
Submitted:
20 March 2024
Posted:
20 March 2024
You are already at the latest version
Abstract
This review summarizes the most recent developments of research on protozoan parasite infections ac-quired through food and drinking water and is aimed at gathering updated knowledge on the risk factors, illnesses caused, and measures for prevention.
Reports included dated from 2019 to the present and regarded recent outbreaks and cases of severe infec-tions, prevalence in different population segments and food handlers, and occurrence in food and drinking water. Cryptosporidium spp., Entamoeba histolytica, Giardia intestinalis, Trypanosoma cruzi and Toxoplasma gondii were the microorganisms most frequently involved in large outbreaks and/or severe infections, while Cyclospora cayetanensis was most frequently involved in single case reports. Cystoisospora belli was implied in severe infections in immunocompromised patients, while Blastocystis homi-nis, Sarcocystis spp., Dientamoeba fragilis, Endolimax nana and Pentatrichomonas hominis appeared to be less relevant for number of symptomatic infections and pathogenic potential.
A progressive improvement in surveillance of protozoan infections and infection sources was highlighted, especially in developed countries. The apparent increase in cases was concomitant with the implementa-tion of reporting systems and the application of molecular diagnostic methods that represent valuable in-struments for establishing epidemiological links and preventing or limiting outbreaks. The pathogenic role and sources of infection should be better elucidated for some species.
Keywords:
pathogenic protozoans
; outbreaks
; cases
; opportunistic infections
; prevalence
; dietary sources
1. Introduction
Protozoans are unicellular eukaryotes once classified in the subkingdom Protozoa [1], and currently reclassified in different clades of the lineage Eukaryota [2]. However, the common names “protozoan” or “protozoans” are still in use for organisms once classified in the Protozoa subkingdom.
Among protozoans some are waterborne and foodborne parasites of humans and animals and their pathogenicity is determined by the ability to invade host intestinal cells by mechanical intrusion, attachment and enzymatic digestion [3]. The oocysts and cysts of protozoa are the infectious stage of these organisms released with feces by infected hosts in the environment. These are very resistant to disinfectants and other external agents, which explains their wide distribution and long term persistence [4,5,6,7,8,9,10,11,12,13]. Despite cysts generally do not multiply outside the host, they may survive in moist foods for months at refrigeration temperatures, enhancing their ability to cause infection [14].
In 1987, the World Health Organization (WHO) declared that protozoan parasites commonly transmitted by contaminated food and water, mainly Entamoeba histolytica, Cryptosporidium spp., and Giardia intestinalis (also denominated G. lamblia or G. duodenalis [15]), represent a heavy public health concern and cause malapsorption in children especially in developing countries [16]. These microorganisms cause intestinal parasitic infections (IPIs) and are a cause of prolonged diarrhea with detrimental effects on children’s growth [17]. Data from 7,800 children positive for intestinal protozoan parasites enrolled in the Global Enteric Multicenter Study (GEMS) in South Asia and Sub-Saharan Africa, showed that growth parameters length/height-for-age (HAZ), weight-for-age (WAZ), and weight-for-length/height (WHZ) determined after 60 days from infection significantly decreased in children with 0–11 months of age infected with Giardia spp., Cryptosporidium spp. or Entamoema histolytica. Other effects of protozoan parasitoses can be intellectual impairment and even death [18].
The Food and Agricultural Organization of the United Nations (FAO) and World Health Organization (WHO) included in the multicriteria-based ranking of 24 foodborne parasites also some of the protozoans transmissible by the oral route, which were, in descending order of pathogenicity, Toxoplasma gondii, Cryptosporidium spp., Entamoeba histolytica, Trypanosoma cruzi, Giardia duodenalis, Cyclospora cayetanensis, and Balantioides coli (new name for Balantidium coli [2]) [19]. Other species with less defined pathogenic roles or lower prevalence in humans are Cystoisospora belli (formerly Isospora belli [20]), Sarcocystis spp., Blastocystis hominis, Dientamoeba fragilis, Endolimax nana, and Pentatrichomonas hominis (formerly Trichomonas hominis [21]) [8,22,23,24,25,26,27,28].
The distribution of intestinal protozoan parasites in different continents was illustrated by a study on prevalence in 2517 international travellers returning with illnesses. Giardiasis accounted for 82.3% cases and was most frequently acquired in South Central Asia (45.8%) and sub-Saharan Africa (22.6%), cryptosporidiosis was acquired for 24.7% in sub-Saharan Africa and for 19.5% in South-Central Asia, cyclosporiasis for 31.3% in South East Asia and for 27.3% in Central America, cystoisosporiasis was acquired for 62.5% in sub-Saharan Africa [29].
Some foodborne protozoans were also associated to cancer development, namely T. gondii infection was associated to pituitary adenoma, ocular tumors, meningioma, leukaemia and lymphoma, brain and breast cancers. T. cruzi was associated to esophageal carcinogenesis, leiomyosarcoma, gastrointestinal cancer, colon cancer and uterine leiomyoma and Cryptosporidium spp. to different cancer types [30,31]. The latter favored the development of colorectal cancer (CRC) in a murine model [32].
This review aimed to collect the latest information on diseases caused, risk factors, infection sources and prevention for protozoan infections acquired through the dietary route. Therefore, scientific articles published since 2019 were searched in the databases Google Scholar (https://scholar.google.com/schhp?hl=it, accessed on 28 December 2023) and Scopus (https://www-scopus-com.bibliosan.idm.oclc.org/search/form.uri?display=basic#basic, accessed on 27 December 2023) firstly with the keyword combinations “protozoan food (or drinking water) human infection case (or outbreak)”. Then, the names of organisms retrieved in the first screening were applied singly in new searches in which the term “protozoan” was substituted with the genus or species name. The search results were ordered by pertinence and screened until not finding further relevant information.
2. Waterborne and Foodborne Protozoan Infections
The first database search led to the identification of the protozoan genera and species of interest for this review and resulted in the collection of systematic reviews and meta-analyses, epidemiological studies in the general population and immunocompromised patients and as well as studies on occurrence in food handlers, water and raw vegetables that considered different parasites at the same time.
Experimental studies considered used different detection methods, i.e. optical microscopy, molecular methods, or both, applied to stool, water and food samples. Most frequently the wet mount observation, but also Ziehl-Neelsen (ZN) acid fast staining and its modifications or other staining methods required for some species were applied [33,34,35,36,37,38,39]. These methods allow identification based on morphological features such as cell dimensions, shape and presence of cilia or flagella. For example, cryptosporidia oocysts are small (4-6 μm in diameter), while Cyclospora, and Cystoisospora form large oocysts with a diameter of 20-33 μm [35].
Microscopic examination is labor-intensive, requires experienced personnel and is poorly sensitive. Moreover, it cannot differentiate pathogenic from non-pathogenic members of some genera, such as Entamoeba [40]. To increase the sensitivity, microscopical examination must be carried out for at least 3 stool samples collected over a 10-day period and PCR should also be performed. Other techniques that were used for protozoan infection diagnosis or detection are immunoassays, such as enzyme linked immunosorbent assay (ELISA), enzymatic immunoassays (EIA), rapid immunochromatographic tests and immunofluorescence assays [8,17,28,41,42,43,44,45,46,47,48,49,50,51], and molecular genotyping methods specific for each organism in most investigations.
The advantage of using molecular methods for protozoan infection diagnosis was demonstrated in different studies [28,52,53].
2.1. Parasitic Protozoans in the General Population
A review of approximately 1000 studies published in the five years preceding 2021 reported a IPI prevalence of 5.9% in children aged 0 - 19 residing in European countries. B. hominis was the protozoan most commonly detected, with a prevalence of 10.7% among positive samples. Cryptosporidium spp., G. duodenalis E. nana and D. fragilis were also reported. Prevalence rates in single studies ranged between 1.3% for Cryptosporidium and 68.3% for D. fragilis [54].
In Brazil, among 1,277 stool samples from a clinical laboratory 12.69% were positive for at least one protozoan with higher percentages for children aged 0 – 10 years and teenagers. The decrease of infection rate with increasing age could be explained with acquired immunity and improvement of the hygiene practices. The most frequently detected pathogenic protozoans were, in order, Endolimax nana, Giardia intestinalis and E. hystolitica/dispar [18].
A cross-sectional study conducted in a rural settlement in Argentina showed that B. hominis was the most prevalent pathogenic protozoan parasite in pre-school and schoolchildren (57.7%), followed by G. intestinalis (24.7%) and the Entamoeba complex (10.1%) [55].
Prevalence of IPIs in Ethiopia was 25.01% with E. histolytica/dispar as the most prevalent protozoans (14.09%) followed by G. intestinalis (10.03%) and Cryptosporidium spp. (5.93%) [56]. E. histolytica and G. duodenalis were the most predominant protozoan parasites in patients with diarrhea in Tigrai, Ethiopia [57,58].
In Jordan among patients with diarrhea E. histolytica/dispar, B. hominis, and C. parvum subtype IIaA15G2R1 showed a prevalence of 12.6%, 6%, and 0.6%, respectively [59].
In Pakistan prevalence of IPIs was higher in rural areas (23%) and the most prevalent protozoan parasites were E. histolytica (48%), G. intestinalis (17%) and P. hominis (5%). The intestinal parasitic infections were more prevalent in children possibly as a result of poorer hygiene [60]. The same parasites and C. parvum were also detected among 3748 children between the age of 1 and 15 years between February 2021 and January 2022 in Karbala, Iraq [61].
Source of drinking water, eating raw vegetables and hygiene related factors such as unavailability of toilets, handwashing habit, and fingernail trimming were strongly associated with IPIs [56,57,60].
The concern of under-reporting of protozoan infections was demonstrated by a prospective study on the prevalence of cryptosporidiosis and giardiasis in children from January to February 2021 in Latvia. In that study prevalence in one month equalized that officially reported annually from year 2000 to 2020 by the Centre for Disease Prevention and Control of Latvia [62].
2.2. Parasitic Protozoans in Patients Immunocompromised or with Predisposing Conditions
Different studies investigated the prevalence of protozoan pathogens in immunocompromised patients since most protozoan intestinal parasites can cause opportunistic infections, defined as “serious, usually progressive infections by a microorganism that has limited or no pathogenic capacity under ordinary circumstances, but which has been able to cause serious disease as a result of the predisposing effect of another disease or of its treatment” [63], in immunocompromised persons [10,20,36,49,50,64,65,66,67,68,69,70,71,72,73].
In three South African hospitals, the prevalence of Cystoisospora and Cryptosporidium spp. was significantly higher in patients infected with the Human Immunodeficiency Virus (HIV) compared to not infected ones among patients of all ages with diarrhea [74].
Among HIV-infected patients in central Ethiopia prevalence of intestinal protozoans was 12.9% decreasing in order for Cryptosporidium spp., E. histolytica, G. intestinalis, and Pentatrichomonas hominis. Multivariate logistic regression indicated that regularly eating uncooked food was a risk factor significantly associated with intestinal parasitic infections [39].
Case-control studies carried out in Lybia and Ghana indicated a higher prevalence of IPIs among diabetic patients compared to not diabetic controls [75,76]. This could be a consequence of the inability of the immune system in these patients to effectively contrast pathogens due to the reduced activity of phagocytes and cytokine production resulting from chronic inflammation. Cryptosporidium spp., C. belli and D. fragilis were isolated more frequently from diabetic patients, while E. histolytica/dispar and Blastocystis spp. showed a higher prevalence in non-diabetic patients. These findings were in accordance with previous studies carried out in areas with lower rates of parasitic infections [76]. Multivariate analysis indicated that IPI was associated with diabetes mellitus, with having diabetes for more than five years and having comorbidities. Coinfection with more than one parasite was more frequent in diabetic patients [75]. In a systematic review and meta-analysis, it was found that the prevalence of IPIs caused by Cryptosporidium spp. and Blastocystis spp., was higher in patients with diabetes than in controls [77].
In children with chronic liver diseases (CLD) of different etiology, protozoan parasites detected by unstained techniques were significantly higher compared to children with no CLD, most probably for immunity impairment in CLD [78].
In a case-control study carried out in Poland, prevalence of pathogenic protozoans G. intestinalis, Cryptosporidium spp. and Blastocystis spp. was found to be significantly higher in patients with hematological malignancies, especially large B-cell lymphoma and plasma cell myeloma, than in healthy controls [79].
Diarrhea in kidney transplant recipients was attributed to infective etiologies in 64.2% cases with G. intestinalis and Cryptosporidium as the most common causative pathogens [80].
2.3. Parasitic Protozoans in Food Handlers
Different studies analyzed the presence of intestinal parasites in food handlers since these persons can transmit the infections to a high number of subjects. According to a recent systematic review and meta-analysis, prevalence of intestinal protozoan parasites among food handlers has been documented in 28 countries worldwide with more reports from Ethiopia (39 studies) and Iran (24 studies). The estimated pooled global prevalence of intestinal pathogenic protozoan parasites in the stool of food handlers was 0.147% decreasing in order for B. hominis, E. histolytica/dispar, Cryptosporidium spp., G. intestinalis, E. nana, and D. fragilis and diminishing from 1990 to 2020. The estimated pooled prevalence in different World Health Organization (WHO) regions [WHO region
Codelist https://apps.who.int/gho/data/node.metadata.REGION?lang=en, accessed on 23 January 2024]. ranged between 0.318% and 0.010%, decreasing in order for Western Pacific Region, Americas, Eastern Mediterranean Region, African region, South-East Asia Region and European Region. Among countries, Gambia had the highest pooled prevalence (0.501%) [81].
A systematic review and meta-analysis on prevalence of intestinal parasites among Ethiopian food handlers estimated that E. histolytica/dispar was the most frequent protozoan with 11.95% prevalence [82]. This was reaffirmed in studies regarding different parts of the country [83,84,85].
Among food handlers in Belgarn province, Saudi Arabia, 52.7% participants were infected with parasites, among which B. hominis was the most frequent (78.4%), followed by G. intestinalis (8.1%), C. parvum and E. histolytica (2.7%). Double infections and triple infections were frequent [86].
Among food handlers preparing and distributing foods in hospitals in Turkey, 59% had intestinal parasites of whom 27% were positive for Blastocystis spp., 25% for T. gondii, 10% for E. histolytica, 7% for Entamoeba coli and 5% for G. intestinalis. The result underlined the importance of investigating the presence of parasites in these employees to protect patients, particularly those who are immunocompromised [87].
2.4. Pathogenic Protozoans in Drinking Water
According to a systematic review of waterborne parasitic infections occurring worldwide from 2017 to 2020, drinking water was the source of infection in about 18 outbreaks and the involved protozoans were Cryptosporidium spp., Giardia spp., Blastocystis spp., C. cayetanensis, E. histolytica, D. fragilis, and T. gondii [88].
A comprehensive review recapitulating 416 outbreaks of illnesses caused by unicellular parasites occurring from 2017 to 2022, found that drinking water was involved in 17.06% instances, while the remainder was associated with recreational water and swimming pools [89]. Reports regarded mainly developed countries, and this was attributed to the efficiency of their national surveillance systems and use of more sensitive detection methods, but also to the fact that many outbreaks in developing countries were not published in peer-reviewed journals. Therefore, the occurrence of waterborne outbreaks in developing countries was probably underestimated also considering the poor hygiene level and inadequate water treatment standards in these countries [88,89].
In a study on the occurrence of pathogenic protozoans in drinking water in African countries, Cryptosporidium spp. was detected in 4% - 100% samples of tap water and sachet water and was reported in Egypt, Ethiopia, Ghana, Uganda, Zambia and Zimbabwe. Giardia was detected in tap water in Egypt, Ethiopia, South Africa, Sudan and Uganda with prevalence ranging between 1 and 38%. C. cayetanensis was detected with a 5% prevalence in tap water in Ghana [14].
2.5. Pathogenic Protozoans in Raw Vegetables
Contamination of leafy greens caused many outbreaks of protozoan infections worldwide [33]. According to a systematic review and meta-analysis, protozoan contamination in vegetables and fruits was documented in 44 countries with a pooled prevalence of 20% for vegetables and 13% for fruits. The pooled prevalence in vegetables was 11% for Cryptosporidium spp., 9% for G. intestinalis, 8% for B. coli and E. histolytica, 6% for T. gondii, C. cayetanensis and B. hominis. In fruits, the pooled prevalence was 9% for E. histolytica, 7% for G. intestinalis and 5% for Cryptosporidium species [90]. The estimated pooled prevalence of parasitic protozoans in different WHO-regions was 37% in South-East Asian region, 26% in the Americas, 22% in the Eastern Mediterranean region, 16% in the African region, 14% in the Western Pacific region and 14% in the European region. At the country level, Nepal had the highest pooled prevalence of 92% [90].
Studies from Egypt, Ethiopia, Ghana, Libya, Nigeria, reported a prevalence in fresh produce of Cryptosporidium spp. and G. intestinalis of 0.8 – 81% and 2 – 99%, respectively, while C. cayetanensis had an overall prevalence of 12 – 21% in Egypt, Ethiopia and Ghana and E. histolytica an overall prevalence of 5 - 99% in Burkina Faso Egypt Ethiopia Nigeria and Sudan [14].
Other studies reported the detection of G. intestinalis in vegetables in Morocco, United Arab Emirates (UAE), Mozambique, Ethiopia, Italy and Spain, Cryptosporidium spp. in Morocco, Ethiopia, Italy and Spain, E. histolytica/dispar/moskowsky in UAE, Mozambique and Italy, T. gondii in Morocco and Spain, C. cayetanensis in Spain, B. hominis in Mozambique and Spain and B. coli and E. nana in Mozambique [13,34,38,40,91,92,93,94] In one study Cryptosporidium spp. was detected only by quantitative PCR (qPCR) and not confirmed by microscope observation [93].
Eating crops irrigated with reused water represents a risk for protozoan infection so a study was carried out on wastewater treatment plants in Japur, India, and found that the risk of infection decreases in the order for moving-bed bioreactor, the activated sludge process and, the sequencing batch reactor [95].
The following sections are dedicated to individual pathogenic protozoans sorted by relevance according to the number of literature sources retrieved in the second search round with a priority for those included in the FAO/WHO ranking [19].
3. Cryptosporidium spp.
The genus Cryptosporidium, clade Sar Alveolata, phylum Apicomplexa, family Cryptosporidiidae [2], comprises 44 validated species and more than 120 genotypes [96,97]. Among these, C. hominis and C. parvum, are responsible of nearly 95% of human infections with a predominance of C. parvum reported in many countries [98,99,100,101,102]. C. hominis was found to be dominant in Lebanon, Israel, Egypt Tunisia and China [103,104].
Other species and genotypes reported in humans are C. andersoni, C. bovis, C. canis, C. cuniculus, C. ditrichi, C. erinacei, C. fayeri, C. felis, C. meleagridis, C. muris, C. scrofarum, C. suis, C. tyzzeri, C. ubiquitum, C. viatorum, C. xiaoi, C. occultus, Cryptosporidium chipmunk genotype I, skunk genotype, horse genotype and mink genotype [10,96,98,101,103,105,106] C. meleagridis can infect both birds and mammals [107]. Cryptosporidium chipmunk genotype I, associated to rodents, was the third cause of human cryptosporidiosis in Sweden. The rodent-associated C. mortiferum was also common in that country [99,106].
Cryptosporidium subtype families are distinguished on the basis of glycoprotein (gp60) gene sequences. Five subtype families of C. hominis with very divergent sequences, Ia, Ib, Id, Ie, and If are common in different countries [67]. The most predominant identified C. hominis subtype was IfA12G1 in the USA, IbA10G2 in the UK, Sweden, and Australia, and IbA9G2 in French Guiana and Germany [10]. The rare subtype C. hominis IeA12G3T3 was isolated in Qatar in asymptomatic food handlers [108]. IaA14R4 was the dominant C. hominis subtype in Chinese children [104].
C. parvum subtype family IIc, followed by IIa, and IId are widespread in different countries [67]. C. parvum subtype family IIa is commonly reported in dairy calves, and subtype family IId in sheeps and goats [105,107]. The latter was reported in humans, in the Middle East and Iran, in Portugal, Ireland, the Netherlands, Australia and Sweden [103].
The C. parvum subtype IIaA15G2R1, associated to calves, predominated in Ontario, Canada, USA and UK [100,101] and became increasingly prevalent in China with the diffusion of intensive farming [102]. C. parvum subtype IIaA19G1R1 predominated in Norway [10] and C. parvum subtype IIdA20G1b predominated among asymptomatic food handlers in Qatar [108].
Sweden was the country with the highest reported diversity of Cryptosporidium species. For C. hominis, four different subtype families and two new subtypes were identified. C. parvum isolates belonged to the subtypes IIdA22G1c, IIdA24G1, IIdA15G2R1 and IIaA16G1R1b and to the new subtype families IIy and IIz. C. mortiferum isolates were of subtype XIVaA20G2T1, and two new subtype families (IIIk and VId) were identified [99].
C. hominis subtype IfA12G1R5 was associated with increased incidence of cryptosporidiosis in the USA and became dominant over C. hominis IbA10G2 that was involved in a massive waterborne outbreak occurring in Milwaukee, Wisconsin, in 1993. It is also common in Australia and New Zealand and, according to comparative genomics, it originated from subtypes in East Africa and Europe recombined with local US subtypes [109].
Cryptosporidium life cycle is divided into asexual and sexual phases. The thick-walled oocyst represents the infectious stage of this organism shed by the feces of infected hosts and resistant to all the available anti-coccidial drugs. The oocyst contains four sporozoites that are released in the host gastrointestinal tract once ingested. The sporozoite adheres to the microvilli, a process favored by GP40, GP900 and circumsporozoite-like (CSL) glycoproteins, attaches to the enterocyte surface and is incorporated in a parasitophorous vacuole derived from the host-cell membrane. Here the parasite forms a feeder organelle and a channel that connects the vacuole with the host cell cytoplasm. In the vacuole the sporozoite divides asexually in trophozoites, and then in type-1 meronts which contain eight merozoites. These begin an asexual reproduction cycle in the epithelial cells. Alternatively, the merozoites initiate the sexual cycle differentiating into type-II meronts from which four merozoites form by asexual division, and after infection of other enterocytes, they differentiate into micro- and macro-gametes. The mature micro-gametes leave their host cell and fertilize the macrogametes forming a zygote that goes through meiosis giving rise to the oocysts. The oocysts are thin- or thick-walled: the thin-walled oocysts cause autoinfection, while the thick-walled oocysts are excreted. A single infected host can shed up to one thousand oocysts [10].
Cryptosporidium spp. can be transmitted by direct contact with contaminated fecal matter from an infected host or through contaminated water or food. Transmission by respiratory secretions or coughing was also documented [110]. The infective dose calculated in healthy volunteers was 132 oocysts for C. parvum and 10 to 83 oocysts for C. hominis [98].
Exposure to 64.2 °C for more than 5 min or to 72.4 °C for 1 min and ultraviolet (UV) light renders the oocysts non-infectious. C. parvum oocysts can survive at -20 °C for long, but not at -70 °C. The most effective disinfectants against Cryptosporidium oocysts are chlorine dioxide, hydrogen peroxide, or ammonia but the needed concentrations and exposure times are unfeasible in practice [10].
3.1. Diseases Caused by Cryptosporidium spp.
Criptosporidia are carried asymptomatically or cause acute diarrhea that is self-limiting in immunocompetent hosts and lasts for 3 - 12 days [10]. However, the infection can be life-threatening in patients who are immunocompromised for different reasons. In HIV infected patients with acquired immunodeficiency syndrome (AIDS) cryptosporidiosis is the primary cause of chronic diarrhea with a high mortality rate. In these patients Cryptosporidium spp. can cause biliary tree inflammation and biliary tract blockage, sclerosing cholangitis, papillary stenosis, and pancreatitis [73]. In AIDS patients Cryptosporidium spp. was isolated from the gallbladder, the ductus pancreaticus and the respiratory tract, including bronchioles [10]. The prevalence of cryptosporidiosis in HIV-infected persons ranged from 5.6% to 25.7% in Africa, 3.7% to 45.0% in Asia, 5.6% to 41.6% in South America, and 2.6% to 15.1% in Europe. Higher infection rates and more severe clinical outcomes were seen in HIV-positive persons with CD4+ cell counts lower than 200 cells/μl [67].
Cryptosporidium infection was frequently seen in children with primary immunodeficiency with the potential to cause sclerosing cholangitis and pulmonary infections [73]. In AIDS patients and children, C. parvum is less virulent than C. hominis and was mostly associated with vomiting and chronic diarrhea [10].
Cryptosporidium infections cannot be treated in immunocompromised subjects since the only approved drug, nitazoxanide, can only be administered to immunocompetent patients. Moreover, there are presently no vaccines available [73,111].
Recent studies have reported the occurrence of cryptosporidiosis in hemodialysis patients and in renal transplant patients in low- and middle-income countries. In contrast, human cryptosporidiosis occurs in persons of various ages and immune statuses in industrialized nations, probably as a reflection of reduced immunity resulting from a less frequent exposure to Cryptosporidium oocysts [67]. In France from 2017 to 2019 cryptosporidiosis occurred for 40% in immunodeficient patients, predominantly those with organ transplantation and were apparently more frequent in these patients. Interestingly, cases observed in immunocompetent patients became dominant after the improvement of cryptosporidiosis monitoring, though still largely underestimated because mild diarrhea forms are rarely investigated [98].
The most common sequelae of cryptosporidiosis were long-lasting diarrhea (25%), abdominal pain (25%), nausea (24%), fatigue (24%) and headache (21%). Symptoms meeting the definition for IBS were described in 10% of cases for up to 36 months, more frequently in children than in adults and occurring for 78% among 5–17 years of age and for 63% between 6 months and 4 years of age [112]. Joint-related symptoms, were also reported [113]. Recently, a case of chronic urticaria associated with Cryptosporidium spp. was described in a 17-month-old baby in Isfahan, Iran [114].
In Poland three cases of middle-aged immunocompetent persons with massive C. hominis infection and chronic diarrhea, large intestine–cryptitis and damage of lamina propria were described [115].
Diarrhea is a common problem in solid organ transplanted patients, for which a prevalence of 20–50% was reported. It was found that a large proportion of diarrhea cases in liver transplanted patients was caused by Cryptosporidium spp. but the lack of vigilance and awareness led to delays in diagnosis resulting in immunorejection and shock. A case recently reported in China regarded a 55-year-old man with diarrhea of food origin lasting for 20 days who had difficulty breathing. Suspension of immunosuppressive treatment and unfruitful treatments with antibiotics and antifungals led to worsened patient conditions with the development of septic shock and multiorgan functional impairment with the need for a ventilator and Continuous Renal Replacement Therapy. Only when, after ten days of hospitalization, a stool specimen showed the presence of a large number of Cryptosporidium oocysts and NGS showed the presence of the organism also in blood, the infection was resolved by treatment with nitazoxanide [73].
The association of Cryptosporidium infection with CRC and other cancers was supported by different studies. In a case-control study carried out in the in Jilin province, China, a correlation was found between Cryptosporidium spp. infection and CRC, liver cancer and, for the first time, esophageal and small intestine cancers. The infection rates in CRC, esophageal, liver and small intestine cancers were 17.24%, 6.25%, 14.29% and 40%, respectively. The isolates were identified as C. parvum with 18S rRNA sequences identical to those of cattle isolates, suggesting zoonotic transmission. C. parvum subtypes IIaA15G2R1 and IIaA15G2R2 were predominant in CRC, while IIaA13G2R2 was identified for the first time in CRC and liver cancers [31].
In a case-control study performed in Lorestan Province, Western Iran, during October 2019–August 2020, a 42.5% prevalence of Cryptosporidium infection was observed in CRC patients and a 12.6% prevalence in controls, with a significant difference (p < 0.001) between the two groups [116].
A study in a murine model showed that C. parvum favored the development of CRC by inducing an upregulation of cancer-promoting genes and pathways in mice orally infected with C. parvum or C. muris oocysts [32].
3.2. Epidemiology of Cryptosporidiosis
Beyond being included among the foodborne parasites of greatest global concern for human health and trade implications by FAO/WHO [19], in 2016, Cryptosporidium spp. was ranked by the European Network for Foodborne Parasites (Euro-FBP) as the second highest priority foodborne parasite in Northern and Western Europe, and the eighth highest priority in Eastern and South-Western Europe [117].
According to the GEMS, Cryptosporidium spp. was among the enteric pathogens associated to an increased risk of death in children aged 12–23 months with moderate to severe diarrhea (MSD), and was among the four major pathogens associated to MSD and growth faltering in developing countries. Indeed, most cases of cryptosporidiosis occurred before the age of two in low income regions such as rural Western Gambia, Kenya, Mali and Mozambique [118,119,120]. The same was confirmed by the MAL-ED (Etiology, Risk Factors and Interactions of Enteric Infections and Malnutrition and the Consequences for Child Health and Development Project) project reported that for eight countries of Africa, Asia, and South America [121], while in industrialized nations it occurred more frequently over the age of 2 years, probably for delayed exposure and better hygienic conditions [112]. According to a case-control study carried out in South West Ethiopia, diarrhea caused by Cryptosporidium in children younger than two years was associated with public tap water use and, as diarrhea from other causes, it was strongly associated with malnutrition [122].
In studies carried out in Cameroon [67], Egypt and Iraq [123], it was found that breastfeeding was protective against Cryptosporidium infection in children within 6 months. In a cryptosporidiosis outbreak in Botswana, hospitalization and mortality in children were associated primarily with nonbreastfeeding. However, prolonged breastfeeding for more than 2 years was found to be a risk factor for pediatric cryptosporidiosis in Malaysia [67].
The Global Burden of Disease Study (GBD) in 2016 listed Cryptosporidium as the fifth leading cause of diarrhoea in children younger than 5 years, with more than 57,000 deaths and more than 12.9 million disability-adjusted life years (DALYs) also considering the effects on growth impairment and the increased risk of subsequent infectious diseases [113].
