME utilization for growth (kg ; 0.42 ± 0.03 vs. 0.44 ± 0.02), and NPm (1.83 ± 0.17 vs. 1.99 ± 0.28 g kg−1 of SBW0.75) did not differ due to gender. They also reported that rams had higher NPg (15.9 to 44.3 g d−1) than those of ewes (13.07 to 33.0 g d−1) at the similar condition of BW and ADG. Oliveira et al. [51] suggested that total ME and MP requirements were lower than those recommended by the NRC and AFRC from a meta-analysis of seven experiments (n = 243) and proposed new equations for nutrient requirements of hair sheep raised in tropical regions. A comparative slaughter experiment on a hair sheep breed Santa Ines lambs (n = 38) suggested that the importance of updating the tables of international committees and of including data obtained from studies with sheep breeds raised in tropical conditions, with the purpose of improving the productive efficiency of the animals [52]. They reported that NEg varied from 1.13 to 2.01 MJ/d for lambs with BW of 15 and 30 kg and ADG of 200 g. The calulated NPg varied from 24.6 to 16.3 g/d for lambs with BW of 15 and 30 kg with ADG of 200 g. The calculated metabolizable energy efficiency for gain (kg) was 0.37, and the metabolizable protein efficiency for gain (kpg) was 0.28. The NEm and NPm did not differ between castrated and non-castrated lambs, with values of 0.241 MJ/kg FBW0.75/d and 1.30 g/kg FBW0.75/d, respectively. The metabolizable energy efficiency for maintenance (km) was 0.60, and the efficiency of metabolizable protein for maintenance (kpm) was 0.57.
Pereira et al. [53] conduct a comparative slaughter experiment to evaluate the energy and protein requirements of intact male, castrated male and female Morada Nova lambs (n = 47). For all sexes, the NEm was 73.0 kcal/kg0.75 empty body weight (EBW)/d. The metabolizable energy efficiency for maintenance (km) was 0.58. The metabolizable energy efficiency utilization for gain (kg) was 0.36; 0.25 and 0.28 for intact males, castrated males and females, respectively. The NEg was 0.191, 0.198 and 0.276 Mcal/kg0.75 EBW/d for intact males, castrated males and females, respectively, with a BW of 20 kg and a ADG of 100 g and differed between sexes. The NPm and MPm were 1.06 g/kg0.75 BW/d and 3.46 g/kg0.75 BW/day, respectively and the NPg differed between sexes. The NPg was 7.08, 7.11 and 6.78 g/d for intact males, castrated males and females, respectively, for animals weighing 20 kg and with ADG of 100 g/d. They concluded that net energy and protein requirements for maintenance of Morada Nova lambs slaughtered between 15 and 28 kg did not vary by sex, and NEg increases and NPg decreased with the increase in body weight in hair lambs. Mendes et al. [54] utilized 35 crossbreed Doper x Santa Ines lambs in a slaughter experiment to estimate energy and protein requirements for maintenance and growth and concluded that NEm for crossbreed Dorper × Santa Ines lambs was similar to those recommended by the international committees but NEg was lower.
4. Energy and Protein Requirements in Goats
Since 2007, there are a number of articles published pertaining to energy and protein requirements in goats. To determine the energy and protein requirements for maintenance and growth using 34 ¾ Boer × ¼ Saanen crossbred, intact male kids [55], the NEm was reported to be 77.3 kcal/kg0.75 EBW or 67.4 kcal/kg0.75 SBW. The MEm were 118.1 kcal/kg0.75 EBW or 103.0 kcal/ kg0.75 of SBW. These requirements were calculated by iteration assuming that the heat produced was equal to the ME intake at maintenance. The partial efficiency of the use of ME for NE for maintenance was 0.65. The NPm was 2.44 g / kg0.75 EBW. The NEg ranged from 2.55 to 3.0 Mcal/kg of EBW gain at 20 and 35 kg of BW, and NPg ranged from 179 to 185 g/kg of EBW gain. They conclude that NEg and NPg for growing meat goats exceeded the requirements previously published for dairy goats. They also suggested that N requirement for maintenance for growing goats was greater than the established recommendations. When goats were included in a mechanistic model [56] termed Cornell Net Carbohydrate and Protein System (CNCPS-S) that predicts nutrient requirements and biological values of feeds for sheep, Tedeschi et al. [57] suggested more data for goats are needed to account for specific differences in nutrition. Medeiros et al. [58] utilized Saanen male kids (n =41) in a slaughtered experiment and suggested that NEm was 417 kJ/kg0.75 EBW/d, while the MEm was 657 kJ/kg0.75 EBW/d. The efficiency of ME use for NE maintenance (km) was 0.64. The NEg ranged from 7.4 to 9.0 MJ/kg of empty weight gain/d at 5 and 20 kg BW, respectively. Their study indicated that the energy requirements in goats were lower than previously published requirements by AFRC [40] and NRC [3] for growing dairy goats.
Results from a comparative slaughter experiment on Saanen goat kids (n = 54) suggested that it was not necessary to formulate diets with different energetic content for intact male, castrated male and female Saanen goat kids weighing from 5 to 15 kg [59]. They found no effect of gender on the energy requirements for maintenance and gain, and overall NEm was 205.6 kJ/kg0.75 EBW gain (EWG) or 170.3 kJ/kg0.75 BW from 5 to 15 kg BW. The MEm (calculated by iteration assuming heat production equal to metabolizable energy intake at maintenance) was 294.3 kJ/kg0.75 EBW with a km of 0.70. As BW increased from 5 to 15 kg for all genders, the NEg increased from 9.5 to 12.0 kJ/g EWG, and assuming kg = 0.47, MEg ranged from 20.2 to 25.5 kJ/g EWG. For growing male and female Saanen goats, MEm was reported to be 412.4 kJ/ kg0.75 BW, with an estimated energy use efficiency for maintenance of 0.627 [60]. During the growth phase, NEg differed between the sexes; intact males, castrated males, and females with an average NEg equal to 15.2, 18.6, and 22.7 MJ/kg EWG, respectively. They concluded that energy requirements for growth differed between the sexes and the difference was attributed to distinct NEg and partial efficiency of ME utilization for growth during the late growth phase. Ferreira et al. [61] conducted a comparative slaughter experiment in castrated male Saanen goats weighing 20-35 Kg and reported that NEm was 261.5 kJ/kg0.75 BW, MEm was 404.2 kJ/kg0.75 BW, and the partial efficiency of the use of ME for NE was calculated to be 0.65. The minimal endogenous N losses were 262 mg N/kg0.75 EBW, corresponding to a NPm of 1.39 g/kg0.75 BW. They reported that NEg increased by 40% (from 12 to 17 MJ/kg EBW gain) and NPg decreased by 3% (from 166 to 160 g/kg EBW gain) as the BW increased from 20 to 35 kg. Almeida et al. [62] proposed the possibility of using body composition to predict maturity as the mature weight is known to affect protein and energy requirement in goats.
The NEm for non-pregnant and non-lactating pubertal female Saanen goats was suggested to be 52 kcal/kg0.75 of BW [63]. The NEg increased from 3.5 to 4.7 Mcal/kg of BW gain as BW increased from 30 to 45 kg. They concluded that that the guidelines of the major feeding systems for the entire growth phase may not be adequate for females at pubertal phase. For weaned male and female Saanen goats, Sex did not affect NEg and NPm (277.8 kJ/kg0.75 BW/d and 2.98 g CP/kg0.75 BW/d respectively), as well as NPg (180.9 g/kg EWG) in Saanen goat kids [64]. However, castrated males and females had similar NEg (varied from 12.6 to 17.9 MJ/kg EWG), but greater than intact males (varied from 9.74 to 10.7 MJ/kg EWG), as the BW increased from 15 to 30 kg. From the slaughter experiments, Teixeira et al. [65] concluded that maintenance requirement for F1 Boer × Saanen goat kids were greater than published values and growth requirements were driven by efficiencies of deposition and largely dependent upon changes in body composition. In their study, NEm, MEm and partial efficiency of use of MEm for NEm were 321.6 kJ/ kg0.75 BW, 525.9 kJ/ kg0.75 BW, and 0.61, respectively. The NPm, MPm and the partial efficiency of MPm for NPm were 2.43 g/ kg0.75 BW, 4.41 g/ kg0.75 BW, and 0.55, respectively. They also reported that the partial efficiency of the utilization of ME to NE for growth was 0.32, and the partial efficiencies of the utilization of ME for the synthesis of protein and fat were 0.19 and 0.59, respectively. Multiparous pregnant goats (n = 66) were utilized in a comparative slaughter experiment to determine the energy and protein requirements for the maintenance of pregnant dairy goats [66]. Days of pregnancy (DOP) did not affect NEm or MEm which were 197 and 315 kJ/kg EBW, respectively, and the efficiency of ME utilization for maintenance (km) was 0.63. The DOP did not affect NPm estimated using the comparative slaughter technique (1.38 g/kg EBW) or using N balance (2.49 g/kg EBW). The MPm estimated using the comparative slaughter technique was not affected by DOP and was 3.22 g MP/kg EBW. The efficiency of MP utilization for maintenance (km) was 0.43. The efficiency of ME utilization for pregnancy (kp) increased with the progress of pregnancy and was 0.058, 0.10, and 0.19 at 80, 110, and 140 DOP, respectively. Similarly, the efficiency of MP utilization for pregnancy (kp) increased with DOP and was 0.12, 0.21, and 0.43 at 80, 110, and 140 DOP, respectively. There was no evidence that pregnancy affected NEm, MEm, NPm, and MPm or km and kp, which were also unaffected by DOP. However, km and kp increased with pregnancy progress as a response to the physiological changes in pregnant females.
