Submitted:
14 March 2024
Posted:
15 March 2024
You are already at the latest version
Abstract
Vibrio parahaemolyticus is an important human pathogen that is currently the leading cause of shellfish-borne gastroenteritis in the world. Particularly, the pandemic strain has the capacity to induce cytotoxicity and enterotoxicity through its Type 3 Secretion System (T3SS2) that leads to massive cell death. However, the specific mechanism by which T3SS2 induces cell death remain unclear and the contribution in mitochondrial stress is not fully understood. In this work, we evaluated the contribution of T3SS2 of V. parahaemolyticus to generate mitochondrial stress and cell death during infection in human intestinal HT-29 cells. To evaluate this, infection assays were carried out to evaluate mitochondrial transition pore opening, mitochondrial fragmentation, ATP quantification, and cell viability during infection. Our results showed that the Δvscn1 (T3SS2+) mutant strain contributes to generating the sustained opening of the mitochondrial transition pore. Furthermore, it generates perturbations in the ATP production in infected cells, leading to a significant decrease in cell viability and loss of membrane integrity. Our results suggest that T3SS2 from V. parahaemolyticus plays a role in generating mitochondrial stress and cell death in human intestinal HT-29 cells. It is important to highlight that this study represents the first report indicating the involvement of T3SS2 from V. parahaemolyticus in generating mitochondrial stress, its impact on the mitochondrial pore, and its effect on ATP production in human cells.
Keywords:
Vibrio parahaemolyticus
; T3SS2
; foodborne illness
; mitochondria
; cell death
1. Introduction
Vibrio parahaemolyticus is a marine pathogenic bacterium that has become the leading cause of gastroenteritis associated with the consumption of raw seafood worldwide [1]. In 1996, emerge the O3:K6 serotype strain of V. parahaemolyticus, now known as the pandemic clone, was responsible for significant gastroenteritis outbreaks globally [2]. In addition to the presence of thermostable direct hemolysin (TDH) and TDH-related hemolysin (TRH), other virulence factors have been described, such a Type III Secretion System on chromosome 1 (T3SS1) present in all V. parahaemolyticus strains [3]. Some strains associated with the pandemic clone, harbor a phylogenetic distinct T3SS named T3SS2 encoded in the pathogenicity island 7 of V. parahaemolyticus (VPaI-7) on chromosome 2 [3,4,5]. T3SSs are multiprotein nanomachines enable the direct delivery of effector proteins from the bacterial cytosol to the eukaryotic cell cytosol, allowing different effects on cells dependent on the repertoire of translocated effectors, which possess the ability to hijack signaling pathways and modify the activity of diverse host cell organelles, including mitochondria involved in crucial cellular functions such as immune response and energy production [6,7,8,9].
Mitochondria play a central role in regulating cellular functions and host responses to bacterial infections. They are dynamic organelles with a double membrane housing electron transport chain complexes and ATP synthase, adapting energy production based on cell needs [10]. Some effects described in bacterial infections are mitochondrial stress, loss of mitochondrial membrane potential (Δψm), depletion of ATP synthesis, high concentrations of reactive oxygen species (mtROS), and mitochondrial calcium overload leading to the opening of mitochondrial permeability transition pore (mPTP), which activates various programmed cell death pathways [11]. Therefore, it is not surprising that pathogenic bacteria use mitochondrially targeted effectors to control host cell death and immunity pathways, hijacking and subverting many different host molecules and organelles, including mitochondria [12,13,14,15].
Surprisingly, despite the above, only a few T3SS effectors that target host mitochondria and impact the mechanisms underlying their activity have been described or characterized. Among them, Escherichia coli EPEC/EHEC T3SS effectors target the mitochondrial pathway. The T3SS effector EspF localizes to mitochondria and is associated with mitochondrial dysfunction and tight junction disruption [12]. Additionally, the effector Map is a mitochondrial association protein imported via the classical TOM/Hsp70 import system and causes mitochondrial dysfunction [13]. In Yersinia pestis, YopH mediates mitochondrial-induced cell death of T lymphocytes and epithelial cells through unclear mechanisms [14]. On the other hand, Salmonella enterica mediates cytochrome c release and macrophage apoptosis through SipB activation of caspase-2 [15]. In the genus Vibrio, only the Vibrio cholerae T3SS effector protein VopE has been reported to be required to induce mitochondrial fragmentation and suppress innate immune responses during infection of cultured mammalian cells [8].
Despite in V. parahaemolyticus it has been described that the contribution of the T3SS2 is key to the colonizing the intestine and causing enteritis and diarrhea [16,17,18], the contribution in mitochondrial stress that leading cell death is not fully understood. To address this, we aimed to assess the contribution of V. parahaemolyticus T3SS2 to mitochondrial stress and cell death during infection in human intestinal cells. In this study, we found that the T3SS2 of V. parahaemolyticus induce sustained opening of the mitochondrial transition pore, altering ATP production and inducing cell death during infection. These observations suggest that this type of mitochondrial stress could be triggering cell death through T3SS2 during infection.
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
All strains of V. parahaemolyticus RIMD2210633 and its mutant of T3SS: Δvscn1 (T3SS2+) and Δvscn1 Δvscn2 (lacking both T3SSs or T3SS-), used in this study were donated by Dr. Carlos Blondel [19,20]. Strains were routinely cultured in Luria Bertani (LB) liquid medium or on LB agar plates at 37°C. Culture medium was supplemented with 0.04% bovine and ovine bile (Sigma, B8381) for the activation of T3SS2 [21] in infection assays.
2.2. Eukaryotic Cell Culture and Maintenance Conditions
HT-29 cells (ATCC HTB-38) were maintained in Dulbecco’s modified Eagle medium (DMEM) (Gibco) supplemented with 10% Fetal Bovine Serum (FBS) (Gibco) (DMEM−10% FBS) at 37°C in 5% CO2. Cells were grown at 37°C with 5% CO2 and routinely passaged at 70 to 80% confluence.
2.3. Mitochondrial Permeability Transition Pore (mPTP) Assay
After infection assay, we will use an Image-iT LIVE Mitochondrial Transition Pore Assay Kit (Thermo Fisher, I35103) to acquire fluorescence images with Lionheart-FX microscopy (Agilent BioTek). This kit utilizes 1 µM of Calcein-AM (green), which accumulates in the mitochondria of live cells and quenches when the mPTP is prolongedly open. As an experimental control, we use 0.5 µM of ionomycin, an ionophore that induces the opening of the mitochondrial pore, and 1 mM of Cobalt (Co+2) to quenches signal of Calcein-AM. The kit was employed according to the manufacturer's specifications, and cells were fixed with 4% formaldehyde (Thermo Fisher, 28908).
