Submitted:
01 March 2024
Posted:
04 March 2024
Read the latest preprint version here
Abstract
Infant mortality remains a major global concern. Sudden unexpected death in infancy (SUDI) is reported globally and accounted for 40% of infant deaths between 2012 and 2016 in the Western Cape. Research highlighting molecular typing of respiratory viruses in SUDI cases is lacking. A total of 116 PCR-positive human rhinovirus (HRV) and respiratory syncytial virus (RSV) swab samples collected from the lungs and trachea of SUDI cases admitted to Tygerberg Medico-legal Mortuary between 2015 and 2019 were included and underwent automated nucleic acid extraction, one-step, nested RT-PCR, and amplification by gel electrophoresis before sequencing and phylogenetic analysis. The median age of the infants were 10.9 weeks. Three distinct HRV species were identified; HRV-A (n=28), followed by -C (n=11), and -B (n=4), including eight HRV-A, one -B and seven -C genotypes. Both RSV-A (n=5) and -B (n=5) were detected. No RSV-A sequences were assigned as ON1, and two samples were assigned BA9 after amino acid alignment indicating characteristic of 20 amino acid duplication, as well as nine substitutions. This study describes the first molecular and phylogenetic characterisation of respiratory viruses in SUDI cases in Africa. Future studies should include identifying circulating HRV and RSV strains associated with clinical disease severity.
Keywords:
respiratory infection
; human rhinovirus
; respiratory syncytial virus
; genotypes
; SUDI / SIDS
; infant death
; Sanger sequencing
; phylogenetic analysis
1. Introduction
T Lower respiratory tract infection (LRTI) is a major barrier to the United Nations Millennium Development Goal to reduce global under-five child mortality [1]. Human rhinovirus (HRV) and respiratory syncytial virus (RSV) are common causes of LRTIs in infants and are associated with wheezing and childhood asthma [2,3,4,5,6,7,8,9,10,11].
HRV was first discovered in the 1950s in effort to identify the aetiology of the common cold [12,13] and is often considered a common cold virus, although it has been implicated as a leading cause of infant hospitalisation [5,14,15]. Primary infection through inhalation of aerosolised droplets or physical inoculation after contact with fomites is followed by a 2-day incubation period [16]. Viral replication and infection occur in the epithelial cells of the upper respiratory tract and LRTI [4,17]. Children are the main reservoir and experience infection four times more frequently than adults [5,12,14,18]. HRV is a member of the family Picornaviridae and the genus Enterovirus. These viruses are positive sense (ps), single-stranded (ss) RNA viruses. The 7 200 base pair (bp) genome consists of structural and non-structural regions, flanked by 5’ and 3’ untranslated regions (UTR). The virus is classified by genotype into HRV-A (80 serotypes), HRV-B (32 serotypes), and HRV-C (57 serotypes), and by cell surface receptor differences into Major and Minor groups. Over 160 genotypes exist [16,19]. Genotype differentiation of HRV is commonly achieved by reverse transcriptase polymerase chain reaction (RT-PCR) targeting the 5′UTR and VP4/2 followed by sequence analysis [20,21,22,23,24,25,26,27].
RSV was first identified in 1956 in a chimpanzee during an outbreak of coryza. The virus was later detected in humans which remain the only host [9,28]. Similar to HRV, RSV is also noted as a leading global cause of LRTI and hospitalisations in infants [8,29,30,31,32,33,34]. Mortality may be lower in healthy infants in developed countries; however, regardless of the socioeconomic status, infants under the age of six months have the same high risk of childhood asthma and recurrent wheeze when infection occurs during peak RSV season [9,35]. In 2016 RSV was reclassified into the family of Pneumoviridae, genus Orthopneumovirus. Pneumoviridae used to be a subfamily within the Paramyxoviridae family [36]. The virus is a non-segmented, negative sense (ns) enveloped ssRNA virus with an approximate genome length of 15 000 bp [28]. Subtypes are based on sequence variations of the second hypervariable region (HVR), or HRV2, of the RSV G-gene [37]. Immune pressure, inherent lack of proof-reading capabilities, and duplication events in the RSV G-gene ectodomain led to the detection of the Buenos Aires (BA) and the Ontario genotypes (ON) in 1999 and 2010 respectively [31,38,39]. The ON1 and BA strains of RSV-A and -B respectively, are differentiated by nucleotide duplications in the genome 31]. The relationship between genotype variation and clinical severity is unclear. Both RSV-A and -B co-circulate with one predominating subtype each, after ON and BA genotypes have rapidly become the predominant subtypes across the globe [28].
All three HRV species circulate within South Africa, commonly infecting young children. HRV-A and -C predominate circulation, similar to Jordan, Hong Kong, and Taiwan 4,6,16,40]. A previous South African study enrolled 220 children presenting with wheeze at Red Cross War Memorial Children’s Hospital, Cape Town. HRV was most common (58%) and all genotypes, with HRV-C (52%) predominating, while HRV-A (37%), and -B (11%), were also detected. More than 70% of these children were under the age of 2 years [4]. Another South African study [42] confirmed this HRV-species-specific dominance, however, neither of these studies investigated any relationship between HRV genotype and disease severity [4,42]. A more recent study conducted in sub-Sahara Africa, specifically Mali, South Africa, and Zambia, found a higher prevalence of HRV-A in infants [25]. Clinical and molecular epidemiology data in low- and middle-income countries in Africa are limited and the specific role of HRV in childhood respiratory disease needs elucidation [4,25].
Both RSV-A and -B subtypes circulate in South Africa and are associated with LRTIs [44,45]. RSV-associated hospitalisations is highest in infants [46]. The Drakenstein Child Health Study is an ongoing birth cohort longitudinal study in the Western Cape, South Africa. Of the 1 143 infants recruited into the study, 690 never developed LRTI and 152 developed LRTI as a result of RSV. Recurrent wheeze was over ten-fold higher in this latter group (43%) than in those with no prior infection (3%). LRTI and hospitalisation as a result of RSV were most common in winter and infants younger than 6 months of age, becoming less common with increasing age. Risk factors included maternal smoking, maternal human immunodeficiency virus status, season of birth, and child age [11].
Globally, respiratory viral infections is a major burden in infants [15,32,47,48,49]. HRV, RSV, human CoV, and human adenovirus are commonly detected in infants with respiratory illness, as well as sudden unexpected death in infancy (SUDI) cases [50,51,52,53].
SUDI is defined as the sudden and unexpected death in an infant with no clinical signs suggesting possible illness up to two days prior to death [54,55]. These cases are mandated to undergo a medico-legal investigation in South Africa to determine the cause of death. If no cause of death can be confirmed after an extensive autopsy, review of medical history, and death scene investigation, such cases are classified as sudden infant death syndrome (SIDS) [56,57,58]. Thus, SIDS is a subset of SUDI [59,60]. Both are regarded as leading causes of infant mortality [60,61].
Research investigating infections in SUDI cases is limited in South Africa. This will be the first report to describe HRV and RSV genotypes in SUDI cases admitted to Tygerberg Hospital in South Africa.
2. Materials and Methods
This study was approved by the Health Research Ethics Committee (HREC) of Stellenbosch University, Cape Town, South Africa (HREC Registration number: N12/02/007).
During autopsy, flocculated FLOQSwabs™ (Copan Flock technologies, Italy) samples were collected from the trachea and left lower lobes of the lungs of SUDI cases between 2015 and 2019 and screened with the Seegene AllplexTM RV Essential multiplex real-time RT-PCR assay (Seegene, South Korea) as per manufacturer’s instructions for a panel of respiratory viruses. All samples positive for HRV or RSV were included in this study.
Samples underwent automated RNA extraction using Microlab STAR Hamilton (Hamilton Company, USA) with the NucleoMag Pathogen Kit (Macherey-Nagel, Germany) as per the manufacturer’s guidelines. A one-step, nested RT-PCR assay was optimised for specific gene amplification of HRV and RSV.
Primers for genotyping were selected from the literature for HRV [22,62] (Accession number: NC_038312) and for the RSV G-gene [63] (Accession number: NC_001803). Primer annealing and target specificity were determined by Geneious Prime® 2021.1.1. Primers were manufactured by Integrated DNA Technologies (USA). The Promega Access RT-PCR (USA) System was used for the pre-nested reaction (cDNA synthesis and PCR) and the Promega GoTaq FlexiKit (USA) for the nested reaction.
The pre-nested and nested mastermix used to sequence the HRV 5′UTR are presented in Table 1 and the primers for the amplification of the HRV 5′UTR and VP4 regions in Table 2 [22].
The master mix for the RSV G-gene pre-nested and nested sequencing is presented in Table 3 and the primers for the amplification of the G-gene region 2016) in Table 4 [63].
The pre-nested and nested thermocycling conditions for HRV and RSV were similar, the changes to the protocol for RSV are written in brackets. In brief, the pre-nested thermocycling conditions were 45 minutes at 48°C, 2 minutes at 94°C, followed by 45 cycles of 30 seconds at 94°C, 90 seconds at 60°C (55°C), and 1 minute at 72°C, concluding with a single cycle of 10 minutes at 72°C. Hold was set at 4°C.
The nested thermocycling conditions were 2 minutes at 94°C, followed by 40 cycles of 30 seconds at 94°C, 1 minute at 55°C (50°C), and 30 seconds at 72°C, concluding with a single cycle of 10 minutes at 72°C. Hold was set at 4°C.
Following extraction, PCR amplicons were visualised on a 2% agarose gel at 200V for 40 minutes. Visualisation was completed using the Bio-Rad ChemiDocTM (US A/N 492085) with Image LabTM Touch Software X (Version 2.4.0.03) set for GelGreen Gel (590/110; UV Trans with auto optimal exposure).

Table 4.
Pre-Nested and Nested Thermocycling conditions.
| Pre-Nested | Cycles | Duration | HRV Assay | RSV Assay |
|---|---|---|---|---|
| Initial denaturation | 1 | 45 minutes | 48°C | |
| PCR activation | 1 | 2 minutes | 94°C | |
| Denaturation | 30 seconds | 94°C | ||
| Annealing | 45 | 90 seconds | 60°C | 55°C |
| Extension | 60 seconds | 72°C | ||
| Final extension | 1 | 10 minutes | 72°C | |
| Final hold | 1 | Infinite | 4°C | |
| Nested | ||||
| PCR activation | 1 | 2 minutes | 94°C | |
| Denaturation | 30 seconds | 94°C | ||
| Annealing | 40 | 60 seconds | 55°C | 50°C |
| Extension | 30 seconds | 72°C | ||
| Final extension | 1 | 10 minutes | 72°C | |
| Final hold | 1 | Infinite | 4°C |
Post-amplification samples were purified using the QIAquick Purification kit (Qiagen, USA) to remove dNTPs, primers, proteins and other impurities that may cause inhibition during the sequencing step. Silica membrane technology is used to absorb the negative DNA to a slightly positively charged column membrane. Centrifugation and wash steps were performed according to the manufacturer’s instructions before elution of the final samples. All samples were then subjected to Sanger Sequencing.
