
Submitted:
22 May 2025
Posted:
23 May 2025
You are already at the latest version
Abstract
Intestinal protozoan parasites represent a significant public and veterinary health burden, especially in low- and middle-income countries, yet their transmission dynamics at the human-animal-environment interface remain poorly characterized in certain countries. This study investigated the prevalence and genetic diversity of key protozoa, including Cryptosporidium spp., Giardia duodenalis, Blastocystis spp., Entamoeba histolytica, and Enterocytozoon bieneusi, in a rural village in Iraq. Samples collected from humans (n=50), livestock (sheep and goats, n=50), water (n=20), and soil (n=20) were analysed using microscopy and molecular methods (qPCR and nested PCR). Blastocystis spp. (78% animals, 16% humans, 45% soil, 5% water) and Cryptosporidium spp. (26% animals, 12% humans, 5% soil, 15% water) were most frequently found using either microscopy and/or molecular detection. Molecular typing identified Cryptosporidium parvum in both humans and sheep, hinting at zoonotic transmission potential. Enterocytozoon bieneusi and Giardia were also found. Cryptosporidium ubiquitum and E. bieneusi genotypes BEB6 and COS-I were detected exclusively in sheep, suggesting roles as potential reservoirs. Blastocystis ST1 was detected in humans, while ST4 and ST10 occurred in sheep. Notably, molecular detection rates of Blastocystis were much lower than those of microscopy. Entamoeba histolytica was not detected. The detection of the same organisms in humans, animal and the environment, suggests zoonotic and environmental transmission pathways, which warrant further investigation using the One Health approach.
Keywords:
infectious diseases
; zoonotic transmission
; environmental contamination
; protozoa
; one health
1. Introduction
Intestinal protozoan parasites are globally significant pathogens, causing considerable morbidity in humans and livestock, particularly in low- and middle-income countries (LMICs)[1]. These organisms, including Cryptosporidium spp., Giardia duodenalis, Entamoeba histolytica, and Enterocytozoon bieneusi, contribute substantially to diarrheal disease, malnutrition, and economic losses in affected communities [2,3]. Beyond their clinical and veterinary impact, these parasites also pose an economic burden due to livestock losses and reduced productivity [4,5].
Cryptosporidium spp. and Giardia duodenalis are known zoonotic agents, frequently transmitted between humans and animals via the fecal-oral route, often through contaminated water or food [6]. Entamoeba histolytica is one of the few invasive amoebae and remains a major cause of dysentery worldwide [7]. Meanwhile, Blastocystis, a genetically diverse organism whose pathogenicity remains controversial, has garnered increasing attention in recent years due to its high prevalence in both healthy and symptomatic individuals, and potential associations with a healthy gut microbiome [8]. Finally, E. bieneusi is an emerging microsporidian parasite found in various hosts, including humans, and is particularly problematic in immuno-compromised individuals [9].
While these eukaryotic microbes have been studied globally, investigations in Iraq remain limited, with most relying on microscopy-based methods [10]. Studies based on molecular methods are scarce, particularly for Blastocystis and E. bieneusi. Prior reports have documented these protozoa in humans and animals in Iraq, but knowledge on their epidemiology and diversity remains limited [11,12,13].
To bridge these gaps, the current study employed both microscopic and molecular diagnostic methods to investigate the occurrence, genetic diversity, environmental contamination and potential zoonotic transmission of intestinal protozoa between humans and small ruminants in a rural village in Iraq.
2. Materials and Methods
2.1. Ethics
The Ethics Committee (ACUC) at the University of Baghdad approved this study on 16-03-2022 under the project "One Health Approach—Iraq" (No.D.A.672).
2.2. Study Area
The study took place at Alissma village, which is located in the northeastern part of Iraq, near the border with Iran (Figure 1). Approximately 1000 individuals reside in this village. The nearest city (Mandli) is about 30 km away. The village is located in a semi-desert area with nearly no tree cover, devoid of ponds and lakes. Oil River runs through the village in the winter, but its water is unfit for drinking and is used solely for irrigation purposes. Groundwater wells and underground well water are typically used for drinking after filtration and purification. The village latrines are basic and located outside the household.
2.3. Sample Collection
The methodology used herein is graphically summarized (Figure 2). A total of 140 samples were collected between February 2022 and July 2022. Fifty of these were stool samples collected from humans. The participants did not have diarrhoea or other gastrointestinal symptoms and were not taking antibiotics at the time of sampling, except four participants who had colitis as diagnosed by a physician. During sample collection, questionnaires were administered, and information about age, gender, type of drinking water, antibiotics, pets, career, and chronic disease was recorded.
Moreover, 50 samples were collected from animals, namely from goats (n=15) and sheep (n=35). These animals grazed near the village and drank unfiltered well water. None of the animals had diarrhea at the time of sampling.
Additionally, 20 water samples were collected from sources used by both humans and animals in 5 ml plastic tubes. These included filtered drinking water (n=5), tap water (n=5), well water (n=5), and river water (n=5). Twenty soil samples were also collected. These included home garden soil (n=5), field soil (n=5), animal grazing field soil (n=5), and river edge soil (n=5). Five grams of soil were collected in plastic tubes after scraping five centimeters from the surface of the soil.
2.4. Microscopic Examination
All samples were examined microscopically using wet mount smears. Iodine staining was used for the human and animal fecal samples. Samples were also observed using the concentration method which entailed mixing the samples with a salt solution, and subsequently examining the concentrated top layer for parasitic forms.
2.5. DNA Extraction and Molecular Detection
For DNA extraction, 200 mg of each fecal and soil sample was used using the PureLink™ Microbiome Genomic DNA Purification Kit (Invitrogen) according to the manufacturer’s protocol. The extracted DNA was used for qPCR and nested PCR (Table 1).
This study screened samples for Blastocystis sp., Cryptosporidium spp., Entamoeba histolytica, Giardia duodenalis and E. bieneusi. The SSU rRNA gene was used to identify Cryptosporidium spp. and gp60 was used for Cryptosporidium subtyping. For Blastocystis, SSU rRNA was used; beta-giardin (bg) and triosephosphate isomerase (tpi) for Giardia; and internal transcribed spacer (ITS) for E. bieneusi. A positive and negative control were included in each PCR run. The reaction conditions differed according to parasite and genetic marker (Table 1). Probe-based qPCR was used to amplify a fragment of the SSU rRNA of G. duodenalis and E. histolytica. The positive PCR products were purified using a Thermo Scientific GeneJET Gel Extraction Kit (Thermo Fisher Scientific, CA, USA) according to the manufacturer’s protocol. Methodology used in this study was according to previous investigations [14,15,16].