2.4. Mitochondrial Fragmentation
After the infection assay, we used 0.2 µM mitotracker for 15 minutes and the cells were fixed with 4% formaldehyde for imaging (Thermo Fisher, 28908) to visualize HT-29 mitochondria and acquire fluorescence images with Lionheart-FX microscopy (Agilent BioTek) to analyze the mitochondrial morphology.
2.5. ATP Determination Assay
The ATP Determination kit (Thermo Fisher, A22066) is a bioluminescence assay designed for the quantitative determination of ATP. It utilizes recombinant firefly luciferase and its substrate D-luciferin. The assay relies on luciferase's dependence on ATP to produce light, with an emission maximum at 560 nm. In the context of an infection study conducted at different time points (0, 1, 2, 3, 4, and 5 h post-infection), cells are lysed using a lysis buffer (HBSS 1x with 1% Triton-X100). The resulting supernatant is then used to measure ATP concentrations. ATP levels are expressed as concentrations in picomoles (pmol) per cell.
2.6. Infection with V. parahaemolyticus and T3SS2-Dependent Cell Death Assay
For cell survival assays, HT-29 cells were prepared by seeding 1.5 × 105 cells/well into 12-well plates with complete media. V. parahaemolyticus strains, cultured overnight, were diluted 1:100 into LB liquid media with 0.04% bile and grown until reaching an OD600 of 0.6. Infection was initiated at an MOI of 1, and cells were incubated at 37°C with 5% CO2. At each time point and individual plates (1, 2, 3, 4, and 5 h post-infection), the medium was replaced with fresh complete DMEM supplemented with 100 μg/mL gentamicin. Following overnight incubation, surviving cells were quantified using trypan blue exclusion (0.4% trypan blue) and counted on a hemocytometer (Neubauer cell chamber). To evaluate T3SS2-mediated cytotoxicity, we used the CellTox Green kit (Promega, G8741) through infection kinetics. HT-29 cells were seeded at 1.0 x 104 cells/well in 96-well plates with complete medium. Green Dye was added to the suspension at a final concentration of 1x. V. parahaemolyticus strains were cultured and T3SS2 was induced as described above. Cells were infected at an MOI of 1 and incubated at 37°C with 5% CO2 until 5 hpi. At each infection time evaluated (1, 2, 3, 4 and 5 hpi) the fluorescence of each condition evaluated was measured and the results were expressed as a Relative Fluorescence Unit (RFU).
3. Results
3.1. V. parahaemolyticus Induces mPTP opening T3SS2-Dependent Manner in Intestinal Cells
To determinate the contribution of the T3SS2 of V. parahaemolyticus in mitochondrial stress and cell death, we used a double mutant strain of RIMD2210633 lacking both T3SSs (Δvscn1 Δvscn2, T3SS-) and a strain containing functional T3SS2 (Δvscn1, T3SS2+), as previously described [20], in infection assay with HT-29 cells to evaluate the prolonged opening of the mPTP during infection (Figure 1).
First, we performed a validation of mPTP assay with imagen analysis by fluorescence with Calcein-AM, a cell permeable fluorophore that diffuses and gets trapped in all subcellular compartments such a mitochondria’s, in combination with treatment with cobalt solution (Co2+) a quenches calcein fluorescence [22]. To evaluate the mPTP in uninfected condition, the HT-29 cells were treated with Calcein-AM, Co+2 solution and ionomycin, a known ionophore to induce mPTP opening (stress condition), in contrast to cells without ionomycin (closed pore or healthy condition) (Figure 1A). Quantification of the intensity was carried out to compare these two states of the mitochondria, producing a decrease in intensity of Calcein-AM in the cells treated with ionomycin, due to the entry of cobalt into the mitochondria or quenching of mitochondrial calcein fluorescence, to demonstrating the opening of the pore (Figure 1B).
Once the mPTP opening assay was validated, we assessed the contribution of the T3SS2 of V. parahaemolyticus to the process of opening or closing the pore. In fluorescence analysis of infection of HT-29 cells with Δvscn1 strain (T3SS2+) led to a noticeable quenching of green fluorescence, starting from one-hour post-infection (hpi) and being progressive over time (Figure 1C). However, when we infected with a mutant strain unable to utilize any T3SSs (Δvscn1 Δvscn2) showed no fluorescence of Calcein-AM differences compared to uninfected cells (Figure 1C). Quantitative analysis of calcein fluorescence intensity from 150 cells images was performed, observing that the quenching of green fluorescence increases over time, these results being significant in comparison to uninfected cells (Figure 1D). These findings suggest that V. parahaemolyticus can induce T3SS2-dependent mitochondrial stress by maintaining prolonged mPTP opening during infection.
3.2. V. parahaemolyticus Has the Ability to Induce Mitochondrial Fragmentation, Disrupt ATP Production, and Trigger T3SS2-Dependent Cell Death during Infection
The maintenance of mitochondrial morphology and function relies on a delicate equilibrium between mitochondrial fusion and fission. Mitochondrial fragmentation can occur under cellular stress conditions, manifesting through an escalation in fission activity, an inhibition of fusion, or a combination of both mechanisms [23]. To investigate the role of V. parahaemolyticus T3SS2-induced mitochondrial fragmentation, we performed an infection assay with microscopically assess whether mitochondria network and morphology, where changes in their structures such as fission or separation of these and the swollen of the cell are signs of mitochondrial fragmentation (Figure 2). The mitochondrial morphology of the cells infected with a functional T3SS2 strain (Δvscn1, T3SS2+) was determined by the phenotype of the mitochondria labeled with mitotracker, which were rounding than uninfected control or double mutant strain (Δvscn1 Δvscn2, T3SS-), in addition a greater rounding and swelling morphology can be observed between fluorescent dots displayed an increased level of mitochondrial fragmentation (marked with the white arrows) (Figure 2A). To quantify these observations, we manually analyzed 50 cells per point from three independent assays and determined the number of cells exhibiting single rounding and swollen mitochondrial morphology. This analysis allowed us to determine the percentage of cells showing mitochondrial fragmentation. In Figure 2B, it is observed that cells treated with ionomycin as a control condition, a known inducer of mitochondrial fragmentation, present a significant increase in the percentage of cells with fragmented mitochondria. For its part, the infection with functional T3SS2 strain (Δvscn1, T3SS2+), showed significant percentage of fragmentation from 2 hpi and with progressive accumulation in compared with uninfected or double mutant strain (Δvscn1 Δvscn2, T3SS-) (Figure 2B).
