
Submitted:
07 March 2024
Posted:
08 March 2024
You are already at the latest version
Abstract
Preventing the rebound of black and odorous water bodies is critical for improving the ecological environment of water bodies. This study investigated the effect and mechanism of in-situ improvement of sediment microenvironment by nitrate in tailwater of wastewater treatment plants combined with aerobic denitrifying bacteria under low-DO regulation (TailN+CFM+LDO). On the 60th day of remediation, the dissolved oxygen and oxidation-reduction potential in the overlying water increased to 5.6 mg/L and 300 mV, respectively, the acid volatile sulfide in the sediment decreased by 70.4%, and the organic matter in the sediment was reduced by 62.7%, in which the HFOM was degraded from 105 g/kg to 56 g/kg, and the potential risk of water reverting to black and odorous conditions significantly decreased. Amplicon sequencing analysis revealed that the relative abundance of electroactive bacteria Thiobacillus and Pseudomonas with denitrification capacity was found to be significantly higher in the TailN+CFM+LDO group than in the other remediation groups. Functional prediction of 16S sequencing results indicated that both the quantity and activity of critical microbial enzymes involved in nitrification and denitrification processes could enhance in the TailN+CFM+LDO group. These results improved our understanding of the improvement of the sediment microenvironment and could thus facilitate its application.
Keywords:
Black and odorous water bodies
; Microenvironment
; Sulfur autotrophic denitrification
; Electroactive Bacteria
; Functional gene
1. Introduction
Tailwater of wastewater treatment plants, as a potential source of pollution in urban receiving water bodies, has a significant impact on urban river ecosystems [1]. Understanding the impact of wastewater treatment plant tailwater on receiving water bodies and the associated mechanisms is crucial for promoting the comprehensive utilization of sewage resources. Sediments in water bodies have a high organic matter content and insufficient hydrodynamics, rendering them susceptible to the proliferation of anaerobic microorganisms such as sulfate-reducing bacteria, anaerobic digestion bacteria, and iron-reducing bacteria. These microorganisms produce substances such as FeS, NH3-N, and acid volatile sulfide (AVS) that cause black and odorous conditions in water bodies [2]. Therefore, reducing organic matter in sediments and improving the sediment microecological environment have emerged as primary strategies to prevent the recurrence of black and odorous water.
The gradual reduction of organic matter in sediments and the production of reducing substances are mainly due to inadequate microbial activity and a lack of electron acceptors in sediments [3,4]. Previous studies have indicated that effective remediation outcomes can be achieved through the utilization of either Ca(NO3)2 or domesticated composite microorganisms in sediments. Ca(NO3)2 can provide electron acceptors for microorganisms, while composite microorganisms can enhance microbial activity and establish stable ecosystems, effectively degrading nitrogen, phosphorus, sulfides, etc [5,6]. Ca(NO3)2, as an economical and efficient electron acceptor, has been extensively used in the remediation of black and odorous water bodies, effectively reducing AVS in water bodies and improving the redox environment [7,8].
Due to the long-term anaerobic or hypoxic conditions in sediments, excess NO3--N under microbial influence can undergo dissimilatory nitrate reduction to ammonium (DNRA). The release of NH3-N may pose potential environmental hazards [9]. Previous studies have indicated that low-intensity or intermittent aeration can can increase DO and ORP and effectively inhibit the DNRA process [10,11]. Aerobic denitrifying bacteria as microorganisms prevalent in water bodies prefer aerobic environments (DO of 0.2 ~ 5.9 mg/L), however, there is limited research reporting the application of tailwater NO3--N + aerobic denitrifying bacteria + low-DO regulation group to prevent the rebound of black and odorous water bodies. Additionally, the influencing mechanism of wastewater treatment plant tailwater on the microbial ecological environment of water bodies has not been reported.
This study aims to investigate the inhibitory effect of using NO3--N in wastewater treatment plant tailwater combined with aerobic denitrifying bacteria under low-DO conditions on the rebound of black and odorous water bodies. We will discuss the impact of various remediation conditions on DO, ORP, NH3-N, NO3--N, NO2--N in the overlying water and AVS in the sediment. Additionally, we will investigate changes in sediment microbial community structure using high-throughput sequencing technology. We aim to explore the changes in the microbial ecological environment of water bodies and the mechanisms that inhibit the rebound of black and odorous water bodies. Through this exploration, we aim to offer practical insights for the remediation of urban water bodies contaminated with black and odorous water.
2. Materials and Methods
2.1. Sediments and Overlying Water Sources
Sediment was collected using a column sampler from the 0~30 cm depth of a river. After collection, any plastic, stone, or glass debris present in the sediment was promptly removed. The sediment was then transferred to plastic buckets, sealed with lids to prevent air interference, and immediately transported to the laboratory. It was stored in the dark at 4°C. Subsequently, overlying water samples were collected from a section of the river that was unaffected by sediment disturbance. The water characteristics of the overlying water were as follows: ORP 100.15 mV, pH 7.45, DO 2.10 mg/L, chemical oxygen demand (COD) 43.00 mg/L, NH3-N 4.20 mg/L, and total nitrogen (TN) 4.50 mg/L. The wastewater treatment plant tailwater exhibited the following characteristics: ORP 200.34 mV, pH 7.12, DO 2.10 mg/L, COD 47.00 mg/L, NH3-N 4.30 mg/L and TN 9.50 mg/L. The primary indicators of the sediment included an organic matter content of 150 g/kg, Heavy fraction organic matters (HFOM) content of 105 g/kg, Light fraction organic matters (LFOM) content of 45 g/kg and an AVS content of 3,010 mg/kg.
2.2. Experimental Design
The experimental setup consisted of a custom-made water tank with dimensions of 40 cm (length) × 40 cm (width) × 100 cm (height) (Figure S1). Approximately 3.2 L of sediment (20 cm in height) was filled into the tank, and about 12.8 L of overlying water (80 cm in height) were gradually added to the tank using the siphoning method. The experimental groups were as follows: blank group (CN), tailwater NO3--N group (TailN), tailwater NO3--N + Aerobic denitrifying bacteria group (TailN+CFM), and tailwater NO3--N + Aerobic denitrifying bacteria + low-DO regulation group (TailN+CFM+LDO). The blank group was supplied with overlying water from the river, whereas the remediation groups were supplied with tailwater from the specified wastewater treatment plant. The water supply had a flow rate of 0.005 m3/s. Aerobic denitrifying bacteria were Citrobacter sp. isolated and screened from sediments. The Aerobic denitrifying bacteria dosage was established at 1 g/month based on preliminary experiment optimization results. In the TailN+CFM+LDO group, intermittent aeration (0.1 m3/h, 3 h/d) was employed. Water samples from the overlying water were collected every 5 days, whereas sediment samples were collected every 15 days, based on the respective indicators.
2.3. Analytical Methods
The DO and ORP in both the overlying water and interstitial water were measured using a multi-parameter water quality analyzer (Multi360 IDS, WTW, China). Prior to analysis, overlying water samples for TN, NH3-N, NO3--N, and NO2--N were centrifuged at 8,000 rpm for 10 minutes, and the resulting supernatant was filtered through a 0.45 μm membrane filter. COD of the overlying water was determined using the potassium dichromate oxidation method. Additionally, the sulfate (SO42-) content in the overlying water was measured using ion chromatography (Dionex 1100, USA). The organic matter in the sediment was determined using the potassium dichromate oxidation method, whereas the total nitrogen in the sediment was measured using the alkaline potassium persulfate method. HFOM and LFOM are separated by the density method, and it is generally accepted that organic matter with a density ρ<1.7 g/cm3 belongs to LFOM, while organic matter with a density ρ>1.7 g/cm3 belongs to HFOM. The AVS in the sediment was determined using the methylene blue spectrophotometric method [12].