2.6. Sequencing Analysis
The purified positive PCR amplicons were sequenced unidirectionally at Eurofins genomics (Cologne, Germany). The obtained raw reads were trimmed manually at both ends to remove ambiguous bases using SnapGeneViewer v.6.0.2. The acquired sequences were used as queries to perform BLAST against the NCBI database. The newly generated nucleotide sequences were submitted to GenBank under accession numbers PV521084, PV521085, PV521086, PV521087, PV504621, PV504622, PV504623, and PV504624.
3. Results
3.1. Microscopic Examination
The stool and environmental samples were initially examined using light microscopy. A total of 12% (6/50) of the human samples were positive for Cryptosporidium spp., 16% (8/50) for Blastocystis sp., and 10% (5/50) for G. duodenalis (Table 2, Figure 3). Moreover, 26% (13/50) of the animal samples were positive for Cryptosporidium spp., 78% (39/50) for Blastocystis sp. [4% (8/50) of the Blastocystis sp. were from goats and 70% (35/50) were from sheep] and 8% (4/50) for G. duodenalis from sheep. Regarding the soil samples, 5% (1/20) tested positive for Cryptosporidium spp., and 45% (9/20) for Blastocystis sp. Regarding the water samples, 15% (3/20) were positive for Cryptosporidium spp. and 5% (1/20) for Blastocystis sp.
Giardia duodenalis and Enterocytozoon spp. were not detected by microscopy in any of the water or soil samples.
3.2. Molecular Detection
Based on qPCR analysis targeting a fragment of the SSU rRNA gene of G. duodenalis, 30% (15/50) of human samples were positive, 14% (7/50) of animals (all positives were from sheep), and 5% (1/20) of soil samples. Nested PCR of longer bg and tpi gene fragments amplified one of the qPCR positive sheep samples, which belonged to assemblage A.
Using nested PCR, one human sample was positive for C. parvum (gp60 gene) and another for Blastocystis ST1. In animals, Cryptosporidium spp. was detected in 4% (2/50); one sheep sample was positive for Cryptosporidium ubiquitum (SSU rRNA gene) and another for Cryptosporidium parvum (SSU rRNA, gp60 genes). 8% (4/50) of animals were positive for Blastocystis (SSU rRNA gene), one with ST4, and three with ST10. Finally, 8% (4/50) of animals were positive for E. bieneusi, with three belonging to E. bieneusi genotype BEB6 and one to genotype COS-I.
All samples were negative for Entamoeba histolytica.
Mixed infections were identified in 8% (4/50) of human samples, when combining microscopy and molecular detection methods (Figure 4). This included two samples that tested positive for Giardia duodenalis by molecular detection and Blastocystis sp. by microscopy. One sample was positive for both Cryptosporidium spp. and G. duodenalis by molecular detection, while another sample was positive for Cryptosporidium spp., G. duodenalis, and Blastocystis sp., all detected by molecular methods (Figure 5).
Regarding animal samples, 34% (17/50) had co-infections. Twelve were infected with Cryptosporidium spp. and Blastocystis sp. by microscopy-based detection, five with G. duodenalis and Blastocystis sp., this included three samples positive for G. duodenalis and Blastocystis sp. microscopically, one sample testing positive for both parasites molecularly and lastly, one tested positive for mixed infection, both microscopically and molecularly. Moreover, two animal samples tested positive for Blastocystis sp. by microscopy and E. bieneusi by PCR (Table 3).
4. Discussion
This study provides insights into the occurrence and genetic diversity of intestinal protozoan parasites in a rural village in Iraq. It highlights the zoonotic potential and environmental presence of species such as Cryptosporidium spp., Giardia duodenalis, Blastocystis spp., and Enterocytozoon bieneusi. Small ruminants are economically important animals in Iraq and are reared primarily on small- and medium-scale herds. Despite this, reports on their intestinal organisms are relatively sparse [17,18]. Herein, the occurrence rate of Cryptosporidium spp. in animals is lower than in previous studies, likely due to the methodologies used or the population examined. The occurrence in sheep is much higher than in goats, matching previous findings in the country [18]. This can be attributed to the free-range nature of goats as opposed to sheep [19]. The detection of Cryptosporidium parvum in both humans and sheep suggests zoonotic transmission within this community. Meanwhile, detection of Cryptosporidium ubiquitum and E. bieneusi in sheep hints at livestock as potential reservoirs for environmental contamination and human infection.
The high prevalence of Blastocystis in both animals and humans align with previous reports [20,21,22]. There was a notable difference between the detection methods used, with microscopy-based detection identifying many more samples as positive, rather than molecular methods, whereby it was not possible to amplify the corresponding gene fragment. The presence of co-infections, many of which were confirmed microscopically, could be a confounding factor here. While it is not possible to compare these findings to other studies in Iraq, previous molecular studies in neighboring Iran showed variable Blastocystis occurrence rates, with one as low 5% [23,24,25,26]. As this is, to the best of our knowledge, the first molecular detection study of Blastocystis in the country, further studies are needed to shed light on the organism’s epidemiology. The detection of ST4 in sheep is intriguing, as this subtype is typically linked to rodents and only infrequently reported in livestock. This is among the first reports of ST4 in sheep in this region, and as such, it could stipulate new insights into subtype distribution in the Middle East.
Interestingly, Entamoeba histolytica was not detected in any sample. Nonetheless, Entamoeba spp. have been detected in both microscopic and molecular-based investigations in ruminants in the country [27,28]. This discrepancy may reflect the increased specificity and sensitivity of molecular methods used in the present study, especially under challenging field conditions that may compromise the detection of fragile organisms such as E. histolytica.
Given the scarcity of molecular epidemiological studies in Iraq, direct comparisons to previous work are limited. Nonetheless, our findings are in line with some earlier reports, such as the 34% Giardia detection rate by microscopy in humans observed by Al-Hasnawy and Idan [29].
While E. bieneusi infections have been reported in birds in Iraq [30], this study contributes new evidence of its presence in livestock, expanding the known host range in the region.
Several limitations should be acknowledged. One major challenge was preserving and storing samples in a remote field setting, where access to cold-chain infrastructure was limited. Inadequate preservation may have reduced the sensitivity of both microscopy and molecular assays, especially for rapidly degrading parasites. This could explain certain species' absence or low detection rates in specific sample types. Additionally, although PCR-based methods were employed, only a subset of positive samples yielded high-quality sequences, limiting the depth of genetic characterization. Environmental samples, in particular, may have contained PCR inhibitors or low DNA concentrations, affecting amplification success. Our findings underline the need for standardized operating procedures for parasite sampling, preservation, and analysis in resource-limited settings. The adoption of field-friendly preservatives compatible with molecular diagnostics, along with optimized DNA extraction protocols for complex environmental matrices, would enhance data quality and comparability [31]. Another limitation of this study is its cross-sectional design, which provides only a single time point and does not capture seasonal trends or temporal changes in infection dynamics. Longitudinal studies incorporating repeated sampling from both hosts and environments are needed to clarify transmission pathways, sources of reinfection, and possible seasonal patterns [32].
