Submitted:
01 March 2024
Posted:
01 March 2024
You are already at the latest version
Abstract
The mechanism of sex determination and differentiation in animals remains a central focus of reproductive and developmental biology research, and the regulation of sex differentiation in amphioxus remains poorly understood. Cytochrome P450 Family 19 Subfamily A member 1 (CYP19A1) is a crucial sex differentiation gene that catalyzes the conversion of androgens into estrogens. In this study, we identified two aromatase-like genes in amphioxus: cyp19-like1 and cyp19-like2. Cyp19-like1 is more primitive and may represent the ancestral form of cyp19 in zebrafish and other vertebrates. Cyp19-like2, on the other hand, is likely the result of gene duplication within am-phioxus suggesting its potential role as a key gene in sex differentiation. To gain further insights into the expression patterns of these two aromatase-like, we examined their expression in different gonad tissues and during different stages of gonad development. While the expression patterns of the two genes differ in gonad tissues, both are highly expressed in the gonad primordium and are primarily localized to microsomal membrane systems. However, as development proceeds, their expression levels decrease significantly. This study enhances our understanding of sex dif-ferentiation mechanisms in amphioxus and provides valuable insights into the formation and evolution of sex determination mechanisms in vertebrates.
Keywords:
Amphioxus
; Invertebrates
; Sex differentiation
; Cyp19
1. Introduction
Sex determination and differentiation, crucial for species continuation, is a complex process influenced by various factors within and outside cells. It involves genetic gonad differentiation shaped by the environment, with multiple cells and organs participating. Gender differentiation builds on this to determine an organism's female or male phenotype. Despite diverse sex determination mechanisms among animals, they all rely on gene expression products. Understanding these molecular mechanisms is crucial. Vertebrates show diverse and uncertain sex determination mechanisms, yet conserved pathways and genes exist across species. For instance, Sry is a switch gene in mammals [1], and other genes related to female and male sex formation have been discovered, such as Cyp19a1, Wnt4, Foxl2, Amh, Sf1, Dmrt1, and Dax1 [2,3,4,5,6]. Signaling molecules like BMP and WNT also regulate sex differentiation [7,8,9,10]. Invertebrates may have sex chromosomes or alternative systems like haploidy. The functions of conserved genes and pathways in genetic sex determination remain unclear, warranting further amphioxus research.
As a Protochordata, the amphioxus bridges invertebrates and vertebrates, playing a key role in evolution [11]. Since its discovery in 1774, it has been a valuable model for vertebrate studies [12,13]. With fewer lineage-specific changes than Urochordate [14], the amphioxus offers a simpler body structure for studying vertebrate origins and invertebrate evolution [15,16]. It exhibits sexual dimorphism with a 1:1 sex ratio [17,18]. The gonads, attached to the inner walls of the peribranchial cavity, are composed of 26 pairs of rectangular sacs with thick walls and its gonads can be clearly observed due to skin transparency. Research on amphioxus gonadal development is fragmented, but studies suggest involvement of estrogen, androgen, and other hormones [19,20]. Chromosome analysis hints at sex chromosome involvement [21], and genome comparisons reveal species-specific sex-determining regions and genes [22]. Amphioxus lacks LH and FSH but has a thyrostimulin [23], suggesting a primitive pituitary-gonadal axis [24,25,26]. However, the sex determination mechanism in amphioxus remains largely unknown.
The CYP superfamily is extensive, with over 13,000 genes across 400 gene families [27]. The Cytochrome P450 Family 19 Subfamily A member 1 (CYP19A1) is crucial for sex differentiation. This enzyme converts androgens to estrogens [28], regulating vertebrate sex development [29,30,31,32]. In most vertebrates, there is only one aromatase gene. However, pigs have three aromatase genes, and the Actinopterygii of Osteichthyes have two aromatase genes. In Actinopterygii, there are two types of aromatase: ovarian and brain aromatase, encoded by the genes cyp19a1a and cyp19a1b, respectively. The ovarian aromatase plays a crucial role in sex differentiation. The Cyp19a1a is primarily expressed in the ovaries, and its knockout in zebrafish leads to female-to-male sex reversal and delayed testis development [33,34,35,36,37]. Cyp19a1b is primarily expressed in the brain and has significant impacts on brain development and function. It is involved in neuroendocrine metabolism during gonad development and also maintains the testis [38,39]. Cyp19a1a and Cyp19a1b differ structurally and functionally, possibly due to genome duplication under selective pressure during evolution [38,40]. Over the course of evolution, vertebrate cyp19a1 have exhibited high sequence conservation and good synteny [41].
Mizuta cloned amphioxus cytochrome P450 members, which encode proteins involved in catalyzing key reactions in the synthesis of progesterone, androgen, and estrogen from cholesterol, such as CYP11A, CYP17, and CYP19 [42,43]. We searched the amphioxus genome and found two cyp19-like on separate chromosomes. These genes don't cluster with zebrafish cyp19a1a or cyp19a1b but sit at the evolutionary tree's base. We cloned these aromatase-like sequences from amphioxus Branchiostoma japonicum and used bioinformatics and molecular techniques to understand their functions, structures, and evolutionary relationships. We also studied their expression in amphioxus tissues and gonads using various methods. This comprehensive approach allowed us to determine the expression pattern of cyp19 in the male and female gonads of amphioxus, providing valuable insights into its role in gonadal differentiation and development.
2. Materials and Methods
2.1. Experimental Materials
The Amphioxus (Branchiostoma japonicum) can be collected near the Shazikou sea area of Qingdao in China in early May every year. Immediately after collection, preliminary processing such as cleaning, weighing, and recording should be performed. When sexually mature, amphioxus can be distinguished by the color of its gonads: the testes appear white, while the ovaries appear pale yellow. Depending on the experimental requirements, the amphioxus can be used fresh or preserved appropriately. The experimental cells are HEK 293T human embryonic kidney cell line, purchased from the American Type Culture Collection (ATCC). Upon receiving the cells, they should be revived, passaged, and cryopreserved according to the operating guidelines provided by ATCC. It is essential to ensure that the cells maintain good growth under suitable conditions. Six-week-old ICR female mice were used for antibody preparation. Before antibody preparation, the mice should undergo adaptive breeding to ensure they are in good health. During the immunization process, strict adherence to experimental animal handling guidelines is crucial to ensure reliable experimental results and animal welfare.
2.2. RNA Extraction and cDNA Synthesis
Prepare and quickly process Amphioxus samples. Use RNA extraction reagents for tissue lysis and RNA isolation, followed by washing, drying, and dissolving the RNA. Quality check the RNA. Then, remove genomic DNA from the RNA and reverse transcribe it into cDNA using reverse transcriptase and primers. Purify the cDNA if needed, and detect its quality by PCR. All steps should be performed in an RNase-free environment, adhering to laboratory safety protocols.
2.3. Cloning of the cyp19
Initially, a comprehensive BLAST search was executed on the NCBI (ncbi.nlm.nih.gov), utilizing the zebrafish cyp19a1a sequence as the query. This meticulous exploration yielded two promising gene sequences from the Florida amphioxus database, designated as aromatase-like (XP_035672840.1) and aromatase-like (XP_035669280.1). Leveraging the transcriptome and genome data from the amphioxus from Qingdao, meticulous sequence alignments were then performed. These alignments facilitated the identification of two homologous sequences, namely cyp19-like1 (corresponding to XP_035672840.1) and cyp19-like2 (corresponding to XP_035669280.1). Subsequently, primers were carefully designed for the cloning of the ORF sequences specific to the cyp19-like1 and cyp19-like2, paving the way for further downstream analyses.

Table 1.
