Submitted:
08 March 2024
Posted:
08 March 2024
You are already at the latest version
Abstract
Salidroside, a valuable phenylethanoid glycoside, is obtained from plants belonging to the Rhodiola genus, known for its diverse biological properties. At present, salidroside is still far from large-scale industrial production due to the lower titer and higher process cost. In this study, we constructed a biosynthetic pathway from L-tyrosine to salidroside in Saccharomyces cerevisiae. To increase sali-droside production, we for the first time constructed an in vivo UDP-glucose regeneration system that sucrose synthase GmSUS worked in conjunction with UDP-glucose transferase RrUGT33 to improve the UDP-glucose availability with inexpensive sucrose as the consumed substrate. Finally, the molar conversion rate of L-tyrosine could reach 95%. To further reduce the cost, co-culture with E. coli that synthesized L-tyrosine de novo resulted in an 81% reduction in the final substrate cost per 1 kg of salidroside compared to the initial cost. The final titer of salidroside obtained 3.8 g/L and was the highest titer achieved at the Shake-Flask-Level in the existing reports. And this marked the first successful synthesis of salidroside in an in situ enhanced UDP-glucose system using sucrose. This accomplishment laid a robust groundwork for further investigations into the synthesis of other notable glycosides and natural compounds.
Keywords:
Salidroside
; Uridine diphosphate glucuronyl-transferase
; UDP-glucose regeneration
; Metabolic engineering
; E. coli-S. cerevisiae co-culture
1. Introduction
Glycosides, widely distributed and exhibiting significant diversity in plants, represent a prevalent and crucial class of compounds in nature. The utilization of plant secondary metabolites as aglycones for glycoside formation via glycosylation frequently results in substantial modifications to solubility, biological effects, or stability [1]. Salidroside is a polyphenolic compound that is soluble in water derived from Rhodiola species with significant biological activities and health-enhancing effects, including anti-fatigue, anti-stress, anti-radiation, anti-inflammatory, and the ability to prevent cardiovascular, neurodegenerative, and liver diseases [2,3,4]. Rhodiola plants are acknowledged as the primary reservoirs of salidroside [2]. However, the increasing commercial demand for salidroside from natural plants is challenging to meet due to the stringent growth requirements, extended growth cycles, and low salidroside concentration [5]. To tackle this issue, a range of biotechnological methods and microbial engineering techniques have been swiftly advanced to boost the production of salidroside.
Numerous successful tactics involving operon genes have been employed within the field of microbial metabolic engineering to elevate the ultimate titer of tyrosol and salidroside. These include pathway reconstruction through the incorporation of foreign genes, augmentation of metabolic flux by overexpressing or co-expressing pivotal genes, or suppression of metabolic flux [6,7,8,9,10]. By conducting these studies, the production levels of tyrosol using sugar (such as glucose, or a mixture of xylose and glucose) as the carbon source have been significantly enhanced, with titers increasing from 69.08 mg/L to 1.47 g/L [11,12]. Tyrosol titers in Saccharomyces cerevisiae were increased to 927.7 mg/L in the shake flask and 8.37 g/L in the bioreactor [6,7]. UDP-glucosyltransferase (UGT) is the critical enzyme in the downstream pathway for the transformation from tyrosol to salidroside. Much work has been done to mine efficient glycosyltransferases because suitable UGT genes are not available in either Escherichia coli or Saccharomyces cerevisiae [7,13]. Torrens-Spence et al. conducted a functional analysis of 34 UGT genes to unveil the entire synthetic route of salidroside biosynthesis in Rhodiola. [14]. Consequently, it was discovered that UGTs (RrUGT 17, 29, 32, and 33) exhibited site-specific T8GT activity, converting tyrosol into salidroside, where RrUGT33 demonstrated the most pronounced activity. To date, the highest titers of tyrosol and salidroside were produced in a 5 L bioreactor by collaborative engineering strategy in S. cerevisiae metabolic engineering, which were about 10 g/L and 26.55 g/L, respectively [15]. In addition to regulating genes with specific functions, fusion engineering of microbial co-culture is also a technology with great potential. Through the utilization of a co-culture strategy, salidroside production increased from 56.9 mg/L to 670.6 mg/L in shake flasks and further escalated to 6.03 g/L in a 5 L bioreactor [11]. Existing studies have focused on increasing salidroside production by increasing tyrosol production and mining efficient UGT enzymes. However, blindly pursuing a high tyrosol titer will produce toxic effects on the strain, negatively affect its growth and metabolism, and ultimately not be conducive to the production of the final product, salidroside. Moreover, it is difficult to fundamentally improve the titer of salidroside by mining only the efficient UGT enzyme without sufficient sugar donor UDP-glucose.
Within the cytoplasm of the engineered E. coli, the inadequate UDP-glucose supply constrains the efficiency of glycosylation. In addition, the low production of glycosylated products was also due to the absence of biosynthetic pathways for other nucleoside sugars or the significantly inadequate supply. Various approaches have been employed to increase the intracellular UDP-glucose reservoir, including the elimination of competitive genes such as pgi, zwf, or ushA [16,17], and the upregulation of crucial genes in the UDP-glucose production pathway. Nevertheless, not every gene overexpression tactic yields favorable outcomes in the buildup of desired products. For instance, a prior research demonstrated that the concurrent overexpression of PGM1 and UGP1 could elevate the UDP-glucose accumulation [18]. But the overexpression of these two genes did not enhance the production of salidroside in S. cerevisiae [19]. Another effective strategy was the utilization of the exogenous UDP-glucose supply pathway for enhancement [20]. For example, Pei et al. introduced the cellobiose phosphorolysis into E. coli and reconstructed a novel UDP-glucose synthesis pathway to increase UDP-glucose supply module and improved the bioconversion efficiency of luteolin to isoorientin [20]. Moreover, UDP, a product resulting from enzymatic glycosylation, acts as a strong inhibitor of UGT activity. Consequently, the rate of the glycosylation reaction diminishes gradually with the accumulation of UDP in the reaction mixture. Aiming at this problem, Thummler et al. found that Sucrose synthase (SUS), as a sucrose lyase, could promote the reversible conversion of sucrose and UDP to UDP-glucose and fructose [21]. Enzymatic approaches enabling the on-site replenishment of UDP-sugars have garnered significant interest due to their ability to lower the expense associated with UDP-sugar substrates and prevent UDP-induced product inhibitions [22,23]. To date, research efforts on salidroside have focused on UGT enzyme expression or optimization of precursor tyrosol production, while little effort has been devoted to the effective production and regeneration of UDP-glucose as a substrate for the glycosylation process. In contrast, efficient UDP-glucose supply has a very important impact on improving the yield of salidroside.
Traditional microbial biosynthesis depends on the control of a single species of microorganism, a process that poses difficulties, especially when dealing with lengthy and intricate foreign synthetic pathways that heavily strain the metabolic capacity of the host organism [24]. In contrast, through the strategy of dividing the complete synthesis pathway into two strains and co-cultivating them (referred to as the division of labor), the metabolic load on each strain can be significantly alleviated [25,26,27]. This approach provides a simple method for balancing different metabolic modules and has proven effective in the production of numerous natural compounds [28,29], including paclitaxel and tanshinone precursors [30], flavanoids [31,32], and resveratrol [33]. Due to their ease of genetic engineering and swift rate of growth, E. coli and S. cerevisiae have been widely utilized as standard prokaryotic and eukaryotic hosts for genetic engineering purposes [34]. The choice of host cells was a crucial prerequisite in the modification of microorganisms' metabolism for the synthesis of natural plant compounds [35]. Saccharomyces cerevisiae emerges as a eukaryotic organism notably more conducive than prokaryotes for the expression of natural product pathways derived from plants [36]. Attempts have also been undertaken to utilize cross-species co-cultures to harness the biosynthetic capabilities of the incorporated microbes for the purpose of biosynthesis. For instance, Yuan et al. utilized S. cerevisiae-E. coli co-cultures to achieve biosynthesis of resveratrol from glucose [37]. Liu et al. designed S. cerevisiae-E. coli co-cultures for biosynthesis of hydroxytyrosol [38]. Instead, scant scholarly investigations have been disseminated presently on the subject of salidroside production utilizing the co-culture system, and the co-culture of E. coli and S. cerevisiae for salidroside biosynthesis is almost blank.
Here, we described the co-expression of a SUS gene from Glycine max as GmSUS [39] and UGT genes from Rhodiola as RrUGT33 [15] in S. cerevisiae cells for enhancing the in vivo synthesis of small molecular tyrosol glucosides. The findings validated that co-expressing SUS enables the on-site regeneration of UDP-sugars, leading to enhanced salidroside production. In order to synthesize salidroside from cheap raw materials, a co-culture system of E. coli-S. cerevisiae brewing synthesized from de novo was established (Figure 1). The upstream strain E. coli was engineered for pathway intermediate L-tyrosine production, whereas the downstream strain S. cerevisiae converted L-tyrosine to salidroside. To accomplish this the downstream strain S. cerevisiae was engineered to utilize sucrose only and to enhance UDP-glucose availability for synthesis of salidroside, while the upstream strain E. coli was consumed to glucose and to produce precursor L-tyrosine. In summary, this work described an efficient biosynthesis of salidroside from inexpensive substrate.