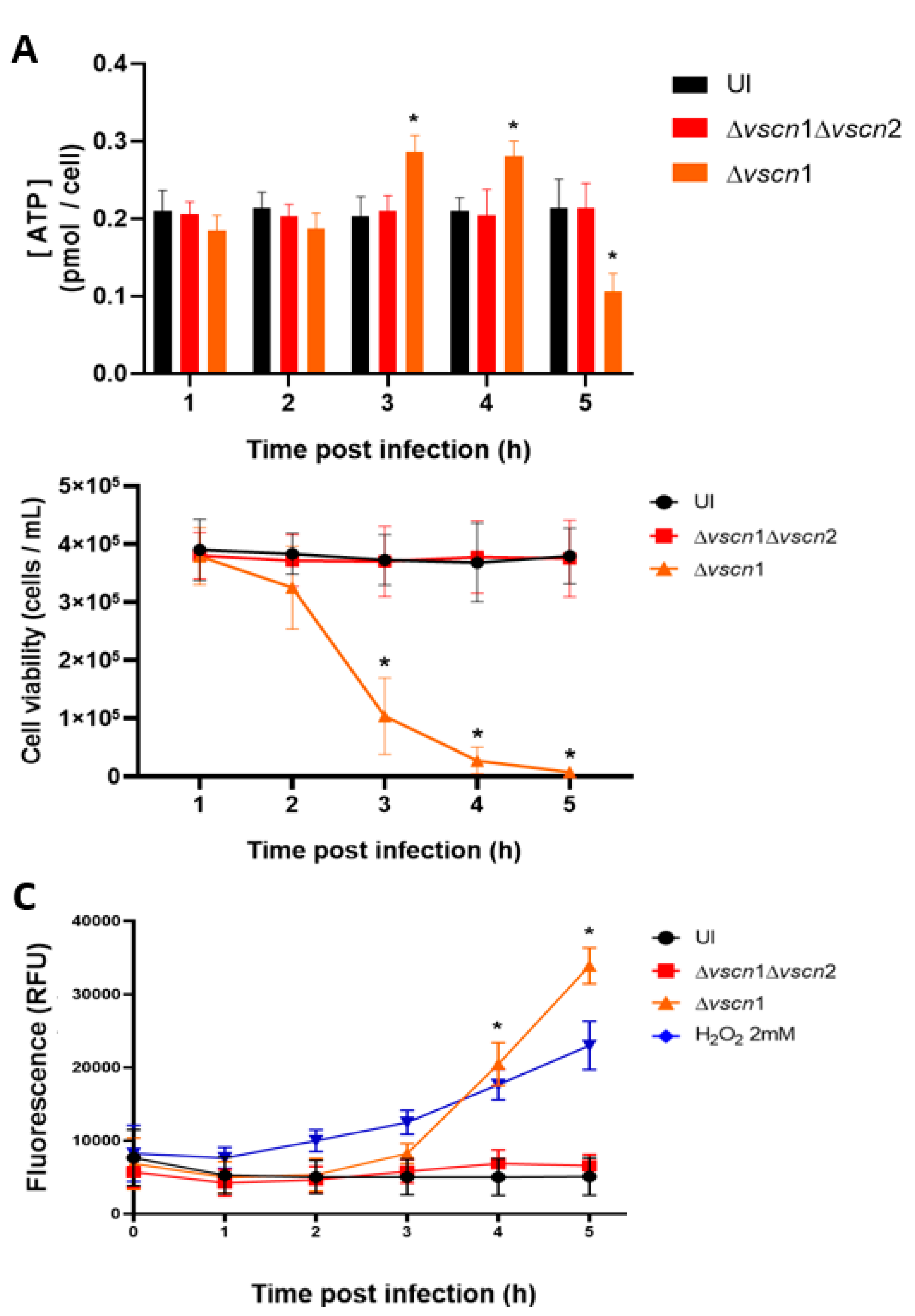
Considering that the main source of energy cell comes from the mitochondria, we aimed to assess whether the mitochondrial stress induced by T3SS2 could disrupt a normal ATP production during the infection process. Our observations revealed that in HT-29 cells infected with the strain possessing a functional T3SS2, there was an increase in ATP production by bioluminescence assay at 3 and 4 hpi, significantly higher than initial ATP levels, followed by a decline at 5 hpi (Figure 3A). Being a classical cellular behavior in the process of cell death, as it has been reported that an increase in ATP production with a subsequent decrease in ATP production is the classic behavior during cell death [24]. On the other hand, the double mutant strain (Δvscn1 Δvscn2, T3SS-) did not exhibit alterations in ATP production during infection, displaying a comparable effect to uninfected cells (Figure 3A).
As observed, V. parahaemolyticus can induce early mitochondrial stress in a T3SS2-dependent manner during infection (Figure 2A,B). This is particularly relevant considering previous reports partially linking V. parahaemolyticus-induced cell death to T3SS2 functionality [20,25]. In our case, we performed an infection kinetics test to evaluate cell viability by exclusion of trypan blue, and we also evaluated the integrity of the membrane using the CellTox Green kit to detect the DNA released into the supernatant, in order to verify viability of the cells and what was observed for mitochondrial stress and ATP generation.
We observed that the contribution of T3SS2 in the generation of cell death was consistent with recently reported data. As expected, HT-29 cells infected with a V. parahaemolyticus double mutant strain (Δvscn1 Δvscn2; T3SS-) did not show differences in cellular viability during the infection process, similar to the uninfected cells (Figure 3B). In turn, fluorescence due to DNA release was not increased in the infection process (Figure 3C). In contrast, a functional V. parahaemolyticus T3SS2 (Δvscn1, T3SS2+) decreases cellular viability (Figure 3B) and induces the release of eukaryotic DNA (reduction of membrane integrity) from infected cells, a phenotype comparable to cells treated with H2O2 as a classical inducer of cell death (Figure 3C). These viability results are consistent with what was observed by other authors, where the contribution of T3SS2 was shown in a significant decrease in cell viability in human intestinal cells [19,26].