High-throughput sequencing of the V3-V4 region of the 16S rDNA gene was conducted on the sediment and activated sludge samples using amplicon sequencing technology. The primer sequences used were ACTCCTACGGGAGGCAGCAG and GGACTACHVGGGTWTCTAAT. The bioinformatics analysis of the sequencing data was conducted using the R language. Microbial alpha (α) diversity and beta (β) diversity were calculated using Mothur and the “vegan” package in the R language, respectively. The microbial co-occurrence network was developed using the “WGCNA” package in R 3.3.2, where nodes represented operational taxonomic units (OTUs) and edges represented correlations between OTUs. All the p-values were adjusted using the Multtest ini and Hochberg false discovery rate (fdr) control procedures for multiple testing.
The abundance of functional genes explored the elemental metabolic pathways in different ecosystems [13,14]. Tax4Fun has become a popular and robust tool for analyzing complex communities' functional genes [15,16]. The normalized bacterial OTUs table was imported to Tax4Fun to predict microbial metabolic function [17]. Tax4Fun function predictions based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) metabolic pathway database analysis have been used to predict functional genes [15].
2.4. Statistical Analysis
To estimate the random error of the measurements, all samples were analyzed in triplicate. The means, standard deviations, and analysis of variance (ANOVA) were calculated using SPSS 22.0. The means were compared using a one-way ANOVA, with the significance level set at P < 0.05.
3. Results and Discussion
3.1. Physiochemical Parameters
DO and ORP are vital indicators for evaluating the presence of black and odorous water. The impact of various remediation strategies on the DO and ORP in the overlying water is illustrated in Figure 1. As shown in Figure 1a, after 60 days of remediation, the DO in the overlying water gradually increased from 2.1 mg/L to 3.0 mg/L (TailN), 3.1 mg/L (TailN+CFM), and 5.6 mg/L (TailN+CFM+LDO). In contrast, the DO in the blank group (CN) exhibited a gradual decrease from the initial 2.1 mg/L to 1.4 mg/L. The changes in ORP followed a similar pattern. After 60 days of remediation, the ORP exhibited a gradual increase from the initial 100.2 mV to 181.6 mV (TailN), 189.5 mV (TailN+CFM), and 300.0 mV (TailN+CFM+LDO)(Figure 1b). In contrast, the ORP in the overlying water of the CN exhibited a gradual decrease from the initial 100.2 mV to -15.2 mV. Based on the changes in DO and ORP, it is evident that the water quality in the CN gradually deteriorated to a mild black and odorous level, while the water quality gradually improved, and the occurrence of black and odorous water was inhibited in the groups receiving the tailwater with NO3--N. NO3--N, as an oxidant, can oxidize reducing substances in the water, including easily oxidizable organic matter, S2-, and Fe2+ through the influence of heterotrophic denitrifying bacteria, sulfur autotrophic denitrifying bacteria, and iron autotrophic denitrifying bacteria, thereby improving water quality [3,4]. Moreover, the introduction of aerobic denitrifying bacteria in the TailN+CFM group may result in the consumption of DO in the water, as evident from the changes in both the DO and ORP in this group. In the TailN+CFM+LDO group, the introduction of oxygen significantly increased both the DO and ORP in the overlying water. The study demonstrates that high oxygen content can enhance the activity of facultative microorganisms at the water-sediment interface, promoting the decomposition and transformation of organic matter or reducible substances, thereby enhancing the self-purification capacity of the water [5]. Therefore, low-oxygen aeration is beneficial for improving the water-sediment microecosystem.
3.2. Changes in Sulfur Forms in Water
The primary cause of water blackening and odour is closely associated with the presence of the sulfur element in water. Sulfides are the primary source of odour in water. Furthermore, S2- combines with manganese and iron in the water to form MnS and FeS, which are the main causes of water blackening [2]. Consequently, to study the inhibitory effects of various remediation strategies on water blackening and odour, the changes in AVS in sediment and the concentration of SO42- in overlying water were analyzed (Figure 2).
The changes in SO42- concentration in the overlying water of different remediation groups are shown in Figure 2a. After 60 days of remediation, the SO42- concentration in the overlying water of the CN decreased by 26.9 mg/L, while in the other remediation groups, it exhibited a gradual increase to 36.5 mg/L (TailN), 35.5 mg/L (TailN+CFM), and 86.5 mg/L (TailN+CFM+LDO). From the perspective of the increased SO42- concentration, under low oxygen conditions, there is a significant increase in SO42 concentration in the overlying water. The study indicates that the impact of NO3--N, SO42- in the overlying water is mainly produced through sulfur autotrophic denitrification [18]. At the same time, with the introduction of oxygen, the DO and ORP in the overlying water are significantly increased, and the activity of sulfur-oxidizing bacteria at the water-sediment interface is further enhanced, increasing the SO42- concentration in the overlying water [5]. In the blank group, as the DO in the overlying water decreases, the concentration of SO42-- also decreases. The study indicates that the activity of anaerobic bacteria in the sediment gradually increases, and sulfate-reducing bacteria, as the main anaerobic bacteria in the sediment, participate in sulfate reduction reactions. This ultimately leads to the reduction of SO42-- in the overlying water [7].
The changes in AVS concentration in the sediment of different remediation groups are illustrated in Figure 2b. After 60 days of remediation, the AVS concentration in the remediation groups decreased to 2,460 g/kg (TailN), 2,560 g/kg (TailN+CFM), and 890 g/kg (TailN+CFM+LDO), reflecting declines of 18.3%, 15.3%, and 70.4% respectively. In contrast, the AVS concentration in the blank group increased from 3,010 g/kg to 3,458 g/kg. According to studies, the reduction of reducible sulfur in sediment is mainly achieved through sulfur autotrophic denitrification reactions with NO3--N [3]. The increase in AVS concentration in the blank group’s sediment is primarily associated with decreases in DO and ORP in the overlying water. As the reducing environment of the water body strengthens, the activity of sulfate-reducing bacteria in the sediment increases, resulting in an increase in AVS concentration [19].
3.3. Changes in the Morphology of Organic Matter in the Sediment
The changes of sediment organic matter concentration in different remediation groups are shown in Figure 3. After 60 days of remediation, the sediment organic matter in the remediation group was reduced to 112 g/kg (TailN), 98 g/kg (TailN+CFM), and 56 g/kg (TailN+CFM+LDO), which were 25.3% (TailN), 34.7% (TailN+CFM), and 62.7%, and 62.7% (TailN+CFM+LDO), respectively, however, sediment organic matter in the CN was reduced by only 10%. In the absence of remediation, the organic matter in the sediments was degraded mainly by sulfate-reducing bacteria, heterotrophic iron-reducing bacteria and electroactive bacteria. Since this study did not use isotopic labelling to calculate the amount of TOM consumed by the sulfate-reducing process and heterotrophic iron-reducing bacteria process, the generated AVS and Fe(II) may be overlooked for re-oxidation [20], and therefore the amount of TOM degraded by the sulfate-reducing process and heterotrophic iron-reducing bacteria process could not be accurately calculated.
In order to deeply investigate the change rule of organic matter, the changes of LFOM and HFOM during the remediation process were analysed, and the results are shown in Figure 4. On the 60th day of remediation, the LFOM in the sediment decreased from the initial 45 g/kg to 10 g/kg (TailN), 2 g/kg (TailN+CFM) and 0 g/kg (TailN+CFM+LDO), while the LFOM in the sediment of the CN group only degraded to 32 g/kg. The HFOM in the sediment decreased from the initial 105 g/kg to 102 g/kg (TailN), 96 g/kg (TailN+CFM) and 56 g/kg (TailN+CFM+LDO), while only 2 g/kg of HFOM was degraded in the CN group. The results showed that LFOM was preferentially degraded in the remediation groups, with significant degradation of HFOM in the TailN+CFM+LDO group. However, the degradation of HFOM could not be achieved without the participation of electroactive bacteria, suggesting that the TailN+CFM+LDO group may promote the degradation of HFOM by inducing electroactive bacteria. The main reason for this is that the extracellular electron transfer of electroactive bacteria enhanced HFOM degradation due to factors such as spatial site resistance or mass transfer [21,22]. Under aeration and denitrification, the DO and ORP in the water column were effectively enhanced, which increased the Fe(III) concentration in the water bodies and induced electroactive bacteria such as Pseudomonas and Thiobacillus.