2. Materials and Methods
2.1. Strains, Plasmids and Primers
The strains and plasmids used were all detailed in the supplementary information. E. Coli BL21 (DE3) was employed as the host to produce L-tyrosine; S. cerevisiae BY4743 was used as the host for salidroside production. For gene editing, E. coli DH5α (Novagen, USA) was utilized. PcDHPAAS (accession number MH710582.1), GmSUS (accession number NM001250596.2) and RrUGT33 (accession number MF674558.1) were codon optimized and chemically synthesized. The nucleic acid sequences of the synthesized genes PcDHPAAS, GmSUS and RrUGT33 were listed in Supplementary Table S3. GmSUS and RrUGT33 cloned into the pTEF426 expression vector to to yield GmSUS-pTEF426 and RrUGT33-pTEF426, respectively. The recombinant plasmid ptDH3-GmSUS-ptDH3-RrUGT33-pTEF426 carrying GmSUS and RrUGT33 was constructed using the pTDH3 promoter and named PDWZ0320 (Supplementary Table S1). The plasmid, AADW01, carrying a PcDHAAS was constructed and cloned into pET28a to form AADW01 (Supplementary Table S1). Then the plasmid RrUGT33-pTEF426 and PZW0320 was introduced into S. cerevisiae BY4743 to form the novel recombinant QH02, QH03, respectively (Supplementary Table S1). Strain QH01 was obtained by integrating PcDHAAS into the genome of S. cerevisiae BY4743. The plasmid AADW01 was introduced into E. coli BL21 (DE3) to form the novel recombinant DWL01. The design of all gene deletion (hxt1–hxt7 and gal2) strains was constructed using the method of Reifenberger et al.[40] and further confirmed by PCR.
2.2. Culture Media
Synthetic complete (SC) medium (6.7 g/L yeast nitrogen base without amino acids, and 2 g/L amino acid drop-out mix). In the coculture system, media containing 50 μg/mL of kanamycin was utilized as supplements. Medium1: 1/2 SC-Ura and 1/2 M9, sucrose: glucose=1:2; medium 2: 1/2 SC-Ura, and 1/2 M9, sucrose: glucose=1:1; medium 3: 1/2 SC-Ura and 1/2 M9 , sucrose: glucose=2:1; medium 4: 1/2 SC-Ura, and 1/2 M9, sucrose: glucose=3:1; medium 4: 1/2 SC-Ura, and 1/2 M9, sucrose: glucose=4:1. The synthetic fermented medium optimization contained 5 g/L yeast extraction, 10 g/L maltose, the composition of inorganic salts was referred to previous studies [36].
2.3. Cultivation Condition
To initiate the cultivation of yeast single-strain cultures, monoclones of the engineered BY4743 strain were inoculated into 5 mL of SC-ura medium. The inoculated cultures were then incubated overnight at 30°C with agitation at 200 rpm to facilitate the growth of seed cultures. Subsequently, 2% (v/v) of the seed cultures were transferred to 50 mL of fresh SC-ura medium and incubated under identical conditions for 96 hours. The medium was supplemented with 2 g/L L-tyrosine when it was needed to provide the substrate. The engineered E. coli BL21 (DE3) monoclonal clones were inoculated in 5 mL M9 medium containing 50 μg/mL kanamycin. The inoculation was carried out at 30 °C and 200 rpm overnight. A volume of one milliliter from overnight cultures was inoculated into 50 mL of M9 medium at 30 °C and agitated at 200 rpm for 4-6 hours. The method used to prepare engineered bacterial seeds in the E. coli - S. cerevisiae co-culture system was the same as in monoculture. To investigate the effect of media composition on the titer of salidroside, 2mL mixed seed cultures were collected, and QH03/QH04 strains were added at a ratio of 1:1 into 50 mL of five synthetic fermentation media (medium 1-5) supplemented with 50 μg/mL kanamycin and 100 μg/mL ampicillin. The mixture was stirred at 200 rpm for 4 hours at 30°C. In order to study the effects of the strain ratio on salidroside productivity, different ratios of QH03 strain to QH04 strain (1:2, 1:1, 2:1, 3:1) were examined using identical protocols. The fermentation medium chosen was medium 4, and the fermentation continued for 96 hours.
2.4. Analytical Methods
To conduct measurements, cells in 1.0 mL of culture were collected through centrifugation at 12000 rpm for 10 minutes, followed by filtration of the supernatants using a microporous membrane (0.22 μm) and subsequent transfer to sample vials for high-performance liquid chromatography (HPLC). The mobile phase conditions and elution procedures for HPLC were referred to previous studies [15]. In order to measure the quantity of salidroside, standard calibration curves were created by dissolving known concentrations of the standard compounds in the culture medium. R2 value for the standard curve was > 0.999. All experiments were conducted in triplicate and repeated a minimum of two times. The titers were presented as the average value with the standard deviation.
3. Results and Discussion
3.1. Construct a Biosynthetic Pathway from L-tyrosine to Salidroside in Saccharomyces cerevisiae
Salidroside is a natural phenolic product derived from the bioactive tyrosine. Previous studies have suggested different pathways involving separate decarboxylated and deamination steps from L-tyrosine to the key intermediate 4-hydroxyphenylacetaldehyde (4-HPAA), and Rhodiola contains phosphopyridoxal-dependent 4-HPAA synthase, which directly converts tyrosine to 4-HPAA. Therefore, here, we screened the gene encoding acetaldehyde synthase AAS to efficiently complete salidroside biosynthesis.
To investigate the application of PcDHPAAS enzymes in natural product engineering, we first compared the role of PcDHPAAS with the previously characterized Rr4HPAAS, PsTyDC and DmAMDr enzymes in tyrostol production. For this purpose, each AAS sequence was cloned into pTEF426 vector and transformed into wild-type S. cerevisiae BY4743. From this screening, we determined that of the various AAS enzymes, PcDHPAAS produces the highest levels of tyrosol 2.5g/L, as shown in Figure 2A. To our acknowledgment, the deduced catalytic reaction was shown in Figure 2B. PcDHPAAS was integrated into the yeast genome to obtain strain QH01. As previously observed with 4-hydroxyphenylacetaldehyde, endogenous yeast alcohol dehydrogenase (ADH) reduces 4-HPAA to tyrosol without the need for a phenylacetaldehyde reductase (PAR) transgene [14]. The outcomes demonstrated that PcDHAAS expression in strains QH01 triggered the tyrosol's biosynthesis (Supplementary Figure S1). Torrens-Spence et al. conducted a functional analysis of 34 UGT genes to unveil the entire the synthetic route of salidroside biosynthesis in Rhodiola [14]. Consequently, it was discovered that UGTs (RrUGT 17, 29, 32, and 33) exhibited site-specific T8GT activity, converting tyrosol into salidroside, where RrUGT33 demonstrated the most pronounced activity. Therefore, we introduced this glycosyltransferase into the synthetic pathway of salidroside. The gene was cloned into the vector pTEF426 and transferred into BY4743 to obtain strain QH02. As shown in Figure 2C, when RrUGT33 was expressed in strain QH02, salidroside was successfully produced with a titer of 1.42 g/L, and residues of bottom L-tyrosine and intermediate tyrosol could be detected in the fermentation solution (Figure 2D).
In this study, we first showed that PcDHPAAS rather than enzymes from other sources could catalyze the transformation of L-tyrosine to tyrosol, with higher efficiency. It was an efficient artificial metabolic pathway for obtaining the potential pharmaceutical chemical salidroside, based on our effort in screening for the “decoration” enzymes to realize the derivatization of the amino acid. The AAS enzyme itself has a valuable multifunctional feature, capable of simultaneous decarboxylation and deamination. The versatility of an enzyme simplifies the metabolic pathway, thereby reducing the metabolic burden of the host bacteria. However, the catalytic efficiency of AAS enzyme that has been studied and characterized are not high, which makes it not widely used. Therefore, it is important to screen for efficient AAS enzymes in synthetic biology.
3.2. Efficient Synthesis of Salidroside by In-Situ UDP-Glucose Regeneration
As can be seen from the previous research results, there was also an accumulation of the intermediate tyrosol in the fermentation solution, indicating that the production of salidroside was limited in the downstream glycosylation process, and its titer needed to be further improved. There were two reasons that may cause the low efficiency of glycosylation, one was insufficient supply of UDP-glucose; The second was the limitation of the catalytic efficiency of glycosyltransferase.
To verify the absence of UDP-glucose in the process, we added 3 mM UDP-glucose to the fermentation broth at 72 h. According to the liquid phase diagram (Figure 3A), the accumulation of the intermediate tyrosol in the fermentation solution decreased significantly, and the amount of salidroside increased. As time goes on, it could be seen that compared with the control group (no exogenous UDP-glucose was added), the production of salidroside in the experimental group (3 mM exogenous UDP-glucose was added) increased rapidly after 72 h, and the final titer was 2.02 g/L (Figure 3B), and the output increased by 42%. It could be seen that the lack of UDP-glucose in late fermentation led to the low titer of salidroside.