All these results suggest that the T3SS2 of V. parahaemolyticus induces changes in the morphology and networks of mitochondria, leading to fragmentation and swollen in the early stages of infection and these alterations have consequences on ATP production at 3 and 4 hpi, promoting cell death during the infection.
4. Discussion
In the present work, we have investigated the contribution of T3SS2 in mitochondrial stress that leads to cell death during V. parahaemolyticus infection, being the first report that supports that T3SS2 is a determinant for generating mitochondrial dysfunction and important for the pathogenesis of V. parahaemolyticus. Mitochondria are intimately involved in the regulation of intracellular Ca2+ fluxes and contain a refined molecular machinery that precipitates regulated cell death via mPTP-dependent regulated [27].
To evaluate the contribution of V. parahaemolyticus T3SS2 to the sustained opening of mPTP (as a mitochondrial stress signal), we employed fluorescence microscopy using HT-29 loaded with Calcein-AM and Co2+ after infection. Calcein-AM is a cell permeable fluorophore that diffuses and gets trapped in all subcellular compartments, including mitochondria [22]. Treatment with Co2+ quenches Calcein fluorescence in all subcellular compartments except the mitochondrial matrix which is enclosed by a Co2+ impermeable inner mitochondrial membrane when mPTP is closed. Thus, the ability of Co2+ to quench mitochondrial Calcein fluorescence only when mPTP is open allows determination of open vs closed status of mPTP in the cell [22]. In our experiments, we observed that the effect of V. parahaemolyticus T3SS2 over de mitochondrial health, demonstrated the ability to induce mitochondrial stress through mPTP opening (Figure 1). Other pathogen such Mycobacterium tuberculosis also generate mPTP-dependent dissipation of Δψm to leading necrosis in macrophages [28].
The central role of mitochondria in the host's response to bacterial infections has become a fundamental area of research because they play critical roles in regulating energy production, proinflammatory response, defense against pathogenic infections, and cell death [10,29,30]. To evaluate the contribution of V. parahaemolyticus T3SS2 in perturbing ATP production by mitochondrial stress, we employed fluorescence microscopy to assess mitochondrial fragmentation (mitochondrial fission) and ATP quantification by bioluminescence using human intestinal HT-29 cells at different time post infection. Our results showed that T3SS2 of V. parahaemolyticus contributes to generating mitochondrial fragmentation (Figure 2) and alterations in ATP production, particularly generating an increase in ATP production and then decreasing over time (Figure 3A). Our results have been consistent with the data reported for Listeria monocytogenes that causes dramatic alterations of mitochondrial dynamics via listeriolysin-O (LLO), generating mitochondrial fragmentation induced by Listeria infection [31,32]. Furthermore, has been reporter that Legionella pneumophila abrogate oxidative phosphorylation (OXPHOS) and then, through unknown mechanisms, enhances cellular glycolysis, thereby promoting a metabolic shift known as the Warburg effect [33]. This alteration favors bacterial replication, possibly by reducing the production of antibacterial mitochondrial reactive oxygen species (ROS) [33]. It has been reported that V. cholerae also induces mitochondrial bioenergetic dysfunction by ROS generation through its virulence factors GbpA and cholix toxin [34,35]. On the other hand, mitochondrial dynamics (Fission and fusion process) are essential for metabolic activity, where it has been well related that the mitochondrial fusion process generates the greatest efficiency of OXPHOS and increases the ATP production, on the contrary, mitochondrial fission would decrease the ability to generate ATP [36]. Based on this background, we can observe that our results are similar, since we have shown that the T3SS2 of V. parahaemolyticus, in addition to generating prolonged opening of the mPTP (Figure 1), generates mitochondrial fission (Figure 2) and a temporary increase in production of ATP from the infected cells, and then its production declines over time (Figure 3A).
The control of mitochondrial health during infection that leads to cell death has already been reported in other pathogenic bacteria. E. coli EHEC secretes the T3SS effectors Map and EspF, which are targeted to the mitochondria though mitochondrial targeting sequences (MASV) to disrupt morphology, perturb calcium homeostasis, and trigger apoptosis [37]. On the other hand, the T3SS effector protein VopE of V. cholerae contains a mitochondrial targeting sequence at its amino terminus that allows precise targeting to mitochondria by hijacking the mitochondrial import machinery [8]. Interestingly, VopE interacts with the GTPase Miro at the mitochondrial outer membrane that was found to perturb mitochondrial perinuclear clustering, which is needed to activate MAVS-mediated NF-kB signaling, an important contributor to the host inflammatory response [8].
Particularly, it has been reported that the T3SS2 of V. parahaemolyticus contributes to generating cell death during infection [6,7,8,9]. However, to date the mechanism involved has not been completely understood, such as the process that leads to cell death, such as the type of cell death generated. In the context of bacterial infections inducing mitochondrial stress, alterations in ATP synthesis and loss of Δψm leading to mPTP opening trigger the activation of various programmed cell death pathways [11]. It has been reported that mitochondrial stress during the infection of various bacterial pathogens can lead to cell death, including sustained opening of the mitochondrial transition pore and mitochondrial fragmentation causing ATP production alterations [38,39,40]. It has been shown that two different types of cell death occur in Jurkat cells, with the type being determined by the intracellular ATP concentration [40]. The authors observed the concentration of intracellular ATP to act as a molecular switch controlling the type of cell death. Indeed, they reported that with a high concentration of intracellular ATP leading to apoptotic cell death, while a low concentration of intracellular ATP leading to necrotic cell death. A high level of ATP is generated by cells in response to stress for the activation of proteins, such as caspases, and DNA repair, necessary conditions for apoptosis generation [24]. In contrast, necrosis is characterized by a significant decrease in ATP generation responding to electron transport chain uncoupling [41]. These features of necrotic cell death are similar to showed by our results (Figure 3), where the T3SS2 of V. parahaemolyticus generates first an increase and then a decrease in ATP concentrations (Figure 3A), generating a decrease in cell viability (Figure 3B) and damaging the integrity of the cell membrane from 3 hpi (Figure 3C), evaluated by increasing the signal of a fluorescent probe that can enter cells with a compromised membrane, suggesting that the T3SS2 of V. parahaemolyticus would generate mitochondrial stress that leads to necrotic cell death.
In summary, our work identifies for the first time a contribution of V. parahaemolyticus T3SS2 in generating mitochondrial stress during infection. Particularly, generating a sustained opening of the mitochondrial transition pore and depletion in the generation of cellular ATP, similar to necrotic cell death. Future molecular studies on the possible T3SS2 effector proteins involved are required and will be valuable to define the role of these virulence factors in the pathogenicity of V. parahaemolyticus.