The recognition of Cryptosporidium as a relevant human pathogen led to the improvement of surveillance in many industrialized countries. In Canada, cryptosporidiosis became a notifiable disease in the year 2000. Since then, about 600 cases have been reported annually with a slight increase from 2011 and were probably underestimated [100]. Human cryptosporidiosis became a notifiable disease in Ireland and Sweden in 2004 and in UK in 2010 [9,99]. Cryptosporidium spp. human infections are notifiable in UK but not mandatorily, apart from those that are considered foodborne [98]. In France, no systematic cryptosporidiosis screening is carried out for diarrheic patients and reporting is not mandatory. However, a National Reference Center-Expert Laboratory (CNR-LE), monitored the national epidemiology of cryptosporidiosis since 2017 on the basis of reporting by private laboratories from the entire French territory. Proactive monitoring led to an increase in outbreak reports, with 11 outbreaks identified in France and French Guiana from 2017 to 2020.
In the Nederlands cryptosporidiosis is not a notifiable disease and no surveillance program exists. However, after a large increase in cryptosporidiosis cases in 2012, the epidemiological trends of the infection were better monitored in the country [124].
In Sweden, the incidence of human cryptosporidiosis increased from 0.8 cases/100,000 inhabitants in 2005 to 6.8 cases/100,000 inhabitants in 2022, possibly as a consequence of using molecular diagnostic tools and increased awareness following two large waterborne outbreaks that occurred in 2010 and 2011 [99].
On the other hand, just 28 cryptosporidiosis outbreaks were reported from five Gulf countries, namely, Kuwaiti, Saudi Arabia, United Arabian Emirates (UAE), Qatar, and Oman, since 1988, with no records for Bahrain [119].
3.3. Cryptosporidium spp. in Drinking Water
Control of Cryptosporidium is a major challenge for water sanitization since oocysts can pass through different types of filters. The conventional filtration methods using coagulation, flocculation, and sedimentation are capable of removing 99% of Cryptosporidium cells. Nevertheless, ubiquitous distribution and resistance to disinfectants have led to large waterborne Cryptosporidium outbreaks [4,9,125,126]. For example, the origin of the Milwakee event was caused by the inefficient sanitization of drinking water catched from Lake Michigan [4].
Indeed, Cryptosporidium oocysts can survive for more than 7 days at concentrations higher than 1 ppm of free available chlorine, the recommended concentration for sanitization of drinking water. Moreover, Cryptosporidium spp. was found to be able, under defined conditions, to aggregate and reproduce outside the host and to be protected by biofilms formed in domestic water networks, so that oocyst could be retained and gradually released [5]. Ozone is effective in killing these microorganism in water [10].
In the years 2010-2020, drinking water was the origin of 7% of waterborne outbreaks caused worldwide by C. hominis [113] with C. hominis IbA10G2 involved in 67.1% of instances [101,113].
In Qatar the prevalence of Cryptosporidium was higher among people with no access to household water or those dependent on river water [107]. The high prevalence of Cryptosporidium in Kuwaiti was linked to the winter desert camping areas, where large numbers of overhead water storage tanks are used to store desalinated potable water transported by tanker trucks [119]. In Gambia an increased risk of Cryptosporidium diarrhea in children below 5 years was associated with the consumption of stored drinking water [121].
In USA drinking water-related outbreaks attributed to Cryptosporidium increased from 7% between 1991 and 2002 to 28% between 2013 and 2014, and in the year 2020 748,000 infection cases were estimated, of which only 2% were officially reported [100]. Of cases notified in France from 2017 until 2019, 60% were associated with tap water and well water consumption [9]. In June 2017, an outbreak involved 100 military trainees in Southwest France and was caused by C. hominis IbA10G2 detected in the water network. The contamination was eliminated by restricting the polluting activities within the perimeter of the water resource and installing an additional ultrafiltration module at the outlet of the water treatment plant [9,125].
Other recent outbreaks occurred in French Guiana, and France, and were caused by tap water contaminated by C. parvum IIdA19G2 and sewage-contaminated tap water, respectively. Moreover, the largest outbreak ever described in France, with several thousand cases, occurred in the Provence-Alpes-Côte d’Azur region from October to December 2019. Water sources were contaminated by different C. parvum subtypes (IIdA22G1 / IIaA15G2R1 / IIdA17G2/ IIdA18G1 / IIaA17G1R1) as a consequence of leaching occurring after intense precipitations. The high number of waterborne cryptosporidiosis outbreaks in France can be explained by leaching from the numerous livestock farms, the ineffectiveness of drinking water treatment plants, mainly based on sedimentation and chlorination, and the large use of tap water for human consumption. Nevertheless, Cryptosporidium spp. is not included among the microbiological criteria for drinking water [9].
In Italy, drinking water contaminated by C. parvum subtype IIdA25G1 caused an outbreak of cryptosporidiosis among tourists staying in two different accommodation structures in a small town in the Tuscan Emilian Appennines. Since water disinfection measures may be insufficient to inactivate these parasites, the authors of the report warned that testing drinking water for protozoan pathogens, which is not commonly included in routine water analysis in Italy, should be implemented [126].
Instead, in China Cryptosporidium was listed as one of the microbial contaminant indicators in Chinese Standards for Drinking Water Quality since 2006, though no waterborne outbreaks were reported in the country. In a retrospective epidemiological analysis, it was reported that Cryptosporidium infection was significantly associated with use of the drinking water from wells versus tap water and hand pump water in Shanghai. Drinking unboiled water was also found to increase Cryptosporidium infection rates [127].
The first global comprehensive scoping review on the presence of Cryptosporidium spp. in drinking water regarded groundwater sources intended for human consumption and included literature records from the year 1992 to July 2019 [5].
Pooled detection rates of Cryptosporidium were 19.6% and 13.3% in supply sources and samples, respectively and a baseline prevalence of Cryptosporidium spp. of 10–20% was estimated in global groundwater supplies. The Greater Middle-East presented the highest detection rates for samples and Europe for supply sources. Private supplies, which are largely unregulated, had higher sample and source contamination levels than public supplies. The highest average oocyst counts were recorded in Saudi Arabia (210 oocyst/L), followed by the USA (25 oocyst/L) and Haiti (12.7 oocyst/L) [5].
Pollution sources were of animal origin in 71.4% of cases, with cattle representing 53.3%, and C. parvum the most prevalent species. The mechanism hypothesized for Cryptosporidium entry was groundwater recharge. Indeed, in-situ and ex-situ investigations demonstrated the ability of oocysts to migrate through soil columns, although substantial soil attenuation was observed. Since hydrogeological characteristics were often not reported, the dynamics of Cryptosporidium transport to groundwater supplies are largely unknown. Protected groundwater sources exhibited high levels of Cryptosporidium spp. prevalence (21.4%) as a consequence of bedrock percolation, casing/liner cracking, well deterioration and insufficient or absent wellhead protection, representing a significant public health concern [5].
A study was carried out in Northern Italy to evaluate the occurrence of Cryptosporidium oocysts in a small drinking water treatment plant (DWTP) distributing surface water after filtration, chlorination and UV treatment. Cryptosporidium oocysts were found in 100% influent and 77.8% of effluent samples, and an equal percentage of tap water samples, except samples collected in May and June 2013. Oocysts in the effluent presented intact nuclei, suggesting no inactivation by the high-power UV treatment inserted in the system. The sequence of the 18S ribosomal DNA from the influent was closely related to a genotype found in bank voles, suggesting water contamination by the feces of wild rodents [4]. In a study conducted in Minnesota, it was found that 40% of the drinking water wells examined were contaminated with Cryptosporidium species [128].
3.4. Cryptosporidium in Food
Until 2020 more than 40 foodborne outbreaks were documented worldwide with unpasteurized milk, unpasteurized apple cider contaminated from manure and salads among the most common foods implicated [108,113] and in the years 2010-2020, C. parvum was responsible for 96.5% of foodborne outbreaks worldwide [113]. Until 2000, most cryptosporidiosis foodborne outbreaks were reported in the USA and UK while, later, the majority of cryptosporidiosis outbreaks, related to fresh produce, were reported from Sweden, Denmark, and Norway [108].
A systematic review and a meta-analysis of case-control and cohort studies revealed significant associations of cryptosporidiosis with meat and dairy foods for the mixed population and composite foods for children. The association of cryptosporidiosis with raw milk was significant for both mixed population and children. Associations with cryptosporidiosis were observed for barbecued foods, meat of non-specified origin, except beef, dishes prepared outside the home and raw milk [129]. In a retrospective study, eating raw foods was found to increase Cryptosporidium infection prevalence in China, despite the absence of reports of foods contaminated with Crysporidium oocysts in the country [127].
The association with meat could reflect fecal contamination of carcasses during the slaughter process though Cryptosporidium spp. was isolated from fecal samples but not from carcasses. The association of dishes prepared outside the home and barbecued foods with infections by Cryptosporidium spp. can be linked to contamination by an infected handler during the preparation [129].
A large, 3-year, population-based case-control study involving 17 regional laboratories initiated in April 2013 in the Netherlands led to the identification of hundreds of cases each year with a predominance of C. parvum infections. In the first and third years, foodborne transmission cases were more likely to have had a picnic or barbeque. In the second year, no association with dietary routes was found, while during the third year cases showed a higher frequency of consuming water from sources other than tap water [124].
In Quebec, from 2016 to 2017, confirmed cases of cryptosporidiosis were linked to raw vegetables (13%), raw fruits (11%), unpasteurized drinks (8%) and raw shellfish (1%), with the majority of cases attributed to C. parvum subtypes IIaA15G2R1, IIaA16G3R1 and IIaA16G2R1 [100].
Outbreaks from unpasteurized milk have been described in the USA, Australia, and the UK before 2020 [113] and C. parvum was listed among the microbiological hazards potentially transmissible through milk in the EU [130,131].
In France, C. parvum IIaA15G2R1 caused an outbreak of gastroenteritis in a high school linked to unpasteurized curd cheese and an outbreak from contaminated milk in the Pays de la Loire region. The contaminated dairy foods could not be analyzed, but C. parvum subtype IIaA15G2R1 was isolated in calves in the dairy farms of origin [9].
Pasteurized milk was, instead, the cause of an outbreak caused by C. parvum subtype IIaA19G1R1 in England. The contaminated milk was distributed from a vending machine present in the producing farm. Identity was demonstrated between isolates from two patients and one calf in the farm by multiple locus variable number of tandem repeats (MLVA) profiles at seven loci. In this case, pasteurization efficiency was demonstrated, thus post-pasteurization contamination of the vending machine was deduced. Bacteriological indicators of post-pasteurization contamination persisted even after sanitization of the vending machine. Since C. parvum is a common cause of enteritis in calves and its oocysts can survive in the farm environment also after cleaning and disinfection, pasteurization efficiency is critical for the control of this pathogen. Therefore, the authors warned that placing vending machines on-farm may expose pasteurized milk to recontamination, thus representing a public health concern [132].
Among foods of plant origin, apple juice was the cause of cryptosporidiosis outbreaks in the USA and of a recent outbreak in Norway where C. parvum subtype IIaA14G1R1 caused illnesses among employees who consumed unpasteurized apple from the same container. After inspection of the producing farm, it was inferred that contamination came from a few contaminated apples accidentally dropped during collection. In spiking experiments, it was observed that even vigorous washing with a detergent did not completely remove Cryptosporidium oocysts from the apple surface and these retained their infectivity for at least 4 weeks, so drinking unpasteurized apple juice can expose to the risk of cryptosporidiosis [110].
Fresh produce washed and not washed was not identified as a risk factor, despite being the main vehicle of foodborne cryptosporidiosis outbreaks. Indeed, several case-control studies suggested that the consumption of vegetables might lead to the acquisition of immunity following repeated exposure to low doses of oocysts [124,129]. Eating chicken, salad, tomatoes, and hard cheese was suggested to be a protective factor against Cryptosporidium infection [124]. Nevertheless, many reported outbreaks were caused by raw vegetables [99,100,108,113,133] and in a study in Ghana it was found that not always washing vegetables was significantly associated with the prevalence of cryptosporidiosis in HIV-infected patients [134].
On the other hand, an outbreak caused by romaine lettuce contaminated with C. parvum occurred in Sweden in 2016 among school teachers [133] and in this country cases of cryptosporidiosis peaked in 2019 and 2022, due mostly to foodborne outbreaks caused by leafy greens [99].
Cryptosporidium contamination was detected in 6% of vegetables and fruits sampled in Norway, with mung bean sprouts significantly more likely to be contaminated, in 5% of vegetable samples collected from an area of high livestock production in Poland, in 5% of cilantro leaves, 8.7% of cilantro roots, 2.5% of lettuce samples and 1.2% in samples of radish, tomato, cucumbers, and carrots in Costa Rica. In ready-to-eat packaged leafy greens sold in Canada and grown in the United States, Canada and Mexico, Cryptosporidium oocysts were detected in 5.9% samples [108].
Higher contamination rates of 8.5%, 14.5% and 19.35% were found in pooled samples collected in three sampling periods in Peru in vegetables sold in markets. Even higher contamination levels were reported in studies from African countries, namely 35% of vegetables in Kaduna State, Nigeria, between 11.7% for green onion and 46.7% for rocket in Egypt, with Cryptosporidium oocysts detected in all tested vegetable types. In the Accra Metropolis, Ghana, the prevalence of C. parvum oocysts in vegetables was 16% with the highest contamination in lettuce and lowest in tomatoes and a 10 times higher contamination of vegetables sold at the open-air markets compared to those sold in supermarkets for possible unhygienic handling.
Cryptosporidium spp. occurrence was observed also in fish and seafood that could represent a novel infection route. The first epidemiological study concerning the detection of Cryptosporidium in edible marine fish was carried out in seas surrounding France and found an overall prevalence of 2.3% and 3.2% in two sampling periods, respectively. The identification by molecular methods showed the presence of C. parvum subtypes IIaA13G1R1, IIaA15G2R1, IIaA17G2R1, and IIaA18G3R1, previously detected in terrestrial mammals, and eight genotypes closely related to C. molnari. Saithe showed the highest prevalence of 19%, but positive samples were found also for cod, ling, mackerel, sardine, anchovy, hake and herring. A higher prevalence of Cryptosporidium spp. was observed in larger fishes, during the spring-summer period, and in the North East Atlantic [135].
In shellfish aquaculture sites on Thailand’s Gulf Coast, 13.8% of oyster samples were contaminated by Cryptosporidium species [136].
In the Western Mediterranean region, positive samples were found among synanthropic fish from four farms, i.e. all those examined, and in cultivated fish from three of these farms in sardinellas, European seabass, Mediterranean horse mackerel, blotched picarel and pompano. The only positive fish from the extractive fisheries was a bogue. The identified species were C. molnari, the zoonotic C. ubiquitum, able to affect a wider range of hosts compared to other species, C. scophthalmi and one isolate highly divergent from known Cryptosporidium species/genotypes. Cryptosporidium spp. was also detected in fish fillets possibly for cross-contamination during evisceration. Fish species consumed not gutted and, occasionally, raw or undercooked, can represent a risk for cryptosporidiosis [137].
4. Entamoeba spp.
The protozoan species Entamoeba histolytica, E. dispar, E. moshkovskii, and E. bangladeshi are referred to as the Entamoeba complex because they are morphologically identical [138]. Entamoeba coli is a non-pathogenic species, also transmitted through fecal-oral route, whose complete name is usually cited to avoid confusion with the bacterium Escherichia coli [139]. These protozoans belong to the Entamoebidae family of the Eukaryota clade Amoebozoa, phylum Evosea [2]. Among these species, E. histolytica can cause invasive intestinal and extraintestinal infections. E. histolytica has a simple life cycle involving a quadrinucleated cyst shed with feces that, when ingested with contaminated water or food undergo excystation in the large intestine and release trophozoites able to reproduce via binary fission. These penetrate the intestinal mucosa, forming typical flask-shaped ulcers, and then access the hepatic portal circulation reaching the liver where they can cause an inflammatory reaction that leads to hepatocyte necrosis and abscess formation. The progression of the disease depends on factors such as intestinal microbiome composition, dysbiosis, and reduced cell-mediated immunity. The intestinal infection is more frequent, and most infected people are asymptomatic [7].
The first step of intestinal invasion by E. histolytica is breaching the mucus layer by modulating the transcription of the MUCIN2 gene in host epithelial cells. Once in contact with epithelial cells, the ameba induces apoptosis causing epithelial damage, tissue invasion and strong upregulation of the inflammatory cytokine interleukin 8 (IL-8), with neutrophil infiltration. The amoeba also actively increases the production of host matrix metalloproteinases (MMPs) that break down the extracellular matrix favoring the translocation of intestinal bacteria into the tissue and their dissemination in organs with risk of systemic diseases [140]. Cysteine proteases, prostaglandin E2 and amoebaepores play important roles in trophozoite colonization and intestinal mucose invasion by E. histolytica, while Gal-lectin and a 21 KDa surface protein is involved in extraintestinal dissemination [141].
4.1. Diseases Caused by Entamoeba spp.
After exposure to the parasite, 80% of patients show symptoms in a few days up to 4–6 weeks, although there are reports of infection manifestation years after exposure to a source of infection. Invasive intestinal amebiasis is an acute event characterized by the presence of diarrhea that lasts for 4-5 weeks accompanied by fever [142,143]. Symptoms similar to ulcerative colitis lasting for several months were also reported after E. histolytica infection as well as IBS-like symptoms with periods of abdominal pain and auto-limited episodes of diarrhea alternated to constipation [142,144].
Extraintestinal infections, such as amebic liver abscesses (ALA), peritonitis, lung abscesses, brain abscesses, purulent pericarditis, pneumonia and skin lesions may develop in some patients with severe and potentially fatal outcomes [144,145]. Other severe forms of E. histolytica infections are fulminant necrotizing colitis, colon ameboma and toxic megacolon. Symptoms, such as severe dysentery, fever above 38°C, tachycardia, hypertension, nausea, and anorexia, are observed in severe forms of intestinal amebiasis [145]. The mortality rates of dysenteric amebiasis are less than 1%, but mortality due to complications increases up to 75%. Fulminant necrotizing amebic colitis occurs in more than 50% of cases with severe colitis in 0.5–3% of cases of amebiasis [69]. The severe forms of invasive amebiasis can be observed in young children, pregnant women, the elderly, and immunocompromised subjects. Fulminant necrotizing amebic colitis was associated with large bowel gangrenous necrosis, perforation, and peritonitis with mucosal and submucosal ulcers. Intestinal amebiasis is easily misdiagnosed as IBD and treated with steroids, thus increasing the possibility of severe complications [69]. Indeed, the destroyed host immunity, combined with tissue damage facilitates invasive amebic infection, leading to a rapid progression of the disease [146].
ALA is the most common extraintestinal manifestation of invasive amebiasis and also E. dispar has been isolated from patients with non-dysenteric colitis and liver abscesses, thus indicating its pathogenic potential [147,148]. E. dispar was significantly associated with abdominal pain [145].
E. moshkovskii was found to infect humans in Yemen, India, Indonesia, Colombia, Malaysia, Tunisia, Tanzania, Australia and Kenia and it was reported not to cause diarrhea but a mild abdominal discomfort [53].
E. gingivalis is the only Entamoeba species known to colonize the human oral cavity. A metagenomics analysis showed that E. gingivalis was significantly increased in inflamed periodontal pockets and in a case-control study in Berlin, E. gingivalis was detected in healthy oral cavities in 15% of controls and 77% of patients in inflamed gingival pockets. In wounded live gingival biopsies, it was observed that the amoeba penetrated the cytoplasm of gingival epithelial cells and ingested fragments of the host cell nuclei suggesting an active role of E. gingivalis in the pathogenesis of periodontitis. Notably, periodontitis is associated with increased risk for cardiovascular disease, rheumatoid arthritis, and oral cancer [149].
ALA has been a major parasitological health concern since 1962 in Sri Lanka (SLK), and though its prevalence declined significantly in the island, it remains a common cause of emergency hospitalization in the north. Population-based studies previously demonstrated that it is associated with the consumption of the alcoholic fermented sap of the Palmyra toddy (Borassus falbellifer) that could be contaminated by E. histolytica through water because cysts are resistant to low doses of chlorination, or unhygienic practices in preparation [7]. Prevention could be improved with the supply of safe drinking water, according to the WHO Guidelines for Drinking-Water Quality [150] and safe consumption of alcoholic beverages [7].
A reported complication of ALA was the development of a right atrial thrombus in a 35-year-old man without a past medical history. The proposed mechanism was the development of a local pro-thrombotic state that led to the formation of a hepatic vein thrombus that extended into the venous circulation and reached the right atrium [151].
ALA caused by E. histolytica was reported in a woman in early pregnancy after seven years from a prolonged episode of intermittently bloody diarrhea that she developed during a journey to Indonesia. Infection with E. histolytica was diagnosed by serological testing and abscess fluid and blood were negative for the parasite. This case was successfully treated with metronidazole and paromomycin [152].
ALAs are more often treated medically since they generally respond well to metronidazole treatment, and percutaneous catheter drainage is required in only 15% of cases [147]. Other amebicidal agents that can be used to treat ALA include ornidazole, tinidazole, nitazoxanide, and chloroquine. At present, no vaccine exists to prevent E. histolytica-induced ALA [7]. Paromycin is effective in treating intestinal infections allowing to eliminate cysts. Metronidazole treatment in invasive amebiasis should always be followed by treatment with an agent able to eliminate cysts and prevent relapse since metronidazole is not effective against cysts [151].
A case of E. histolytica infection in a 59-year-old Indonesian man occurred in form of infiltrative appendicitis with a wide area of necrosis, an almost perforated caecum, and inflamed ascending colon. The patient necessitated hemicolectomy with the removal of caecum and ascending colon and an anastomosis between the terminal ileum and transverse colon. He recovered after treatment with metronidazole [69].
Misdiagnosis as Crohn’s disease and administration of corticosteroids led to life-threatening worsening of E. histolytica infections [47,146]. One case occurred in a 49-year-old man with a two-month history of diarrhea, occasionally bloody, accompanied by abdominal pain, who developed multiple ulcerations in the entire colon. He underwent total colectomy, small bowel resections and other interventions and died of sepsis and multiple organ failure. E. histolytica with phagocytosed intracellular red cells was late identified in ulcers of the colon mucosa. In another patient, a 40-year-old man with persistent diarrhea and patchy colitis, misdiagnosed as Crohn’s disease and treated with steroids, the symptoms worsened but E. histolytica trophozoites were seen during endoscopy, so he was treated with metronidazole and paromomycin and recovered. The infection probably originated from a coffee consumed in a street store in India [146]. Another case regarded a 30-year-old male patient hospitalized for suspected malignancy and misdiagnosed with Crohn’s disease. Two months later, the patient developed a large liver abscess and was treated with ceftriaxone and metronidazole. Upon suspicion of ALA, specific tests for E. histolytica were carried out and a positive serum ELISA and positive E. histolytica PCR tests were obtained, so the patient received therapy with metronbidazole and paromomycin and recovered [47].
In a 35-year-old German patient who had recently traveled to Indonesia E. histolytica infection was initially misidentified as colitis associated to non-steroidal anti-inflammatory-drug use because initial stool examination by microscopy and Entamoeba fecal antigen ELISA did not reveal the infection. The patient did not improve with the appropriate therapy so reinvestigation was carried out by examining colon biopsies by real-time PCR and fluorescence in situ hybridization (FISH) that proved the diagnosis of E. histolytica infection. Metronidazole therapy allowed the patient’s recovery [45].
These cases indicated the need to consider infectious etiology before diagnosing IBD or Crohn’s disease and to doubt these diagnoses if a patient does not respond to immunosuppression. Moreover, since a greater prevalence of amebiasis was observed in patients with IBD (16%), compared to the normal population (1.7%), the amebic infection should be ruled out in IBD patients before the administration of corticosteroids [146]. Diagnosis of amoebic infections can be obtained with several ELISA or immunofluorescence antibody tests. However, serology is unable to distinguish active from past infections and is prone to false negatives in immunocompromised patients. Therefore, a commercially available PCR assay for the detection of E. histolytica DNA in stool targeted on the small subunit rRNA genes should be applied [47].
4.2. Epidemiology of Entamoeba spp.
E. histolytica has been listed as one of the top-15 causes of diarrhea in the first 2 years of life and breastfeeding is protective also against this parasite [7].
Amebiasis affects about 50 million people and causes 100,000 deaths annually with a worldwide distribution, though more commonly in tropical and subtropical countries. Infections most often occur in people living in groups as in orphanages, shelters, and prisons. Foodborne exposure infection is the most common route of transmission, especially in case of inadequate hygiene of food handlers [143].
Entamoeba spp. prevalence in the world was found to vary between 0.43% in Belgium and 82.64% in Malaysia and infections by E. histolytica occurred mostly in developing countries or in returning travelers and immigrants from endemic areas [7]. E. histolytica infection is endemic in Iraq and prevalence does not vary seasonally. It was detected in 31.3% samples from patients younger than 12 years of age with diarrhea and E. dispar was detected in 17.5% of those samples by nested PCR able to differentiate the two species. By using real-time PCR, E. histolytica and E. dispar were detected in 35.0% and 18.8% of samples, respectively [153]. The prevalence of E. histolytica infection in children from 1 to 10 years old Sulaimani province, Iraq, was 19.3%, and was significantly associated with raw vegetable consumption and parent’s education level [154]. E. histolytica was the most prevalent parasite, accounting for 80.1% of samples positive for parasites in the stools of food handlers in Erbil city, Iraq [155].
In Abuja, Nigeria, the prevalence of E. histolytica in children was 12% [139] and in the same country, E. histolytica was the most prevalent parasite (5.6%) in food handlers in Kano, with occurrence significantly associated with lack of personal hygiene [156]. In Northern South Africa, E. histolytica was found to be significantly associated with diarrhea and to be more prevalent among HIV patients [145].
In Brazilian regions, a systematic review and meta-analysis reported a pooled prevalence of Entamoeba spp. between 2% and 72% and 18% in patients immunosuppressed for cancer, HIV infection and hemodialysis. Single species prevalence was 7.9% for E. dispar, 3.1% for E. histolytica, and 0.6% for E. gingivalis. E. histolytica frequently caused opportunistic infections in immunosuppressed patients and was one of the most common causes of morbidity in this group, so it should be considered for routine diagnosis in these patients [157].
A prevalence of 19.8%, was found among school children in Arsi Town, West Zone, Oromia Regional State, Ethiopia[158] and in Perak, Malaysia among children aged between 7 and 12 years old, the overall prevalence of Entamoeba complex infections was 21.3% of which E. moshkovskii, E. dispar, and E. histolytica represented 10.7%, 9.0%, and 5.0%, respectively [159]. Hand washing habits were significantly associated with E. histolytica infection [158,159]. A systematic review and meta-analysis based on 29 articles with 36 studies from Brazil (7), India (6), Ethiopia (3), Cote d’lvoire, Kenya, Lesotho, Mexico and Vietnam (2 for each country), Colombia, Ecuador, Cuba, Chile, Cambodia, Iraq, Nigeria, South Africa, Uganda and Yemen (1 for each country) found that the prevalence of intestinal Entamoeba spp. infection was positively, though not significantly, correlated with the lack of safe drinking water, while the lack of toilets had a significant positive correlation with prevalence. A significant positive regression between Entamoeba spp infection and year of study was found for both lack of safe drinking water and lack of toilets [160].
E. histolytica was detected in 12.8% of immunocompromised patients in Sana’s City Hospital, Yemen [161].
4.3. Drinking Water Involvement in Entamoeba spp. Infections
In an outbreak involving 250 cases in Idahluye Bozorg village, Iran, from January 19th to 31st, 2018 the source of infection was network drinking water contaminated by sewage pipe erosion [162].
E. dispar was reported to be a pioneering agent in the formation of biofilms, as observed in an industrial wastewater deep injection disposal well in Florida where it was the most abundant microorganism. This aspect is probably implied in the occurrence and persistence of this microorganism in water [163].
5. Toxoplasma gondii
Toxoplasmosis is a zoonotic parasitic infection with a worldwide distribution caused by the protozoan Toxoplasma gondii, clade Sar Alveolata, family Sarcocystidae [2] and is one of the most important opportunistic parasitic diseases affecting humans and animals [44,64,164].
It was estimated that about one-third of the global population is infected with T. gondii which is one of the most important foodborne parasites worldwide ranged fourth globally and second in Europe [44]. Felids are the definitive hosts in which the sexual reproduction of the parasite occurs and shed the infective oocysts, while other animals and humans, act as intermediate hosts. Up to 70% of the cat population is infected with T. gondii, and the infected cats can shed millions of oocysts in their feces. Oocysts may survive for several years in the environment and infect intermediate hosts through water and food [13,49,64,165,166]. Upon infection, the parasites differentiate into tachyzoites that rapidly divide asexually and spread in the body causing toxoplasmosis [165,167]. Humans and animals can become infected with T. gondii also through the ingestion of the parasite in the form of slowly replicating bradyzoites that originate from tachyzoites. These form cysts in a host’s heart, liver, kidney, brain or skeletal muscle, and, due to the high affinity for nerve cells, also in the brain. This persistent chronic form of the pathogen may persist for a long time in the host, perhaps during the whole life, and it may reactivate and lead to a severe pathology in case of weakened immune response. Patients with dormant toxoplasmosis who receive allogeneic hematopoietic stem cell transplants, or those with graft, are at risk of severe or fatal toxoplasmosis by reactivation of a latent infection [70]. A global mortality rate of 43% was reported for toxoplasmosis in nonallografted hematopoietic stem cell transplant patients [64].
