Submitted:
27 March 2024
Posted:
29 March 2024
You are already at the latest version
Abstract
Abstract Termites are eusocial arthropod decomposers that increase crop productivity and soil fertility. Because of the various benefits they possess, they are being exploited by humans. Termite infestations cause significant damage to structures and ecosystems. There are various physical and chemical methods for termite removal and control, but the use of chemical insecticides has a detrimental effect on human health and the environment, so efforts are directed towards the development of biological approaches to reduce their presence. Various effective and dependable biological approaches focused on using bacteria, fungi, and viruses have been explored, but termites continue to pose a major global threat. Recent advancements in molecular biology may provide a better solution for termites control. This review summarizes various biological methods to reduce the presence of termites in a sustainable manner.
Keywords:
Termites
; Pests
; Biocontrol
; Suistanable
1. Introduction
Termites (order Isoptera) are detriphagous eusocial pests that can cause detrimental effect to wooden infrastructure, buildings, crops, and forestry plantations. Despite their positive contribution to nutrient cycling via organic matter decomposition, improving soil fertility, water infiltrability, providing food and biofuel sources, termites are commonly known as economical pests (Figueirêdo et al., 2015; Siddiqui et al., 2023; Aidoo et al., 2023; Muon et al., 2023).
Annually, termites account for a global economic loss of USD 50 billion (Subekti et al., 2015). Over 3000 species of termites are known to exist today throughout the world.
According to Subekti et al., 2015, the USA has identified 50 species, of which 20 are classified as structural pests. In India, 337 species have been documented, of which 35 are known to cause harm to structures and agricultural products (Murthy et al., 2015). Termites are often classified into four types on the basis of their habitat and behaviour: subterranean, drywood, dampwood, and formosan. According to Rust and Su, 80% of the total damage is caused by subterranean termites.
To address these challenges, many physical and chemical termite management strategies are utilized around the world, but bulk of them rely on the use of chemical termiticides. However, excessive use of chemical insecticides has a negative influence on human life, animal health and non-target creatures such as beneficial insects and pollinators, as well as soil micro and macrofauna (Liu et al., 2011). Furthermore, the indiscriminate use of these chemical insecticides contributes to the emergence of insecticide resistance to bifenthrin, fipronil, cypermethrin, and deltamethrin, as well as soil and groundwater contamination, resulting in toxic residues being retained and bioaccumulated in the food chain for a long time. Thus, there is an urgent need to develop sustainable and environment friendly termite control methods as an alternative to chemical methods.
In recent years, the use of plant extracts and natural antagonists like fungi and bacteria as biocontrol agents (BCAs) has garnered significant interest as a sustainable termite management strategy. Nowadays, RNA interference (RNAi) has become a viable option for creating biological control tactics since it offers superior target-specificity for many species. Biological alternatives are highly efficient, inexpensive, safer for humans and create no pollution in the environment. An overview of the various biological agents used to combat termites is provided in this review.
2. Termites as a Noxious Agricultural and Industrial Economic Agent
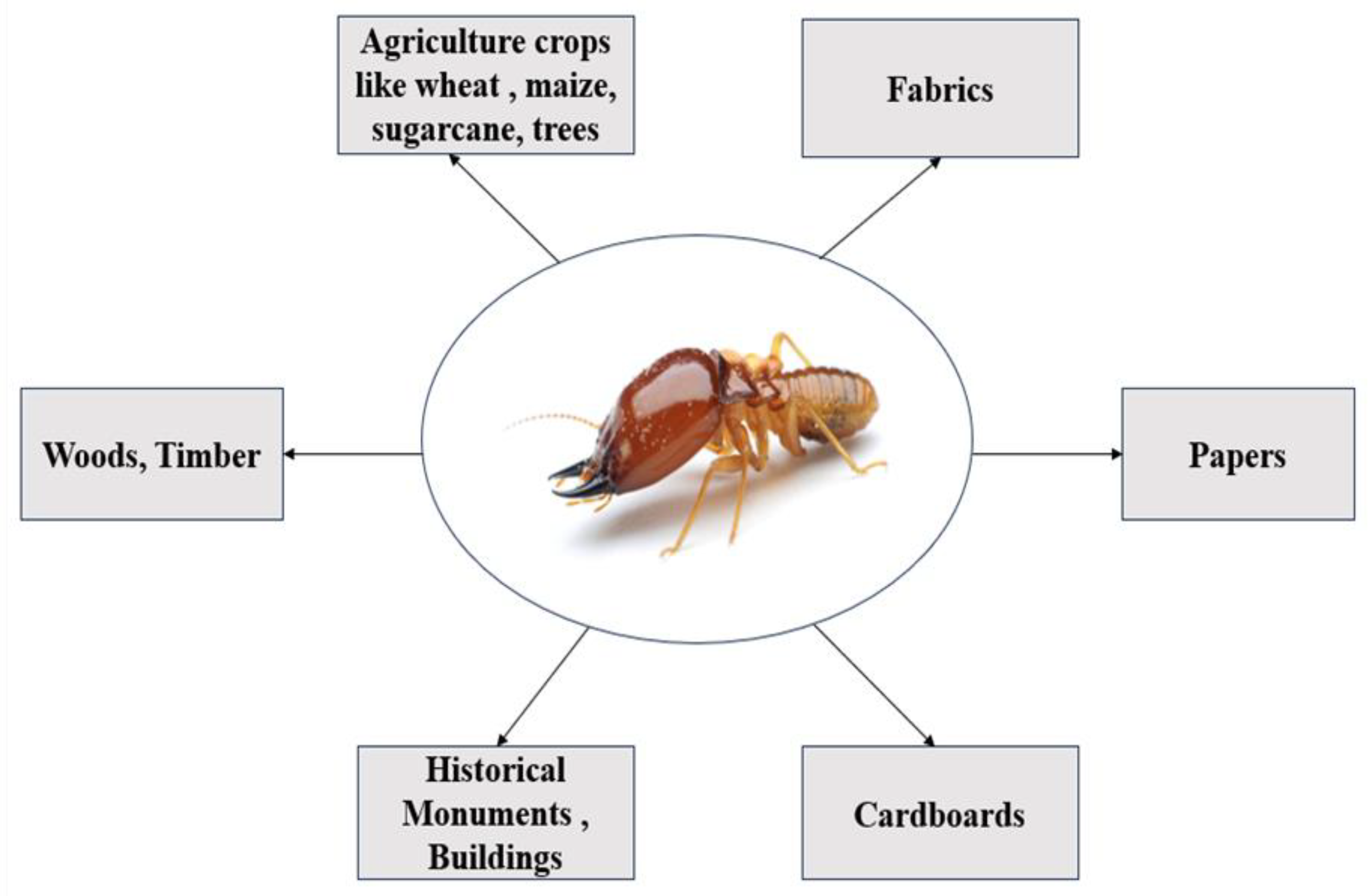
Termites are the primary decomposers of dead plant materials and they significantly contribute to soil nutrient turnover. When termites feed on buildings, electric cables, agriculture crops or plant materials used by humans, they act as noxious pests (Scharf et al., 2015; Wekhe et al., 2019; Azam et al., 2015). Termite management and economic losses cost hundreds of millions of dollars annually worldwide, as demonstrated in Figure 1
According to Smagghe, the world population is predicted to reach over 9 billion by 2050, which would also lead to 70% increase in agricultural and food output. Termites are highly destructive pests that attack various annual and perennial crops. Various crops damaged by termites in India are shown in Table 1.
Termites are the leading cause of wood decay in the natural environment. Heterotermes indicola is the primary subterranean termite in India that impact buildings and other infrastructure (Mahapatro & Chatterjee, 2018). According to recent studies, Heterotermes indicola caused 90% of instances of substantial damage to timber structures between 2009 and 2021 in a variety of areas, including Chandigarh, Delhi, Haryana, Jammu & Kashmir, West Bengal, Punjab, and Orrisa.
In India, it was also discovered that termites were attacking river dams present in the vicinity of Hirakund dam on the Mahanadi River in Orissa. Mahapatro & Chatterjee, 2018 stated that termites also damaged a whole village in Rajasthan near Kota. A very recent report from the Times of India reported that in U.P., termites devoured around Rs. 18 lakhs that were kept in a Bank of Baroda.
Different types of drywood and subterranean termites invaded botanical gardens, museums, and libraries in tropical regions. Drywood termites, such as Cryptotermes and Kalotermes, have been seen to live in books and paper stacks and dig tunnels under timber constructions (Nagaraju et al., 2021). Twenty percent of homes have termite damage, according to Kalleshwaraswamy et al., 2023. Nine termite species belonging to the families
Rhinotermitidae, Termitidae, and Kalotermitidae were responsible for 89% of the termite infestation in historic buildings and 62% of the infestation in residential buildings in India (Mello et al., 2014; Gusmao et al., 2014).
Figure 3.
Damages inflicted due to termites (Source: This study).
3. Management of Termites
In recent decades, a number of approaches for managing termites in diverse fields have been developed. Traditional termite treatment methods include physical and chemical treatments.
Figure 4.
Different approaches of Termites control (Source: This study).
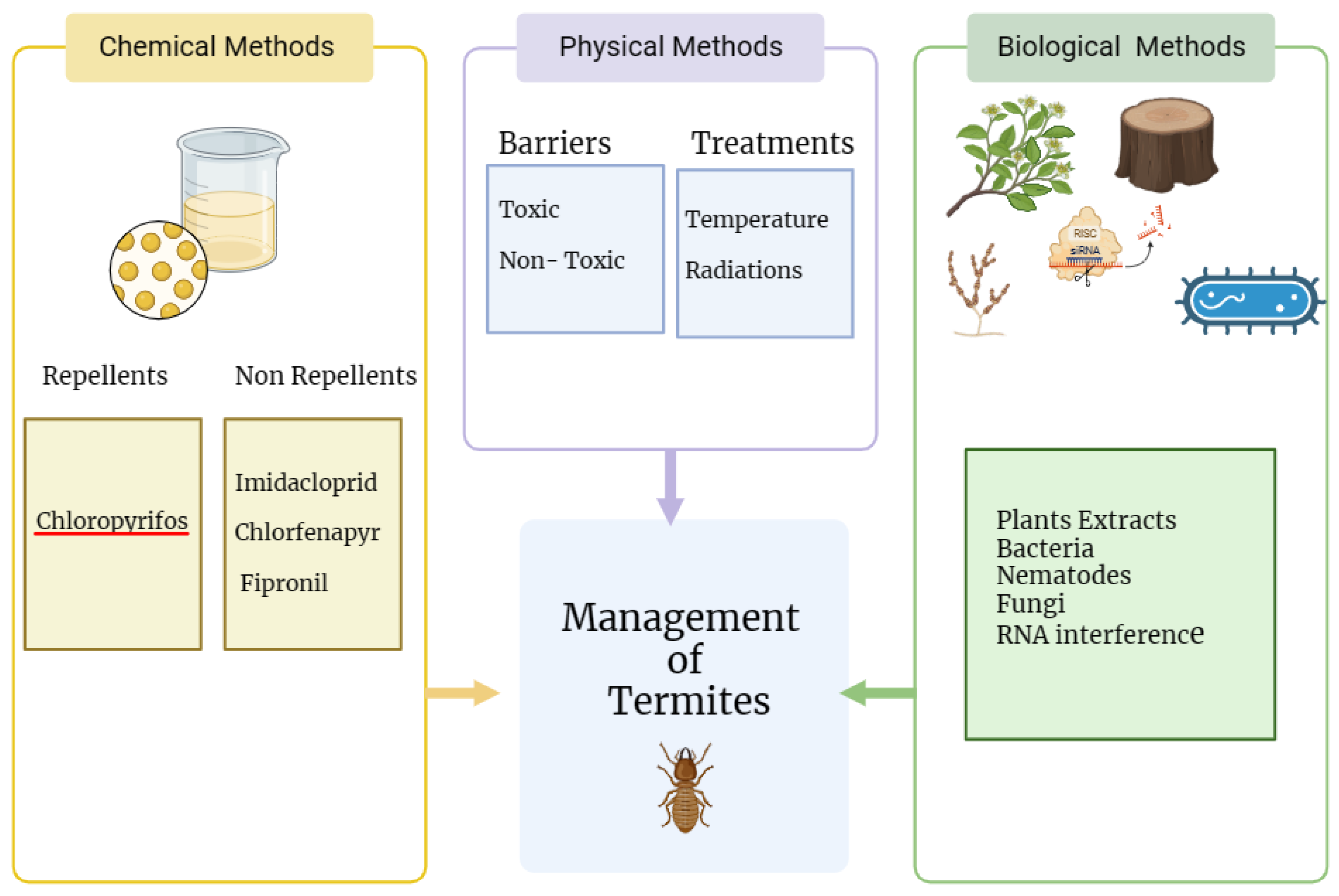
3.1. Physical Methods
Physical methods of termite removal involve the dequeening or removal of kings from the colony (Atsbha & Hintsa, 2018), the incorporation of materials like sand etc., freezing, heating, and the use of microwave radiation. Humidity also led to the removal of termites (Cao et al., 2016).