5. Conclusions
Overall, our results indicate that the T3SS2 of V. parahaemolyticus is essential to generate mitochondrial stress and cell death during infection. To the best of our knowledge, this is the first report of the relevance of the T3SS2 of V. parahaemolyticus in the mitochondrial stress generation.
Author Contributions
Conceptualization: Í.M.U. and N.P.; methodology and formal analysis: Í.M.U. and N.P.; Sample recollection: Í.M.U. and N.P. Data curation: Í.M.U, N.P. D.P-R, G.C. and K.G; writing original draft preparation Í.M.U. and N.P.; writing review and editing: Í.M.U, N.P. D.P-R, G.C. and K.G Funding acquisition, project administration, and supervision: Í.M.U. and N.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by FONDECYT grant 3200874 (ANID) to I.M.U. N.P. is supported by FONDECYT grant 11231192 (ANID). K.G. is supported by FONDECYT grant 1190957 (ANID). D.P-R is supported by Research Direction grant DIUAV 02-2022 (VRID of Universidad Autónoma de Chile). GC is supported by Research Direction grant DIU 260-2023 (VRID of Universidad Autónoma de Chile).
Acknowledgments
We want to thank Dr. Carlos Blondel's laboratory for kindly providing us the different strains of Vibrio parahaemolyticus used in this work.
References
- Letchumanan, V.; Chan, K.-G.; Lee, L.-H. Vibrio parahaemolyticus: a review on the pathogenesis, prevalence, and advance molecular identification techniques. Front. Microbiol. 2014, 5, 705. [Google Scholar] [CrossRef]
- Velazquez-Roman, J.; León-Sicairos, N.; Hernández-Díaz, L.d.J.; Canizalez-Roman, A. Pandemic Vibrio parahaemolyticus O3:K6 on the American continent. Front. Cell. Infect. Microbiol. 2014, 3, 110. [Google Scholar] [CrossRef]
- Makino, K.; Oshima, K.; Kurokawa, K.; Yokoyama, K.; Uda, T.; Tagomori, K.; Iijima, Y.; Najima, M.; Nakano, M.; Yamashita, A.; et al. Genome sequence of Vibrio parahaemolyticus: a pathogenic mechanism distinct from that of V cholerae. Lancet 2003, 361, 743–749. [Google Scholar] [CrossRef]
- Abby, S.S.; Rocha, E.P.C. The Non-Flagellar Type III Secretion System Evolved from the Bacterial Flagellum and Diversified into Host-Cell Adapted Systems. PLOS Genet. 2012, 8, e1002983. [Google Scholar] [CrossRef]
- Xu, F.; Gonzalez-Escalona, N.; Drees, K.P.; Sebra, R.P.; Cooper, V.S.; Jones, S.H.; Whistler, C.A. Parallel Evolution of Two Clades of an Atlantic-Endemic Pathogenic Lineage of Vibrio parahaemolyticus by Independent Acquisition of Related Pathogenicity Islands. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef]
- Lara-Tejero, M.; Galán, J.E. The Injectisome, a Complex Nanomachine for Protein Injection into Mammalian Cells. EcoSal Plus 2019, 8, 245–259. [Google Scholar] [CrossRef]
- Portaliou, A.G.; Tsolis, K.C.; Loos, M.S.; Zorzini, V.; Economou, A. Type III Secretion: Building and Operating a Remarkable Nanomachine. Trends Biochem. Sci. 2015, 41, 175–189. [Google Scholar] [CrossRef]
- Suzuki, M.; Danilchanka, O.; Mekalanos, J.J. Vibrio cholerae T3SS Effector VopE Modulates Mitochondrial Dynamics and Innate Immune Signaling by Targeting Miro GTPases. Cell Host Microbe 2014, 16, 581–591. [Google Scholar] [CrossRef]
- Nandi, I.; Aroeti, L.; Ramachandran, R.P.; Kassa, E.G.; Zlotkin-Rivkin, E.; Aroeti, B. Type III secreted effectors that target mitochondria. Cell. Microbiol. 2021, 23, e13352. [Google Scholar] [CrossRef]
- H. Chen, D.C. H. Chen, D.C. Chan, Mitochondrial dynamics in mammals, Curr Top Dev Biol 59 (2004) 119-44.
- Ramaccini, D.; Montoya-Uribe, V.; Aan, F.J.; Modesti, L.; Potes, Y.; Wieckowski, M.R.; Krga, I.; Glibetić, M.; Pinton, P.; Giorgi, C.; et al. Mitochondrial Function and Dysfunction in Dilated Cardiomyopathy. Front. Cell Dev. Biol. 2021, 8. [Google Scholar] [CrossRef]
- P. Dean, M. P. Dean, M. Maresca, S. Schuller, A.D. Phillips, B. Kenny, Potent diarrheagenic mechanism mediated by the cooperative action of three enteropathogenic Escherichia coli-injected effector proteins, Proc Natl Acad Sci U S A 103(6) (2006) 1876-81.
- Dean, P.; Kenny, B. The effector repertoire of enteropathogenic E. coli: ganging up on the host cell. Curr. Opin. Microbiol. 2009, 12, 101–109. [Google Scholar] [CrossRef]
- Bruckner, S.; Rhamouni, S.; Tautz, L.; Denault, J.-B.; Alonso, A.; Becattini, B.; Salvesen, G.S.; Mustelin, T. Yersinia Phosphatase Induces Mitochondrially Dependent Apoptosis of T Cells. J. Biol. Chem. 2005, 280, 10388–10394. [Google Scholar] [CrossRef]
- V. Jesenberger, K.J. V. Jesenberger, K.J. Procyk, J. Yuan, S. Reipert, M. Baccarini, Salmonella-induced caspase-2 activation in macrophages: a novel mechanism in pathogen-mediated apoptosis, J Exp Med 192(7) (2000) 1035-46.