5.1. Diseases Caused by T. gondii
In immunocompetent individuals, Toxoplasma infection is mostly asymptomatic or mild and self-limiting but it can be life-threatening in fetuses and immunocompromised individuals [44,70,165]. Latent infection may be associated with specific neuropsychiatric conditions [46]. Primary infection during pregnancy can cause congenital infection and malformations that may lead to miscarriage, stillbirth in both humans and animals, prematurity, neonatal death and a wide spectrum of clinical manifestations in the newborn. In Europe, 75% of children with congenital T. gondii infection were asymptomatic at birth. However, if left untreated, these children could develop symptoms later in life. Manifestations in congenitally infected children at birth include chorioretinitis, intracranial calcifications, or hydrocephalus. Severe toxoplasmosis in immunocompromised persons mainly manifests as central nervous system (CNS) disease, myocarditis, or pneumonitis [49,50,64].
When the organism is disseminated throughout the body via macrophages, toxoplasmosis can manifest with fever, pneumonia, or brain cysts. Clinical manifestations include myelopathy, encephalitis, brain abscesses, hydrocephalus, short-term memory loss, cognitive impairment, altered mental status, cachexia, hypercalcemia, and stroke. Rare voluminous lesions mimic primary and metastatic tumors. Central nervous system (CNS) involvement, most commonly acute or chronic meningitis, has been reported in 5–10% of cases and usually occurs both in disseminated or focal CNS infection [70]. In patients with AIDS toxoplasmosis can manifest as encephalitis, which is mostly caused by the bradyzoite form of T. gondii [168].
The diagnosis of T. gondii infection is carried out by serological tests, with detection in serum of anti-toxoplasma-specific antibodies. Presence of T. gondii-specific IgM in combination with an avidity of T. gondii-specific IgG antibodies of less than 60% are indicative of acute infection [49]. Laboratory tests are less reliable in immunosuppressed patients so clinical symptoms are evaluated and investigation on possible foodborne transmission through consumption of rare and cured meat, raw shellfish and drinking unpasteurized milk should be considered [64].
In addition, PCR, histological exams, isolation of the organisms and imaging are carried out. Acute infection can be diagnosed by visualization of tachyzoites and infiltration of inflammatory cells in infected tissues or body fluids with immunohistochemical or Giemsa staining [70].
Also, patients treated with immunosuppressants can develop symptomatic toxoplasmosis. A case was reported for a 34-year-old woman under chronic psoriasis therapy with ixekizumab who manifested dry cough and rhinorrhea associated with tender left axillary lymphadenopathy. Serological values indicated that she was affected by a recent primary T. gondii infection most probably caused by regular consumption of rare meat. She was treated with pyrimethamine, leucovorin, and clindamycin [64].
A case-control study based on serological diagnosis showed that rheumatic patients had a significantly higher T. gondii seroprevalence than control subjects. Consumption of raw shellfish was identified through multivariate analysis among the risk factors that affected the T. gondii seroprevalence in these patients [48].
5.2. Epidemiology of T. gondii Infections and Involvement of Food and Drinking Water
Brazil has one of the highest rates of T. gondii infection, and the severity of toxoplasmosis in congenitally infected children is also considered to be the highest in the world. Outbreak notification and epidemiological investigations are compulsory in the country so since 1967, among the more than 35 outbreak reports published worldwide most were from Brazil. The main transmission routes were water, vegetables, fruits, raw or undercooked meats and unpasteurized goat’s milk. The largest global outbreak of toxoplasmosis occurred in 2018 in the municipality of Santa Maria, Rio Grande do Sul, affecting more than 900 people and the main suspected transmission route was water.
A recent outbreak involved 73 employees of the same institution diagnosed with acute toxoplasmosis over 4 months. A case-control study revealed significant associations between cases and eating raw salad at the institution’s restaurant [169].
In June 2016, acute cases of toxoplasmosis were reported in Montes Claros de Goiás, Brazil, in patients attending health facilities, with fever, lymphadenomegaly, ophthalmic alteration, seizure and myalgia. Vegetables and artisan fresh cheese from raw cow’s milk were consumed by 100.0% and 78.6% of the involved patients, respectively. These sources were evaluated in a case-control study and it emerged that the only significant variable was the consumption of artisan fresh cheese from raw cow’s milk. Two samples of artisan fresh cheese and one irrigation water sample used in a small horticultural property tested positive for T. gondii by PCR. In one of the inspected cheese manufacturers, an inadequate factory structure allowed access to cats, which were probably the source of the parasite [170].
The largest described outbreak of toxoplasmosis in Brazil, with 960 confirmed cases, was reported in Santa Maria, southern Brazil, in 2018. The outbreak investigation began when physicians reported increased cases of a syndrome characterized by fever, myalgia, headache, rash and mild gastrointestinal and respiratory symptoms. Pregnant women represented 15% of cases and three fetal deaths, nine abortions and 28 cases of congenital toxoplasmosis occurred. Drinking water was found to be the source of infection by bioassay in piglets and mice carried out with the contaminated water. The necessity of continuous monitoring of public water supplies for T. gondii contamination emerged [171].
Based on the determination of antibodies against T. gondii IgM, unpasteurised milk consumption was found to be a major risk factor for infection in a case-control study comparing females of childbearing age with a previous history of recurrent pregnancy loss and controls in the province of Khyber Pakhtunkhwa, Pakistan [43].
In a systematic review and meta-analysis on seroprevalence of T. gondii in pregnant women in eighteen countries of the WHO Eastern Mediterranean region, a pooled prevalence was 36.5% was found. From the studies that examined foodborne transmission, it emerged that 32.9% of positive women drank unprocessed milk, 43.7% ate raw or undercooked meat, and 40.8% ate unwashed raw vegetables [50].
Multiple outbreaks and cases of toxoplasmosis have been reported recently and regarded also immunocompetent persons. One involved hunters who consumed undercooked venison in Wisconsin, USA. An atypical genotype (haplogroup 12, polymerase chain reaction restriction fragment length polymorphism genotype 5) common in North America, was isolated and characterized for the first time concerning human clinical manifestations [172].
In a case–control study in the Netherlands, it was found that in the cases ascertained by fourteen regional medical laboratories consumed beef, veal and raw/undercooked beef such as steak, steak tartare, and roast beef more frequently than the controls, in the nine months preceding the analysis. Consumption of lamb, duck/goose, big game animals and undercooked pork prepared as raw bacon, spreadable sausages, or toppings was also more often reported by cases. Furthermore, consumption of raw or undercooked crustaceans or shellfish was also reported more often by cases. After adjustment for age, gender, and pregnancy, two factors remained as risk factors that are consumption of meat from large game animals and washing hands occasionally or never before preparing food. Results confirmed those obtained in a study in England and Wales, concerning beef consumption and washing hand habits before food preparation that cross-contamination of food via sources other than food as cause of infection [49]. In a meta-analysis including cohort, case-control, and cross-sectional studies it was observed that individuals who eat raw or undercooked meat have, respectively, 1.2-1.3 times the risk of T. gondii infection compared to those who thoroughly cook meat [166].
Marine mammals such as whales, dolphins, and seals are also parasitized by T. gondii. In June 2020, in a suspected food poisoning case reported in Tokyo, Japan, five of nine people who ingested the raw meat of a common minke whale (Balaenoptera acutorostrata) not previously frozen before consumption showed symptoms such as diarrhea and fever up to 39 °C after 12 h - 5 days post-ingestion. The anti-Toxoplasma antibodies could not be investigated in the patients due to a lack of cooperation. However, molecular and histopathological examinations of whale meat indicated the presence of Sarcocystis spp. and T. gondii of atypical type II genotype. The latter belonged to the ToxoDB genotype #39 isolated from sea otters, sheep, and goats in the USA. Occurrence of T. gondii has already been reported in marine mammalians, namely, genotype ToxoDB-RFLP genotype #300 in Bryde’s whale (Balaenoptera edeni) in Brazil and type II strain in Fin Whales (B. physalus) in Italy. Minke whales in Scotland were positive for T. gondii serum antibodies and a seroprevalence of T. gondii of 60% was found in the Inuits in Canada and linked to the consumption of seal meat [173].
According to a systematic review of studies carried out up to March 2018, 44.1% of documented worldwide outbreaks were oocyst-related. Waterborne infections gave rise to large-scale outbreaks since oocysts can survive exposure to sodium hypochlorite and chlorine. T. gondii oocysts were found in soil in 28 out of 34 studies, in water in 25 out of 40 studies, in fresh produce comprising leafy greens, non-leafy vegetables including roots, herbs, and fruits in 13 out of 23 studies and bivalve mollusks in 19 out of 22 studies. For soil and water some studies reported a 100% detection rate, while for fresh produce and bivalve mollusks the maximum detection rates were 46% and 50%, respectively. Only in 13 of the selected articles, the One Health concept was applied to establish links between soil, water and food contamination with infection cases. These mostly regarded North and South America [13].
A recent monitoring activity in Northern Italy showed that in the Emilia-Romagna region the prevalence of human toxoplasmosis was between 20.0% and 20.8%, higher than in other European countries. Acute infection was diagnosed in 161 patients, of whom 113 were pregnant women. During three years 22.3% of women tested positive for toxoplasmosis during early pregnancy with a higher likelihood of infection for women of African, Asian, Eastern European, and South American origin compared to Italian women. The frequence of active toxoplasmosis in this group was 0.39% [46].
In a cross-sectional study on pregnant women the overall seroprevalence of anti-T. gondii antibodies (IgG and/or IgM) was 21.2% and a significant association was found with raw vegetable consumption [41].
5.3. Distribution of T. gondii in Food Producing Animals
In a molecular epidemiological study carried out in Belgium, T. gondii could be isolated and detected by magnetic capture-qPCR in 14 out of 92 pig hearts collected during 2016 and 2017 from pigs raised in organic farms [164]. Mouse bioassay demonstrated the viability of T. gondii in 9 of the positive animals. Isolate genotyping with a multiplex PCR on microsatellite markers which allows to discriminate between the main T. gondii genotypes I, II, and III and different strains, showed that pig isolates belonged to type II, which is predominant in Europe [49], and genotypes were highly related to those of human isolates obtained during 2015 and 2016, leading to hypothesize either transmission of T. gondii from pigs to humans or that both species were infected via a common source of infection. These findings are in agreement with other studies carried out in France, Portugal, and Poland. In contrast, one study performed in North-Western Italy found genotype I as the most prevalent in different species. In Denmark, more than one-third of the isolates were attributed to genotypes Africa 1, HG12-like, and other atypical genotypes. Also in Portugal, 12.5% and 20.8% of isolates were identified as type I and mix/recombinant, respectively. In Poland also the circulation in humans of type III strains associated with wildlife was reported [164].
Particularly pork and lamb are a major source of T. gondii infection while tissue cysts are less common in poultry, rabbits, cattle and horses. T. gondii-infected pork is considered an important source of infection for humans in Europe and the United States. Regarding swine farming conventional finisher herds may be classified as low-risk, organic finisher herds as medium-risk, and conventional and organic sow herds as high-risk. Recently, the global T. gondii seroprevalence in pigs was reported to be 19%, with the lowest in Europe at (13%) and the highest in Africa and North America (25%). The seroprevalence in Asian and South American regions was (21%) and (23%), respectively [44].
Seroepidemiological studies reported that farmed and free-ranging wild boars are commonly exposed to T. gondii in many countries with a seroprevalence of 32% in North America, 26% in Europe, 13% in Asia, and 5% in South America.
In general, processed pork products pose a lower infection risk than fresh raw pork because salting, freezing at −12°C, hot smoking, long fermentation, heating above 67°C, irradiation and high hydrostatic pressure can reduce or inactivate T. gondii cysts in meat. Several studies confirmed the safety of ready-to-eat pork prepared using typical NaCl concentrations of 13% or higher, as well as industry standard fermentation and drying procedures [44].
In the Emilia Romagna region, Italy, a high prevalence of toxoplasmosis of 41.9% in sheep flocks and 14% among wild water birds hunted in the region was observed. Seroprevalence was 29.9% in sheeps, 18.7% in goats, 15.5% in wild boar and 25% in roe deer. Moreover, infected cats were widespread in the territory. The significance of toxoplasmosis in these animal species is of concern for public health since some of the animal species are used to produce raw gastronomic preparations. Its significance is also economic as it represents a primary cause of abortion in sheeps and goats [46].
Occurrence in wild animals in Southern Italy also indicated a wide distribution of the pathogen, with a prevalence determined by qPCR of 28.6% in wolves, 27.3% in badgers, 23.9% in foxes, 18.9% in wild boars, and 14.3% in roe deer [166]. In wild boars of Sardinia island, Italy, anti-Toxoplasma IgGs were detected in 24.6% of animals [51]. Out of 11 blue crabs, 6 were found positive for T. gondii and out of 55 tissues samples, 12.7% were positive for T. gondii with hemolymph and gills being the most infected tissues [174].
Raw milk can pose a risk of infection due to tachyzoites of T. gondii that can be shed in the milk of naturally infected animals. T. gondii DNA was detected in 8% ewes and one 4% goat raw milk samples, while none of the cheese samples were positive in Turkey [175].
6. Giardia Intestinalis
Giardia genus of flagellated protozoans of the phylum Fornicata, clade Metamonada [2] comprises the eight species G. agilis, G. ardeae, G. cricetidarum, G. microti, G. muris, G. peramelis, G. psittaci, associated with amphibians, herons, hamsters, voles and muskrats, rodents, in bandicoots, and budgerigars, respectively, and G. intestinalis, alternatively designated G. duodenalis or G. intestinalis, responsible for human infections. These organisms colonize the small intestine of humans and other mammals. G. intestinalis is divided into assemblages, or subtypes, A‒H, and sub-assemblages. Assemblages A and B have a wide host range and cause most infections in humans with the latter responsible for the majority of human cases. The remaining assemblages have narrow host ranges, namely, C and D are associated with dogs and other canids, E with livestock, F with cats, G with rodents, and H with seals. Metabarcoding at the glutamate dehydrogenase gdh locus showed that assemblage B comprises multiple variants [176]. Currently, the possibility that Giardia assemblages could represent distinct species is under consideration [15].
G. duodenalis Assemblage A can be subtyped into sub-assemblages AI, AII, and AIII by multilocus sequence typing (MLST), with mainly sub-assemblages AI and AII involved human and animal giardiasis worldwide, and sub-assemblage AIII more commonly infecting hoofed animals. Assemblage B has previously been subtyped into BIII, BIV but later a new typing system for Assemblage B was developed. However, BIII and BIV subtypes must still be referred to since they have been amply reported in the literature [103]. In addition, an assemblage A-specific multilocus sequence typing (MLST) method based on six genetic loci has provided increased levels of polymorphism facilitating source tracking in foodborne outbreaks [177].
G. intestinalis assemblages A and B were reported in 58 and 57 out of the 68 subtyping studies in the MENA region, respectively, and 41 studies reported co-infections with both. Moreover, in human infections assemblage C, commonly found in dogs, was detected in one study in Egypt and assemblage E, commonly found in livestock, in three studies in Egypt and in Iraq. Assemblage F which usually infects cats was reported in two studies in Iraq. Subtyping of G. duodenalis identified sub-assemblages AI, AII, BIII, BIV and a novel sub-assemblage B in Iran. G. duodenalis sub-assemblage AII was reported in drinking water in Egypt [103]. Sub-assemblages AII and BIV were detected both in food handlers and in students in public schools in Angulo, Brazil, and sub-assemblage BIII caused an outbreak in Long Island, New York, USA transmitted by infected food handlers in a grocery store chain. G. intestinalis assemblages A, B, C and D were reported in the hemolymph of shellfish [177].
G. intestinalis has a simple life cycle comprising the trophozoite, which causes the symptoms, and the cyst, which represents the infective form shed with the stools of the host [177]. The pathogenic mechanism of Giardia spp. includes disruption of the mucus layer and the gastrointestinal epithelium [176,179].
Characteristics of G. duodenalis that favor infection are the low infective dose, of 10 to 100 cysts in humans with even a single cyst having a 2% probability to cause giardiasis, the large number of cysts that can be shed by a single individual, reported to be 2.5 × 107 Giardia cysts annually in a study in the Netherlands, contribute to the high prevalence of giardiasis [180]. High-level shedding by a wide range of animal hosts, including insects, determines high environmental contamination. Moreover, a high survival ability of cysts from weeks to months was reported in drinking water and food. The small size of Giardia cysts, 8–12 μm in length, allows them to penetrate and survive in water filters such as sand filters. In addition, their ability to survive at low temperatures determines their persistence in refrigerated conditions [177].
6.1. Diseases Caused by G. intestinalis
Symptoms of giardiasis include diarrhea, abdominal bloating and cramps, malabsorption and weight loss. Some individuals can experience chronic and sometimes severe disease that is unresponsive to treatment [177]. If left untreated the infection can become chronic and asymptomatic over time. Chronic infection is associated with malabsorption which causes growth retardation, weight loss, iron deficiency, anemia, and sometimes cognitive impairment in children [179]. In the GEMS study, Giardia spp. was more frequently detected among children older than two years and it was associated with growth deficit [17]. Chronic Giardia infections are also associated with food allergies, irritable bowel syndrome (IBS), chronic fatigue syndrome and arthritis [177,181]. G. intestinalis was implicated in stunting at 2 years of age and presented two peaks of morbidity, at ages 1-9 years and 45-49 years. Long-term sequelae include cognitive deficits [182].
Two cohort studies showed that, beyond Clostridium difficile, Helicobacter pylori, Norovirus and Candida, which are implicated in the initiation and exacerbation of the IBS syndrome, also G. intestinalis is significantly associated with IBS incidence and to an increased risk of IBS, though not significantly [183].
The GEMS reported that Giardia was not associated with severe diarrhea [184]. On the other hand, volunteer cyst challenge studies showed that G. duodenalis is capable of causing diarrhea in immunocompetent adults and a meta-analysis demonstrated that giardiasis offered protection from acute diarrhea but increased the risk of persistent diarrhea in children [177].
Symptoms of giardiasis described in a recently reported case in a 59-year-old man with alcoholic cirrhosis, chronic hepatitis C, and hepatocellular carcinoma were watery diarrhea lasting for 2 weeks and rectum bleeding. The infection was accidentally discovered in duodenal biopsies carried out to exclude celiachy. Symptoms resolved spontaneously [185]. G. intestinalis infection was discovered accidentally also in a 67-year-old patient with pancreatic cancer in the histological section. The association of G. intestinalis infection and pancreatic or gallbladder cancer was reported also previously [186].
Treatment of giardiasis is carried out with nitroheterocyclic antiparasitic drugs including the 5-nitroimidazoles secnidazole and tinidazole whose activity depends on the reduction by ferredoxin or flavodoxin in organisms lacking mitochondria like G. duodenalis [179,187]. The emergence of resistance mediated by epigenetic or posttranslational modifications led to the evaluation of alternative substances for the treatment of giardiasis, including different nitroheterocyclic substances, benzimidazoles, nitazoxanide, quinacrine anti-malarian drug, aminoglycosides hygromycin and paromomycin, ciprofloxacin, bacitracin, anti-viral protease inhibitors, anti-rheumatic and anti-tumoral substances, molecule combinations and hybrid molecules. Omeprazole, which inhibits the glycolytic enzyme triosephosphate isomerase of G. intestinalis leading the organism to death, had effects in vitro [187].
6.2. Epidemiology of Giardiasis
G. intestinalis is ranked 11th In the FAO/WHO risk management list of intestinal parasites [19], is responsible for approximately 180 million symptomatic infections annually and several studies indicated that its prevalence in the stool of asymptomatic children was between 17% and 18% in Spain and between 18% and 64% in subtropical countries such as Brazil, Ethiopia, Argentina, and Mozambique. Asymptomatic colonization occurs also in animals and a higher prevalence was observed in individuals with animal contact regardless of the type of animal [15].
Between 2011 and 2017 over 140 waterborne outbreaks of giardiasis occurred globally, while foodborne transmission occurred in approximately 23.2 million cases annually, but these could be underestimated since the detection of G. intestinalis in food is inefficient and reporting and surveillance are carried out mainly in countries in which giardiasis is notifiable, namely the USA, European Union (EU) [188], some Australian states and New Zealand [176,177].
Giardiasis was the most frequently reported food- and waterborne parasitic disease in the EU according to the report on giardiasis of the European Centre of Disease and Prevention (ECDC) for 2019, with 5.2 cases per 100,000 inhabitants, highest for Belgium and Bulgaria in the age group 0–4 years [189].
Studies in single countries reported that G. intestinalis was the predominant parasite, with a 37% prevalence, in marginalized rural areas in the North of Palestinian West Bank Region [190], had an estimated prevalence in Colombia between 0.9 and 48.1% when analyzed with classical microscopy and between 4.2 and 87% when analyzed by qPCR and nested PCR. Prevalence of G. intestinalis among patients with 5-8 years of age treated in health centers in Sheno, Ethiopia, was 53.3% in 2020 and 56% in 2021 [191].
In Côte d’Ivoire, quantitative microbial risk assessment (QMRA), which estimates the risk of infection from exposure to a microorganism, assessed an annual risk of infection with Giardia of 0.36 and a probability of becoming ill from eating vegetables grown locally of 1.0%. In many countries, the risk of giardiasis from eating vegetables irrigated with wastewater was higher than acceptable, with a maximum of 100% in Thailand. An “adjusted likelihood ratio” statistical tool that examines the association between outbreak cases and food distribution was developed to improve the identification of food products that should be analyzed for Giardia cysts [177].
Prevalence of G. intestinalis in asymptomatic school children was 10% in Lusaka, Zambia [192], and 52% in Zambézia province, Mozambique [193], where one of the associated risk factors was drinking untreated river/spring water [194].
G. intestinalis accounted for 13.8% of intestinal infections in immunocompromised patients with intestinal illnesses in Yemen, a country with limited access to water resources [161].
According to a meta-analysis of cases reported globally between 1977 and 2016, impaired immunity was associated with giardiasis, while breastfeeding was a protective factor in African countries, possibly for the inhibitory activity of unsaturated fatty acids and of specific secretory IgA antibodies present in human milk in areas of endemicity. Breastfeeding also protects from exposure to contaminated milk. Not washing hands after the toilet and before eating was significantly associated with giardiasis in children mostly, while not washing hands with soap was associated with giardiasis in the general population in Asia, Africa, and South America. Consumption of fresh produce was identified as a risk factor in children and consumption of composite ready-to-eat foods represented a highly significant risk factor in Canada. The consumption of unwashed vegetables increased the risk of giardiasis in Cuba and Malaysia [182].
The results of a recent meta-analysis showed significant positive associations between intestinal Giardia spp. infection and lack of safe drinking water, no access to the toilet and age, and a positive association, though not significant, between intestinal Giardia spp infection risk and no hand-wash. The results of meta-regression showed that Giardia spp. infection risk decreased significantly over time [179].
6.3. G. intestinalis Infections from Food and Drinking Water
Foodborne transmission of giardiasis has become more widely recognized in recent years and it was reported that in the USA 7–15% of Giardia infections were probably foodborne [177]. Foodborne outbreaks of giardiasis reported from 1971 to 2011 were almost all from the USA, with a total of 38 outbreaks [195]. The type of food involved was fresh produce, including salads, raw vegetables and fresh fruits, but also canned salmon, raw oysters, ice cream, and processed foods such as noodle salad, chicken salad, dairy products, sandwiches and tripe soup. In the USA some outbreaks until 2018 were caused also by unpasteurized milk, shellfish and not identified foods served in restaurants or communities such as offices, schools and camps. In twelve cases infected food handlers were the cause of food contamination and G. duodenalis was frequently identified in fecal samples and under their nails [177].
A study in an urban area of southern Brazil, which screened humans, animals, water samples and vegetables, identified the same genotype of assemblage B, with 100% gdh gene sequence similarity, in humans, one dog and two lettuce samples, suggesting a linkage with irrigation water possibly contaminated from septic tanks. In another study, whole genome sequencing (WGS) showed that Giardia from beavers was the cause of two waterborne outbreaks in a small community [177].
Waterborne outbreaks of giardiasis in Europe were mainly reported by Nordic countries, including a large outbreak in Bergen, Norway, which involved around 6,000 cases in 2004. In January 2019 14 cases of giardiasis, were diagnosed in a single day, in Bologna, Italy. The outbreak involved more than 200 cases and lasted for several months. The first case interviews indicated that consumption of tap water was most likely the source of infection. However, Giardia cysts were not detected in water samples collected from the distribution network during the outbreak and the additional monitoring period. It was hypothesized that water contamination occurred during working operations carried out on the water supply network before the outbreak and was no longer detectable [178].
The pooled prevalence estimate of Giardia spp. contamination from waterbodies in Africa was 11.9% from a total of 7950 samples that were detected using microscopy, with the highest infection rate of 37.3% in Tunisia [196].
Giardia cysts have been detected on fresh produce, including leafy greens, herbs, berries, fruits, green onions, carrots and tomatoes, dairy products, meat, shellfish and processed foods [177,195]. A few studies examined washed and ready-to-eat vegetables and salads at supermarkets and percentages of contamination in ready-to-eat salads ranged between 0.5% observed in Palermo, Italy, and 1.8% in Canada. On unwashed vegetables the highest percentage was 55% in Ilam City, Iran, followed by field-collected vegetables in Valencia, Spain (52.6%). Generally, the numbers of cysts recovered from fruit and vegetables were low but the real contamination rates were probably higher since recoveries of Giardia cysts from food can be low. Washing fruit and vegetables can reduce the risk of contamination but in a study in Nepal it was reported that even when treated water was used for washing vegetables, the contamination levels were higher than the infectious dose of 10 cysts [177]. Contamination of fresh produce may occur for use of contaminated fertilizer and access to crops by livestock and other animals or directly from the hands of infected workers or equipment [195].
The detection of Giardia in food was improved by the use of immunomagnetic separation (IMS) methods, before DNA extraction and PCR targeted on widely used loci such as genes for the glutamate dehydrogenase (gdh), triose phosphate isomerase (tpi), beta-giardin (bg) and 18S rRNA, to isolate and elute cysts, from the food sample and a standardized method for the detection and enumeration of Giardia cysts on berry fruits and fresh leafy green vegetables based on IMS became available in 2016. However, the cost of IMS beads limits the implementation of this method in developing countries [177].
7. Trypanosoma cruzi
Trypanosoma cruzi is a flagellated protozoan of the clade Discoba [2], that causes Chagas disease (CD), endemic in Latin America an infection that can be severe and lead to disability and death. Among the mechanisms of transmission, the oral route is the most important for the maintenance of the zoonotic cycle of the parasite. Confirmed human cases of transmission through food were reported in Brazil, Bolivia, Ecuador, Venezuela, Colombia, Argentina and French Guiana and were mostly attributed to unpasteurized fruit juices, fruits, other food and water infected with feces of triatomine (Triatomine subfamily, Hemiptera, Reduviidae) vectors of the parasite or with secretions of animal reservoirs [197]. The infection can be acquired also by the ingestion of raw meat of infected animals containing metacyclic trypomastigotes. These are the infective forms of the parasite and in oral infection enter the bloodstream through the mucosal membranes after binding to the gastric epithelium, a process modulated by glycoproteins gp82 and gp90. The metacyclic trypomastigotes are covered with mucin-like molecules that make them highly resistant to proteolysis in the gastric tract [42].
In the host cell, the trypomastigotes transform into proliferative forms, the amastigotes, that multiply by binary division. These burst the host cells, infecting other cells and entering the bloodstream thus reaching many organs. In the oral infection, the intensity of the inflammatory response depends on the parasitic load and determines mild or severe gastrointestinal manifestations [42].
The genera Triatoma, Rhodnius and Panstrongylus are the most epidemiologically relevant vectors of T. cruzi [11].
7.1. Epidemiology and Symptoms of T. cruzi Infection
T. cruzi was rated tenth in the FAO/WHO list of most important foodborne parasites [19].
In Brazil between 2012 and 2018 1340 oral cases of CD were reported of which 1172 in the State of Pará, the largest producer and consumer of açai that is frequently contaminated by T. cruzi. Acute CD cases were attributed for 68.4% to oral transmission and were characterized by a highly severe acute myocarditis with an 8-35% lethality rate [42,198]. Another cause of death following T. cruzi infection can be meningoencephalitis [197]. Beyond acai (Euterpe oleracea) also sugarcane (Saccharum spp.) in Brazil, guanabana or soursop (Annona muricata) and mandarin orange (Citrus reticulata) in Colombia, guava (Psidium guajava) in Venezuela, majo or patawa (Oenocarpus bataua) in Bolivia and milpesillo or bacaba (Oenocarpus bacaba) in French Guiana can be contaminated [11].
In the Americas, CD shows an annual average incidence of 30,000 new cases, 12,000 deaths per year, and approximately 9000 newborns become infected during gestation. It is estimated that around 70 million people in the Americas are exposed to the risk of contracting CD [11].
CD has spread to other regions in the world in the last 2 decades, affecting immigrants in Europe and in the USA. A meta-analysis summarized 2470 orally-transmitted acute CD cases that resulted in 97 deaths all occurred in Latin America, specifically in Venezuela, Colombia, Bolivia, French Guiana, and Brazil. The food source was unreported in many studies, but açaí was the most common, followed by sugar cane juice, palm fruit, majo fruit, mango juice, orange juice, and other juices. Through meta-regression, it was found that the lethality from orally acquired CD displayed a statistically significant reduction over the years. This may reflect increased awareness, more rapid diagnosis, and prompt institution of appropriate therapy [197].
A recent outbreak occurred in Southwest Amazonas in 2021 and involved 27 individuals who consumed açaí collected and prepared by a single person in a house with a palm leaf roof, and surrounded by palm trees, which are typical habitats for triatomines. All exposed individuals, regardless of the presence of symptoms, underwent diagnostic procedures and five individuals had a confirmed diagnosis of CD. All symptomatic patients had prolonged fever. Other symptoms varied among patients and were lower limb and face edema, headache, abdominal pain, vomiting, back pain, polyarthralgia, asthenia, exanthema, retro-orbital pain, generalized rash and itching. Symptoms appeared after a few days from ingestion of the juice. None of the patients had severe manifestations, such as acute myocarditis or digestive hemorrhage. However, the youngest patient, a 10-year-old boy, had an allergic reaction to the drug benznidazole, used to treat CD and needed a longer treatment along with antiallergic drugs. As in this outbreak, inadequate hygiene in food preparation at home caused CD in many other instances [199].