Incorporation of fine materials like sand crushed volcanic cinders, gravel aggregate, and Termi-Mesh, a tiny mesh constructed of stainless steel, has the ability to operate as a barrier against subterranean termites (Li et al., 2017). Several limitations of physical approaches have been identified, including their inability to eliminate termites quickly, the cost and time required to create physical barriers, and the need for regular inspections and maintenance.
3.2 Chemical Methods
The major way of managing subterranean termites is to treat the soil with chemical insecticides near the active infestation. Chemical barriers fall into two categories: repellent and non-repellent. Toxicity in non-repellents cannot be detected in advance, therefore the lethal impact is passed on to successive colony members (Ahmed et al., 2006). Non-repellent termiticides include chloranthraniliprole, imidacloprid, chlorfenapyr, and fipronil. Termites may detect the repellent compounds chlorpyriphos, bifenthrin, and permethrin and relocate away from the treated area, preventing toxicity transfer to other colony members (Iqbal and Saeed, 2013). Chlorpyrifos 20 EC is sprayed in various crops like groundnut, maize, sugarcane, and also in soil for effective termite management.
Chemical fumigation is also used to control drywood termite infestations. Methyl bromide phosphine, carbon dioxide and sulfuryl fluoride are the active components in several fumigants. Methyl bromide is a frequently employed fumigant. It damages insects’ nervous system and penetrates swiftly and profoundly at room pressure.
Chemical pesticides were used as a preventative approach to offset the losses brought on by pests and other insects, but their widespread usage had deadly effects all over the world, including diminished soil fertility, low productivity of land air, and water pollution, and threats to human health (Liu et al., 2011). As environmental preservation and safety become more and more important, these chemical termite control products will soon be phased out and replaced by biopesticides.
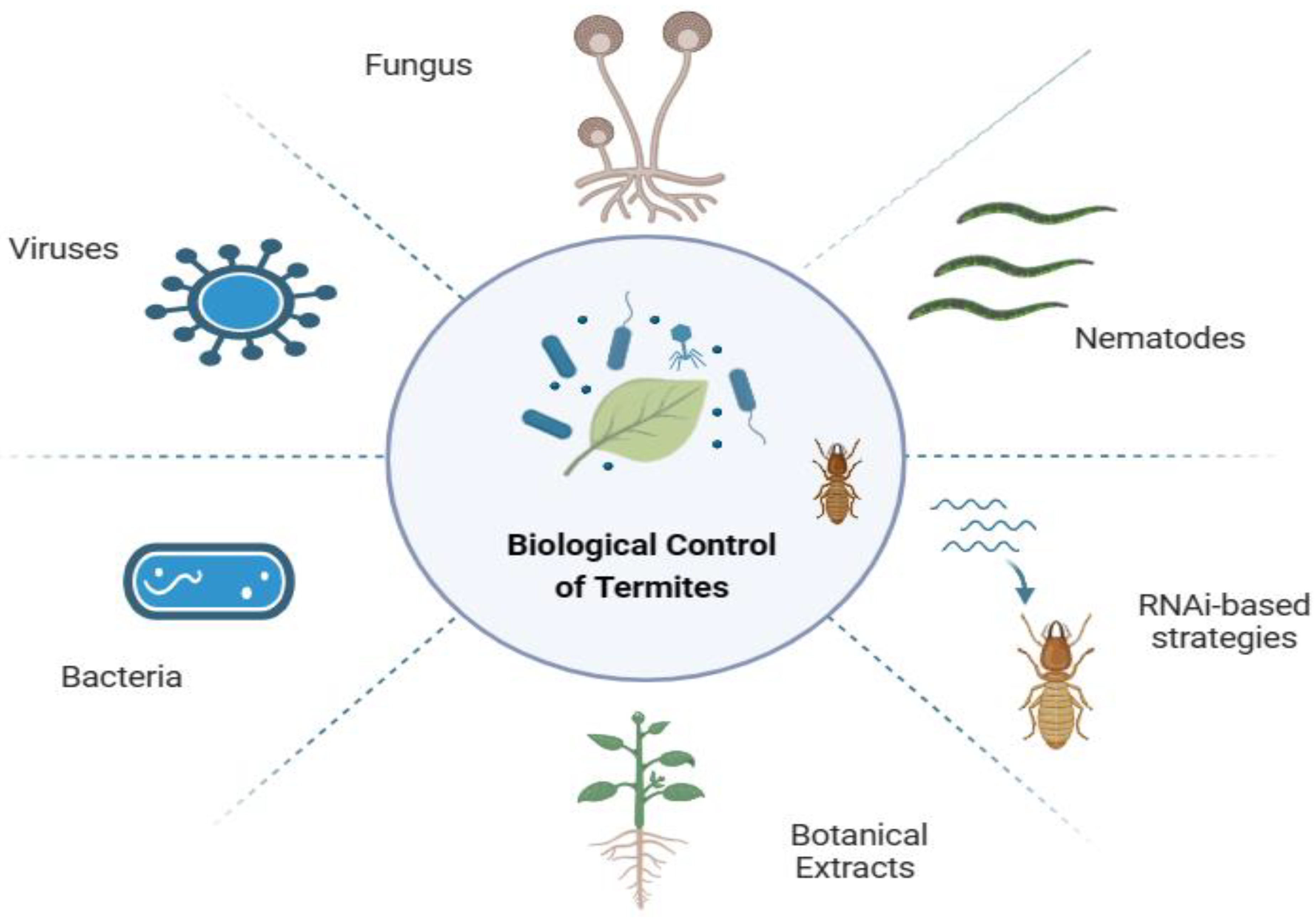
3.3. Biological Warfare Against Termites
Biological termite control uses natural antagonists such as bacteria, viruses, fungi, and nematodes instead of man-made chemical pesticides, which can be harmful to human health and the environment. Many plants produce biologically active compounds, and molecular approaches such as RNA interference (RNAi) can be used to control termites. RNA interference is a biological mechanism that is conserved in eukaryotes, including insects, in which mRNA degradation and protein synthesis disruption turn off gene function (Zhu & Palli, 2020). As a result, biological management has several advantages, as it is a sustainable, environmentally friendly technique, cost-effective, reduces health risks, and improves crop quality.
Figure 5.
Biocontrol of Termites (Source: This study).
3.3.1. Plant Extracts for Termite Management
Botanicals are phyto-based products that are seen to be the most promising substitutes for chemical pesticides. They are generated from plant stems, roots, leaves, fruits, flowers, seeds, and wood. Pesticidal activity is found in around 2000 plant species from 60 families, and many of them are exploited as insect repellents, attractants, growth regulators, behavior modulators, and so on (Verma et al., 2018). Chemicals, such as terpenoids, flavonoids, and saponins, or combinations of chemicals, are naturally found in plants and can repel and kill termites, as well as disrupt their gut flora.
Pre-treating building materials such as wooden frames, pillars, logs, and timbers with anti-termite herbal products like Datura alba, Ricinus communis, Curcuma amada, Cannabis sativus, and Asafoetida spp. in areas afflicted by termites has proven effective in India. However, more research is necessary to determine the effectiveness of these remedies in treating soil and preventing subterranean termite infestations. Plants are safer for the environment, biodegradable, and non-toxic, even though they are less effective than chemicals.
Figure 6.
Termites control via plants secondary metabolites (Source: This study).
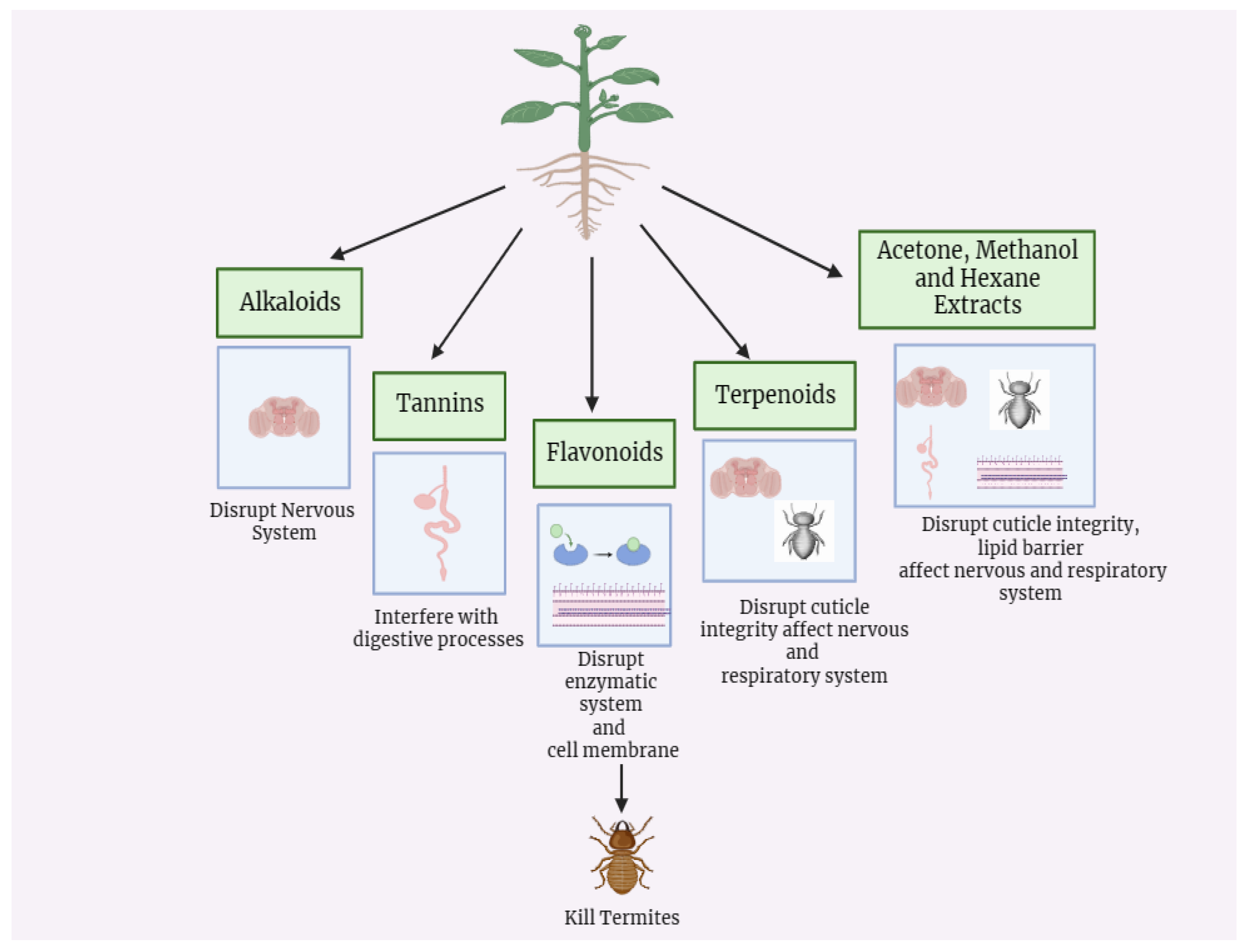

Table 2.