Despite including asymptomatic individuals and animals, future research should aim to expand the number of sampling sites, include larger sample sizes, and integrate clinical and immunological data. Such approaches would allow a better assessment of the pathogenic potential and health impacts of these parasites.
Looking ahead, the high prevalence of Blastocystis a common yet enigmatic member of the gut eukaryome presents a valuable opportunity to study its interaction with the bacterial microbiota and host immune system. Future studies should consider using 16s rRNA sequencing or shotgun metagenomics to explore microbe-parasite interactions, particularly in communities with frequent co-infections [33]. These approaches could inform new strategies for diagnostics, surveillance, and intervention.
5. Conclusions
In conclusion, this study enriches the limited molecular data on intestinal protozoan parasites in Iraq, offering a comprehensive view of their prevalence in humans, livestock, and the environment. The findings emphasize the importance of enhanced sampling protocols, environmental monitoring, and capacity building to improve parasite detection and control in rural, resource-limited settings.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Map of Iraq and neighbouring countries. The study area is located in the eastern part of Iraq, within the Diali Province. The village location is marked with the red pin and lies about 10 km from the Iranian border. Figure S2: Methodology summary for the samples and approaches used in the study. Figure S3: Cryptosporidium (A) and Blastocystis (B) from fecal samples. Table S1: Parasites, genes, primer sequences, Amplification Procedures, and the expected fragment size (bp), which were used in the study.; Table S2: Number and percentage of positive samples in the microscopic examination for all sources and parasites used in the study: Table S3: Number and percentage of positive samples that achieved co-infection in microscopic and molecular examination for all sources and parasites used in the study.
Author Contributions
Conceptualization, A.D.T, M.M.S and D.A.K.; methodology, Y.M.S.A and S.M.; software, Y.M.S.A and E.G.; validation, Y.M.S.A, E.A.O, E.G and A.D.T.; formal analysis, Y.M.S.A, and E.G.; investigation, Y.M.S.A, E.G and A.D.T.; resources, A.D.T.; data curation, Y.M.S.A.; writing—original draft preparation, Y.M.S.A.; writing—review and editing, Y.M.S.A, S.M, A.D.T, E.G, M.M.S and D.A.K.; visualization, Y.M.S.A.; supervision, A.D.T, E.G, M.M.S and D.A.K. and E.A.O,; project administration, A.D.T.; funding acquisition, A.D.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of the University of Baghdad (protocol code No. D.A. 672 and 16-03-2022).” for studies involving humans. The animal study protocol was approved by the Institutional Review Board (or Ethics Committee) of the University of Baghdad (protocol code No. D.A. 672 and 16-03-2022) for studies involving animals.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The original contributions presented in this study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author(s).
Acknowledgments
The authors would like to extend their deepest thanks to the residents and community of Alissma Village for their help and participation in collecting samples, which made this research successful. We also wish to acknowledge the exceptional guidance and support provided by the Parasitology Lab at the University of Baghdad.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- A. Abdoli, ‘Prevalence of intestinal protozoan parasites among Asian schoolchildren: a systematic review and meta-analysis’, Infection, vol. 52, no. 6, Dec. 2024. [CrossRef]
- S. T. Ghebremichael et al., ‘First identification and coinfection detection of Enterocytozoon bieneusi, Encephalitozoon spp., Cryptosporidium spp. and Giardia duodenalis in diarrheic pigs in Southwest China’, BMC Microbiol, vol. 23, no. 1, pp. 1–14, Dec. 2023. [CrossRef]
- R. Das et al., ‘Symptomatic and asymptomatic enteric protozoan parasitic infection and their association with subsequent growth parameters in under five children in South Asia and sub-Saharan Africa’, PLoS Negl Trop Dis, vol. 17, no. 10, p. e0011687, Oct. 2023. [CrossRef]
- D. C. De Graaf, E. Vanopdenbosch, L. M. Ortega-Mora, H. Abbassi, and J. E. Peeters, ‘A review of the importance of cryptosporidiosis in farm animals’, Int J Parasitol, vol. 29, no. 8, pp. 1269–1287, Aug. 1999. [CrossRef]
- H. Gao et al., ‘Prevalence and Molecular Characterization of Cryptosporidium spp., Giardia duodenalis, and Enterocytozoon bieneusi in Diarrheic and Non-Diarrheic Calves from Ningxia, Northwestern China’, Animals 2023, Vol. 13, Page 1983, vol. 13, no. 12, p. 1983, Jun. 2023. [CrossRef]
- H. Hsu et al., ‘An Epidemiological Assessment of Cryptosporidium and Giardia spp. Infection in Pet Animals from Taiwan’, Animals 2023, Vol. 13, Page 3373, vol. 13, no. 21, p. 3373, Oct. 2023. [CrossRef]
- R. Haque, C. D. Huston, M. Hughes, E. Houpt, and W. A. Petri, ‘Amebiasis’, N Engl J Med, vol. 348, no. 16, pp. 1565–1573, Apr. 2003. [CrossRef]
- M. Aykur et al., ‘Blastocystis: A Mysterious Member of the Gut Microbiome’, Microorganisms, vol. 12, no. 3, p. 461, Mar. 2024. [CrossRef]
- J. Mena et al., ‘Enterocytozoon bieneusi Infection after Hematopoietic Stem Cell Transplant in Child, Argentina - Volume 30, Number 3—March 2024 - Emerging Infectious Diseases journal - CDC’, Emerg Infect Dis, vol. 30, no. 3, pp. 613–616, Mar. 2024. [CrossRef]
- N. Hassan Bedair and I. Naif Zeki, ‘Prevalence of Some Parasitic Infections in Iraq from 2019 to 2020’, Bedair and Zeki Iraqi Journal of Science, vol. 64, no. 7, pp. 4181– 4191, 2023. [CrossRef]
- M. I. Alberfkani, ‘Molecular characterization of Blastocystis hominis in irritable bowel syndrome patients and nursing staff in public and private clinic in Iraq’, Ann Parasitol, vol. 68, no. 4. 2022; 715–720. [CrossRef]
- S. A. Khabisi, A. N. Najafi, and A. S. Khorashad, ‘Molecular diagnosis of intestinal microsporidia infection in HIV/AIDS-patients in Zahedan city, Southeast of Iran’, Ann Parasitol, vol. 68. 2022; 87–92. [CrossRef]
- Al-Sadi Hafidh and Al-Mahmood aevan, ‘MICROSPORIDIAL INFECTION IN SOME DOMESTICAND LABORATORY ANIMALS IN IRAQ’, International Journal of Basic and Applied Medical Sciences, vol. Vol. 3 (3), no. ISSN: 2277-2103, pp. 78–91, Sep. 2013.