An acute CD case occurred in the municipality of Santarem, Para State, Brazil, in a 22-months-old infant, after the oral ingestion of bacaba palm fruit’s wine. The infant was admitted in a fair general condition, with bipalpebral edema and anasarca. The parents reported that she had had fever, vomiting, diarrhea and upper abdominal pain thirteen days before the hospital admission. She was the first patient detected in an outbreak with more than eight infected individuals. On day 24, treatment was initiated with orally administrated benznidazole and no side effects were noticed. She was discharged asymptomatic and with a negative blood smear and a prescribed 60-day treatment with benznidazole, but the patient suddenly died three days later for a not ascertained reason. Fatality due to stroke and upper gastrointestinal bleeding has been described in CD and delay in treatment, low platelets and altered liver enzymes with a very high parasitemia might have all contributed to the fatal evolution in this case [200].
T. cruzi is classified into at least seven discrete typing units (DTUs) TcI to TcVI and and Tcbat. TcI is the most common DTU found in oral outbreaks and the one associated with severe cardiopathy [42], while TcII, TcV and TcVI, also associated with cardiopathy, give milder or no symptomatology. DTUs related to oral outbreaks in Brazil, Colombia, Venezuela, Bolivia, and French Guiana were TcI, TcV, TcIII, and TcIV. TcI presents an extensive genetic diversity and is divided, according to the transmission route, into domestic (TcIDom) and sylvatic (TcISylv) genotypes. Methods for DTU distinction are specific PCR on genetic markers, including Spliced-leader intergenic region (SL-IR), microsatellites, kinetoplast DNA (kDNA), heat shock proteins (HSP), 18S rRNA gene, cytochrome c oxidase subunit 2 (COII), cytochrome b (Cytb), Glycosylphosphatidylinositol (GPI) and 24Sα rDNA/rDNA subunits (24Sα). Older PCR-RFLP methods are still in use. A map of DTU distribution in different countries of parasites isolated from clinical samples, insects and food is available [201].
In the acute phase, when the parasitemia is elevated, trypomastigotes can be visualized in peripheral blood by direct microscopy, while in the chronic phase, with low parasitemia, serological tests to detect IgG antibodies against the antigens of T. cruzi by ELISA (Enzyme-linked immunosorbent Assay), IFA (Indirect Immunofluorescence Assay) and HAI (Hemagglutination Inhibition Test). Molecular methods such as qPCR are suitable for molecular diagnosis both for the acute and chronic phase. For oral cases of CD one of the most important diagnostic phase is the epidemiological inquiry which must consider travel history, raw-food consumption, and geographical location [42].
T. cruzi cannot multiply out of its host but can survive at room temperature for 10 h on melon and tomato, 18 h on papaya and banana, 24 h on apple and 48 h on potato and carrot. T. cruzi can survive in açai juice for 144 h at 4 °C, for 26 h at −20 °C, and for 24 h at room temperature. Appropriate prevention technologies to eliminate the possibility of the ingestion of the infectious parasites include inactivation by blanching fruits at 70 /80± 1 °C for 10 s or pasteurizing juice at 82.5 °C for 1 min. Triatomine contact with food can be prevented with the implementation of good manufacturing practices (GMP) and hazard analysis critical control points (HACCP) [200,202]. These rules are recommended by Brazilian regulations but their observance is not surveilled [200].
T. cruzi detection in food matrices is important for the identification of potentially infectious food and for the implementation of prevention procedures. Traditional detection methods, such as culture and microscopic observation are difficult, labor and time-intensive and not enough sensitive so these were replaced by molecular techniques [202].
8. Cyclospora Cayetanensis
Cyclospora is a cyst-forming protozoan belonging to the clade Sar Alveolata, phylum Apicomplexa, family Eimeriidae [2]. Currently, nineteen Cyclospora species have been reported to cause disease in various animals but only C. cayetanensis was associated with acute and chronic diarrhea in humans [6,203]. However, a recent study on the genotyping of thousands Cyclospora isolates from the USA and one from China based on sequencing of two mitochondrial and six nuclear genetic loci reported that at least three genetic lineages of Cyclospora were responsible for human cyclosporiasis; lineages A and B causing outbreaks in North America and lineage C from Henan Province, China. Epidemiological data demonstrated the associations between lineage and the geographical and temporal distribution in USA infections. The authors proposed to consider lineage A as C. cayetanensis species and to classify lineage B as the novel species C. ashfordi and lineage C as the novel species C. henanensis [204].
The life cycle of C. cayetanensis starts with the release of non-infective oocysts with feces by infected individuals. These sporulate in the environment after days or weeks at 22 °C to 32 °C. The sporulated oocysts contain two ovoid sporocysts, each containing two sporozoites, that are released in the gastrointestinal tract when ingested [72,203]. These invade the epithelial cells of the small intestine undergoing asexual multiplication into Type I and Type II meronts. Merozoites from Type I meronts perpetuate the asexual cycle and autoinfection of the host, while merozoites from Type II meronts undergo sexual development into macrogametocytes and microgametocytes upon invasion of another host cell. These form a zygote that develops into an oocyst which is released from the host cell and is shed in stools [203]. There is still little information on the infective dose and the conditions that favor oocyst persistence and sporulation, the role of water and soil in transmission, and the existence of reservoirs [72,203].
8.1. Diseases Caused by C. cayetanensis
Enterocyte invasion causes damage to the epithelium of the small intestine with disruption of the brush border, loss of membrane-bound digestive enzymes, and atrophy of villi. As a consequence, the uptake of water, nutrients, and electrolytes in the small intestine decreases determining symptoms such as watery diarrhea, abdominal cramps, bloating, nausea, fatigue, low-grade fever, anorexia and weight loss [72,203]. Prolonged diarrhea may cause death in rare cases involving infants and individuals with co-morbidities [203]. Infection is often mild or asymptomatic in residents of countries in which C. cayetanensis is endemic, while a more severe disease occurs in infants, the elderly, and immunocompromised patients, such as those with HIV/AIDS, and travelers from non-endemic countries [72,203].
Late identification of etiology of C. cayetanensis diarrhea can lead to complications such as Acute Kidney Injury (AKI), which can turn into Chronic Kidney Disease (CKD). A recent case of Acute Kidney Injury complication involved a 35-year-old Mexican male with a kidney transplant who was most probably infected after eating fresh vegetables in a Dutch restaurant during a trip [205].
The first case of C. cayetanensis infection reported in India though this parasite is endemic in the Indian subcontinent, was a 30-year-old woman in Kolkata in 2022. She was in complete clinical remission from Hodgkin’s Lymphoma and was affected by diarrhea for two months unfruitfully treated with quinolones. Later C. cayetanensis was isolated from stool and identified by PCR and DNA sequencing, so she was treated with Tab Nitazoxanide and recovered. The source of infection was suspected to be contaminated water or fruits and fresh vegetables available locally [72].
A 76-year-old man with diffuse large B cell lymphoma in San Sebastian, Spain, was affected by diarrhea for 4 weeks and treated with ciprofloxacin for one month without success. A multiplex one-step real-time PCR on stool samples was positive for Campylobacter jejuni and C. cayetanensis but only C. jejuni infection was initially treated with azithromycin. After initial improvement the patient relapsed and, at this time, only C. cayetanensis was detected in stools, so treatment with TMP–STX led to the improvement of diarrhea symptoms [68].
TMP–STX treatment is effective against cyclosporiasis and in immunocompromised patients, such as those with AIDS and transplant recipients, it should be administered for longer or at higher doses, and as prophylaxis for relapse prevention [203]. A retrospective review in Florida, USA, regarding years 2012 – 2022, showed that, beyond Cryptosporidium (18 cases) also Cyclospora spp. (4 cases) can infect cancer patients [206].
Infection prevention measures that should be followed especially by immunocompromised patients, include avoiding eating uncooked raw vegetables and unpeeled fruits, eating fully cooked foods since C. cayetanensis oocysts are inactivated by cooking and avoiding to consume untreated well water and surface water [203].
8.2. Epidemiology of C. cayetanensis
Latin America, Central and Southeast Asia, the Middle East, and North Africa are hot spots of endemicity for C. cayetanensis and there it caused several outbreaks linked to the consumption of fresh fruits, berries, leafy greens and aromatic herbs. Since Cyclospora is highly resistant to disinfectants used in the food industry, the contamination of fresh vegetables and fruits persists even after sanitation [6].
In the USA outbreaks of cyclosporiasis have been documented since the 1990s and have affected thousands of individuals annually in the past decade. Outbreaks were linked to berries, cilantro, basil, and, more recently, ready-to-eat bagged salads imported or grown in the USA [207]. The first reported outbreak of cyclosporiasis in the USA was in Chicago and was possibly linked to a hospital water supply [208]. From 2018 to 2022 annual summertime outbreaks became regular in people who had no recent history of international travel and laboratory-confirmed cases of cyclosporiasis were linked to both imported and locally grown fresh produce [203,208]. Outbreaks occurred between 1 May and 31 August 2021, with 1123 laboratory-confirmed cases in 36 states and 561 isolates were distinguished in 31 temporal genetic clusters (TGC) and 10 of these were associated with epidemiological clusters. Two epidemiological clusters were multistate and by isolate typing and patient interviews allowed to attribute one of these clusters to lettuce from a single brand, and the other to a lettuce type not associated to a brand [209].
In a case–control study of an outbreak in Canada, with a total of 87 cases and 1 hospitalization, blackberries imported from Mexico were identified as the most likely source of infection based on case exposure. In South Korea an outbreak of cyclosporiasis reported among travelers returning from Nepal was the first in the country and was probably caused by infected vegetables, water, or fruits from the visited country [207].
In Europe and Australia, most of the reported cases of cyclosporiasis were linked to international travel to endemic areas. A foodborne outbreak in Stockholm, Sweden, was linked to sugar snap peas imported from Guatemala based on information obtained from laboratory-confirmed C. cayetanensis infected patients. In Europe, the sources of a foodborne outbreak in Germany were epidemiologically traced to lettuce and herbs from Germany, France, and Italy; the contamination of food crops probably derived from agricultural workers who had no access to sanitary facilities [208]. The long incubation period of up to two weeks and intermittent remissions confound case recognition and make it difficult to identify the infection source in the outbreaks [72].
The prevalence of C. cayetanensis among populations reported in different studies varied greatly. In China, it was 0.2% among patients with diarrheal illnesses in Zhenjiang City, Shanghai Municipality, and 10.0% in Yongfu country, also in Southeast China, among workers in a laboratory animal facility. In children younger than 5 years of age in African countries prevalence of C. cayetanensis was 3.3% in Gabon, 2.7%, in Ghana and 0.9% - 1.5% In Tanzania. In Iraq, prevalence of C. cayetanensis varied between 1.0% in a hospital in Baqdad and 14.5% in rural areas, and a high rate of infection was observed in children 1–9 years of age (25.8%) and in the age group 10–29 years (21.8% - 16.6%), while it was comprised between 1.0 and 3.0% in older persons. In Thailand, C. cayetanensis was not detected by molecular methods in 254 schoolchildren in Ratchaburi Province [207].
A significantly higher prevalence of 8.7% was reported in Ghana in HIV-positive individuals compared to 1.2% in HIV-negative individuals. The prevalence reached 13.6% in patients with CD4+ T cell counts below 200 cells/µL. A pooled prevalence of C. cayetanensis infection of 3.9% was estimated in HIV-infected persons in North Western Iran [210]. In Sweden, in patients with gastroenteritis, the parasite was only detected in 2 among 803 fecal samples by microscopy and real-time PCR. The incidence and suspected origin of those infections were unknown [211].
Beyond being associated with poverty as a social factor, cyclosporiasis was associated with soil transmission in two Venezuelan communities [212].
8.3. Dietary Sources of C. cayetanensis
Contamination by C. cayetanensis was recently observed in imported blueberries in Italy [213], in imported and locally produced raspberry, blueberry, and strawberry in Norway, with 8.7%, 5.5%, and 4.8% occurrence, respectively, in central Mexico, in blueberry and blackberry from commercial farms in 16.6% and 27.3% samples, respectively, and in one strawberry sample among 120 analyzed in Bogota, Colombia [207,214].
C. cayetanensis was detected in 0.3% samples of imported leafy greens, herbs, and berry samples in the USA and [208] in 0.28% of samples tested in Canada, of which spinach, cilantro, and blueberries were grown in the USA [208,215].
In Iraq C. cayetanensis was detected in 3.7% of fresh produce in Baghdad Province and in washing water from at least one sample of garden cress, radish, leek, green onions, and purslane with a number of oocysts per liter of 6, 7.8, 7.2, 4.4, and 3.2, respectively in Anbar province. In China, C. cayetanensis was only identified in two vegetable samples, 0.2% of those analyzed, isolates showed identity of the 18S ribosomal RNA gene with a C. cayetanensis human isolate from Shanghai. In a dry area of Northwest Mexico Cyclospora spp. was observed in 3% of samples of fresh asparagus and in 1.0%, 5.0%, 8.0%, and 30.0% of melon, peach, asparagus, and grape samples with no differences in the percentage of positive samples between those collected in open-air markets or closed markets. In Venezuela, 3 out of 77 samples of fresh produce collected at local markets were positive for Cyclospora species [207].
In Southern Ethiopia, Cyclospora spp. was detected in 4, 2, and 1 out of the 100, 66, and 23 tomatoes, green peppers, and salad samples, respectively, and in eight types of fruits and vegetables (lettuce, cabbage, carrot, tomato, green pepper, banana, orange, and spinach) obtained from local markets in Dire Dawa City. In Egypt, Cyclospora spp. showed a prevalence rate of 2.9% in vegetables with positive samples of watercress, parsley, radish and green onion, but not lettuce or coriander. A higher detection rate was reported for El-Kharga Oasis, Upper Egypt, with 20% positive samples of arugula and radish in public markets. In Korean fresh-cut fruit products at retail no Cyclospora-positives were found by real-time PCR [207].
Cyclospora cayetanensis was found to occur in marine crustaceans and mollusks such as bivalve shellfish and wild clams in Tunisia, Egypt, and Turkey [208], mussels (Mytilus galloprovincialis) near the west coast of Turkey and blue crabs (Callinectes sapidus) from Lesina lake, Italy [174], thus showing its occurrence in the marine ecosystem.
Cyclospora spp. in drinking water was reported in a major watershed in the Philippines after rainfalls and in a watershed in Bamenda, Cameroon, among other protozoan parasites, in a concentration of 141.31 ± 143.19 oocysts/l in the rainy season and 471.42 ± 216.32 oocysts/l in the dry season. In Turkey, Cyclospora was found in agricultural irrigation water samples collected in Denizli City Center but not in drinking water [207].
In Italy, 30% (3/10) of train tap water samples were C. cayetanensis-positive, representing a cause of concern. In Apulia, Italy, the parasite was identified in treated wastewater (21.3%), well water (6.2%), soil (11.8%), and vegetables (12.2%) [208].
The difficulty of detecting the parasite in fresh produce and environmental samples is determined by the low numbers of oocysts present. Therefore, molecular techniques, including quantitative PCRs (qPCR), were recently developed for their detection in these samples and water [208,216] to improve outbreak investigation and prevention. The genotyping system based on eight genetic markers applied to human clinical samples was not yet successfully employed for food samples. However, a workflow was recently developed and successfully applied to obtain complete mitochondrial genome sequences from produce samples spiked with low numbers of C. cayetanensis [208].
9. Balantioides coli
Balantioides coli [2], clade Sar Alveolata, phylum Ciliophora, the only ciliated protozoan and the largest among those infecting humans [218], causes balantidiasis, a neglected parasitic infection of zoonotic significance and worldwide distribution, but most frequent in subtropical and tropical regions [217].
B. coli is considered a commensal of the intestine in several mammalian hosts and the main reservoirs are domestic, wild pigs and wild boars, which carry the parasite in the large intestine. Its life cycle comprises the two stages of cyst and trophozoite [217,218].
The prevalence of B. coli in human hosts was reported to be 0.02% - 1% but real values may be underestimated since it usually remains asymptomatic [218]. The highest prevalences were reported in Latin America, the Philippines, Papua New Guinea, and the Middle East, and 29% of reports regarded pig farmers. Transmission may occur by the ingestion of food and water contaminated with cysts, beyond contact with animals. Predisposing factors for illness manifestation are malnutrition, concomitant infections, and debilitating diseases [217]. Symptomatic cases are characterized by loose feces or watery diarrhea, loss of appetite and body condition, dehydration, and retarded growth. Severe cases are characterized by intestinal mucosa ulceration and bloody diarrhea with possible fatal outcomes and also liver abscess, vertebral osteomyelitis and myelopathy [217,218]. The parasite may cause extra intestinal infections through dissemination into the appendix, liver, peritoneum, lung, and genitourinary tract especially in immunocompromised individuals. Uterine infection, vaginitis, and cystitis may occur through direct spread from the anal area or rectovaginal fistula [218].
A case of urinary tract infection caused by B. coli was casually discovered during routine analysis for preeclampsia and preterm delivery in a 24-year-old pregnant woman in North West Ethiopia. The patient might have contracted the infection by contact with goats, sheep and other domestic animals or by eating raw vegetables and it was probably facilitated by immune suppression in pregnancy [218].
B. coli is frequently found in pigs and in a recent study carried out in Southern Italy, among 177 samples from swines reared and slaughtered locally, 46.89% tested positive for B. coli with an infection rate significantly higher in commercial hybrid races (64.84%) compared to the autochthonous breed (27.91%) and significantly higher in pigs reared in intensive system, possibly for the increased difficulty to maintain hygienic conditions. The data underlined that B. coli is widespread in the area considered as previously found also in other national and European areas. However, this parasitosis has been poorly considered with respect to public health significance, despite its high prevalence in animals and risk of zoonotic transmission. Since B. coli can survive on carcass surfaces, good slaughtering hygiene practices are essential to prevent its spread [217].
10. Cystoisospora belli
Cystoisospora belli, is an obligate intracellular parasite of the family Sarcocystidae, phylum Apicomplexa [2] that occurs globally, but is more common in tropical and subtropical regions and is most frequently transmitted by contaminated food and water. The life cycle of this parasite is similar to that of Cryptosporidium spp. and its oocysts can survive for months in the environment [66].
10.1. Diseases Caused by C. belli
C. belli infection in immunocompetent hosts can be asymptomatic or cause self-limiting watery diarrhea but it causes prolonged diarrhea or chronic diarrhea with relapses in immunocompromised patients, such as those infected with HIV, sometimes associated with headache, fever, malaise, abdominal pain, vomiting, dehydration, and weight loss. Moreover, in these patients, it can cause extra-intestinal complications with monozoic tissue cyst-like stages involving the liver, spleen, gallbladder, biliary tract, trachea-bronchial tract, and lymph nodes. This parasite can also infect epithelial cells of bile ducts and gallbladder, especially in severely immunocompromised patients. Other patients at risk are transplant recipients under immunosuppressive therapy [36,65,66,71].
Often, patients infected with C. belli are asymptomatic carriers before contracting HIV and become symptomatic once infected with HIV. Symptoms can occur in HIV-positive patients regardless of CD4 count, suggesting the absence of a protective CD4 count threshold [35]. Moreover, cystoisosporiasis recurrences are increasingly observed in HIV/AIDS patients and its chronicity could be related to the reactivation of the tissue cysts formed in extra-intestinal organs or the lamina propria of the intestine. Diagnosis of this parasitic infection is made difficult by intermittent shedding in stools. There is no reported serological test at present [66].
Infections and even fatal cases have been sporadically reported in immunocompromised patients, including those with Hodgkin’s and non-Hodgkin’s lymphoma, and acute lymphoblastic leukemia. Infections with C. belli have were reported also in association with human T-cell leukemia virus type 1 (HTLV-1) [65].
Recent cases in immunocompromised patients regarded a 66-year-old male patient in İstanbul, Turkey, with myeloma relapse and severe and persistent watery diarrhea who was treated empirically with metronidazole and ciprofloxacin before C. belli oocysts detection in the duodenum [36], and 6 HIV infected patients including a 25-year-old kidney transplanted male patient with deteriorated renal function who experienced alternating diarrhea and constipation for one year [66], a 45-year-old Nigerian woman with disseminated Kaposi sarcoma, watery diarrhea and gastrointestinal bleeding [19], a 57-year-old man in Antioquia, Colombia, who experienced a four-month history of diarrhea, other intestinal symptoms and weight loss [35], a 34-year-old woman with also chronic hepatitis B, latent tuberculosis, and Helicobacter pylori infection hospitalized in Mayo Clinic for abdominal pain, nausea, vomiting, and diarrhea lasting for one year, a 61-year-old male, with a very low CD4 cell count, with watery diarrhea for one-month and a dramatic weight loss who presented multiple erosions and ulcers in colon and C. oocysts belli detected in stools [220], and a 27-year-old male patient in Cameroon who contracted persistent diarrhea in Morocco and required hemodynamic support in a intensive care unit (ICU) and presented C. belli oocysts in the duodenum [221].
In all mentioned cases, C. belli infection was successfully treated with TMP-SMX [35,36,65,66,220,221] which should be administered as a long-term maintenance therapy in HIV patients [20].
Misdiagnoses were also reported for this parasite, as in the case of ovoid perinuclear cytoplasmic structures composed of dense fibrillar aggregates in the gallbladder of pediatric patients [222] and inclusions formed by aggregates of degenerated cytoplasmic components in the gallbladder in immunocompetent individuals, reported as Cystoisospora infection in a large series of studies, but not confirmed to be infectious by molecular and ultrastructural analysis [223].
10.2. Epidemiology of C. belli
In a study investigating the prevalence of C. belli in resected human gallbladders in a large cohort, the parasite was identified in 9.7% of specimens in a retrospective analysis and 27.3% of specimens in a prospective analysis and this organism appeared more prevalent among immunocompetent humans than previously recognized. The unexpectedly high prevalence in gallbladder specimens indicates that C. belli might be conceived as a commensal organism with a latent presence in the gallbladder, only giving rise to symptoms in case of pronounced immunodeficiency or immunosuppression [21]. Prevalence rates in HIV-infected patients range between 0.4% and 28% [66].
In a cross-sectional study, involving 156 children suffering from diarrhea or abdominal discomfort, in Sulaimaniyah, Iraq, from July to August 2021 C. belli oocyst, missed by direct wet mount technique, were detected in 26.92% of cases with the modified ZN staining method. The highest infection rate, 15.38%, was found among male children aged 4-6 years, who lived in an urban area (23.08%) and used bottled water (15.38%) [224].
10.3. Dietary Sources of C. belli
Vegetables are the main food source of C. belli that was the parasite with highest contamination rate (16.67%) and mean density (13 oocysts/g) in vegetables collected from March to August 2018 in the wet market and supermarkets of San Jose City, Nueva Ecija, Philippines [225] and the most frequent among parasites contaminating fruit preparations (tamarind water, sliced fruits and fruit juices) sold in schools in Dhaka city, Bangladesh [226].
11. Sarcocystis
Sarcocystis genus belongs to the clade Sar Alveolata, phylum Apicomplexa, family Sarcocistidae [2] and includes over 200 species of intracellular protozoan parasites, of which some species are zoonotic. The complete lifecycle is known for only 26 species and requires two hosts, generally an herbivore or carnivore intermediate host and a carnivore or omnivore definitive host [227]. The 18S rDNA sequence can differentiate members of the genus, while ribosomal ITS-1 and mitochondrial cytochrome oxidase (COI) sequences better highlight parasite diversity, relationships, and transmission cycles [8].
The life cycle comprises a sporocyst stage that, when ingested by the intermediate host, releases the sporozoites in the gastrointestinal tract. These enter the endothelial cells of blood vessels and undergo schizogony, resulting in first-generation schizonts or merozoites that invade small capillaries and blood vessels. Here these become second-generation schizonts and then second-generation merozoites that invade skeletal and heart myocytes, as well as neurons. Second-generation merozoites develop into metrocytes and undergo internal mitotic divisions until becoming filled with bradyzoites and finally a sarcocyst resistant to digestion. Sarcocysts in muscle do not spread to new cells, thus there seems to be no risk of recrudescence [227].
Oocysts containing two sporocysts are formed in the small intestine of the intermediate host and are excreted with the stool into the environment either as oocysts or sporocysts when the oocysts rupture. Sarcocystis hominis and S. heydorni produce thick-walled cysts or thin-walled cysts, respectively, in cattle tissue, preferentially in the heart muscle. After consumption of undercooked beef containing cysts humans can become definitive hosts of these Sarcocystis species. Also the definitive host eliminates with feces sporulated oocysts containing two sporocysts, each with four sporozoites [8].
11.1. Diseases Caused by Sarcocystis spp.
Most individuals with intestinal sarcocystosis remain asymptomatic but symptomatic infections may manifest with nausea, abdominal discomfort, and self-limiting diarrhea with severity varying with the amount of meat consumed. Diarrhea occurs between 3 and 48 h post-ingestion and resolves within 36 h. Segmental, eosinophilic, necrotizing enteritis attributed to sexual forms of Sarcocystis has been reported but not defined with certainty. Cases of extraintestinal sarcocystosis are most often asymptomatic but can give symptoms when merozoites invade striated muscles inducing vasculitis and musculoskeletal symptoms. In this case, painful swelling of muscle from 1 to 3 cm initially associated with erythema of the overlying skin that lasts from 2 days to 4 weeks, fever, diffuse myalgia, muscle tenderness, weakness, eosinophilia, and bronchospasm can be present. Treatments are directed toward the replicating stages of Sarcocystis spp. but do not affect sarcocystis in muscles. Therefore, symptomatic Sarcocystis infection is treated by palliative therapies with corticosteroids or other drugs that reduce the allergic inflammatory reactions that may occur. Diagnosis of Sarcocystis muscle infection by biopsy must exclude other muscle cyst-forming organisms, such as Toxoplasma gondii and Trypanosoma cruzi. Eosinophilia, compatible symptoms and epidemiologic exposure can also allow a diagnosis [8].
S. suihominis can cause a transient infection restricted to the gastrointestinal tract after consumption of undercooked pork. S. nesbitti, associated with macaques and snakes, was also reported as a cause of outbreaks in humans with symptoms including fever, myalgia, headache, and cough. Indeed, Sarcocystis infection can also be transmitted by water or food contaminated with feces of predatory carnivores [8].
A clinical report described marked symptoms in a 31-year-old AIDS patient, indicating that Sarcocystis should be considered an opportunistic pathogen in those persons [8].
S. fayeri, found in horses and deer, can cause acute food poisoning with vomiting and diarrhea after a short post-consumption incubation period for the production of a 15 kDa proteinaceous diarrheal toxin [8]. A similar toxin is probably produced by S. sybillensis and S. wapiti, which caused food poisoning from venison and in S. truncata, identified in raw deer meat that caused abdominal pain, vomiting, watery diarrhea, and fever in three Japanese persons [228]. Whale meat caused similar symptoms in four patients 12 h post-consumption and it was suggested that a Sarcocystis species associated with whale meat also produces a toxin [173].
11.2. Epidemiology of Sarcocystis spp.
The epidemiology of human sarcocystosis relies primarily on case reports and outbreaks from Southeast Asia. In Malaysia and Thailand seroprevalence studies and stool surveys showed widespread distribution and exposures. In one study on routine autopsy specimens, prevalence was estimated at more than 20%. A seroprevalence of 10% - 20% was reported in adults in rural areas of Laos and Tibet. An unidentified Sarcocystis species caused an outbreak of eosinophilic myositis in a USA military unit operating in Malaysia and one patient had serious, chronic sequelae. The parasite was detected in a muscle biopsy. A recent outbreak in Malaysia, involving 93 suspected cases, was caused by S. nesbitti [8].
11.3. Prevalence of Sarcocystis spp. in Food Producing Animals
Cattle are the intermediate host of several Sarcocystis species and bovine eosinophilic myositis (BEM) is an inflammatory myopathy that can be caused by S. cruzi and S. hominis, S. bovifelis and S. hirsuta [227,229]. It appears as diffuse grey-green patches on masticatory muscles, tongue, heart, and diaphragm. In severe cases all striated muscles can be involved [230] and, though clinical symptoms are not usually observed, BEM lesions can cause carcass depreciation and economic losses [227].
In a recent study, on cattle carcasses in a Belgian slaughterhouse it was found that female dairy cattle had the highest Sarcocystis occurrence rate (91%) that reached 100% in animals older than 7 years. Beef cattle males had the lowest Sarcocystis occurrence rate (22%) which might be partially explained by the lower age of animals. S. cruzi was the most prevalent species, followed by S. hominis and S. bovifelis. S. heydorni, S. bovini and S. hirsuta were found in female dairy cattle only. Other species occurring in cattle are S. rommeli, S. bovini, S. sinensis, S. gigantea, S. fusiformis, S. hjorti and S. tenella [230].
Though a high prevalence of Sarcocystis in cattle, the prevalence of BEM is quite low, ranging from 0.002% to 5% worldwide. However, the association of Sarcocystis with BEM was confirmed in a study in which Sarcocystis DNA was detected in 91.7% of the lesions [230].
S. miescheriana is associated with wild boars and in those hunted in Northern Italy Sarcocystis spp. showed a prevalence of 97% [231,232,233].
Species associated with goats and sheeps, S. capracanis and S. tenella which do not have humans as the definitive host can cause anorexia, weight loss, fever, anemia, hair loss, abortion, premature birth, neurologic signs, myositis, and death in sheeps [234,235,236]. In Lithuania a high prevalence of infection was established in cattle (44.9–98.1%) and sheep (100%), and a moderate prevalence in pigs (30.1–50.0%) and horses (34.7–63.9%) [237].