Phylogenetic analysis was performed by assembling patient consensus sequences with De Novo Assemble tool of forward and reverse sequences in Geneious Prime® 2021.1.1. Each chromatogram was assessed for overlapping fluorescent peaks prior to trimming. The Basic Local Alignment Search Tool (BLAST) within Geneious Prime was used to identify reference sequences for each nucleotide sequence. For HRV, Coxsackie virus (M16560) and Poliovirus 1 (V01149), and for RSV, Bovine orthopneumovirus (AF092942), listed by the International Committee on Taxonomy of Viruses website, were selected as outgroups for phylogenetic analysis. The accession numbers were searched on GenBank and sequences downloaded. All files were uploaded to Multiple Alignment using Fast Fourier Transformation version 7 software for multiple sequence alignment. Randomised Axelerated Maximum Likelihood (RAxMLGUI 2.0) was used to determine optimal model test (HRV: General Time Reversible substitution model with gamma distributed rate variation and invariant sites; RSV: Hasegawa-Kishino-Yano model with gamma distributed rates), maximum likelihood tree, and bootstrapping support for phylogenetic analysis achieved with 1 000 bootstrap replicates. Where applicable, species and genotype assignments were assigned based on clustering on the phylogenetic tree and BLAST results from the National Centre for Biotechnology Information (NCBI) website (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
RSV-A and -B nucleotide sequences were translated to amino acids sequences using standard code in Geneious Prime® 2021.1.1. Thereafter, files were uploaded to Multiple Alignment using Fast Fourier Transformation version 7 software for multiple sequence alignment. The file was viewed using DNASTAR Version 17.4.1.17. For comparison between the SUDI sample sequences and RSV subtypes, the HVR2 of RSV-A patient sequences were aligned with prototype strain RSV ON1 (Accession number: JN257693; ON67-1210A), and RSV-B sequences were aligned with prototype strain RSV-BA (Accession number: AY333364; BA/4128/99B). N-Glycosylation sites were predicted by uploading translated amino acid sequences to online program NetNGlyc 1.0.
3. Results
HRV and RSV samples with Ct values below 40 and 30 respectively as confirmed by Seegene AllplexTM RV Essential testing in this study were included in manual PCR. Of the 106 expected HRV PCR-positive samples detected by the previous screening studies (2015/2016, n=68; 2018/2019, n=38), 64 (60%) were PCR-positive on repeat testing (2015/2016, n=49; 2018/2019, n=15) and all were selected for manual PCR.
Of the 56 expected RSV-positive samples detected by the previous screening studies (2015/2016, n=11; 2018/2019, n=45), only 15 (26%) were PCR-positive on repeat testing (2015/2016, n=10; 2018/2019, n=5), of which 10 were selected for manual PCR as the Ct values were within the RSV manual PCR assay amplification limit determined by Ct value. The Ct values for RSV-positive samples G03 (Ct: 29.30) and G08 (Ct: 27.99) were higher than the detection limit previously determined by the RSV manual PCR assay, but these samples were included, and amplification was successful.
Table 5 summarises the number of positive samples for HRV and RSV for the previous collection periods (Expected), as well as HRV and RSV results obtained by the confirmatory Seegene Allplex™ RV Essential kit (Detected). Samples selected for manual PCR met inclusion criteria for manual PCR detection limit by measure of Ct value (HRV: <40; RSV: <30). PCR products were obtained from a total of 60 HRV and 10 RSV positive samples which successfully amplified, of which 43 HRV and all 10 RSV samples were successfully sequenced.
HRV samples produced three distinct species , HRV-A (n=28), followed by -C (n=11), and -B (n=4). In total, eight HRV-A (A28, A80, A10, A82, A43, A56, A11, and A56), one -B (B84), and seven -C (C22, C19, C24, C35, C29, C38, and C2) genotypes were identified. HRV-A was detected in all seasons, while -C was detected in the cooler seasons, and -B was only detected in winter.
RSV-A and RSV-B were confirmed in five samples each. None of the RSV-A sequences were assigned as ON1, and three samples were assigned BA9 after phylogenetic analysis. RSV-A was detected in samples from all seasons, except summer, while RSV-B was detected throughout the year. Some samples from the same SUDI case clustered together and had a bootstrap value of 100 (HRV samples 4A and 10B and 1 and 7, and RSV samples 5 and 9).
The in Figure 1 tree was constructed using the Maximum Likelihood Method in Randomised Axelerated Maximum Likelihood (RAxML) program with 1000 bootstrap replicates with values over 70% shown. The General Time Reversible substitution model with gamma distributed rate variation and invariant sites (GTR + G + I). The tree was rooted with Coxsackievirus B1 (M16560) and Poliovirus 1 (V01149), and represent the outgroups in this phylogenetic analysis, as both viruses are closely related to the in-group (HRV) as per the ICTV website. Blue shading indicates HRV-A species, purple shading indicated HRV-B species, and pink shading indicates HRV-C species. The Samples successfully genotyped of each species are shaded in darker colour and are labelled with the respective genotype following the tip label, where appropriate.
The tree was constructed using the Maximum Likelihood Method in Randomised Axelerated Maximum Likelihood program with 1000 bootstrap replicates with values over 70% shown. The Hasegawa-Kishino-Yano model with gamma distributed rates (HKY + G) was used. The tree was rooted with Bovine orthopneumovirus (AF092942), and represents the outgroup selected for phylogenetic analysis. Blue shading indicates RSA-A genotype and purple shading indicated RSV-B genotype. The Samples successfully subtyped of each genotype are shaded in darker colour and are labelled with the respective subtyped following the tip label, where appropriate.
All RSV-A deduced amino acid sequences were aligned and compared to prototype ON1 strain (Accession number: JN257693; ON67-1210A), which is characterised by a nucleotide insertion of a 72-nucleotide duplication in the second hypervariable region of the RSV G-gene, otherwise a 23 amino acid duplication (QEETLHSTTSEGYLSPSQVYTTS) beginning at amino acid 261 to 283 and 285 to 307. None of the RSV-A samples presented with this duplication or matching N-glycosylation sites.
BA9 genotypes display a characteristic 60 nucleotide duplication, otherwise a duplication of 20 amino acids (TERDTSTSQSTVLDTTTSKH) beginning at position 240 to 259 and 260 to 279 of the RSV G-gene. Though sample 8 was closely related to LR699741—genotype BA9—and Samples 5 and 9 diverge from KY249662—also genotype BA9—as depicted in phylogenetic analysis following amino acid alignment of all RSV-B sequences obtained in this study to sequences of prototype BA1 (Accession number: AY333364; BA/4128/99B) and the inclusion of a BA9 subtype (Accession number: DQ227395; BA/100/04), only samples 9, and 5 displayed the insertion of 20 amino acid duplication, and amino acid substitutions K218T, L223P, K225I, V251T, D253N, S267P, H287Y, S288L, and E292G. All samples showed S247P, and four showed L286P substitutions. Additionally, outside of the second hypervariable region, all samples had I200T substitution. Two NST N-Glycosylation sites are reported within the RSV-B second hypervariable region of the G- gene, at codons 296 to 298 and 310 to 312. N-Glycosylation sites were determined by scores over 0.5. Sample 9 retained both N-Glycosylation sites, at codon 296 (0.52) and 310 (N-Glyc: 0.57). Sample 5 also retained both N-Glycosylation sites at codons 296 (0.51) and 310 (0.56). Sample 8 matched the first glycosylation site (N-Glyc: 0.59) with N296L substitution.
Deduced amino acid alignment of RSV G-gene of RSV-A and RSV-B samples are presented as Figure 3 and Figure 4. RSV-A sample sequences were aligned and are shown relative to the sequence of RSV ON1 (Accession number: JN257693; ON67-1210A), while RSV-B samples sequences were aligned and are shown relative to sequence of prototype RSV-BA1 (Accession number: AY333364; BA/4128/99B) and BA9 (Accession number: DQ227395; BA/100/04).
The amino acid numbers of the sequence ruler (in blue) within the orange braces correspond to the positions of 212 and 321 of the second hypervariable region of the G-gene of the RSV ON1 sequence. The red outlined rectangular boxes depict the 23 amino acid duplication (QEETLHSTTSEGYLSPSQVYTTS) of the 24 amino acid insertion, characteristic to RSV ON1. Dots represent matching amino acid to reference sequence (RSV ON1). Dashes represent missing amino acid. Grey shading depicts predicted N-glycosylation sites.
The amino acid numbers of the sequence ruler (in blue) within the orange braces correspond to the positions of 213 and 315 of the second hypervariable region of the G-gene of the RSV-BA1 sequence. The red outlined rectangular boxes depict the 20 amino acid duplication (TERDTSTSQSTVLDTTTSKH) insertion, characteristic to RSV-BA. Dots represent matching amino acid to reference sequence (RSV-BA1). Dashes represent missing amino acid. Grey shading depicts predicted N-glycosylation sites.
4. Discussion
This study used retrospective respiratory tract samples collected from SUDI cases admitted to the Tygerberg Medico-legal Mortuary in Cape Town to determine the different HRV and RSV genotypes. The phylogenetic findings and analysis of this study were influenced by successful retrieval, and amplification of previously identified PCR-positive HRV and/or RSV samples, successful confirmatory PCR testing in this study, followed by successful sequencing. The literature describes genotypic characterisation of HRV-A, -B, and -C, as well as RSV-A and -B in infants and children admitted for hospital care in South Africa [4,44,64]; however, this is the first study to genotype respiratory viruses in SUDI cases at Tygerberg Medico-legal Mortuary, and South Africa.
Sample 11C from the 2015/2016 study, was from a male infant, but the age was not captured in the SUDI database or case file. While the results from this sample closely related to that of a 9-month-old infant as per the GenBank record (Accession number: KF543881), the exact age of Infant 11C could not be confirmed. HRV was confirmed in a 3-month-old SUDI case (HRV-A10, Sample 6A; and HRV-A11, Samples 3A and 7B). The same HRV genotypes were confirmed in an infant presenting with manifestations of acute respiratory illness in the literature [65]. Although HRV-A10 is less frequently reported than HRV-A28 (which was present in Sample 11C), HRV-A56 (Samples 1, 7, and 12), and HRV-A80 (Samples 10B and 4A from the same SUDI case) the genotype is commonly detected in infants presenting with respiratory disease in other sub-Saharan Africa countries [25,66,67].
Genotypes HRV-A11 and HRV-A80 were also identified in some of the 2015/2016 SUDI samples. HRV-A80 (Samples 10B and 4A from the same SUDI case) was also detected in Tunis, Tunisia, between 2015 and 2017 and in Guangzhou, China between 2018 and 2019 in hospitalised infants presenting with severe and acute respiratory tract infection [68,69]. Certain HRV genotypes, such as HRV-A82 and HRV-A43, are not exclusively detected in infants, but also in adolescents and adults, who may transmit the virus to infants when in close contact [70], such as bed-sharing. Most infants in this study had reported a history of bed-sharing, in which the infant is in close contact with other individuals for prolonged periods of time. If such individuals present with HRV or any other viral infection, it may increase the risk of viral transmission.