- S. Maxamhud et al., ‘Molecular Identification of Cryptosporidium spp., and Giardia duodenalis in Dromedary Camels (Camelus dromedarius) from the Algerian Sahara’, Parasitologia 2023, Vol. 3, Pages 151-159, vol. 3, no. 2, pp. 151–159, Apr. 2023. [CrossRef]
- V. Jinatham, S. Maxamhud, S. Popluechai, A. D. Tsaousis, and E. Gentekaki, ‘Blastocystis One Health Approach in a Rural Community of Northern Thailand: Prevalence, Subtypes and Novel Transmission Routes’, Front Microbiol, vol. 12, p. 746340, Dec. 2021. [CrossRef]
- S. Muadica et al., ‘First identification of genotypes of Enterocytozoon bieneusi (Microsporidia) among symptomatic and asymptomatic children in Mozambique’, PLoS Negl Trop Dis, vol. 14, no. 6, p. e0008419, Jun. 2020. [CrossRef]
- A. Hraiga, ‘Investigation of Cryptosporidium infection in Lambs and Goat Kids at Al-kut city,wasit province’, Journal of Health, Medicine and Nursing, vol. 43, no. 0, pp. 130–134, 2017, Accessed: May 08, 2025. [Online]. Available: https://iiste.org/Journals/index.php/JHMN/article/view/39540.
- M. Jadaan, A. Alkhaled, and W. A. Hamad, ‘Molecular characterization of Cryptosporidium spp. in sheep and goat in Al-Qadisiyah province/ Iraq: Mansoor Jadaan Ali Alkhaled and Weam Abbas Hamad’, The Iraqi Journal of Veterinary Medicine, vol. 41, no. 2, pp. 31–37, Feb. 2017. [CrossRef]
- K. Hatam-Nahavandi, E. Ahmadpour, D. Carmena, A. Spotin, B. Bangoura, and L. Xiao, ‘Cryptosporidium infections in terrestrial ungulates with focus on livestock: a systematic review and meta-analysis’, Parasites & Vectors 2019 12:1, vol. 12, no. 1, pp. 1–23, Sep. 2019. [CrossRef]
- J. Rehena, A. Bin Harun, and M. R. Karim, ‘Epidemiology of Blastocystis in farm animals: A review’, Vet Parasitol, vol. 334, Feb. 2025. [CrossRef]
- A. Rauff-Adedotun, F. H. M. Termizi, N. Shaari, and I. L. Lee, ‘The coexistence of Blastocystis spp. In humans, animals and environmental sources from 2010–2021 in Asia’, Biology (Basel), vol. 10, no. 10, Oct. 2021. [CrossRef]
- R. Salehi et al., ‘Prevalence and subtype identification of Blastocystis isolated from humans in Ahvaz, Southwestern Iran’, Gastroenterol Hepatol Bed Bench, vol. 10, no. 3, 2017; 235. [CrossRef]
- K. Hatam-Nahavandi, H. Mohammad Rahimi, M. Rezaeian, E. Ahmadpour, M. Badri, and H. Mirjalali, ‘Detection and molecular characterization of Blastocystis sp., Enterocytozoon bieneusi and Giardia duodenalis in asymptomatic animals in southeastern Iran’, Sci Rep, vol. 15, no. 1, p. 6143, Dec. 2025. [CrossRef]
- M. Heydarian, K. Manouchehri Naeini, S. Kheiri, and R. Abdizadeh, ‘Prevalence and subtyping of Blastocystis sp. in ruminants in Southwestern, Iran’, Scientific Reports 2024 14:1, vol. 14, no. 1, pp. 1–15, Aug. 2024. [CrossRef]
- H. Mohammad Rahimi, H. Mirjalali, and M. R. Zali, ‘Molecular epidemiology and genotype/subtype distribution of Blastocystis sp., Enterocytozoon bieneusi, and Encephalitozoon spp. in livestock: concern for emerging zoonotic infections’, Scientific Reports 2021 11:1, vol. 11, no. 1, pp. 1–16, Sep. 2021. [CrossRef]
- M. Shams et al., ‘First molecular characterization of Blastocystis subtypes from domestic animals (sheep and cattle) and their animal-keepers in Ilam, western Iran: A zoonotic concern’, Journal of Eukaryotic Microbiology, vol. 71, no. 3, p. e13019, May 2024. [CrossRef]
- R. W. Kadhum, N. Jaber, H. Al-Asadi, Z. Abbas, and J. Al-Maliki, ‘STUDY THE PREVALENCE OF ENTAMOEBA SPECIES IN CHILDREN AND FARM ANIMALS IN WAIST PROVINCE’, Web of Scientists and Scholars: Journal of Multidisciplinary Research, vol. 2, no. 4, pp. 14–23, Apr. 2024, Accessed: 8 May, 2025. [Online]. Available: https://webofjournals.com/index.php/12/article/view/1240.