S. cruzi is the most pathogenic species in cattle, and acute infection can lead to weakness, reduced milk yield [238].
Measures for prevention of Sarcocystis spp. infection include boiling water that could be contaminated and avoiding the consumption of raw or undercooked beef and pork. Indeed, sarcocysts in pork can be destroyed by heat treatment at 60 °C for 20 min, 70 °C for 15 min or 100 °C for 5 min, or by freezing at −4 °C for 2 days or − 20 °C for 24 h. Sporocysts die when heated to 60 °C for 1 min, 55 °C for 15 min, or 50 °C for one hour, but can survive freezing [8]. These cysts lose viability in 2.0% salt after 1 day [228]. Commonly used disinfectants, e.g., 1% iodine, 10% formalin, 12% phenol and 2% chlorhexidine fail to kill sporocysts, but 5.25% sodium hydroxide is effective [8].
12. Protozoan Parasites with a Poorly Defined Impact on Health
12.1. Blastocystis
Blastocystis hominis, which many studies cited here designate Blastocystis spp., is an anaerobic intestinal protozoan of the Sar Stramenophiles clade, phylum Bigyra [2]. It has a broad host range and is one of the most prevalent microeukaryotes worldwide [24,26,239,240]. It is highly polymorphic and has four development stages, namely, cyst, granular, vacuolar and ameboid with a complex life cycle not yet fully described [239,241].
Sequence analysis of the small subunit ribosomal RNA (SSU-rRNA) allowed the distinction of more than 30 subtypes (STs) of B. hominis of which STs ST1 to ST8 were found both in humans and animals, ST9 was exclusively identified in humans, and ST10 to ST17 were identified only in animals [242]. The most frequently isolated STs in humans were ST1 to ST4, with ST3 being the most common worldwide [24,241,243,244,245,246,247,248] and the most pathogenic. This is able to invade the intestine by releasing a cysteine protease, which increases the production of interleukin-8 (IL-8) in human intestinal epithelial cells [246] and was associated with 53.3% of the abnormal findings from colonoscopies, including colitis, CRC, inflammatory ulcer and ileitis, and with 57.7% histopathological abnormal findings. ST1 was associated with 4.4% of abnormal findings from colonoscopies and 2.7% of abnormal findings from histopathology [246]. Subtype ST3 was significantly associated with chronic spontaneous urticaria and a statistically significant association was found between total IgE value and subtypes ST2 and ST3 [249]. Subtypes ST2 and ST1 showed higher intra-subtype variability than other subtypes [247,250].
The acquisition of these microorganisms occurs via the ingestion of cysts present in water or food or by direct contact with animal reservoirs. Symptoms are primarily gastrointestinal and manifest as lack of appetite, nausea, loose stools, abdominal pain, flatulence, bloating, constipation, and, less frequently, skin lesions or pruritus [251]. Symptoms are self-limiting [26] and many carriers are asymptomatic [247]. Blastocystis sp. was considered one of the main causes of traveler’s diarrhea, though this was not confirmed by recent findings [26]. Its pathogenicity might be attributed to increased oxidative damage and invasive activity, as demonstrated in a rat model.
It was estimated that about 1 billion people are infected with Blastocystis in the world and 50% of them remain asymptomatic carriers for months or years. B. hominis is the most commonly detected eukaryote in human fecal samples, especially in hot climates, with reported prevalence values of 58% in Alexandria, Egypt, 67% in Morocco [244,246,248,252], and 100% in 25 children below five years of age examined in Medellin Colombia [27].
In a large case-control study carried out in Zaragoza, Spain, B. hominis was detected in 9.18% of patients with gastrointestinal illnesses and symptoms such as abdominal pain, nausea, anorexia, aerophagia, halitosis, urticaria, anal itching and dyspepsia were significantly more common in patients positive for B. hominis than in controls negative for the parasite. Significant associations of B. hominis infection were observed with type 2 diabetes, bacterial co-infection and immunosuppressive treatments, except for treatment with corticosteroids [25].
B. hominis infection was associated with cutaneous manifestations such as urticaria, rash, and itching and these symptoms ceased when the infection was treated [253]. A recent case of urticaria complicated by arthritis and tenosynovitis occurred in an 18-year-old woman and did not improve with antihistamine treatment. The patient was affected by recurrent diarrhea in the past months and Blastocystis spp. was detected in the patient’s feces. She completely recovered after administration of metronidazole, naproxen and prednisolone [254].
A 3-year-old boy recently diagnosed with HIV at stage three and under treatment for pulmonary tuberculosis, suffered from diarrhea for about seven days, with also nausea, vomiting, sunken eyes, decreased skin turgor, and a rash on the extremities. Blastocystis hominis was identified in a stool specimen, thus he was treated with TMP-STX and symptoms disappeared. The infection was probably caused by close contact with pets, eating unwashed fruits and vegetables, and not washing hands [255].
An association of Blastocystis infection with the development of irritable bowel syndrome (IBS) and inflammatory bowel disease (IBD) was observed [181,251,253,256].
Evidence that B. hominis might play a role in the development of autoimmune disorders was achieved in a case–control study that included patients with different autoimmune diseases divided in a group with gastrointestinal problems (cases) and a group with no gastrointestinal symptoms (controls). B. hominis was identified more in cases (16.6%) than in controls (5.2%), and the difference was significant [240]. B. hominis infection can exacerbate symptoms of Hashimoto’s thyroiditis (HT), an autoimmune disease, by increasing serum levels of interleukin (IL-17) which plays a relevant role in pathogenesis. HT patients improved after treatment of B. hominis infection which led to lower levels of IL-17, thyroid stimulating hormone (TSH), and thyroid peroxidase antibodies (anti-TPO) [257]. Some studies found an association between B. hominis and CRC proposing that its presence was favored by immunosuppression. It was also suggested that the Blastocystis antigen can induce the proliferation of HCT116 colon carcinoma cells through the downregulation of p53 and proapoptotic genes and upregulation of IL-6 [246].
A case-control study carried out in Egypt found that patients with CRC infection did not have a higher prevalence of B. hominis infection but a significantly higher frequency of urticaria in the case of B. hominis infection. Moreover, B. hominis infection rates increased with CRC stage advancement. Subtype ST7 was isolated only from CRC stool samples [249].
B. hominis infection is frequently found in immunocompromised patients, showing characteristics of an opportunistic pathogen. The global estimated pooled prevalence of Blastocystis infection in immunocompromised people was 10% and, specifically, 21% in Australia 12% in America, 11% in Europe, 10% in Asia, and 7% in Africa [239]. In a cross-sectional study involving patients who were immunocompromised for different reasons, i.e. cancer, uncontrolled diabetes mellitus (DM), chronic renal diseases, and healthy controls, it was found that the percentage of Blastocystis spp. infection was higher in immunocompromised patients with no statistically significant difference among the different origins of immunodeficiency [258]. However, contrasting results were registered in Turkey where Blastocystis spp. was detected in a higher percentage in immunocompetent patients (21.8%) compared to immunocompromised patients (12.7%) [37].
In a rural community in Mexico B. hominis was the microorganism found with the highest frequency (44%) in schoolchildren aged 3 to 15 years old with the presence of subtypes ST1, ST2, and ST3. The only statistically significant risk factor was “eating sweets, snacks, and handmade food on the way home”. Therefore, it was concluded that they become infected mainly outside their homes, perhaps by eating contaminated handmade food [259].
Fresh vegetables have been identified as contributors to foodborne transmission [260] and B. hominis was the most frequently detected pathogenic protozoan in vegetable samples collected from Lebanese markets, with some samples contaminated also by Entamoeba spp., Balantidium spp., and G. intestinalis [261].
B. hominis has been detected in drinking water, tap water and drinking water treatment facilities worldwide and it is not destroyed by chlorine treatment at the concentrations of 0.2 - 5.0 ppm used to treat drinking water. It was observed that eleven isolates exposed to chlorine showed recovery after 24 h of incubation without chlorine exposure, suggesting that resistant forms, i.e. cysts, were formed during exposure. Concentrations ranging between 178 and 2,857 ppm were required to completely inhibit recovery and ST1 and ST7 were the most resistant subtypes. All isolates were also resistant to hydrogen peroxide, with ST8 and ST9 showing the highest resistance, and concentrations ranging from 103 ppm to 3,338 ppm were necessary to impede recovery after 24 without exposure. Therefore, hydrogen peroxide at concentrations used for disinfection, 5 - 30 ppm, is adequate to eliminate the organism from surfaces and objects [12].
12.2. Dientamoeba Fragilis
Dientamoeba fragilis belongs to the clade Metamonada, phylum Parabasalia [2] and has been reported in humans with a worldwide distribution. There are two described genotypes of D. fragilis, 1 and 2, that can be distinguished by restriction fragment length polymorphisms of the SSU rRNA gene [37].
Most studies on this protozoan parasite were conducted in industrialized countries and reported prevalence values of 0.3% up to 82.9%. Less is known about its occurrence in developing countries, but the few studies available reported a prevalence ranging between 0% and 60.6% [37]. The possible zoonotic transmission was suggested based on the detection of D. fragilis in pigs, cats and dogs but spreading by the fecal-oral route appears most likely [23].
The life cycle and clinical significance of this parasite are not well defined. Recently, a cyst stage was discovered and its transmission and association of the protozoan with symptomatic disease was shown in a rat model. Acquisition together with pinworms (Enterobius vermicularis) was also suggested. The advent of PCR methods determined an apparent increase in D. fragilis prevalence and the proportion of asymptomatic carriage, ranging between 11% and 39%, raised doubts about its pathogenicity. However, symptoms such as abdominal pain and diarrhea or loose stools and a chronic course lasting up to several years have been reported for 2% to 32% of the patients. In a retrospective study carried out in Finland, 85% of Dientamoeba patients, of whom 54% were also infected with B. hominis, reported loose stools, abdominal pain, constipation and fecal urgency indicative of colon irritation. Symptoms persisted for a median duration of 180 days [23]. Of 438 patients with diarrhea 11.9% were infected with D. fragilis in Turkey [37].
The most common examination methods used in clinical laboratories to detect D. fragilis are based on microscopy but these do not allow a definitive diagnosis so these are progressively replaced by molecular techniques [37].
12.3. Endolimax Nana
The genus Endolimax, like Entamoeba, belongs to the clade Amoebozoa, Endamoebidae family [2]. Endolimax spp. occur in mammalians, birds, reptiles, amphibians, fishes and insects and have two life cycle stages, an amoeboid trophozoite and a cyst. Intestinal colonization with Endolimax occurs after ingestion of mature cysts via the oral-fecal route. The trophozoite originates from the cyst and multiplies by binary fission in the host [28] where it feeds exclusively on bacteria [262]. The cysts are excreted in feces and can survive for two weeks at room temperature and for two months at lower temperatures [262].
E. nana is considered a commensal in the colon and appendix [262] but rare cases of abdominal pain, diarrhea, polyarthritis and urticaria were associated with infection with this protozoan. In a systematic review of E. nana prevalence most data were obtained from general studies on intestinal parasites and very few case reports are available [28]. A recent one involved a 34-year-old Italian woman who was affected by chronic urticaria, occasional abdominal pain and diarrhea with weight loss for four months after a journey to Vietnam. A stool examination showed the presence of numerous cysts of E. nana. She completely recovered after treatment with metronidazole with no need for using antihistamines. Cysts of E. nana were detected also in other patients with acute or chronic urticaria treated in the same dermatological center who traveled to tropical and subtropical countries [262].
12.4. Pentatrichomonas Hominis
The parasite Pentatrichomonas hominis belongs to the clade Metamonada, phylum Parabasalia [2]. It can be transmitted via the fecal-oral route, it is potentially zoonotic [20] and some studies suggest that it can cause disease symptoms such as diarrhea and gastrointestinal or pulmonary diseases in children and older people but its pathological role is still unclear [5,263]. This protozoan was detected in 13.8% of schoolchildren examined in Egypt [263,264].
P. hominis was recently detected in the stool of immunocompromised patients and it was reported that P. hominis infections significantly increased CRC incidence, though this observation was based on only 25 participants including 8 infected patients with CRC and 8 not infected patients with CRC and 9 controls. NGS of the 16S rRNA gene sequences of the V3 and V4 regions showed that in the infected patients the richness of the gut microbiota was reduced while it was not significantly altered in patients without P. hominis infection. Moreover, bacterial genera Flavonifractor sp., Lachnoclostridium sp., and the R. gnavus group, which are associated with the development of CRC, significantly increased in infected patients so the authors concluded that the infection with P. hominis can aggravate CRC by inducing an increase of intestinal bacteria associated with its development [265].
13. Conclusions
This literature survey highlighted the current trends in research regarding waterborne and foodborne pathogenic protozoans, with the number of published outbreaks and cases reflecting both the relevance of the pathogen for public health and the advancement in knowledge regarding the organisms, albeit representing only a small part of the real number of infections occurring. An overview of the number of cases and outbreaks of waterborne and foodborne protozoan infections published per protozoan pathogen since 2019 is shown in Figure 1.
The paucity or lack of reports for B. hominis, E. nana, D. fragilis and P. hominis reflects the poorly defined role of these species as pathogens. However, further investigation should consider evaluating their occurrence in cases with symptoms found to be associated with these organisms, such as urticaria-like reactions [253,254,262].
In the case of G. intestinalis, the low number of case/outbreak reports retrieved was probably biased by the type of investigations carried out for this pathogen, which was more focused on large epidemiological studies and less on single cases or outbreaks. This could also reflect a minor role of this pathogen in causing severe opportunistic infections.
In the case of B. coli, which is listed as a priority pathogen by FAO/WHO [18], the low number of reported outbreaks is explained by the rarity of severe infections and the high frequency of asymptomatic carriage. Indeed, the only case reported in this review was discovered fortuitously and was asymptomatic [218].
The identified sources of the infection cases and outbreaks most recently reported for waterborne and foodborne parasitic protozoans are shown in Table 1.
In Table 1, it is possible to observe how for some pathogens ascertained links to infection sources are still lacking. Also those reported were defined on the basis of epidemiological investigations and the organisms were not found in the food or water samples causing the illnesses, either because no more available at the time of the outbreaks, or for the lack of sensitivity of the analytical methods [9,113].
Indeed, the long incubation period that can be of some weeks before symptom manifestation resulted in difficulty in identifying the source in the majority of the foodborne outbreaks [9,130,132,178,266]. In addition, methods available for the detection of protozoans in food matrices are considered inefficient. These involve the separation of the parasite from the food matrix followed by detection with microscopy or molecular methods [9]. A guideline from the International Organization for Standardization (ISO), ISO 18744:2016 [267] is available only for the detection and enumeration of Cryptosporidium and G. intestinalis in vegetables and berry fruits, but it does not evaluate viability and does not allow species or genotype identification. Therefore, the development of standardized analytical methods is still needed for other food matrices from which the recovery of the parasites is difficult, as in the case of Cryptosporidium from milk and meat [9,129,169]. The development and standardization of reliable detection methods would greatly improve prevention and could represent a valuable tool for surveillance.
The implementation of surveillance programs is another critical aspect to be considered since even protozoans with a high pathogenic potential are still not included in national surveillance plans [164].
Most of the pathogens considered in this review are zoonotic, so One Health plans for a better definition of the prevalence and efficient treatments of the disease in farm animals, an increase of public health organism awareness and cooperation among professionals in veterinary and human medicine as well as decision-makers, would greatly reduce parasite transmission [10,13,113]. To this end, the development of new genotyping methods, still unavailable for some species and subtypes, would be essential for pathogen source tracking.
Also, metagenomics techniques can be exploited to elucidate the protozoan pathogen transmission chain, as shown by a study that used metataxonomic and metagenomic techniques, coupled with traditional outbreak investigation, to trace the origin of a foodborne outbreak caused by C. parvum in Sweden. In that study, case interviews, sampling of patients, and back-tracing identified romaine lettuce from a specific farm as the source of the outbreak. Then, the microbiota of romaine lettuce and other crops collected in the farm was sequenced with both 16S rRNA gene V4 hypervariable region amplicon metagenomics and shotgun sequencing. C. parvum of the same subtype of the outbreak, IIdA24G1, was found in two more samples of romaine lettuce. Moreover, microbiome sequences associated with fecal environments of farm animals, and untreated sewage were identified in those samples. In addition, shotgun sequencing of the bacteria present identified some species highly associated with human and animal fecal environments. Since the farm used only chemical fertilizers and irrigation water was taken from a deep well that was not contaminated at the time of the outbreak, it was concluded that the most likely source of fecal pollution was human and it could derive either from wastewater contamination of the irrigation water or from water used for washing the lettuce, as well as from infected people handling the lettuce [133].
Nearly all the studies on prevalence considered in this review indicated that improving people’s hygiene habits is essential to reduce the prevalence of infections caused by parasitic protozoans. Worth of note is the association of not washing hands with an increased risk of infection found in many epidemiological studies.
In this respect, the application of the WHO WASH collection of integrated prevention and control strategies for infectious diseases, including handwashing among the most effective prevention measures [67,112,120,268], represents a valuable tool to decrease the public health burden of protozoan infections. The WASH strategies also include making available safe water sources and clean toilets that can decrease the occurrence of diarrhea by 15 to 50%. Handwashing with soap can reduce up to 40% of nonemergency diarrheal cases. WASH application allowed a 27 to 56% reduction in diarrhea occurrence in children of less than 5 years of age in low-income countries, e.g. giardiasis in rural Bangladesh, with handwashing as the most effective long-term intervention. Moreover, a point-of-use drinking water filter reduced the occurrence of Cryptosporidium infection in children in Rwanda.
Author Contributions
G.C. and F.R.: conceptualization; F.R.: methodology, data curation, and original draft preparation; S.S., C.A., L.M. and S.S.: investigation, review, and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yaeger, R.G. Protozoa: Structure, Classification, Growth, and of the Development. Chapter 77 in: Medical Microbiology. Editor: Samuel Baron. 4th edition. University of Texas Medical Branch at Galveston. 1996. Available online: https://www.ncbi.nlm.nih.gov/books/NBK7627/ (accessed on 1 December 2023).
- National Center for Biotechnology Information. Taxonomy browser. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?mode=Info&id=2698737&lvl=3&lin=f&keep=1&srchmode=1&unlock (accessed on 1 March 2024).
- Baig, A.M.; Suo, X.; Liu, D. Chapter 148 - Pathogenesis of protozoan infections, Editor(s): Yi-Wei Tang, Musa Y. Hindiyeh, Dongyou Liu, Andrew Sails, Paul Spearman, Jing-Ren Zhang, Molecular Medical Microbiology (Third Edition), Academic Press, 2024, Pages 2921-2940.
- Pignata, C.; Bonetta, S.; Bonetta, S.; Cacciò, S.M.; Sannella, A.R.; Gilli, G.; Carraro, E. Cryptosporidium Oocyst Contamination in Drinking Water: A Case Study in Italy. Int. J. Environ. Res. Public Health 2019, 16, 2055. [Google Scholar] [CrossRef]
- Chique, C.; Hynds, P.D.; Andrade, L.; Burke, L.; Morris, D.; Ryan, M.P.; O’Dwyer, J. Cryptosporidium Spp. in Groundwater Supplies Intended for Human Consumption - A Descriptive Review of Global Prevalence, Risk Factors and Knowledge Gaps. Water Res. 2020, 176, 115726. [Google Scholar] [CrossRef]
- Hadjilouka, A.; Tsaltas, D. Cyclospora Cayetanensis-Major Outbreaks from Ready to Eat Fresh Fruits and Vegetables. Foods 2020, 9. [Google Scholar] [CrossRef]
- Tharmaratnam, T.; Kumanan, T.; Iskandar, M.A.; D’Urzo, K.; Gopee-Ramanan, P.; Loganathan, M.; Tabobondung, T.; Tabobondung, T.A.; Sivagurunathan, S.; Patel, M.; Tobbia, I. Entamoeba Histolytica and Amoebic Liver Abscess in Northern Sri Lanka: A Public Health Problem. Trop. Med. Health 2020, 48, 2. [Google Scholar] [CrossRef]
- Rosenthal, B.M. Zoonotic Sarcocystis. Res. Vet. Sci. 2021, 136, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.; Razakandrainibe, R.; Basmaciyan, L.; Raibaut, J.; Delaunay, P.; Morio, F.; Gargala, G.; Villier, V.; Mouhajir, A.; Levy, B.; Rieder, C.; Larreche, S.; Lesthelle, S.; Coron, N.; Menu, E.; Demar, M.; de Santi, V.P.; Blanc, V.; Valot, S.; Dalle, F.; Favennec, L. A Summary of Cryptosporidiosis Outbreaks Reported in France and Overseas Departments, 2017-2020. Food Waterborne Parasitol. 2022, 27, e00160. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Hafez, H.M. Cryptosporidiosis: From Prevention to Treatment, a Narrative review. Microorganisms 2022, 10, 2456. [Google Scholar] [CrossRef]
- López-García, A.; Gilabert, J.A. Oral Transmission of Chagas Disease from a One Health Approach: A Systematic Review. Trop. Med. Int. Health 2023, 28, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Martín-Escolano, R.; Ng, G.C.; Tan, K.S.W.; Stensvold, C.R.; Gentekaki, E.; Tsaousis, A.D. Resistance of Blastocystis to Chlorine and Hydrogen Peroxide. Parasitol. Res. 2023, 122, 167–176. [Google Scholar] [CrossRef]
- López Ureña, N.M.; Chaudhry, U.; Calero Bernal, R.; Cano Alsua, S.; Messina, D.; Evangelista, F.; Betson, M.; Lalle, M.; Jokelainen, P.; Ortega Mora, L.M.; Álvarez García, G. Contamination of Soil, Water, Fresh Produce, and Bivalve Mollusks with Toxoplasma Gondii Oocysts: A Systematic Review. Microorganisms 2022, 10, 517. [Google Scholar] [CrossRef]
- Siwila, J.; Mwaba, F.; Chidumayo, N.; Mubanga, C. Food and Waterborne Protozoan Parasites: The African Perspective. Food Waterborne Parasitol. 2020, 20, e00088. [Google Scholar] [CrossRef]
- Brožová, K.; Jirků, M.; Lhotská, Z.; Květoňová, D.; Kadlecová, O.; Stensvold, C.R.; Samaš, P.; Petrželková, K.J.; Jirků, K. The Opportunistic Protist, Giardia Intestinalis, Occurs in Gut-Healthy Humans in a High-Income Country. Emerg. Microbes Infect. 2023, 12, 2270077. [Google Scholar] [CrossRef]
- World Health Organization Team Control of Neglected Tropical Diseases. Prevention and control of intestinal parasitic infections: WHO Technical Report Series N°749. Guideline. Available online: https://www.who.int/publications/i/item/WHO-TRS-749 (accessed on 16 November 2023).
- Das, R.; Palit, P.; Haque, M.A.; Levine, M.M.; Kotloff, K.L.; Nasrin, D.; Hossain, M.J.; Sur, D.; Ahmed, T.; Breiman, R.F.; Freeman, M.C.; Faruque, A.S.G. Symptomatic and Asymptomatic Enteric Protozoan Parasitic Infection and Their Association with Subsequent Growth Parameters in under Five Children in South Asia and Sub-Saharan Africa. PLoS Negl. Trop. Dis. 2023, 17, e0011687. [Google Scholar] [CrossRef]
- Dos Santos, T.L.; Medeiros de Carvalho Neto, A.; Da Silva Ferreira, J.R.; Azevedo, P.V.M.; Wallas Lins Da Silva, K.; Alencar dos Nascimento, C.M.; Calheiros, C.M.L.; Wanderley, F.S. G. Dos Santos Cavalcanti, M.; Alencar Do Nascimento Rocha, M.; Rocha, T.J.M. Frequency of Intestinal Protozoan Infections Diagnosed in Patients from a Clinical Analysis Laboratory. Biosci. J. 2022, 38, e38001. [Google Scholar]
- FAO/WHO. Multicriteria-Based Ranking for Risk Management of Food-Borne Parasites. Rome. 2014. Available online: http://www.who.int/iris/handle/10665/112672 (accessed on 16 February 2024).
- Iordanov, R.B.; Leining, L.M.; Wu, M.; Chan, G.; DiNardo, A.R.; Mejia, R. Case Report: Molecular Diagnosis of Cystoisospora Belli in a Severely Immunocompromised Patient with HIV and Kaposi Sarcoma. Am. J. Trop. Med. Hyg. 2021, 106, 678–680. [Google Scholar] [CrossRef]
- Mahittikorn, A.; Udonsom, R.; Koompapong, K.; Chiabchalard, R.; Sutthikornchai, C.; Sreepian, P.M.; Mori, H.; Popruk, S. Molecular Identification of Pentatrichomonas Hominis in Animals in Central and Western Thailand. BMC Vet. Res. 2021, 17, 203. [Google Scholar] [CrossRef] [PubMed]
- Noor, M.; Katzman, P.J.; Huber, A.R.; Findeis-Hosey, J.J.; Whitney-Miller, C.; Gonzalez, R.S.; Zhou, Z.; N’kodia, H.D.; Skonick, K.; Abell, R.L.; Saubermann, L.J.; Lamps, L.W.; Drage, M.G. Unexpectedly High Prevalence of Cystoisospora Belli Infection in Acalculous Gallbladders of Immunocompetent Patients. Am. J. Clin. Pathol. 2019, 151, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, J.-P.; Meri, T.; Siikamäki, H.; Tyyni, E.; Kerttula, A.-M.; Pakarinen, L.; Jokiranta, T.S.; Kantele, A. Dientamoeba Fragilis - the Most Common Intestinal Protozoan in the Helsinki Metropolitan Area, Finland, 2007 to 2017. Euro Surveill. 2019, 24. [Google Scholar] [CrossRef] [PubMed]
- Popruk, S.; Adao, D.E.V.; Rivera, W.L. Epidemiology and Subtype Distribution of Blastocystis in Humans: A Review. Infect. Genet. Evol. 2021, 95, 105085. [Google Scholar] [CrossRef] [PubMed]
- Matovelle, C.; Tejedor, M.T.; Monteagudo, L.V.; Beltrán, A.; Quílez, J. Prevalence and Associated Factors of Blastocystis Sp. Infection in Patients with Gastrointestinal Symptoms in Spain: A Case-Control Study. Trop. Med. Infect. Dis. 2022, 7, 226. [Google Scholar] [CrossRef] [PubMed]
- Cobuccio, L.G.; Laurent, M.; Gardiol, C.; Wampfler, R.; Poppert, S.; Senn, N.; Eperon, G.; Genton, B.; Locatelli, I.; de Vallière, S. Should We Treat Blastocystis Sp.? A Double-Blind Placebo-Controlled Randomized Pilot Trial. J. Travel Med. 2023, 30. [Google Scholar] [CrossRef]
- Garcia-Montoya, G.M.; Galvan-Diaz, A.L.; Alzate, J.F. Metataxomics Reveals Blastocystis Subtypes Mixed Infections in Colombian Children. Infect. Genet. Evol. 2023, 113, 105478. [Google Scholar] [CrossRef]
- Hooshyar, H.; Arbabi, M.; Rostamkhani, P. Checklist of Endolimax Species Recognized in Human and Animals, a Review on a Neglected Intestinal Parasitic Amoeba. Iraq Med. J. 2023, 7, 28–31. [Google Scholar] [CrossRef]
- Weitzel, T.; Brown, A.; Libman, M.; Perret, C.; Huits, R.; Chen, L.; Leung, D.; Leder, K.; Connor, B.A.; Menéndez, M.D.; Asgeirsson, H.; Schwartz, E.; Salvador, F.; Malvy, D.; Saio, M.; Norman, F.F.; Amatya, B.; Duvingnaud, A.; Vaughan, S.; Glynn, M.; Angelo, K.M.; GeoSentinel Network. Intestinal protozoa in returning travellers: a GeoSentinel analysis from 2007 to 2019. J Travel Med. 2024 Jan 21:taae010.