The addition of exogenous UDP-glucose has been shown to increase the production of salidroside, but the price of UDP-glucose was expensive, which made the cost high and did not meet the industrial scale production. Enzymatic approaches enabling the on-site replenishment of UDP-sugars have garnered significant interest due to their ability to lower the expense associated with UDP-sugar substrates and prevent UDP-induced product inhibition [22,23]. In this study, the UDP-glucose in situ circulation system was introduced into S. cerevisiae cells for the first time. We constructed the co-expression of a SUS gene from Glycine max as GmSUS [39] and UGT genes from Rhodiola as RrUGT33 [15] in S. cerevisiae cells for constructing a in situ UDP-glucose recycling system with inexpensive sucrose as the consumed substrate. GmSUS-pTEF426, RrUGT33-pTEF426 and PZW0320 were transformed into S. cerevisiae BY4743, and the successfully transformed strains were selected on Ura-deficient medium. Staining gels showed that the molecular sizes of GmSUS and RrUGT33 proteins were approximately 92 kD and 56 kD, respectively (Figure S2). The findings showed that GmSUS and RrUGT33 were effectively co-expressed in S. cerevisiae BY4743 cells. To verify the action of GmSUS, the recombinant QH02 and the recombinant QH03 were co-incubated with L-tyrosine and sucrose, respectively (Figure 3C). The results showed that strain QH03 containing GmSUS obtained higher salidroside titer.
In the present study, both QH02 and QH03 were able to form salidroside when incubated with L-tyrosine and glucose or L-tyrosine and sucrose, respectively (Figure 3C). A small amount of glucose could be detected in the fermentation broth of QH02 and QH03 with L-tyrosine and sucrose, respectively. This implied that sucrose did undergo hydrolysis to glucose and fructose before entering the cell [41]. Compared to glucose, the fermentation rate of sucrose was slightly behind, probably because of the hydrolysis process required to utilize the carbon source (Figure 3C). In addition, the salidroside production of strain QH02 gradually slowed down after 72 h of fermentation, while the salidroside production of strain QH03 (incubation with sucrose) continued to increase, suggesting that strain QH03 had more sugar donors at the later stage of fermentation. When sucrose was used as the sole carbon source, the final salidroside titer was 1.13 g/L for strain QH02 and 2.16 g/L for strain QH03. As expected, GmSUS were able to enhance the supply of UDP-glucose, thereby increasing salidroside production.
Finally, the highest content of salidroside was obtained when QH03 was incubated with L-tyrosine and sucrose. The molar conversion rate of L-tyrosine reached 70%, which was about 36% higher than that of QH02 (Figure 3D). However, the growth of strain QH03 seemed to be affected to a certain extent, and it did not grow as well in sucrose as glucose (Figure 3E).
The results indicated that GmSUS had the capability to increase UDP-glucose, which was a sugar donor essential for the enzymatic glycosylation reaction. This also reconfirmed the idea that sucrose could be ingested directly by S. cerevisiae [42,43,44,45,46], allowing in situ recycling of UDP-glucose to be achieved within the cell. Hence, it was crucial for the microbial synthesis of glycosides to optimize the production of UDP-sugars in the host organisms through a range of regulations and enhancements. The high accumulation of UDP-glucose (UDPG) in microbial hosts was not always guaranteed by the overexpression of endogenous pathway genes, as this overexpression frequently induces a negative cellular response, resulting in repression of gene expression or enzyme deactivation [47]. Glycosyltransferase converted UDPG and aglycones into glycosides and UDP. The activity of glycosyltransferase would be inhibited by UDP, thereby impacting the production of glycosides. The in situ recycling strategy of intracellular UDP used in this study could effectively increase the storage of UDPG in vivo without affecting the production of glycosides, which had great application prospects for the production of glycoside compounds.
3.3. Optimize the UDP-Glucose Regeneration System for Effective Salidroside Production
Because the growth of strain QH03 was affected to some extent, it did not grow as well in sucrose as in glucose. Therefore, considering that if the growth of strain QH03 can be improved, the titer of the target product salidroside can be further improved. We modified the QH03 strain based on previous studies. Batista et al. found that [48] a yeast strain deficient in key hexose transporters (hxt1–hxt7 and gal2) was incapable of growing on or fermenting glucose. The hxt-null strain was capable of fermenting sucrose better through the direct absorption of the sucrose into the cells. We constructed strain QH03-hxt-null by deleting hxt1–hxt7 and gal2 in strain QH03, and expected to have minimal uptake of glucose. As shown in Figure 4A, when the QH03-hxt-null strain was provided with glucose as the only carbon source, it exhibited slow consumption, and no additional consumption was noted after 24 hours. In other words, strain QH03-hxt-null grew very poorly on only glucose (Figure 4A). In comparison to glucose alone, strain QH03-hxt-null utilized sucrose more rapidly and grew better (Figure 4A). Finally, the content of salidroside obtained by incubation of QH03-hxt-null with L-tyrosine and sucrose was increased to 2.67 g/L (Figure 4B). The molar conversion rate of L-tyrosine was 81%.
In S. cerevisiae, direct sucrose intake was linked to maltose metabolism [48]. Indeed, higher sucrose fermentation rates have been observed when the S. cerevisiae-hxt-null strain was pre-grown on maltose. Therefore, maltose was used as an additive component of the medium to improve sucrose uptake by strain QH03-hxt-null. To further optimize the medium, the effects of maltose, inorganic salt mixture, and yeast extract on titer were explored using orthogonal studies. The 9th condition, which consisted of 10 g/L maltose, 5 g/L yeast extract, and 1.5 times the inorganic salt content, yielded the highest salidroside production at 3.14 g/L after 96 hours; the molar conversion rate of L-tyrosine was as high as 95% (Figure 4C). The significant impact of maltose and inorganic salt on salidroside production was clearly evident. As the quantity of yeast extract demonstrated an inverse relationship, the concentration of yeast extract was maintained at 5 g/L, maltose at 10 g/L, and inorganic salt at 1.5 times the original level.
It could be seen from the experimental results that the key to improving salidroside production in the UDP-glucose circulation system was the better absorption and utilization of sucrose by host strains. Whether through strain modification or medium optimization, the ultimate goal was to make the host bacteria have a strong metabolic ability for sucrose. For this problem, some scholars have done related research before. They found that continuous passage of host bacteria in sucrose environments can effectively promote the metabolic utilization of sucrose [45]. It also had an important reference value for increasing salidroside production in UDP-glucose circulation system.
3.4. Construction of a Co-Culture System Used for De Novo Synthesis of Salidroside
So far, although there has been a lot of research on the synthesis of salidroside, but the high cost is an important factor limiting the scale of industrialization. In order to reduce the cost of biosynthesis of salidroside, a co-culture strategy was adopted in this study to introduce tyrosine-producing QH04 into the fermentation system instead of adding the exogenous substrate L-tyrosine. This allowed the synthetic substrate of salidroside to be derived from inexpensive glucose and sucrose. In this system, the QH04 strain was used to produce L-tyrosine, while the QH03-hxt-null strain was responsible for converting L-tyrosine into salidroside (Figure 1). In this study, salidroside was successfully synthesized de novo by co-culturing strain QH04 with engineered yeast QH03-hxt-null and adding sucrose and glucose, respectively, at a final concentration of 20 g/L in the medium mixed 1:1 with M9 medium and SC-URA medium (Figure S3).