Management of Termites using Plants extracts.
| Sr. No. | Plants Species | Parts | Active compounds | Mechanism | MortalityRate | Country | References |
|---|---|---|---|---|---|---|---|
| 1 | Cymbopogon citratus | Leaves | Alkaloids, Flavonoids, Phenolics, Tannins, Saponins, Glycosides and Citrals | Disrupt cell membrane and cytoskeleton structure Cause blockage of neuromuscular junction and respiratory failure |
75% | Africa | Essien et al., 2023 |
| 2 | Dioscorea bulbifera | Leaves | Methanolic extracts | Inhibit enzymatic activity and affect the metabolism of termites | 70.97% | Indonesia | Oksari et al., 2023 |
| 3 | Rhazya stricta, Lantana camara, Ruta chalepensi and Heliotropium bacciferum | Leaves | n- hexane extracts(Ethyl ester, Octadecadienoic acid) | Disrupt nervous system Interfere with metabolic pathways | 70-76.3% | Saudi Arabia | Alshehry et al., 2014 |
| 4 | `Azadirachta excelsa | Seed Kernel | n-hexane (Hexadecanoic acid, Ethyl ester, Octadecadienoic acid) and methanolic extracts | Disrupt the growth, reproduction and hormonal regulation of termites | 80% | Indonesia | Adfa et al., 2023 |
| 5 | Lavandula latifolia, Origanum vulgare and Syzygium aromaticum | Dry Buds | Lavandula latifolia (Linalool, Lavandulol, β-terpinyl acetate) Origanum vulgare (Thymol, m-cymene, Linalool and Terpinen) Syzygium aromaticum (Eugenol) | Affect nervous system | 75% | Egypt | Salem et al., 2020 |
| 6 | Odontotermes assamensis | Oil | Eugenol, α-pinene, terpinen-4-ol and β-trans-caryophyllene | Affect nervous system and other physiological process like digestion, respiration | 70-90% | India | Pandey et al., 2012 |
| 7 | Syzygium aromaticum | Oil | Flavonoids, Glycosides, Tannins and terpenoids | Affect feeding behaviour of termites by inhibiting enzymes essential for digestion also alter smell and taste of food Blockage of neuromuscular junction | 100% | Africa | Sadiq et al., 2019 |
| 8 | Carica papaya | Leaves | Papain enzyme, Saponins, Flavonoids and Alkaloids Karpain | Proteolytic action of papain disrupt internal proteins affect digestive processes Disrupt cell membrane, leading to cell leakage, also affect nervous system | 40% | Malaysia | Zahtamal et al., 2017 |
| 9 | Eucalyptus globulus | Leaves | Ethanolic Extracts | Disrupt social organization of termite colony Also interfere with termite nervous system, digestive and respiratory processes | 90% | India | Kaundal et al., 2023 |
| 10 | Bintaro | Seeds | Quercetin | Has antioxidant activity may influence cellular processes, interfere with cell cycle | 80% | Japan | Tarmadi et al., 2014 |
| 11 | Azadirachta indica | Oil | Azadirachtin | Disrupt foraging, social behaviour, antifeedant properties and also impact reproductive capability | 74% | Japan | Himmi et al., 2013 |
| 12 | Cissusquadrangularis,Pennisetum purpureum, Vetiveria zizanoides | Leaves | Acetone and Hexane extracts | Disrupt cuticle, neural transmission, has antifeedant properties, interfere with chitin synthesis | 90% | Africa | Kasseney et al., 2016 |
| 13 | L. leucocephala, A. paniculata, A. indica and P. niruri | Leaves | Flavonoids and Methanolic extracts | Antioxidant properties lead to disruption of oxidative balance, disrupt cellular membranes and nervous system | 70-100% | Malaysia | Bakaruddin et al., 2018 |
3.3.2. Management of Termites using Bacteria
Bacterial pathogens were the first microorganisms to be tested as a biological control agent against various rural nuisances. According to Kalha et al., 2014, the majority of bacteria used for biocontrol are found in the families Enterobacteriaceae, Streptococcaceae, Pseudomonadaceae, and Bacillaceae.
Figure 7.
Bacterial compounds as an effective biocontrol for termites (Source: This study).
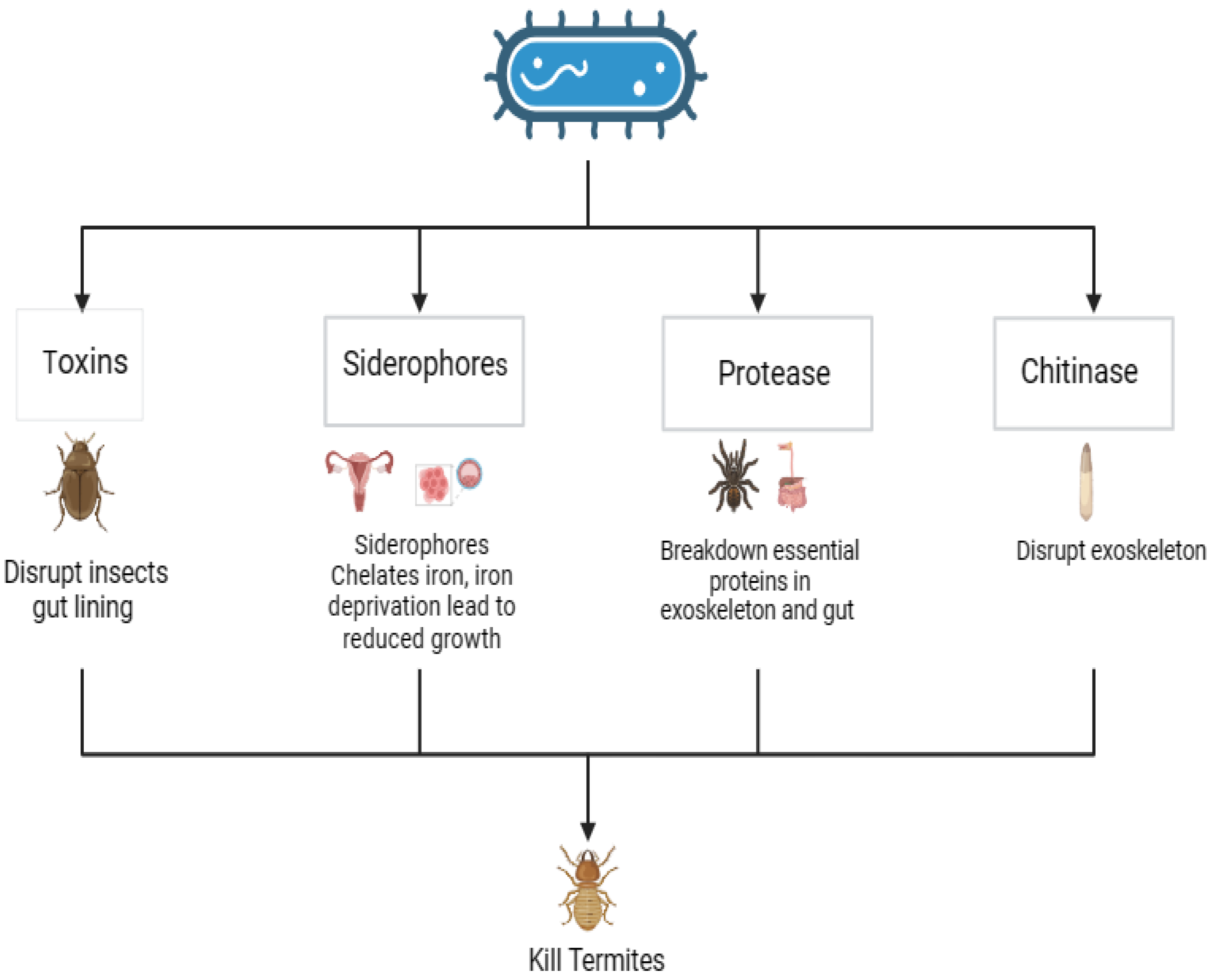
Over 80% of termites (Microtermes obesi and Microcerotermes beesoni) are killed by Bacillus thuringiensis subsp. israelensis, making it a very efficient pathogen against termites. Bacillus thuringiensis produces a crystal-clear protein that is toxic to termites and other insects. When these toxins enter the termite's digestive system, they become active due to the alkaline condition of the insect's gut. These toxins disrupt the termite's gut lining, leading to starvation or infection and eventually, death. Bacillus licheniformis PR2 has been found to produce chitinase and protease activity up to 82.3 units/mL and 35.9 units/mL, respectively. These activities resulted in setae loss, epicuticle disintegration, procuticle rupturing, and socket enlargement in Reticulitermes speratus (Moon et al., 2023).
Some antagonistic bacteria inhibit the growth of termites by producing secondary metabolites like hydrogen cyanide. In addition to producing HCN, P. fluorescens also inhibits the termite respiratory chain's Cytochrome C Oxidase enzyme, killing Odontotermes obesus (Devi et al., 2009).
Table 3.