- Salim, M.; Mohammad Shahid, M.; Shagufta, P. The Possible Involvement of Protozoans in Causing Cancer in Human. Egypt. Acad. J. Biol. Sci., Med. Entomol. Parasitol. 2022, 14, 71–86. [Google Scholar] [CrossRef]
- Zhang, N.; Yu, X.; Zhang, H.; Cui, L.; Li, X.; Zhang, X.; Gong, P.; Li, J.; Li, Z.; Wang, X.; Li, X.; Li, T.; Liu, X.; Yu, Y.; Zhang, X. Prevalence and Genotyping of Cryptosporidium Parvum in Gastrointestinal Cancer Patients. J. Cancer 2020, 11, 3334–3339. [Google Scholar] [CrossRef]
- Certad, G. Is Cryptosporidium a Hijacker Able to Drive Cancer Cell Proliferation? Food Waterborne Parasitol. 2022, 27, e00153. [Google Scholar] [CrossRef]
- Berrouch, S.; Escotte-Binet, S.; Harrak, R.; Huguenin, A.; Flori, P.; Favennec, L.; Villena, I.; Hafid, J. Detection Methods and Prevalence of Transmission Stages of Toxoplasma Gondii, Giardia Duodenalis and Cryptosporidium Spp. in Fresh Vegetables: A Review. Parasitology 2020, 147, 516–532. [Google Scholar] [CrossRef]
- Berrouch, S.; Escotte-Binet, S.; Amraouza, Y.; Flori, P.; Aubert, D.; Villena, I.; Hafid, J. Cryptosporidium Spp., Giardia Duodenalis and Toxoplasma Gondii Detection in Fresh Vegetables Consumed in Marrakech, Morocco. Afr. Health Sci. 2020, 20, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Blake, H.; Fortes, B.H.; Dilmaghani, S.; Ryan, S.M. 34-Year-Old Woman with Human Immunodeficiency Virus and Chronic Diarrhea. Mayo Clin. Proc. 2021, 96, 2474–2479. [Google Scholar]
- Özmen-Çapın, B.B.; Gürsoy, G.; Tortop, S.; Jabrayilov, J.; İnkaya, A.Ç.; Ergüven, S. Cystoisospora Belli Infection in a Renal Transplant Recipient: A Case Report and Review of Literature. J. Infect. Dev. Ctries. 2021, 15, 599–602. [Google Scholar] [CrossRef] [PubMed]
- Sarzhanov, F.; Dogruman-Al, F.; Santin, M.; Maloney, J.G.; Gureser, A.S.; Karasartova, D.; Taylan-Ozkan, A. Investigation of Neglected Protists Blastocystis Sp. and Dientamoeba fragilis in Immunocompetent and Immunodeficient Diarrheal Patients Using Both Conventional and Molecular Methods. PLoS Negl. Trop. Dis. 2021, 15, e0009779. [Google Scholar] [CrossRef]
- Barlaam, A.; Sannella, A.R.; Ferrari, N.; Temesgen, T.T.; Rinaldi, L.; Normanno, G.; Cacciò, S.M.; Robertson, L.J.; Giangaspero, A. Ready-to-Eat Salads and Berry Fruits Purchased in Italy Contaminated by Cryptosporidium Spp., Giardia Duodenalis, and Entamoeba Histolytica. Int. J. Food Microbiol. 2022, 370, 109634. [Google Scholar] [CrossRef]
- Mesfun, M.G.; Fuchs, A.; Holtfreter, M.C.; Tufa, T.B.; Orth, H.M.; Luedde, T.; Feldt, T. The Implementation of the Kinyoun Staining Technique in a Resource-Limited Setting Is Feasible and Reveals a High Prevalence of Intestinal Cryptosporidiosis in Patients with HIV. Int. J. Infect. Dis. 2022, 122, 130–135. [Google Scholar] [CrossRef]
- El Bakri, A.; Hussein, N.M.; Ibrahim, Z.A.; Hasan, H.; AbuOdeh, R. Intestinal Parasite Detection in Assorted Vegetables in the United Arab Emirates. Oman Med. J. 2020, 35, e128. [Google Scholar] [CrossRef]
- Al-Adhroey, A.H.; Mehrass, A.A.-K.O.; Al-Shammakh, A.A.; Ali, A.D.; Akabat, M.Y.M.; Al-Mekhlafi, H.M. Prevalence and Predictors of Toxoplasma Gondii Infection in Pregnant Women from Dhamar, Yemen. BMC Infect. Dis. 2019, 19, 1089. [Google Scholar] [CrossRef]
- Franco-Paredes, C.; Villamil-Gómez, W.E.; Schultz, J.; Henao-Martínez, A.F.; Parra-Henao, G.; Rassi, A., Jr; Rodríguez-Morales, A.J.; Suarez, J.A. A Deadly Feast: Elucidating the Burden of Orally Acquired Acute Chagas Disease in Latin America - Public Health and Travel Medicine Importance. Travel Med. Infect. Dis. 2020, 36, 101565. [Google Scholar] [CrossRef] [PubMed]
- Rehman, F.; Shah, M.; Ali, A.; Ahmad, I.; Sarwar, M.T.; Rapisarda, A.M.C.; Cianci, A. Unpasteurised Milk Consumption as a Potential Risk Factor for Toxoplasmosis in Females with Recurrent Pregnancy Loss. J. Obstet. Gynaecol. 2020, 40, 1106–1110. [Google Scholar] [CrossRef] [PubMed]
- Papatsiros, V.G.; Athanasiou, L.V.; Kostoulas, P.; Giannakopoulos, A.; Tzika, E.; Billinis, C. Toxoplasma Gondii Infection in Swine: Implications for Public Health. Foodborne Pathog. Dis. 2021, 18, 823–840. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Frickmann, H.; Tannich, E.; Poppert, S.; Hagen, R.M. Colitis Caused by Entamoeba Histolytica Identified by Real-Time-PCR and Fluorescence in Situ Hybridization from Formalin-Fixed, Paraffin-Embedded Tissue. Eur. J. Microbiol. Immunol. (Bp.) 2022. [Google Scholar] [CrossRef] [PubMed]
- Dini, F.M.; Morselli, S.; Marangoni, A.; Taddei, R.; Maioli, G.; Roncarati, G.; Balboni, A.; Dondi, F.; Lunetta, F.; Galuppi, R. Spread of Toxoplasma Gondii among Animals and Humans in Northern Italy: A Retrospective Analysis in a One-Health Framework. Food Waterborne Parasitol. 2023, 32, e00197. [Google Scholar] [CrossRef] [PubMed]
- Dominic, C.; Kelly, S.; Melzer, M. Refractory Inflammatory Bowel Disease – Entamoeba Histolytica, the Forgotten Suspect. Clin. Infect. Pract. 2023, 20, 100245. [Google Scholar] [CrossRef]
- Dong, W.; Zhong, Q.; Gu, Y.-L.; Liang, N.; Zhou, Y.-H.; Cong, X.-M.; Liang, J.-Y.; Wang, X.-M. Is Toxoplasma Gondii Infection a Concern in Individuals with Rheumatic Diseases? Evidence from a Case-Control Study Based on Serological Diagnosis. Microb. Pathog. 2023, 182, 106257. [Google Scholar] [CrossRef]
- Friesema, I.H.M.; Hofhuis, A.; Hoek-van Deursen, D.; Jansz, A.R.; Ott, A.; van Hellemond, J.J.; van der Giessen, J.; Kortbeek, L.M.; Opsteegh, M. Risk Factors for Acute Toxoplasmosis in the Netherlands. Epidemiol. Infect. 2023, 151, e95. [Google Scholar] [CrossRef]
- Rabaan, A.A.; Uzairue, L.I.; Alfaraj, A.H.; Halwani, M.A.; Muzaheed, *!!! REPLACE !!!*; Alawfi, A.; Alshengeti, A.; Al Kaabi, N.A.; Alawad, E.; Alhajri, M.; Alwarthan, S.; Alshukairi, A.N.; Almuthree, S.A.; Alsubki, R.A.; Alshehri, N.N.; Alissa, M.; Albayat, H.; Zaidan, T.I.; Alagoul, H.; Fraij, A.A.; Alestad, J.H. Seroprevalence, Risk Factors and Maternal-Fetal Outcomes of Toxoplasma Gondii in Pregnant Women from WHO Eastern Mediterranean Region: Systematic Review and Meta-Analysis. Pathogens 2023, 12, 1157. [Google Scholar] [CrossRef] [PubMed]
- Sini, M.F.; Manconi, M.; Varcasia, A.; Massei, G.; Sandu, R.; Mehmood, N.; Ahmed, F.; Carta, C.; Cantacessi, C.; Scarano, C.; Scala, A.; Tamponi, C. Seroepidemiological and biomolecular survey on Toxoplasma gondii in Sardinian wild boar (Sus scrofa). Food Waterborne Parasitol. 2024, 34, e00222. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, H.; Rostamkhani, P.; Arbabi, M.; Delavari, M. Giardia Lamblia Infection: Review of Current Diagnostic Strategies. Gastroenterol. Hepatol. Bed Bench 2019, 12, 3–12. [Google Scholar]
- Kyany’a, C.; Eyase, F.; Odundo, E.; Kipkirui, E.; Kipkemoi, N.; Kirera, R.; Philip, C.; Ndonye, J.; Kirui, M.; Ombogo, A.; Koech, M.; Bulimo, W.; Hulseberg, C.E. First Report of Entamoeba Moshkovskii in Human Stool Samples from Symptomatic and Asymptomatic Participants in Kenya. Trop. Dis. Travel Med. Vaccines 2019, 5, 23. [Google Scholar] [CrossRef]
- Kantzanou, M.; Karalexi, M.A.; Vrioni, G.; Tsakris, A. Prevalence of Intestinal Parasitic Infections among Children in Europe over the Last Five Years. Trop. Med. Infect. Dis. 2021, 6, 160. [Google Scholar] [CrossRef]
- Candela, E.; Goizueta, C.; Periago, M.V.; Muñoz-Antoli, C. Prevalence of Intestinal Parasites and Molecular Characterization of Giardia Intestinalis, Blastocystis Spp. and Entamoeba Histolytica in the Village of Fortín Mbororé (Puerto Iguazú, Misiones, Argentina). Parasit. Vectors 2021, 14, 510. [Google Scholar] [CrossRef]
- Tegen, D.; Damtie, D.; Hailegebriel, T. Prevalence and Associated Risk Factors of Human Intestinal Protozoan Parasitic Infections in Ethiopia: A Systematic Review and Meta-Analysis. J. Parasitol. Res. 2020, 2020, 8884064. [Google Scholar] [CrossRef]
- Berhe, B.; Mardu, F.; Tesfay, K.; Legese, H.; Adhanom, G.; Haileslasie, H.; Gebremichail, G.; Tesfanchal, B.; Shishay, N.; Negash, H. More than Half Prevalence of Protozoan Parasitic Infections among Diarrheic Outpatients in Eastern Tigrai, Ethiopia, 2019; A Cross-Sectional Study. Infect. Drug Resist. 2020, 13, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Tilahun, Y.; Gebreananiya, T.A. Prevalence of human intestinal parasitic infections and associated risk factors at Lay Armachiho District Tikildingay town health center, Northwest Ethiopia. Research Square 2023. [Google Scholar] [CrossRef]
- Hijjawi, N.; Zahedi, A.; Ryan, U. Molecular Characterization of Entamoeba, Blastocystis and Cryptosporidium Species in Stool Samples Collected from Jordanian Patients Suffering from Gastroenteritis. Trop. Parasitol. 2021, 11, 122–125. [Google Scholar] [PubMed]
- Karim, A.; Zartashia, B.; Khwaja, S.; Akhter, A.; Raza, A.A.; Parveen, S. Prevalence and Risk Factors Associated with Human Intestinal Parasitic Infections (IPIs) in Rural and Urban Areas of Quetta, Pakistan. Braz. J. Biol. 2023, 84, e266898. [Google Scholar] [CrossRef]
- Al-Sultany, A.K.; Al-Morshidy, K.A.H. An Epidemiological Study of Intestinal Parasites in Children Attending the Pediatric Teaching Hospital in the Holy City of Karbala, Iraq. Medical Journal of Babylon 2023, 20, 95–100. [Google Scholar]
- Deksne, G.; Krūmiņš, A.; Mateusa, M.; Morozovs, V.; Šveisberga, D.P.; Korotinska, R.; Bormane, A.; Vīksna, L.; Krūmiņa, A. Occurrence of Cryptosporidium spp. and Giardia spp. Infection in Humans in Latvia: Evidence of Underdiagnosed and Underreported Cases. Medicina (Kaunas) 2022, 58, 471. [Google Scholar] [CrossRef]
- Symmers, W.S.C. Opportunistic Infections: The Concept of ‘Opportunistic Infections. ’ Proc. R. Soc. Med. 1965, 58, 341–346. [Google Scholar] [CrossRef]
- Lobo, Y.; Holland, T.; Spelman, L. Toxoplasmosis in a Patient Receiving Ixekizumab for Psoriasis. JAAD Case Rep. 2020, 6, 204–206. [Google Scholar] [CrossRef]
- Tiryaki, T.O.; Anıl, K.U.; Büyük, M.; Yıldırım, A.Y.; Atasoy, A.; Çiftçibaşı Örmeci, A.; Kalayoğlu Beşışık, S. Prolonged Severe Watery Diarrhea in a Long-Term Myeloma Survivor: An Unforeseen Infection with Cystoisospora Belli. Turk. J. Hematol. 2021, 38, 171–173. [Google Scholar] [CrossRef]
- Galván-Díaz, A.L.; Alzate, J.C.; Villegas, E.; Giraldo, S.; Botero, J.; García-Montoya, G. Chronic Cystoisospora Belli Infection in a Colombian Patient Living with HIV and Poor Adherence to Highly Active Antiretroviral Therapy. Biomedica 2021, 41 (Suppl. 1), 17–22. [Google Scholar] [CrossRef]
- Yang, X.; Guo, Y.; Xiao, L.; Feng, Y. Molecular Epidemiology of Human Cryptosporidiosis in Low- and Middle-Income Countries. Clin. Microbiol. Rev. 2021, 34. [Google Scholar] [CrossRef]
- Alkorta, M.; Manzanal, A.; Zeberio, I. A Case of Severe Diarrhoea Caused by Cyclospora Cayetanensis in an Immunocompromised Patient in Northern Spain. Access Microbiol. 2022, 4, 000313. [Google Scholar] [CrossRef]
- Rahadiani, N.; Habiburrahman, M.; Putranto, A.S.; Handjari, D.R.; Stephanie, M.; Krisnuhoni, E. Fulminant Necrotizing Amoebic Colitis Presenting as Acute Appendicitis: A Case Report and Comprehensive Literature Review. J. Infect. Dev. Ctries. 2022, 16, 717–725. [Google Scholar] [CrossRef]
- Tawfik, O.; Hassan, A.; Mohamed, W.; Essmyer, C.; Ewing, E.; Lem, V.; Grobelny, B.; Breshears, J.; Akhtar, N. Disseminated Opportunistic Infections Masquerading as Central Nervous System Malignancies. Human Pathology Reports 2022, 28, 300629. [Google Scholar] [CrossRef]
- Kanaujia, R.; Mewara, A. Cystoisospora Belli: A Cause of Chronic Diarrhea in Immunocompromised Patients. Am. J. Trop. Med. Hyg. 2023, 108, 1084–1085. [Google Scholar] [CrossRef] [PubMed]
- Sardar, S.K.; Goel, G.; Ghosal, A.; Deshmukh, R.; Bhattacharya, S.; Haldar, T.; Maruf, M.M.; Mahto, R.; Kumar, J.; Bhave, S.J.; Dutta, S.; Ganguly, S. First Case Report of Cyclosporiasis from Eastern India: Incidence of Cyclospora Cayetanensis in a Patient with Unusual Diarrheal Symptoms. J. Infect. Dev. Ctries. 2023, 17, 1037–1040. [Google Scholar] [CrossRef]
- Tie, X.; Zhang, Z.; Zhou, R.; Li, Y.; Xu, J.; Yin, W. A Case of Septic Shock Due to Delayed Diagnosis of Cryptosporidium Infection after Liver Transplantation. BMC Infect. Dis. 2023, 23, 260. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, S.L.; Erasmus, L.; Thomas, J.; Groome, M.J.; Plessis, N.M.D.; Avenant, T.; de Villiers, M.; Page, N.A. Epidemiology and aetiology of moderate to severe diarrhoea in hospitalised HIV-infected patients ≥5 years old in South Africa, 2018-2021: a case-control analysis. Available online: https://www.medrxiv.org/content/10.1101/2023.02.23.23286353v1.
- Younis, E.Z.; Elamami, A.H.; El-Salem, R.M. Intestinal Parasitic Infections among Patients with Type 2 Diabetes Mellitus in Benghazi, Libya. JCDO 2021, 1, 1–4. [Google Scholar] [CrossRef]
- Konadu, E.; Essuman, M.A.; Amponsah, A.; Agroh, W.X.K.; Badu-Boateng, E.; Gbedema, S.Y.; Boakye, Y.D. Enteric Protozoan Parasitosis and Associated Factors among Patients with and without Diabetes Mellitus in a Teaching Hospital in Ghana. Int. J. Microbiol. 2023, 2023, 5569262. [Google Scholar] [CrossRef]
- Taghipour, A.; Javanmard, E.; Rahimi, H.M.; Abdoli, A.; Matin, S.; Haghbin, M.; Olfatifar, M.; Mirjalali, H.; Zali, M.R. Prevalence of Intestinal Parasitic Infections in Patients with Diabetes: A Systematic Review and Meta-Analysis. Int. Health 2023. [Google Scholar] [CrossRef]
- Sror, S.M.; Galal, S.M.; Mohamed, Y.M.; Dyab, A.K. Prevalence of Intestinal Parasitic Infection among Children with Chronic Liver Diseases, Assiut Governorate, Egypt. Aljouf Univ. Med. J. 2019, 6, 1–8. [Google Scholar] [CrossRef]
- Łanocha, A.; Łanocha-Arendarczyk, N.; Wilczyńska, D.; Zdziarska, B.; Kosik-Bogacka, D. Protozoan Intestinal Parasitic Infection in Patients with Hematological Malignancies. J. Clin. Med. 2022, 11, 2847. [Google Scholar] [CrossRef] [PubMed]
- Patil, A.D.; Saxena, N.G.; Thakare, S.B.; Pajai, A.E.; Bajpai, D.; Jamale, T.E. Diarrhea after Kidney Transplantation: A Study of Risk Factors and Outcomes. J. Postgrad. Med. 2023, 69, 205–214. [Google Scholar] [CrossRef]
- Eslahi, A.V.; Olfatifar, M.; Zaki, L.; Karimipour Saryazdi, A.; Barikbin, F.; Maleki, A.; Abdoli, A.; Badri, M.; Karanis, P. Global Prevalence of Intestinal Protozoan Parasites among Food Handlers: A Systematic Review and Meta-Analysis. Food Control 2023, 145, 109466. [Google Scholar] [CrossRef]
- Yimam, Y.; Woreta, A.; Mohebali, M. Intestinal Parasites among Food Handlers of Food Service Establishments in Ethiopia: A Systematic Review and Meta-Analysis. BMC Public Health 2020, 20, 73. [Google Scholar] [CrossRef] [PubMed]
- Hajare, S.T.; Gobena, R.K.; Chauhan, N.M.; Erniso, F. Prevalence of Intestinal Parasite Infections and Their Associated Factors among Food Handlers Working in Selected Catering Establishments from Bule Hora, Ethiopia. Biomed Res. Int. 2021, 2021, 6669742. [Google Scholar] [CrossRef] [PubMed]
- Lette, A.; Negash, G.; Kumbi, M.; Hussen, A.; Kassim, J.; Zenbaba, D.; Gezahgn, H.; Bonsa, M.; Aman, R.; Abdulkadir, A. Predictors of Intestinal Parasites among Food Handlers in, Southeast Ethiopia, 2020. J. Parasitol. Res. 2022, 2022, 3329237. [Google Scholar] [CrossRef]
- Wondimu, H.; Mihret, M. Prevalence and Associated Factors of Intestinal Parasites among Food Handlers Working in Food Service Establishments in Northwest Ethiopia, 2022. J. Parasitol. Res. 2023, 2023, 3230139. [Google Scholar] [CrossRef]
- Alqarni, A.S.; Wakid, M.H.; Gattan, H.S. Prevalence, Type of Infections and Comparative Analysis of Detection Techniques of Intestinal Parasites in the Province of Belgarn, Saudi Arabia. PeerJ 2022, 10, e13889. [Google Scholar] [CrossRef]
- Şahin, M.; Ödemiş, N.; Yılmaz, H.; Beyhan, Y.E. Investigation of Parasites in Food Handlers in Turkey. Foodborne Pathog. Dis. 2023, 20, 381–387. [Google Scholar] [CrossRef]
- Ma, J.-Y.; Li, M.-Y.; Qi, Z.-Z.; Fu, M.; Sun, T.-F.; Elsheikha, H.M.; Cong, W. Waterborne Protozoan Outbreaks: An Update on the Global, Regional, and National Prevalence from 2017 to 2020 and Sources of Contamination. Sci. Total Environ. 2022, 806 Pt 2, 150562. [Google Scholar] [CrossRef] [PubMed]
- Bourli, P.; Eslahi, A.V.; Tzoraki, O.; Karanis, P. Waterborne Transmission of Protozoan Parasites: A Review of Worldwide Outbreaks - an Update 2017-2022. J. Water Health 2023, 21, 1421–1447. [Google Scholar] [CrossRef] [PubMed]
- Badri, M.; Olfatifar, M.; Karim, M.R.; Modirian, E.; Houshmand, E.; Abdoli, A.; Nikoonejad, A.; Sotoodeh, S.; Zargar, A.; Samimi, R.; Hashemipour, S.; Mahmoudi, R.; Harandi, M.F.; Hajialilo, E.; Piri, H.; Bijani, B.; Eslahi, A.V. Global Prevalence of Intestinal Protozoan Contamination in Vegetables and Fruits: A Systematic Review and Meta-Analysis. Food Control 2022, 133, 108656. [Google Scholar] [CrossRef]
- Kifleyohannes, T.; Debenham, J.J.; Robertson, L.J. Is Fresh Produce in Tigray, Ethiopia a Potential Transmission Vehicle for Cryptosporidium and Giardia? Foods 2021, 10, 1979. [Google Scholar] [CrossRef] [PubMed]
- Salamandane, C.; Lobo, M.L.; Afonso, S.; Miambo, R.; Matos, O. Occurrence of Intestinal Parasites of Public Health Significance in Fresh Horticultural Products Sold in Maputo Markets and Supermarkets, Mozambique. Microorganisms 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Trelis, M.; Sáez-Durán, S.; Puchades, P.; Castro, N.; Miquel, A.; Gozalbo, M.; Fuentes, M.V. Survey of the Occurrence of Giardia Duodenalis Cysts and Cryptosporidium Spp. Oocysts in Green Leafy Vegetables Marketed in the City of Valencia (Spain). Int. J. Food Microbiol. 2022, 379, 109847. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mesonero, L.; Soler, L.; Amorós, I.; Moreno, Y.; Ferrús, M.A.; Alonso, J.L. Protozoan Parasites and Free-Living Amoebae Contamination in Organic Leafy Green Vegetables and Strawberries from Spain. Food Waterborne Parasitol. 2023, 32, e00200. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Rana, S.; Singh, R.; Gurian, P.L.; Betancourt, W.; Kumar, A.; Kumar, A. Non-Potable Water Reuse and the Public Health Risks from Protozoa and Helminths: A Case Study from a City with a Semi-Arid Climate. J. Water Health 2023, 21, 981–994. [Google Scholar] [CrossRef]
- Ryan, U.M.; Feng, Y.; Fayer, R.; Xiao, L. Taxonomy and Molecular Epidemiology of Cryptosporidium and Giardia - a 50 Year Perspective (1971-2021). Int. J. Parasitol. 2021, 51, 1099–1119. [Google Scholar] [CrossRef]
- Ryan, U.; Zahedi, A.; Feng, Y.; Xiao, L. An Update on Zoonotic Cryptosporidium Species and Genotypes in Humans. Animals (Basel) 2021, 11, 3307. [Google Scholar] [CrossRef]
- Costa, D.; Razakandrainibe, R.; Valot, S.; Vannier, M.; Sautour, M.; Basmaciyan, L.; Gargala, G.; Viller, V.; Lemeteil, D.; French National Network on Surveillance of Human Cryptosporidiosis; Ballet, J.-J.; Dalle, F. Epidemiology of Cryptosporidiosis in France from 2017 to 2019. Microorganisms 2020, 8, 1358. [Google Scholar] [CrossRef]
- Bujila, I.; Troell, K.; Ögren, J.; Hansen, A.; Killander, G.; Agudelo, L.; Lebbad, M.; Beser, J. Cryptosporidium species and subtypes identified in human domestic cases through the national microbiological surveillance programme in Sweden from 2018 to 2022. BMC Infect Dis. 2024, 24, 146. [Google Scholar] [CrossRef]
- Ayres Hutter, J.; Dion, R.; Irace-Cima, A.; Fiset, M.; Guy, R.; Dixon, B.; Aguilar, J.L.; Trépanier, J.; Thivierge, K. Cryptosporidium Spp.: Human Incidence, Molecular Characterization and Associated Exposures in Québec, Canada (2016-2017). PLoS One 2020, 15, e0228986. [Google Scholar] [CrossRef] [PubMed]
- Guy, R.A.; Yanta, C.A.; Muchaal, P.K.; Rankin, M.A.; Thivierge, K.; Lau, R.; Boggild, A.K. Molecular Characterization of Cryptosporidium Isolates from Humans in Ontario, Canada. Parasit. Vectors 2021, 14, 69–9. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ryan, U.; Feng, Y.; Xiao, L. Emergence of Zoonotic Cryptosporidium Parvum in China. Trends Parasitol. 2022, 38, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Hijjawi, N.; Zahedi, A.; Al-Falah, M.; Ryan, U. A Review of the Molecular Epidemiology of Cryptosporidium Spp. and Giardia Duodenalis in the Middle East and North Africa (MENA) Region. Infect. Genet. Evol. 2022, 98, 105212. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, Y.; Qin, H.; Xu, H.; Zhang, L. Prevalence of Cryptosporidium Infection in Children from China: A Systematic Review and Meta-Analysis. Acta Trop. 2023, 244, 106958. [Google Scholar] [CrossRef]
- Lebbad, M.; Winiecka-Krusnell, J.; Stensvold, C.R.; Beser, J. High Diversity of Cryptosporidium Species and Subtypes Identified in Cryptosporidiosis Acquired in Sweden and Abroad. Pathogens 2021, 10, 523. [Google Scholar] [CrossRef]
- Bujila, I.; Troell, K.; Fischerström, K.; Nordahl, M.; Killander, G.; Hansen, A.; Söderlund, R.; Lebbad, M.; Beser, J. Cryptosporidium Chipmunk Genotype I - An Emerging Cause of Human Cryptosporidiosis in Sweden. Infect. Genet. Evol. 2021, 92, 104895. [Google Scholar] [CrossRef]
- Boughattas, S.; Behnke, J.M.; Al-Sadeq, D.; Ismail, A.; Abu-Madi, M. Cryptosporidium Spp., Prevalence, Molecular Characterisation and Socio-Demographic Risk Factors among Immigrants in Qatar. PLoS Negl. Trop. Dis. 2019, 13, e0007750. [Google Scholar] [CrossRef]
- Nasser, A.M. Transmission of Cryptosporidium by Fresh Vegetables. J. Food Prot. 2022, 85, 1737–1744. [Google Scholar] [CrossRef]
- Huang, W.; Guo, Y.; Lysen, C.; Wang, Y.; Tang, K.; Seabolt, M.H.; Yang, F.; Cebelinski, E.; Gonzalez-Moreno, O.; Hou, T.; Chen, C.; Chen, M.; Wan, M.; Li, N.; Hlavsa, M.C.; Roellig, D.M.; Feng, Y.; Xiao, L. Multiple Introductions and Recombination Events Underlie the Emergence of a Hyper-Transmissible Cryptosporidium Hominis Subtype in the USA. Cell Host Microbe 2023, 31, 112–123. [Google Scholar] [CrossRef]
- Robertson, L.J.; Temesgen, T.T.; Tysnes, K.R.; Eikås, J.E. An Apple a Day: An Outbreak of Cryptosporidiosis in Norway Associated with Self-Pressed Apple Juice. Epidemiol. Infect. 2019, 147, e139. [Google Scholar] [CrossRef]
- Khan, S.M.; Witola, W.H. Past, Current, and Potential Treatments for Cryptosporidiosis in Humans and Farm Animals: A Comprehensive Review. Front. Cell. Infect. Microbiol. 2023, 13, 1115522. [Google Scholar] [CrossRef]
- Carter, B.L.; Chalmers, R.M.; Davies, A.P. Health Sequelae of Human Cryptosporidiosis in Industrialised Countries: A Systematic Review. Parasit. Vectors 2020, 13, 443. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, A.; Ryan, U. Cryptosporidium - An Update with an Emphasis on Foodborne and Waterborne Transmission. Res. Vet. Sci. 2020, 132, 500–512. [Google Scholar] [CrossRef] [PubMed]
- Azami, M.; Amini Rarani, S.; Kiani, F. Treatment of Urticaria Caused by Severe Cryptosporidiosis in a 17-Month-Old Child - a Case Report. BMC Infect. Dis. 2023, 23, 461. [Google Scholar] [CrossRef] [PubMed]
- Pielok, Ł.; Nowak, S.; Kłudkowska, M.; Frąckowiak, K.; Kuszel, Ł.; Zmora, P.; Stefaniak, J. Massive Cryptosporidium Infections and Chronic Diarrhea in HIV-Negative Patients. Parasitol. Res. 2019, 118, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Ghanadi, K.; Khalaf, A.K.; Jafrasteh, A.; Anbari, K.; Mahmoudvand, H. High Prevalence of Cryptosporidium Infection in Iranian Patients Suffering from Colorectal Cancer. Parasite Epidemiol. Control 2022, 19, e00271. [Google Scholar] [CrossRef] [PubMed]
- Bouwknegt, M.; Devleesschauwer, B.; Graham, H.; Robertson, L.J.; van der Giessen, J.W.; Euro-FBP workshop participants. Prioritisation of Food-Borne Parasites in Europe, 2016. Euro Surveill. 2018, 23. [Google Scholar] [CrossRef]
- Levine, M.M.; Nasrin, D.; Acácio, S.; Bassat, Q.; Powell, H.; Tennant, S.M.; Sow, S.O.; Sur, D.; Zaidi, A.K.M.; Faruque, A.S.G.; Hossain, M.J.; Alonso, P.L.; Breiman, R.F.; O’Reilly, C.E.; Mintz, E.D.; Omore, R.; Ochieng, J.B.; Oundo, J.O.; Tamboura, B.; Sanogo, D.; Onwuchekwa, U.; Manna, B.; Ramamurthy, T.; Kanungo, S.; Ahmed, S.; Qureshi, S.; Quadri, F.; Hossain, A.; Das, S.K.; Antonio, M.; Saha, D.; Mandomando, I.; Blackwelder, W.C.; Farag, T.; Wu, Y.; Houpt, E.R.; Verweiij, J.J.; Sommerfelt, H.; Nataro, J.P.; Robins-Browne, R.M.; Kotloff, K.L. Diarrhoeal Disease and Subsequent Risk of Death in Infants and Children Residing in Low-Income and Middle-Income Countries: Analysis of the GEMS Case-Control Study and 12-Month GEMS-1A Follow-on Study. Lancet Glob. Health 2020, 8, e204–e214. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Karanis, P. Cryptosporidium and Cryptosporidiosis: The Perspective from the Gulf Countries. Int. J. Environ. Res. Public Health 2020, 17, 6824. [Google Scholar] [CrossRef]
- Korpe, P.; Ni, Z.; Kabir, M.; Alam, M.; Ferdous, T.; Ara, R.; Munday, R.M.; Haque, R.; Duggal, P. Prospective Cohort Study of Cryptosporidium Infection and Shedding in Infants and Their Households. Clin. Infect. Dis. 2023, 76, 2178–2186. [Google Scholar] [CrossRef]
- Hossain, M.J.; Saha, D.; Antonio, M.; Nasrin, D.; Blackwelder, W.C.; Ikumapayi, U.N.; Mackenzie, G.A.; Adeyemi, M.; Jasseh, M.; Adegbola, R.A.; Roose, A.W.; Kotloff, K.L.; Levine, M.M. Cryptosporidium Infection in Rural Gambian Children: Epidemiology and Risk Factors. PLoS Negl. Trop. Dis. 2019, 13, e0007607. [Google Scholar] [CrossRef]
- Johansen, Ø.H.; Abdissa, A.; Zangenberg, M.; Mekonnen, Z.; Eshetu, B.; Sharew, B.; Moyo, S.; Sommerfelt, H.; Langeland, N.; Robertson, L.J.; Hanevik, K. A Comparison of Risk Factors for Cryptosporidiosis and Non-Cryptosporidiosis Diarrhoea: A Case-Case-Control Study in Ethiopian Children. PLoS Negl. Trop. Dis. 2022, 16, e0010508. [Google Scholar] [CrossRef]
- Alali, F.; Abbas, I.; Jawad, M.; Hijjawi, N. Cryptosporidium Infection in Humans and Animals from Iraq: A Review. Acta Trop. 2021, 220, 105946. [Google Scholar] [CrossRef] [PubMed]
- Nic Lochlainn, L.M.; Sane, J.; Schimmer, B.; Mooij, S.; Roelfsema, J.; van Pelt, W.; Kortbeek, T. Risk Factors for Sporadic Cryptosporidiosis in the Netherlands: Analysis of a 3-Year Population Based Case-Control Study Coupled with Genotyping, 2013-2016. J. Infect. Dis. 2019, 219, 1121–1129. [Google Scholar] [CrossRef]
- Watier-Grillot, S.; Costa, D.; Petit, C.; Razakandrainibe, R.; Larréché, S.; Tong, C.; Demont, G.; Billetorte, D.; Mouly, D.; Fontan, D.; Velut, G.; Le Corre, A.; Beauvir, J.-C.; Mérens, A.; Favennec, L.; Pommier de Santi, V. Cryptosporidiosis Outbreaks Linked to the Public Water Supply in a Military Camp, France. PLoS Negl. Trop. Dis. 2022, 16, e0010776. [Google Scholar] [CrossRef]
- Franceschelli, A.; Bonadonna, L.; Cacciò, S.M.; Sannella, A.R.; Cintori, C.; Gargiulo, R.; Coccia, A.M.; Paradiso, R.; Iaconelli, M.; Briancesco, R.; Tripodi, A. An Outbreak of Cryptosporidiosis Associated with Drinking Water in North-Eastern Italy, August 2019: Microbiological and Environmental Investigations. Euro Surveill. 2022, 27. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Gong, B.; Liu, X.; Shen, Y.; Wu, Y.; Zhang, W.; Cao, J. A Retrospective Epidemiological Analysis of Human Cryptosporidium Infection in China during the Past Three Decades (1987-2018). PLoS Negl. Trop. Dis. 2020, 14, e0008146. [Google Scholar] [CrossRef] [PubMed]
- Stokdyk, J.P.; Spencer, S.K.; Walsh, J.F.; de Lamber, J.R.; Firnstahl, A.D.; Anderson, A.C.; Rezania, L.I.W.; Borchardt, M.A. Cryptosporidium incidence and surface water influence of groundwater supplying public water systems in Minnesota, USA. Environ. Sci. Technol. 2019, 53, 3391–3398. [Google Scholar] [CrossRef]
- Kooh, P.; Thébault, A.; Cadavez, V.; Gonzales-Barron, U.; Villena, I. Risk Factors for Sporadic Cryptosporidiosis: A Systematic Review and Meta-Analysis. Microb. Risk Anal. 2021, 17, 100116. [Google Scholar] [CrossRef]
- Ursini, T.; Moro, L.; Requena-Méndez, A.; Bertoli, G.; Buonfrate, D. A Review of Outbreaks of Cryptosporidiosis Due to Unpasteurized Milk. Infection 2020, 48, 659–663. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion. The efficacy and safety of high-pressure processing of food. EFSA Journal 2022. https://doi.org/10.2903/j.efsa.2022.7128. Available online: https://www.efsa.europa.eu/sites/default/files/2022-03/7128.pdf (accessed on 27 February 2024).