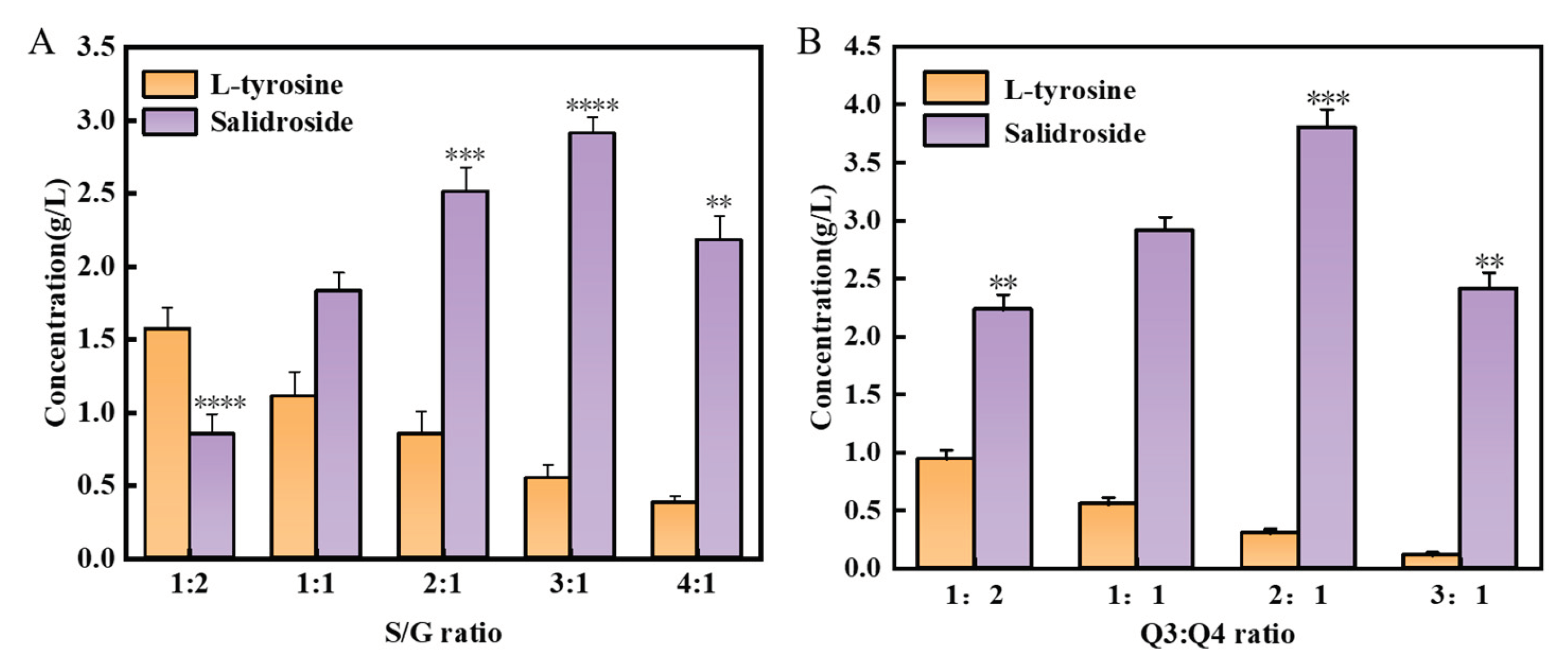
In order to increase the titer of de novo synthesis of salidroside, we optimized the co-culture conditions. Based on previous research, the performance of the microbial consortia was greatly impacted by the formulation medium's composition [36,49,50]. When engineering the QH03-hxt-null and QH04 strains in the symbiotic co-culture to utilize glucose and sucrose with varying efficiencies, our initial focus was on adjusting the metabolic capabilities of the two strains by manipulating the sucrose-to-glucose ratio (referred to as the S/G ratio). A sugar blend with varying sucrose and glucose proportions was examined using an inoculation ratio of 1:1 for the QH03-hxt-null and QH04 strains. When the S/G ratio was raised in the product profile (Figure 5A), there was a decrease in L-tyrosine accumulation, while salidroside production increased rapidly until the S/G ratio reached 3/1, resulting in the attainment of a high salidroside titer of 3.02 g/L. Increasing the S/G ratio to 4/1 not only reduced the accumulation of L-tyrosine but also hindered the production of salidroside. Next, we tried to adjust the initial inoculation ratio between the QH03-hxt-null and QH04 strains in the co-culture system. Various ratios of QH03-hxt-null and QH04 strains (referred to as the Q3/Q4 ratio, ranging from 1/2 to 3/1) were introduced into the coculture system at a S/G ratio of 3/1 (15 g/L sucrose and 5 g/L glucose) and cultivated together. As depicted in Figure 5B, when the initial Q3/Q4 ratio was 2/1 in the co-culture, the production of salidroside reached 3.8 g/L with minimal accumulation of L-tyrosine. Co-cultivation of the QH03-hxt-null strain with the QH04 strain reduced the cost by 81% compared with the QH02 strain (Table S4).
Consistently, a primary goal in microbial coculture engineering has been to improve the regulation of population dynamics and behavior. This was due to the highly dynamic nature of the interactions between strains and the considerable difficulty of maintaining balanced growth and metabolic function [51]. When two strains were co-cultivated on the same carbon source, competition for growth would lead to the incompatibility and instability of the consortium [52]. Implementing the strategy of engineering the co-culture strains to utilize distinct carbon sources, such as glucose or sucrose, aimed to decrease growth competition and enhance growth compatibility. The production of salidroside in the modified co-culture system was increased, which again proved the importance of strain compatibility in co-culture. These findings demonstrate that the synergistic interaction between QH03-hxt-null and QH04 strains was compatible, resulting in a robust coculture enabling high salidroside titers.
4. Conclusions
In this study, a synthetic pathway from L-tyrosine to salidroside was constructed by screening the highly efficient acetaldehyde synthetase AAS first. Next, the supply of UDP-glucose was enhanced by coupling the UDP-glucose regeneration system with UDP-glucosyltransferase from Rhodiola (RrUGT33) to establish an in situ UDP-glucose recycling system to using inexpensive sucrose as the substrate. After condition optimization, the molar conversion of L-tyrosine reached 95% at the shake flask level. Additionally, to reduce substrate costs, a co-culture system for salidroside production was developed by leveraging the interaction between E. coli and S. cerevisiae, with sucrose and glucose serving as carbon sources. Finally, 3.8 g/L of salidroside was obtained by fermentation in a cheap substrate. Notably, this study successfully implemented the recycling of in situ UDP-glucose in vivo to compensate for the inadequate supply of UDP-glucose. Moreover, it was the first time that sucrose was utilized as a carbon source for the production of the high-value product, salidroside. These findings suggested that in situ UDP-glucose enhancement holds significant potential for future salidroside production. Additionally, it provided valuable insights for the production of other glycosylated products.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: A. HPLC analysis of fermentation broth of QH04 strains. B. The LC–MS/MS of the fermentation supernatant sample of strain QH04. C. The LC–MS/MS of the standard tyrosol. Strain QH04 produced a major product that has identical retention time and yields identical ([M]-) ions to standard tyrosol and the primary ion fragment at m/z 137([M]-) corresponds to tyrosol with molecular weight of 138. Figure S2: Co-expression of RrUGT33 genes and GmSUS in S. cerevisiae. Analysis of GmSUS and the proteins obtained from crude extraction GmSUS and RrUGT33 co-expressing S. cerevisiae cells by coomassie-stained polyacrylamide gel. M: Marker, (1) GmSUS singly expressed; (2) RrUGT33 singly expressed; (3) RrUGT33 co-expressed with GmSUS. Figure S3: A. HPLC analysis of fermentation broth of QH03 and QH04 strains coculture. B. The LC–MS/MS of the standard salidroside. C. The LC–MS/MS of the fermentation supernatant sample of QH03 and QH04 strains coculture. Strain coculture produced a major product that has identical retention time and yields identical ([M]-) ions to standard salidroside and the primary ion fragment at m/z 299([M]-) corresponds to salidroside with molecular weight of 300; Table S1: Strains and plasmids used in this study; Table S2: Nucleotide sequences of codon optimized genes; Table S3: Nucleotide sequences of codon optimized genes; Table S4: Comparison of cost with different strains. Strain QH02 was fermented with 20 g/L glucose and 2 g/L L-tyrosine for 96 h to obtain the cost of 1 kg salidroside for comparison. The QH03-hxt-null strain was incubated with 20 g/L glucose and 2 g/L L-tyrosine for 96 h. The QH03-hxt-null and QH04 co-culture systems were fermented with 15 g/L sucrose and 5 g/L glucose as substrates for 96 h. Note: Cost was calculated at shaker level.
Author Contributions
DW conceived and supervised the project. XjZ and XxZ designed and performed themain experiments. Data wasevaluated by all authors. XjZ wrote the manuscript, D W, ZQ and RL et al. revised themanuscript. All authors have reviewed and endorsed the final manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (2022YFC2105700, 2021YFC2103300); the National Natural Science Foundation of China (22378032); Chongqing Outstanding Youth Fund (cstc2021jcyj-jqX0013), Youth project of science and technology research program of Chongqing Education Commission of China (KJQN201900112), the Human Resources and Social Security Bureau of Chongqing (cx2023036).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data produced or examined in this study are incorporated within this published article [and its Supplementary data].