Entomopathogenic bacteria as a biocontrol for termites.
| Bacteria | Termites | Enzymeformulations | Mode of Action | Country | Mortality Rate | References |
|---|---|---|---|---|---|---|
| Stentrophomons maltophilia | Coptotermes heimi and Heterotermes indicola | Chitinase | Disruption of cuticle | Pakistan | 53 % | Jabeen et al., 2018 |
| Methicillin Resistant Staphylococcus aureus | Reticulitermes flavipes | Protease | Alter haemolymph protein that affect cellular immune response, Disrupt essential processes like digestion, respiration | Korea | 70% | Zeng et al., 2016 |
| Serratia marcescens (SM1) | Odontotermes formosanus (Shiraki) | Chitinase enzymes | Hydrolyzes chitin in locust midgut and perforates intestinal membranes, making insecticidal proteins more likely to penetrate the insect body and destroy the digestive tract | China | NA | Fu et al., 2020 |
| Rhizobium radiobacter, Alcaligenes latus, Aeromonas caviae | Odontotermes obesus | HCN production | Disrupt electron transport chain and preventing cells from utilizing oxygen | India | 70% mortality followed by 1 hr incubation | Devi et al., 2007 |
| Serratia marcescens | Odontotermes formosans | Protease | Cuticle damage, disrupt microbial balance in termite gut | China | 80 % mortality after 72 hours | Fu et al., 2020 |
| Entrobacter cloacae | Odontotermes obesus | Insecticidal Proteins | Affects the cellular immune response | China | NA | Zhang et al., 2010 |
| Citrobacter freundii | Coptotermes spp. | CellulaseEnzyme | Breaks cellulose present in cell wall so disrupt cuticle | Africa | NA | Omoya and Kelley, 2014 |
| Bacillus licheniformis PR2 | Reticulitermes speratus kyushuensis Morimoto | Chitinase | Cuticle degradation | Korea | 88.9% after 3 hours | Moon et al., 2023 |
| B. licheniformis USMW10IK | G. sulphureus | Chitinase | Cuticle degradation | Malaysia | 22.8% after 48 h | Hussin and Mazid, 2020 |
| Pseudomonas fluorescens CHA0 | O. obesus | Cytochrome C Oxidase enzyme | Inhibit respiratory chain | India | NA | Devi et al., 2009 |
| B. thuringiensis subsp. israelensis |
Microtermes obesi and Microcerotermes Beesoni |
Chitinase | Cuticle degradation | India | 80% mortality | Singha et al., 2010; Wang & Henderson, 2013 |
| Bacillus thuringiensis KJ3P1 | Macrotermes gilvus | Chitinase | Cuticle degradation | Indonesia | 50% after treatment | Pujiastuti et al., 2021 |
3.3.3. Fungal Species as a Biocontrol Agent for Termites
Entomopathogenic fungi plays a vital role in biological pest control and can cause mortality at all stages of development. The pathogenesis of termite fungal diseases appears to involve the following phases. Conidium first attaches and penetrates the insect cuticle, causing the germination of fungal hyphae in the haemocoel and production of toxins, which ultimately leads to termite death.
Several EPF strains, including Beauveria spp., Metarhizium spp., and Lecanicillium spp., were evaluated against several insect pests and shown to be efficient termite biocontrol agents (Pandey et al., 2013). The Metarhizium and Beauveria genera have parasitic interactions with insects like termites.
Yii et al., 2015 used fipronil termiticide in combination with M. anisopliae spores to control Coptotermes curvignathus. They concluded that combining conidia with sublethal fipronil kills C. curvignathus in more than 99% of cases. In Iraq, a commercial formulation of the fungus Metarhizium anisopliae was used successfully to protect olive trees from termite species such as M. diversus (Hussain et al. 2011, Zaidawi et al., 2020). Table 4 shows a list of common fungal species that are harmful to termites.
Numerous studies have demonstrated the effectiveness of entomopathogenic fungi as biological control microorganisms for agricultural termite management, although fewer have demonstrated their ineffectiveness. Liu et al., 2019 observed that termites have highly sophisticated multidefence tactics against fungi. Their defence mechanisms include avoiding fungal-contaminated areas, secreting antifungal substances and using symbiotic bacteria as the foundation of their nests to prevent infections from entering their colonies. The dispersal, transmission, germination, penetration into the host, and environmental factors all contribute to the efficacy of fungal biocontrol (Yii et al., 2015). Fungus-based termite management can be a successful IPM program tactic as a result of these parameters being optimized.
Table 4.
Fungal species pathogenic to termites.
| Fungal Species | Termites | Mortality rate | LD50 | Country | References |
|---|---|---|---|---|---|
| Aspergillus spp. |
Microcerotermes beesoni Snyder |
55 % | 3.69 × 107 conidia/ml | India | Pandey et al., 2013 |
| Beauveria bassiana | Microtermes obesi Holmgreen | 50% | 2 ×105 conidia /ml | India | Singha and Dutta,2011 |
| Isaria fumosorose | Coptotermes formosanus | 100% | Not Known | Malaysia | Jessica et al., 2019 |
| Metarhizium anisopliae | Coptotermes formosanus | 62.8% | 2.5 ×104 conidia /ml | USA | Wright et al., 2008 |
| Metarhizium anisoploae | Odontotermes formasanus | 100% | 3×108 conidia/ml | USA | Dong et al., 2007 |
| Metarhizium brunneum Cb15-III | Odontotermes formasanus | 100% | 1×108 conidia /ml | Kenya | Ambele et al., 2020 |
| Aspergillus auricomus | Coptotermes curvignathus Holmgren | NA | 1.49 × 105 conidia/ml | India | Kamarudin and Lau, 2022 |
| Metarhizium anisoploae | Coptotermes curvignathus | 95% | 1×107 conidia/ml | Malaysia | Samsuddin et al., 2016 |
| M. anisopliae, B. bassiana and A. niger | Psammotermeshypostoma | 61.52, 35.54 and 20.06% | 4×106 spores/ml | Egypt | Somalian et al., 2019 |
| Metarhizium spp. B2.2 | Coptotermes spp. | 100% | 8.3 x 109 conidia/ml | Indonesia | Zulfiana et al., 2020 |
| Metarhizium anisopliae | Microtermes obesi (Holmgren) | 90% | 2 × 107 conidia/ml | India | Deka et al., 2021 |
| Isaria farinosa | Nasutitermes cornige | 85% | 6.6 × 104 conidia/ml | Brazil | Lopes et al., 2017 |
| Paecilomyces fumosoroseus | Coptotermes sp. | NA | NA | USA | Dunlap et al., 2007 |
| Beauveria bassiana | Microtermes obesi | 25-40% | 4 ×106 conidia /ml | India | Padmaji and Kaur 2001 |
| Metarhizium anisopliae TK29 | Coptotermes formosanus | 75% | 1 × 108 conidia/ml | China | Keppanan et al., 2018 |
3.3.4. Nematodes Species as Biological Control for Termites
A significant amount of termite’s population is naturally killed by roundworms belonging to the phylum Nematoda. Four nematode families have the potential to control termites Heterorhabditidae, Allantonematidae, Steinernematidae, and Mermithidae. The Steinernematidae and Heterorhabditidae families have been used to manage insect pests primarily (Poinar et al., 1990). By penetrating the target through natural openings like the mouth, anus, and cuticle, nematodes can infect and destroy insect pests. Table 5 below provides a list of frequent nematodal species that are harmful to termites.
Despite several advantages, nematodes as biocontrol offers limited effectiveness, specificity, and slow action. The physical and chemical characteristics of soil (such as moisture, temperature, oxygen, pore size, carbon dioxide levels, pH, salinity, and the presence of artificial chemicals) as well as biotic factors (such as competitive interactions with other soil species, restricted motility, and termite behaviors) all affect how effective nematodes are in biocontrol programs. Additional investigation into nematode ecology and biology, genetic manipulation, and combinations with other control agents is needed to fully realize their insecticidal potential
Table 5.
Nematodes species parasitic to termites.
| Nematodes | Termites | Mortality | LD50 | Country | References |
|---|---|---|---|---|---|
| H. bacteriophora | Microcerotermes diversus | 43.6% after 48 hours | 57.9 IJ/ml | Iraq | Zaidawi et al., 2020 |
| Steinernema pakistanense,S. Bifurcatum | Microtermes obesi | 100% after 48 hours | 350 -650 IJ/ml). | Pakistan | Javed et al., 2021 |
| Steinernema carpocapsae, Heterorhabditis bacteriophora and Heterorhabditis indica | Odontotermes obesus | S. carpocapsae (58.46%), followed by H. bacteriophora (45.45%) and H. indica (32.39%) after 48 hours | 1000 IJ/ml | Pakistan | Aslam et al., 2023 |
| Heterorhabditis indica | Odontotermes obesus | 87.98 % after 48 hours | 600 IJ/ml | India | Afroz et al., 2023 |
| Steinernema siamkayai, S. pakistanense and Heterorhabditis indica | Reticulitermes flavipes and Odontotermis hornei | 80% mortality S. pakistanense in 15.5 hrs, S. siamkayai in 16.3 hrs H. indica in 19.8 hrs | 5.84 IJ/ml, 5.68 IJ/ml, 5.00 IJ/ml | Pakistan | Razia and Sivaramakrishnan, 2016 |
| Steinernema karii | Coptotermes formosanus | 100% mortality after 96 hours | Not Known | Kenya | Wagutu et al., 2017 |
| Steinernema feltiae | Reticulitermes tibialis | Not Known | Not Known | USA | Epsky and Capinera,1988 |
| Steinernema thermophilum | Odontotermes obesus | 42 - 48 % mortality after 72 hours | Not Known | India | Rathour et al., 2014 |
| Heterorhabditis sonorensis Azohoue2 | G. mellonella | 63.2% mortality after 48 hours | Not Known | India | Rahman et al., 2011 |
| Steinernema riobrave Cabanillas | H. aureus | 80% mortality after 3 days | Not known | USA | Yu et al., 2006 |
3.3.5. Potential of Viruses as Biocontrol Agent for Termites
Some viruses act as biocontrol agents for termites by infecting and killing them. Until now, very little research has been conducted on virus activity against termites. Among viruses, the Baculoviridae family, which includes granuloviruses and nuclear polyhedrosis viruses, has the highest potential for biocontrol. Baculoviruses have been shown to reside in more than 400 insect species, the majority of which are found in the Lepidoptera and Hymenoptera orders. Gibbs et al., 1970 found a virus that attacked Coptotermes lacteus and was related to the virus that causes acute paralysis in honey bees, Apis mellifera.
Al Fazairy et al.,1988 discovered that a nuclear polyhedrosis virus isolated from Spodoptera littoralis Boisduval was infectious against the K. flavicollis (Kalotermitidae). Termites died 2–10 days after infection in the lab, leading scientists to believe that using NPV (Nuclear Polyhedrosis Virus) to control K. flavicollis would be possible.
Baculovirus is a useful supplement to broad-spectrum insecticides due to its effectiveness, specificity, and capacity to generate secondary inoculum. They are a crucial component of integrated pest management (IPM) because, unlike other biological control agents, they do not negatively impact beneficial insects. Nevertheless, employing the virus to manage pest populations has many drawbacks. For instance, large production of the virus is challenging due to its demand for a living host or tissue culture. Additionally, they eventually eradicate their hosts.
3.3.6. RNAi for Termite Management
In an effort to shield plants against pest infestations, scientists have recently focused on RNA interference (RNAi) technology. RNAi is a natural biological process that regulates gene expression by inhibiting the translation of particular mRNA molecules. In the context of termite management, researchers have investigated the use of RNAi to target critical genes in termites, potentially affecting their biological processes and eventually controlling their populations (Zhu et al., 2020).
Figure 8.
RNA interference against Termites (Source: This study).