During the COVID-19 pandemic the implemented “lockdown” forced individuals to spend prolonged hours within confined spaces at home and/or other areas. There was a reported increase in HRV infection in multiple countries, which may be the result of the high transmissibility of HRV. In particular, HRV-A82 (Samples 4 and 5) was commonly detected in children and adolescents presenting with respiratory disease during the COVID-19 pandemic [71,72]. Furthermore, publications have reported genotype HRV-A43 (which was also present in Sample 10), reinfecting children, adults and the elderly with acute respiratory infections, evidenced by prolonged viral shedding [73,74,75]. This evidence suggests that HRV may either play a contributory role in infant death, or the detection during post-mortem analysis may not infer causation of SUDI, but rather transmission from close-contact with other infected individuals. While the results of this study could not support or exclude this theory, or evaluated overcrowding in the infant population, a study from the Salt River Mortuary has identified the consequences of overcrowded living conditions and infection-related causes of death in SUDI cases [76]. Finally, subtypes HRV-A56 (Samples 1, 7, and 12), HRV-A28 (Sample 11C) and HRV-A80 (Samples 10B and 4A) were among the most frequently reported in clinical studies reporting on HRV diversity Asia and Africa between 2013 and 2021 [16]. Only Genotype HV-A63 (Samples 5C and 10C) has been detected in another SUDI case in the literature [77]. This may be attributed to the lack of SUDI investigations assessing viral genotypes in forensic settings in South Africa and globally. Due to limited resources and research capacity at the Tygerberg Medico-legal Mortuary, the sample collection periods of SUDI cases are intermittent and it is often not possible to identify other HRV genotypes to compare to the findings in the literature.
HRV-B was least often detected in this study, which is a reported trend by molecular epidemiology research in Tanzania, Nigeria, Kenya, Mozambique and South Africa [25,67,78,79,80]. This species is not associated with severe disease and parents may therefore not seek medical attention and laboratory testing, suggesting that the true burden may be underrepresented [78]. However, genotype HRV-B84 (Samples 13A and 16C) was one of the top 25 genotypes reported by the seven studies from Africa [16]. The low detection of HRV-B in this study might not be a true representation of its diversity in SUDI cases in this population due to the significant sample loss (43%) during confirmatory testing and limited sequencing success of previously collected HRV samples.
HRV-C is commonly detected in Cape Town, especially in children with respiratory disease [4]. Genotype C22 (Sample 1B) confirmed in samples from the 2015/2016 study was also detected in children in Nigeria and Tanzania suspected of respiratory illness, and hospitalised Swedish children [16,67,78]. HRV-C38 and HRV-C2 were identified in samples from the 2015/2016 and 2018/2019 studies respectively. Both genotypes were among the top three identified in Kenya from 2007 to 2018 in children presenting with SARI. C2 was previously detected in Amsterdam between 2007 and 2012 [16,81]. The burden of HRV infection in infants living in Africa and hospitalised in the Cape Town Metropole has previously been well described [4,42,67].
The findings of this study suggest that certain HRV genotypes were present in the SUDI cases at the time of death, but the studies to date were not designed to assess a causal relationship between PCR-results and infant death. The mechanisms by which infection may lead to death, especially in infants, may be supplemented by genotype identification in the presence of clinical features of respiratory disease to further elucidate the role of HRV genotypes in infant death [15,82,83].
Studies from, China and Saudi Arabia report an RSV-A predominance [84,85]. While RSV-A predominated in South Africa between 2017 and 2018, RSV-B predominated in eight other countries, possibly due to virulence and circulation of specific strains at the time [86]. However, RSV-A remains the most common and diverse group in South Africa [64,86,87,88]. Three samples collected in 2018/2019 closely related to other RSV-A isolates collected in South Africa in 2015 (Sample 2; Accession number: MN516963) and in 2018 (Samples 6 and 7; Accession number: OK299916). Despite a high prevalence of ON1 in South Africa between 2015 and 2017, none of the RSV-A samples from this study closely related to ON1. The reason could be two-fold. Either ON1 was truly absent or it could have been present in the previous PCR-positive samples that could not be reconfirmed with retesting.
RSV-BA was first identified in 1999 in Buenos Aires and first detected in South Africa in 2006 during a nosocomial outbreak [37,85,88,89]. One study sample (Sample 8), collected in 2015/2016 from a 1-month-old infant closely related to RSV-BA9 first isolated from a male infant in Spain as per the GenBank record. RSV-BA9 was first identified in Japan during the 2006 and 2007 epidemic season and is commonly detected in infants presenting with LRTI [84,90,91,92,93]. In 2021, RSV-BA9 was detected in an outbreak in Shenyang, China at a postpartum care centre in neonates which required subsequent hospitalisation and presented with respiratory failure [94]. RSV-BA9 has previously been detected in infants and other regions of South Africa, however recently published amino acid substitutions are limited [37,44,64,87,95]. The amino acid substitutions noted in RSV-BA9 this study were characteristics of RSV-BA9 strains also reported by other authors [9,92,96]. Substitution I281T and P291L differentiating between sub-genotypes RSV-BA9a and RSV-BA9b were not noted in this study [93]. Outside the RSV-B duplication site, Samples 5 and 9 had commonly reported RSV-BA9 K218T, L233P, and H287Y substitutions [98,99,100]. S247P, also a common substitution of RSV-BA9 strains, was present in all samples, while L223P was only noted in Samples 5 and 9. S247P substitution has been reported from Taiwan, Panama, Thailand, and other cities [92,100,101,102,103]. Furthermore, 247P with L223P and H287Y substitutions of Samples 5 and 9 were noted in strains identified in Central African Republic between 2015 and 2018, which was a similar sample collection period of both previous screening studies included in this analysis [104].
In the RSV-B duplication site, D253N amino acid substitution was reported in one strain during between in the 2017 to 2019 RSV seasonal epidemics in Shanghai, China and multiple studies in India from children presenting with respiratory disease from children presenting with LRTI [91,105,106]. Additionally, the substitution may lead to a possible third N-glycosylation site [107], but this was not present in the current study. Interestingly, E292G substitution in samples 5 and 9, were reported in RSV-BA10 detected in children admitted for acute respiratory infection in Vietnam and is confirmed by other publications in RSV-BA10 genotypes [108,109]. Although Sample 10 did not present with the amino acid duplication insertion, but it had T254I substitution, which was reported as a characteristic of RSV-BA9 [92]. In combination with mutation K314R, not noted in this study, the substitution may have played a role in BA9 predominance in 2015/2016 in Taiwan at the same time period the sample from this study was collected [100]. When RSV-B sequences were compared to DQ227395 (BA/100/04), A269V substitution of Samples 5 and 9 were noted and reported in strains identified in children in Shanghai, China during the COVID-19 pandemic [110].
5. Conclusions
The phylogenetic analysis of HRV and RSV in this SUDI population was severely constrained by the much lower than anticipated sample number and manual PCR amplification limit (determined by Ct value). Thus, the findings in this study do not extensively characterise the viral diversity of these viruses in SUDI cases.
This was the first study at the Tygerberg Medico-legal Mortuary to investigate specific viruses on genetic level and this forms the foundation to continue investigation to further identify and describe distribution patterns, severity of disease, and possible associations between the identified risk factors and genotypes for the most prevalent respiratory viruses in infants and children. Additionally, this study describes the first findings of viral genotypes in SUDI cases. The discussion is limited to other research which includes infants and children with respiratory disease and subsequently HRV and/or RSV infection was detected. While the findings of this study are incomparable to such research, the subtypes within the SUDI cases are linked to infants presenting with severe respiratory illness in paediatric cases, which in combination with anatomical, immune, and socio-demographic factors, may result in SUDI.
Author Contributions
Conceptualization, Protocol and Methodology, Institutional Review Application and Approval, Funding acquisition, Resources, Provided training and supervision, Project administration, Visualisation, Overseeing laboratory work, results, data analysis and interpretation, Writing—review, editing and submission of manuscript—C.D.B.; Methodology, Data analysis and interpretation, writing—original draft preparation, H.D.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the South African Medical Research Council [MRC] (Self-initiated research), Poliomyelitis Research Foundation [PRF] (Grant numbers 22/03, 17/05, 15/07, 12/06, 09/29 and Student Bursaries 21/47, 18/53, 18/33, 16/35, 15/53, 14/31, 12/42, 11/37,09/33); National Health Laboratory Service [NHLS] Research Grants (Grant numbers 94795, 94603, 94380, 94172, KNC 151).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of Stellenbosch University Health Research Ethics Committee [HREC] (Registration Number N12/02/007, first registration 07/03/2012, with annual renewal).
Informed Consent Statement
HREC approved a waiver of consent under Protocol N12/02/007. Consent is provided for by Section 3(a) of the Inquests Act (58 of 1959) and the Criminal Procedure Act (1977) as all SUDI cases must be subjected to a full post-mortem investigation according to the Act (58 of 1959), as it is regarded as unnatural deaths, and samples are collected to ascertain the cause(s) of death.
Data Availability Statement
No publicly archived datasets were analyzed or generated during the study. Datasets are available on request from the authors.