To investigate the effects of sex on the requirements for maintenance and efficiency of energy utilization in growing Saanen goats, Souza et al. [67] analyzed data (n=238) from 7 slaughter experiments and concluded that NEm and MEm estimated by the comparative slaughter technique were greater in males than in females (75.0 kcal/kg0.75 EBW for males and 63.6 kcal/kg0.75 EBW for females). Sex did not affected NEM when degree of maturity was considered. Different sexes of Saanen goats have different energy utilization for growth (kg =0.31 for castrated males and females, and 0.26 for intact males), but similar retained energy as protein (kp = 0.21) and as fat (kf = 0.80), and efficiency of ME utilization for maintenance (km=0.63). Using both the comparative slaughter technique (n = 185) and N balance method (n = 136) under a meta-analytical approach, Souza et al. [68] suggested that there was no evidence that sex affects the protein requirements for maintenance and efficiencies of protein utilization and proposes new equations for NPm. The daily NPm estimated was 1.23 g/kg0.75 BW when using the comparative slaughter technique, while it was 3.18 g/kg0.75 BW when using the N balance methos for growing Saanen goats. The MPm estimated was 3.8 g/ kg0.75 BW, the kpm calculated as NPm/MPm was 0.33, and the kpg calculated as NPg/MPg was 0.52. Santos et al. [69] proposed prediction equations to estimate microbial protein synthesis to be used for the calculation of rumen degradable protein (RDP) requirements from MP in sheep and goats. They found no differences to fit metabolizable crude protein (MCP) efficiency between sheep and goats, and suggested that MCP synthesis in the rumen can be predicted from energy intake using combined equations that encompass both sheep and goats. With the Akaike Information Criterion Index ranged from 2,755 to 3,007, they proposed that the prediction could be based on total digestible nutrients (TDN), digestible organic matter (DOM), or ME intake by the following equations: MCP (g/day) = 12.7311 + 59.2956 × TDN intake; MCP (g/day) = 15.7764 + 62.2612 × DOM intake; and MCP (g/day) = 12.7311 + 15.3000 × ME intake.
5. Macromineral Requirements in Sheep and Goats
A meta-analysis (n = 154 from comparative slaughter experiments, n = 160 from minimum endogenous losses experiments) was conducted to evaluate the effects of sex on the daily net requirements of calcium (NCam), phosphorus (NPhm), potassium (NKm), and magnesium (NMgm) for maintenance in Saanen goats from 5 to 45 kg BW [70]. It was concluded that using the comparative slaughter technique, the results revealed that sex did not affect NCam, NPhm, or NKm (21.1, 22.8, and 4.0 mg/kg BW, respectively), but NMgm of intact males was greater than castrated males or females (2.6 versus 1.4 mg/kg BW). When minimum endogenous losses method was used, sex did not affect the NCam, NKm, or NMgm (38.0, 25.2, and 7.4 mg/kg BW, respectively). They concluded that Mg requirement for maintenance should consider sex difference, but not Ca, P, and K requirements for maintenance in goats. They also pointed out that P requirements for maintenance were significantly lower than current feeding system recommendations. Daily maintenance requirements, calculated using the comparative slaughter technique, were estimated as 32.3 mg Ca, 30.8 mg P, 1.31 mg Mg, 8.41 mg K, and 5.14 mg Na/kg of empty EBW [70]. Net requirements for growth increased from 6.2 to 6.6 g Ca, 5.3 to 5.4 g P, and 0.29 to 0.30 g Mg and decreased from 1.20 to 1.07 g K and 0.65 to 0.59 g Na/kg EWG for kids from 20 to 35 kg BW. This study indicated that the net mineral requirements for Boer crossbred goat kids may be different from those of purebred or other genotypes, and more data are needed for goats in general [71]. The Ca, P, Mg, K, and Na requirements of Saanen goats were estimated from two experiments (n = 75 in comparative slaughter experiment, and n = 58 in growth trial) [72]. They reported that the daily net macromineral requirements for maintenance did not differ among the sexes, and the average values obtained were 35.4 mg Ca, 24.7 mg P, 2.5 mg Mg, 5.0 mg K, and 3.30 mg Na /kg BW/day. The net Ca, P, and Mg requirements for growth were not different among the sexes, but sex could affect net K and Na requirements for growth. They also reported that the net K requirements for growth (g/kg ADG) of intact males were greater and increased approximately 16%, whereas females and castrated males decreased approximately 11%, as BW increased from 15 to 30 kg BW. The net Na requirements for growth (g/kg ADG) increased 9.5% for intact males and decreased 22% for females when the goats grew from 15 to 30 kg BW. They concluded that sex affected net K and Na requirements for growth, but it did not affect net macromineral requirements for maintenance in Saanen goats.
Vargas et al. [73] compiled data from six comparative slaughter experiments (n= 209) and analyzed the effects of sex on the net requirements of growth for Ca (NCag), P (NPg), Na (NNag), K (NKg), and Mg (NMgg) in Saanen goats from 5 to 45 kg BW, with or without the consideration of the degree of maturity. Without considering the degree of maturity, they reported that sex did not affect NCag, NPg, and NNag. When the degree of maturity was considered, NCag and NPg of intact males were greater than those of castrated males and females, and NNag of males (castrated and intact) was greater than that of females. The NCag and NPg remained constant, whereas NNag decreased by 32% as BW changed from 5 to 45 kg. The NMgg of castrated and intact males were greater than that of female goats, regardless of maturity. The NMgg of castrated and intact males increased by 8% and 15%, respectively, whereas that of females decreased by 8% as BW ranged from 5 to 45 kg. They concluded that elucidation of sex effects on macromineral requirements for growth may be useful for improving the accuracy of recommendations for mineral requirements for dairy goats, and the consideration of maturity stage across sexes in should be considered [73]. There were indications that mineral requirements may be different between sheep and goats. Wilkens et al. [74] provided an overview of similarities and differences between sheep and goats. They concluded that ruminal Ca absorption and renal Ca excretion were not affected by dietary Ca supply in both sheep and goats, but goats were able to compensate for the low Ca availability and sheep could not. As a response to dietary Ca restriction, sheep had a smaller increase in plasma calcitriol but a greater increase in the circulating concentration of a bone resorption marker than goats. The Ca and P concentrations in ruminal and abomasal fluids and in saliva were reported being different between sheep and goats, with a greater salivary P secretion in goats than in sheep. In the peripartal period, differences in the contribution of gastrointestinal Ca absorption and bone mobilization to the maintenance of Ca homeostasis were observed in lactating and non-lactating animals of both species.