The sequences of primers and peptides used in this study.
| Experiment | Description | Sequences (5′–3′) |
|---|---|---|
| 5′RACE | P1(cyp19-like1 F) | ATGTACGGAGTGATCTCTCTCCTTA |
| P2(cyp19-like1 R) | CTAGTTTCTCTCTTCAAAGTACATG | |
| P3(cyp19-like2 F) P4(cyp19-like2 R) |
ATGGACCTAGGCGAAGGCTGGGACG TCAGCTGTTGTCCACCCTTGGGTAC |
|
| Real-time PCR | P5(actin F) | TGCTGATTGTGGCTGCTGGTACTG |
| P6(actin F) P7(cyp19-like1 F) P8(cyp19-like1 R) P9(cyp19-like2 F) P10(cyp19-like2 R) |
GGTGTAGGCCAGCAGGGCGTG GCTCAGGAGGACGACAGGATTG GCAGCAGCGTACACATGATGG TTCGCCGCTGCTCTCATCCA CGGTCTCCGACGACTTCTCTGA |
|
| In situ hybridization | P11(cyp19-like1 F) | GCGTGGTCGCCGTTGTCGTT |
| P12(cyp19-like1 R) | CGCCGCAAGAAATCCAGAGCT | |
| P13(cyp19-like2 F) | GTGTATCCGCCATTGCTACC | |
| P14(cyp19-like2 R) | TCTCCGACGACTTCTCTGATT | |
| Subcellular Localization | P15(cyp19-like1 F) P16(cyp19-like1 R) P17(cyp19-like2 F) |
gcacagtggcggccgctcgagATGTACGGAGTGATCTCTCTCCTTACC gctcaccattctagactcgagGTTTCTCTCTTCAAAGTACATGTAACTAGC gcacagtggcggccgctcgagATGGACCTAGGCGAAGGCTG |
| P18(cyp19-like2 R) |
gctcaccattctagactcgagGCTGTTGTCCACCCTTGGG | |
| Polyclonal antibody preparation (mouse) |
Cyp19-like1 Cyp19-like2 |
N'-CREELKTAPPSDKPD-C' N'-CPSRDHKSLDVSRNL-C' |
2.4. Bioinformatics Analysis
Commence by acquiring the genome sequences of both organisms from the NCBI website. Make use of TBtools-, a versatile bioinformatics toolbox, to extract the exon and intron sequences pertaining specifically to the cyp19. These sequences can then be uploaded to the GSDS2.0 (Gene Structure Display Server 2.0 (gao-lab.org)) to generate illustrative gene structure diagrams that elucidate the intricacies of the gene's organization. In parallel, clone the cyp19-like1 and cyp19-like2 cDNA sequences from the amphioxus and translate them into protein sequences with the aid of EditSeq. Subsequently, align these protein sequences with the zebrafish Cyp19a1a and Cyp19a1b amino acid sequences retrieved from NCBI using Megalign7.1.0, enabling a comparative assessment of sequence similarity and an analysis of molecular parameters such as molecular weight. Additionally, harness the capabilities of SMART7.1.0 (SMART: Main page (embl.de)) and PHYRE2.0 (PHYRE2 ) to predict signal peptides, transmembrane regions, functional domains, and three-dimensional structures of the Cyp19 in both organisms, providing valuable insights into their structural and functional attributes. Lastly, embark on a phylogenetic analysis using MEGA10.1.6 to elucidate the evolutionary relationships within the Cyp superfamily. Gather amino acid sequences for Cyp19 from a diverse array of species and steroid biosynthesis-related Cyp subfamilies (Cyp3a, Cyp17a, Cyp21) from NCBI. Align these sequences using Clustal W and construct a phylogenetic tree employing the Maximum Likelihood method, revealing the evolutionary history and domain architecture of the Cyp19.
2.5. Real-Time PCR
Real-Time PCR is a technique for quantifying nucleic acid sequences in real-time during PCR amplification. It involves sample preparation, primer design, PCR setup with fluorescent probes or dyes, and real-time detection using a dedicated instrument. Data analysis provides quantitative results of the target nucleic acid in the sample. This method offers sensitivity, specificity, and direct quantification, making it useful in genetics, molecular biology, and diagnostics.
2.6. Paraffin Section In Situ Hybridization
The paraffin section in situ hybridization (ISH) technique involves several steps to localize specific mRNA sequences within histological specimens. Firstly, the paraffin-embedded sections undergo preprocessing, including dewaxing, hydration, and protease treatment to enhance probe accessibility. Next, the probe, which is a labeled oligonucleotide complementary to the target mRNA, is prepared and denatured. The denatured probe is then applied to the sections and incubated under controlled temperature conditions, allowing for hybridization between the probe and the target mRNA. After hybridization, unbound probe is removed, and the bound probe is detected using appropriate methods such as immunohistochemical staining. This technique enables the direct visualization of gene expression patterns within cells or tissues, providing valuable insights into gene regulation and function.
2.7. Hematoxylin and Eosin staining
Begin with tissue sample preparation, including dehydration, clearing, and embedding in paraffin. Slice the paraffin-embedded tissue into thin sections using a microtome. Then, remove the paraffin with xylene and hydrate the sections through graded alcohol concentrations. Next, stain the nuclear DNA blue with hematoxylin and the cytoplasm red with eosin. Post-processing involves dewatering, clearing, and mounting the sections on slides(0.4μm~0.8μm). Finally, observe the stained sections under a microscope, where the nuclei should appear blue and the cytoplasm red, providing a clear view of the tissue's morphology.
2.8. Protocol for Generating Mouse Polyclonal Antibodies
Select and prepare the antigen, mix it with an immune adjuvant, and emulsify. Immunize mice via intraperitoneal injection. Monitor antibody production periodically, collect mouse sera, and purify the antibodies. Identify the specificity and purity of the antibodies before storing them for future experiments.
2.9. Western Blot
Prepare protein samples, determine their concentration, and load them onto an SDS-PAGE gel for electrophoresis. After electrophoresis, transfer the proteins to a nitrocellulose membrane. Incubate the membrane with specific antibodies, wash to remove unbound antibodies, and visualize the target protein using a suitable substrate. Analyze the results, observing band positions and intensities to assess protein expression.
2.10. Immunohistochemical Staining
Obtain the tissue sample, perform dehydration, defatting, and fixation. Slice the sample into thin sections and place them on slides. Bind specific antibodies to the tissue sample, followed by washing to remove unbound antibodies. Then, bind specific secondary antibodies to the bound primary antibodies and wash again. Subsequently, conjugate the marker to the secondary antibodies and add a chromogen for color development, revealing the specific protein. Finally, mount the slides and observe the results under a microscope.
2.11. Subcellular Localization
Cultivate target cells and transfect them with plasmids labeled for the protein of interest. Fix and permeabilize the cells to allow antibody access. Add specific antibodies to recognize the target protein, followed by washing to remove unbound antibodies. Then, conjugate fluorescently labeled secondary antibodies to the primary antibodies. Observe the cells under a fluorescence microscope to localize the protein of interest within subcellular structures.
2.12. Statistical Analysis
Statistical analysis was performed using GraphPad Prism 9, with all assays conducted in triplicate technical and three times biologic replicates. Data were analyzed using One-way ANOVA or two-tailed Student's t-test and presented as means ± SD. Significance levels were set at *p < 0.05, **p < 0.01, and ***p < 0.001, with 'ns' indicating non-significance.
3. Results
3.1. Cloning of the cyp19
Using female amphioxus cDNA as a template, the complete ORF sequence of the cyp19-like gene in amphioxus B. japonicum was cloned through 5’RACE technology. The cyp19-like1 has a total length of 1491bp, encoding 496 amino acids (Figure A). The cyp19-like2 has a total length of 1482bp, encoding 494 amino acids (Figure B). Sequencing results confirmed the accuracy of the sequences.
Figure 1.
Complete ORF nucleotide sequences of two cyp19-like fragments in amphioxus. Figure A shows the cyp19-like1 sequence fragment in amphioxus, with a size of 1491bp. Figure B displays the cyp19-like2 sequence fragment in amphioxus, with a size of 1482bp.
Figure 1.
Complete ORF nucleotide sequences of two cyp19-like fragments in amphioxus. Figure A shows the cyp19-like1 sequence fragment in amphioxus, with a size of 1491bp. Figure B displays the cyp19-like2 sequence fragment in amphioxus, with a size of 1482bp.