- S. M. K. Al-Dabbagh, H. H. Alseady, and E. J. Alhadad, ‘Molecular identification of Entamoeba spp. in humans and cattle in Baghdad, Iraq’, Vet World, vol. 17, no. 6, p. 1348, Jun. 2024. [CrossRef]
- M. H. Al-Hasnawy and S. R. Idan, ‘International Journal of Pharmaceutical and Bio-Medical Science Microscopic and Molecular Diagnosis of Giardia duodenalis in Human in Babylon Province, Iraq’. [CrossRef]
- A. Abdullah, W. A. Alobaidii, Y. N. M. Alkateb, F. F. Ali, S. D. Ola-Fadunsin, and F. I. Gimba, ‘Molecular detection and prevalence of human-pathologic Enterocytozoon bieneusi among pet birds in Mosul, Iraq’, Comp Immunol Microbiol Infect Dis, vol. 95, p. 101964, Apr. 2023. [CrossRef]
- T. P. Vital, G. S. V. P. Raju, I. S. S. Rao, and A. D. P. Kumar, ‘Data collection, statistical analysis and machine learning studies of cancer dataset from North Costal Districts of AP, India’, Procedia Comput Sci, vol. 48, no. C. 2015; 706–714. [CrossRef]
- C. R. Stensvold, ‘Blastocystis in stool: friend, foe or both?’, J Travel Med, vol. 32, no. 2, p. 11, Mar. 2025. [CrossRef]
- A. D. Tsaousis, E. Gentekaki, and C. R. Stensvold, ‘Advancing research on Blastocystis through a One Health approach’, Open Research Europe, vol. 4, p. 2024; 145. [CrossRef]
- N. Reghaissia et al., ‘First Epidemiological Report on the Prevalence and Associated Risk Factors of Cryptosporidium spp. in Farmed Marine and Wild Freshwater Fish in Central and Eastern of Algeria’, Acta Parasitol, vol. 67, no. 3, pp. 1152–1161, Sep. 2022. [CrossRef]
- G. Lindergard, D. V. Nydam, S. E. Wade, S. L. Schaaf, and H. O. Mohammed, ‘The sensitivity of PCR detection of Cryptosporidium oocysts in fecal samples using two DNA extraction methods’, Mol Diagn, vol. 7, no. 3–4. 2003; 147–153. [CrossRef]
- P. Pinto et al., ‘Cross-Border Investigations on the Prevalence and Transmission Dynamics of Cryptosporidium Species in Dairy Cattle Farms in Western Mainland Europe’, Microorganisms 2021, Vol. 9, Page 2394, vol. 9, no. 11, p. 2394, Nov. 2021. [CrossRef]
- R. ten Hove, T. Schuurman, M. Kooistra, L. Möller, L. Van Lieshout, and J. J. Verweij, ‘Detection of diarrhoea-causing protozoa in general practice patients in The Netherlands by multiplex real-time PCR’, Clinical Microbiology and Infection, vol. 13, no. 10, pp. 1001–1007, Oct. 2007. [CrossRef]
- J. J. Verweij et al., ‘Simultaneous Detection of Entamoeba histolytica, Giardia lamblia, and Cryptosporidium parvum in Fecal Samples by Using Multiplex Real-Time PCR’, J Clin Microbiol, vol. 42, no. 3, pp. 1220–1223, Mar. 2004. [CrossRef]
- S. M. Cacciò, M. De Giacomo, and E. Pozio, ‘Sequence analysis of the β-giardin gene and development of a polymerase chain reaction–restriction fragment length polymorphism assay to genotype Giardiaduodenalis cysts from human faecal samples’, Int J Parasitol, vol. 32, no. 8, pp. 1023–1030, Jul. 2002. [CrossRef]
- M. Sulaiman et al., ‘Triosephosphate Isomerase Gene Characterization and Potential Zoonotic Transmission of Giardia duodenalis - Volume 9, Number 11—November 2003 - Emerging Infectious Diseases journal - CDC’, Emerg Infect Dis, vol. 9, no. 11, pp. 1444––1452, 2003. [CrossRef]
- S. M. Scicluna, B. Tawari, and C. G. Clark, ‘DNA Barcoding of Blastocystis’, Protist, vol. 157, no. 1, pp. 77–85, Feb. 2006. [CrossRef]
- M. A. Buckholt, J. H. Lee, and S. Tzipori, ‘Prevalence of Enterocytozoon bieneusi in swine: An 18-month survey at a slaughterhouse in Massachusetts’, Appl Environ Microbiol, vol. 68, no. 5, pp. 2595––2599, 2002. [CrossRef]
Figure 1.
Map of Iraq and neighbouring countries. The study area is located in the eastern part of Iraq, within the Diali Province. The village location is marked with the red pin and lies about 10 km from the Iranian border.
Figure 1.
Map of Iraq and neighbouring countries. The study area is located in the eastern part of Iraq, within the Diali Province. The village location is marked with the red pin and lies about 10 km from the Iranian border.
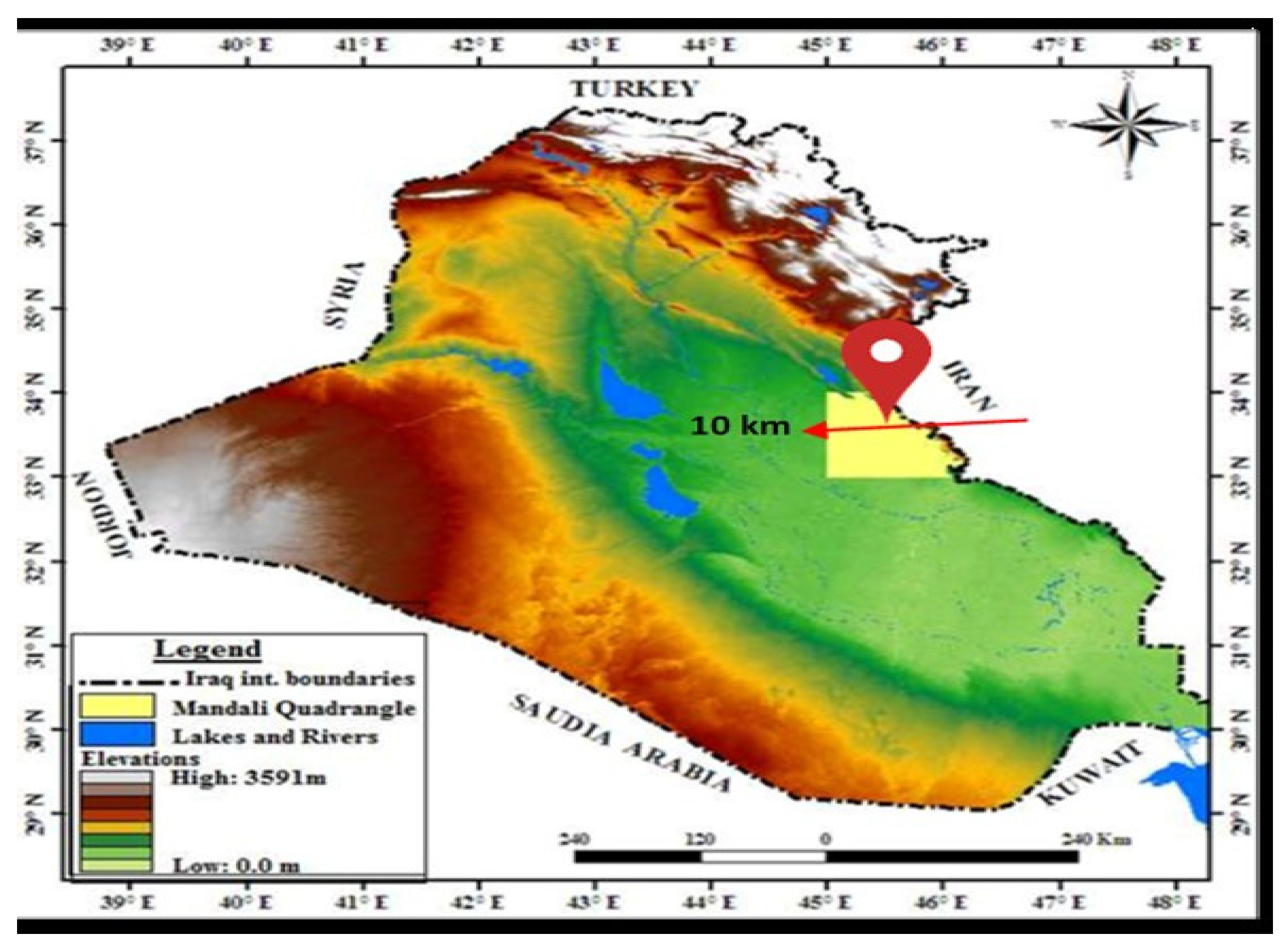
Figure 2.