6. Estimating Nutrient Requirements
6.1. Physiological Functions and Classifications
Maintenance, growth, lactation, pregnancy, activity, breeding, and fiber growth are major physiological and/or productive functions considered for the determination of nutrient requirements for sheep and goats. In sheep, NRC [3] classifications of nutrient requirements include mature ewes, yearling farm ewes, yearling range ewes, rams (maintenance and prebreeding), growing lambs and yearling (early and late maturing). Mature ewes include four additional subclassifications: maintenance, breeding, gestation (early, late; single, twin and triplet), and lactation (early, mid, and late; single, twin, triplet or more) [3]. In goats, NRC classifications of nutrient requirements differentiate Angora goats from dairy and nondairy goats [3]. For dairy and non-dairy mature does, it is further classified to maintenance, growth, breeding, gestation (early, late; single, twin and triplet) and lactation (early, mid, and late; single, twin, triplet or more). For dairy and non-dairy mature bucks, it is classified as maintenance and prebreeding. For Angora goats, the classification include growing Angora kids (male and female), mature Angora males (maintenance and breeding), and mature angora females (maintenance, breeding, gestation and lactation). Gestation is further classified as early and late gestation and fetal size (single, twin and triplet); and lactation as early (single, twin) and late lactation.
A number of factors affecting nutrient requirements in sheep and goats have been described [3]. The list includes species, level of Intake, genotype, mature size, age, gender, body composition, use of tissue energy, activity, season, acclimatization, heat and cold extremes. Furthermore, parasitism can affect nutrient requirements of sheep and goats [3]. It is known that nutrient requirements in sheep and goats can be influenced by the environment [75]. As the effect of climate changes on the environment intensifies, it is likely that precisely estimation of nutrient requirements in sheep and goats will becoming even more important to minimized greenhouse gas emission and carbon footprint in the future.
6.2. Estimating Energy Requirements
Estimation of energy requirements in sheep and goats are based on two fundamental laws of thermodynamic and portion of Einstein’s theory of relativity. Firstly, energy input must equal to energy output plus or minus any changes in body energy (Energy cannot be created or destroyed). Secondly, no transformation of energy is 100% efficient and the inefficiency is lost as heat (Entropy of the universe always increases). By applying these two principles it affords the equivalence between mass and energy and allow the conversion of body mass (kg) to energy (calories). Energy utilization by ruminants including sheep and goats can be partitioned into intake energy (IE), digestible energy(DE), metabolizable energy (ME), and net energy (NE) (Figure 1), and has been described and discussed [76]. A number of conversions have used in establishing energy requirement in sheep and goats [3]. Those include: DE (Mcal) = 4.4 TDN (kg), ME (Mcal) = DE (Mcal) x 0.82, estimate of heat production, conversion of ME to NEM, conversion of ME to NEg, conversion of ME to NEL, and 1 Mcal = 4.184 MJ.
6.3. NRC Equations for Predicting Energy Requirements for Sheep and Goats
A number of important equations have been used to estimate energy requirements in sheep [3]:
For maintenance, unadjusted:
0.062 Mcal NE/kg SBW0.75 ; (MEm x 0.644 = NEm)
For growth:
NEg, Mcal/d = ADG, kg x TEC avg; TEC avg, Mcal/kg = (TEfin – TE init)/(FBWfin – FBWinit); (MEg x 0.6 = NEg)
For lactation:
NEl, Mcal/d = (251.73 + (89.64 x MFC, %) + (37.85 x (MPC, %)/0.95))) x 0.001 x MY, kg/d ; (MEl x 0.644 = NEl)
A number of important equations have been used to estimate energy requirements in goats [3]:
For maintenance, unadjusted:
101 - 149 kcal ME/kg FBW0.75 ; (ME x 0.644 = NE)
For growth:
MEg = 3.20 - 6.81 kcal/g ADG; MEtg = 8.89 kcal/g ADG (Angora); MEf = 37.5 kcal/g ADG (Angora)
For lactation:
where SBW is shrunk body weight, MEm is metabolizable energy required for maintenance, NEm is net energy for maintenance, NEg is net energy for gain, ADG is average daily gain, TECavg is average energy concentration on a full body weight basis during the feeding period, TEfin is final tissue energy, TE init is initial tissue energy, FBWfin final full body weight, FBWinit is initial full or unshrunk body weight, MEg is metabolizble energy used for tissue gain, NEl is net energy for lactation, MFC is milk fat concentration, MPC is milk true protein concentration, MEl is metabolizable energy used for lactation, MEtg is metabolizable energy used for nonfiber tissue gain, FCM is fat-corrected milk.
MEl = 1.25 Mcal/Kg 4%FCM
6.4. Estimating Protein Requirements
Nitrogen utilization in ruminants has been described [77]. Protein digestion in sheep and goats (Figure 1) is largely adopted from the model developed by Satter and Roffler [78], although there may be small difference in digestion kinectics such as rumen turnover rate between small and large ruminants [79,80].
Figure 2.
Protein digestion in Sheep and Goats (Adapted from [78]).
Figure 2.
Protein digestion in Sheep and Goats (Adapted from [78]).
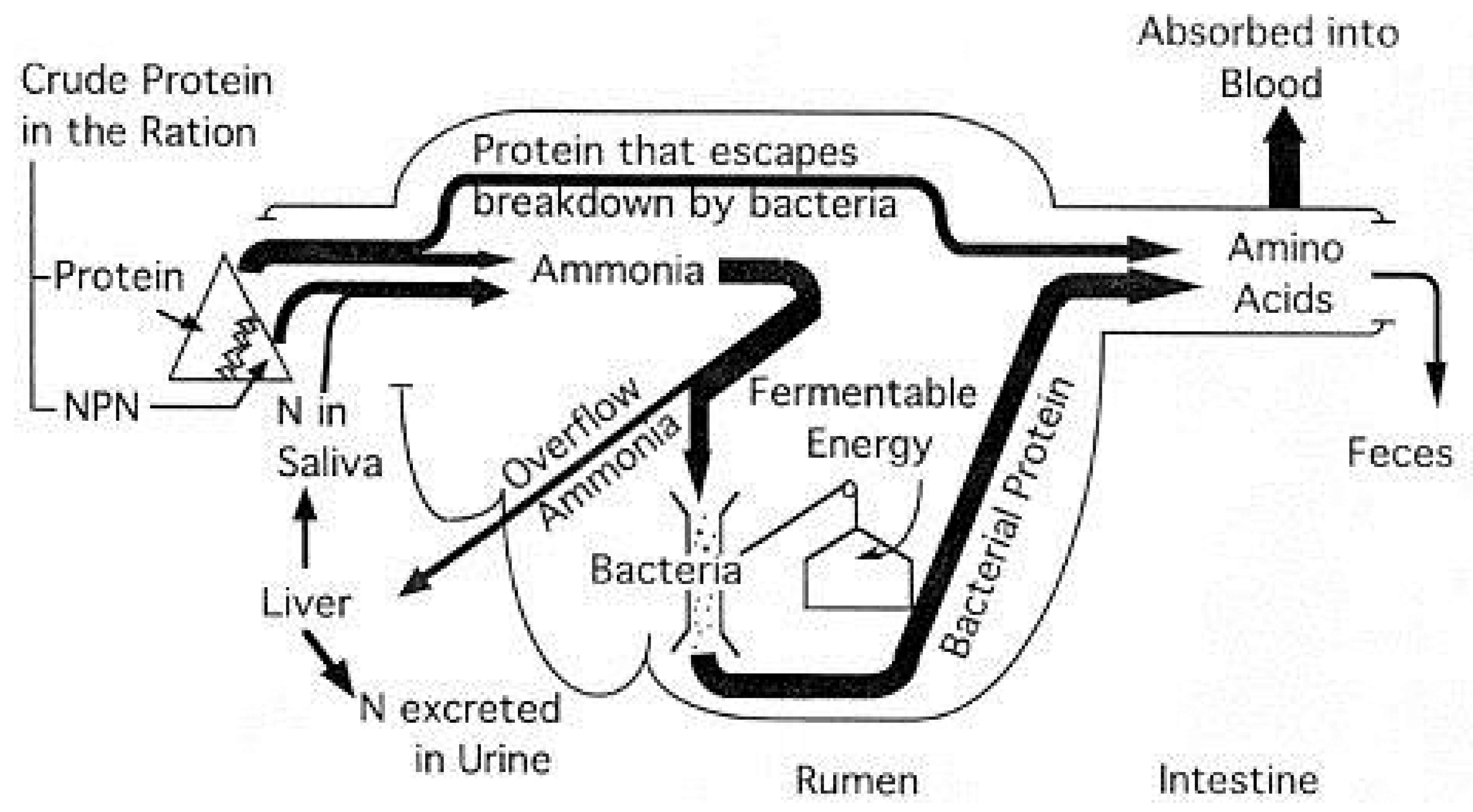
Intake protein (IP) is consisted of true protein and nonprotein nitrogen (NPN). Undegradable Intake Protein (UIP) is defined as 80% digestible and termed as Degradable Intake Protein (DIP) [3]. Digestible Protein (DP) is the difference between IP and Fecal Protein (FP) and is less useful to predict productive performance in ruminants due to microbial digestion and synthesis. Metabolizable protein (MP, true protein and AA that is digested postruminally) is the summation of UIP and Microbial Protein. Crude protein (CP) or IP is the summation of Ruminal Degradable Protein (RDP or DIP) and Ruminal Undegradable Protein (RUP or UIP). Therefore, RUP requirement = (MP required – MP bacteria – MP endogenous) / Diet RUP digestibility [3].