3.2. Bioinformatic Analysis of the cyp19
3.2.1. Comparison of cyp19 Structures between Zebrafish and Amphioxus
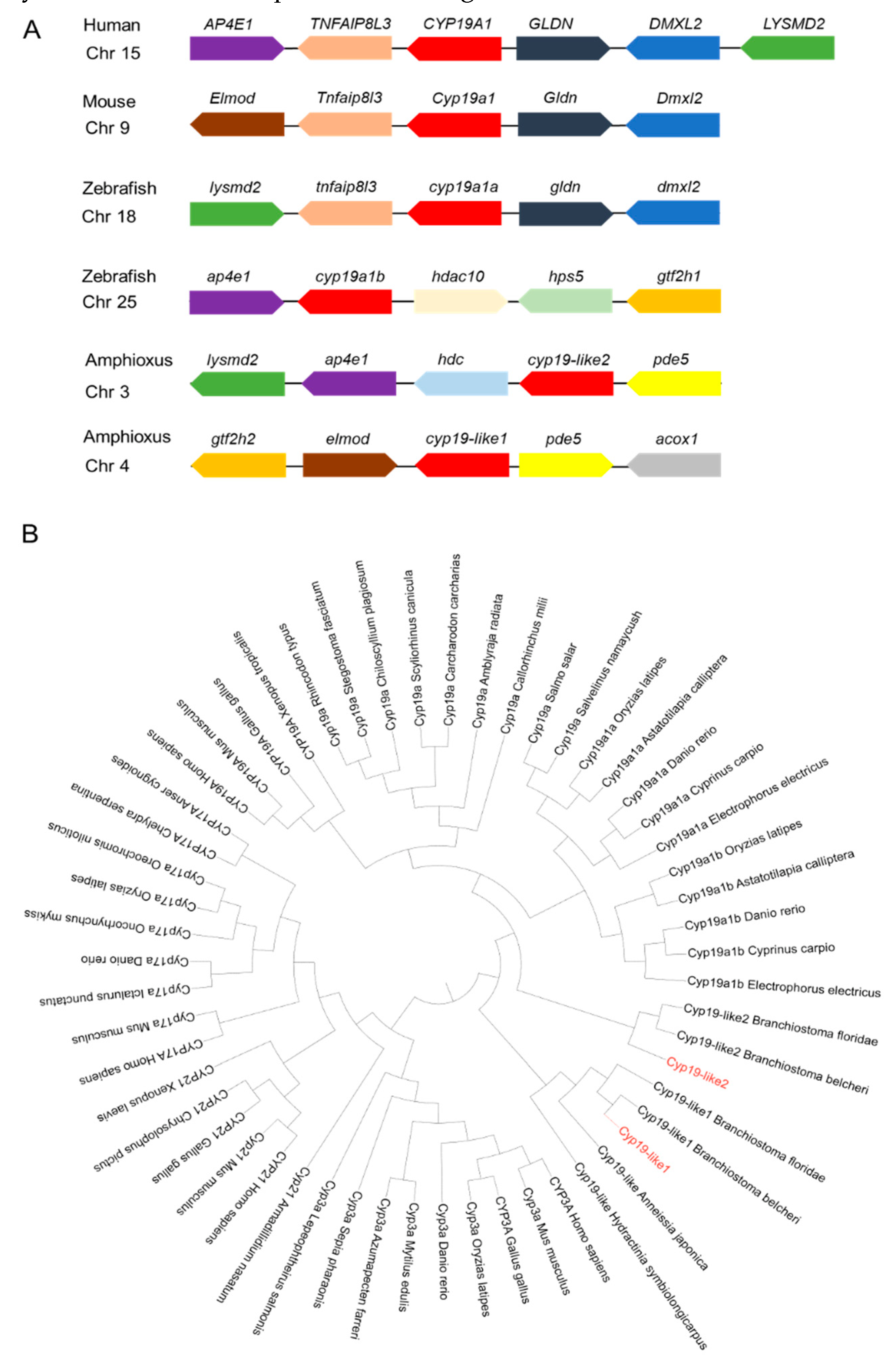
The zebrafish cyp19a1a gene has 9 exons located on chromosome 18, while cyp19a1b has 9 exons on chromosome 25. In amphioxus, cyp19-like1 has 10 exons on chromosome 4, and cyp19-like2 has 9 exons on chromosome 3. Since the zebrafish cyp19a1a and cyp19a1b genes are located on different chromosomes and have different functions, we hypothesize that the two cyp19-like in amphioxus, located on different chromosomes, may also exhibit functional differences.
Figure 2.
Comparison of the gene structures of cyp19a1a, cyp19a1b, and aromatase-like genes in zebrafish and Florida amphioxus.
Figure 2.
Comparison of the gene structures of cyp19a1a, cyp19a1b, and aromatase-like genes in zebrafish and Florida amphioxus.

3.2.2. Comparison of Sequence Similarity between Zebrafish and Amphioxus Cyp19
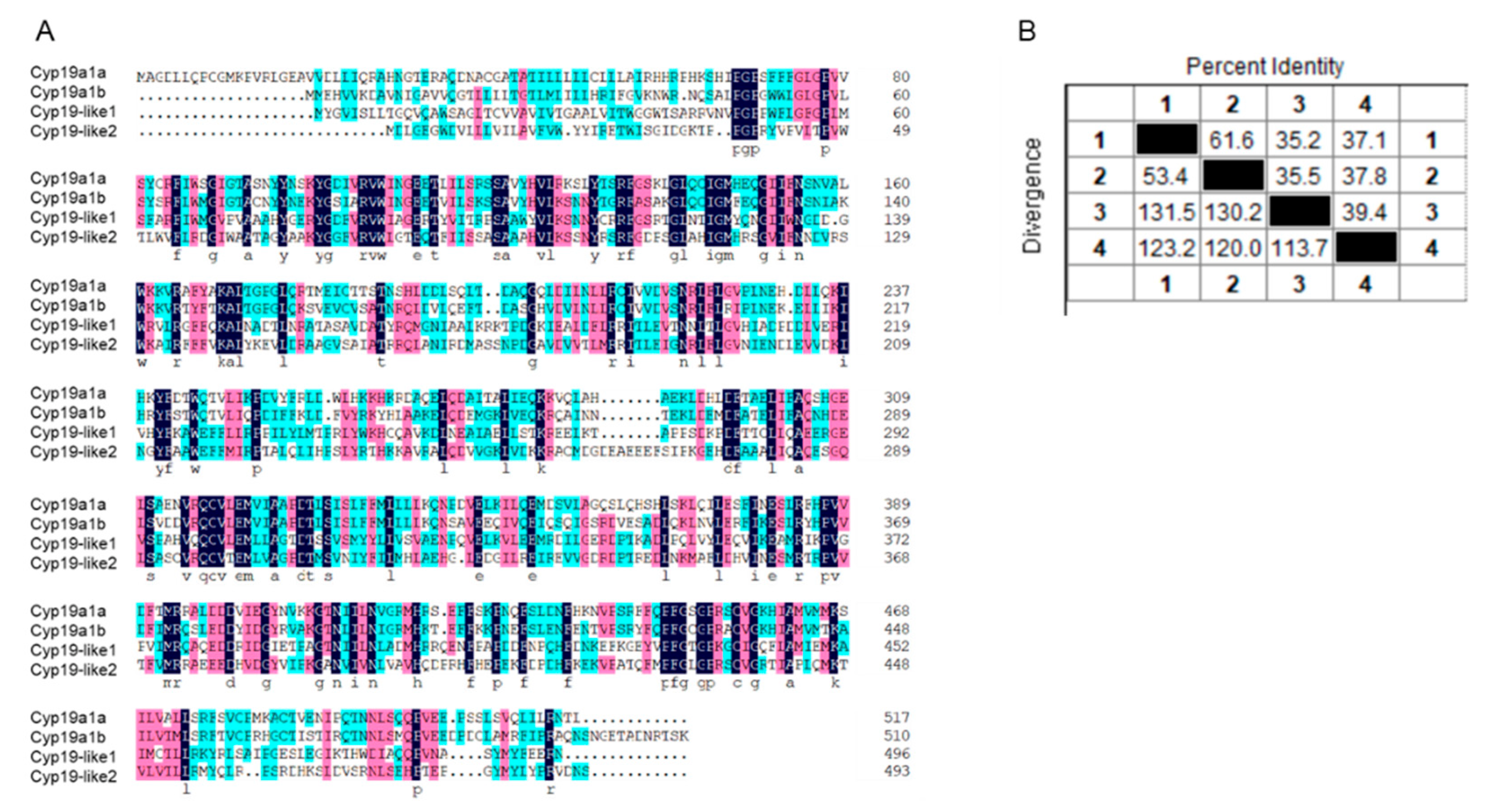
The amino acid sequence similarity between zebrafish cyp19a1a, cyp19a1b, and amphioxus cyp19-like1, cyp19-like2 is low (Figure A). Sequence alignment revealed that the similarity between the two amphioxus cyp19-like is only 39.4%. The sequence similarity between cyp19-like1 and cyp19a1a, cyp19a1b is 35.2% and 35.5%, respectively. The sequence similarity between cyp19-like2 and cyp19a1a, cyp19a1b is 37.1% and 37.8%, respectively (Figure B).