Methodology summary for the samples and approaches used in the study.
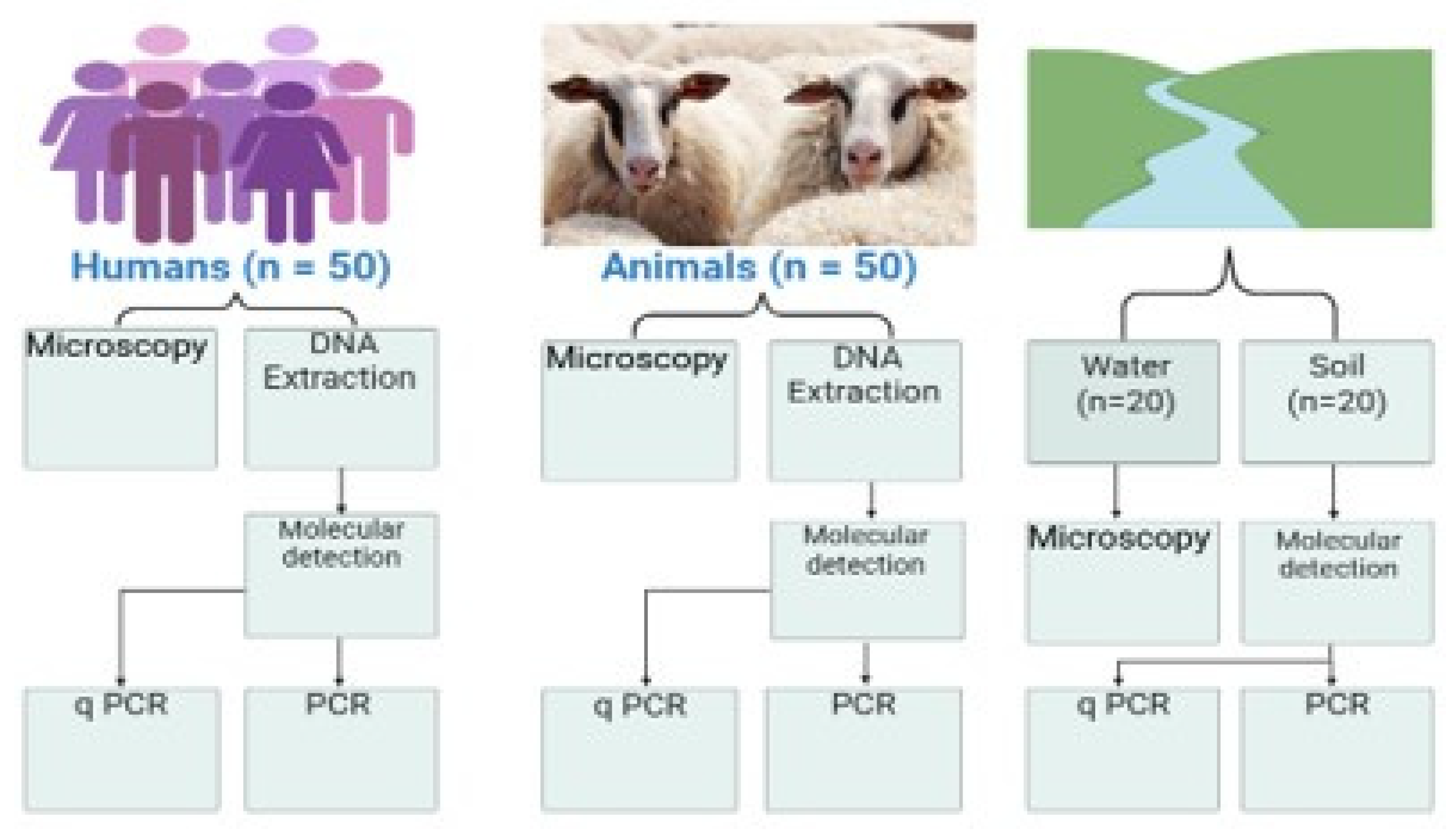
Figure 3.
Cryptosporidium (A) and Blastocystis (B) from faecal samples.
Figure 4.
Venn diagram showing the overlap of intestinal protozoan infections among 187 human stool samples. Blastocystis was identified by microscopy, Giardia duodenalis by qPCR, and Cryptosporidium spp. by nested PCR. Only a limited number of co-infections were detected, and no triple infections were observed.
Figure 4.
Venn diagram showing the overlap of intestinal protozoan infections among 187 human stool samples. Blastocystis was identified by microscopy, Giardia duodenalis by qPCR, and Cryptosporidium spp. by nested PCR. Only a limited number of co-infections were detected, and no triple infections were observed.