There are a number of assumptions used pertaining to the efficiency of conversation in estimate protein requirements in sheep and goats [3]. Efficiency of dietary MP for Maintenance (Kpm) is equal to 1.0. Efficiency of MP for Maintenance (Kpm) is similar for Metabolic Fecal Crude Protein (MFCP), Endogenous Urinary Crude Protein Loss (EUCP), and scurf CP. Range of Kpm from 0.67 to 1.00 is due to correction of MFCP for microbial CP. Kpm may vary with age and may be lower with greater intake. Efficiency of MP for Growth (Kpg) ranges from 0.5 [15] to 0.7 [81]. Kpg may vary with age, BW and body condition score. Efficiency of dietary MP for tissue gain in lactating goats is equal to 0.59. Efficiency of MP for Lactation (Kpl) varies: 0.58 [43], 0.59, 0.64 [33], 0.67 [82], 0.68 [39], 0.7 [81]. Efficiency of mobilized tissue protein for lactation (Kpl-t) is yet to be defined. Efficiency of mobilized MP for milk protein synthesis is equal to 0.69. Efficiency of MP for Fiber Growth (Kpf) varies: 0.26 [39], 0.48 [83], 0.5 [15], 0.6 [81]. Kpf may not be the same for wool sheep and angora goats and may be different between diet and mobilized tissue. Efficiency of mobilized MP for clean mohair fiber synthesis is equal to 0.61. Estimation of scurf and fiber crude protein requirements varied from 0.1125 g/kg BW0.75 [39] to 0.2 g/kg BW0.60 [82,84], as fiber production can be affected by season, breed, species, and frequency of shedding. Efficiency of MP for Pregnancy and Gestation (Kppreg) varies: 0.33 [34], 0.4 [43], 0.65 [85], 0.70 [81], 0.85 [39,40]. Kppreg may not be the same for sheep and goats and may be influenced by the development of mammary gland.
It is worthy to mention that evidences affecting the estimation of protein requirements have been emerging. Plant secondary metabolites can decrease activities of intestinal enzymes and interacting with epithelium lining that may ultimately affect the estimation of protein requirement [3]. Internal parasites can result greater use of AA for the synthesis of immunoglobulins and cytokines, increase replacement and repair of GI tract tissues with higher rates of liver protein synthesis can also affect the estimation of protein requirement [3].
6.5. NRC Equations for Predicting Protein Requirements in Sheep and Goats
A number of important equations have been used to estimate metabolizable protein requirements in sheep [3]:
For maintenance, plus fiber:
MPm, g/d = (SF – CPE/0.6) + (U-CPE/0.67) + (F-CPE/0.67)
For growth:
MPg, g/d = NPg /0.7; NPg, g/d = ADG, g x TPFr x 0.92
For lactation:
MPl-d, g/d = NPl-d /0.58: NPl-d, g/d = (10 x MTPC, % x MY, kg/d) – (ADG, g x TPFr)
A number of important equations have been used to estimate protein requirements in goats [3]:
For maintenance, plus fiber:
MPm, g/d = MFCP + EUCP + (0.2 g/kg BW0.60)
For growth:
MPg, g/d = 0.290g/g ADG (Dairy and Indigenous); MPg, g/d = 0.404g/g ADG (Meat); MPg, g/d = 0.281g/g tissue gain (Angora)
For lactation:
MPl = 1.45 g/g
For Fiber:
where MPm is metabolizable protein required for maintenance, SF – CPE is scurf and fiber protein, U-CPE endogenous urinary crude protein, F-CPE is metabolic fecal crude protein, MPg is metabolizable protein available for gain and growth, NPg net protein for gain, ADG is average daily gain, TPFr protein concentration in tissue accreted or mobilized on an empty BW basis, MPl-d is dietary metabolizable protein required for lactation, NPl-d is net protein for lactation from the diet, MTPC is milk true protein concentration, MY is milk yield, MPl is metabolizable protein used for lactation.
MPclean mohair = 1.65 g/g
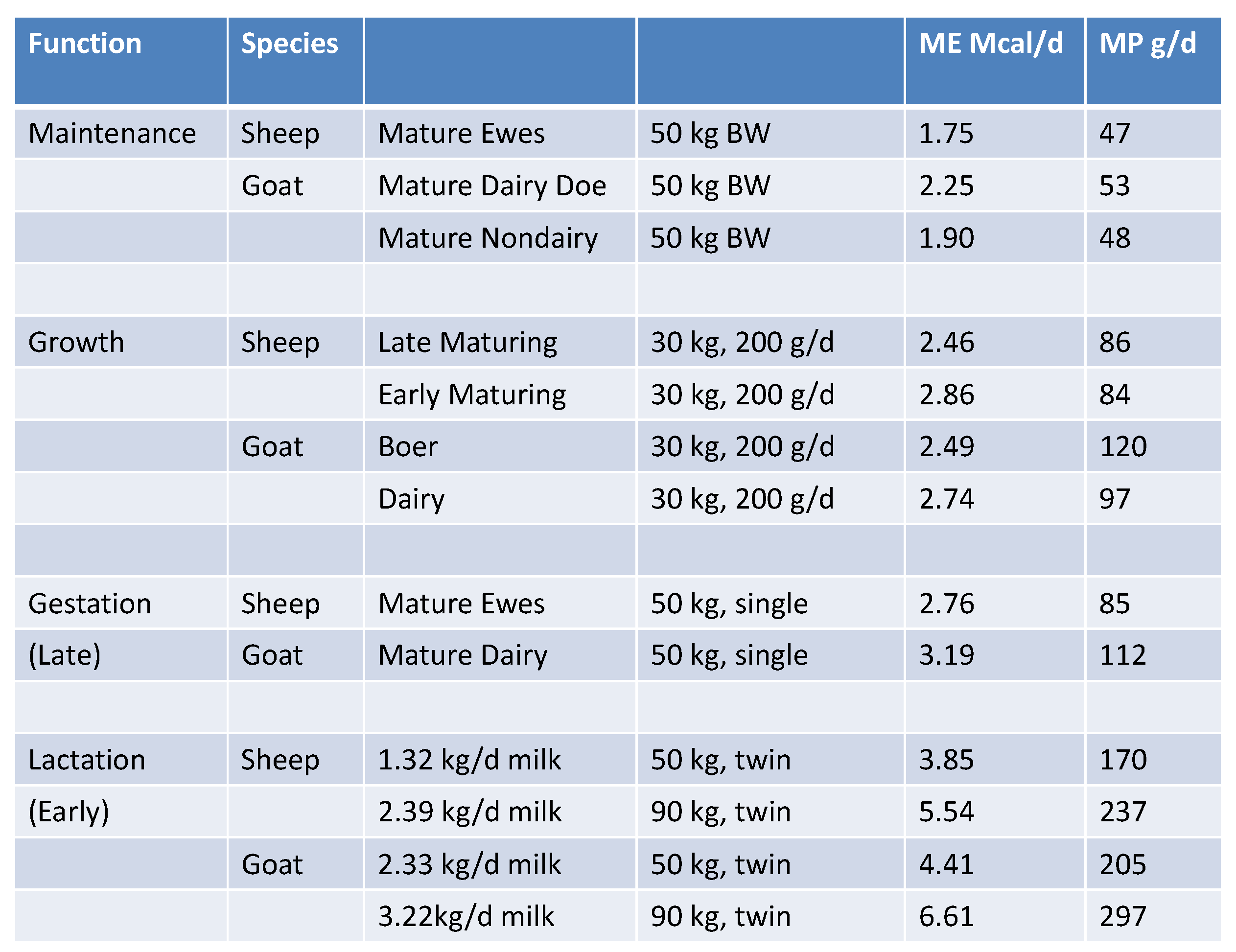
A simplified energy and protein requirements has been tabulated (Figure 3) for the purpose of comparison between sheep and goats. There are notable differences in both energy and protein requirements between these two species.