Figure 3.
Alignment of amino acid sequences of cyp19 in zebrafish and amphioxus. A: Amino acid sequence alignment; B: Sequence similarity comparison. 1 represents cyp19a1a, 2 represents cyp19a1b, 3 represents cyp19-like1, and 4 represents cyp19-like2.
Figure 3.
Alignment of amino acid sequences of cyp19 in zebrafish and amphioxus. A: Amino acid sequence alignment; B: Sequence similarity comparison. 1 represents cyp19a1a, 2 represents cyp19a1b, 3 represents cyp19-like1, and 4 represents cyp19-like2.
3.2.3. Prediction and Comparison of Cyp19 Structures between Zebrafish and Amphioxus
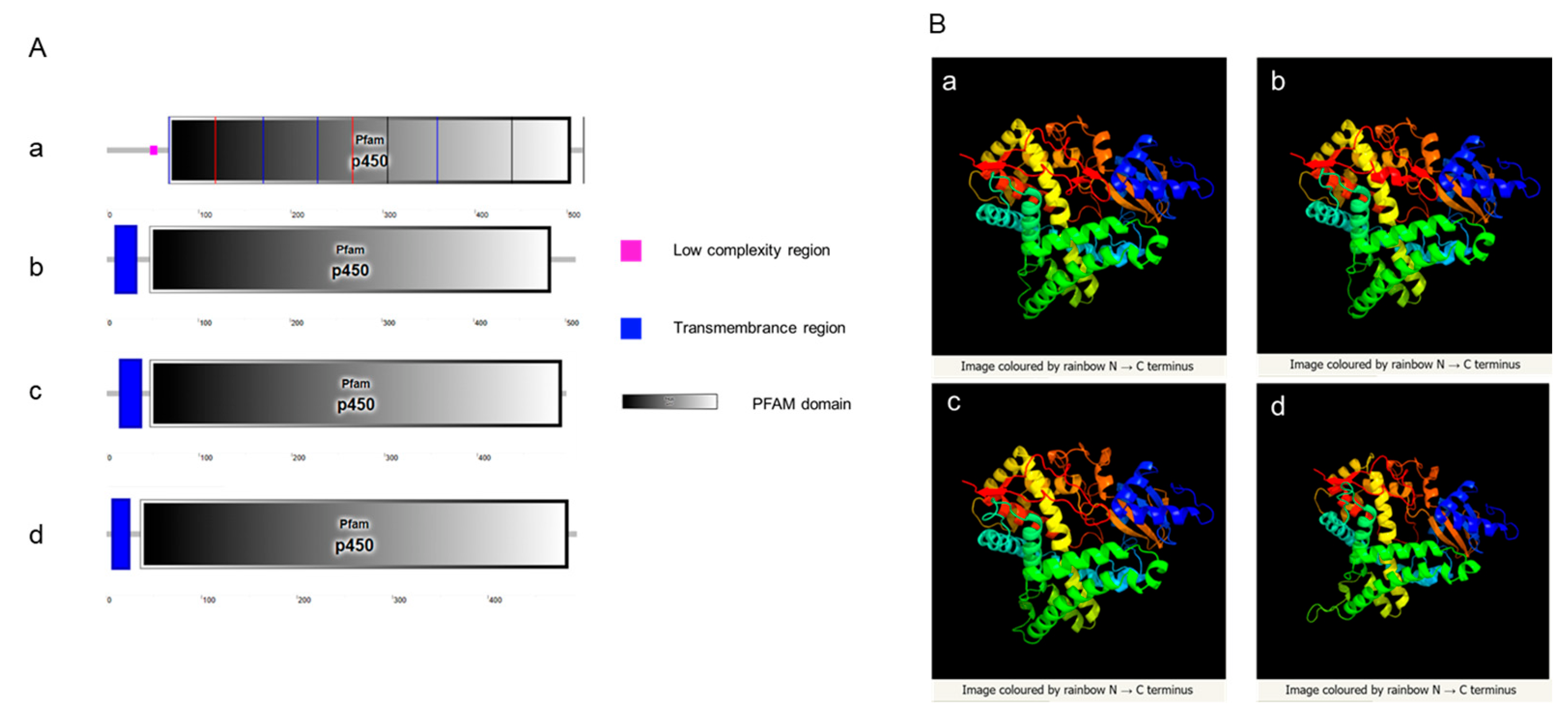
SMART website was used to predict signal peptides, transmembrane regions, and domains of Cyp19 in zebrafish and amphioxus. The results showed that both amphioxus and zebrafish possess a conserved cytochrome p450 domain. Zebrafish Cyp19a1a does not have a transmembrane region, while Cyp19a1b, Cyp19-like1, and Cyp19-like2 have a transmembrane region at the N-terminus (Figure A). Three-dimensional structure prediction of Cyp19 encoded by zebrafish and amphioxus revealed a high degree of similarity in their three-dimensional structures (Figure B), indicating potential similar functions.
Figure 4.
Prediction of Cyp19 structures in zebrafish and amphioxus. A: SMART prediction of domains in zebrafish Cyp19a1a (a), Cyp19a1b (b), and amphioxus Cyp19-like1 (c), Cyp19-like2 (d); B: PHYRE2 prediction of three-dimensional structures in zebrafish Cyp19a1a (a), Cyp19a1b (b), and amphioxus Cyp19-like1 (c), Cyp19-like2 (d).
Figure 4.
Prediction of Cyp19 structures in zebrafish and amphioxus. A: SMART prediction of domains in zebrafish Cyp19a1a (a), Cyp19a1b (b), and amphioxus Cyp19-like1 (c), Cyp19-like2 (d); B: PHYRE2 prediction of three-dimensional structures in zebrafish Cyp19a1a (a), Cyp19a1b (b), and amphioxus Cyp19-like1 (c), Cyp19-like2 (d).
3.2.4. Synteny and Evolutionary Analysis of Cyp19
Through Cyp19 synteny analysis, it was found that the protein has synteny in humans, mice, zebrafish, and amphioxus. Cyp19-like1 and cyp19-like2 exhibit synteny, but cyp19-like2 has stronger synteny with humans, mice, and zebrafish, suggesting it may play a more important role in sex determination (Figure A). The main function of Cyp19 is to participate in steroid synthesis. Among the Cyp superfamily, other subfamilies related to steroid biosynthesis include Cyp3a, Cyp17a, and Cyp21. The results showed that the three amphioxus Cyp19-like sequences belong to the Cyp19 subfamily. Amphioxus Cyp19-like1 clusters with invertebrates such as hydroids and sea lilies, indicating a more primitive evolutionary status. In contrast, Cyp19-like2 clusters with vertebrates, suggesting it may be a key gene in sex determination (Figure B). Analysis revealed that the two cyp19-like sequences in amphioxus do not have a one-to-one correspondence with cyp19a1a and cyp19a1b. Amphioxus cyp19-like1 may be the primitive form before the evolution of other vertebrates, while cyp19-like2 may be the result of amphioxus's own genome
Figure 5.
Analysis of synteny and phylogeny of Cyp superfamily across different species. A: Location maps of Cyp19 on amphioxus and other vertebrate chromosomes. Boxes represent genes, and the direction of the boxes indicates the orientation of the genes. B: Phylogenetic analysis of Cyp superfamily across different species. Sequence sources: see Supplemental Table S1.
Figure 5.
Analysis of synteny and phylogeny of Cyp superfamily across different species. A: Location maps of Cyp19 on amphioxus and other vertebrate chromosomes. Boxes represent genes, and the direction of the boxes indicates the orientation of the genes. B: Phylogenetic analysis of Cyp superfamily across different species. Sequence sources: see Supplemental Table S1.