Acknowledgments
Prof Johan Dempers (deceased) for his involvement in earlier SUDI studies, Ms Mathilda Claassen for training and assistance with sequencing and analysis.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Geoghegan, S., Erviti, A., Caballero, M.T., Vallone, F., Zanone, S.M., Losada, J.V., Bianchi, A., Acosta, P.L., Talarico, L.B., Ferretti, A., Grimaldi, L.A., Sancilio, A., Dueñas, K., Sastre, G., Rodriguez, A., Ferrero, F., Barboza, E., Gago, G.F., Nocito, C., Flamenco, E., Perez, A.R., Rebec, B., Ferrola, F.M., Libser, R., Karron, R.A., Bergel, El., Polack, F.P. Mortality due to Respiratory Syncytial Virus. Burden and Risk Factors. Am J Respir Crit Care Med 2017, 195(1), 96–103. [CrossRef]
- Kotaniemi-Syrjänen, A., Vainionpää, R., Reijonen, T.M., Waris, M., Korhonen, K., Korppi, M. Rhinovirus-induced wheezing in infancy--the first sign of childhood asthma?. J Allergy Clin Immunol 2003, 111(1), 66–71. [CrossRef]
- Linsuwanon, P., Payungporn, S., Suwannakarn, K., Chieochansin, T., Theamboonlers, A., Poovorawan, Y. Complete coding sequence characterization and comparative analysis of the putative novel human rhinovirus (HRV) species C and B. Virology J 2003, 8, 5. [CrossRef]
- Smuts, H.E., Workman, L.J., Zar, H.J. Human rhinovirus infection in young African children with acute wheezing. BMC Infect Dis 2011, 11, 65. [CrossRef]
- Drysdale, S.B., Alcazar, M., Wilson, T., Smith, M., Zuckerman, M., Lauinger, I.L., Tong, C.Y., Broughton, S., Rafferty, G.F., Johnston, S.L., Greenough, A. Respiratory outcome of prematurely born infants following human rhinovirus A and C infections. Eur J Pediatr 2014, 173(7), 913–919. [CrossRef]
- Stone, C.A., Jr, Miller, E.K. Understanding the Association of Human Rhinovirus with Asthma. Clin Vaccine Immunol 2015, CVI, 23(1), 6–10. [CrossRef]
- Jartti, T., Gern, J.E. Role of viral infections in the development and exacerbation of asthma in children. J Allergy Clinical Immunol 2017, 140(4), 895–906. [CrossRef]
- Shi, T., McAllister, D.A., O’Brien, K.L., Simoes, E.A.F., Madhi, S.A., Gessner, B.D., Polack, F.P., Balsells, E., Acacio, S., Aguayo, C., Alassani, I., Ali, A., Antonio, M., Awasthi, S., Awori, J.O., Azziz-Baumgartner, E., Baggett, H.C., Baillie, V.L., Balmaseda, A., Barahona, A., Basnet, S., Bassat, Q., Basualdo, W., Bigogo, G., Bont, L., Breiman, R.F., Brooks, W.A., Broor, S., Bruce, N., Bruden, D., Buchy, P., Campbell, S., Carosone-Link, P., Chadha, M., Chipeta, J., Chou, M., Clara, W., Cohen, C., de Cuellar, E., Dang, D.A., Dash-Yandag, B., Deloria-Knoll, M., Dherani, M., Eap, T., Ebruke, B.E., Echavarria, M., de Freitas Lázaro Emediato, C.C., Fasce, R.A., Feikin, D.R., Feng, L., Gentile, A., Gordon, A., Goswami, D., Goyet, S., Groome, M., Halasa, N., Hirve, S., Homaira, N., Howie, S.R.C., Jara, J., Jroundi, I., Kartasasmita, C.B., Khuri-Bulos, N., Kotloff, K.L., Krishnan, A., Libster, R., Lopez, O., Lucero, M.G., Lucion, F., Lupisan, S.P., Marcone, D.N., McCracken, J.P., Mejia, M., Moisi, J.C., Montgomery, J.M., Moore, D.P., Moraleda, C., Moyes, J., Munywoki, P., Mutyara, K., Nicol, M.P., Nokes, D.J., Nymadawa, P., da Costa Oliveira, M.T., Oshitani, H., Pandey, N., Paranhos-Baccalà, G., Phillips, L.N., Picot, V.S., Rahman, M., Rakoto-Andrianarivelo, M., Rasmussen, Z.A., Rath, B.A., Robinson, A., Romero, C., Russomando, G., Salimi, V., Sawatwong, P., Scheltema, N., Schweiger, B., Scott, J.A.G., Seidenberg, P., Shen, K., Singleton, R., Sotomayor, V., Strand, T.A., Sutanto, A., Sylla, M., Tapia, M.D., Thamthitiwat, S., Thomas, E.D., Tokarz, R., Turner, C., Venter, M., Waicharoen, S., Wang, J., Watthanaworawit, W., Yoshida, L.M., Yu, H., Zar, H.J., Campbell, H., Nair, H.; RSV Global Epidemiology Network. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: a systematic review and modelling study. Lancet (London, England) 2017, 390(10098), 946–958. [CrossRef]
- Borchers, A.T., Chang, C., Gershwin, M.E., Gershwin, L.J. Respiratory syncytial virus--a comprehensive review. Clin Rev Allergy Immunol 2013, 45(3), 331–379. [CrossRef]
- Laudanno, S.L., Sánchez Yanotti, C.I., Polack, F.P. RSV Lower Respiratory Tract Illness in Infants of Low- and Middle-income Countries. Acta Med Acad 2020, 49(2), 191–197. [CrossRef]
- Zar, H.J., Nduru, P., Stadler, J.A.M., Gray, D., Barnett, W., Lesosky, M., Myer, L., Nicol, M.P. Early-life respiratory syncytial virus lower respiratory tract infection in a South African birth cohort: epidemiology and effect on lung health. Lancet Glob Health 2020, 8(10), e1316–e1325. [CrossRef]
- Jacobs, S.E., Lamson, D.M., St George, K., Walsh, T.J. Human rhinoviruses. Clin Microbiol Rev 2013, 26(1), 135–162. [CrossRef]
- Nakauchi, M., Nagata, N., Takayama, I., Saito, S., Kubo, H., Kaida, A., Oba, K., Odagiri, T., Kageyama, T. Propagation of Rhinovirus C in Differentiated Immortalized Human Airway HBEC3-KT Epithelial Cells. Viruses 2019, 11(3), 216. [CrossRef]
- Ashraf, S., Brockman-Schneider, R., Bochkov, Y.A., Pasic, T.R., Gern, J.E. Biological characteristics and propagation of human rhinovirus-C in differentiated sinus epithelial cells. Virology 2013, 436(1), 143–149. [CrossRef]
- Gomez-Perosanz, M., Sanchez-Trincado, J.L., Fernandez-Arquero, M., Sidney, J., Sette, A., Lafuente, E.M., Reche, P.A. Human rhinovirus-specific CD8 T cell responses target conserved and unusual epitopes. FASEB J 2021, 35(1), e21208. [CrossRef]
- Esneau, C., Duff, A.C., Bartlett, N.W. Understanding Rhinovirus Circulation and Impact on Illness. Viruses 2022, 14(1), 141. [CrossRef]
- Clementi, N., Ghosh, S., De Santis, M., Castelli, M., Criscuolo, E., Zanoni, I., Clementi, M., Mancini, N. Viral Respiratory Pathogens and Lung Injury. Clin Microbiol Rev 2021, 34(3), e00103-20. [CrossRef]
- Han, M., Rajput, C., Hershenson, M.B. Rhinovirus Attributes that Contribute to Asthma Development. Immunol Allergy Clin North Am 2019, 39(3), 345–359. [CrossRef]
- Makris, S., Johnston, S. Recent advances in understanding rhinovirus immunity. F1000Res 2018, 7, F1000 Faculty Rev-1537. [CrossRef]
- Santti, J., Hyypiä, T., Kinnunen, L., Salminen, M. Evidence of recombination among enteroviruses. J Virol 1999, 73(10), 8741–8749. [CrossRef]
- Savolainen, C., Laine, P., Mulders, M.N., Hovi, T. Sequence analysis of human rhinoviruses in the RNA-dependent RNA polymerase coding region reveals large within-species variation. J Gen Virol 2004, 85(Pt 8), 2271–2277. [CrossRef]
- Arden, K.E., McErlean, P., Nissen, M.D., Sloots, T.P., Mackay, I.M. Frequent detection of human rhinoviruses, paramyxoviruses, coronaviruses, and bocavirus during acute respiratory tract infections. J Med Virol 2006, 78(9), 1232–1240. [CrossRef]
- Ren, L., Yang, D., Ren, X., Li, M., Mu, X., Wang, Q., Cao, J., Hu, K., Yan, C., Fan, H., Li, X., Chen, Y., Wang, R., An, F., An, S., Luo, M., Wang, Y., Xiao, Y., Xiang, Z., Xiao, Y., Li, L., Huang, F., Gao, Z., Wang, J. Genotyping of human rhinovirus in adult patients with acute respiratory infections identified predominant infections of genotype A21. Sci Rep 2017, 7, 41601. [CrossRef]
- Zhao, Y., Shen, J., Wu, B., Liu, G., Lu, R., Tan, W. Genotypic Diversity and Epidemiology of Human Rhinovirus Among Children With Severe Acute Respiratory Tract Infection in Shanghai, 2013-2015. Front Microbiol 2018, 9, 1836. [CrossRef]
- Baillie, V.L., Moore, D.P., Mathunjwa, A., Morailane, P., Simões, E.A.F., Madhi, S.A. Molecular Subtyping of Human Rhinovirus in Children from Three Sub-Saharan African Countries. J Clin Microbiol 2019, 57(9), e00723-19. [CrossRef]
- Luka, M.M., Kamau, E., Adema, I., Munywoki, P.K., Otieno, G.P., Gicheru, E., Gichuki, A., Kibinge, N., Agoti, C.N., Nokes, D.J. Molecular Epidemiology of Human Rhinovirus From 1-Year Surveillance Within a School Setting in Rural Coastal Kenya. Open Forum Infect Dis 2020, 7(10), ofaa385. [CrossRef]
- da Costa Souza, L., Bello, E.J.M., Dos Santos, E.M., Nagata, T. Molecular and clinical characteristics related to rhinovirus infection in Brasília, Brazil. Braz J Microbiol 2021, 52(1), 289–298. [CrossRef]
- Pandya, M.C., Callahan, S.M., Savchenko, K.G., Stobart, C.C. A Contemporary View of Respiratory Syncytial Virus (RSV) Biology and Strain-Specific Differences. Pathogens 2019, 8(2), 67. [CrossRef]
- Garcia-Garcia, M.L., Calvo Rey, C., Del Rosal Rabes, T. Pediatric Asthma and Viral Infection. Asma y virus en el niño. Arch Bronconeumol 2016, 52(5), 269–273. [CrossRef]
- Karron, R.A., Zar, H.J. Determining the outcomes of interventions to prevent respiratory syncytial virus disease in children: what to measure?. Lancet Respir Med 2018, 6(1), 65–74. [CrossRef]
- Battles, M.B., McLellan, J.S. Respiratory syncytial virus entry and how to block it. Nat Rev Microbiol 2019, 17(4), 233–245. [CrossRef]
- Ruckwardt, T.J., Morabito, K.M., Graham, B.S. Immunological Lessons from Respiratory Syncytial Virus Vaccine Development. Immunity 2019, 51(3), 429–442. [CrossRef]
- Driscoll, A.J., Arshad, S.H., Bont, L., Brunwasser, S.M., Cherian, T., Englund, J.A., Fell, D.B., Hammitt, L.L., Hartert, T.V., Innis, B.L., Karron, R.A., Langley, G.E., Mulholland, E.K., Munywoki, P.K., Nair, H., Ortiz, J.R., Savitz, D.A., Scheltema, N.M., Simões, E.A.F., Smith, P.G., Were, F., Zar, H.J., Feikin, D.R. Does respiratory syncytial virus lower respiratory illness in early life cause recurrent wheeze of early childhood and asthma? Critical review of the evidence and guidance for future studies from a World Health Organization-sponsored meeting. Vaccine 2020, 38(11), 2435–2448. [CrossRef]
- Bergeron, H.C., Tripp, R.A. Immunopathology of RSV: An Updated Review. Viruses 2021, 13(12), 2478. [CrossRef]
- Malinczak, C.A., Lukacs, N.W., Fonseca, W. Early-Life Respiratory Syncytial Virus Infection, Trained Immunity and Subsequent Pulmonary Diseases. Viruses 2020, 12(5), 505. [CrossRef]
- Nam, H.H., Ison, M.G. Respiratory syncytial virus infection in adults. Br Med J 2019, 366, l5021. [CrossRef]
- Liu, H., Lu, B., Tabor, D.E., Tovchigrechko, A., Wilkins, D., Jin, H., Madhi, S.A., Soofie, N., Esser, M.T., Nunes, M.C. Characterization of human respiratory syncytial virus (RSV) isolated from HIV-exposed-uninfected and HIV-unexposed infants in South Africa during 2015-2017. Influenza Other Respir Viruses 2020, 14(4), 403–411. [CrossRef]
- Bin Lu, Liu, H., Tabor, D.E., Tovchigrechko, A., Qi, Y., Ruzin, A., Esser, M.T., Jin, H. Emergence of new antigenic epitopes in the glycoproteins of human respiratory syncytial virus collected from a US surveillance study, 2015-17. Sci Rep 2019, 9(1), 3898. [CrossRef]
- Yun, K.W., Choi, E.H., Lee, H.J. Molecular epidemiology of respiratory syncytial virus for 28 consecutive seasons (1990-2018) and genetic variability of the duplication region in the G gene of genotypes ON1 and BA in South Korea. Arch Virol 2020, 165(5), 1069–1077. [CrossRef]
- Lau, S.K., Yip, C.C., Lin, A.W., Lee, R.A., So, L.Y., Lau, Y.L., Chan, K.H., Woo, P.C., Yuen, K.Y. Clinical and molecular epidemiology of human rhinovirus C in children and adults in Hong Kong reveals a possible distinct human rhinovirus C subgroup. J Infect Dis 2009, 200(7), 1096–1103. [CrossRef]
- Miller, E.K., Khuri-Bulos, N., Williams, J.V., Shehabi, A.A., Faouri, S., Al Jundi, I., Chen, Q., Heil, L., Mohamed, Y., Morin, L.L., Ali, A., Halasa, N.B. Human rhinovirus C associated with wheezing in hospitalised children in the Middle East. J Clin Virol 2009, 46(1), 85–89. [CrossRef]
- Pretorius, M.A., Tempia, S., Treurnicht, F.K., Walaza, S., Cohen, A.L., Moyes, J., Hellferscee, O., Variava, E., Dawood, H., Chhagan, M., Haffjee, S., Madhi, S.A., Cohen, C., Venter, M. Genetic diversity and molecular epidemiology of human rhinoviruses in South Africa. Influenza Other Respir Viruses 2014, 8(5), 567–573. [CrossRef]
- Hung, H.M., Yang, S.L., Chen, C.J., Chiu, C.H., Kuo, C.Y., Huang, K.A., Lin, T.Y., Hsieh, Y.C., Gong, Y. N., Tsao, K.C., Huang, Y.C. Molecular epidemiology and clinical features of rhinovirus infections among hospitalized patients in a medical center in Taiwan. J Microbiol Immunol Infect 2019, 52(2), 233–241. [CrossRef]
- Venter, M., Madhi, S.A., Tiemessen, C.T., Schoub, B.D. Genetic diversity and molecular epidemiology of respiratory syncytial virus over four consecutive seasons in South Africa: identification of new subgroup A and B genotypes. J Gen Virol 2001, 82(Pt 9), 2117–2124. [CrossRef]
- Robertson, S.E., Roca, A., Alonso, P., Simoes, E.A., Kartasasmita, C.B., Olaleye, D.O., Odaibo, G.N., Collinson, M., Venter, M., Zhu, Y., Wright, P.F. Respiratory syncytial virus infection: denominator-based studies in Indonesia, Mozambique, Nigeria and South Africa. Bull World Health Organ 2004, 82(12), 914–922.