7. Limitations in Establishing Nutritional Standards
Because of a number of factors that has been identified to influence the determination of nutrient requirements, it is apparent that nutritional standards are relative, not absolute. It is targeted to a population but not to specific individual or a subset of group of animals, unless it is well defined at the beginning of the experiment. Nutritional standards can be determined through deduction or empirical methods. They are not always verified by additional animal experiments once established. These standards reflect the professional judgement or opinion of a group of experts during a particular period of time. In order to apply these standards effectively, the users must recognize the main sources of variation such as individual, genetic, climatic, dietary, age and physiological stages.
The basic methodology in the evaluation of nutrient requirements includes empirical and factorial approaches. Empirical method is based on observation or experience and is verifiable. Factorial method is related to a factor or integral parts of a factor. In the empirical method, nutritional requirements can be defined as the minimal amount of nutrients needed to maximize or minimize population responses for one or several performance criteria such as growth during a given time period [86]. In the factorial method, daily requirements can be calculated for an individual animal at a specific point in time by combining the estimated requirements for maintenance and production such as growth and milk production. The empirical method estimates optimal nutrient allowances from a population perspective, whereas the factorial method estimates the needs of a reference animal during a very short period of time [86]. There is always a need to reconcile the difference in nutrient requirements between empirical and factorial methods.
Advanced methodology in the evaluation of nutrient requirement can involve calorimetry in measuring energy intake and expenditure (direct and indirect), whole body dynamic non-invasive detection of body composition such as fat and lean mass imaging (magnetic resonance imaging, computed tomography, positron emission tomography, single photon emission tomography), system biology using mathematical and statistical modeling, and Integrating large data sets to interpret and understand complex physiological systems [5], and nutrigenomics. Application of Artificial intelligence (AI), with powerful machine and deep learnings in data analysis, has contributed to problem solving in agriculture, including predicting nutrient requirements in animals. Tedeschi [87] discussed the usefulness and limitation of mathematical modeling in predicting nutrient requirements in ruminants. He cautioned the less mechanistic modeling approaches such as AI for the replacement of mechanistic learning and system-thinking approaches.
8. Nutrigenomics and Nutrient Requirements
Advancement in molecular nutrition and especially nutrigenomics can have important impact on nutrient requirements. Molecular nutrition may be defined as a science concerned with the effect of nutrients and foods/food components on whole body physiology and health status at a molecular and cellular level [5]. Descriptive and mechanistic studies using state of the art epidemiology, food intake registration, genomics with single nucleotide polymorphisms (SNPs) and epigenomics, transcriptomics, proteomics, metabolomics, advanced biostatistics, imaging, calorimetry, cell biology, challenge tests (meals, exercise, etc.), and integration of all data by systems biology, will provide insight on a much higher level than today in a field we may name molecular nutrition research [5]. The concept of molecular nutrition research is broader than nutrigenomics.
The precise determination of molecular mechanisms underlying animal health and disease offers a great potential for promoting health, and lowering mortality and morbidity, and give to the rise of the science of nutrigenomics. Nutrigenomics is the study the effect of food constituents on gene expression and is broadly defined as the relationship between nutrients, diet, and gene expression [4]. It can identify molecular interaction between nutrients and other dietary bioactives with the genome. It also focus on the influence of genetic variation on nutrition and correlate gene expression or SNP (a DNA sequence variation occurring commonly within a population in which a single nucleotide — A, T, C or G — in the genome or other shared sequence differs between members of a biological species or paired chromosomes) with a nutrient’s absorption, metabolism, elimination or biological effects. It has the potential to optimize and customize nutrition with respect to subject’s genotype. It is promising and can be useful for disease prevention, health promotion and anti-aging. Although nutrigenomics in sheep and goats is considered as its infancy, there are recent contributions to the subject [88,89].
Nutrigenomics and other omics (genomics, epigenomics, transcriptomics, proteomics and metabolomics) and bioinformatics tools are poised to accelerate our understanding of the multiple levels of regulation induced in small ruminants by dietary nutrients during their utilization for milk, meat, wool, or reproduction [6]. Initial data indicate that the nutrigenomics approach may eventually lead to more precise management of goats and sheep, hence, helping to improve utilization of feed resources in a more optimal fashion [6], therefore, affecting the estimation of nutrient requirements. Nature largest gene depository resides in bacteria. Rumen microbes is an integrated part of nutrient digestion and utilization in sheep and goats. Understand of microorganism in the rumen can enhance the understanding of nutrient utilization and subsequently improve the precision of determination of nutrient requirements in sheep and goats. Total genetic composition of rumen microbes is consisted of core genome and accessory genome. Separation of commonly share genes (core genome) from strain specific genes (accessory genome) will certainly contribute to a better understanding of microbial interaction with nutrients. Precision nutrition is another arena that can deliver nutrients precisely to individual animal and have potential to meet nutrient requirements for the maintenance and production of meat, milk and fiver in sheep and goats. The Use of EID (ear tag that can be read by handheld reader) and software (feed formulation, livestock administration, reproductive optimization, quality management) make it possible to address individual needs of a particular animal. Readers are connected to the internet to process the data immediately for individual anima. The system recognizes individual properties of each animal and uses advanced technologies to optimize animal production. It enables the possibility to customize individual animal’s nutritional needs and render application of nutrient requirements effective.
9. Conclusions
Nutrition is complicated. Defining nutrient requirements is a gateway to streamlining the mass nutritional information and enabling the practical application to animals. Therefore, it contributes to simplifying the complexity of nutrition at times. In spite of many variables, nutrient requirements of sheep and goats will continue to be an important focus for years to come. To justify the enormous investment, animal molecular nutrition or nutrigenomics will have to be linked more directly to the improvement of production efficiency via more precise prediction of nutrient requirement. To conserve resources and maintain long term sustainability, precision nutrition will continue to play a role. Meaningful research and discussion pertaining to implication of nutrigenomics and application of AI to nutrient requirements in sheep and goats can be useful in defining nutritional needs of not only the population but also individuals.
Funding
Not applicable.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lu, C.D. The role of goats in the world: Society, science, and sustainability. Small Rumin. Res. 2023, 227, 107056. [Google Scholar] [CrossRef]
- Gonza ́lez, L.A.; Kyriazakis, I.; Tedeschi, L.O. Review: Precision nutrition of ruminants: Approaches, challenges and potential gains. Animal 2018, 12, s246–s261. [Google Scholar] [CrossRef] [PubMed]
- NRC (National Research Council). Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids; National Academy Press: Washington, DC, 2007. [Google Scholar] [CrossRef]
- Chadwick, R. Nutrigenomics, individualism and public health. Proc. Nutr. Soc. 2004, 63, 161–166. [Google Scholar] [CrossRef]
- Norheim, F.; Gjelstad, I.M.F.; Hjorth, M.; Vinknes, K.J.; Langleite, T.M.; Holen, T.; Jensen, J.; Dalen, K.T.; Karlsen, A.S.; Kielland, A.; et al. Molecular Nutrition Research—The Modern Way Of Performing Nutritional Science. Nutrients 2012, 4, 1898–1944. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.S.; Vailati-Ribonib, M.; Palladinoc, A.; Luo, J.; Loor, J.J. Application of nutrigenomics in small ruminants: Lactation, growth, and beyond. Small Rumin. Res. 2017, 154, 29–44. [Google Scholar] [CrossRef]
- ul Haq, Z.; Saleem, A.; Khan, A.A.; Dar, M.A.; Ganaie, A.M.; Beigh, Y.A.; Hamadani, H.; Ahmad, S.M. Nutrigenomics in livestock sector and its human-animal interface-a review. Vet. Anim. Sci. 2022, 17, 100262. [Google Scholar] [CrossRef]
- National Academic of Science. The organization of National Research Council. Retrieved on March 14, 2024 from: http://www.nasonline.org/about-nas/history/archives/milestones-in-NAS-history/organization-of-the-nrc.html.