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zha, J.; Wu, X.; Gong, G.; Koffas, M.A.G. Pathway enzyme engineering for flavonoid production in recombinant microbes. Metab Eng Commun 2019, 9, e00104. [Google Scholar] [CrossRef]
- Chiang, H.M.; Chen, H.C.; Wu, C.S.; Wu, P.Y.; Wen, K.C. Rhodiola plants: Chemistry and biological activity. J Food Drug Anal 2015, 23, 359–369. [Google Scholar] [CrossRef]
- Li, M.; Xu, T.; Zhou, F.; Wang, M.; Song, H.; Xiao, X.; Lu, B. Neuroprotective Effects of Four Phenylethanoid Glycosides on H2O2-Induced Apoptosis on PC12 Cells via the Nrf2/ARE Pathway. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Liu, X.; Wen, S.; Yan, F.; Liu, K.; Liu, L.; Wang, L.; Zhao, S.; Ji, X. Salidroside provides neuroprotection by modulating microglial polarization after cerebral ischemia. J Neuroinflammation 2018, 15, 39. [Google Scholar] [CrossRef] [PubMed]
- Marchev, A.S.; Dinkova-Kostova, A.T.; Gyorgy, Z.; Mirmazloum, I.; Aneva, I.Y.; Georgiev, M.I. Rhodiola rosea L.: from golden root to green cell factories. Phytochem Rev 2016, 15, 515–536. [Google Scholar] [CrossRef]
- Guo, W.; Huang, Q.L.; Feng, Y.H.; Tan, T.C.; Niu, S.H.; Hou, S.L.; Chen, Z.G.; Du, Z.Q.; Shen, Y.; Fang, X. Rewiring central carbon metabolism for tyrosol and salidroside production in " (Vol. 117, pg 2410, 2020). Biotechnol Bioeng 2022, 119, 2010–2010. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.J.; Yin, H.; Wang, S.A.; Zhuang, Y.B.; Liu, S.W.; Liu, T.; Ma, Y.H. Metabolic Engineering of Saccharomyces cerevisiae for High-Level Production of Salidroside from Glucose. J Agr Food Chem 2018, 66, 4431–4438. [Google Scholar] [CrossRef]
- Kallscheuer, N.; Menezes, R.; Foito, A.; da Silva, M.H.; Braga, A.; Dekker, W.; Sevillano, D.M.; Rosado-Ramos, R.; Jardim, C.; Oliveira, J.; et al. Identification and Microbial Production of the Raspberry Phenol Salidroside that Is Active against Huntington's Disease. Plant Physiol 2019, 179, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.X.; Chen, X.Z.; Yang, C.; Chang, J.Z.; Shen, W.; Fan, Y. Engineering for Enhanced Tyrosol Production. J Agr Food Chem 2017, 65, 4708–4714. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.; Kim, S.Y.; Ahn, J.-H. Production of three phenylethanoids, tyrosol, hydroxytyrosol, and salidroside, using plant genes expressing in Escherichia coli. Scientific Reports 2017, 7, 2578. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.B.; Jiang, J.L.; Liu, Z.N.; Qiao, B.; Li, F.F.; Cheng, J.S.; Sun, X.C.; Yuan, Y.J.; Qiao, J.J.; et al. Convergent engineering of syntrophic coculture for efficient production of glycosides. Metabolic Engineering 2018, 47, 243–253. [Google Scholar] [CrossRef]
- Yang, H.Q.; Xue, Y.X.; Yang, C.; Shen, W.; Fan, Y.; Chen, X.Z. Modular Engineering of Tyrosol Production in. J Agr Food Chem 2019, 67, 3900–3908. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.S.; Ma, L.Q.; Zhang, J.X.; Shi, G.L.; Hu, Y.H.; Wang, Y.N. Characterization of glycosyltransferases responsible for salidroside biosynthesis in. Phytochemistry 2011, 72, 862–870. [Google Scholar] [CrossRef]
- Torrens-Spence, M.P.; Pluskal, T.; Li, F.S.; Carballo, V.; Weng, J.K. Complete Pathway Elucidation and Heterologous Reconstitution of Biosynthesis. Mol Plant 2018, 11, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tian, Y.; Zhou, Y.; Kan, Y.; Wu, T.; Xiao, W.; Luo, Y. Multi-modular engineering of Saccharomyces cerevisiae for high-titre production of tyrosol and salidroside. Microbial Biotechnology 2021, 14, 2605–2616. [Google Scholar] [CrossRef]
- Thuan, N.H.; Chaudhary, A.K.; Van Cuong, D.; Cuong, N.X. Engineering co-culture system for production of apigetrin in Escherichia coli. J Ind Microbiol Biotechnol 2018, 45, 175–185. [Google Scholar] [CrossRef]
- Zhang, X.; Xie, J.; Cao, S.; Zhang, H.; Pei, J.; Bu, S.; Zhao, L. Efficient production of the glycosylated derivatives of baicalein in engineered Escherichia coli. Appl Microbiol Biotechnol 2023, 107, 2831–2842. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Yang, G.Y.; Chen, X.H.; Liu, Q.; Zhang, X.L.; Deng, Z.X.; Feng, Y. Biosynthesis of plant-derived ginsenoside Rh2 in yeast via repurposing a key promiscuous microbial enzyme. Metabolic Engineering 2017, 42, 25–32. [Google Scholar] [CrossRef]
- Liu, H.; Tian, Y.; Zhou, Y.; Kan, Y.; Wu, T.; Xiao, W.; Luo, Y. Multi-modular engineering of Saccharomyces cerevisiae for high-titre production of tyrosol and salidroside. Microb Biotechnol 2021, 14, 2605–2616. [Google Scholar] [CrossRef]
- Pei, J.; Sun, Q.; Zhao, L.; Shi, H.; Tang, F.; Cao, F. Efficient Biotransformation of Luteolin to Isoorientin through Adjusting Induction Strategy, Controlling Acetic Acid, and Increasing UDP-Glucose Supply in Escherichia coli. J Agric Food Chem 2019, 67, 331–340. [Google Scholar] [CrossRef]
- Thummler, F.; Verma, D.P.S. Nodulin-100 of Soybean Is the Subunit of Sucrose Synthase Regulated by the Availability of Free Heme in Nodules. Journal of Biological Chemistry 1987, 262, 14730–14736. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Fang, J.W.; Zhang, J.B.; Liu, Z.Y.; Shao, J.; Kowal, P.; Andreana, P.; Wang, P.G. Sugar nucleotide regeneration beads (superbeads): A versatile tool for the practical synthesis of oligosaccharides. Journal of the American Chemical Society 2001, 123, 2081–2082. [Google Scholar] [CrossRef] [PubMed]
- Masada, S.; Kawase, Y.; Nagatoshi, M.; Oguchi, Y.; Terasaka, K.; Mizukami, H. An efficient chemoenzymatic production of small molecule glucosides with in situ UDP-glucose recycling. Febs Lett 2007, 581, 2562–2566. [Google Scholar] [CrossRef] [PubMed]
- Roell, G.W.; Zha, J.; Carr, R.R.; Koffas, M.A.; Fong, S.S.; Tang, Y.J.J. Engineering microbial consortia by division of labor. Microb Cell Fact 2019, 18. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Wang, X.N.; Zhang, H.R. Balancing the non-linear rosmarinic acid biosynthetic pathway by modular co-culture engineering. Metabolic Engineering 2019, 54, 1–11. [Google Scholar] [CrossRef]
- Johnston, T.G.; Yuan, S.F.; Wagner, J.M.; Yi, X.; Saha, A.; Smith, P.; Nelson, A.; Alper, H.S. Compartmentalized microbes and co-cultures in hydrogels for on-demand bioproduction and preservation. Nat Commun 2020, 11, 563. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X. Modular co-culture engineering, a new approach for metabolic engineering. Metab Eng 2016, 37, 114–121. [Google Scholar] [CrossRef]
- Jawed, K. Advances in the development and application of microbial consortia for metabolic engineering (vol 9, e00095, 2019). Metabolic Engineering Communications 2020, 11. [Google Scholar]