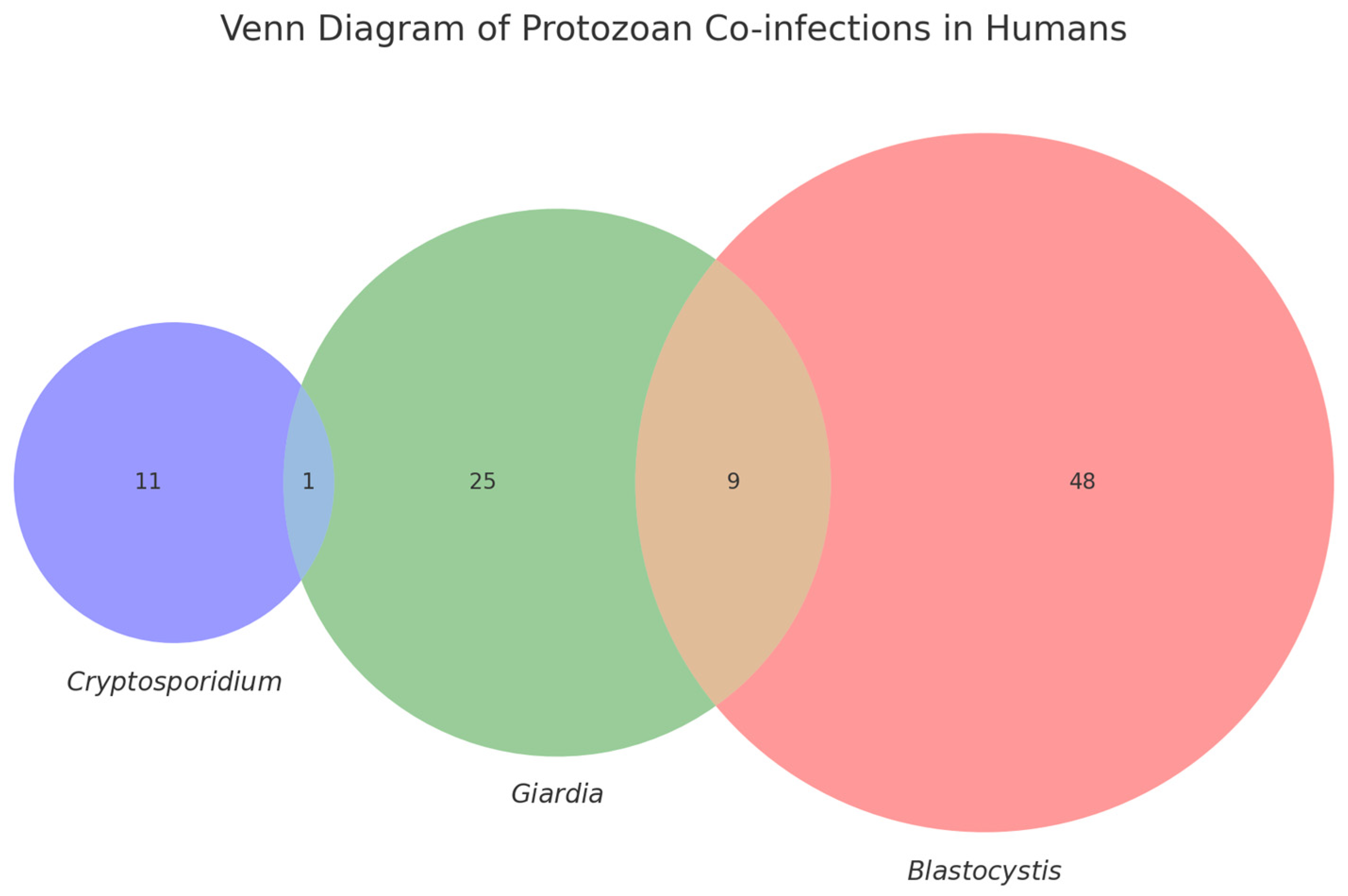
Figure 5.
Bar chart showing the number of positive detections for each protozoan parasite using different detection methods. Microscopy detected more Blastocystis and Cryptosporidium spp. compared to PCR, highlighting potential false positives or DNA degradation.
Figure 5.
Bar chart showing the number of positive detections for each protozoan parasite using different detection methods. Microscopy detected more Blastocystis and Cryptosporidium spp. compared to PCR, highlighting potential false positives or DNA degradation.
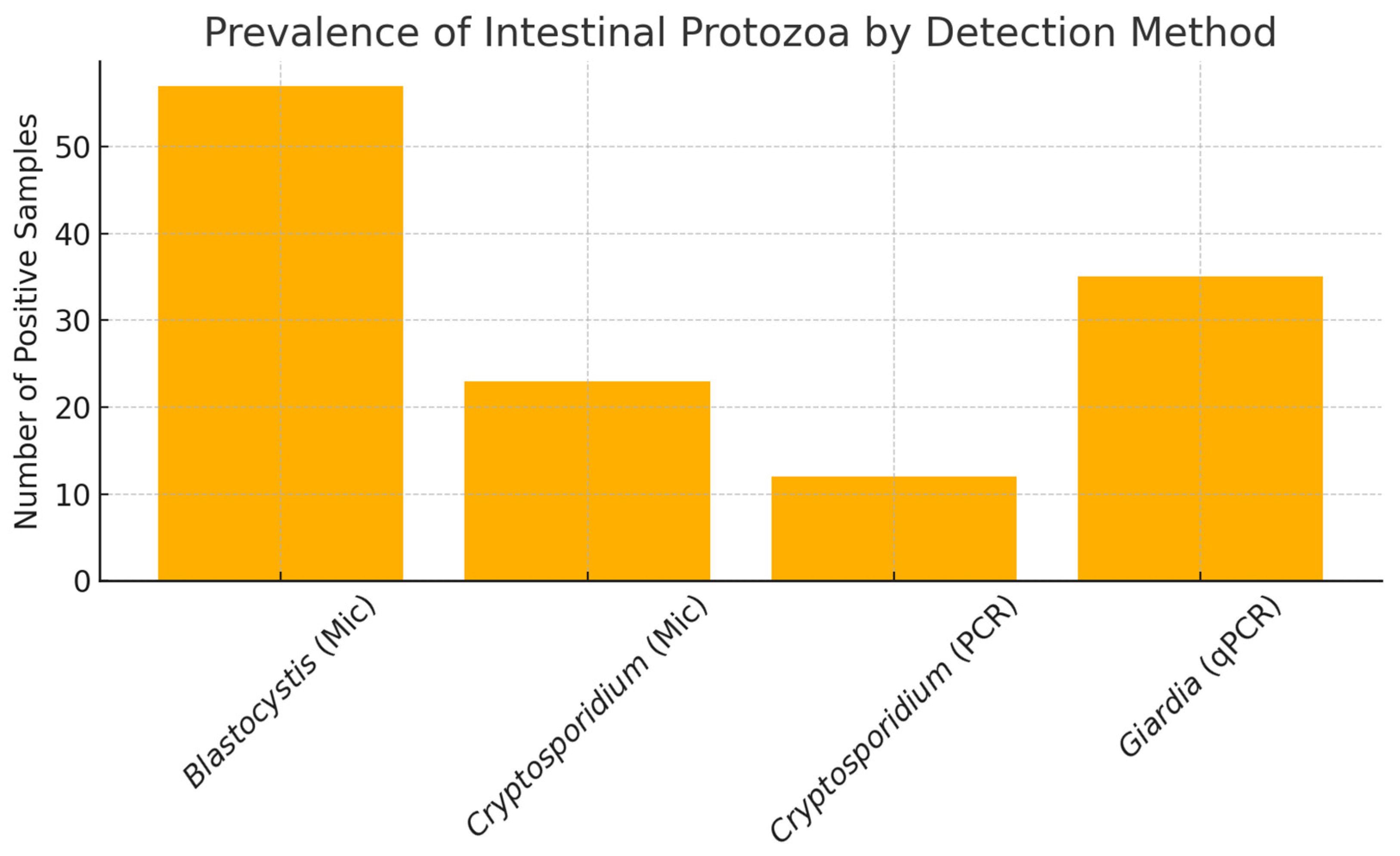

Table 1.
Parasites, genes, primer sequences, Amplification Procedures, and the expected fragment size (bp), which were used in the study.