- McMorrow, M.L., Tempia, S., Walaza, S., Treurnicht, F.K., Moyes, J., Cohen, A.L., Pretorius, M., Hellferscee, O., Wolter, N., von Gottberg, A., Nguweneza, A., McAnerney, J.M., Naby, F., Mekgoe, O., Venter, M., Madhi, S.A., Cohen, C. The Role of Human Immunodeficiency Virus in Influenza- and Respiratory Syncytial Virus-associated Hospitalizations in South African Children, 2011-2016. Clin Infect Dis 2019, 68(5), 773–780. [CrossRef]
- Xu, L., Gao, H., Zeng, J., Liu, J., Lu, C., Guan, X., Qian, S., Xie, Z. A fatal case associated with respiratory syncytial virus infection in a young child. BMC Infect Dis 2018, 18(1), 217. [CrossRef]
- Kubale, J., Kuan, G., Gresh, L., Ojeda, S., Azziz-Baumgartner, E., Sanchez, N., Lopez, R., Harris, E., Balmaseda, A., Gordon, A. Assessing the Incidence of Symptomatic Respiratory Syncytial Virus Illness Within a Prospective Birth Cohort in Managua, Nicaragua. Clin Infect Dis 2020, 70(10), 2029–2035. [CrossRef]
- Winterbach M, Hattingh C, Heathfield, LJ. Retrospective study of sudden unexpected death of infants in the Garden Route and Central Karoo districts of South Africa: Causes of death and epidemiological factors. S Afr J Child Health 2021, 15(2):74-82. [CrossRef]
- la Grange, H. Respiratory Pathogens in Cases of Sudden Unexpected Death in Infancy (SUDI) at Tygerberg Forensic Pathology Service Mortuary. Master of Science thesis, Stellenbosch University, Cape Town; 2013.
- Luoto, R., Jartti, T., Ruuskanen, O., Waris, M., Lehtonen, L., Heikkinen, T. Review of the clinical significance of respiratory virus infections in newborn infants. Acta Paediatr 2016, 105(10), 1132–1139. [CrossRef]
- Toizumi, M., Suzuki, M., Nguyen, H.A.T., Le, M.N., Ariyoshi, K., Moriuchi, H., Hashizume, M., Dang, D.A., Yoshida, L.M. Viral Acute Respiratory Illnesses in Young Infants Increase the Risk of Respiratory Readmission. Pediatr Infect Dis 2018, 37(12), 1217–1222. [CrossRef]
- Boonyaratanakornkit, J., Englund, J.A., Magaret, A.S., Bu, Y., Tielsch, J.M., Khatry, S.K., Katz, J., Kuypers, J., Shrestha, L., LeClerq, S.C., Steinhoff, M.C., Chu, H.Y. Primary and Repeated Respiratory Viral Infections Among Infants in Rural Nepal. J Pediatr Infect Dis Soc 2020, 9(1), 21–29. [CrossRef]
- Dempers, J.J., Burger, E.H., Toit-Prinsloo, L.D., Verster, J. A South African Perspective. In: SIDS Sudden Infant and Early Childhood Death: The Past, the Present and the Future. Duncan, J.R., Byard, R.W., Eds. University of Adelaide Press: Adelaide, Australia, 2018. Chapter 17. PMID: 30035954.
- Garstang, J., Watson, D., Pease, A., Ellis, C., Blair, P.S., Fleming, P. Improving engagement with services to prevent Sudden Unexpected Death in Infancy (SUDI) in families with children at risk of significant harm: A systematic review of evidence. Child Care Health Dev 2021, 47(5), 713–731. [CrossRef]
- Weber, M.A., Ashworth, M.T., Risdon, R.A., Hartley, J.C., Malone, M., Sebire, N.J. The role of post-mortem investigations in determining the cause of sudden unexpected death in infancy. Arch Dis Child 2008, 93(12), 1048–1053. [CrossRef]
- la Grange, H., Verster, J., Dempers, J.J., de Beer, C. Review of immunological and virological aspects as contributory factors in Sudden Unexpected Death in Infancy (SUDI). For Sci Int 2014, 245, 12–16. [CrossRef]
- Dempers, J.J., Coldrey, J., Burger, E.H., Thompson, V., Wadee, S.A., Odendaal, H.J., Sens, M.A., Randall, B.B., Folkerth, R.D., Kinney, H.C., PASS Network. The Institution of a Standardized Investigation Protocol for Sudden Infant Death in the Eastern Metropole, Cape Town, South Africa. J Forensic Sci 2016, 61(6), 1508–1514. [CrossRef]
- du Toit-Prinsloo, L., Dempers, J., Verster, J., Hattingh, C., Nel, H., Brandt, V.D., Jordaan, J., Saayman, G. Toward a standardized investigation protocol in sudden unexpected deaths in infancy in South Africa: a multicenter study of medico-legal investigation procedures and outcomes. Forensic Sci Med Pathol 2013, 9(3), 344–350. [CrossRef]
- Osei-Poku, G.K., Thomas, S., Mwananyanda, L., Lapidot, R., Elliott, P.A., Macleod, W.B., Somwe, S.W., Gill, C.J. A systematic review of the burden and risk factors of sudden infant death syndrome (SIDS) in Africa. J Glob Health 2021, 11, 04075. [CrossRef]
- Shipstone, R.A., Young, J., Kearney, L., Thompson, J.M.D. Applying a Social Exclusion Framework to Explore the Relationship Between Sudden Unexpected Deaths in Infancy (SUDI) and Social Vulnerability. Front Public Health 2020, 8, 563573. [CrossRef]
- Coiras, M.T., Aguilar, J.C., García, M.L., Casas, I., Pérez-Breña, P. Simultaneous detection of fourteen respiratory viruses in clinical specimens by two multiplex reverse transcription nested-PCR assays. J Med Virol 2004, 72(3), 484–495. [CrossRef]
- Slovic, A., Ivancic-Jelecki, J., Ljubin-Sternak, S., Galinović, G.M., Forcic, D. A molecular epidemiological study of human respiratory syncytial virus in Croatia, 2011-2014. Infect Genet Evol 2016, 44, 76–84. [CrossRef]
- Venter, M., Collinson, M., Schoub, B.D. Molecular epidemiological analysis of community circulating respiratory syncytial virus in rural South Africa: Comparison of viruses and genotypes responsible for different disease manifestations. J Med Virol 2002, 68(3), 452–461. [CrossRef]
- Fawkner-Corbett, D.W., Khoo, S.K., Duarte, C.M., Bezerra, P.G., Bochkov, Y.A., Gern, J.E., Le Souef, P.N., McNamara, P.S. Rhinovirus-C detection in children presenting with acute respiratory infection to hospital in Brazil. J Med Virol 2016, 88(1), 58–63. [CrossRef]
- Daleno, C., Piralla, A., Scala, A., Senatore, L., Principi, N., Esposito, S. Phylogenetic analysis of human rhinovirus isolates collected from otherwise healthy children with community-acquired pneumonia during five successive years. PloS One 2013, 8(11), e80614. [CrossRef]
- Oluwasemowo, O.O., Nejo, Y.T., Abokede, J.O., Lawson, M., Motayo, B.O. Genotypes of rhinovirus detected among children in two communities of South-West Nigeria. Virus genes 2021, 57(3), 276–279. [CrossRef]
- Haddad-Boubaker, S., Mefteh, K., Mejri, C., Bouaffsoun, A., El Moussi, A., Boutiba, I., Mnif, K., Slim, A., Kechrid, A., Smaoui, H. High genotypic diversity of Rhinoviruses obtained from Tunisian children with severe acute respiratory infection. J Infect Dev Ctries 2021, 15(5), 726–735. [CrossRef]
- Li, W., Yu, B., Zhou, J., Wang, Y., Xue, B., Pan, J., Ran, Y., Yang, X., Wang, X., Yang, F., Li, H. Genetic diversity and epidemiology of human rhinovirus among children with severe acute respiratory tract infection in Guangzhou, China. Virol J 2021, 18(1), 174. [CrossRef]
- Panda, S., Mohakud, N.K., Panda, S., Kumar, S. Epidemiology and phylogenetic analysis of human rhinovirus/Enterovirus in Odisha, Eastern India. Indian J Med Microbiol 2019, 37(4), 569–573. [CrossRef]
- Peltola, V., Waris, M., Osterback, R., Susi, P., Ruuskanen, O., Hyypiä, T. Rhinovirus transmission within families with children: incidence of symptomatic and asymptomatic infections. J Infect Dis 2008, 197(3), 382–389. [CrossRef]
- Jia, R., Lu, L., Li, S., Liu, P., Xu, M., Cao, L., Su, L., Xu, J. Human rhinoviruses prevailed among children in the setting of wearing face masks in Shanghai, 2020. BMC Infect Dis 2022, 22(1), 253. [CrossRef]
- Zlateva, K.T., de Vries, J.J., Coenjaerts, F.E., van Loon, A.M., Verheij, T., Little, P., Butler, C.C., Goossens, H., Ieven, M., Claas, E.C., GRACE Study Group. Prolonged shedding of rhinovirus and re-infection in adults with respiratory tract illness. Eur Respir J 2014, 44(1), 169–177. [CrossRef]
- Prasetyo, A.A., Desyardi, M.N., Tanamas, J., Suradi, Reviono, Harsini, Kageyama, S., Chikumi, H., Shimizu, E. Respiratory viruses and torque teno virus in adults with acute respiratory infections. Intervirology 2015, 58(1), 57–68. [CrossRef]
- Kamau, E., Onyango, C.O., Otieno, G.P., Kiyuka, P.K., Agoti, C.N., Medley, G.F., Cane, P.A., Nokes, D.J., Munywoki, P.K. An Intensive, Active Surveillance Reveals Continuous Invasion and High Diversity of Rhinovirus in Households. J Infect Dis 2019, 219(7), 1049–1057. [CrossRef]
- Heathfield, L., Martin, L., Ramesar, R. A 5-year retrospective analysis of infant death at Salt River Mortuary, Cape Town. S Afr J Child Health 2020, 14(3), 148-154. [CrossRef]
- Baker, T., Schandl, C., Presnell, S.E., Madory, J., Nolte, F.S., Batalis, N. Use of an Automated Nested Multiplex Respiratory Pathogen PCR Panel Postmortem in the Pediatric Forensic Setting. J For Sci 2017, 62(5), 1223–1228. [CrossRef]
- L’Huillier, A.G., Kaiser, L., Petty, T.J., Kilowoko, M., Kyungu, E., Hongoa, P., Vieille, G., Turin, L., Genton, B., D’Acremont, V., Tapparel, C. Molecular Epidemiology of Human Rhinoviruses and Enteroviruses Highlights Their Diversity in Sub-Saharan Africa. Viruses 2015, 7(12), 6412–6423. [CrossRef]