- Akdemir, H.; Silva, A.; Zha, J.; Zagorevski, D.V.; Koffas, M.A.G. Production of pyranoanthocyanins using Escherichia coli co-cultures. Metabolic Engineering 2019, 55, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Qiao, K.J.; Edgar, S.; Stephanopoulos, G. Distributing a metabolic pathway among a microbial consortium enhances production of natural products. Nature Biotechnology 2015, 33, 377-U157. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Vernacchio Victoria, R.; Collins Shannon, M.; Shirke Abhijit, N.; Xiu, Y.; Englaender Jacob, A.; Cress Brady, F.; McCutcheon Catherine, C.; Linhardt Robert, J.; Gross Richard, A.; et al. Complete Biosynthesis of Anthocyanins Using E. coli Polycultures. mBio 2017, 8. [Google Scholar] [CrossRef]
- Wang, X.; Li, Z.; Policarpio, L.; Koffas, M.A.G.; Zhang, H. De novo biosynthesis of complex natural product sakuranetin using modular co-culture engineering. Appl Microbiol Biotechnol 2020, 104, 4849–4861. [Google Scholar] [CrossRef]
- Dwibedi, V.; Saxena, S. Effect of precursor feeding, dietary supplementation, chemical elicitors and co-culturing on resveratrol production by Arcopilus aureus. Prep Biochem Biotechnol 2022, 52, 404–412. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Nielsen, J. Production of natural products through metabolic engineering of Saccharomyces cerevisiae. Curr Opin Biotechnol 2015, 35, 7–15. [Google Scholar] [CrossRef]
- O'Connor, S.E. Engineering of Secondary Metabolism. Annu Rev Genet 2015, 49, 71–94. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.; Li, X.; Liu, D.; Dong, X.T.; Li, F.F.; Wang, E.X.; Wang, E.X.; Li, B.Z.; Yuan, Y.J. Production of naringenin from D-xylose with co-culture of E-coli and S-cerevisiae. Engineering in Life Sciences 2017, 17, 1021–1029. [Google Scholar] [CrossRef]
- Yuan, S.F.; Yi, X.; Johnston, T.G.; Alper, H.S. De novo resveratrol production through modular engineering of an Escherichia coli-Saccharomyces cerevisiae co-culture. Microb Cell Fact 2020, 19, 143. [Google Scholar] [CrossRef]
- Liu, Y.; Song, D.; Hu, H.; Yang, R.; Lyu, X. De Novo Production of Hydroxytyrosol by Saccharomyces cerevisiae-Escherichia coli Coculture Engineering. ACS Synth Biol 2022, 11, 3067–3077. [Google Scholar] [CrossRef]
- Huang, F.C.; Hinkelmann, J.; Hermenau, A.; Schwab, W. Enhanced production of β-glucosides by in-situ UDP-glucose regeneration. Journal of Biotechnology 2016, 224, 35–44. [Google Scholar] [CrossRef]
- Reifenberger, E.; Freidel, K.; Ciriacy, M. Identification of Novel Hxt Genes in Saccharomyces-Cerevisiae Reveals the Impact of Individual Hexose Transporters on Glycolytic Flux. Mol Microbiol 1995, 16, 157–167. [Google Scholar] [CrossRef]
- Lagunas, R. Sugar-Transport in Saccharomyces-Cerevisiae. Fems Microbiol Lett 1993, 104, (3–4). [Google Scholar] [CrossRef]
- Orlowski, J.H.; Barford, J.P. Direct Uptake of Sucrose by Saccharomyces-Cerevisiae in Batch and Continuous Culture. J Gen Appl Microbiol 1991, 37, 215–218. [Google Scholar] [CrossRef]
- Barford, J.P.; Phillips, P.J.; Orlowski, J.H. A New Model of Uptake of Multiple Sugars by Saccharomyces-Cerevisiae.1. Bioprocess Eng 1992, 7, 297–302. [Google Scholar] [CrossRef]
- Friis, J.; Ottolenghi, P. Localization of Invertase in a Strain of Yeast. Cr Trav Lab Carlsb 1958, 31, 259–271. [Google Scholar]
- Barford, J.P.; Mwesigye, P.K.; Phillips, P.J.; Jayasuriya, D.; Blom, I. Further Evidence of Direct Uptake of Sucrose by Saccharomyces-Cerevisiae in Batch Culture. J Gen Appl Microbiol 1993, 39, 389–394. [Google Scholar] [CrossRef]
- Rodrigues, C.I.S.; Wahl, A.; Gombert, A.K. Aerobic growth physiology of Saccharomyces cerevisiae on sucrose is strain-dependent. FEMS Yeast Research 2021, 21, foab021. [Google Scholar] [CrossRef]
- Zha, J.; Koffas, M.A.G. Production of anthocyanins in metabolically engineered microorganisms: Current status and perspectives. Syn Syst Biotechno 2017, 2, 259–266. [Google Scholar] [CrossRef]
- Batista, A.S.; Miletti, L.C.; Stambuk, B.U. Sucrose Fermentation by Saccharomyces cerevisiae Lacking Hexose Transport. Journal of Molecular Microbiology and Biotechnology 2005, 8, 26–33. [Google Scholar] [CrossRef]
- Yuan, S.-F.; Yi, X.; Johnston, T.G.; Alper, H.S. De novo resveratrol production through modular engineering of an Escherichia coli–Saccharomyces cerevisiae co-culture. Microbial Cell Factories 2020, 19, 143. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, X.; Li, Y.; Yan, Y.; Yuan, Q. Metabolic engineering of Escherichia coli for microbial synthesis of monolignols. Metabolic Engineering 2017, 39, 102–109. [Google Scholar] [CrossRef]
- Lindemann, S.R.; Bernstein, H.C.; Song, H.S.; Fredrickson, J.K.; Fields, M.W.; Shou, W.Y.; Johnson, D.R.; Beliaev, A.S. Engineering microbial consortia for controllable outputs. Isme J 2016, 10, 2077–2084. [Google Scholar] [CrossRef]
- Song, H.; Ding, M.Z.; Jia, X.Q.; Ma, Q.; Yuan, Y.J. Synthetic microbial consortia: from systematic analysis to construction and applications. Chemical Society Reviews 2014, 43, 6954–6981. [Google Scholar] [CrossRef]
Figure 1.
Diagram showcasing the synthetic coculture of S. cerevisiae and E. coli designed for the production of salidroside using a blend of glucose and sucrose. E. coli QH04 synthesizes L-tyrosine from glucose. Sucrose synthase GmSUS from Glycine max and glycosyltransferase RrUGT33 from Rhodiola were introduced into the endogenous UDP-glucose-producing S. cerevisiae to form strain QH03-hxt-null, which could achieve in situ UDP-glucose circulation and salidroside synthesis. QH03-hxt-null and QH04 strains ingested sucrose and glucose, respectively, and cocultured stably. G6P, glucose 6-phosphate; F6P, fructose 6-phosphate; 6PG, 6-phosphogluconic acid; PEP, phosphoenolpyruvate; E4P, Erythrose-4-phosphate; DAHP, 3-deoxy-arabino-heptulonate 7-phosphate; CHA, chorismate; L-TYR, l-tyrosine; ×, gene deletion; PcDHPAAS, acetaldehyde synthases enzymes; 4-HPAA, 4-Hydroxyphenylacetaldehyde, ADH, alcohol dehydrogenase, UDP, uridine diphosphate; UDP-glucose, uridine 5, 9-diphosphoglucose; RrUGT33, UDP-glucosyltransferase from Rhodiola; GmSUS, sucrose synthase from Glycine max; Agt1, Alpha-Glucoside Transporter.
Figure 1.
Diagram showcasing the synthetic coculture of S. cerevisiae and E. coli designed for the production of salidroside using a blend of glucose and sucrose. E. coli QH04 synthesizes L-tyrosine from glucose. Sucrose synthase GmSUS from Glycine max and glycosyltransferase RrUGT33 from Rhodiola were introduced into the endogenous UDP-glucose-producing S. cerevisiae to form strain QH03-hxt-null, which could achieve in situ UDP-glucose circulation and salidroside synthesis. QH03-hxt-null and QH04 strains ingested sucrose and glucose, respectively, and cocultured stably. G6P, glucose 6-phosphate; F6P, fructose 6-phosphate; 6PG, 6-phosphogluconic acid; PEP, phosphoenolpyruvate; E4P, Erythrose-4-phosphate; DAHP, 3-deoxy-arabino-heptulonate 7-phosphate; CHA, chorismate; L-TYR, l-tyrosine; ×, gene deletion; PcDHPAAS, acetaldehyde synthases enzymes; 4-HPAA, 4-Hydroxyphenylacetaldehyde, ADH, alcohol dehydrogenase, UDP, uridine diphosphate; UDP-glucose, uridine 5, 9-diphosphoglucose; RrUGT33, UDP-glucosyltransferase from Rhodiola; GmSUS, sucrose synthase from Glycine max; Agt1, Alpha-Glucoside Transporter.
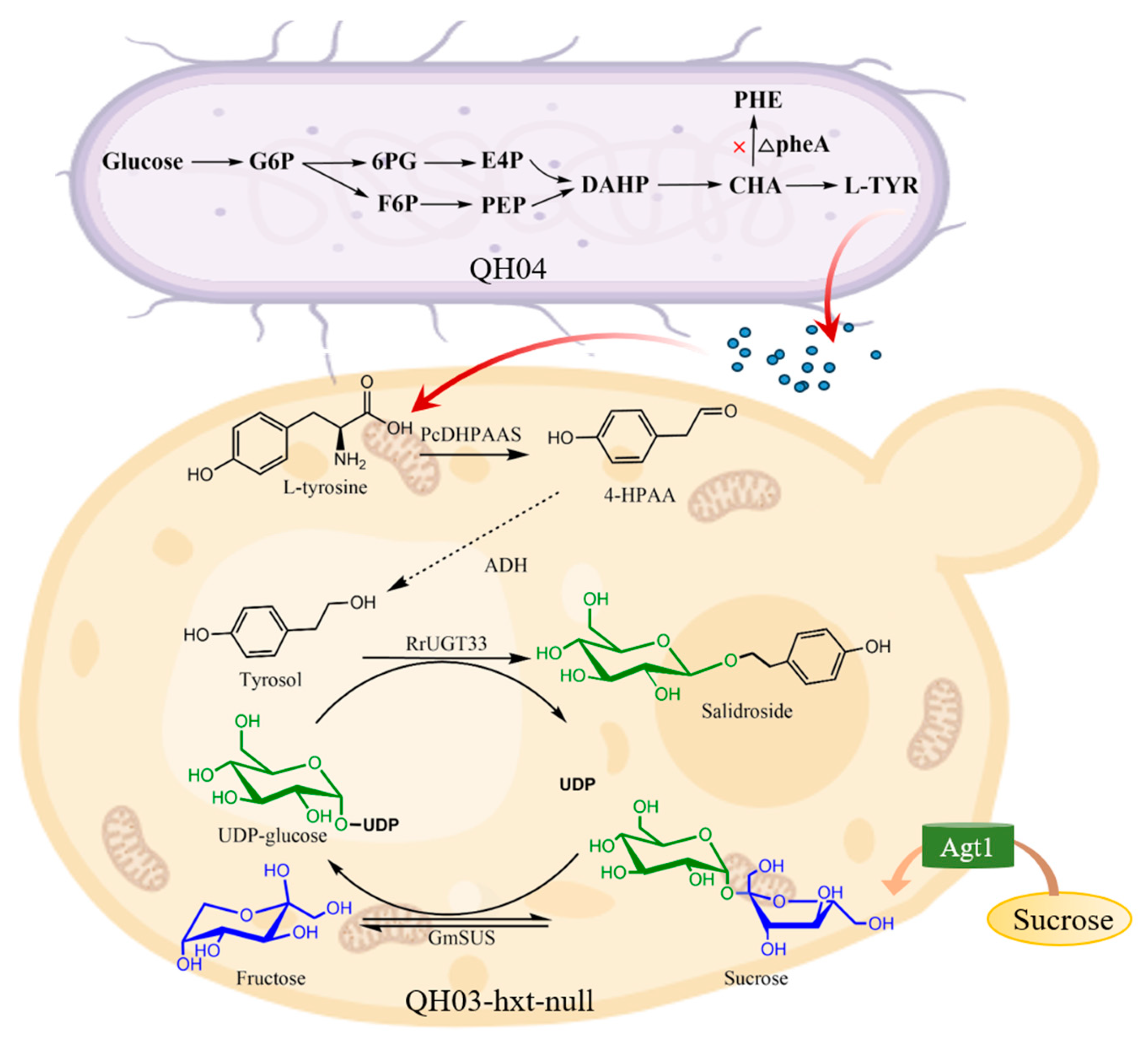
Figure 2.