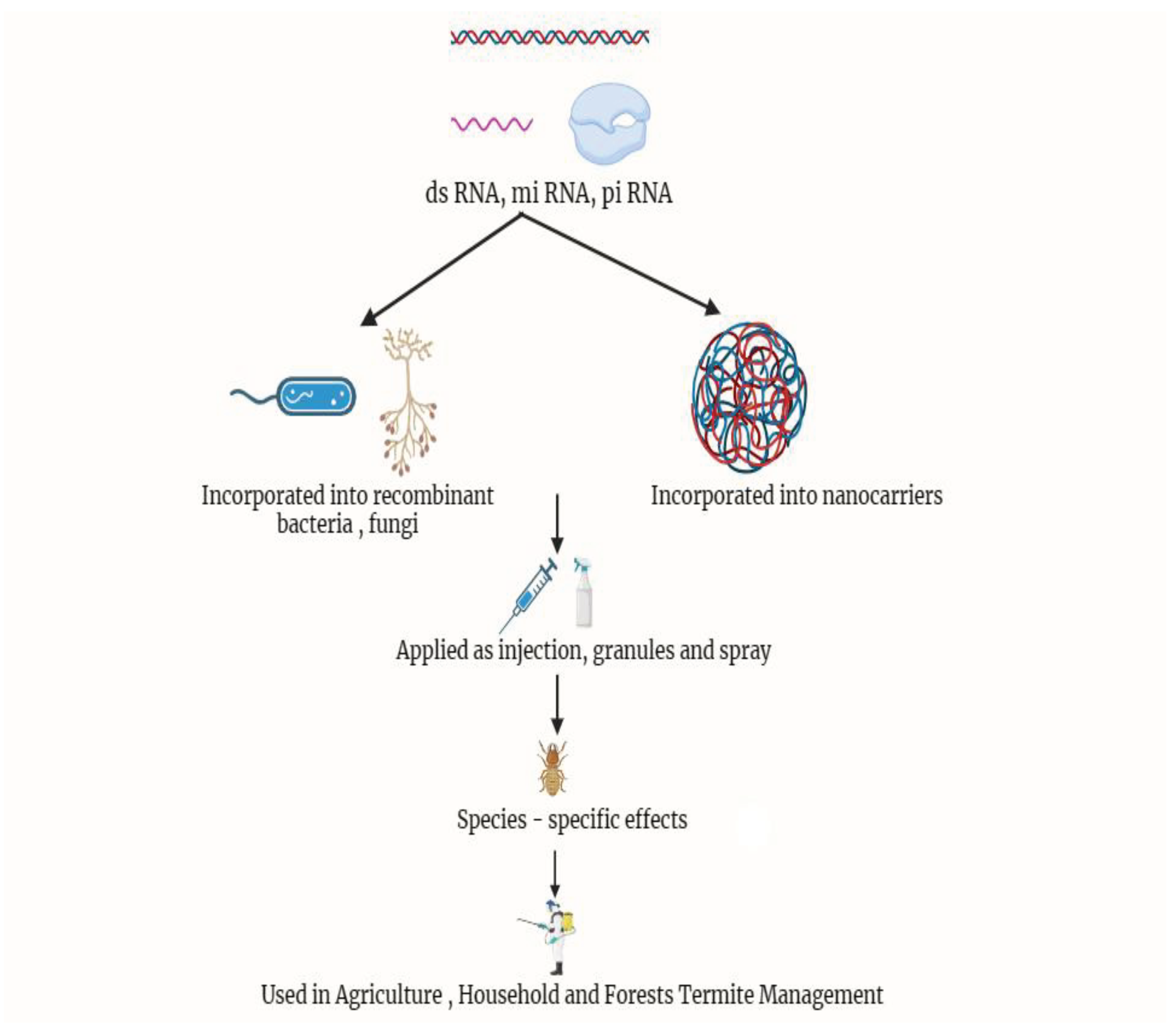
The initial study on RNA interference (RNAi) was carried out on R. flavipes through the injection of short interfering RNA (siRNA), which produced siRNA against two hexamerin genes (Hex-1 and Hex-2) linked to the regulation of caste polymorphism. Later, two termite genes in R. flavipes were targeted by Zhou et al., 2008 using the dsRNA feeding approach. These genes were for the cellulase-digesting enzyme endoglucanase and the caste-regulating hexamerin storage protein hexamerin. The selenoprotein T (SELT) gene's capacity for active immunization against R. chinensis termites was investigated by Zhao et al. in 2020. After SELT and TG were knocked down, Zhao et al., 2020 found that both proteins are crucial for producing active immunity against the entomopathogenic fungus M. anisopliae in R. chinensis.
Termite metabolism may be affected by isotrate dehydrogenase (IDH), and dysregulation of IDH leads to an increase in apoptotic lesions and high infection and mortality rates (Liu et al., 2020). According to Zhao et al., 2020 and Esparza-Mora et al., 2020, GNBP2 and TG also play a role in termite behavioural defence by controlling cannibalistic and grooming behaviours respectively. According to Liu et al., 2022, termites with Dicer-1-mediated miRNA dysregulation expressed fewer genes related to antioxidant defence and glucose metabolism, which significantly reduced the termite's ability to combat fungal growth. Additionally, termites treated with miR-71-5 stimulant showed a reduction in overall antifungal activity, suggesting a detrimental effect of miRNA dysregulation on termite physiological defences.
The main barriers to the broad use of RNAi for termite pest management are its effectiveness, delivery challenges, off-target and non-target effects, immune responses and potential resistance development.
Table 6.
RNAi for controlling Termites.
| Species | Target gene | Target Gene Function | Type of RNA | Delivery Method | Country | References |
|---|---|---|---|---|---|---|
| Reticulitermes chinensis and Odontotermes formosanus | Olfactory coreceptor | Necessary for odorant detection | dsRNA | Injection | China | Gao et al., 2020 |
| Odontotermes formosanus | Termicin | Antifungal activity | dsRNA | Feeding | China | Feng et.al, 2022 |
| C. formosanus | Protistan gene | Importantfor protists lignocellulosic process | siRNA | Feeding | China | Liu et al., 2017 |
| Zootermopsis nevadensis | Met, 20E signaling and nuclear receptor, Hormone receptor 39 | Methoprene Tolerant maintains the action of juvenile hormone 20 E signaling and nuclear receptor regulates egg development Hormone Receptor 39 helps in reproductive gland development | dsRNA | Injection | Japan | Masuoka et al., 2018 |
| Hodotermopsis holmgreen | Dac and Distal less | Dac helps in egg and legs development Distal less involved in egg development | ds RNA | Injection | Japan | Sugime et al., 2019 |
| Nasutitermes takasagoensis | Deformed gene | Involved in determining mandibular positional information during pre-soldier differentiation | siRNA | Injection | Japan | Toga et al., 2013 |
4. Conclusion
Termites are becoming a more serious menace to the world economy. Many Integrated Termites Management (ITM) components have received a lot of attention, but more focused efforts are still needed. Furthermore, several physical and chemical termite treatment options are available. The widespread use of chemical-based pesticides harm crops and increase the risk of secondary pollution.
Biopesticides have been demonstrated to be the most effective and necessary method of treating termites. Many plant extracts were discovered to be useful against termites, while some botanicals were not as powerful as chemicals. Microorganisms such as fungus, nematodes, bacteria, and viruses are effective termiticides. They kill termites effectively by creating siderophore, poisons, lytic enzymes such as protease, chitinase, and secondary metabolites like HCN. Nowadays, indoor termiticides based on RNAi are probably used to protect wooden structures. In conclusion safer, eco- friendly and cost- effective mode of strategy to reduce the termites can be practised using microorganism, plants extracts and RNA interference for restoring the biological and physiochemical composition of our ecosystems.
However, there are some issues with the use of microorganisms, such as the fact that they become inactive under certain extreme environments. Furthermore, these microbial entities have some leaching issues, which have become a source of contamination of ground water and have caused various water-borne diseases. Long-term pest management control cannot usually be achieved by a single microbial control agent. As part of a comprehensive plan in all farming methods, entomopathogens and their enzymes could offer cost effective, safer, substantial and targeted pest control without compromising the efficacy of other methods.
5. Future Prospectives
The efficacy and sustainability of integrated control strategies should be enhanced in the near future by the synergistic pairing of microbial control agents with other technologies. Technologies including improved biological control agents, precise targeting, and genetically modified organisms (GMOs) may provide environmentally acceptable and long-lasting pest management solutions. The majority of investigations on checking biocontrol efficacy are conducted at the research level or on a lab scale; further studies are required to determine the effectiveness of these methods in real-world settings. More research is also needed to assure the long-term release and efficacy of microbial application methods used as biocontrol. Many of the difficulties that microbial control is currently facing should be resolved by advances in microbial products and increased public understanding regarding the advantages of biological control through government support, media outreach, and educational programs.
Abbreviations
BCA, Biological Control Agents; USD, United States Dollar; RNAi, RNA Interference; CCO, Cytochrome C Oxidase Enzyme; HCN, Hydrogen Cyanide; IJ, Intra jugular; siRNA, Short Interfering RNA; dsRNA Double stranded RNA; Hex, Hexamerin; IDH, Isotrate Dehydrogenase; SELT, Selenoprotein T; NPV, Nuclear Polyhedrovirus; IPM, Integrated Pest Management; GMOs, Genetically Modified Organisms; ITM, Integrated Termites Management
References
- de Figueirêdo, R.E.C.R.; Vasconcellos, A.; Policarpo, I.S.; Alves, R.R.N. Edible and medicinal termites: a global overview. J. Ethnobiol. Ethnomedicine 2015, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.; Aidoo, O.; Ghisletta, M.; Osei-Owusu, J.; Saraswati, Y.; Bhardwaj, K.; Khalid, W.; Fernando, I.; Golik, A.; Nagdalian, A.; et al. African edible insects as human food – a comprehensive review. J. Insects Food Feed. 2023, 10, 51–78. [Google Scholar] [CrossRef]
- Aidoo, O.F.; Osei-Owusu, J.; Asante, K.; Dofuor, A.K.; Boateng, B.O.; Debrah, S.K.; Ninsin, K.D.; Siddiqui, S.A.; Chia, S.Y. Insects as food and medicine: a sustainable solution for global health and environmental challenges. Front. Nutr. 2023, 10, 1113219. [Google Scholar] [CrossRef] [PubMed]
- Muon, R.; Ket, P.; Sebag, D.; Boukbida, H.A.; Podwojewski, P.; Hervé, V.; Ann, V.; Jouquet, P. Termite constructions as patches of soil fertility in Cambodian paddy fields. Geoderma Reg. 2023, 33. [Google Scholar] [CrossRef]
- Subekti, N., Yoshimura, T., Rokhman, F., & Mastur, Z. (2015). Potential for subterranean termite attack against five bamboo species in correlation with chemical components. Procedia Environmental Sciences, 28, 783-788.
- Murthy, K.S.; Resources, P.B.N. 2.N.B.O.A.I. Diversity and abundance of Subterranean Termites in South India. Indian J. Pure Appl. Biosci. 2020, 8, 141–149. [Google Scholar] [CrossRef]
- Rust, M.K.; Su, N.-Y. Managing Social Insects of Urban Importance. Annu. Rev. Èntomol. 2012, 57, 355–375. [Google Scholar] [CrossRef]
- Liu Ning, L. N. , Yan Xing, Y. X., Zhang MeiLing, Z. M., Xie Lei, X. L., Wang Qian, W. Q., Huang YongPing, H. Y., & Zhou ZhiHua, Z. Z. (2011). a: Microbiome of fungus-growing termites.