- Annamalay, A.A., Lanaspa, M., Khoo, S.K., Madrid, L., Acácio, S., Zhang, G., Laing, I.A., Gern, J., Goldblatt, J., Bizzintino, J., Lehmann, D., Le Souëf, P.N., Bassat, Q. Rhinovirus species and clinical features in children hospitalised with pneumonia from Mozambique. Trop Med Int Health 2016, 21(9), 1171–1180. [CrossRef]
- Milanoi, S., Ongus, J.R., Gachara, G., Coldren, R., Bulimo, W. Serotype and genetic diversity of human rhinovirus strains that circulated in Kenya in 2008. Influenza Other Respir Viruses 2016, 10(3), 185–191. [CrossRef]
- Morobe, J.M., Nyiro, J.U., Brand, S., Kamau, E., Gicheru, E., Eyase, F., Otieno, G.P., Munywoki, P.K., Agoti, C.N., Nokes, D.J. Human rhinovirus spatial-temporal epidemiology in rural coastal Kenya, 2015-2016, observed through outpatient surveillance. Wellcome Open Res 2019, 3, 128. [CrossRef]
- Basnet, S., Palmenberg, A.C., Gern, J.E. Rhinoviruses and Their Receptors. Chest 2019, 155(5), 1018–1025. [CrossRef]
- Lamborn, I.T., Su, H.C. Genetic determinants of host immunity against human rhinovirus infections. Hum Genet 2020, 139(6-7), 949–959. [CrossRef]
- Song, J., Wang, H., Shi, J., Cui, A., Huang, Y., Sun, L., Xiang, X., Ma, C., Yu, P., Yang, Z., Li, Q., Ng, T.I., Zhang, Y., Zhang, R., Xu, W. Emergence of BA9 genotype of human respiratory syncytial virus subgroup B in China from 2006 to 2014. Sci Rep 2017, 7(1), 16765. [CrossRef]
- A Al-Sharif, H., El-Kafrawy, S.A., Yousef, J.M., Kumosani, T.A., Kamal, M.A., Khathlan, N.A., Kaki, R.M., Alnajjar, A.A., Azhar, E.I. Dominance of the ON1 Genotype of RSV-A and BA9 Genotype of RSV-B in Respiratory Cases from Jeddah, Saudi Arabia. Genes 2020, 11(11), 1323. [CrossRef]
- Tabor, D.E., Fernandes, F., Langedijk, A.C., Wilkins, D., Lebbink, R.J., Tovchigrechko, A., Ruzin, A., Kragten-Tabatabaie, L., Jin, H., Esser, M.T., Bont, L.J., Abram, M.E., INFORM-RSV Study Group. Global Molecular Epidemiology of Respiratory Syncytial Virus from the 2017-2018 INFORM-RSV Study. J Clin Microbiol 2020, 59(1), e01828-20. [CrossRef]
- Madhi, S.A., Venter, M., Alexandra, R., Lewis, H., Kara, Y., Karshagen, W.F., Greef, M., Lassen, C. Respiratory syncytial virus associated illness in high-risk children and national characterisation of the circulating virus genotype in South Africa. J Clin Virol 2003, 27(2), 180–189. [CrossRef]
- van Niekerk, S., Venter, M. Replacement of previously circulating respiratory syncytial virus subtype B strains with the BA genotype in South Africa. J Virol 2011, 85(17), 8789–8797. [CrossRef]
- Ihling, C.M., Schnitzler, P., Heinrich, N., Mangu, C., Sudi, L., Souares, A., Gies, S., Sié, A., Coulibaly, B., Ouédraogo, A.T., Mordmüller, B., Held, J., Adegnika, A.A., Fernandes, J.F., Eckerle, I., May, J., Hogan, B., Eibach, D., Tabatabai, J. Molecular epidemiology of respiratory syncytial virus in children in sub-Saharan Africa. Trop Med Int Health 2021, 26(7), 810–822. [CrossRef]
- Pangesti, K.N.A., Abd El Ghany, M., Walsh, M.G., Kesson, A.M., Hill-Cawthorne, G.A. Molecular epidemiology of respiratory syncytial virus. Rev Med Virol 2018, 28(2), 10.1002/rmv.1968. [CrossRef]
- Kang, H.M., Park, K.C., Park, J., Kil, H.R., Yang, E.A. Circulating Respiratory Syncytial Virus Genotypes and Genetic Variability of the G Gene during 2017 and 2018/2019 Seasonal Epidemics Isolated from Children with Lower Respiratory Tract Infections in Daejeon, Korea. Journal of Korean medical science 2020, 35(49), e422. [CrossRef]
- Korsun, N., Angelova, S., Trifonova, I., Voleva, S., Grigorova, I., Tzotcheva, I., Mileva, S., Alexiev, I., Perenovska, P. Predominance of ON1 and BA9 genotypes of respiratory syncytial virus (RSV) in Bulgaria, 2016-2018. J Med Virol 2021, 93(6), 3401–3411. [CrossRef]
- Razanajatovo Rahombanjanahary, N.H., Rybkina, K., Randriambolamanantsoa, T.H., Razafimanjato, H., Heraud, J.M. Genetic diversity and molecular epidemiology of respiratory syncytial virus circulated in Antananarivo, Madagascar, from 2011 to 2017: Predominance of ON1 and BA9 genotypes. J Clin Virol 2020, 129, 104506. [CrossRef]
- Wang, B., Song, J., Song, J., Mao, N., Liang, J., Chen, Y., Qi, Y., Bai, L., Xie, Z., Zhang, Y. An Outbreak of Severe Neonatal Pneumonia Caused by Human Respiratory Syncytial Virus BA9 in a Postpartum Care Centre in Shenyang, China. Microbiol Spectr 2022, 10(4), e0097422. [CrossRef]
- Valley-Omar, Z., Muloiwa, R., Hu, N.C., Eley, B., Hsiao, N.Y. Novel respiratory syncytial virus subtype ON1 among children, Cape Town, South Africa, 2012. Emerg Infect Dis 2013, 19(4), 668–670. [CrossRef]
- Tabatabai, J., Prifert, C., Pfeil, J., Grulich-Henn, J., Schnitzler, P. Novel respiratory syncytial virus (RSV) genotype ON1 predominates in Germany during winter season 2012-13. PloS One 2014, 9(10), e109191. [CrossRef]
- Tavakoli, F., Izadi, A., Yavarian, J., Sharifi-Zarchi, A., Salimi, V., Mokhtari-Azad, T. Determination of genetic characterization and circulation pattern of Respiratory Syncytial Virus (RSV) in children with a respiratory infection, Tehran, Iran, during 2018-2019. Virus Res 2021, 305, 198564. [CrossRef]
- Kenmoe, S., Vernet, M.A., Miszczak, F., Dina, J., Schoenhals, M., Beng, V.P., Vabret, A., Njouom, R. Genetic diversity of human respiratory syncytial virus isolated among children with acute respiratory infections in Southern Cameroon during three consecutive epidemic seasons, 2011-2013. Trop Med Health 2018, 46, 7. [CrossRef]
- Luo, H.J., Huang, X.B., Zhong, H.L., Ye, C.X., Tan, X., Zhou, K., Yuan, L., Zhang, S.F., Zhu, X., Lin, C.J., Wang, W.J., Xu, L., Cao, K.Y. Epidemiological characteristics and phylogenic analysis of human respiratory syncytial virus in patients with respiratory infections during 2011-2016 in southern China. Int J Infect Dis 2020, 90, 5–17. [CrossRef]
- Lee, C.Y., Fang, Y.P., Wang, L.C., Chou, T.Y., Liu, H.F. Genetic Diversity and Molecular Epidemiology of Circulating Respiratory Syncytial Virus in Central Taiwan, 2008-2017. Viruses 2021, 14(1), 32. [CrossRef]
- Auksornkitti, V., Kamprasert, N., Thongkomplew, S., Suwannakarn, K., Theamboonlers, A., Samransamruajkij, R., Poovorawan, Y. Molecular characterization of human respiratory syncytial virus, 2010-2011: identification of genotype ON1 and a new subgroup B genotype in Thailand. Arch Virol 2014, 159(3), 499–507. [CrossRef]
- Etemadi, M.R., Sekawi, Z., Othman, N., Lye, M.S., Moghaddam, F.Y. Circulation of human respiratory syncytial virus strains among hospitalized children with acute lower respiratory infection in malaysia. Evol Bioinform Online 2013, 9, 151–161. [CrossRef]
- Ábrego, L.E., Delfraro, A., Franco, D., Castillo, J., Castillo, M., Moreno, B., López-Vergès, S., Pascale, J. M., Arbiza, J. Genetic variability of human respiratory syncytial virus group B in Panama reveals a novel genotype BA14. J Med Virol 2017, 89(10), 1734–1742. [CrossRef]
- Komoyo, G.F., Yambiyo, B.M., Manirakiza, A., Gody, J.C., Muller, C.P., Hübschen, J.M., Nakoune, E., Snoeck, C.J. Epidemiology and genetic characterization of respiratory syncytial virus in children with acute respiratory infections: Findings from the influenza sentinel surveillance network in Central African Republic, 2015 to 2018. Health Sci Rep 2021, 4(2), e298. [CrossRef]
- Raghuram S,V., Khan, W.H., Deeba, F., Sullender, W., Broor, S., Parveen, S. Retrospective phylogenetic analysis of circulating BA genotype of human respiratory syncytial virus with 60 bp duplication from New Delhi, India during 2007-2010. Virusdisease 2015, 26(4), 276–281. [CrossRef]
- Bandla, S.S., Devadiga, S., Bhatt, R., Dsa, O.C., Govindakarnavar, A. Molecular epidemiology of respiratory syncytial virus among children and adults in India 2016 to 2018. Virus Genes 2021, 57(6), 489–501. [CrossRef]
- Hindupur, A., Menon, T., Dhandapani, P. Genetic diversity of human respiratory syncytial virus in children with acute respiratory infections in Chennai, South India. Ind J Med Microbiol 2019, 37(2), 248–254. [CrossRef]
- Do, L.A.H., Wilm, A., van Doorn, H.R., Lam, H.M., Sim, S., Sukumaran, R., Tran, A.T., Nguyen, B.H., Tran, T.T.L., Tran, Q.H., Vo, Q.B., Dac, N.A.T., Trinh, H.N., Nguyen, T.T.H., Binh, B.T.L., Le, K., Nguyen, M.T., Thai, Q.T., Vo, T.V., Ngo, N.Q.M., Dang, T.K.H., Cao, N.H., Tran, T.V., Ho, L.V., Farrar, J., de Jong, M., Chen, S., Nagarajan, N., Bryant, J.E., Hibberd, M.L. Direct whole-genome deep-sequencing of human respiratory syncytial virus A and B from Vietnamese children identifies distinct patterns of inter- and intra-host evolution. J Gen Virol 2015, 96(12), 3470–3483. [CrossRef]
- Thongpan, I., Mauleekoonphairoj, J., Vichiwattana, P., Korkong, S., Wasitthankasem, R., Vongpunsawad, S., Poovorawan, Y. Respiratory syncytial virus genotypes NA1, ON1, and BA9 are prevalent in Thailand, 2012-2015. PeerJ 2017, 5, e3970. [CrossRef]
- Jia, R., Lu, L., Su, L., Lin, Z., Gao, D., Lv, H., Xu, M., Liu, P., Cao, L., Xu, J. Resurgence of Respiratory Syncytial Virus Infection During COVID-19 Pandemic Among Children in Shanghai, China. Front Microbiol 2022, 13, 938372. [CrossRef]
Figure 1.