Construction of biosynthetic pathway from L-tyrosine to salidroside. A. Compared the catalytic efficiency of the four enzymes PcDHPAAS, Rr4HPAAS, PsTyDC and DmAMDr enzymes in the production of 4-HPAA. B. Reaction process from L-tyrosine to 4-HPAA. C. The HPLC of fermentation broth. Recombinant QH02 (expressing PcDHPAAS, RrUGT33 in S. cerevisiae) were incubated with L-tyrosine and glucose. D. Concentration changes of substrate L-tyrosine, intermediate tyrosol and target salidroside in fermentation solution with time.
Figure 2.
Construction of biosynthetic pathway from L-tyrosine to salidroside. A. Compared the catalytic efficiency of the four enzymes PcDHPAAS, Rr4HPAAS, PsTyDC and DmAMDr enzymes in the production of 4-HPAA. B. Reaction process from L-tyrosine to 4-HPAA. C. The HPLC of fermentation broth. Recombinant QH02 (expressing PcDHPAAS, RrUGT33 in S. cerevisiae) were incubated with L-tyrosine and glucose. D. Concentration changes of substrate L-tyrosine, intermediate tyrosol and target salidroside in fermentation solution with time.
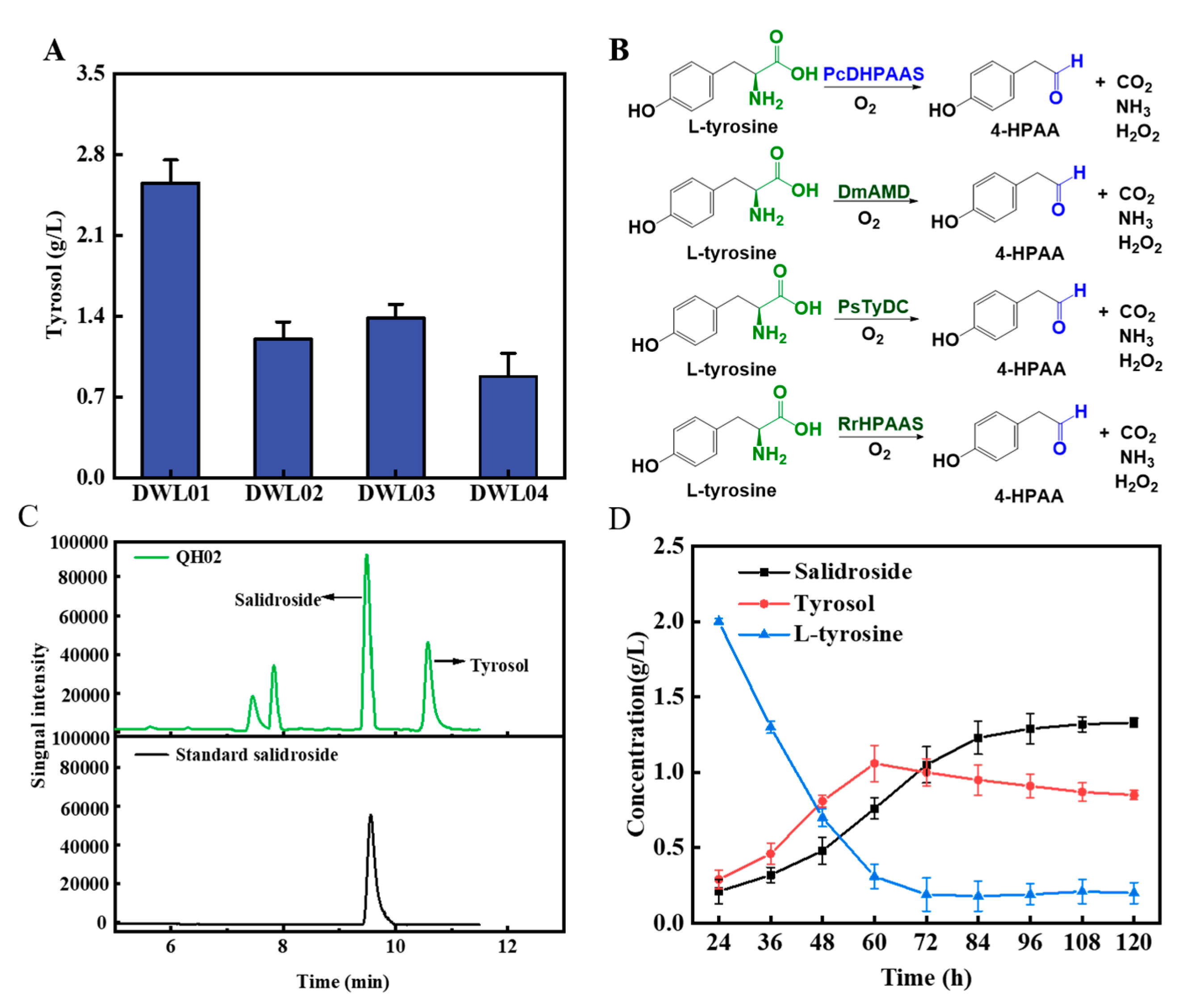
Figure 3.
Strain QH02 used L-tyrosine as substrate to produce salidroside. A. The HPLC of fermentation broth. Recombinant QH02 (expressing PcDHPAAS, RrUGT33 in S. cerevisiae) were incubated with 11mM L-tyrosine and 0 mM or 3 mM UDP-glucose, respectively. B. Plot of the titer variation of strains QH02 produce salidroside by fermentation with UDP-glucose. C. Plot of the titer variation of salidroside produced by QH02 and QH03 with different carbon sources. D. The final titer of salidroside and molar conversion of L-tyrosine produced by QH02 and QH03 with different carbon sources. E. OD600nm of QH03 under different carbon sources. The trials were conducted three times, with the error bars indicating the standard deviations. “*” p < 0.05 vs. compared group, “***” p < 0.001 vs. compared group, “ns” stands for “not significant”.
Figure 3.
Strain QH02 used L-tyrosine as substrate to produce salidroside. A. The HPLC of fermentation broth. Recombinant QH02 (expressing PcDHPAAS, RrUGT33 in S. cerevisiae) were incubated with 11mM L-tyrosine and 0 mM or 3 mM UDP-glucose, respectively. B. Plot of the titer variation of strains QH02 produce salidroside by fermentation with UDP-glucose. C. Plot of the titer variation of salidroside produced by QH02 and QH03 with different carbon sources. D. The final titer of salidroside and molar conversion of L-tyrosine produced by QH02 and QH03 with different carbon sources. E. OD600nm of QH03 under different carbon sources. The trials were conducted three times, with the error bars indicating the standard deviations. “*” p < 0.05 vs. compared group, “***” p < 0.001 vs. compared group, “ns” stands for “not significant”.