3.3. Expression patterns of two cyp19-like genes in the gonads of male and female amphioxus
3.3.1. Differential tissue expression of cyp19
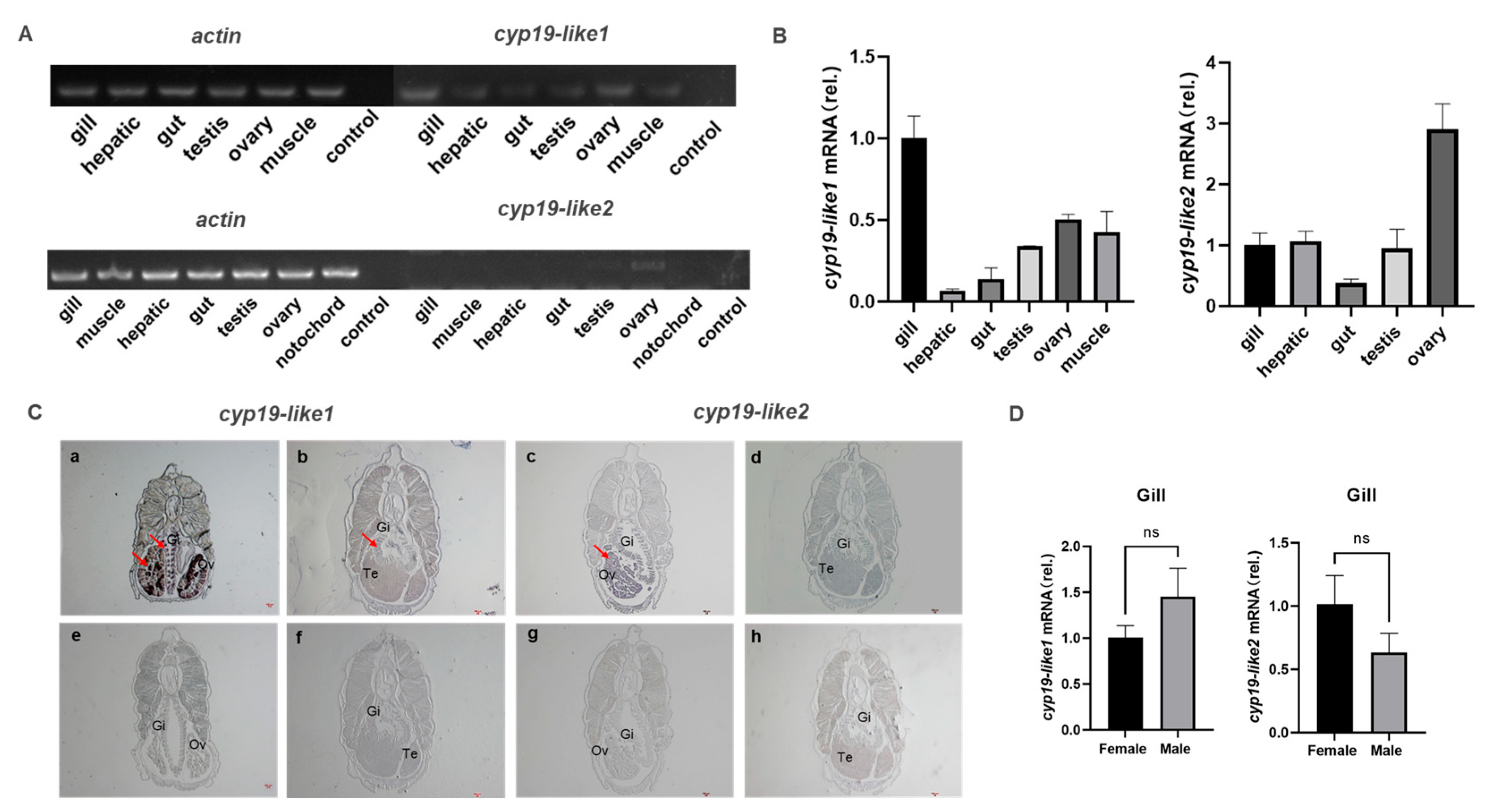
We have demonstrated through semi-quantitative, Real-time PCR, and in situ hybridization techniques that there are differences in the expression of cyp19-like1 and cyp19-like2 in different tissues of adult amphioxus. Semi-quantitative and Real-time PCR results show that cyp19-like1 is highest expressed in the gills, followed by the ovaries in amphioxus. Cyp19-like2 is highly expressed in the ovaries but is also present in tissues such as the gills (Figures A and B). In situ hybridization results show that both cyp19-like1 and cyp19-like2 have significant positive signals in the gills and ovaries (Figure C). The Real-time PCR and in situ hybridization results are similar to the characteristic of high expression of cyp19a1 in the ovaries of vertebrates. However, unlike vertebrates, cyp19-like are also highly expressed in the gills of amphioxus. To further investigate the expression of cyp19-like in the gills of male and female amphioxus, we conducted Real-time PCR using male and female gill templates. The results showed no significant difference in the expression of cyp19-like1 and cyp19-like2 in the gills (Figure D).
Figure 6.
Tissue expression and localization of cyp19-like in amphioxus. A: Semi-quantitative PCR detection of cyp19-like gene expression in different tissues of amphioxus. B: Real-time PCR detection of cyp19-like gene expression in different tissues of amphioxus, including gill, hepatic, gut, testis, ovary, muscle and notochord; C: In situ hybridization results of cyp19-like on paraffin sections of amphioxus. a-d show antisense probe hybridization results; e-h show sense probe hybridization results (negative control). Gi: Gill; Ov: Ovary; Te: Testis. Arrows indicate positive signals. Scale bar: 100μm. D: Expression of cyp19-like in the gills of male and female amphioxus. ns indicates no significant difference.
Figure 6.
Tissue expression and localization of cyp19-like in amphioxus. A: Semi-quantitative PCR detection of cyp19-like gene expression in different tissues of amphioxus. B: Real-time PCR detection of cyp19-like gene expression in different tissues of amphioxus, including gill, hepatic, gut, testis, ovary, muscle and notochord; C: In situ hybridization results of cyp19-like on paraffin sections of amphioxus. a-d show antisense probe hybridization results; e-h show sense probe hybridization results (negative control). Gi: Gill; Ov: Ovary; Te: Testis. Arrows indicate positive signals. Scale bar: 100μm. D: Expression of cyp19-like in the gills of male and female amphioxus. ns indicates no significant difference.
3.3.2. Expression of cyp19 at different stages
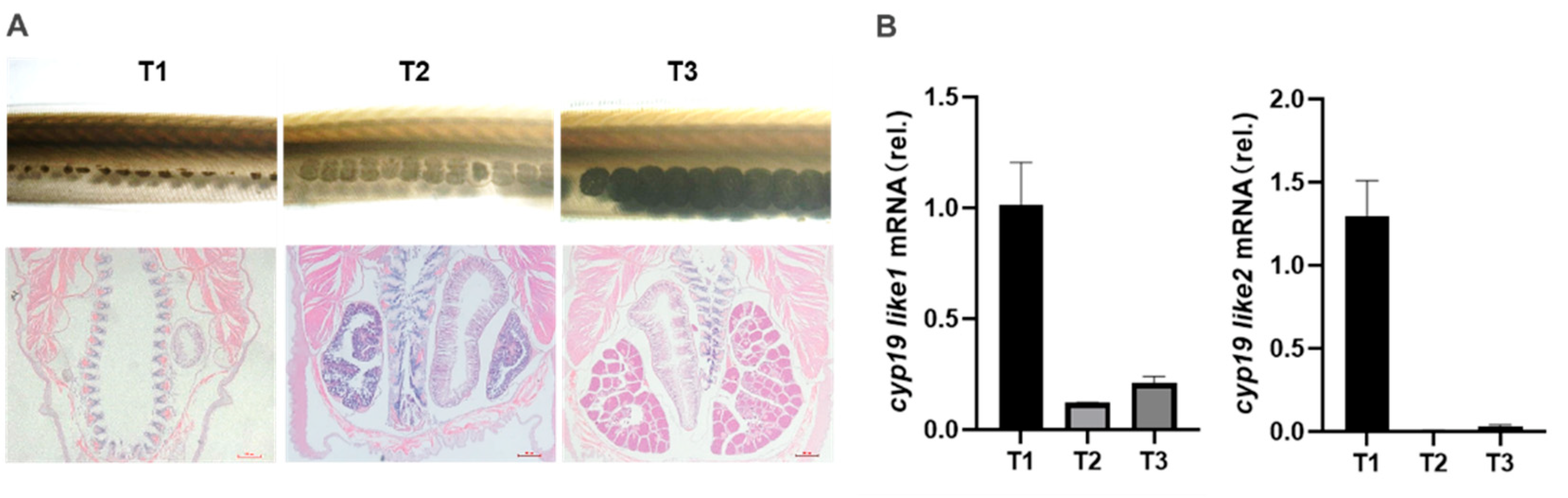
During our laboratory breeding period, we observed the gonadal development of amphioxus and found that the gonads begin to develop in April, with the appearance of small black particles (gonadal primordia) on both sides of the amphioxus body. The development time of male and female gonads and both sides of the gonads is relatively consistent. In May, the gonads begin to mature, and at this time, they are transparent vesicular structures, and the black particles become larger and lighter with development. June to July is the peak period of maturity, with large and full gonads, rectangular shape, white testes in males, and yellow ovaries in females. The spawning peak occurs in July.