- Hubbard, T.P.; Chao, M.C.; Abel, S.; Blondel, C.J.; Wiesch, P.A.Z.; Zhou, X.; Davis, B.M.; Waldor, M.K. Genetic analysis of Vibrio parahaemolyticus intestinal colonization. Proc. Natl. Acad. Sci. 2016, 113, 6283–6288. [Google Scholar] [CrossRef]
- Piñeyro, P.; Zhou, X.; Orfe, L.H.; Friel, P.J.; Lahmers, K.; Call, D.R. Development of Two Animal Models To Study the Function of Vibrio parahaemolyticus Type III Secretion Systems. Infect. Immun. 2010, 78, 4551–4559. [Google Scholar] [CrossRef]
- Ritchie, J.M.; Rui, H.; Zhou, X.; Iida, T.; Kodoma, T.; Ito, S.; Davis, B.M.; Bronson, R.T.; Waldor, M.K. Inflammation and Disintegration of Intestinal Villi in an Experimental Model for Vibrio parahaemolyticus-Induced Diarrhea. PLOS Pathog. 2012, 8, e1002593. [Google Scholar] [CrossRef] [PubMed]
- Blondel, C.J.; Park, J.S.; Hubbard, T.P.; Pacheco, A.R.; Kuehl, C.J.; Walsh, M.J.; Davis, B.M.; Gewurz, B.E.; Doench, J.G.; Waldor, M.K. CRISPR/Cas9 Screens Reveal Requirements for Host Cell Sulfation and Fucosylation in Bacterial Type III Secretion System-Mediated Cytotoxicity. Cell Host Microbe 2016, 20, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Gewurz, B.E.; Ritchie, J.M.; Takasaki, K.; Greenfeld, H.; Kieff, E.; Davis, B.M.; Waldor, M.K. A Vibrio parahaemolyticus T3SS Effector Mediates Pathogenesis by Independently Enabling Intestinal Colonization and Inhibiting TAK1 Activation. Cell Rep. 2013, 3, 1690–1702. [Google Scholar] [CrossRef]
- Gotoh, K.; Kodama, T.; Hiyoshi, H.; Izutsu, K.; Park, K.-S.; Dryselius, R.; Akeda, Y.; Honda, T.; Iida, T. Bile Acid-Induced Virulence Gene Expression of Vibrio parahaemolyticus Reveals a Novel Therapeutic Potential for Bile Acid Sequestrants. PLOS ONE 2010, 5, e13365. [Google Scholar] [CrossRef] [PubMed]
- Petronilli, V.; Miotto, G.; Canton, M.; Colonna, R.; Bernardi, P.; Di Lisa, F. Imaging the mitochondrial permeability transition pore in intact cells. BioFactors 1998, 8, 263–272. [Google Scholar] [CrossRef]
- Sprenger, H.-G.; Langer, T. The Good and the Bad of Mitochondrial Breakups. Trends Cell Biol. 2019, 29, 888–900. [Google Scholar] [CrossRef]
- Zamaraeva, M.V.; Sabirov, R.Z.; Maeno, E.; Ando-Akatsuka, Y.; Bessonova, S.V.; Okada, Y. Cells die with increased cytosolic ATP during apoptosis: a bioluminescence study with intracellular luciferase. Cell Death Differ. 2005, 12, 1390–1397. [Google Scholar] [CrossRef]
- Matlawska-Wasowska, K.; Finn, R.; Mustel, A.; O'Byrne, C.P.; Baird, A.W.; Coffey, E.T.; Boyd, A. The Vibrio parahaemolyticus Type III Secretion Systems manipulate host cell MAPK for critical steps in pathogenesis. BMC Microbiol. 2010, 10, 329–329. [Google Scholar] [CrossRef]
- Plaza, N.; Urrutia, I.M.; Garcia, K.; Waldor, M.K.; Blondel, C.J. Identification of a Family of Vibrio Type III Secretion System Effectors That Contain a Conserved Serine/Threonine Kinase Domain. mSphere 2021, 6, e0059921. [Google Scholar] [CrossRef]
- Marchi, S.; Morroni, G.; Pinton, P.; Galluzzi, L. Control of host mitochondria by bacterial pathogens. Trends Microbiol. 2021, 30, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Gan, H.; Remold, H.G. A Mechanism of Virulence: Virulent Mycobacterium tuberculosis Strain H37Rv, but Not Attenuated H37Ra, Causes Significant Mitochondrial Inner Membrane Disruption in Macrophages Leading to Necrosis. J. Immunol. 2006, 176, 3707–3716. [Google Scholar] [CrossRef]
- Stavru, F.; Palmer, A.E.; Wang, C.; Youle, R.J.; Cossart, P. Atypical mitochondrial fission upon bacterial infection. Proc. Natl. Acad. Sci. USA 2013, 110, 16003–16008. [Google Scholar] [CrossRef]
- Vringer, E.; Tait, S.W.G. Mitochondria and cell death-associated inflammation. Cell Death Differ. 2022, 30, 304–312. [Google Scholar] [CrossRef]
- Carvalho, F.; Spier, A.; Chaze, T.; Matondo, M.; Cossart, P.; Stavru, F. Listeria monocytogenes Exploits Mitochondrial Contact Site and Cristae Organizing System Complex Subunit Mic10 To Promote Mitochondrial Fragmentation and Cellular Infection. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Stavru, F.; Cossart, P. Listeria infection modulates mitochondrial dynamics. Commun. Integr. Biol. 2011, 4, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Escoll, P.; Song, O.-R.; Viana, F.; Steiner, B.; Lagache, T.; Olivo-Marin, J.-C.; Impens, F.; Brodin, P.; Hilbi, H.; Buchrieser, C. Legionella pneumophila Modulates Mitochondrial Dynamics to Trigger Metabolic Repurposing of Infected Macrophages. Cell Host Microbe 2017, 22, 302–316. [Google Scholar] [CrossRef]
- Mandal, S.; Chatterjee, N.S. Vibrio cholerae GbpA elicits necrotic cell death in intestinal cells. J. Med Microbiol. 2016, 65, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Ogura, K.; Terasaki, Y.; Miyoshi-Akiyama, T.; Terasaki, M.; Moss, J.; Noda, M.; Yahiro, K. Vibrio cholerae Cholix toxin-induced HepG2 cell death is enhanced by tumor necrosis factor-alpha through ROS and intracellular signal-regulated kinases. Toxicol. Sci. 2017, 156, 455–468. [Google Scholar] [CrossRef]
- Chen, W.; Zhao, H.; Li, Y. Mitochondrial dynamics in health and disease: mechanisms and potential targets. Signal Transduct. Target. Ther. 2023, 8, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, R.P.; Spiegel, C.; Keren, Y.; Danieli, T.; Melamed-Book, N.; Pal, R.R.; Zlotkin-Rivkin, E.; Rosenshine, I.; Aroeti, B. Mitochondrial Targeting of the Enteropathogenic Escherichia coli Map Triggers Calcium Mobilization, ADAM10-MAP Kinase Signaling, and Host Cell Apoptosis. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Karbowski, M.; Youle, R.J. Dynamics of mitochondrial morphology in healthy cells and during apoptosis. Cell Death Differ. 2003, 10, 870–880. [Google Scholar] [CrossRef]
- K.W. Kinnally, P.M. K.W. Kinnally, P.M. Peixoto, S.Y. Ryu, L.M. Dejean, Is mPTP the gatekeeper for necrosis, apoptosis, or both?, Biochim Biophys Acta 1813(4) (2011) 616-22.