3.4. Analysis of Sediment Microbial Community Structure
To investigate the changes in the microbial ecological environment of the sediment under various remediation measures, 16S amplicon sequencing analysis was performed on sediment samples collected on day 60 to identify the changes in the sediment microbial community structure. The results are shown in Figure 5. The relative abundance of Sterolibacterium [23], Brevundimonas [24], Silanimonas [25], Thiobacillus [19] and Denitratisoma [23], which are bacterial genera with denitrification functions, increased in the TailN, TailN+CFM, and TailN+CFM+LDO groups. Among them, the relative abundance of Sterolibacterium increased from 0.64% (CN) to 1.37% (TailN), 1.00% (TailN+CFM), and 3.21% (TailN+CFM+LDO). The results indicate that the addition of NO3--N in the tailwater increased the relative abundance of denitrifying bacteria genera. It is worth noting that the relative abundance of Sterolibacterium increased the most under low oxygen conditions, and previous studies have shown that low oxygen conditions can effectively stimulate the activity of denitrifying bacteria [5]. Additionally, two microorganisms capable of eliminating black and odour, Rhizobium [26] and Mycobacterium [27], were detected in the remediation groups. Their relative abundance increased compared to the blank group, with the highest increase observed under low oxygen conditions. The relative abundance of the sulfate-reducing bacterium Syntrophus [28] in the sediment decreased significantly with the introduction of nitrate in the tailwater, dropping from 1.39% (CN) to 1.27% (TailN), 1.37% (TailN+CFM), and 0.90% (TailN+CFM+LDO).
The relative abundance of electroactive bacteria Thiobacillus [19] and Pseudomonas [29,30] with denitrification ability increased in the remediation group, with the relative abundance of Thiobacillusincreasing 0.45% (CN) to 1.79% (TailN), 2.35% (TailN+CFM), and 3.84% (TailN+CFM+LDO). The microbial community structure indicates that the introduction of NO3--N in the tailwater greatly enhanced the microbial community structure of the sediment, with the greatest improvement observed under low oxygen conditions.
To further analyze the impact of TailN+CFM+LDO on the microbial ecological environment of the sediment, stratified sampling of the TailN+CFM+LDO sediment was conducted for microbial community structure analysis. The results are shown in Figure 6. In the shallow layer (0~4 cm), the relative abundance of bacteria genera with denitrification function, including Sterolibacterium [23]、Brevundimonas [24]、Silanimonas [25]、Thiobacillus [19] and Denitratisoma [23], was significantly higher than that of bacteria genera with denitrification function in the deep layer. Furthermore, it exhibited a decreasing trend with increasing depth. Additionally, Rhizobium [26] and Mycobacterium [27], which are microorganisms capable of eliminating black and odour, and Thiobacillus [19], a sulfur autotrophic denitrifying bacterium, exhibited similar patterns of relative abundance changes. The relative abundance of Syntrophus [28], a sulfate-reducing bacterium, in the sediment exhibited an opposite pattern of change. Combining the changes in concentrations of DO, ORP, NH3-N, NO3--N, NO2--N, and SO42- at different depths in the sediment (Figure S2), it can be inferred that denitrification mainly occurs within the range of -8 cm to 0 cm, whereas sulfate reduction mainly occurs within the range of -16 cm to -8 cm. The results indicate that the microbial community structure in the sediment within the range of -8 to 0 cm is significantly improved by TailN+CFM+LDO treatment.
3.5. Mechanisms of Improvement in Sediment Microbial Ecological Environment
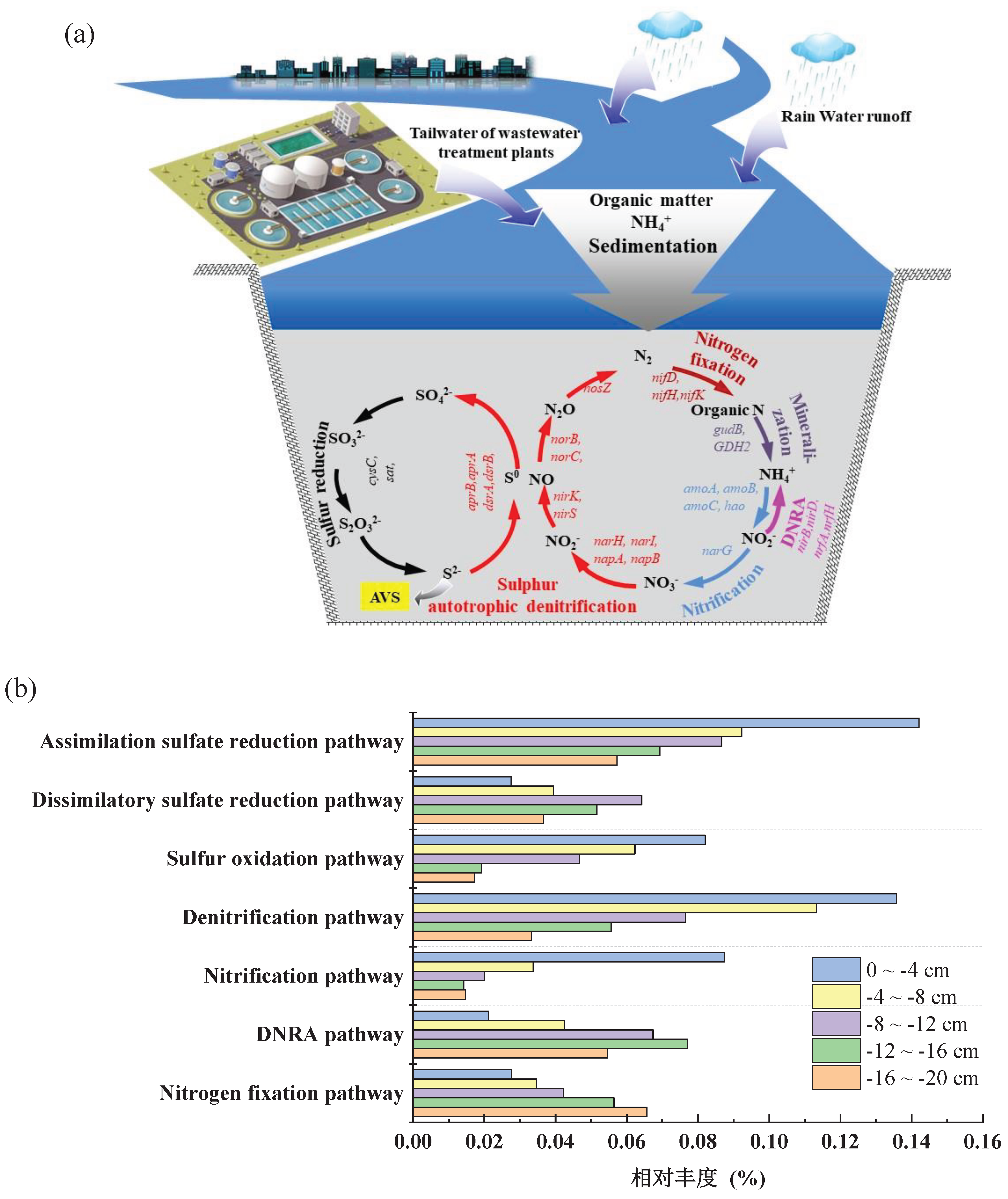
To further analyze the mechanisms of improvement in the sediment microbial ecological environment, the PICRUSt software was used to predict the relative abundance of nitrogen and sulfur metabolism pathways, as well as essential genes, in various sediment layers of the TailN+CFM+LDO group. The results are shown in Figure 7. Key genes involved in nitrogen cycling include denitrification genes (such as narH, napA, nirK, nosZ, and norB), nitrogen fixation genes (such as nifD and nifH), ammonia oxidation genes (such as amoA and amoB), and DNRA genes (such as nirB and nirD) [31,32]. Nitrogen fixation and DNRA genes promote the formation of NH3-N, whereas denitrification and ammonia oxidation genes promote nitrogen removal [33,34]. The relative abundance of nitrogen fixation genes and DNRA genes in the deep sediment layer (-20~-12 cm) was significantly higher than that in the shallow sediment layer (-12~0 cm). The relative abundance of denitrification and ammonia oxidation genes in the shallow sediment layer (-12~0 cm) was significantly higher than that in the deep sediment layer (-20~-12 cm). Key sulfur cycling processes include sulfur oxidation (such as Sox, SoxA, SoxB, SoxD, and SoxG), dissimilatory sulfate reduction (such as dsrB, dsrC, aprA, and aprB), and assimilatory sulfate reduction (such as sat, cysC, cysN, and sir) [35]. The increase in the relative abundance of Thiobacillus, a sulfur autotrophic denitrifying bacterium, indicates that sulfur autotrophic denitrification is the primary mechanism in the shallow sediment layer (-4~0 cm) [36,37]. The presence of a high concentration of sulfide in sediment promotes the utilization of nitrogen and sulfur by sulfur bacteria [5,38]. Researchers discovered that the degradation of AVS in the sediment is primarily driven by sulfur-driven autotrophic denitrification [39,40]. The relative abundance of sulfur oxidation genes in the shallow sediment layer (-12~0 cm) was significantly higher than that in the deep sediment layer (-20~-12 cm). The relative abundance of dissimilatory sulfate reduction genes in the shallow sediment layer (-12~0 cm) was significantly lower than that in the deep sediment layer (-20~12 cm). Dissimilatory sulfate reduction is the primary biochemical reaction responsible for H2S production in anaerobic environments, while assimilatory sulfate reduction results in the production of organic sulfur compounds such as cysteine and methionine. In summary, under the conditions of TailN+CFM+LDO, the enhancement of sulfur oxidation in the shallow sediment layer (-4~0 cm) inhibits dissimilatory sulfate reduction, thereby promoting the oxidation of AVS in the sediment. This process promotes sulfur-driven autotrophic denitrification [37,41].