Based on observations of gonadal morphology and histology, we divided the gonadal development cycle of amphioxus into four stages: undifferentiated gonad (T0), gonadal primordium (T1), gonadal development (T2), and gonadal maturity (T3) (Figure A). We used Real-time PCR to detect the expression patterns of the two cyp19-like at different stages of gonadal development in amphioxus. The results showed that both cyp19-like1 and cyp19-like2 are highly expressed at the beginning of gonadal development, i.e., the gonadal primordium stage, and their expression decreases with the maturation of the gonads (Figure B).
Figure 7.
Expression of cyp19-like in amphioxus at different developmental stages. A: Tissue morphology and HE staining results at different stages of gonad development in amphioxus. B: Real-time PCR detection of cyp19-like expression at different stages of gonad development in amphioxus. T1, T2, and T3 represent the primordial, developing, and mature gonad stages, respectively.
Figure 7.
Expression of cyp19-like in amphioxus at different developmental stages. A: Tissue morphology and HE staining results at different stages of gonad development in amphioxus. B: Real-time PCR detection of cyp19-like expression at different stages of gonad development in amphioxus. T1, T2, and T3 represent the primordial, developing, and mature gonad stages, respectively.
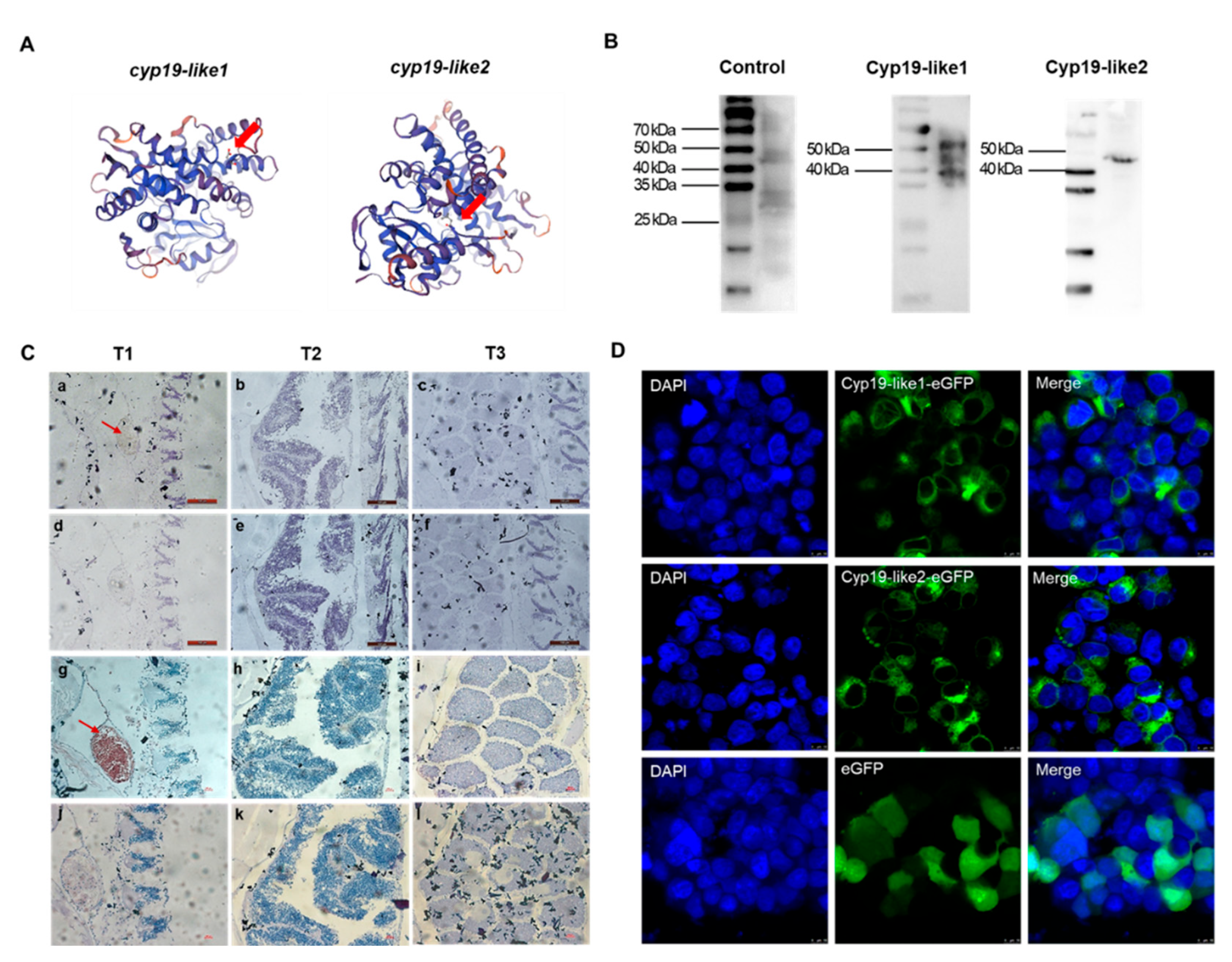
3.3.3. Temporal expression and cellular localization of Cyp19
The immunogenic peptide sequences of Cyp19-like1 and Cyp19-like2 are N'-CREELKTAPPSDKPD-C' and N'-CPSRDHKSLDVSRNL-C', respectively. These immunogenic peptide sequences are located on the outer side of the predicted Cyp19 protein three-dimensional structure (Figure A). The antibody preparation effect was enhanced by KLH coupling. We used WB to detect the specificity of the prepared Cyp19 polyclonal antibody and the size of the Cyp19. The results showed good specificity of the Cyp19 antibody (Figure B).
We selected amphioxus individuals at three different stages and detected the protein expression levels of the two Cyp19-like at different stages of gonadal development using immunohistochemical staining. The immunohistochemical staining results were consistent with the Real-time PCR results, showing that Cyp19-like1 and Cyp19-like2 are highly expressed at the beginning of gonadal development, i.e., the gonadal primordium stage, and their expression gradually decreases with the maturation of the gonads. Additionally, the expression level of Cyp19-like2 was higher (Figure C). Furthermore, we examined the localization of Cyp19 in cells and found that the protein is a membrane protein mainly localized to mitochondrial membranes, endoplasmic reticulum membranes, and other microsomal membrane systems (Figure D).
Figure 8.
Antibody preparation and protein expression of amphioxus Cyp19-like. A: Designated sites for immunogenic peptides which were indicated with arrows. B: Western blot results of Cyp19-like in Amphioxus. C: Immunohistochemical results of Cyp19-like in Amphioxus. T1, T2, and T3 represent the primordial, developing, and mature gonad stages, respectively. a-f show Cyp19-like1 immunohistochemical results, while g-l show Cyp19-like2 immunohistochemical results. a-c, g-i are experimental groups, while d-f, j-l are control groups. Scale bar: 100μm. D: Subcellular localization of amphioxus Cyp19-like in HEK cellls using LSCM. Scale bar: 10μm.
Figure 8.
Antibody preparation and protein expression of amphioxus Cyp19-like. A: Designated sites for immunogenic peptides which were indicated with arrows. B: Western blot results of Cyp19-like in Amphioxus. C: Immunohistochemical results of Cyp19-like in Amphioxus. T1, T2, and T3 represent the primordial, developing, and mature gonad stages, respectively. a-f show Cyp19-like1 immunohistochemical results, while g-l show Cyp19-like2 immunohistochemical results. a-c, g-i are experimental groups, while d-f, j-l are control groups. Scale bar: 100μm. D: Subcellular localization of amphioxus Cyp19-like in HEK cellls using LSCM. Scale bar: 10μm.