- Leist, M.; Single, B.; Castoldi, A.F.; Kühnle, S.; Nicotera, P. Intracellular Adenosine Triphosphate (ATP) Concentration: A Switch in the Decision Between Apoptosis and Necrosis. J. Exp. Med. 1997, 185, 1481–1486. [Google Scholar] [CrossRef]
- Chen, D.; Yu, J.; Zhang, L. Necroptosis: an alternative cell death program defending against cancer. Biochim. et Biophys. Acta (BBA) - Rev. Cancer 2016, 1865, 228–236. [Google Scholar] [CrossRef]
Figure 1.
V. parahaemolyticus induce mPTP opening in intestinal cells T3SS2-dependent. (A, C) The HT-29 cells were treated with ionomycin (a mPTP opening inducer), uninfected as a control condition (UI) or infected with V. parahaemolyticus strains at different hour post infection. In the figures, green color indicates the mPTP are closed (healthy mitochondria), red color shows the mitochondrial mass (stress condition), and blue color represents the DAPI stained nuclei. (B, D) indicate the mean fluorescence intensity units of green channel. Representative images from 3 independent trials are shown. Statistical significance of differences in calcein fluorescence comparing different conditions with uninfected cells was determined by one-way ANOVA with Dunnett's test (∗P < 0.05).
Figure 1.
V. parahaemolyticus induce mPTP opening in intestinal cells T3SS2-dependent. (A, C) The HT-29 cells were treated with ionomycin (a mPTP opening inducer), uninfected as a control condition (UI) or infected with V. parahaemolyticus strains at different hour post infection. In the figures, green color indicates the mPTP are closed (healthy mitochondria), red color shows the mitochondrial mass (stress condition), and blue color represents the DAPI stained nuclei. (B, D) indicate the mean fluorescence intensity units of green channel. Representative images from 3 independent trials are shown. Statistical significance of differences in calcein fluorescence comparing different conditions with uninfected cells was determined by one-way ANOVA with Dunnett's test (∗P < 0.05).
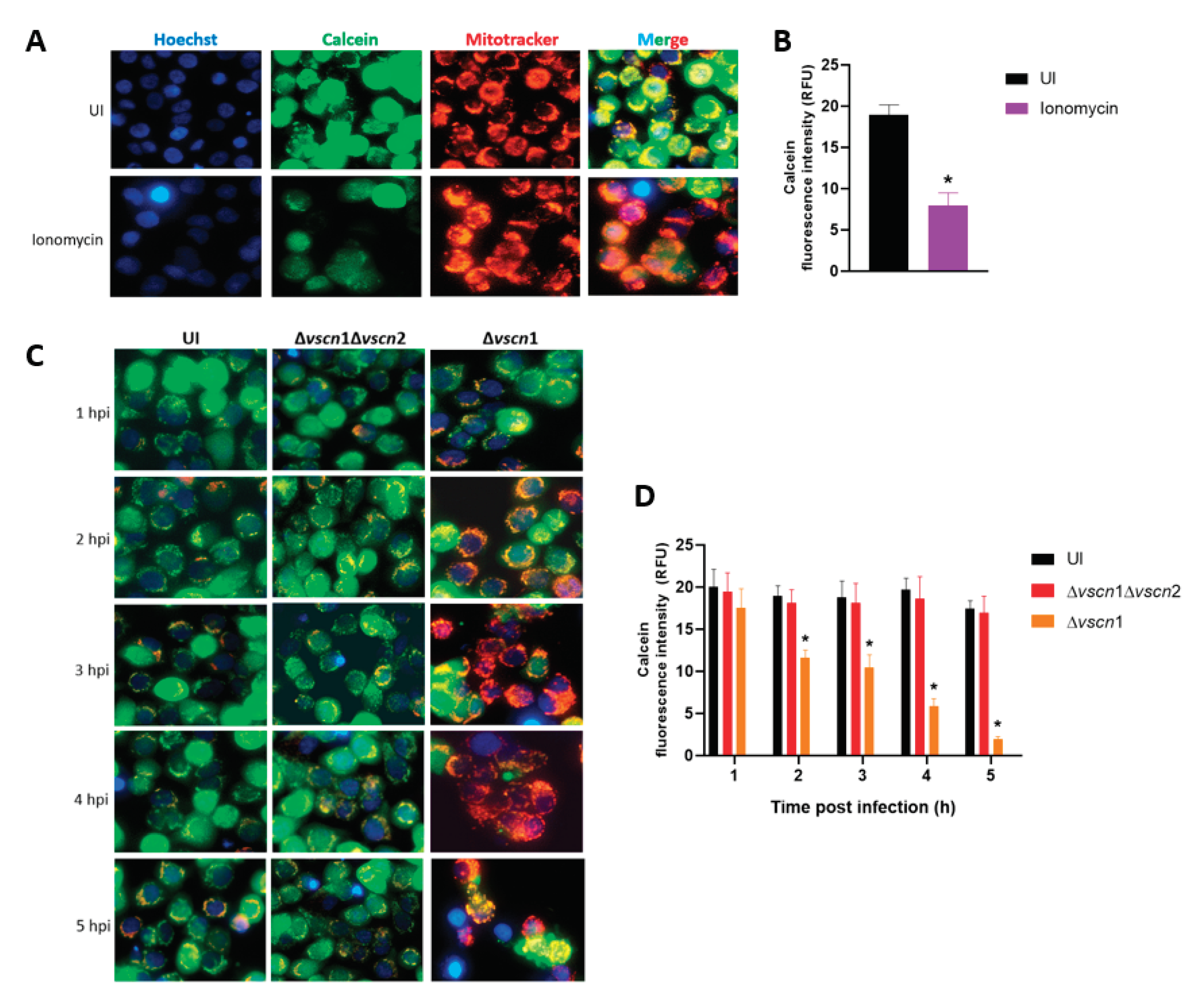
Figure 2.