- Scharf, M.E. Omic research in termites: an overview and a roadmap. Front. Genet. 2015, 6, 76–76. [Google Scholar] [CrossRef] [PubMed]
- Wekhe, C.; Ugbomeh, A.P.; Ebere, N.; Bawo, D.D.S. Subterranean Termites of a University Environment in Port Harcourt, Nigeria. Asian J. Biol. 2019, 1–10. [Google Scholar] [CrossRef]
- Subekti, N., Yoshimura, T., Rokhman, F., & Mastur, Z. (2015). Potential for subterranean termite attack against five bamboo species in correlation with chemical components. Procedia Environmental Sciences, 28, 783-788.
- Smagghe, F.; Spooner-Hart, R.; Chen, Z.-H.; Donovan-Mak, M. Biological control of arthropod pests in protected cropping by employing entomopathogens: Efficiency, production and safety. Biol. Control. 2023, 186. [Google Scholar] [CrossRef]
- Farook, U. B., Khan, Z. H., Ahad, I., Maqbool, S., Yaqoob, M., Rafieq, I., & Sultan, N. (2019). A review on insect pest complex of wheat (Triticum aestivum ). Journal of Entomology and Zoology Studies, 7(1), 1292-1298.
- Sharma, A. K., Babu, K. S., Nagarajan, S., Singh, S. P., & Manoj, K. (2004). Distribution and status of termite damage to wheat crop in India. Indian Journal of Entomology, 66(3), 235-237.
- Parween, T.; Jan, S.; Mahmooduzzafar, S.; Fatma, T.; Siddiqui, Z.H. Selective Effect of Pesticides on Plant—A Review. Crit. Rev. Food Sci. Nutr. 2014, 56, 160–179. [Google Scholar] [CrossRef]
- Rana, A.; Chandel, R.S.; Verma, K.S.; Joshi, M.J. Termites in Important Crops and Their Management. Indian J. Èntomol. 2021, 83, 486–504. [Google Scholar] [CrossRef]
- Paul, B. , Khan, M. A., Paul, S., Shankarganesh, K., & Chakravorty, S. (2018). Termites and Indian agriculture. Termites and Sustainable Management: Volume 2-Economic Losses and Management, 51-96.
- Parween, T., Bhandari, P., & Raza, S. K. (2016). Survey and identification of termite in some selected parts of India. Res J Life Sci Bioinform Pharm Chem Sci, 2, 122-135.
- Roy, S.; Prasad, A.K.; Neave, S.; Bhattacharyya, P.N.; Nagpal, A.; Borah, K.; Rahman, A.; Sarmah, M.; Sarmah, S.R.; Pandit, V. Nonchemical based integrated management package for live-wood eating termites in tea plantations of north-east India. Int. J. Trop. Insect Sci. 2020, 40, 435–440. [Google Scholar] [CrossRef]
- Mahapatro, G.K.; Chatterjee, D. Termites as Structural Pest: Status in Indian Scenario. Proc. Natl. Acad. Sci. India Sect. B: Biol. Sci. 2017, 88, 977–994. [Google Scholar] [CrossRef]
- Nagaraju, D.K.; Kalleshwaraswamy, C.M.; Iyyanar, D.; Singh, M.; Jain, R.K.; Kasturi, N.; Ranjith, M.; Mahadevaswamy, H.M.; Asokan, R. First interception of two wood feeding potential invasive Coptotermes termite species in India. Int. J. Trop. Insect Sci. 2020, 41, 1043–1052. [Google Scholar] [CrossRef]
- Kalleshwaraswamy, C.M. Potential Invasive Termites in India and Importance of Integrative Taxonomy. Indian J. Èntomol. 17. [CrossRef]
- de Mello, A.P.; da Costa, B.G.; Da Silva, A.C.; Silva, A.M.B.; Bezerra-Gusmão, M.A. Termites in historical buildings and residences in the semiarid region of Brazil. Sociobiology 2014, 61, 318–323. [Google Scholar] [CrossRef]
- Gusmão, M.A.B.; Barbosa, A.M.S.; Mello, A.P.; Oliveira, J.V.; Barros, A.T. Perceptions of termites in urban areas of semiarid Brazil. Biotemas 2014, 27, 117. [Google Scholar] [CrossRef]
- Atsbha Gebreslasie, A. G., & Hintsa Meressa, H.M. (2018). Evaluation of chemical, botanical and cultural management options of termite in Tanqua Abergelle district, Ethiopia.
- Cao, R.; Su, N.-Y. Temperature Preferences of Four Subterranean Termite Species (Isoptera: Rhinotermitidae) and Temperature-Dependent Survivorship and Wood-Consumption Rate. Ann. Èntomol. Soc. Am. 2015, 109, 64–71. [Google Scholar] [CrossRef]
- Li, Y.; Dong, Z.-Y.; Pan, D.-Z.; Pan, C.-H.; Chen, L.-H. Effects of Termites on Soil pH and Its Application for Termite Control in Zhejiang Province, China. Sociobiology 2017, 64, 317–326. [Google Scholar] [CrossRef]
- Ahmed, Sohail, Mustafa, Tariq, Riaz, M. A., & Hussain, A. B. I. D. (2006). Efficacy of insecticides against subterranean termites in sugarcane. Int. J. Agric. Biol, 8(4), 508-510.
- Iqbal, N. , & Saeed, S. (2013). Toxicity of six new chemical insecticides against the termite, Microtermes mycophagus D.(Isoptera: Termitidae: Macrotermitinae). Pakistan Journal of Zoology, 45(3).
- Liu, N.; Yan, X.; Zhang, M.; Xie, L.; Wang, Q.; Huang, Y.; Zhou, X.; Wang, S.; Zhou, Z. Microbiome of Fungus-Growing Termites: a New Reservoir for Lignocellulase Genes. Appl. Environ. Microbiol. 2011, 77, 48–56. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Palli, S.R. Mechanisms, Applications, and Challenges of Insect RNA Interference. Annu. Rev. Èntomol. 2020, 65, 293–311. [Google Scholar] [CrossRef]
- Verma, M. , Verma, S., & Sharma, S. (2018). Eco-friendly termite management in tropical conditions. Termites and Sustainable Management: Volume 2-Economic Losses and Management, 137-164.
- Essien, R. A. , Oboho, D. E., Imakwu, C. A., Atteh, I., & Okwor, J. (2023). Laboratory Evaluation of Termiticidal Efficacy of Cymbopogon citratus Stapf leaf Extract against Termites, Macrotermes bellicosus (Blattodea: Termitidae) in Obio Akpa Community, Oruk Anam Akwa Ibom State-Nigeria. Journal of Experimental Research, 11(2).
- A Oksari, A.; Susanty, D.; Rizki, F.H.; Wanda, I.F.; Arinana; Dadang Potential of Dioscorea bulbifera L. as a bio-insecticide in controlling dry wood termites (Cryptotermes cynocephalus Ligh.). IOP Conf. Series: Earth Environ. Sci. 2023, 1133. [Google Scholar] [CrossRef]
- Alshehry, A. Z. , Zaitoun, A. A., & Abo-Hassan, R. A. (2014). Insecticidal activities of some plant extracts against subterranean termites, Psammotermes hybostoma (Desneux)(Isoptera: Rhinotermitidae). International Journal of Agricultural Sciences, 4(9), 257-260.
- Adfa, M.; Wiradimafan, K.; Pratama, R.F.; Sanjaya, A.; Triawan, D.A.; S. , S.Y.; Ninomiya, M.; Rafi, M.; Koketsu, M. Anti-Termite Activity of Azadirachta excelsa Seed Kernel and Its Isolated Compound against Coptotermes curvignathus. J. Korean Wood Sci. Technol. 2023, 51, 157–172. [Google Scholar] [CrossRef]
- Salem, M. Z. , Ali, M. F., Mansour, M. M., Ali, H. M., Abdel Moneim, E. M., & Abdel-Megeed, A. (2020). Anti-termitic activity of three plant extracts, chlorpyrifos, and a bioagent compound (protecto) against termite Microcerotermes eugnathus silvestri (Blattodea: Termitidae) in Egypt. Insects, 11(11), 756.
- Pandey, A.; Chattopadhyay, P.; Banerjee, S.; Pakshirajan, K.; Singh, L. Antitermitic activity of plant essential oils and their major constituents against termite Odontotermes assamensis Holmgren (Isoptera: Termitidae) of North East India. Int. Biodeterior. Biodegradation 2012, 75, 63–67. [Google Scholar] [CrossRef]
- Sadiq, S. I., Adeboye, A. G., Yalli, F. A., & Dikwa, M. A. (2019). Phytochemical constituents and termicidal activity of essential oils from Syzygium Aromaticum (Clove bud. Fudma Journal of sciences, 3(2), 220-225.
- Zahtamal, S. A. , Marsifah, T., Soesilohadi, R. H., Handayani, S., & Handayani, S. M. Toxic Potential of Papaya (Carica papaya) Leaves as Termicidal Against Coptotermes curvignathus Holmgren.
- Umang, U.; Kaundal, A.K.; Kumari, P.; Kaur, S. Investigations On Anti- Termite Activity Of Eucalyptus Globulus Leaf Extract. 2023, 3397–3405. [CrossRef]
- Tarmadi, D.; Himmi, S.K.; Yusuf, S. The Efficacy of the Oleic Acid Isolated from Cerbera Manghas L. Seed Against a Subterranean Termite, Coptotermes Gestroi Wasmann and a Drywood Termite, Cryptotermes Cynocephalus Light. Procedia Environ. Sci. 2014, 20, 772–777. [Google Scholar] [CrossRef]
- Himmi, S.K.; Tarmadi, D.; Ismayati, M.; Yusuf, S. Bioefficacy Performance of Neem-based Formulation on Wood Protection and Soil Barrier against Subterranean Termite, Coptotermes Gestroi Wasmann (Isoptera: Rhinotermitidae). Procedia Environ. Sci. 2013, 17, 135–141. [Google Scholar] [CrossRef]
- Kasseney, B. D., Nyamador, W. S., Abla, D. Ã., Ketoh, G. K., & AdolÃ, I. (2016). Termiticidal activities of few plant extracts against Macrotermes subhyalinus smeathman and Trinervitermes geminatus wasmann (Isoptera: Termitidae) survival. African Journal of Agricultural Research, 11(28), 2475-2480.
- Bakaruddin, N.H.; Dieng, H.; Sulaiman, S.F.; Ab Majid, A.H. Evaluation of the toxicity and repellency of tropical plant extract against subterranean termites, Globitermes sulphureus and Coptotermes gestroi. Inf. Process. Agric. 2018, 5, 298–307. [Google Scholar] [CrossRef]
- Kalha, C.S.; Singh, P.P.; Kang, S.S.; Hunjan, M.S.; Gupta, V.; Sharma, R. Entomopathogenic Viruses and Bacteria for Insect-Pest Control. In Integrated Pest Management; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 978-0-12-398529-3. [Google Scholar]
- Moon, J. H. , Ajuna, H. B., Won, S. J., Choub, V., Choi, S. I., Yun, J. Y.,... & Ahn, Y. S. (2023). The Anti-Termite Activity of Bacillus licheniformis PR2 against the Subterranean Termite, Reticulitermes speratus kyushuensis Morimoto (Isoptera: Rhinotermitidae). Forests, 14(5), 1000.