Phylogeny of HRV 5′UTR and capsid coding region sequences of samples from SUDI cases and reference sequences obtained from NCBI BLAST.
Figure 1.
Phylogeny of HRV 5′UTR and capsid coding region sequences of samples from SUDI cases and reference sequences obtained from NCBI BLAST.
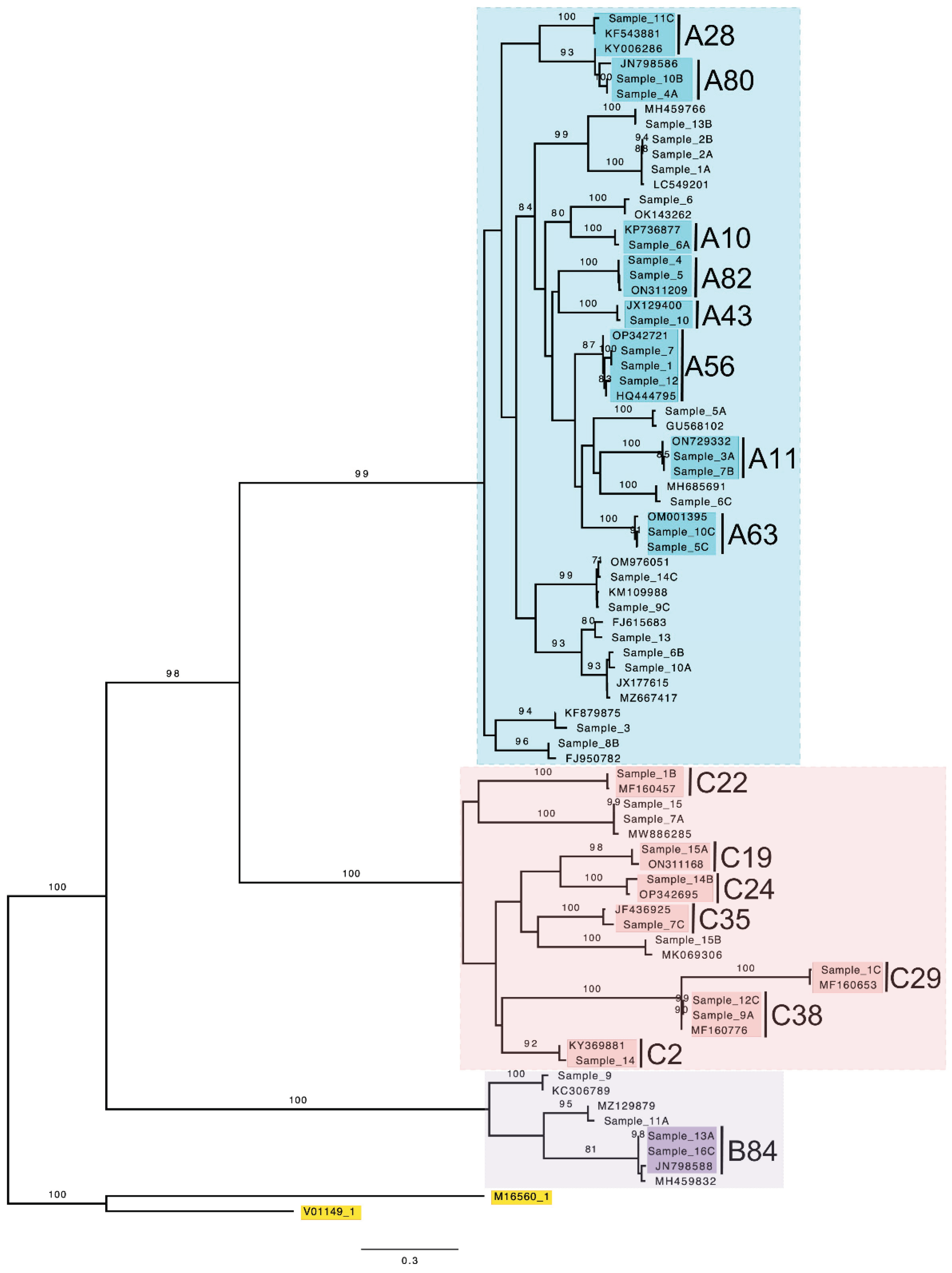
Figure 2.
Phylogeny of RSV G-gene sequences of samples from SUDI cases and reference sequences obtained from NCBI BLAST.
Figure 2.
Phylogeny of RSV G-gene sequences of samples from SUDI cases and reference sequences obtained from NCBI BLAST.
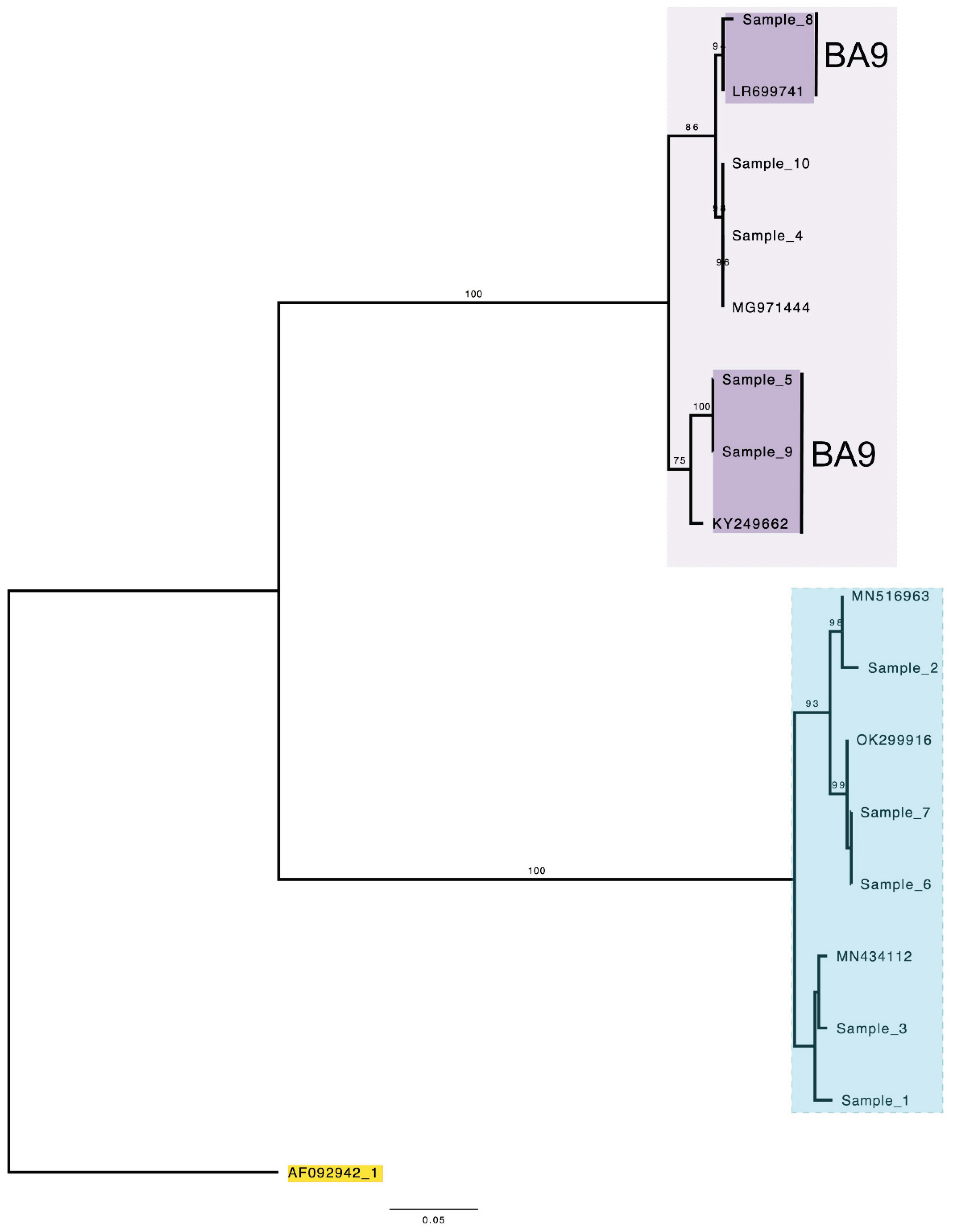
Figure 3.
Deduced amino acid alignment of RSV G-gene of RSV-A samples. Sample sequences were aligned and are shown relative to the sequence of RSV ON1 (Accession number: JN257693; ON67-1210A).
Figure 3.
Deduced amino acid alignment of RSV G-gene of RSV-A samples. Sample sequences were aligned and are shown relative to the sequence of RSV ON1 (Accession number: JN257693; ON67-1210A).