- NRC. Recommended nutrient allowances for sheep. Recommended nutrient allowances for domestic animals. National Academy Press: Washington, DC, USA, 1945. [CrossRef]
- NRC. Recommended nutrient allowances for sheep. (1st ed.). Recommended nutrient allowances for domestic animals. National Academy Press: Washington, DC, USA, 1949. [CrossRef]
- NRC. Nutrient requirements of sheep, 2nd ed.; National Academies Press: Washington, DC, USA, 1957. [Google Scholar] [CrossRef]
- NRC. Nutrient requirements of sheep. Nutrient requirements of domestic animals, 3rd ed.; National Academy Press: Washington, DC, USA, 1964. [Google Scholar]
- NRC. Nutrient requirements of sheep. Nutrient requirements of domestic animals, 4th ed.; National Academy Press: Washington, DC, USA, 1968. [Google Scholar]
- NRC. Nutrient requirements of sheep. Nutrient requirements of domestic animals, 5th ed.; National Academy Press: Washington, DC, USA, 1975. [Google Scholar] [CrossRef]
- NRC. Nutrient requirements of sheep. Nutrient requirements of domestic animals, 6th ed.; National Academy Press: Washington, DC, USA, 1985. [Google Scholar]
- NRC. Nutrient Requirements of Goats: Angora, Dairy, and Meat Goats in Temperate and Tropical Countries, 1981. [CrossRef]
- Lu, C.D.; Sahlu, T.; Fernandez, J.M. Assessment of energy and protein requirements for growth and lactation in goats. In Proceedings of the 4th International Conference on Goats, Brasilia, Brazil, 8–13 March 1987; Volume 2, pp. 1229–1248. [Google Scholar]
- Lu, C.D. Energy and protein nutrition in lactating dairy goats. In Proceedings of the 24th Pacific Northwest Animal Nutrition Conference, Boise, Idaho, USA; 1989; pp. 133–142. [Google Scholar]
- Lu, C.D.; Potchoiba, M.J. Feed intake and weight gain of growing goats fed diets of various energy and protein levels. J. Anim. Sci. 1990, 68, 1751–1759. [Google Scholar] [CrossRef]
- Sahlu, T.; Fernandez, J.M.; Lu, C.D.; Potchoiba, M.J. Influence of dietary protein on performance of dairy goats during pregnancy. J. Dairy Sci. 1992, 75, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Sahlu, T.; Fernandez, J.M.; Lu, C.D.; Manning, R. Dietary protein level and ruminal degradability for mohair production in Angora goats. J. Anim. Sci. 1992, 70, 1526–1533. [Google Scholar] [CrossRef]
- Fernandez, J.M.; Sahlu, T.; Lu, C.D.; Ivey, D.; Potchoiba, M.J. Production and metabolic aspects of nonprotein nitrogen incorporation in lactation rations of dairy goats. Small Rumin. Res. 1997, 26, 105–117. [Google Scholar] [CrossRef]
- Qi, K.; Lu, C.D. Sulfur and sulfur-containing amino acid requirements for meat, milk and mohair production in goats. In Proceeding of the 6th International Conference on Goats. Beijing, China. 1996, 2:537-548. Available at: https://www.researchgate.net/publication/307902682.
- Qi, K.; Lu, C.D.; Owens, F.N. Sulfate supplementation of Alpine goats: Effects on milk yield and composition, metabolites, nutrient digestibilities and acid-base balance. J. Anim. Sci. 1992, 70, 3551–3560. [Google Scholar] [CrossRef] [PubMed]
- Qi, K.; Lu, C.D.; Owens, F.N. Sulfate supplementation of Angora goats: Sulfur metabolism and interactions with zinc, copper and molybdenum. Small Rumin. Res. 1993, 11, 209–225. [Google Scholar] [CrossRef]
- Qi, K.; Lu, C.D.; Owens, F.N. Sulfate supplementation of growing goats: Effects on performance, acid-base balance, and nutrient digestibilities. J. Anim. Sci. 1993, 71, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Qi, K.; Lu, C.D.; Owens, F.N. Effects of sulfate supplementation on performance, acid-base balance, and nutrient metabolism in Alpine kids. Small Rumin. Res. 1994, 15, 9–18. [Google Scholar] [CrossRef]
- Qi, K.; Lu, C.D.; Owens, F.N.; Lupton, C.J. Sulfate supplementation of Angora goats: Metabolic and mohair responses. J. Anim. Sci. 1992, 70, 2828–2837. [Google Scholar] [CrossRef] [PubMed]
- Qi, K.; Lu, C.D.; Owens, F.N.; Lupton, C.J. Effects of sulfate supplementation on performance, acid-base balance, and nutrient metabolism in Angora kids. Small Rumin. Res. 1994, 15, 19–29. [Google Scholar] [CrossRef]
- Luo, J.; Goetsch, A.L.; Nsahlai, I.V.; Johnson, Z.B.; Sahlu, T.; Moore, J.E.; Ferrell, C.L.; Galyean, M.L.; Owens, F.N. Maintenance energy requirements of goats: Predictions based on observations of heat and recovered energy. Small Rumin. Re. 2004, 53, 221–230. [Google Scholar] [CrossRef]
- Luo, J.; Goetsch, A.L.; Sahlu, T.; Nsahlai, I.V.; Johnson, Z.B.; Moore, J.E.; Galyean, M.L.; Owens, F.N.; Ferrell, C.L. Prediction of metabolizable energy requirements for maintenance and gain of preweaning, growing and mature goats. Small Rumin. Res. 2004, 53, 231–252. [Google Scholar] [CrossRef]
- Luo, J.; Goetsch, A.L.; Nsahlai, I.V.; Sahlu, T.; Ferrell, C.L.; Owens, F.N.; Galyean, M.L.; Moore, J.E.; Johnson, Z.B. Prediction of metabolizable energy and protein requirements for maintenance, gain and fiber growth of Angora goats. Small Rumin. Res. 2004, 53, 339–356. [Google Scholar] [CrossRef]
- Nsahlai, I.V.; Goetsch, A.L.; Luo, J.; Johnson, Z.B.; Moore, J.E.; Sahlu, T.; Ferrell, C.L.; Galyean, M.L.; Owens, F.N. Metabolizable energy requirements of lactating goats. Small Rumin. Res. 2004. 53, 253–273. [CrossRef]
- Sahlu, T.; Goetsch, A.L.; Luo, J.; Nsahlai, I.V.; Moore, J.E.; Galyean, M.L.; Owens, F.N.; Ferrel, C.L.; Johnson, Z.B. Nutrient requirements of goats: Developed equations, other considerations and future research to improve them. Small Rumin. Res. 2004, 53, 191–219. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Beef Cattle, 8th rev. ed. National Academy Press, Washington, DC, 2016. [CrossRef]
- NRC. Nutrient Requirements of Dairy Cattle, 8th rev. ed. National Academy Press, Washington, DC, 2021. [CrossRef]
- AFRC (Agricultural and Food Research Council). Nutritive requirements of ruminant animals: Energy. AFRC Technical Committee on Responses to Nutrients, Report No. 5, Nutr. Abstr. Rev. Series B. 1990, 60:729-804.
- AFRC. Nutritive requirements of ruminant animals: Protein. AFRC Technical Committee on Responses to Nutrients, Report No. 9, Nutr. Abstr. Rev. Series B. 1992, 62:787-835.
- AFRC. Energy and protein requirements of ruminants. Advisory Manual, AFRC Technical Committee on Responses to Nutrients. 1993. CABI Publishing, Wallingford, Oxon, UK.
- AFRC. The nutrition of goats. AFRC Technical Committee on Responses to Nutrients. Report 10. 1998. CABI Publishing, Wallingford, UK, OCLC: 769705109.
- INRA (National Institute for Agricultural Research) Alimentation des Ruminants. R. Jarrige, ed. Versailles, France, 1978. INRA Publications.
- INRA (Institut National de la Recherche Agronomique) Alimentation des Bovins, Ovins et Caprins. INRA, Paris, France, 1988. INRA Publications.
- INRA (Institut National de la Recherche Agronomique). Ruminant Nutrition. Recommended Allowance and Feed Tables. R. Jarrige, ed. INRA, Paris, France, 1989. http://hdl.handle.net/102.100.100/256672?index=1.
- CSIRO. Feeding Standards for Australian Livestock. Ruminants. Commonwealth Scientific and Industrial Research Organization Publications: Melbourne, Australia. 1990.
- CSIRO. Nutrient requirements of domesticated ruminants. Commonwealth Scientific and Industrial Research Organization Publications: Collingwood, Victoria, Australia. 2007.