- Gopfert, A.; Chalmers, R.M.; Whittingham, S.; Wilson, L.; van Hove, M.; Ferraro, C.F.; Robinson, G.; Young, N.; Nozad, B. An Outbreak of Cryptosporidium Parvum Linked to Pasteurised Milk from a Vending Machine in England: A Descriptive Study, March 2021. Epidemiol. Infect. 2022, 150, e185. [Google Scholar] [CrossRef]
- Ahlinder, J.; Svedberg, A.-L.; Nystedt, A.; Dryselius, R.; Jacobsson, K.; Hägglund, M.; Brindefalk, B.; Forsman, M.; Ottoson, J.; Troell, K. Use of Metagenomic Microbial Source Tracking to Investigate the Source of a Foodborne Outbreak of Cryptosporidiosis. Food Waterborne Parasitol. 2022, 26, e00142. [Google Scholar] [CrossRef] [PubMed]
- Dankwa, K.; Nuvor, S.V.; Obiri-Yeboah, D.; Feglo, P.K.; Mutocheluh, M. Occurrence of Cryptosporidium Infection and Associated Risk Factors among HIV-Infected Patients Attending ART Clinics in the Central Region of Ghana. Trop. Med. Infect. Dis. 2021, 6, 210. [Google Scholar] [CrossRef] [PubMed]
- Certad, G.; Follet, J.; Gantois, N.; Hammouma-Ghelboun, O.; Guyot, K.; Benamrouz-Vanneste, S.; Fréalle, E.; Seesao, Y.; Delaire, B.; Creusy, C.; Even, G.; Verrez-Bagnis, V.; Ryan, U.; Gay, M.; Aliouat-Denis, C.; Viscogliosi, E. Prevalence, Molecular Identification, and Risk Factors for Cryptosporidium Infection in Edible Marine Fish: A Survey across Sea Areas Surrounding France. Front. Microbiol. 2019, 10, 1037. [Google Scholar] [CrossRef]
- Srisuphanunt, M.; Wilairatana, P.; Kooltheat, N.; Damrongwatanapokin, T.; Karanis, P. Occurrence of Cryptosporidium Oocysts in Commercial Oysters in Southern Thailand. Food Waterborne Parasitol. 2023, 32, e00205. [Google Scholar] [CrossRef]
- Moratal, S.; Dea-Ayuela, M.A.; Martí-Marco, A.; Puigcercós, S.; Marco-Hirs, N.M.; Doménech, C.; Corcuera, E.; Cardells, J.; Lizana, V.; López-Ramon, J. Molecular Characterization of Cryptosporidium Spp. In Cultivated and Wild Marine Fishes from Western Mediterranean with the First Detection of Zoonotic Cryptosporidium Ubiquitum. Animals (Basel) 2022, 12, 1052. [Google Scholar] [CrossRef]
- Servián, A.; Lorena Zonta, M.; Navone, G.T. Differential Diagnosis of Human Entamoeba Infections: Morphological and Molecular Characterization of New Isolates in Argentina. Rev. Argent. Microbiol. 2023. [Google Scholar] [CrossRef]
- Balarabe-Musa, B.; Onyeagba, K.D. Prevalence of Entamoeba Histolytica amongst Infants with Diarrhoea Visiting the University of Abuja Teaching Hospital. Journal of Veterinary and Biomedical Sciences 2020, 2, 128–134. [Google Scholar] [CrossRef]
- Leon-Coria, A.; Kumar, M.; Chadee, K. The Delicate Balance between Entamoeba Histolytica, Mucus and Microbiota. Gut Microbes 2020, 11, 118–125. [Google Scholar] [CrossRef]
- Singh, A.; Banerjee, T. Host-Parasite Interactions in Infections Due to Entamoeba Histolytica: A Tale of Known and Unknown. Trop. Parasitol. 2022, 12, 69–77. [Google Scholar] [PubMed]
- Morán, P.; Serrano-Vázquez, A.; Rojas-Velázquez, L.; González, E.; Pérez-Juárez, H.; Hernández, E.G.; Padilla, M. de L.A.; Zaragoza, M.E.; Portillo-Bobadilla, T.; Ramiro, M.; Ximénez, C. Amoebiasis: Advances in Diagnosis, Treatment, Immunology Features and the Interaction with the Intestinal Ecosystem. Int. J. Mol. Sci. 2023, 24, 11755. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.-H.; Chen, B.-C.; Chou, Y.-C.; Chien, W.-C.; Chung, C.-H.; Hsieh, C.-J.; Yu, C.-P. The Epidemiology of Entamoeba histolytica Infection and Its Associated Risk Factors among Domestic and Imported Patients in Taiwan during the 2011–2020 Period. Medicina 2022, 58, 820. [Google Scholar] [CrossRef]
- Tahir, M.; Sendzischew Shane, M.; Deshpande Amar, R.; Milikowski, C. An Unusual Case of Entamoeba Histolytica Infection in Miami Fl: Case Report and Review of the Literature. Res. Rev. Infect. Dis. 2020, 3. [Google Scholar] [CrossRef]
- Samie, A.; Mahlaule, L.; Mbati, P.; Nozaki, T.; ElBakri, A. Prevalence and Distribution of Entamoeba Species in a Rural Community in Northern South Africa. Food Waterborne Parasitol. 2020, 18, e00076. [Google Scholar] [CrossRef]
- Wang, H.; Kanthan, R. Multiple Colonic and Ileal Perforations Due to Unsuspected Intestinal Amoebiasis-Case Report and Review. Pathol. Res. Pract. 2020, 216, 152608. [Google Scholar] [CrossRef]
- Usuda, D.; Tsuge, S.; Sakurai, R.; Kawai, K.; Matsubara, S.; Tanaka, R.; Suzuki, M.; Takano, H.; Shimozawa, S.; Hotchi, Y.; Tokunaga, S.; Osugi, I.; Katou, R.; Ito, S.; Mishima, K.; Kondo, A.; Mizuno, K.; Takami, H.; Komatsu, T.; Oba, J.; Nomura, T.; Sugita, M. Amebic Liver Abscess by Entamoeba Histolytica. World J. Clin. Cases 2022, 10, 13157–13166. [Google Scholar] [CrossRef] [PubMed]
- Carrero, J.C.; Reyes-López, M.; Serrano-Luna, J.; Shibayama, M.; Unzueta, J.; León-Sicairos, N.; de la Garza, M. Intestinal Amoebiasis: 160 Years of Its First Detection and Still Remains as a Health Problem in Developing Countries. Int. J. Med. Microbiol. 2020, 310, 151358. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Wiehe, R.; Dommisch, H.; Schaefer, A.S. Entamoeba Gingivalis Causes Oral Inflammation and Tissue Destruction. J. Dent. Res. 2020, 99, 561–567. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Guidelines for drinking-water quality. Fourth edition incorporating the first and second addenda. 2022 ISBN 978-92-4-004506-4. Available online: https://iris.who.int/bitstream/handle/10665/352532/9789240045064-eng.pdf?sequence=1&isAllowed=y#page=153 (accessed on 15 February 2024).
- Fichardt, H.J.; Rocha, J. S2741 Entamoeba Histolytica Dysentery Complicated by Amebic Liver Abscess and Type a Right Heart Thrombus: A Case Report. Am. J. Gastroenterol. 2020, 115, S1432–S1432. [Google Scholar] [CrossRef]
- Kaiser, R.W.J.; Allgeier, J.; Philipp, A.B.; Mayerle, J.; Rothe, C.; Wallrauch, C.; Op den Winkel, M. Development of Amoebic Liver Abscess in Early Pregnancy Years after Initial Amoebic Exposure: A Case Report. BMC Gastroenterol. 2020, 20, 424. [Google Scholar] [CrossRef] [PubMed]
- Flaih, M.H.; Khazaal, R.M.; Kadhim, M.K.; Hussein, K.R.; Alhamadani, F.A.B. The Epidemiology of Amoebiasis in Thi-Qar Province, Iraq (2015-2020): Differentiation of Entamoeba Histolytica and Entamoeba Dispar Using Nested and Real-Time Polymerase Chain Reaction. Epidemiol. Health 2021, 43, e2021034. [Google Scholar] [CrossRef] [PubMed]
- Aziz, K.T.; Rostam, R.A.; Nasir, K. ِِ.; Qasem, A.O.; Azize, P.M. Epidemiological Study for Entamoeba Histolytica Infection and Their Risk Factors among Children in Pediatric Hospital in, Sulaimani Province-Iraq. Kufa Jour. Nurs. Sci. 2022, 12. [Google Scholar]
- Al-Daoody, A.A.K.; A. Ali, F.; Sadiq, L.B.; Samer Mamand, A.; Salah Ismail, R.; Fatah Muhammed, H. Investigation of Intestinal Protozoan Infections among Food-Handlers in Erbil City, Iraq. Plant Arch. 2021, 21, 1367–1372. [Google Scholar] [CrossRef]
- Muhammad Bunza, N.; Sale Kumurya, A.; Muhammad, A. Prevalence and Associated Risk Factors of Intestinal Parasitic Infections among Food Handlers in Kano Metropolis, Kano State, Nigeria. Microbes and Infectious Diseases 2020, 0, 0–0. [Google Scholar] [CrossRef]
- Dos Santos Zanetti, A.; Malheiros, A.F.; de Matos, T.A.; Dos Santos, C.; Battaglini, P.F.; Moreira, L.M.; Lemos, L.M.S.; Castrillon, S.K.I.; da Costa Boamorte Cortela, D.; Ignotti, E.; Espinosa, O.A. Diversity, Geographical Distribution, and Prevalence of Entamoeba Spp. in Brazil: A Systematic Review and Meta-Analysis. Parasite 2021, 28, 17. [Google Scholar] [CrossRef]
- Roro, G.B.; Eriso, F.; Al-Hazimi, A.M.; Kuddus, M.; Singh, S.C.; Upadhye, V.; Hajare, S.T. Prevalence and Associated Risk Factors of Entamoeba Histolytica Infection among School Children from Three Primary Schools in Arsi Town, West Zone, Ethiopia. J. Parasit. Dis. 2022, 46, 776–784. [Google Scholar] [CrossRef]
- Tokijoh, N.I.; Bakar, A.A.; Othman, N.; Noordin, R.; Saidin, S. Assessing the Prevalence and Risk Factors Associated with Entamoeba Complex Infection among the Orang Asli School Children in Perak, Malaysia through Molecular Approach. Parasitol. Int. 2022, 91, 102638. [Google Scholar] [CrossRef]
- Atabati, H.; Kassiri, H.; Shamloo, E.; Akbari, M.; Atamaleki, A.; Sahlabadi, F.; Linh, N.T.T.; Rostami, A.; Fakhri, Y.; Khaneghah, A.M. The Association between the Lack of Safe Drinking Water and Sanitation Facilities with Intestinal Entamoeba Spp Infection Risk: A Systematic Review and Meta-Analysis. PLoS One 2020, 15, e0237102. [Google Scholar] [CrossRef]
- Al-Yousofi, A.; Yan, Y.; Al Mekhlafi, A.M.; Hezam, K.; Abouelnazar, F.A.; Al-Rateb, B.; Almamary, H.; Abdulwase, R. Prevalence of Intestinal Parasites among Immunocompromised Patients, Children, and Adults in Sana’a, Yemen. J. Trop. Med. 2022, 2022, 5976640. [Google Scholar] [CrossRef]
- Azizi, H.; Davtalab-Esmaeili, E.; Mirzapour, M.; Karimi, G.; Rostampour, M.; Mirzaei, Y. A Case-Control Study of Timely Control and Investigation of an Entamoeba Histolytica Outbreak by Primary Health Care in Idahluy-e Bozorg Village, Iran. Int. J. Epidemiol. Res. 2019, 6, 120–127. [Google Scholar] [CrossRef]
- Chowdhury, R.; Esiobu, N.; Meeroff, D.; Bloetscher, F. Different Detection and Treatment Methods for Entamoeba histolytica and Entamoeba dispar in Water/Wastewater: A Review. Journal of Environmental Protection, 2022, 13, 126–149. [Google Scholar] [CrossRef]
- Gisbert Algaba, I.; Verhaegen, B.; Murat, J.B.; Coucke, W.; Mercier, A.; Cox, E.; Dorny, P.; Dierick, K.; De Craeye, S. Molecular Study of Toxoplasma Gondii Isolates Originating from Humans and Organic Pigs in Belgium. Foodborne Pathog. Dis. 2020, 17, 316–321. [Google Scholar] [CrossRef]
- Cerutti, A.; Blanchard, N.; Besteiro, S. The Bradyzoite: A Key Developmental Stage for the Persistence and Pathogenesis of Toxoplasmosis. Pathogens 2020, 9, 234. [Google Scholar] [CrossRef]
- Dakroub, H.; Sgroi, G.; D’Alessio, N.; Russo, D.; Serra, F.; Veneziano, V.; Rea, S.; Pucciarelli, A.; Lucibelli, M.G.; De Carlo, E.; Fusco, G.; Amoroso, M.G. Molecular Survey of Toxoplasma Gondii in Wild Mammals of Southern Italy. Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Ducrocq, J.; Simon, A.; Lemire, M.; De Serres, G.; Lévesque, B. Exposure to Toxoplasma Gondii through Consumption of Raw or Undercooked Meat: A Systematic Review and Meta-Analysis. Vector Borne Zoonotic Dis. 2021, 21, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Hoshina, T.; Horino, T.; Saiki, E.; Aonuma, H.; Sawaki, K.; Miyajima, M.; Lee, K.; Nakaharai, K.; Shimizu, A.; Hosaka, Y.; Kato, T.; Sato, F.; Nakazawa, Y.; Yoshikawa, K.; Yoshida, M.; Hori, S.; Kanuka, H. Seroprevalence and Associated Factors of Toxoplasma Gondii among HIV-Infected Patients in Tokyo: A Cross Sectional Study. J. Infect. Chemother. 2020, 26, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Cabral Monica, T.; Pinto-Ferreira, F.; Martins, F.D.C.; de Matos, R.L.N.; de Matos, A.M.R.N.; Santos, A.C.; Nino, B. de S. L.; Pereira, L.; Narciso, S.G.; Garcia, J.L.; Freire, R.L.; Navarro, I.T.; Mitsuka-Bregano, R. Epidemiology of a Toxoplasmosis Outbreak in a Research Institution in Northern Paraná, Brazil. Zoonoses Public Health 2020, 67, 760–764. [Google Scholar] [CrossRef] [PubMed]
- da Costa, M.A.; Pinto-Ferreira, F.; de Almeida, R.P.A.; Martins, F.D.C.; Pires, A.L.; Mareze, M.; Mitsuka-Breganó, R.; Freire, R.L.; da Rocha Moreira, R.V.; Borges, J.M.; Navarro, I.T. Artisan Fresh Cheese from Raw Cow’s Milk as a Possible Route of Transmission in a Toxoplasmosis Outbreak, in Brazil. Zoonoses Public Health 2020, 67, 122–129. [Google Scholar] [CrossRef]
- Minuzzi, C.E.; Fernandes, F.D.; Portella, L.P.; Bräunig, P.; Sturza, D.A.F.; Giacomini, L.; Salvagni, E.; Ribeiro, J.D.S.; Silva, C.R.; Difante, C.M.; Farinha, L.B.; Menegolla, I.A.; Gehrke, G.; Dilkin, P.; Sangioni, L.A.; Mallmann, C.A.; Vogel, F.S.F. Contaminated Water Confirmed as Source of Infection by Bioassay in an Outbreak of Toxoplasmosis in South Brazil. Transbound. Emerg. Dis. 2021, 68, 767–772. [Google Scholar] [CrossRef]
- Schumacher, A.C.; Elbadawi, L.I.; DeSalvo, T.; Straily, A.; Ajzenberg, D.; Letzer, D.; Moldenhauer, E.; Handly, T.L.; Hill, D.; Dardé, M.-L.; Pomares, C.; Passebosc-Faure, K.; Bisgard, K.; Gomez, C.A.; Press, C.; Smiley, S.; Montoya, J.G.; Kazmierczak, J.J. Toxoplasmosis Outbreak Associated with Toxoplasma Gondii-Contaminated Venison-High Attack Rate, Unusual Clinical Presentation, and Atypical Genotype. Clin. Infect. Dis. 2021, 72, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Murata, R.; Kodo, Y.; Maeno, A.; Suzuki, J.; Mori, K.; Sadamasu, K.; Kawahara, F.; Nagamune, K. Detection of Toxoplasma Gondii and Sarcocystis Sp. in the Meat of Common Minke Whale (Balaenoptera Acutorostrata): A Case of Suspected Food Poisoning in Japan. Parasitol. Int. 2023, 99, 102832. [Google Scholar] [CrossRef]
- Marangi, M.; Lago, N.; Mancinelli, G.; Antonio, O.L.; Scirocco, T.; Sinigaglia, M.; Specchiulli, A.; Cilenti, L. Occurrence of the protozoan parasites Toxoplasma gondii and Cyclospora cayetanensis in the invasive Atlantic blue crab Callinectes sapidus from the Lesina Lagoon (SE Italy). Mar. Pollut. Bull. 2022, 176, 113428. [Google Scholar] [CrossRef]
- Çulbasan, V.; Gungor, C.; Gundog, D.A.; Koskeroglu, K.; Onmaz, N.E. Molecular surveillance of Toxoplasma gondii in raw milk and Artisan cheese of sheep, goat, cow and water buffalo origin. Int J Dairy Technol, 2023, 76, 948–954. [Google Scholar] [CrossRef]
- Ogbuigwe, P.; Biggs, P.J.; Garcia-Ramirez, J.C.; Knox, M.A.; Pita, A.; Velathanthiri, N.; French, N.P.; Hayman, D.T.S. Uncovering the Genetic Diversity of Giardia Intestinalis in Isolates from Outbreaks in New Zealand. Infect. Dis. Poverty 2022, 11, 49. [Google Scholar] [CrossRef]
- Ryan, U.; Hijjawi, N.; Feng, Y.; Xiao, L. Giardia: An under-Reported Foodborne Parasite. Int. J. Parasitol. 2019, 49, 1–11. [Google Scholar] [CrossRef]
- Resi, D.; Varani, S.; Sannella, A.R.; De Pascali, A.M.; Ortalli, M.; Liguori, G.; Benvenuti, M.; Re, M.C.; Pirani, R.; Prete, L.; Mazzetti, C.; Musti, M.; Pizzi, L.; Sanna, T.; Cacciò, S.M. A Large Outbreak of Giardiasis in a Municipality of the Bologna Province, North-Eastern Italy, November 2018 to April 2019. Euro Surveill. 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Fakhri, Y.; Daraei, H.; Ghaffari, H.R.; Rezapour-Nasrabad, R.; Soleimani-Ahmadi, M.; Khedher, K.M.; Rostami, A.; Thai, V.N. The Risk Factors for Intestinal Giardia Spp Infection: Global Systematic Review and Meta-Analysis and Meta-Regression. Acta Trop. 2021, 220, 105968. [Google Scholar] [CrossRef] [PubMed]
- Hoseinzadeh, E.; Rostamian, A.; Razaghi, M.; Wei, C. Waterborne Transmission of Protozoan Parasites: A Review of Water Resources in Iran - an Update 2020. Desalination And Water Treatment 2021, 213, 91–105. [Google Scholar] [CrossRef]
- Wadi, W.F.; Rathi, M.H.; Molan, A.-L. The Possible Link between Intestinal Parasites and Irritable Bowel Syndrome (IBS) in Diyala Province, Iraq. Ann. Parasitol. 2021, 67, 505–513. [Google Scholar] [PubMed]
- Thébault, A.; Favennec, L.; Kooh, P.; Cadavez, V.; Gonzales-Barron, U.; Villena, I. Risk Factors for Sporadic Giardiasis: A Systematic Review and Meta-Analysis. Microb. Risk Anal. 2021, 17, 100158. [Google Scholar] [CrossRef]
- Abedi, S.H.; Fazlzadeh, A.; Mollalo, A.; Sartip, B.; Mahjour, S.; Bahadory, S.; Taghipour, A.; Rostami, A. The Neglected Role of Blastocystis Sp. and Giardia Lamblia in Development of Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis. Microb. Pathog. 2022, 162, 105215. [Google Scholar] [CrossRef] [PubMed]
- Shaima, S.N.; Das, S.K.; Ahmed, S.; Jahan, Y.; Khan, S.H.; Mamun, G.M.S.; Shahid, A.S.M.S.B.; Parvin, I.; Ahmed, T.; Faruque, A.S.G.; Chisti, M.J. Anthropometric Indices of Giardia-Infected under-Five Children Presenting with Moderate-to-Severe Diarrhea and Their Healthy Community Controls: Data from the Global Enteric Multicenter Study. Children (Basel) 2021, 8, 1186. [Google Scholar] [CrossRef] [PubMed]
- Groudan, K.; Gupta, K.; Chalhoub, J.; Singhania, R. Giardia Lamblia Diagnosed Incidentally by Duodenal Biopsy. J. Investig. Med. High Impact Case Rep. 2021, 9, 23247096211001649. [Google Scholar] [CrossRef]
- Hurník, P.; Žiak, D.; Dluhošová, J.; Židlík, V.; Šustíková, J.; Uvírová, M.; Urban, O.; Dvořáčková, J.; Nohýnková, E. Another Case of Coincidental Giardia Infection and Pancreatic Cancer. Parasitol. Int. 2019, 71, 160–162. [Google Scholar] [CrossRef]
- Loderstädt, U.; Frickmann, H. Antimicrobial Resistance of the Enteric Protozoon Giardia Duodenalis - A Narrative Review. Eur. J. Microbiol. Immunol. (Bp.) 2021, 11, 29–43. [Google Scholar] [CrossRef]
- European Commission. Commission Implementing Decision (EU) 2018/945 of 22 June 2018 on the communicable diseases and related special health issues to be covered by epidemiological surveillance as well as relevant case definitions (Text with EEA relevance.). Official Journal of the European Union. 2018 L 170/1. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=uriserv%3AOJ.L_.2018.170.01.0001.01.ENG (accessed on 16 January 2024).
- European Centre for Disease Prevention and Control (ECDC) Giardiasis (lambliasis) - Annual Epidemiological Report for 2019. 2022. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/giardiasis-%20annual-epidemiological-report-2019_0.pdf (accessed on 22 February 2024).