4. Conclusion
In this study, it was discovered that the nitrate remediation measure in the tailwater of wastewater treatment plants combined with aerobic denitrifying bacteria under low-DO regulation effectively increased the DO concentration and ORP in the overlying water, thereby enhancing the activity of electroactive bacteria, denitrifying and sulfur-oxidizing microorganisms in the sediment. This resulted in an increase in the quantity and activity of key enzymes involved in the nitrification and denitrification processes. With the improvement of the microbial ecological environment, the acid volatile sulfide in the sediment decreased by 70.4%, and the organic matter in the sediment was reduced by 62.7%, in which the HFOM was degraded from 105 g/kg to 56 g/kg, and the potential risk of water reverting to black and odorous conditions significantly decreased. The results indicate that TailN+CFM+LDO has a promising remediation effect on the sediment microenvironment.
Acknowledgements
This study was supported by the National Natural Science Foundation of China(40871232);Beijing Natural Science Foundation-Joint key projects of the Education Commission (KZ201810020025).
References
- Gong, S.; Donde, O.O.; Cai, Q.; Wu, X.; Song, K.; Wang, C.; Hong, P.; Xiao, B.; Tian, C. Improved lakeshore sediment microenvironment and enhanced denitrification efficiency by natural solid carbon sources. Int. J. Sediment Res. 2022, 37, 823–832. [Google Scholar] [CrossRef]
- Cao, J.; Sun, Q.; Zhao, D.; Xu, M.; Shen, Q.; Wang, D.; Wang, Y.; Ding, S. A critical review of the appearance of black-odorous waterbodies in China and treatment methods. J. Hazard. Mater. 2020, 385, 121511. [Google Scholar] [CrossRef]
- Yin, H.; Yang, P.; Kong, M. Effects of nitrate dosing on the migration of reduced sulfur in black odorous river sediment and the influencing factors. Chem. Eng. J. 2019, 371, 516–523. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.-H.; Yan, F.-L.; Ding, Z.; Feng, L.-L.; Zhao, J.-C. Effects and mechanisms of calcium peroxide on purification of severely eutrophic water. Sci. Total Environ. 2019, 650, 2796–2806. [Google Scholar] [CrossRef]
- Li, W.; Zhang, S.; Zhang, L.; Li, X.; Wang, F.; Li, G.; Li, J.; Li, W. In-situ remediation of sediment by calcium nitrate combined with composite microorganisms under low-DO regulation. Sci. Total Environ. 2019, 697, 134109. [Google Scholar] [CrossRef]
- Zhang, C.; Li, M.; Sun, J.; Zhang, S.; Huang, J. The mechanism of C-N-S interconnection degradation in organic-rich sediments by Ca(NO3)(2)-Ca-O2 synergistic remediation. Environ. Res. 2022, 214, 1–12. [Google Scholar] [CrossRef]
- Liu, X.; Tao, Y.; Zhou, K.; Zhang, Q.; Chen, G.; Zhang, X. Effect of water quality improvement on the remediation of river sediment due to the addition of calcium nitrate. Sci. Total Environ. 2017, 575, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Zhang, X.; Xue, Y. Application of calcium peroxide in water and soil treatment: A review. J. Hazard. Mater. 2017, 337, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Roberts, K.L.; Grace, M.R.; Kessler, A.J.; Cook, P.L.M. Role of organic carbon, nitrate and ferrous iron on the partitioning between denitrification and DNRA in constructed stormwater urban wetlands. Sci. Total Environ. 2019, 666, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Hou, L.; Gao, D.; Liu, M.; Liu, C.; Zhang, Z. Organic matter degradation state affects dissimilatory nitrate reduction processes in Knysna estuarine sediment, South Africa. J. Soils Sediments 2021, 21, 3202–3212. [Google Scholar] [CrossRef]
- Wang, F.; Yang, F.; Gao, H.; Bai, Y.; Liao, H.; Li, H. Simulation of Denitrification Process of Calcium Nitrate Combined with Low Oxygen Aeration Based on Double Logarithm Mode. Water 2022, 14, 269. [Google Scholar] [CrossRef]
- Zhang, C.; Li, M.; Zhang, S.; Qi, Z.; Huang, J.; Sun, J. An improved method of fluorescein diacetate determination for assessing the effects of pollutants on microbial activity in urban river sediments. J. Soils Sediments 2022, 22, 2792–2801. [Google Scholar] [CrossRef]
- Shu, D.; He, Y.; Yue, H.; Wang, Q. Metagenomic and quantitative insights into microbial communities and functional genes of nitrogen and iron cycling in twelve wastewater treatment systems. Chem. Eng. J. 2016, 290, 21–30. [Google Scholar] [CrossRef]
- Nelson, M.B.; Martiny, A.C.; Martiny, J. Global biogeography of microbial nitrogen-cycling traits in soil. Proc. Natl. Acad. Sci. USA 2016, 113, 8033–8040. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sun, F.; Zhao, H.; Mi, H.; He, S.; Chen, Y.; Liu, Y.; Lan, H.; Zhang, M.; Wang, Z. Compositional changes of sedimentary microbes in the Yangtze River Estuary and their roles in the biochemical cycle. Sci. Total Environ. 2021, 760, 143383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, M.; Sun, J.; Huang, J.; Chang, S. Fluorescein diacetate hydrolytic activity as a sensitive tool to quantify nitrogen/sulfur gene content in urban river sediments in China. Environ. Sci. Pollut. Res. 2023, 30, 62544–62552. [Google Scholar] [CrossRef]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef]
- He, Z.; Huang, R.; Liang, Y.; Yu, G.; Chong, Y.; Wang, L. Index for nitrate dosage calculation on sediment odor control using nitrate-dependent ferrous and sulfide oxidation interactions. J. Environ. Manag. 2018, 226, 289–297. [Google Scholar] [CrossRef]