- Ma, T.; Deng, K.; Tu, Y.; Zhang, N.; Zhao, Q.; Li, C.; Jin, H.; Diao, Q. Recent advances in nutrient requirements of meat-type sheep in China: A review. J Integrative Agri 2022, 21, 1–14. [Google Scholar] [CrossRef]
- Galvani, D.B.; Pires, C.C.; Kozloski, G.V.; Wommer, T.P. Energy requirements of Texel crossbred lambs. J Anim Sci 2008, 86, 3480–3490. [Google Scholar] [CrossRef]
- Galvani, D.B.; Pires, C.C.; Kozloski, G.V.; Sanchez, L.M.B. Protein requirements of Texel crossbred lambs. Small Rumin Res 2009, 81, 55–62. [Google Scholar] [CrossRef]
- Salah, N.; Sauvant, D.; Archimède, H. Nutritional requirements of sheep, goats and cattle in warm climates: A meta-analysis. Animal 2014, 8, 1439–1447. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.T.; You, J.H.; Zhang, H.; Wang, F. Determination of energy and protein requirement for maintenance and growth and evaluation for the effects of gender upon nutrient requirement in Dorper × Hu crossbred lambs. Trop. Anim. Health Prod 2015, 47, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.P.; Pereira, E.S.; Biffani, S.; Medeiros, A.N.; Silva, A.M.A.; Oliveira, R.L.; Marcondes, M.I. Meta-analysis of the energy and protein requirements of hair sheep raised in the tropical region of Brazil. J. Anim. Physio. Anim Nutr. 2018, 101, e52–e60. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.S.; Lima, F.W.R.; Marcondes, M.I.; Rodrigues, J.P.P.; Campos, A.C.N.; Silva, L.P.; Bezerra, L.R.; Pereira, M.W.F.; Oliveira, R.L. Energy and protein requirements of Santa Ines lambs, a breed of hair sheep. Animal 2017, 11, 2165–2174. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.S.; Pereira, M.W.F.; Marcondes, M.I.; Medeiros, N.A.; Oliveira, R.L.; Silva, L.P.; Mizubuti, I.Y.; Campos, A.C.N.; Heinzen, E.L.; Veras, A.S.C.; et al. Maintenance and growth requirements in male and female hair lambs. Small Rumin Res 2018, 159, 75–83. [Google Scholar] [CrossRef]
- Mendes, M.S.; Souza, J.G.; Herbster, C.J.L.; Brito, N.A.S.; Silva, L.P.; Rodrigues, J.P.P.; Marcondes, M.I.; Oliveira, R.L.; Bezerra, L.R.; Pereira, E.S. Maintenance and Growth Requirements in Male Dorper × Santa Ines Lambs. Front Vet Sci 2021, 8, 676956. [Google Scholar] [CrossRef]
- Fernandes, M.H.M.R.; Resende, K.T.; Tedeschi, L.O.; Fernandes, J.S.; Silva, H.M.; Carstens, G.E.; Berchielli, T.T.; Teixeira, I.A.M.A.; Akinaga, L. Energy and protein requirements for maintenance and growth of Boer crossbred kids. J Anim Sci 2006, 85, 1014–1023. [Google Scholar] [CrossRef]
- Cannas, A.; Tedeschi, L.O.; Fox, D.G.; Pell, A.N.; Van Soest, P.J. A mechanistic model for predicting the nutrient requirements and feed biological values for sheep. J Anim Sci 2004, 82, 149–169. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, L.O.; Cannas, A.; Fox, D.G. A nutrition mathematical model to account for dietary supply and requirements of energy and other nutrients for domesticated small ruminants: The development and evaluation of a Small Ruminant Nutrition System. Small Rumin Res 2010, 89, 174–184. [Google Scholar] [CrossRef]
- Medeiros, A.; Resende, K.; Teixeira, I.; Araújo, M.; Yáñez, E.; Ferreira, A. Energy Requirements for Maintenance and Growth of Male Saanen Goat Kids. Anim Biosci 2014, 27, 1293–1302. [Google Scholar] [CrossRef]
- Bompadre, T.F.; Neto, O.B.; Resende, K.T.; Teixeira, I.A.M.A. Energy requirements in early life are similar for male and female goat kids. Asian-Australas J Anim Sci 2014, 27, 1712–1720. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.K.; Resende, K.T.; St Pierre, N.; Silva, S.P.; Soares, D.C.; Fernandes, M.H.M.R.; Souza, A.P.; Silva, N.C.D.; Lima, A.R.C.; Teixeira, I.A.M.A. Energy requirements for growth in male and female Saanen goats. J. Anim. Sci 2015, 93, 3932–3940. [Google Scholar] [CrossRef]
- Ferreira, A.C.D.; Yáñez, E.A.; Medeiros, A.N.; Resende, K.T.; Filho, J.M.P.; Fernandes, M.H.M.R.; Almeida, A.K.; Teixeira, I.A.M.A. Protein and energy requirements of castrated male Saanen goats. Small Rumin Res 2015, 123, 1, 88–94. [Google Scholar] [CrossRef]
- Almeida, A.K.; Resende, K.T.; Tedeschi, L.O.; Fernandes, M.H.M.R.; Regadas Filho, J.G.L.; Teixeira, I.A.M.A. Using body composition to determine weight at maturity of male and female Saanen goats. J. Anim. Sci. 2016, 94, 2564–2571. [Google Scholar] [CrossRef]
- Figueiredo, F.O.M.; Berchielli, T.T.; Resende, K.T.; Gomes, H.F.B.; Almeida, A.K.; Sakomura, N.K.; Teixeira, I.A.M.A. Energy requirements for growth of pubertal female Saanen goats. J. Anim. Physiol. Anim. Nutr. (Berl.) 2016, 100, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, F.O.M.; Leite, R.F.; St- Pierre, N.R.; Resende, K.T.; Almeida, A.K.; Souza, A.P.; Teixeira, I.A.M.A. Energy and protein requirements of weaned male and female Saanen goats. J. Anim. Physiol. Anim. Nutr. (Berl.) 2017, 101, 1014–1025. [Google Scholar] [CrossRef]
- Teixeira, I.A.M.A.; Fernandes, M.H.M.R.; Filho, J.M.P.; Canesin, R.C.; Gomes, R.A.; Resende, K.T. Body composition, protein and energy efficiencies, and requirements for growth of F1 Boer x Saanen goat kids. J. Anim. Sci. 2017, 95, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Harter, C.J.; Lima, L.D.; Silva, H.G.O.; Castagnino, D.S.; Rivera, A.R.; Resende, K.T.; Teixeira, I.A.M.A. Energy and protein requirements for maintenance of dairy goats during pregnancy and their efficiencies of use. J. Anim. Sci. 2017, 95, 4181–4193. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.P.; St-Pierre, N.R.; Fernandes, M.H.R.M.; Almeida, A.K.; Vargas, J.A.C.; Resende, K.T.; Teixeira, I.A.M.A. Energy requirements and efficiency of energy utilization in growing dairy goats of different sexes. J Dairy Sci 2020, 103, 272–281. [Google Scholar] [CrossRef]
- Souza, A.P.; Vargas, J.A.C.; Fernandes, M.H.M.R.; Almeida, A.K.; Resende, K.T.; Teixeira, I.A.M.A. Metabolizable Protein: 2. Requirements for Maintenance in Growing Saanen Goats. Front Vet Sci 2021, 8, 650203. [Google Scholar] [CrossRef]
- Santos, S.A.; Carvalho, G.G.P.; Azevedo, J.A.G.; Zanetti, D.; Santos, E.M.; Pereira, M.L.A.; Pereira, E.S.; Pires, A.J.V.; Valadares, F.S.C.; Teixeira, I.A.M.A.; et al. Metabolizable protein: 1. Predicting equations to estimate microbial crude protein synthesis in small ruminants. Front Vet Sci 2021, 8, 650248. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.A.C.; Almeida, A.K.; Souza, A.P.; Fernandes, M.H.M.R.; Harter, C.J.; Resende, K.T.; Teixeira, I.A.M.A. Macromineral requirements for maintenance in male and female Saanen goats: A meta-analytical approach. Livestock Sci 2020, 235, 104003. [Google Scholar] [CrossRef]
- Fernandes, M.H.M.R.; Resende, K.T.; Tedeschi, L.O.; Teixeira, I.A.M.A.; Fernandes, J.S. Macromineral requirements for the maintenance and growth of Boer crossbred kids. J. Anim. Sci. 2012, 90, 4458–4466. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.M.; Resende, K.T.; Teixeira, I.A.M.A.; Vargas, J.A.C.; Lima, A.R.; Leite, R.F.; Figueiredo, F.O.; Tedeschi, L.O.; Fernandes, M.H. Net macromineral requirements in male and female Saanen goats. J Anim Sci 2016, 9, 3409–3419. [Google Scholar] [CrossRef]
- Vargas, J.A.C.; Almeida, A.K.; Souza, A.P.; Fernandes, M.H.M.R.; Resende, K.T.; Teixeira, I.A.M.A. Sex effects on macromineral requirements for growth in Saanen goats: A meta-analysis. J Anim Sci 2017, 95, 4646–4657. [Google Scholar] [CrossRef]
- Wilkens, M.R.; Breves, G.; Schröder, B. A goat is not a sheep: Physiological similarities and differences observed in two ruminant species facing a challenge of calcium homeostatic mechanisms. Anim Prod Sci 2014, 54, 1507–1511. [Google Scholar] [CrossRef]
- NRC. Effect of Environment on Nutrient Requirements of Domestic Animals. National Academy Press, Washington, DC, 1981. [CrossRef]
- NRC. Nutritional Energetics of Domestic Animals and Glossary of Energy Terms. National Academy Press, Washington, DC, 1981. [CrossRef]
- NRC. Ruminant Nitrogen Usage. National Academy Press, Washington, DC, 1985. [CrossRef]
- Satter, L.D.; Roffler, R.E. Nitrogen requirement and utilization in dairy cattle. J. Dairy Sci. 1975, 58, 1219–1237. [Google Scholar] [CrossRef] [PubMed]
- Schlecht, E.; Richter, H.; Fernandez-Rivera, S.; Becker, K. Gastrointestinal passage of Sahelian roughages in cattle, sheep and goats, and implications for livestock-mediated nutrient transfers. Anim. Feed Sci. and Tech. 2007, 137, 93–114. [Google Scholar] [CrossRef]
- Ferreira, L.M.M.; Hervas, G.; Belenguer, A.; Celaya, R.; Rodrigues, M.A.M.; Garcia, U.; Frutos, P.; Osoro, K. Comparison of feed intake, digestion and rumen function among domestic ruminant species grazing in upland vegetation communities. J. Anim. Physiol. Anim. Nutr. (Berl). 2017, 101, 846–856. [Google Scholar] [CrossRef] [PubMed]
- SCA (Standing Committee on Agriculture, Ruminants Subcommittee). Feeding Standards for Australian Livestock. Ruminants. CSIRO Publications, East Melbourne, Australia, 1990.