Mitochondrial fragmentation T3SS2-dependent during V. parahaemolyticus infection. HT-29 cells were infected with different V. parahaemolyticus strains or uninfected as a control condition. (A) Mitochondria network morphology was analysis by microscopy. White arrows show cells with mitochondrial fragmentation. (B) Percentage of cells with fragmented mitochondria as described for panel A. Representative images from three independent assays are shown. Statistical significance of differences in percentage of cellular with fragmented mitochondria was determined using a one-way ANOVA with Dunnett’s test (∗: P < 0.05 compared each condition with uninfected cells at corresponding times).
Figure 2.
Mitochondrial fragmentation T3SS2-dependent during V. parahaemolyticus infection. HT-29 cells were infected with different V. parahaemolyticus strains or uninfected as a control condition. (A) Mitochondria network morphology was analysis by microscopy. White arrows show cells with mitochondrial fragmentation. (B) Percentage of cells with fragmented mitochondria as described for panel A. Representative images from three independent assays are shown. Statistical significance of differences in percentage of cellular with fragmented mitochondria was determined using a one-way ANOVA with Dunnett’s test (∗: P < 0.05 compared each condition with uninfected cells at corresponding times).
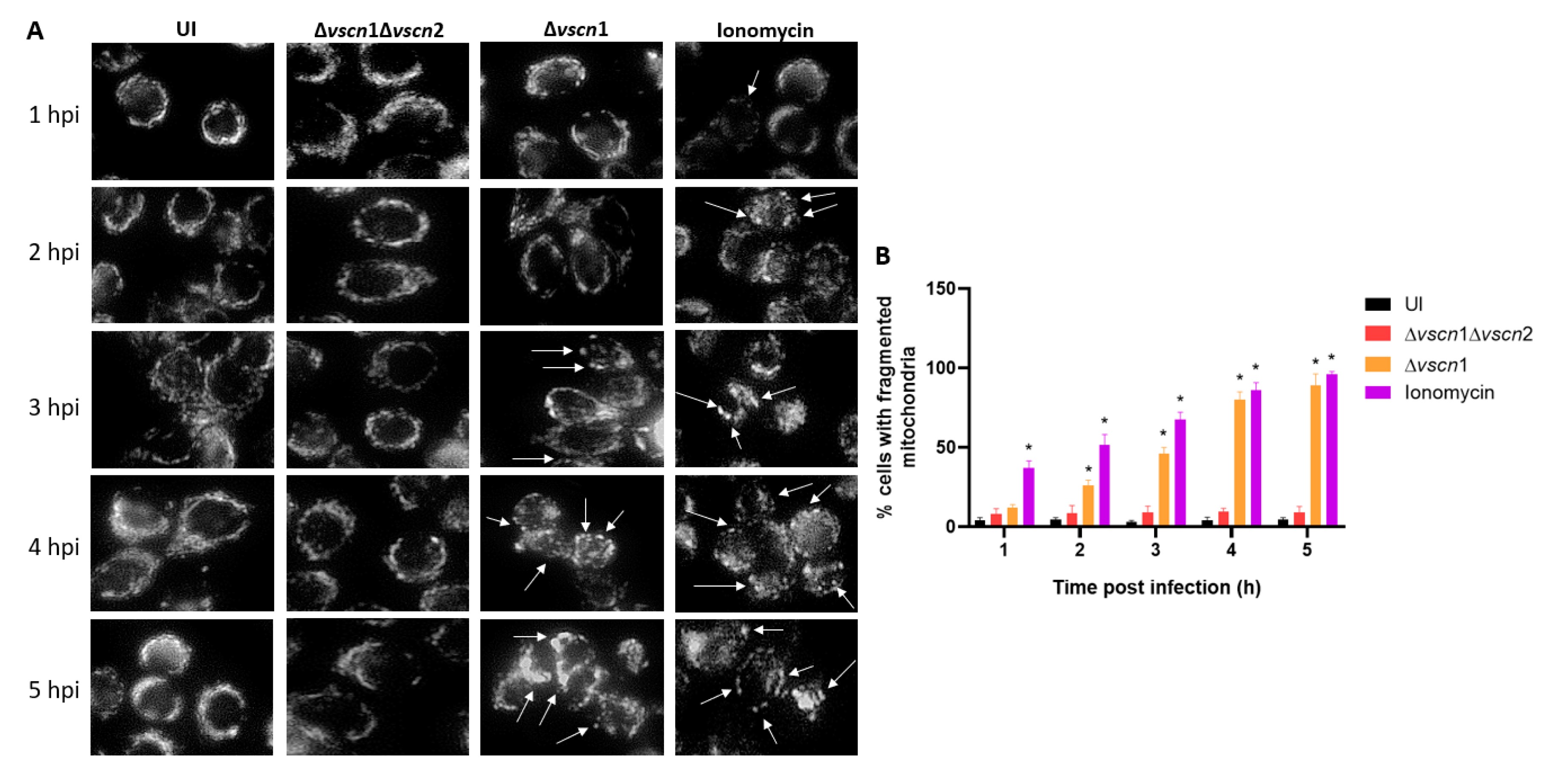
Figure 3.
ATP production and T3SS2-dependent cell death during V. parahaemolyticus infection. HT-29 cells were infected with different V. parahaemolyticus strains or uninfected to evaluate ATP production and cell death. (A) The ATP assay was conducted using a bioluminescent assay in cell lysates at the end of each hpi. The values were normalized based on total cell viability. (B) Cell viability was evaluated by trypan blue exclusion assay at the indicated times post infection. (C) Membrane integrity was evaluated by fluorescence tests at the indicated times post infection. Statistical significance comparing different conditions with uninfected cells at corresponding times was determined by one-way ANOVA with Dunnett's test (∗: P < 0.05).
Figure 3.
ATP production and T3SS2-dependent cell death during V. parahaemolyticus infection. HT-29 cells were infected with different V. parahaemolyticus strains or uninfected to evaluate ATP production and cell death. (A) The ATP assay was conducted using a bioluminescent assay in cell lysates at the end of each hpi. The values were normalized based on total cell viability. (B) Cell viability was evaluated by trypan blue exclusion assay at the indicated times post infection. (C) Membrane integrity was evaluated by fluorescence tests at the indicated times post infection. Statistical significance comparing different conditions with uninfected cells at corresponding times was determined by one-way ANOVA with Dunnett's test (∗: P < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