- Al-Jawabreh, A.; Ereqat, S.; Dumaidi, K.; Al-Jawabreh, H.; Abdeen, Z.; Nasereddin, A. Prevalence of Selected Intestinal Protozoan Infections in Marginalized Rural Communities in Palestine. BMC Public Health 2019, 19, 1667. [Google Scholar] [CrossRef]
- Fusaro, C.; Chávez-Romero, Y.A.; Prada, S.L.G.; Serrano-Silva, N.; Bernal, J.E.; González-Jiménez, F.E.; Sarria-Guzmán, Y. Burden and Epidemiology of Human Intestinal Giardia Duodenalis Infection in Colombia: A Systematic Review. Trop. Med. Infect. Dis. 2022, 7. [Google Scholar] [CrossRef]
- Bekele, D.; Reta, T. Prevalence and distribution of Giardia lamblia infection among patients of children aged 5-11years treated in Sheno town health centers, North Shoa Oromia region, Ethiopia. ClinicSearch 2023. [Google Scholar] [CrossRef]
- Tembo, S.J.; Mutengo, M.M.; Sitali, L.; Changula, K.; Takada, A.; Mweene, A.S.; Simulundu, E.; Chitanga, S. Prevalence and Genotypic Characterization of Giardia Duodenalis Isolates from Asymptomatic School-Going Children in Lusaka, Zambia. Food Waterborne Parasitol. 2020, 19, e00072. [Google Scholar] [CrossRef] [PubMed]
- Muadica, A.S.; Balasegaram, S.; Beebeejaun, K.; Köster, P.C.; Bailo, B.; Hernández-de-Mingo, M.; Dashti, A.; Dacal, E.; Saugar, J.M.; Fuentes, I.; Carmena, D. Risk Associations for Intestinal Parasites in Symptomatic and Asymptomatic Schoolchildren in Central Mozambique. Clin. Microbiol. Infect. 2021, 27, 624–629. [Google Scholar] [CrossRef] [PubMed]
- Dixon, B.R. Giardia Duodenalis in Humans and Animals - Transmission and Disease. Res. Vet. Sci. 2021, 135, 283–289. [Google Scholar] [CrossRef]
- Tawana, M.; Onyiche, T.E.; Ramatla, T.; Thekisoe, O. A “One Health” Perspective of Africa-Wide Distribution and Prevalence of Giardia Species in Humans, Animals and Waterbodies: A Systematic Review and Meta-Analysis. Parasitology 2023, 150, 769–780. [Google Scholar] [CrossRef]
- Bruneto, E.G.; Fernandes-Silva, M.M.; Toledo-Cornell, C.; Martins, S.; Ferreira, J.M.B.; Corrêa, V.R.; da Costa, J.M.; Pinto, A.Y. das N.; de Souza, D. do S. M.; Pinto, M.C.G.; Neto, J.A. de F.; Ramos, A.N.; Maguire, J.H.; Silvestre, O.M. Case-Fatality from Orally-Transmitted Acute Chagas Disease: A Systematic Review and Meta-Analysis. Clin. Infect. Dis. 2021, 72, 1084–1092. [Google Scholar] [CrossRef]
- Velásquez-Ortiz, N.; Ramírez, J.D. Understanding the Oral Transmission of Trypanosoma Cruzi as a Veterinary and Medical Foodborne Zoonosis. Res. Vet. Sci. 2020, 132, 448–461. [Google Scholar] [CrossRef]
- Brito, A.K.S.B. de; Sousa, D.R.T. de; Silva Junior, E.F. da; Ruiz, H.J. da S.; Arcanjo, A.R.L.; Ortiz, J.V.; Brito, S.S. de; Jesus, D.V.; Lima, J.R.C. de; Couceiro, K. do N.; Silva, M.R.H. da S. E.; Ferreira, J.M.B.B.; Guerra, J.A.O.; Guerra, M. das G. V. B. Acute Micro-Outbreak of Chagas Disease in the Southeastern Amazon: A Report of Five Cases. Rev. Soc. Bras. Med. Trop. 2022, 55, e0687. [Google Scholar]
- Esper, H.R.; Freitas, V.L.T. de; Assy, J.G.P.L.; Shimoda, E.Y.; Berreta, O.C.P.; Lopes, M.H.; França, F.O.S. Fatal Evolution of Acute Chagas Disease in a Child from Northern Brazil: Factors That Determine Poor Prognosis. Rev. Inst. Med. Trop. Sao Paulo 2019, 61, e27. [Google Scholar] [CrossRef]
- Velásquez-Ortiz, N.; Herrera, G.; Hernández, C.; Muñoz, M.; Ramírez, J.D. Discrete Typing Units of Trypanosoma Cruzi: Geographical and Biological Distribution in the Americas. Sci. Data 2022, 9, 360. [Google Scholar] [CrossRef]
- De Oliveira, A.C.; Soccol, V.T.; Rogez, H. Prevention Methods of Foodborne Chagas Disease: Disinfection, Heat Treatment and Quality Control by RT-PCR. Int. J. Food Microbiol. 2019, 301, 34–40. [Google Scholar] [CrossRef]
- Mathison, B.A.; Pritt, B.S. Cyclosporiasis-Updates on Clinical Presentation, Pathology, Clinical Diagnosis, and Treatment. Microorganisms 2021, 9, 1863. [Google Scholar] [CrossRef]
- Barratt, J.L.N.; Shen, J.; Houghton, K.; Richins, T.; Sapp, S.G.H.; Cama, V.; Arrowood, M.J.; Straily, A.; Qvarnstrom, Y. Cyclospora Cayetanensis Comprises at Least 3 Species That Cause Human Cyclosporiasis. Parasitology 2023, 150, 269–285. [Google Scholar] [CrossRef]
- Lugo, R.; Angulo-Várguez, F.; Ávila-Nava, A.; Gutiérrez-Solis, A.L.; Reyes-Sosa, M.; Medina-Escobedo, M. Acute Kidney Injury Associated with Intestinal Infection by Cyclospora Cayetanensis in a Kidney Transplant Patient. A Case Report. Parasitol. Int. 2021, 80, 102212. [Google Scholar] [CrossRef]
- Einhorn, N.; Lamphier, I.; Klinkova, O.; Baluch, A.; Pasikhova, Y.; Greene, J. Intestinal Coccidian Infections in Cancer Patients: A Case Series. Cureus 2023, 15, e38256. [Google Scholar] [CrossRef] [PubMed]
- Almeria, S.; Chacin-Bonilla, L.; Maloney, J.G.; Santin, M. Cyclospora Cayetanensis: A Perspective (2020-2023) with Emphasis on Epidemiology and Detection Methods. Microorganisms 2023, 11. [Google Scholar] [CrossRef]
- Chacin-Bonilla, L.; Santin, M. Cyclospora Cayetanensis Infection in Developed Countries: Potential Endemic Foci? Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Ahart, L.; Jacobson, D.; Rice, M.; Richins, T.; Peterson, A.; Zheng, Y.; Barratt, J.; Cama, V.; Qvarnstrom, Y.; Montgomery, S.; Straily, A. Retrospective Evaluation of an Integrated Molecular-Epidemiological Approach to Cyclosporiasis Outbreak Investigations - United States, 2021. Epidemiol. Infect. 2023, 151, e131. [Google Scholar] [CrossRef]
- Ramezanzadeh, S.; Barzegar, G.; Osquee, H.O.; Pirestani, M.; Mahami-Oskouei, M.; Hajizadeh, M.; Hosseini, S.A.; Rodrigues Oliveira, S.M.; Agholi, M.; de Lourdes Pereira, M.; Ahmadpour, E. Microscopic and Molecular Identification of Cyclospora Cayetanensis and Cystoisospora Belli in HIV-Infected People in Tabriz, Northwest of Iran. Trop. Med. Infect. Dis. 2023, 8. [Google Scholar] [CrossRef] [PubMed]
- Ögren, J.; Dienus, O.; Beser, J.; Henningsson, A.J.; Matussek, A. Protozoan Infections Are Under-Recognized in Swedish Patients with Gastrointestinal Symptoms. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2153–2160. [Google Scholar] [CrossRef]
- Chacin-Bonilla, L.; Sanchez, Y.; Cardenas, R. Factors Associated with Cyclospora Infection in a Venezuelan Community: Extreme Poverty and Soil Transmission Relate to Cyclosporiasis. Trans. R. Soc. Trop. Med. Hyg. 2023, 117, 83–90. [Google Scholar] [CrossRef]
- Barlaam, A.; Temesgen, T.T.; Tysnes, K.R.; Rinaldi, L.; Ferrari, N.; Sannella, A.R.; Normanno, G.; Cacciò, S.M.; Robertson, L.J.; Giangaspero, A. Contamination of Fresh Produce Sold on the Italian Market with Cyclospora Cayetanensis and Echinococcus Multilocularis. Food Microbiol. 2021, 98, 103792. [Google Scholar] [CrossRef] [PubMed]
- Temesgen, T.T.; Stigum, V.M.; Robertson, L.J. Surveillance of berries sold on the Norwegian market for parasite contamination using molecular methods. Food Microbiol. 2022, 104, 103980. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, L.; Oakley, J.; Fries, P. Verification and Use of the US-FDA BAM 19b Method for Detection of Cyclospora cayetanensis in a Survey of Fresh Produce by CFIA Laboratory. Microorganisms 2022, 10, 559. [Google Scholar] [CrossRef] [PubMed]
- Almeria, S.; Shipley, A. Detection of Cyclospora cayetanensis on bagged pre-cut salad mixes within their shelf-life and after sell by date by the U.S. food and drug administration validated method. Food Microbiol. 2021, 98, 103802. [Google Scholar] [CrossRef] [PubMed]
- Giarratana, F.; Nalbone, L.; Napoli, E.; Lanzo, V.; Panebianco, A. Prevalence of Balantidium coli (Malmsten, 1857) Infection in Swine Reared in South Italy: A Widespread Neglected Zoonosis. Vet. World 2021, 14, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Almaw, A.; Berhan, A.; Solomon, Y.; Malkamu, B.; Eyayu, T.; Workineh, L.; Mekete, G.; Yayehrad, A.T. Balantidium coli; Rare and Accidental Finding in the Urine of Pregnant Woman: Case Report. Int. Med. Case Rep. J. 2022, 15, 105–109. [Google Scholar] [CrossRef]
- Kennedy Souza Araújo, R.; Rios da Silva, A.; Souza, N.R.; Santos de Melo, K. Parasitological Analysis of Vegetables Sold in Supermarkets and Free Markets in the City of Taguatinga, Federal District, Brazil. Rev. Patol. Trop. 2022, 51, 225–234. [Google Scholar] [CrossRef]
- Yi, F.; Li, F.; Chen, D.; Ma, Y. Cystoisospora Belli Infection in an AIDS Patient in China: Need for Cautious Interpretation of mNGS. Indian J. Med. Microbiol. 2022, 40, 163–165. [Google Scholar] [CrossRef]
- Rial-Crestelo, D.; Otero, B.; Pérez-Ayala, A.; Pinto, A.; Pulido, F. Severe Diarrhea by Cystoisospora Belli in a Well Controlled HIV-Infected Patient. AIDS 2022, 36, 485–486. [Google Scholar] [CrossRef]
- Rose, G.S.; Arnold, C.A.; Badizadegan, K.; Carter, C.M.; Conces, M.R.; Kahwash, S.B.; Nicol, K.K.; Arnold, M.A. Cytoplasmic Fibrillar Aggregates in Gallbladder Epithelium Are a Frequent Mimic of Cystoisospora in Pediatric Cholecystectomy Specimens. Arch. Pathol. Lab. Med. 2019, 143, 1259–1264. [Google Scholar] [CrossRef]
- Gopinath, A.; Alkhasawneh, A.; Mubeen, A.; Makary, R.; Mohammed, I.; Baskovich, B. Pitfalls in Morphologic Diagnosis of Pathogens: Lessons Learned from the Pseudo-Cystoisospora Epidemic. Int. J. Surg. Pathol. 2021, 29, 169–173. [Google Scholar] [CrossRef]
- Ali, F.M. Detection of Cystoisospora belli among Children in Sulaimaniyah, Iraq. Research Square. [CrossRef]
- Vizon, K.C.C.; Battad, Z.G., 2nd; Castillo, D.S.C. Contamination of Food-Borne Parasites from Green-Leafy Vegetables Sold in Public Markets of San Jose City, Nueva Ecija, Philippines. J. Parasit. Dis. 2019, 43, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Barua, P.; Banik, K.S.; Saha, S.; Musa, S. Parasitic contamination of street food samples from schoolbased food vendors of Dhaka city, Bangladesh. Bangladesh J. Zool. 2023, 51, 217–229. [Google Scholar] [CrossRef]
- Zeng, H.; Van Damme, I.; Kabi, T.W.; Šoba, B.; Gabriël, S. Sarcocystis Species in Bovine Carcasses from a Belgian Abattoir: A Cross-Sectional Study. Parasit. Vectors 2021, 14, 271. [Google Scholar] [CrossRef]
- Ota, T.; Nakano, Y.; Mizuno, T.; Shiozaki, A.; Hori, Y.; Yamanishi, K.; Hayakawa, K.; Hayakawa, T.; Fujimoto, T.; Nakamoto, C.; Maejima, K.; Wada, Y.; Terasoma, F.; Ohnishi, T. First Case Report of Possible Sarcocystis Truncata-Induced Food Poisoning in Venison. Intern. Med. 2019, 58, 2727–2730. [Google Scholar] [CrossRef]
- Rubiola, S.; Moré, G.; Civera, T.; Hemphill, A.; Frey, C.F.; Basso, W.; Colasanto, I.; Vercellino, D.; Fidelio, M.; Lovisone, M.; Chiesa, F. Detection of Sarcocystis Hominis, Sarcocystis Bovifelis, Sarcocystis Cruzi, Sarcocystis Hirsuta and Sarcocystis Sigmoideus Sp. Nov. in Carcasses Affected by Bovine Eosinophilic Myositis. Food Waterborne Parasitol. 2024, 34, e00220. [Google Scholar] [CrossRef]
- Shams, M.; Shamsi, L.; Asghari, A.; Motazedian, M.H.; Mohammadi-Ghalehbin, B.; Omidian, M.; Nazari, N.; Sadrebazzaz, A. Molecular Epidemiology, Species Distribution, and Zoonotic Importance of the Neglected Meat-Borne Pathogen Sarcocystis Spp. In Cattle (Bos Taurus): A Global Systematic Review and Meta-Analysis. Acta Parasitol. 2022, 67, 1055–1072. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Gjerde, B.; Villa, L.; Minazzi, S.; Zanzani, S.A.; Riccaboni, P.; Sironi, G.; Manfredi, M.T. Prevalence and Molecular Characterisation of Sarcocystis Miescheriana and Sarcocystis in Wild Boars (Sus Scrofa) in Italy. Parasitol. Res. 2019, 118, 1271–1287. [Google Scholar] [CrossRef]
- Prakas, P.; Kirillova, V.; Dzerkale, A.; Kirjušina, M.; Butkauskas, D.; Gavarāne, I.; Rudaitytė-Lukošienė, E.; Šulinskas, G. First Molecular Characterization of Sarcocystis Miescheriana in Wild Boars (Sus Scrofa) from Latvia. Parasitol. Res. 2020, 119, 3777–3783. [Google Scholar] [CrossRef] [PubMed]
- Helman, E.; Dellarupe, A.; Cifuentes, S.; Chang Reissig, E.; Moré, G. Identification of Sarcocystis Spp. in Wild Boars (Sus Scrofa) from Argentina. Parasitol. Res. 2023, 122, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Metwally, D.M.; Al-Damigh, M.A.; Al-Turaiki, I.M.; El-Khadragy, M.F. Molecular Characterization of Sarcocystis Species Isolated from Sheep and Goats in Riyadh, Saudi Arabia. Animals (Basel) 2019, 9, 256. [Google Scholar] [CrossRef]
- Salehi, M.; Spotin, A.; Rostamian, M.; Adami, M. Prevalence and Molecular Assessment of Sarcocystis Infection in Livestock in Northeast Iran. Comp. Immunol. Microbiol. Infect. Dis. 2022, 80, 101738. [Google Scholar] [CrossRef]
- Feng, Y.; Guo, R.; Sang, X.; Zhang, X.; Li, M.; Li, X.; Yang, N.; Jiang, T. A Systematic Meta-Analysis of Global Sarcocystis Infection in Sheep and Goats. Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Januskevicius, V.; Januskeviciene, G.; Prakas, P.; Butkauskas, D.; Petkevicius, S. Prevalence and Intensity of Sarcocystis Spp. Infection in Animals Slaughtered for Food in Lithuania. Vet. Med. (Praha) 2019, 64, 149–157. [Google Scholar] [CrossRef]
- Zhu, Z.; Ying, Z.; Feng, Z.; Liu, Q.; Liu, J. The Occurrence and Meta-Analysis of Investigations on Sarcocystis Infection among Ruminants (Ruminantia) in Mainland China. Animals (Basel) 2022, 13, 149. [Google Scholar] [CrossRef]
- Khorshidvand, Z.; Khazaei, S.; Amiri, M.; Taherkhani, H.; Mirzaei, A. Worldwide Prevalence of Emerging Parasite Blastocystis in Immunocompromised Patients: A Systematic Review and Meta-Analysis. Microb. Pathog. 2021, 152, 104615. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Ismail, K.A.; Khalifa, O.M.; Abdel-wahab, M.M.; Hagag, H.M.; Mahmoud, M.K. Molecular Identification of Blastocystis hominis Isolates in Patients with Autoimmune Diseases. Appl. Microbiol. 2023, 3, 417–428. [Google Scholar] [CrossRef]
- Wakid, M.H.; Aldahhasi, W.T.; Alsulami, M.N.; El-Kady, A.M.; Elshabrawy, H.A. Identification and Genetic Characterization of Blastocystis Species in Patients from Makkah, Saudi Arabia. Infect. Drug Resist. 2022, 15, 491–501. [Google Scholar] [CrossRef]
- Maloney, J.G.; Molokin, A.; Seguí, R.; Maravilla, P.; Martínez-Hernández, F.; Villalobos, G.; Tsaousis, A.D.; Gentekaki, E.; Muñoz-Antolí, C.; Klisiowicz, D.R.; et al. Identification and Molecular Characterization of Four New Blastocystis Subtypes Designated ST35-ST38. Microorganisms 2023, 11, 46. [Google Scholar] [CrossRef]
- Piubelli, C.; Soleymanpoor, H.; Giorli, G.; Formenti, F.; Buonfrate, D.; Bisoffi, Z.; Perandin, F. Blastocystis Prevalence and Subtypes in Autochthonous and Immigrant Patients in a Referral Centre for Parasitic Infections in Italy. PLoS One 2019, 14, e0210171. [Google Scholar] [CrossRef]
- Cinek, O.; Polackova, K.; Odeh, R.; Alassaf, A.; Kramná, L.; Ibekwe, M.U.; Majaliwa, E.S.; Ahmadov, G.; Elmahi, B.M.E.; Mekki, H.; Oikarinen, S.; Lebl, J.; Abdullah, M.A. Blastocystis in the Faeces of Children from Six Distant Countries: Prevalence, Quantity, Subtypes and the Relation to the Gut Bacteriome. Parasit. Vectors 2021, 14, 399. [Google Scholar] [CrossRef] [PubMed]
- Rauff-Adedotun, A.A.; Meor Termizi, F.H.; Shaari, N.; Lee, I.L. The Coexistence of Blastocystis Spp. In Humans, Animals and Environmental Sources from 2010-2021 in Asia. Biology (Basel) 2021, 10, 990. [Google Scholar] [CrossRef]
- Issa, Y.A.; Ooda, S.A.; Salem, A.I.; Idris, S.N.; Elderbawy, M.M.; Tolba, M.M. Molecular Diagnosis and Subtyping of Blastocystis Sp.: Association with Clinical, Colonoscopic, and Histopathological Findings. Trop. Parasitol. 2023, 13, 46–53. [Google Scholar] [PubMed]
- Gureser, A.S.; Karasartova, D.; Sarzhanov, F.; Kosar, N.; Taylan-Ozkan, A.; Dogruman-Al, F. Prevalence of Blastocystis and Dientamoeba Fragilis in Diarrheal Patients in Corum, Türkiye. Parasitol. Res. 2023. [Google Scholar] [CrossRef]
- Marangi, M.; Boughattas, S.; De Nittis, R.; Pisanelli, D.; Delli Carri, V.; Lipsi, M.R.; La Bella, G.; Serviddio, G.; Niglio, M.; Lo Caputo, S.; Margaglione, M.; Arena, F. Prevalence and Genetic Diversity of Blastocystis Sp. among Autochthonous and Immigrant Patients in Italy. Microb. Pathog. 2023, 185, 106377. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.H.; Ismail, M.A.M.; El-Badry, A.A.; Abu-Sarea, E.Y.; Dewidar, A.M.; Hamdy, D.A. An Association between Blastocystis Subtypes and Colorectal Cancer Patients: A Significant Different Profile from Non-Cancer Individuals. Acta Parasitol. 2022, 67, 752–763. [Google Scholar] [CrossRef]
- Rojas-Velázquez, L.; Maloney, J.G.; Molokin, A.; Morán, P.; Serrano-Vázquez, A.; González, E.; Pérez-Juárez, H.; Ximénez, C.; Santin, M. Use of Next-Generation Amplicon Sequencing to Study Blastocystis Genetic Diversity in a Rural Human Population from Mexico. Parasit. Vectors 2019, 12, 566. [Google Scholar] [CrossRef]
- Rudzińska, M.; Sikorska, K. Epidemiology of Blastocystis Infection: A Review of Data from Poland in Relation to Other Reports. Pathogens 2023, 12, 1050. [Google Scholar] [CrossRef]
- Boutahar, M.; Belaouni, M.; Ibrahimi, A.; Eljaoudi, R.; Aanniz, T.; Er-Rami, M. Prevalence of Blastocystis Sp. in Morocco: Comparative Assessment of Three Diagnostic Methods and Characterization of Parasite Forms in Jones’ Culture Medium. Parasite 2023, 30, 64. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, F.; Babaei, E.; Badirzadeh, A.; Riabi, T.R.; Abdoli, A. Blastocystis, Urticaria, and Skin Disorders: Review of the Current Evidences. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1027–1042. [Google Scholar] [CrossRef]
- Alamlih, L.; Abufaied, M.; Al-Allaf, A.-W. An Unusual Cause of Reactive Arthritis with Urticarial: A Case Report. Qatar Med. J. 2020, 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Permasutha, M.B.; Pranata, I.W.A.; Diptyanusa, A.; Murhandarwati, E.E.H. Blastocystis Hominis Infection in HIV/AIDS Children with Extraintestinal Symptom: A Case Report. Bali Med. J. 2022, 11, 498–500. [Google Scholar] [CrossRef]
- Ascuña-Durand, K.; Salazar-Sánchez, R.S.; Cartillo-Neyra, R.; Ballón-Echegaray, J. Molecular Epidemiology of Blastocystis in Urban and Periurban Human Populations in Arequipa, Peru. bioRxiv 2020. [Google Scholar] [CrossRef]
- El-Zawawy, H.T.; Farag, H.F.; Tolba, M.M.; Abdalsamea, H.A. Improving Hashimoto’s Thyroiditis by Eradicating Blastocystis Hominis: Relation to IL-17. Ther. Adv. Endocrinol. Metab. 2020, 11, 2042018820907013. [Google Scholar] [CrossRef]
- Sayed Ahmed Nasr, M.; Mohammed Abdellatif, M.; Belal, U.; Abdel-Hafeez, E.; Abdalgelil, N. Detection of Blastocystis Species in the Stool of Immuno-Compromised Patients by Using Two Different Diagnostic Techniques. Minia Journal of Medical Research 2023, 34, 103–110. [Google Scholar] [CrossRef]
- Hidalgo-Gonzalez, L.A.; Salgado-Lopez, J.; Pineda-Rodriguez, S.A.; Martinez, A.; Romero-Valdovinos, M.; Martinez-Hernandez, F.; Rendon-Franco, E.; Olivo-Diaz, A.; Maravilla, P.; Rodriguez-Bataz, E. Identification of Blastocystis Sp. in School Children from a Rural Mexican Village: Subtypes and Risk Factors Analysis. Parasitol. Res. 2023, 122, 1701–1707. [Google Scholar] [CrossRef]
- Jinatham, V.; Wandee, T.; Nonebudsri, C.; Popluechai, S.; Tsaousis, A.D.; Gentekaki, E. Blastocystis Subtypes in Raw Vegetables from Street Markets in Northern Thailand. Parasitol. Res. 2023, 122, 1027–1031. [Google Scholar] [CrossRef]
- El Safadi, D.; Osman, M.; Hanna, A.; Hajar, I.; Kassem, I.I.; Khalife, S.; Dabboussi, F.; Hamze, M. Parasitic Contamination of Fresh Leafy Green Vegetables Sold in Northern Lebanon. Pathogens 2023, 12. [Google Scholar] [CrossRef]
- Veraldi, S.; Angileri, L.; Rossi, L.C.; Nazzaro, G. Endolimax Nana and Urticaria. J. Infect. Dev. Ctries. 2020, 14, 321–322. [Google Scholar] [CrossRef] [PubMed]
- Al-Hasnawy, M.H.; Rabee, A.H. A Review on Trichomonas Species Infection in Humans and Animals in Iraq. Iraqi J. Vet. Sci. 2023, 37, 305–313. [Google Scholar] [CrossRef]
- Abdo, S.M.; Ghallab, M.M.I.; Elhawary, N.M.; Elhadad, H. Pentatrichomonas hominis and other intestinal parasites in school-aged children: coproscopic survey. J Parasit Dis. 2022, 46, 896–900. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, Y.; Li, J.; Gong, P.; Wang, X.; Li, X.; Cheng, Y.; Yu, X.; Zhang, N.; Zhang, X. Changes of Gut Microbiota in Colorectal Cancer Patients with Pentatrichomonas Hominis Infection. Front. Cell. Infect. Microbiol. 2022, 12, 961974. [Google Scholar] [CrossRef]
- Withworth, J. Unpasteurized spinach drink tied to increase of cryptosporidium cases. Food Safety News. 2019. Available online: https://www.foodsafetynews.com/2019/12/unpasteurized-spinach-drink-tied-to-increase-of-cryptosporidium-cases/.
- Technical Committee: ISO/TC 34/SC 9. Microbiology of the food chain. Detection and enumeration of Cryptosporidium and Giardia in fresh leafy green. Available online: https://www.iso.org/standard/63252.html (accessed on 28 February 2024).
- World Health Organization. Ending the neglect to attain the sustainable development goals: a global strategy on water, sanitation and hygiene to combat neglected tropical diseases, 2021–2030. World Health Organization. 2021. Available online: https://iris.who.int/handle/10665/340240 (accessed on February 2024).
Figure 1.
Number of waterborne or foodborne acute cases/outbreaks reported for pathogenic protozoans since 2019 [8,9,19,35,36,45,47,64,66,68,69,72,110,125,126,132,133,146,151,152,162,169,170,171,172,173,178,185,186,199,200,205,207,209,211,218,220,221,228,254,255,262].
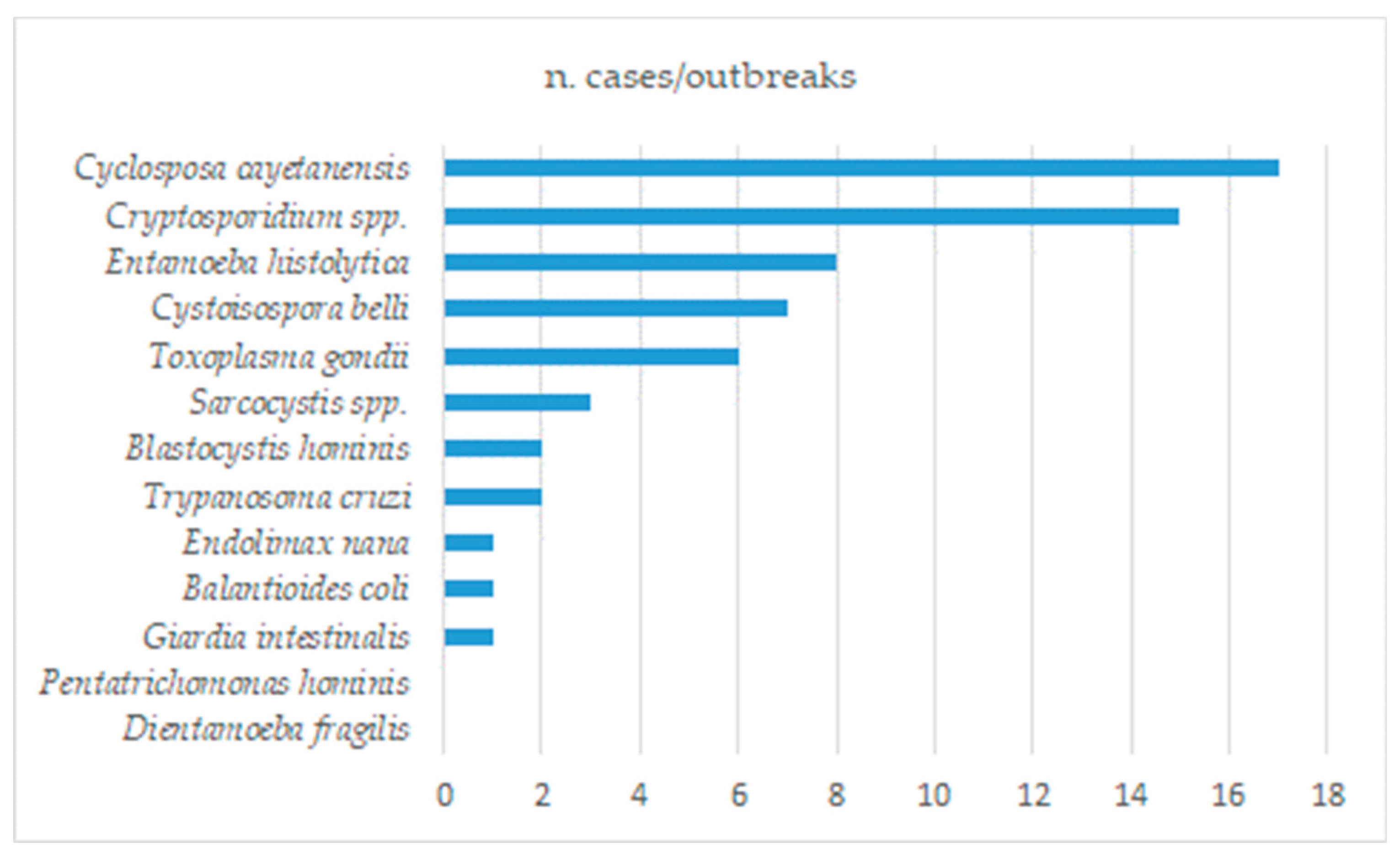

Table 1.
Dietary sources of pathogenic protozoans involved in cases and outbreaks according to reports and reviews published since 2019.
Table 1.
Dietary sources of pathogenic protozoans involved in cases and outbreaks according to reports and reviews published since 2019.
| Infectious agent | Dietary infection sources in cases and outbreaks* |
|---|---|
| Cryptosporidium spp. | Drinking water, unpasteurized or re-contaminated pasteurized milk, meat, dairy foods, dishes prepared outdoors, barbecued foods, raw foods, raw vegetables, raw fruits, unpasteurized drinks, apple juice, raw shellfish [9,99,100,107,108,110,113,121,124,125,126,127,129,130,131,132,133,266] |
| Entamoeba spp. | Drinking water, alcoholic fermented sap of the Palmyra toddy [7,162] |
| Toxoplasma gondii | Raw vegetables, fruits, raw or undercooked meats, unpasteurized milk, raw cow’s milk cheese, raw or undercooked crustaceans or shellfish, whale meat, drinking water [13,41,43,49,50,64,167,169,170,171,172,173] |
| Giardia intestinalis | Drinking water, fresh produce, composite ready to eat food, canned salmon, raw oysters, ice cream, noodle salad, chicken salad, dairy products, sandwiches, tripe soup, unpasteurized milk, shellfish, unidentified foods [177,178,179,182,194,195] |
| Trypanosoma cruzi | açaí juice, sugar cane juice, palm, guanabana, guava, milpesillo, majo, mango, mandarin, orange juices, raw meat [11,197] |
| Balantioides coli | None reported |
| Cyclospora cayetanensis | berries, cilantro, basil, lettuce, ready-to-eat bagged salads, sugar snap peas [6,207,208,209] |
| Cystoisospora belli | None reported |
| Sarcocystis spp. | Beef, pork, venison, whale meat [8,173,228] |
| Blastocystis hominis | None reported |
| Endolimax nana | None reported |
| Dientamoeba fragilis | None reported |
| Pentatrichomonas hominis | None reported |
*ascertained or most probable, determined by statistical association or by direct evidence in cases and outbreaks.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