- Devi, K.K.; Kothamasi, D. Pseudomonas fluorescens CHA0 can kill subterranean termite Odontotermes obesus by inhibiting cytochrome c oxidase of the termite respiratory chain. FEMS Microbiol. Lett. 2009, 301, 147–147. [Google Scholar] [CrossRef]
- Jabeen, F., Hussain, A., Manzoor, M., Younis, T., Rasul, A., & Qazi, J. I. (2018). Potential of bacterial chitinolytic, Stenotrophomonas maltophilia, in biological control of termites. Egyptian Journal of Biological Pest Control, 28, 1-10.
- Zeng, Y.; Hu, X.P.; Suh, S.-J. Characterization of Antibacterial Activities of Eastern Subterranean Termite, Reticulitermes flavipes, against Human Pathogens. PLOS ONE 2016, 11, e0162249–e0162249. [Google Scholar] [CrossRef]
- Fu, R., Qi, X., Feng, K., Xia, X., & Tang, F. (2019). Identification and characteristics of a strain of Serratia marcescens isolated from the termites, Odontotermes formosanus. Journal of Nanjing Forestry University, 62(01), 76.
- Devi, K.K.; Seth, N.; Kothamasi, S.; Kothamasi, D. Hydrogen Cyanide-Producing Rhizobacteria Kill Subterranean Termite Odontotermes obesus (Rambur) by Cyanide Poisoning Under In Vitro Conditions. Curr. Microbiol. 2007, 54, 74–78. [Google Scholar] [CrossRef]
- Zhang PengBo, Z. P. , Yan Xun, Y. X., Qiu XueHong, Q. X., & Han RiChou, H. R. (2010). Application of transgenic Enterobacter cloacae with the insecticidal tcdA1B1 genes for control of Coptotermes formosanus (Isoptera: Rhinotermitidae) in the field.
- Omoya, F.; Kelly, B. Variability of the potency of some selected entomopathogenic bacteria (Bacillus spp. and Serratia spp.) on termites, Macrotermes bellicosus (Isoptera: Termitidae) after exposure to magnetic fields. Int. J. Trop. Insect Sci. 2014, 34, 98–105. [Google Scholar] [CrossRef]
- Hussin, N.A.; Ab Majid, A.H. Termiticidal activity of chitinase enzyme of Bacillus licheniformis, a symbiont isolated from the gut of Globitermes sulphureus worker. Biocatal. Agric. Biotechnol. 2020, 24. [Google Scholar] [CrossRef]
- Singha, D. I. P. E. N. D. R. A. , Singha, B. K. ( 2010). In vitro pathogenicity of Bacillus thuringiensis against tea termites. Journal of Biological Control, 279–281.
- Wang, C.; Henderson, G. Evidence of Formosan Subterranean Termite Group Size and Associated Bacteria in the Suppression of Entomopathogenic Bacteria, Bacillus thuringiensis subspecies israelensis and thuringiensis. Ann. Èntomol. Soc. Am. 2013, 106, 454–462. [Google Scholar] [CrossRef]
- Pujiastuti, Y.; Riskal, A.; Suparman; Arsi, A. ; Gunawan, B.; Sulistyani, D.P. Effectiveness of Proteins and Supernatants Isolated from Bacillus Thuringiensis-Based Bio-Insecticides Against Termites Macrotermes Gilvus (Isoptera: Termitidae). IOP Conf. Series: Earth Environ. Sci. 2021, 810, 012049. [Google Scholar] [CrossRef]
- Pandey, P., Singha, L. P., & Singha, B. (2013). Colonization and antagonistic activity of entomopathogenic Aspergillus sp. against tea termite (Microcerotermes beesoni Snyder). Current Science, 105(9), 1216-1219.
- Yii, J.E.; Bong, C.F.J.; King, J.H.P.; Kadir, J. Synergism of entomopathogenic fungus, Metarhizium anisopliae incorporated with fipronil against oil palm pest subterranean termite, Coptotermes curvignathus. Plant Prot. Sci. 2016, 52, 35–44. [Google Scholar] [CrossRef]
- Hussain, A.; Ahmed, S.; Shahid, M. Laboratory and field evaluation of Metarhizium anisopliae var. anisopliae for controlling subterranean termites.. 2011, 40, 244–50. [Google Scholar]
- Al-Zaidawi, J.B.; Karimi, J.; Moghadam, E.M. Entomopathogenic Nematodes as Potential Biological Control Agents of Subterranean Termite, Microcerotermes diversus (Blattodea: Termitidae) in Iraq. Environ. Èntomol. 2020, 49, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, W.; Liu, Y.; Sun, P.; Lei, C.; Huang, Q. The Influence of Allogrooming Behavior on Individual Innate Immunity in the Subterranean TermiteReticulitermes chinensis(Isoptera: Rhinotermitidae). J. Insect Sci. 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Singha, D.; Singha, B.; Dutta, B.K. Potential of Metarhizium anisopliae and Beauveria bassiana in the control of tea termite Microtermes obesi Holmgren in vitro and under field conditions. J. Pest Sci. 2010, 84, 69–75. [Google Scholar] [CrossRef]
- Jessica, J.J.; Peng, T.L.; Sajap, A.S.; Lee, S.H.; Syazwan, S.A. Evaluation of the virulence of entomopathogenic fungus, Isaria fumosorosea isolates against subterranean termites Coptotermes spp. (Isoptera: Rhinotermitidae). J. For. Res. 2018, 30, 213–218. [Google Scholar] [CrossRef]
- Wright, M.S.; Lax, A.R. Combined effect of microbial and chemical control agents on subterranean termites. J. Microbiol. 2013, 51, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Zhang, J.; Chen, W.; Huang, H.; Hu, Y. Characterization of a newly discovered China variety of Metarhizium anisopliae (M. anisopliae var. dcjhyium) for virulence to termites, isoenzyme, and phylogenic analysis. Microbiol. Res. 2007, 162, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Ambele, C.F.; Ekesi, S.; Bisseleua, H.D.B.; Babalola, O.O.; Khamis, F.M.; Djuideu, C.T.L.; Akutse, K.S. Entomopathogenic Fungi as Endophytes for Biological Control of Subterranean Termite Pests Attacking Cocoa Seedlings. J. Fungi 2020, 6, 126. [Google Scholar] [CrossRef] [PubMed]
- Kamarudin, M.A.; Abdullah, S.; Lau, W.H. Efficacy of soil-borne entomopathogenic fungi against subterranean termite, Coptotermes curvignathus Holmgren (Isoptera: Rhinotermitidae). Egypt. J. Biol. Pest Control. 2022, 32, 1–16. [Google Scholar] [CrossRef]
- Samsuddin, A. S. B. (2017). Interaction between an entomopathogenic fungus Metarhizium anisopliae and its host subterranean termites Coptotermes curvignathus during infection process.
- Somalian, M.; Allah, R.K.; Hammad, M.; Ebnalwaled, K. Biological control of subterranean termites (Psammotermes hypostoma ) by entomopathogenic fungi. Sci. J. Agric. Sci. 2019, 1, 21–29. [Google Scholar] [CrossRef]
- Zulfiana, D.; Zulfitri, A.; Lestari, A.S.; Krishanti, N.P.R.A.; Meisyara, D. Production of Conidia by Entomopathogenic Fungi and Their Pathogenicity Against Coptotermes sp. Biosaintifika: J. Biol. Biol. Educ. 2020, 12, 1–9. [Google Scholar] [CrossRef]
- Deka, B. , Babu, A., Peter, A. J., Kumhar, K. C., Sarkar, S., Rajbongshi, H.,... & Talluri, V. R. (2021). Potential of the entomopathogenic fungus, Metarhizium anisopliae in controlling live-wood eating termite, Microtermes obesi (Holmgren)(Blattodea: Termitidae) infesting tea crop. Egyptian Journal of Biological Pest Control, 31(1), 1-8.
- Lopes, R. D. S. , Lima, G. D., Correia, M. T. D. S., da Costa, A. F., Lima, E. Á. D. L. A., & Lima, V. L. D. M. (2017). The potential of Isaria spp. as a bioinsecticide for the biological control of Nasutitermes corniger. Biocontrol Science and Technology, 27(9), 1038-1048.
- Dunlap, C. A., Jackson, M. A., & Wright, M. S. (2007). A foam formulation of Paecilomyces fumosoroseus, an entomopathogenic biocontrol agent. Biocontrol Science and Technology, 17(5), 513-523.
- Padmaja, V. , & Kaur, G. (2001). Use of the fungus Beauveria bassiana (Bals.) Vuill (Moniliales: Deuteromycetes) for controlling termites. Current Science, 81(6), 645-647.
- Keppanan, R.; Sivaperumal, S.; Aguila, L.C.R.; Hussain, M.; Bamisile, B.S.; Dash, C.K.; Wang, L. Isolation and characterization of Metarhizium anisopliae TK29 and its mycoinsecticide effects against subterranean termite Coptotermes formosanus. Microb. Pathog. 2018, 123, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Poinar Jr, George O. "Taxonomy and biology of Steinernematidae and Heterorhabditidae." Entomopathogenic nematodes in biological control 54 (1990).
- Al-Zaidawi, J.B.; Karimi, J.; Moghadam, E.M. Entomopathogenic Nematodes as Potential Biological Control Agents of Subterranean Termite, Microcerotermes diversus (Blattodea: Termitidae) in Iraq. Environ. Èntomol. 2020, 49, 412–421. [Google Scholar] [CrossRef]
- Javed, S.; Ali, A.; Khanum, T.A. Biocontrol potential of the entomopathogenic nematodes (Rhabditida: Steinernematidae and Heterorhabditidae) against the termite species, Microtermes obesi (Holmgren) (Blattodea: Termitidae). Egypt. J. Biol. Pest Control. 2021, 31, 1–5. [Google Scholar] [CrossRef]
- Aslam, A.; Chi, D.-F.; Abbasi, A.; Arshad, M. Biocontrol Potential of Entomopathogenic Nematodes against Odontotermes obesus (Blattodea: Termitidae) under Laboratory and Field Conditions. Forests 2023, 14, 580. [Google Scholar] [CrossRef]
- Afroz, A.; Shaw, S.S.; Naralasetti, R.; Saxena, R.R. Comparative bio-efficacy of indigenous isolates of Heterorhabditis indica from Chhattisgarh against Odontotermes obesus under laboratory conditions. J. Exp. Zoöl. India 2022, 26, 1245–1248. [Google Scholar] [CrossRef]
- Razia, M., & Sivaramakrishnan, S. (2016). Evaluation of entomopathogenic nematodes against termites. Journal of Entomology and Zoology Studies, 4(4), 324-327.
- Wagutu, G. K., Kang’ethe, L. N., & Waturu, C. N. (2017). Efficacy of entomopathogenic nematode (Steinernema karii) in control of termites (Coptotermes formosanus). Journal of Agriculture, Science and Technology, 18(1), 55-64.
- Epsky, N.D.; Capinera, J.L. Efficacy of the Entomogenous Nematode Steinernema feltiae Against a Subterranean Termite, Reticulitermes tibialis (Isoptera: Rhinotermitidae). J. Econ. Èntomol. 1988, 81, 1313–1317. [Google Scholar] [CrossRef]
- Rathour, K. S., Ganguly, S., Das, T. K., Singh, P., Kumar, A., & Somvanshi, V. S. (2014). Biological management of subterranean termites (Odontotermes obesus) infesting wheat and pearl millet crops by entomopathogenic nematodes. Indian Journal of Nematology, 44(1), 97-100.