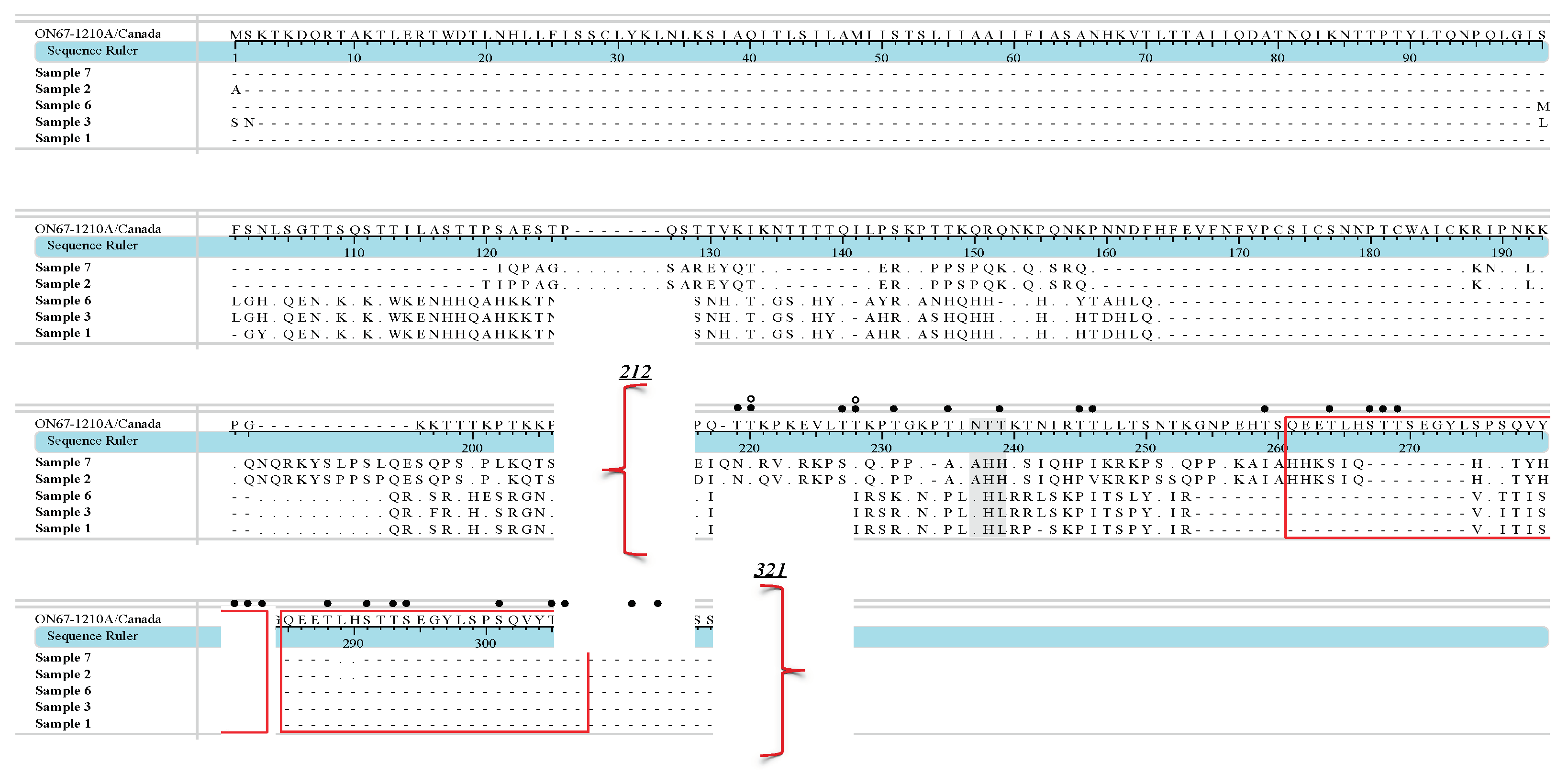
Figure 4.
Deduced amino acid alignments of RSV G-gene sequences of RSV-B sample. Samples sequences were aligned and are shown relative to sequence of prototype RSV-BA1 (Accession number: AY333364; BA/4128/99B) and BA9 (Accession number: DQ227395; BA/100/04).
Figure 4.
Deduced amino acid alignments of RSV G-gene sequences of RSV-B sample. Samples sequences were aligned and are shown relative to sequence of prototype RSV-BA1 (Accession number: AY333364; BA/4128/99B) and BA9 (Accession number: DQ227395; BA/100/04).
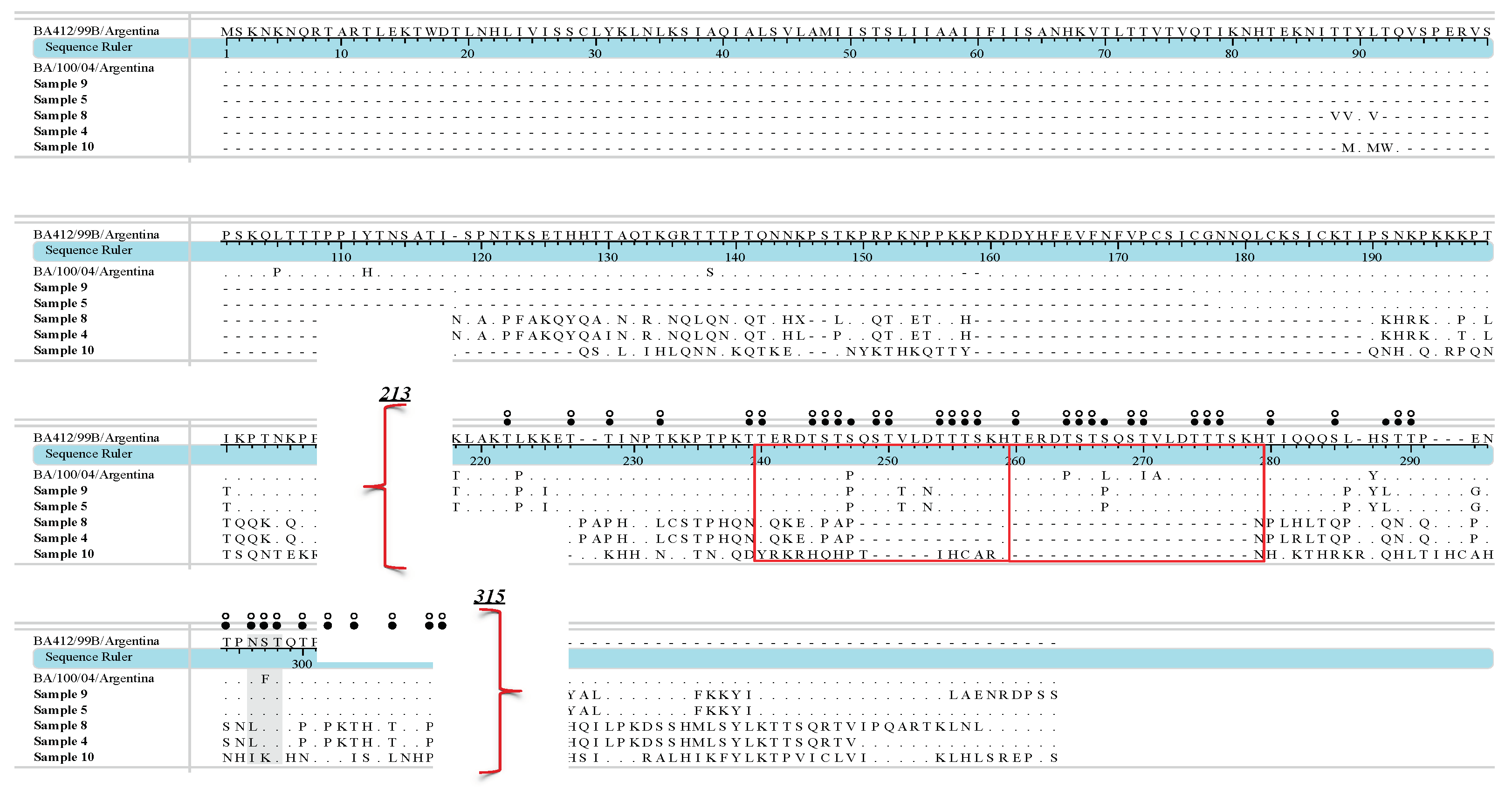
Table 1.
Pre-Nested and Nested Master Mix used for HRV 5′UTR Sequencing.
| Pre-Nested Master Mix | Final concentration | Volume (μl) |
|---|---|---|
| Nuclease Free Water | - | 26 |
| AMV/Tfl 5X Reaction Buffer | 1X | 10 |
| dNTP Mix | 0.2 mM of each | 1 |
| Forward Primer | 0.2 pmol/μl | 1 |
| Reverse Primer | 0.2 pmol/μl | 1 |
| MgSO4 | 1 mM | 4 |
| AMV Reverse Transcriptase | 0.1 u/μl | 1 |
| Tfl DNA Polymerase | 0.1 u/μl | 1 |
| Template RNA | - | 5 |
| Nested Master Mix | ||
| Nuclease Free Water | - | 30.5 |
| 5X GoTaq® Flexi Buffer | 1X | 10 |
| PCR nucleotide mix | 0.2 mM of each | 1 |
| Forward Primer | 0.2 pmol/μl | 1 |
| Reverse Primer | 0.2 pmol/μl | 1 |
| MgCl2 | 2 mM | 4 |
| Taq DNA Polymerase | 1.25 u | 0.5 |
| Pre-Nested PCR product | - | 2 |
Table 2.
HRV primers used for the amplification of the HRV5′UTR and VP4 regions [22].
| PCR Stage | Name | Sequence 5′ → 3′ |
|---|---|---|
| Pre-Nested | RV-Forward 1 | CTC CGG CCC CTG AAT RYG GCT AA |
| RV-Reverse 1 | TCI GGI ARY TTC CAS YAC CAI CC | |
| Nested | HRV 01.3 | TAC TTT GGG TGT CCG TGT TTC |
| HRV 02.3 | GGC AAC TTC CAC CAC CC |
Table 3.
Pre-Nested and Nested Master Mix.
| Pre-Nested Master Mix | Final concentration | Volume (μl) |
|---|---|---|
| Nuclease Free Water | - | 20.6 |
| AMV/Tfl 5X Reaction Buffer | 1X | 10 |
| dNTP Mix | 0.2 mM of each | 1 |
| Forward Primer | 0.2 pmol/μl | 1 |
| Reverse Primer | 0.2 pmol/μl | 1 |
| MgSO4 | 1 mM | 4 |
| AMV Reverse Transcriptase | 0.1 u/μl | 1 |
| Tfl DNA Polymerase | 0.1 u/μl | 1 |
| Dithiothreitol | 0.4 | |
| Template RNA | - | 10 |
| Nested Master Mix | ||
| Nuclease Free Water | - | 30.5 |
| 5X GoTaq® Flexi Buffer | 1X | 10 |
| PCR nucleotide mix | 0.2 mM of each | 1 |
| Forward Primer | 0.2 pmol/μl | 1 |
| Reverse Primer | 0.2 pmol/μl | 1 |
| MgCl2 | 2 mM | 4 |
| Taq DNA Polymerase | 1.25u | 0.5 |
| Pre-Nested PCR product | - | 2 |
Table 4.
RSV primers used for the amplification of the G-gene region [63].
| PCR Stage | Name | Sequence 5′ → 3′ |
|---|---|---|
| Pre-Nested | SH1 | CAC AGT KAC TGA CAA YAA AGG AGC |
| F164 | GTT ATG ACA CTG GTA TAC CAA CC | |
| Nested | ABG490 | ATG ATT WYC AYT TTG AAG TGT TC |
| F9AB | CAA CTC CAT KRT TAT TTG CC |
Table 5.
PCR-positive HRV and RSV results from previous study samples (Expected) vs. confirmatory AllplexTM RV Essential results (Detected).
Table 5.
PCR-positive HRV and RSV results from previous study samples (Expected) vs. confirmatory AllplexTM RV Essential results (Detected).
| Study | Virus | Expected | Detected | Selected for sequencing | PCR products obtained |
|---|---|---|---|---|---|
| 2015/2016 | HRV | 68 | 49 | 49 | 45 |
| RSV | 11 | 10 | 7 | 7 | |
| 2018/2019 | HRV | 38 | 15 | 15 | 15 |
| RSV | 45 | 5 | 3 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