- NRC. Nutrient Requirements of Dairy Cattle, 7th rev. ed. National Academy Press, Washington, DC, 2001. [CrossRef]
- Luo, J.; Goetsch, A.L.; Nsahlai, I.V.; Sahlu, T.; Ferrell, C.L.; Owens, F.N.; Galyean, M.L.; Moore, J.E.; Johnson, Z.B. Metabolizable protein requirements for maintenance and gain of growing goats. Small Ruminant Res. 2004, 53, 309–326. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Beef Cattle, 6th rev. ed. National Academy Press, Washington, DC, 1984. [CrossRef]
- NRC. Nutrient Requirements of Beef Cattle, 7th rev. ed. (Updated). National Academy Press, Washington, DC, 2000. [CrossRef]
- Hauschild, L.; Pomar, C.; Lovatto, P.A. Systematic comparison of the empirical and factorial methods used to estimate the nutrient requirements of growing pigs. Animal 2010, 4, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, L.O. Mathematical modeling in ruminant nutrition: Approaches and paradigms, extant models, and thoughts for upcoming predictive analytics. Journal of Anim. Sci. 2019, 97, 1921–1944. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.; Shao, Y.; Yu, Y.; Sha, K.; Jiang, Y.; Xian Zhang, X.; Zhong, Y.; Shi, H.; Li, W. The miR-214-5p/lactoferrin/miR-224-5p/ADAM17 axis is involved in goat mammary epithelial cell’s immune regulation. Animals 2023, 13, 2835. [Google Scholar] [CrossRef]
- Bilal Akyüz, B.; Sohel, M.M.H.; Konca, Y.; Korhan Arslan, K.; Gürbulak, K.; Abay, M.; Kaliber, M.; White, S.N.; Cinar, M.U. Effects of low and high maternal protein intake on fetal skeletal muscle miRNAome in sheep. Animals Accepted for publication. 2024, 14. [Google Scholar]
Figure 1.
Schematic Diagram of Energy Utilization by Sheep and Goats (Modified from [76]).
Figure 1.
Schematic Diagram of Energy Utilization by Sheep and Goats (Modified from [76]).
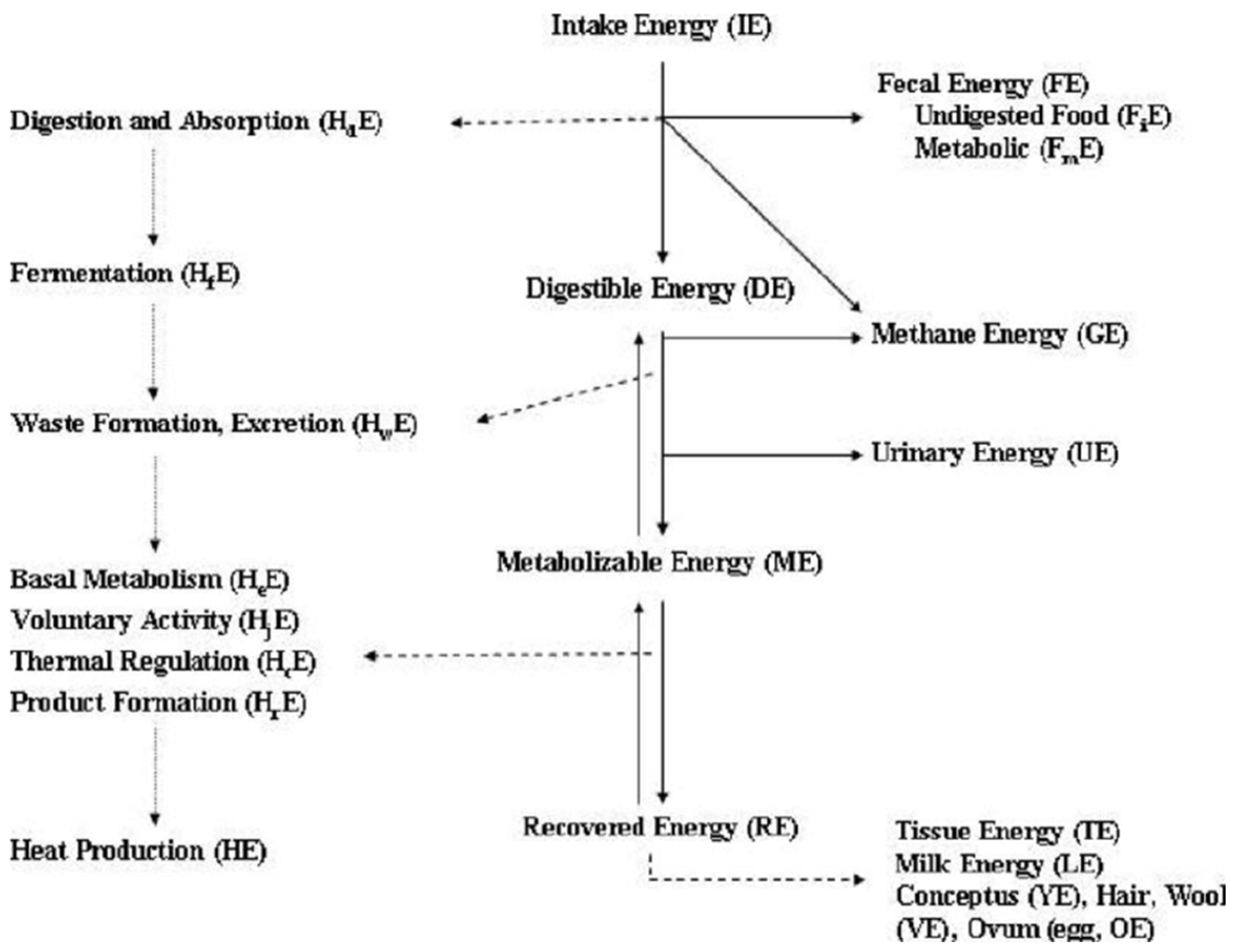
Figure 3.
Comparison of NRC [3] energy and protein requirements between Sheep and Goats.
Figure 3.
Comparison of NRC [3] energy and protein requirements between Sheep and Goats.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