- El-Bassiouny, A.R.; El-Rahman, R.M.A. SUSCEPTIBILITY OF EGYPTIAN SUBTERRANEAN TERMITE TO SOME ENTOMOPATHOGENIC NEMATODES. Egypt. J. Agric. Res. 2011, 89, 121–135. [Google Scholar] [CrossRef]
- Yu, H. , Gouge, D. H., & Baker, P. (2006). Parasitism of subterranean termites (Isoptera: Rhinotermitidae: Termitidae) by entomopathogenic nematodes (Rhabditida: Steinernematidae; Heterorhabditidae). Journal of Economic Entomology, 99(4), 1112-1119.
- Gibbs, A.; Gay, F.; Wetherly, A. A possible paralysis virus of termites. Virology 1970, 40, 1063–1065. [Google Scholar] [CrossRef] [PubMed]
- Al Fazairy, A.A.; Hassan, F.A. Infection of Termites by Spodoptera littoralis Nuclear Polyhedrosis Virus. Int. J. Trop. Insect Sci. 1988, 9, 37–39. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Palli, S.R. Mechanisms, Applications, and Challenges of Insect RNA Interference. Annu. Rev. Èntomol. 2020, 65, 293–311. [Google Scholar] [CrossRef]
- Zhou, X.; Wheeler, M.M.; Oi, F.M.; Scharf, M.E. RNA interference in the termite Reticulitermes flavipes through ingestion of double-stranded RNA. Insect Biochem. Mol. Biol. 2008, 38, 805–815. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, L.; Zhou, W.; Cai, Q.; Huang, Q. Roles of selenoprotein T and transglutaminase in active immunization against entomopathogenic fungi in the termite Reticulitermes chinensis. J. Insect Physiol. 2020, 125, 104085. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, C.-C.; Zhao, X.-Y.; Guan, J.-X.; Lei, C.-L.; Huang, Q.-Y. Isocitrate dehydrogenase-mediated metabolic disorders disrupt active immunization against fungal pathogens in eusocial termites. J. Pest Sci. 2019, 93, 291–301. [Google Scholar] [CrossRef]
- Esparza-Mora, M.A.; Davis, H.E.; Meconcelli, S.; Plarre, R.; McMahon, D.P. Inhibition of a Secreted Immune Molecule Interferes With Termite Social Immunity. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef]
- Liu, S.; Xia, S.; Wu, D.; Behm, J.E.; Meng, Y.; Yuan, H.; Wen, P.; Hughes, A.C.; Yang, X. Understanding global and regional patterns of termite diversity and regional functional traits. iScience 2022, 25, 105538. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Huang, Q.; Xu, H. Silencing Orco Impaired the Ability to Perceive Trail Pheromones and Affected Locomotion Behavior in Two Termite Species. J. Econ. Èntomol. 2020, 113, 2941–2949. [Google Scholar] [CrossRef]
- Feng, K.; Li, W.; Tang, X.; Luo, J.; Tang, F. Termicin silencing enhances the toxicity of Serratia marcescens Bizio (SM1) to Odontotermes formosanus (Shiraki). Pestic. Biochem. Physiol. 2022, 185, 105120. [Google Scholar] [CrossRef] [PubMed]
- Liu, X. J., Xie, L., Liu, N., Zhan, S., Zhou, X. G., & Wang, Q. (2017). RNA interference unveils the importance of Pseudotrichonympha grassii cellobiohydrolase, a protozoan exoglucanase, in termite cellulose degradation. Insect Molecular Biology, 26(2), 233-242.
- Masuoka, Y. , Yaguchi, H., Toga, K., Shigenobu, S., & Maekawa, K. (2018). TGF β signaling related genes are involved in hormonal mediation during termite soldier differentiation. PLoS genetics, 14(4), e1007338.
- Sugime, Y.; Oguchi, K.; Gotoh, H.; Hayashi, Y.; Matsunami, M.; Shigenobu, S.; Koshikawa, S.; Miura, T. Termite soldier mandibles are elongated by dachshund under hormonal and Hox gene controls. Development 2019, 146, dev171942. [Google Scholar] [CrossRef]
- Toga, K.; Saiki, R.; Maekawa, K. Hox Gene Deformed is likely involved in mandibular regression during presoldier differentiation in the nasute termite Nasutitermes takasagoensis. J. Exp. Zoöl. Part B: Mol. Dev. Evol. 2013, 320, 385–392. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Loss caused by termites annually (Source: This study).
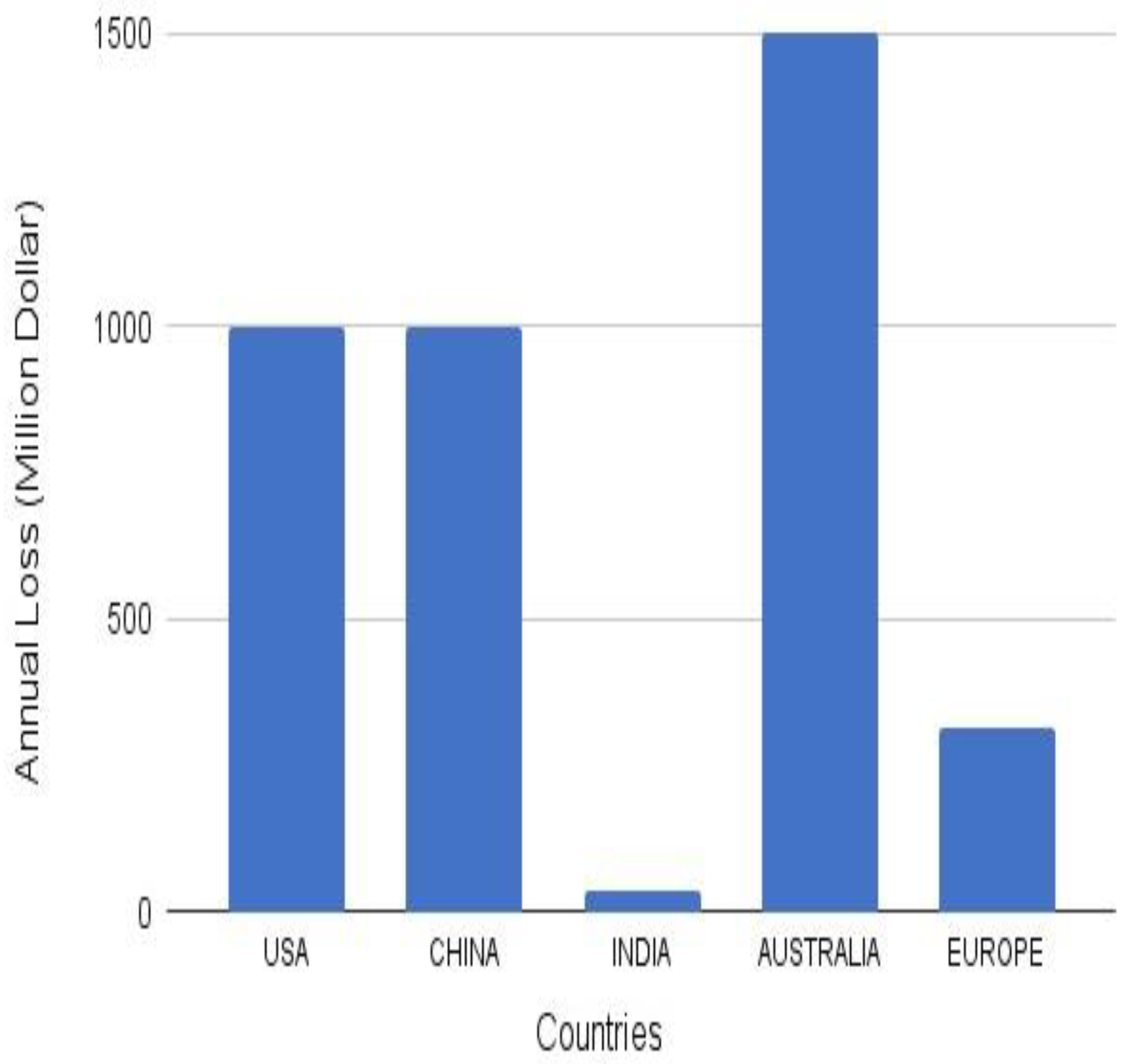
Figure 2.
Most attacked states by Termites in India (Source: This study).

Table 1.
Crops yield reduction caused by Termites.
| Agriculture Crops | Termites | Yield Loss (%) | Place | Symptoms of Damage | Year | References |
|---|---|---|---|---|---|---|
| Wheat | Cryptotermes heimi,Odontotermes obesus,O. redemanni,M obesi, O. feae,T. biformis | 43%- 80% | Rajasthan, Madhya Pradesh, Bihar, Delhi, Punjab, Madhya Pradesh, Andhra Pradesh, Gujarat, Maharashtra, Karnataka, Haryana, Kerala and Tamil Nadu | Affected plants dry up completely and may give rise to white ears | 1995 -2016 | Farook et al., 2019 Sharma et al., 2004 |
| Maize | O. obesus,M. obesi | 30% - 75% | Rajasthan, Madhya Pradesh, Haryana, Punjab | From seedling emergence to ear growth, termites target every stage of the maize crop | 1984-2010 | Parween et al., 2016 Rana et al., 2021 |
| Pulses | Odontotermes obesus, Odontotermes parvide | 25%-90% | Rajasthan, Madhya Pradesh, Haryana, Punjab | Bore holes on buds, flowers or pods Silvery patches and streaks on leaves |
1990-2012 | Paul et al., 2018 |
| Groundnut | Microtermes spp., Odontotermes spp., Amitermes spp., Microtermes spp., Odontotermes spp. | 15%-30% | Punjab, U.P., Rajasthan, Delhi, Haryana, Karnataka, Gujarat, M.P., Maharashtra, Kerala | Root and stem hollowingPod bore holes | 1984-2012 | Paul et al., 2018 |
| Soyabean | M. albopartitus,M. redenianus,A. latinotus,A. tenax | 20%-25% | Punjab, Haryana | Stem lesions, Wilted or curled leaves | 1980-2012 | Paul et al., 2018 |
| Coconut | Microtermes spp., Microcerotermes spp., Coptotermes spp. | 20%-30% | Western coast of India | Yellowing of leavesStunted growth | 1990-2016 | Paul et al., 2018 |
| Sugarcane | Odontotermes redemanni | 74 % | Punjab, Haryana | Yellowing and wilting of leaves | 1986-2015 | Parween et al., 2016 |
| Tea | M. beesoni | 11%- 55 % | North East India | Galls on the roots affecting nutrient uptake Shoot growth is retarded and photosynthetic rates may decrease |
1980-2016 | Roy et al., 2020 |
| Chilli Pepper | O. obesus andM. obesi | 45 % | Rajasthan | Infested leaves develop crinkles and curl upwards Elongated petioles (leaf stalks) Buds become brittle and may drop down. Early-stage infestations lead to stunted growth | 1984-2012 | Parween et al., 2016 |
| Cotton | M. mycophagus,O. obesi | 25% | Rajasthan, Gujarat,Haryana, Punjab and Madhya Pradesh | Drying and drooping of terminal shoots Shedding of squares (immature flower buds) and young bolls (developing cotton capsules). | 1980-2014 | Parween et al., 2016 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