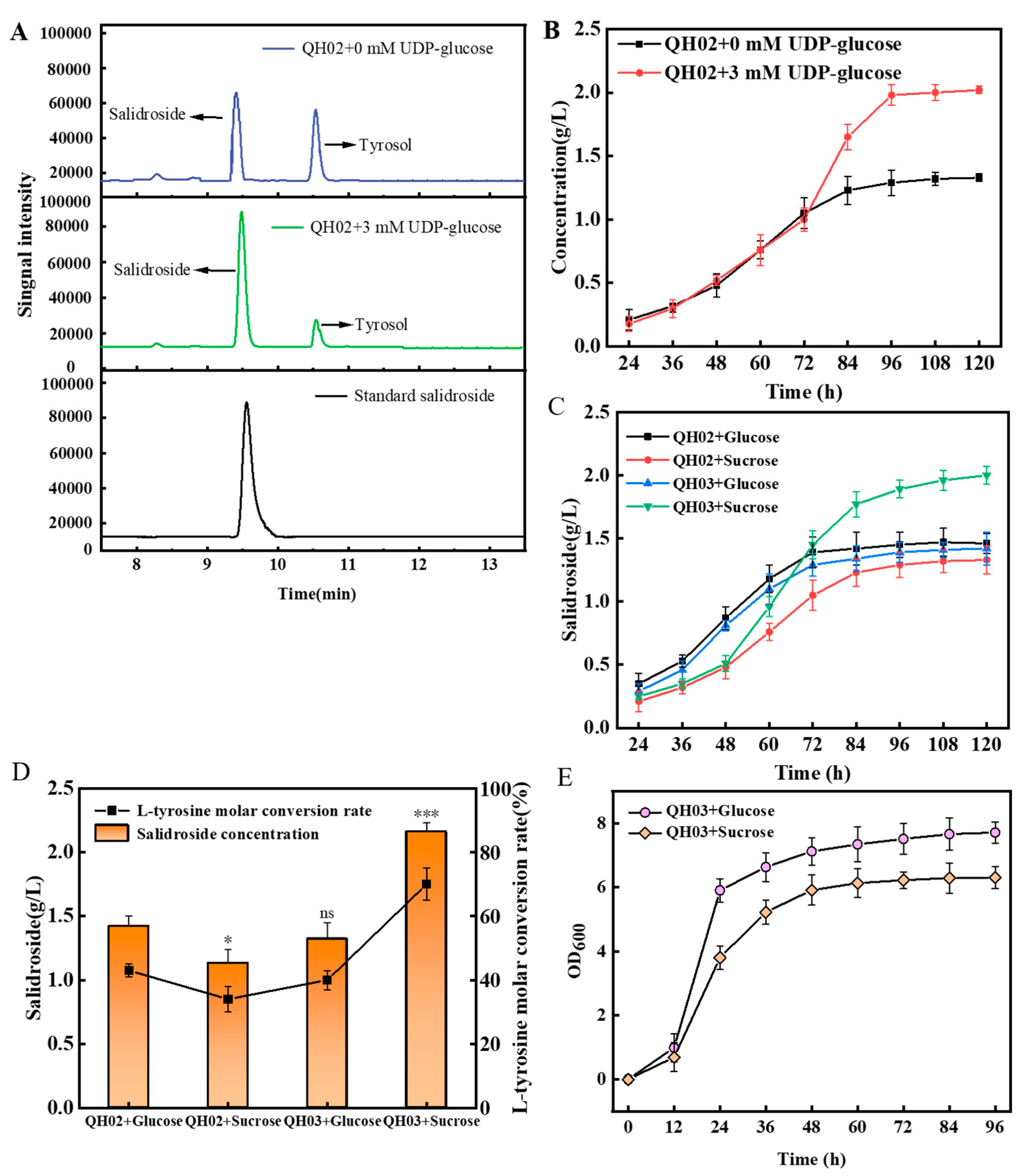
Figure 4.
Optimization of the UDP-glucose circulation System. A. Comparison of sugar consumption and growth of strain QH03-hxt-null in single glucose and sucrose, respectively. B. The concentration change of salidroside produced by strain QH03 and strain QH03-hxt-null when sucrose was carbon source. C. Optimization of salidroside production by altering the content of maltose, inorganic salt mixture, and yeast extract. The trials were conducted three times, with the error bars indicating the standard deviations.
Figure 4.
Optimization of the UDP-glucose circulation System. A. Comparison of sugar consumption and growth of strain QH03-hxt-null in single glucose and sucrose, respectively. B. The concentration change of salidroside produced by strain QH03 and strain QH03-hxt-null when sucrose was carbon source. C. Optimization of salidroside production by altering the content of maltose, inorganic salt mixture, and yeast extract. The trials were conducted three times, with the error bars indicating the standard deviations.
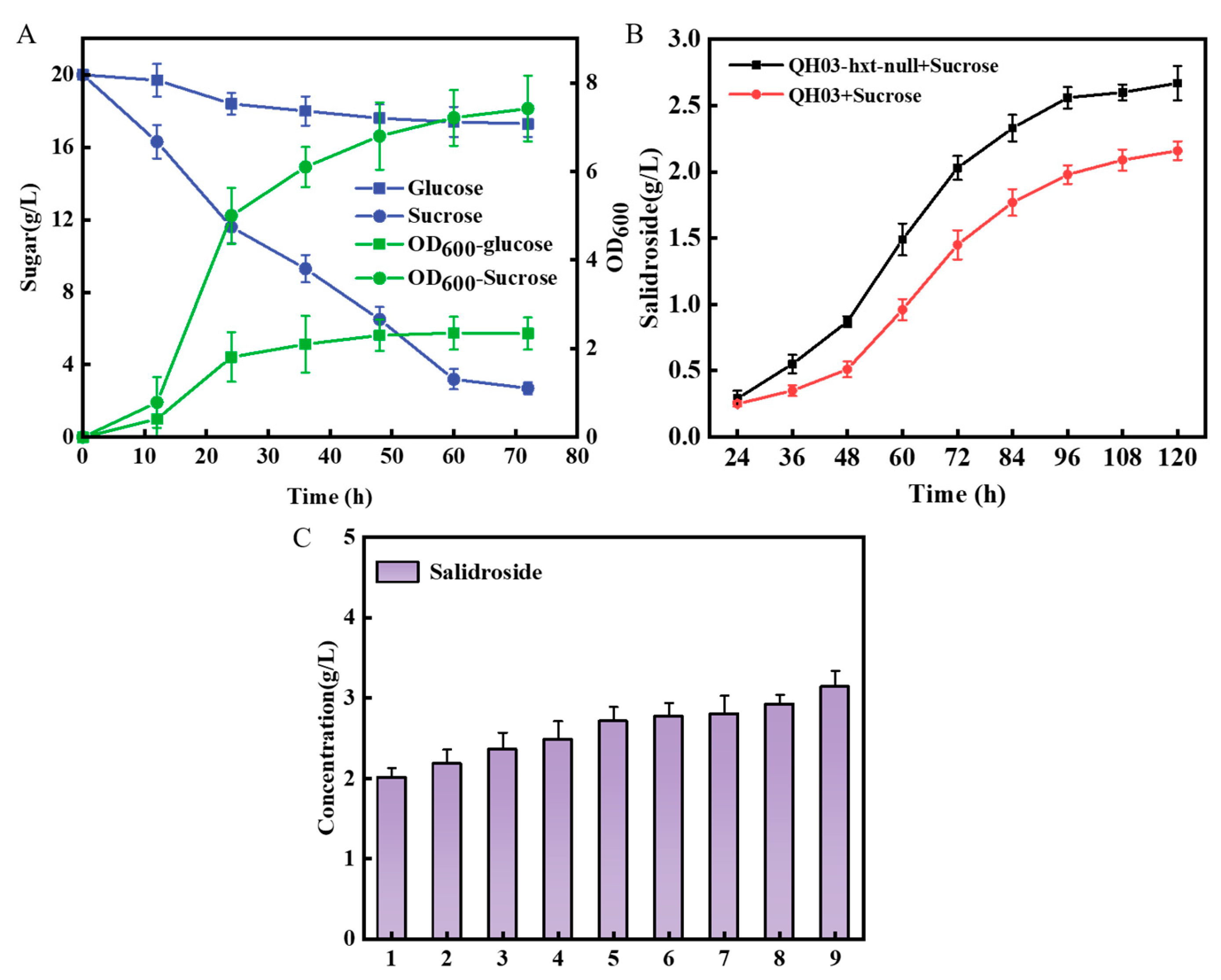
Figure 5.
Salidroside production through metabolic balancing of QH03-hxt-null and QH04 strains in a synergistic coculture. A. Enhancing salidroside production by adjusting the sucrose-to-glucose ratio, starting with an initial S/G ratio of 1/1. B. Improving salidroside production by adjusting the inoculation ratio of the QH03-hxt-null strain and QH04 strain, using an S/G ratio of 3/1. The trials were conducted three times, with the error bars indicating the standard deviations. “*” p < 0.05 vs. compared group, “**” p < 0.01 vs. compared group, “***” p < 0.001 vs. compared group, and “****” p < 0.0001 vs. compared group.
Figure 5.
Salidroside production through metabolic balancing of QH03-hxt-null and QH04 strains in a synergistic coculture. A. Enhancing salidroside production by adjusting the sucrose-to-glucose ratio, starting with an initial S/G ratio of 1/1. B. Improving salidroside production by adjusting the inoculation ratio of the QH03-hxt-null strain and QH04 strain, using an S/G ratio of 3/1. The trials were conducted three times, with the error bars indicating the standard deviations. “*” p < 0.05 vs. compared group, “**” p < 0.01 vs. compared group, “***” p < 0.001 vs. compared group, and “****” p < 0.0001 vs. compared group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