4. Discussion
The aromatase, encoded by the cyp19a1, plays a crucial role in vertebrate reproduction by catalyzing the conversion of androgens to estrogens. While aromatase is known to be critical in sex determination and differentiation in animals, the origin of the cyp19a1 remains enigmatic. In vertebrates, the majority of animals possess a single aromatase gene. However, fish belonging to the Actinopterygii subclass of Osteichthyes exhibit a unique feature: they possess two aromatase genes, ovarian aromatase and brain aromatase. These genes are expressed in distinct locations and exhibit structural and functional differences. Through sequence alignment, we discovered the existence of two aromatase-like in Florida amphioxus, situated on chromosomes 4 and 3, respectively. Utilizing amphioxus B. japonicum as our material, we successfully cloned these two homologous sequences, naming them cyp19-like1 and cyp19-like2.
Given the presence of amphioxus cyp19-like1 and cyp19-like2, which are also situated on different chromosomes and display structural and sequence variations, it raises intriguing questions regarding their specific functions in amphioxus and whether both genes play a role in sex differentiation. cyp19-like1 has 10 exons, while the other sequences possess 9 exons, which aligns with the basic structure of vertebrate cyp19 having 9 exons [42,43]. Amphioxus and zebrafish share a conserved cytochrome p450 domain, indicating a common ancestral origin. Notably, zebrafish Cyp19a1a lacks a transmembrane region, whereas zebrafish Cyp19a1b, amphioxus Cyp19-like1, and amphioxus Cyp19-like2 all possess a transmembrane region at their N-termini. This feature suggests potential differences in their subcellular localization and function.
The question at hand concerns is the functional relationship between the two cyp19-like in amphioxus and the cyp19a1a and cyp19a1b in fish, particularly in the context of their evolutionary history. Zhang and colleagues' observations about the strong synteny and sequence conservation of vertebrate cyp19a1 during evolution provide a valuable baseline for comparison. Despite this conservation, amphioxus Cyp19 (ABA47317.1) does not share synteny with vertebrate cyp19a1, suggesting that the direct evolutionary link between these two genes is tenuous. In their opinion, it appears that the cyp19a1 in bony fish may have originated from an ancestor that evolved alongside amphioxus, rather than directly descending from it. Alternatively, significant chromosomal rearrangements in the region surrounding the cyp19a1 locus could have occurred in the basal vertebrate ancestor closely related to amphioxus [40]. Our findings, however, introduce a new element to this discussion. Our analysis indicates that amphioxus cyp19-like exhibit some synteny with vertebrate cyp19a1. Yet, cyp19-like2 exhibits stronger synteny with humans, mice, and zebrafish. This suggests that cyp19-like2 may have a closer evolutionary affinity with these vertebrate genes. In addition, phylogenetic analysis provides valuable insights into the evolutionary history and potential functions of genes. In the context of cyp19, the finding that cyp19-like1 represents a more primitive and ancestral form suggests that it played a crucial role in the early stages of vertebrate evolution. The proposed origin of cyp19-like2 from genome duplication within amphioxus itself highlights the dynamic nature of genome evolution and the potential for novel functions to arise from such duplications.
In bony fish, the observed differences in tissue expression patterns [44,45], affinities for substrates [46,47], and inducibility by estrogens and xenoestrogens [45,48,49,50] between the cyp19a1a and cyp19a1b are consistent with the idea that duplicated genes can diverge in function. Similar observations in pigs further support this notion [51,52]. Given these findings, it is intriguing to explore whether the two amphioxus cyp19-like exhibit differences in expression patterns, particularly in male and female gonads. Our study using real-time PCR and in situ hybridization techniques has revealed interesting patterns of cyp19-like expression in amphioxus. The high expression of cyp19-like1 in the gills, followed by the ovaries, suggests a role for this gene in reproductive and other functions. In contrast, the predominant expression of cyp19-like2 in the ovaries is consistent with its putative role in sex differentiation. We investigated whether there are differences in cyp19-like expression in the gills of males and females using Real-time PCR and found no sex-specific differences in expression in the gills. The absence of sex-specific differences in cyp19-like1 expression in the gills is noteworthy, indicating that this gene may play a more general role in amphioxus physiology. However, the expression pattern of cyp19-like in the heads of adult amphioxus remains enigmatic and deserves further investigation.
The examination of cyp19 expression during gonad development in amphioxus provides further insights into the potential roles of these genes in sex differentiation and reproduction. The high expression of both cyp19-like at the onset of gonad development suggests their involvement in the initial stages of gonad formation. However, the subsequent downregulation of expression as development progresses might indicate that these genes are not essential for maintaining gonad function but rather play a crucial role in initiating gonad development. Immunohistochemical staining also confirmed that the proteins encoded by these two genes were most abundant in the early stages of gonad development, with subsequent downregulation. Overall, the expression of cyp19-like2 and its encoded protein was more prominent in the gonad primordium. Aromatase proteins are monomeric and anchored within the endoplasmic reticulum via a transmembrane domain at their amino-terminal end [53,54]. We investigated the subcellular localization of Cyp19 and found that the protein is likely localized primarily to microsomal membrane systems such as mitochondrial and endoplasmic reticulum membranes, consistent with its expression pattern in vertebrate cells.
Future studies should aim to further investigate the molecular mechanisms underlying the observed differences in gene expression levels during gonad development and elucidate the precise functions of cyp19-like1 and cyp19-like2 in amphioxus, including their potential roles in sex differentiation, reproduction, and other physiological processes.
5. Conclusions
We comprehensively examined the expression of two cyp19-like genes in amphioxus using techniques such as Real-time PCR, ISH, and IHC. This study clarifies Cyp19 expression patterns during amphioxus gonad development and highlights its importance in vertebrate reproduction evolution. Differences in gene expression and protein localization of cyp19-like1 and cyp19-like2 offer insights into sex determination and gonad development mechanisms. However, further research is needed to fully understand these complex mechanisms.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: The GenBank accession numbers.
Author Contributions
Investigation, methodology, visualization, validation, writing—original draft: Y.W.; visualization, validation, writing—review and editing: J.L., W.L. and G.J.; conceptualization, formal analysis, methodology, visualization, validation, writing—review and editing, project administration, funding acquisition: Z.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the grants of the National Key Research and Development Project of the Ministry of Science and Technology (2023YFE0199500), Shandong Provincial Natural Science Foundation (ZR2022MC032) and Science & Technology Innovation Project of Laoshan Laboratory (LSKJ202203204).
Institutional Review Board Statement
The experimental animal zebrafish follow the ethical guidelines formulated by the Animal Protection and Use Committee of Ocean University of China (permit number, SD2007695).