- Wang, L.; Long, X.; Chong, Y.; Yu, G. Potential risk assessment of heavy metals in sediments during the denitrification process enhanced by calcium nitrate addition: Effect of AVS residual. Ecol. Eng. 2016, 87, 333–339. [Google Scholar] [CrossRef]
- Luo, M.; Zhu, W.; Huang, J.; Liu, Y.; Duan, X.; Wu, J.; Tong, C. Anaerobic organic carbon mineralization in tidal wetlands along a low-level salinity gradient of a subtropical estuary: Rates, pathways, and controls. Geoderma 2019, 337, 1245–1257. [Google Scholar] [CrossRef]
- Tan, W.; Zhao, X.; Dang, Q.; Cui, D.; Xi, B. Microbially reducible extent of solid-phase humic substances is governed by their physico-chemical protection in soils: Evidence from electrochemical measurements. Sci. Total Environ. 2020, 708, 134683. [Google Scholar] [CrossRef]
- Li, M.; Sun, J.; Liu, C.; Tang, Y.; Huang, J. The remediation of urban freshwater sediment by humic-reducing activated sludge. Environ. Pollut. 2020, 265, 115038. [Google Scholar] [CrossRef]
- Yu, J.; Fan, C.; Zhong, J.; Zhang, Y.; Wang, C.; Zhang, L. Evaluation of in situ simulated dredging to reduce internal nitrogen flux across the sediment-water interface in Lake Taihu, China. Environ. Pollut. 2016, 214, 866–877. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, Y.; Huang, T.; Liu, Y.; Fang, K.; Zhang, C. Microbial aerobic denitrification dominates nitrogen losses from reservoir ecosystem in the spring of Zhoucun reservoir. Sci. Total Environ. 2019, 651, 998–1010. [Google Scholar] [CrossRef]
- Manucharova, N.A.; Vlasenko, A.N.; Tourova, T.P.; Panteleeva, A.N.; Stepanov, A.L.; Zenova, G.M. Thermophilic chitinolytic microorganisms of brown semidesert soil. Microbiology 2008, 77, 610–614. [Google Scholar] [CrossRef]
- Su, J.F.; Zhang, Y.M.; Liang, D.H.; Wang, J.X.; Wang, Z.; Li, M. Performance and microbial community of an immobilized biofilm reactor (IBR) for Mn(II)-based autotrophic and mixotrophic denitrification. Bioresour. Technol. 2019, 286, 121407. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Oguchi, H.; Kobayashi, T.; Kusama, S.; Sugiura, R.; Moriya, K.; Hirata, T.; Yukioka, Y.; Takaya, N.; Yajima, S.; et al. Nitrogen oxide cycle regulates nitric oxide levels and bacterial cell signaling. Sci. Rep. 2016, 6, 22038. [Google Scholar] [CrossRef]
- Keshri, J.; Ram, A.S.P.; Sime-Ngando, T. Distinctive Patterns in the Taxonomical Resolution of Bacterioplankton in the Sediment and Pore Waters of Contrasted Freshwater Lakes. Microb. Ecol. 2018, 75, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Zhang, L.; Li, D.; Zhan, G.; Qian, J.; Tao, Y. Dissimilatory nitrate reduction by Pseudomonas alcaliphila with an electrode as the sole electron donor. Biotechnol. Bioeng. 2012, 109, 2904–2910. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, Y.; Su, W.; Jiang, Y.; Su, M.; Gao, P.; Li, D. Effects of cathode potentials and nitrate concentrations on dissimilatory nitrate reductions by Pseudomonas alcaliphila in bioelectrochemical systems. J. Environ. Sci. 2014, 26, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Peng, Y.; Fan, L.; Zhang, L.; Ni, B.; Kartal, B.; Feng, X.; Jetten, M.S.M.; Yuan, Z. Metagenomic analysis of anammox communities in three different microbial aggregates. Environ. Microbiol. 2016, 18, 2979–2993. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, L.; Li, Y.; Zhang, W.; Niu, L.; Wang, L. The bacterial community structure and N-cycling gene abundance in response to dam construction in a riparian zone. Environ. Res. 2021, 194, 110717. [Google Scholar] [CrossRef]
- Hu, Y.; Hong, Y.; Ye, J.; Wu, J.; Wang, Y.; Ye, F.; Chang, X.; Long, A. Shift of DNRA bacterial community composition in sediment cores of the Pearl River Estuary and the impact of environmental factors. Ecotoxicology 2021, 30, 1704–1704. [Google Scholar] [CrossRef]
- Wang, S.; Wu, B.; Wang, Y.; Wang, X. Nitrate Migration and Transformation in Low Permeability Sediments: Laboratory Experiments and Modeling. Water 2023, 15, 2528. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.; Sun, Q.; Chen, P.; Wei, X. Correlations of functional genes involved in methane, nitrogen and sulfur cycling in river sediments. Ecol. Indic. 2020, 115, 106411. [Google Scholar] [CrossRef]
- Chen, M.; Zhou, X.-F.; Yu, Y.-Q.; Liu, X.; Zeng, R.J.-X.; Zhou, S.-G.; He, Z. Light-driven nitrous oxide production via autotrophic denitrification by self-photosensitized Thiobacillus denitrificans. Environ. Int. 2019, 127, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Li, Z.; Hou, Y.; Wang, A.; Liu, Q.; Huang, C. Effects of different carbon sources on the efficiency of sulfur-oxidizing denitrifying microorganisms. Environ. Res. 2022, 204, 111946. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Ekama, G.A.; Wang, Y.; Dai, J.; Biswal, B.K.; Chen, G.; Wu, D. Advances in sulfur conversion-associated enhanced biological phosphorus removal in sulfate-rich wastewater treatment: A review. Bioresour. Technol. 2019, 285, 121303. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Pang, Y.; Ji, G. Increase of N2O production during nitrate reduction after long-term sulfide addition in lake sediment microcosms. Environ. Pollut. 2021, 291, 118231. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Wang, J.; Li, S.; Ji, G. Long-term sulfide input enhances chemoautotrophic denitrification rather than DNRA in freshwater lake sediments. Environ. Pollut. 2021, 270, 116201. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Liu, Q.; Wang, H.; Gao, L.; Hou, Y.-N.; Nan, J.; Ren, N.; Li, Z.-L. Influence of microbial spatial distribution and activity in an EGSB reactor under high- and low-loading denitrification desulfurization. Environ. Res. 2021, 195, 110311. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Variation of DO (a) and ORP (b) in the overlying water.