Table 1.
Parasites, genes, primer sequences, Amplification Procedures, and the expected fragment size (bp), which were used in the study.
| Parasite of interest | Target gene | Detection Method | PrimerSequences (5’-3’) | AmplificationCondition | Amplicon Size(bp) | Reference |
| Cryptosporidium | SSU |
Npcr |
CRY-SSU-F1: GATTAAGCCATGCATGTCTAA | 95 C: 2 min; 24 cycles: (94 C: 50 s, 53 C: 50 s, 72 C:1 min); 72 C: 10 min | 723 bp |
[14,34] [35] [36] |
| CRY-SSU-R1: CTTGAATACTCCAGCATGGAA | ||||||
| CRY-SSU- F2 F2:CAGTTATAGTTTACTTGATAATC | 94 C: 2 min; 30 cycles: (94 C: 50 s, 56 C: 30 s, 72 C:1 min); 72 C: 10 min | 631 bp | ||||
| CRY-SSU- R2 R2:GAAAATTAGAGTGCTTAAAGCAGG | ||||||
| GP60 |
nPCR |
F1: AL3531: ATAGTCTCCGCTGTATTC | 94 C: 3 min; 35 cycles: (94 C: 45 s, 50 C: 45 s, 72 C:1 min); 72 C: 7 min | 1000 bp | ||
| R1: AL3535: GCAAGGAACGATGTATCT | ||||||
| F2 AL3532: TCCGCTGTATTCTCAGCC | 94 C: 3 min; 35 cycles: (94 C: 45 s, 50 C: 45 s, 72 C:1 min); 72 C: 7 min | 850 bp | ||||
| R2 AL3534: GCAGAGGAACCAGCATC | ||||||
| Giardia duodenalis | SSU | qPCR | GIARDIA-80-F: GACGGCTCAGGACAACGGTT | 95 C: 2 min; 50 cycles: (95 C: 15 s); 50 cycles: (58 C: 30 s), 50 cycles: (72 C: 30 s). | 62 bp | [14,37,38] |
| GIARDIA-127-R:TTGCCAGCGGTGTCCG | ||||||
| Probe: FAM-5'-CCCGCGGCGGTCCCTGCTAG-3' | ||||||
| Bg beta-giardin |
nPCR |
F1(G7F): AAGCCCGACCTCACCCGCAGTGC | 94 C: 5 min; 35 cycles: (94 C: 30 s, 66 C: 30 s, 72 C:1 min); 72 C: 7 min | 292 bp | [39] | |
| F2(G376): CATAAGGACGCCATCGCGGCTCTGAGG | 94 C: 3 min; 30 cycles: (94 C: 30 s, 65 C: 15 s, 72 C:30 s); 72 C: 7 min | |||||
| R (G759R): GAGGCCGCCCTGGATCTTCGAGACGAC | ||||||
| Tpi triosephosphate isomerase |
nPCR |
Tpi_AL3543_F1: AAAT/IDEOXYL/ATGCCTGGTCG | 94 C: 3 min; 35 cycles: (94 C: 45 s, 50 C: 35 s, 72 C:30 s); 72 C: 10 min | 605 bp | [40] | |
| Tpi_AL3546_R1: CAAACCTT/IDEOXYL/TCCGCAAACC | ||||||
| Tpi_AL3544_F2: CCCTTGATCGG/IDEXYL/GGTAACTT | 94 C: 3 min; 35 cycles: (94 C: 35 s, 47 C: 35 s, 72 C:30 s); 72 C: 10 min | |||||
| Tpi_AL3545_R2: GTGGCCACCAC/IDEOXYL/CCCGTGCC | ||||||
|
Blastocystis |
SSU |
nPCR |
RD3 – F1 5′-GGGATCCTGA TCCTTCCGCAGGTTCACCTAC-3′ |
3 min at 94°C, 35 cycles at 94°C for 1 min, annealing 60°C for 1 min, and extension at 72°C for 100 s, with a final elongation step at 72°C for 7 min. | 650 bp |
[15] (Clark, 1997) [41] |
| RD5 – R1 5′-GGAAGC TTATCTGGTTGATCCTGCCAGTA-3′ | ||||||
| BsRD5F – F2 (5′-ATCTGGTTGATCCTGCCAGT-3′) |
3 min at 94°C, 35 cycles at 94°C for 1 min, annealing 60°C for 1 min, and extension at 72°C for 100 s, with a final elongation step at 72°C for 10 min. | |||||
| BhRDr – R2 (5′-GAGCTTTTTAACTGCAACAACG-3′) | ||||||
| Entamoeba histolytica | SSU |
qPCR |
End-239F – 5’-ATT GTC GTG GCA TCC TAA CTC A-3’ | 95 °C: 2 min; 50 cycles: (95 °C: 15 s); 50 cycles: (58 °C: 30 s), 50 cycles: (72 °C: 30 s). | 172 bp | [38] |
| End-88R – 5’. GCG GAC GGC TCA TTA TAA CA.3 | ||||||
| probe (VIC-5′-TCATTGAATGAATTGGCCATTT-3′-NFQ) | ||||||
| Enterocytozoonbieneusi | ITS Internal Transcribed Spacer |
nPCR |
EBITS3 (5´‒GGTCATAGGGATGAAGAG‒3´) |
95 °C 5 min 35 Cycles: 94 °C 40s 53 °C 45s 72 °C 45s 72 °C 4 min |
390 bp | [16] [42] |
| EBITS4 (5´‒TTCGAGTTCTTTCGCGCTC‒3´) | ||||||
| EBITS1 (5´‒GCTCTGAATATCTATGGCT‒3´) |
95 °C 5 min 30 Cycles: 94 °C 35s 55 °C 40s 72 °C 40s 72 °C 5 min |
|||||
| EBITS2.4 (5´‒ATCGCCGACGGATCCAAGTG‒3´) |
Table 2.
Number and percentage of positive samples in the microscopic examination for all sources and parasites used in the study.
Table 2.
Number and percentage of positive samples in the microscopic examination for all sources and parasites used in the study.
| Type of source | Human | Animals | soil | water | Total | |||||
| Name of Parasites | +ve | % | +ve | % | +ve | % | +ve | % | +ve | % |
| Cryptosporidium spp | 6 | 12% | 13 | 26% | 1 | 5% | 3 | 15% | 23 | 19.16% |
| Blastocystis sp | 8 | 16% | 39 | 78% | 9 | 45% | 1 | 5% | 57 | 47.5% |
| Goat: 8 | 4% | |||||||||
| Giardia spp | 5 | 10% | 4 | 8% | - | - | - | - | 9 | 7.5% |
| Total | 19 | 56 | 10 | 4 | 89 | 63.57% | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