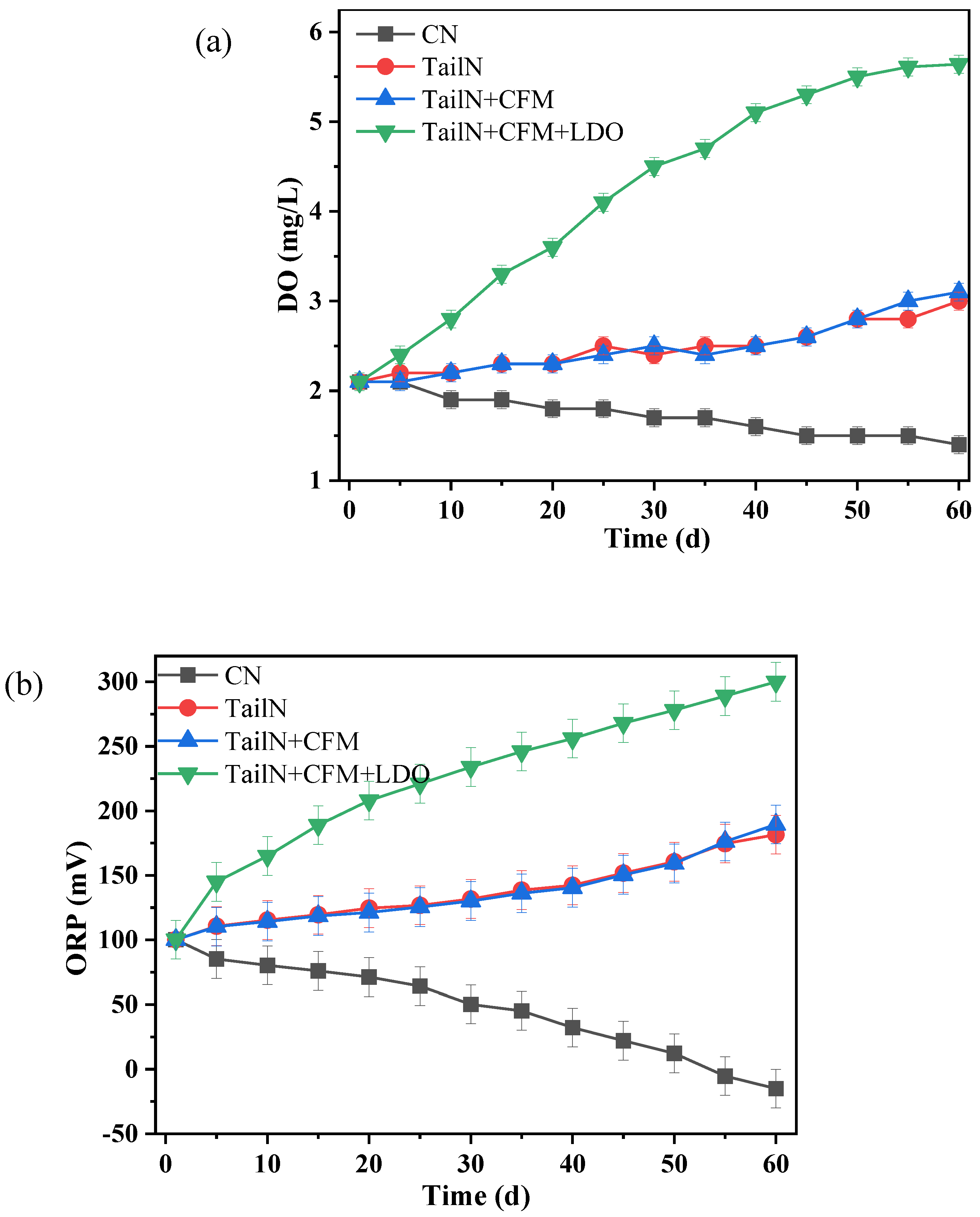
Figure 2.
Variation of overlying water SO42- (a) and sediment AVS (b) during restoration.
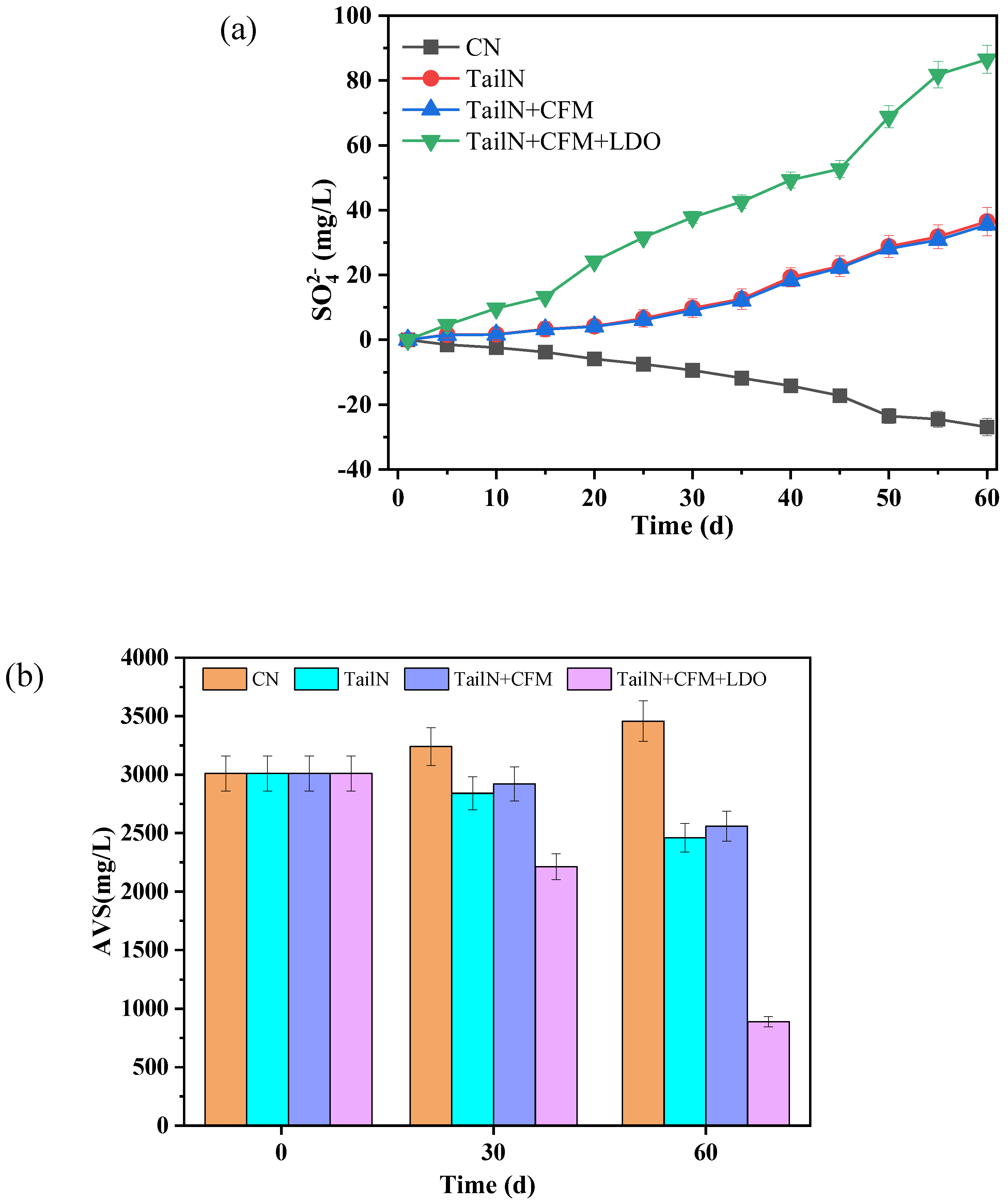
Figure 3.
Concentration of TOM removal.
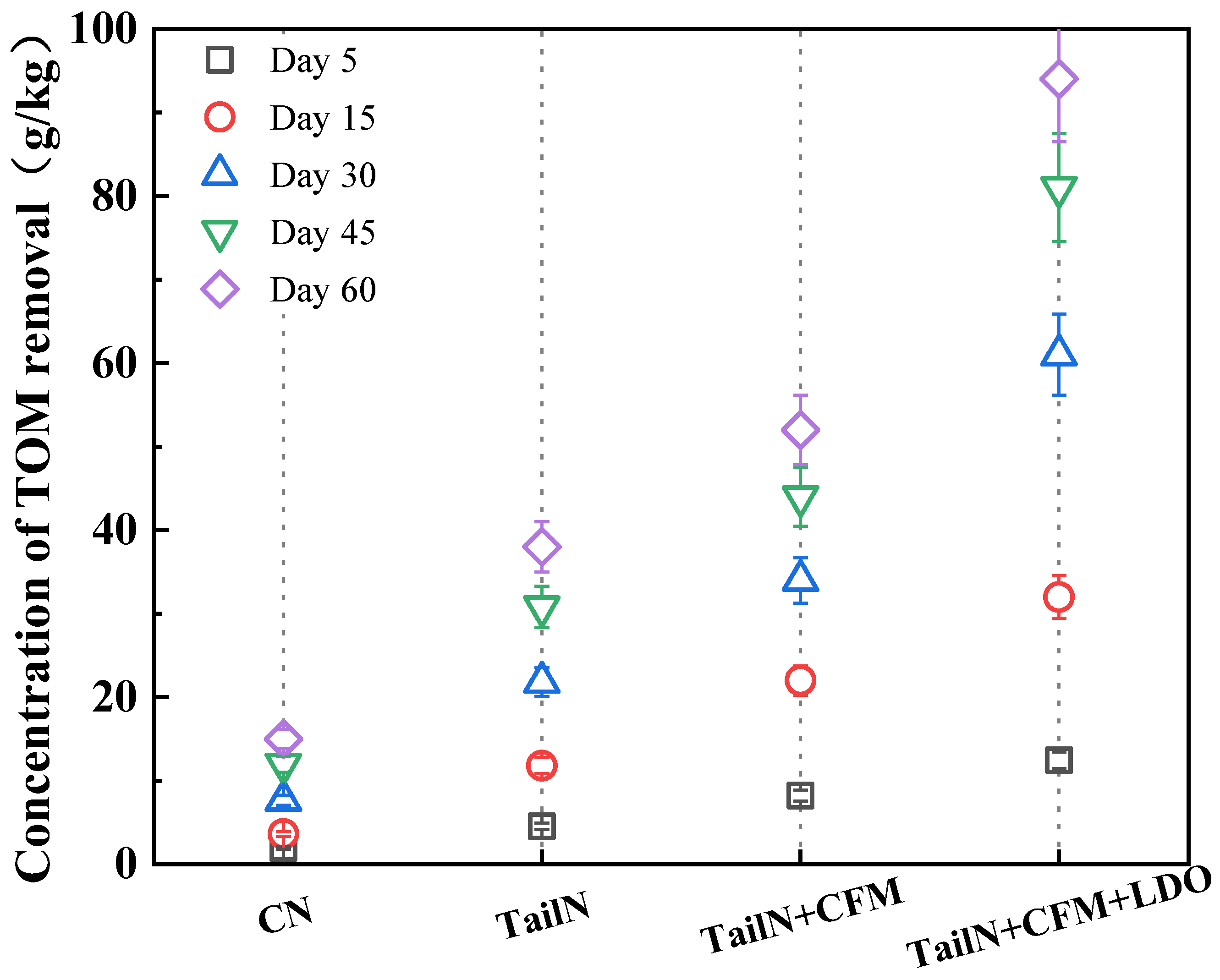
Figure 4.
Concentration of LFOM/HFOM.
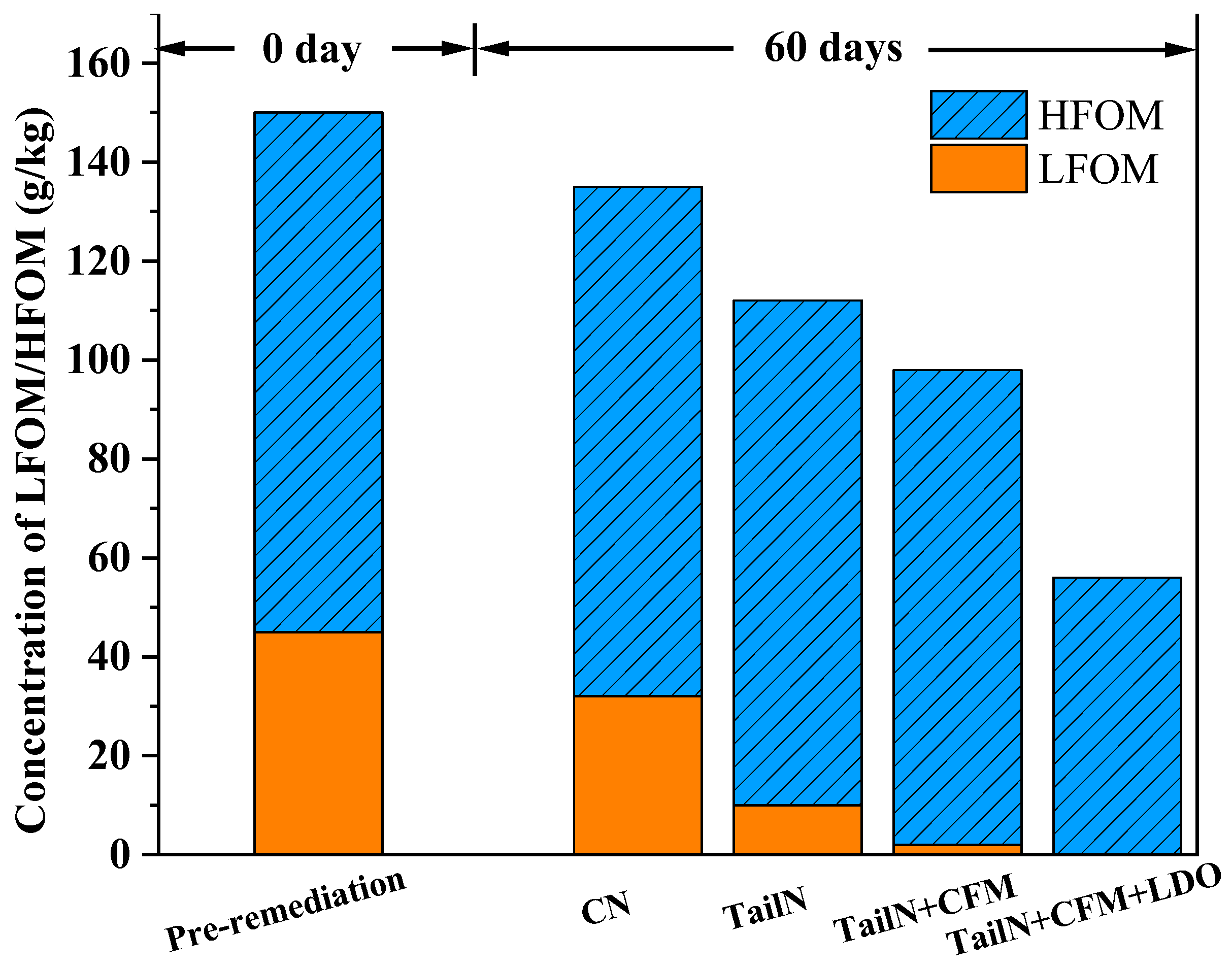
Figure 5.
Composition and abundance of the sediment samples at genus levels.
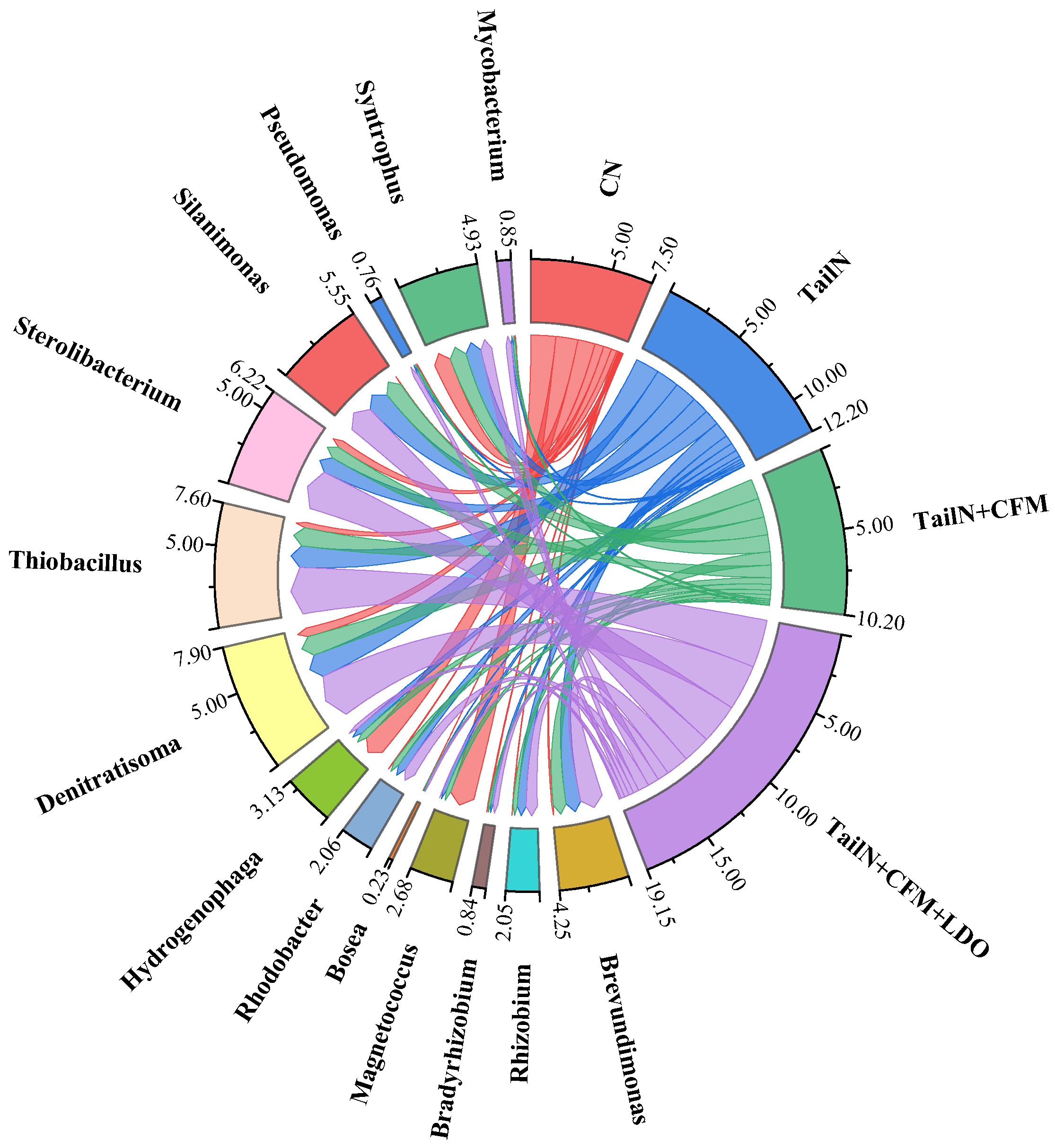
Figure 6.
Downcore distribution of composition and abundance in different sediment sections at genus levels.
Figure 6.
Downcore distribution of composition and abundance in different sediment sections at genus levels.
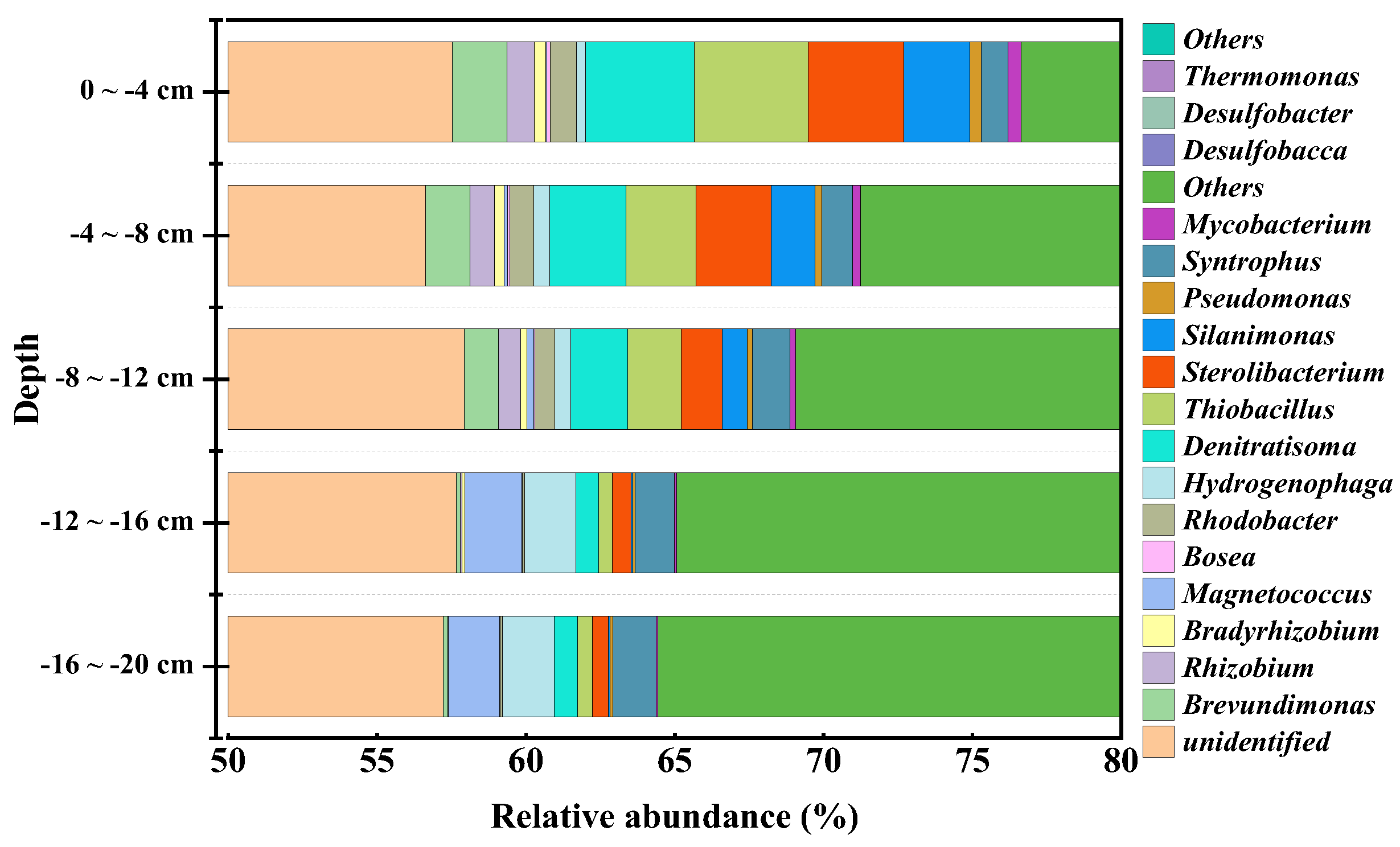
Figure 7.
Nitrogen and sulfur metabolic pathways (a) and relative abundance of key genes (b) in sediments.
Figure 7.
Nitrogen and sulfur metabolic pathways (a) and relative abundance of key genes (b) in sediments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