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data are available from the authors upon request and the corresponding author will be responsible for replying to the request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Koopman, P.; Gubbay, J.; Vivian, N.; et al. Male development of chromosomally female mice transgenic for Sry. Nature 1991, 351, 117–121. [Google Scholar] [CrossRef]
- Wright, E.; Hargrave, M.R.; Christiansen, J.; et al. The Sry-related gene Sox9 is expressed during chondrogenesis in mouse embryos. Nat. Genet. 1995, 9, 15–20. [Google Scholar] [CrossRef]
- Swain, A.; Narvaez, V.; Burgoyne, P.; et al. Dax1 antagonizes Sry action in mammalian sex determination. Nature 1998, 391, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Shinomiya, A.; Kinoshita, M.; et al. DMY gene induces male development in genetically female (XX) medaka fish. Proc. Natl. Acad. Sci. USA 2007, 104, 3865–3870. [Google Scholar] [CrossRef] [PubMed]
- Mar, N.I.; Baker, B.S. The evolutionary dynamics of sex determination. Science (New York, NY) 1998, 281, 1990–1994. [Google Scholar] [CrossRef]
- Raymond, C.S.; Shamu, C.E.; Shen, M.M.; et al. Evidence for evolutionary conservation of sex-determining genes. Nature 1998, 391, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Ottolenghi, C.; Pelosi, E.; Tran, J.; et al. Loss of Wnt4 and Foxl2 leads to female-to-male sex reversal extending to germ cells. Hum. Mol. Genet. 2007, 16, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Dranow, D.B.; Hu, K.; Bird, A.M. Bmp15 Is an Oocyte-Produced Signal Required for Maintenance of the Adult Female Sexual Phenotype in Zebrafish. PLoS Genet. 2016, 12, e1006323. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.I.; Nakaki, F. ZGLP1 is a determinant for the oogenic fate in mice. Science 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, R.; Jiang, J.; Wang, X.; et al. Gonad differentiation in zebrafish is regulated by the canonical Wnt signaling pathway [J]. Biol. Reprod. 2014, 90, 45. [Google Scholar] [CrossRef]
- Holland, L. Z.; Laudet, V.; Schubert, M. The chordate amphioxus: an emerging model organism for developmental biology. Cellular and molecular life sciences: CMLS 2004, 61, 2290–2308. [Google Scholar] [CrossRef]
- Bourlat S J, Juliusdottir T, Lowe C J; et al. Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida. Nature 2006, 444, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; et al. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Putnam, N.H.; Butts, T.; Ferrier, D.E.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 2008, 453, 1064–1071. [Google Scholar] [CrossRef]
- Bertrand, S.; Escriva, H. Evolutionary crossroads in developmental biology: amphioxus. Development 2011, 138, 4819–4830. [Google Scholar] [CrossRef]
- Holland, P. The dawn of amphioxus molecular biology - a personal perspective. Int. J. Dev. Biol. 2017, 61, 585–590. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Henmi, Y. Biology of the amphioxus, Branchiostoma belcheri in the Ariake Sea, Japan II. Reproduction. Zool. Sci. 2003, 20, 907–918. [Google Scholar] [CrossRef]
- Kubokawa, K.; Mizuta, T.; Morisawa, M.; et al. Gonadal state of wild amphioxus populations and spawning success in captive conditions during the breeding period in Japan. Zool. Sci. 2003, 20, 889–895. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, S.; Chu, J. G-banding patterns of the chromosomes of amphioxus Branchiostoma belcheri tsingtauense. Hereditas 2004, 141, 2–7. [Google Scholar] [CrossRef]
- Guraya, S.S. Recent advances in the morphology, histochemistry, and biochemistry of steroid-synthesizing cellular sites in the nonmammalian vertebrate ovary. Int. Rev. Cytol. 1976, 44, 365–409. [Google Scholar]
- Zhang, S.; Wang, C.; Chu, J. C-banding pattern and nucleolar organizer regions of amphioxus Branchiostoma belcheri tsingtauense Tchang et Koo, 1936. Genet. 2004, 121, 101–105. [Google Scholar] [CrossRef]
- Huang, Z.; Xu, L.; Cai, C. Three amphioxus reference genomes reveal gene and chromosome evolution of chordates. Proc. Natl. Acad. Sci. 2023, 120, e2201504120. [Google Scholar] [CrossRef] [PubMed]
- Kubokawa, K.; Tando, Y.; Roy, S. Evolution of the reproductive endocrine system in chordates. Integrative and comparative biology 2010, 50, 53–62. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, L.; Li, H.; et al. Characterization of GRP as a functional neuropeptide in basal chordate amphioxus. Int. J. Biol. Macromol. 2020, 142, 384–94. [Google Scholar] [CrossRef]
- Wang, P.; Wang, M.; Zhang, L.; et al. Functional characterization of an orexin neuropeptide in amphioxus reveals an ancient origin of orexin/orexin receptor system in chordate. Sci. China Life Sci. 2019, 62, 1655–1669. [Google Scholar] [CrossRef]
- Wang, P.; Wang, M.; Ji, G.; et al. Demonstration of a Functional Kisspeptin/Kisspeptin Receptor System in Amphioxus With Implications for Origin of Neuroendocrine Regulation. Endocrinol. 2017, 158, 1461–1473. [Google Scholar] [CrossRef]
- Manikandan, P.; Nagini, S. Cytochrome P450 Structure, Function and Clinical Significance: A Review. Curr. Drug Targets 2018, 19, 38–54. [Google Scholar] [CrossRef]
- Simpson, E.R.; Mahendroo, M.S.; Means, G.D.; et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 1994, 15, 342–355. [Google Scholar]
- Crews, D.; Bergeron, J.M. Role of reductase and aromatase in sex determination in the red-eared slider (Trachemys scripta), a turtle with temperature-dependent sex determination. J. Endocrinol. 1994, 143, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Sakata, N.; Tamori, Y.; Wakahara, M. P450 aromatase expression in the temperature-sensitive sexual differentiation of salamander (Hynobius retardatus) gonads. Int. J. Dev. Biol. 2005, 49, 417–425. [Google Scholar] [CrossRef]
- Kitano, T.; Takamune, K.; Nagahama, Y.; et al. Aromatase inhibitor and 17alpha-methyltestosterone cause sex-reversal from genetical females to phenotypic males and suppression of P450 aromatase gene expression in Japanese flounder (Paralichthys olivaceus). Mol. Reprod. Dev. 2000, 56, 1–5. [Google Scholar] [CrossRef]
- Elbrecht, A.; Smith, R.G. Aromatase enzyme activity and sex determination in chickens. Science (New York, NY) 1992, 255, 467–470. [Google Scholar] [CrossRef]
- Fan, Z.; Zou, Y.; Jiao, S.; et al. Significant association of cyp19a promoter methylation with environmental factors and gonadal differentiation in olive flounder Paralichthys olivaceus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 208, 70–79. [Google Scholar] [CrossRef]
- Lau, S.W.; Zhang, Z.; Qin, M.; et al. Knockout of Zebrafish Ovarian Aromatase Gene (cyp19a1a) by TALEN and CRISPR/Cas9 Leads to All-male Offspring Due to Failed Ovarian Differentiation. Sci. Rep. 2016, 6, 37357. [Google Scholar] [CrossRef]
- Nakamoto, M.; Shibata, Y.; Ohno, K.; et al. Ovarian aromatase loss-of-function mutant medaka undergo ovary degeneration and partial female-to-male sex reversal after puberty. Mol. Cell. Endocrinol. 2018, 460, 104–122. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Liu, Z.; et al. Epigenetic modifications during sex change repress gonadotropin stimulation of cyp19a1a in a teleost ricefield eel (Monopterus albus). Endocrinology 2013, 154, 2881–2890. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Tang, H.; Liu, Y.; et al. Targeted Disruption of Aromatase Reveals Dual Functions of cyp19a1a During Sex Differentiation in Zebrafish. Endocrinology 2017, 158, 3030–3041. [Google Scholar] [CrossRef] [PubMed]
- B Hne, A.; Heule, C.; Boileau, N.; et al. Expression and sequence evolution of aromatase cyp19a1 and other sexual development genes in East African cichlid fishes. Mol. Biol. Evol. 2013, 30, 2268–2285. [Google Scholar] [CrossRef] [PubMed]
- Ramallo, M.R.; Morandini, L.; Birba, A.; et al. From molecule to behavior: Brain aromatase (cyp19a1b) characterization, expression analysis and its relation with social status and male agonistic behavior in a Neotropical cichlid fish. Horm. Behav. 2017, 89, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.; Lu, H.; et al. Genes encoding aromatases in teleosts: evolution and expression regulation. Gen. Comp. Endocrinol. 2014, 205, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.F.; Santos, M.M.; Reis-Henriques, M.A. The genomic environment around the Aromatase gene: evolutionary insights. BMC Evol. Biol. 2005, 5, 43. [Google Scholar] [CrossRef]
- Mizuta, T.; Kubokawa, K. Presence of Sex Steroids and Cytochrome P450 Genes in Amphioxus. Endocrinology 2007, 148, 3554–3565. [Google Scholar] [CrossRef]
- Baker, M.E. Co-evolution of steroidogenic and steroid-inactivating enzymes and adrenal and sex steroid receptors. Mol. Cell. Endocrinol. 2004, 215, 55–62. [Google Scholar] [CrossRef]
- Tchoudakova, A.; Kishida, M.; Wood, E.; et al. Promoter characteristics of two cyp19 genes differentially expressed in the brain and ovary of teleost fish. J. Steroid Biochem. Mol. Biol. 2001, 78, 427–439. [Google Scholar] [CrossRef]
- Kishida, M.; Mclellan, M.; Miranda, J.A.; et al. Estrogen and xenoestrogens upregulate the brain aromatase isoform (P450aromB) and perturb markers of early development in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 129, 261–8. [Google Scholar] [CrossRef]
- Zhao, J.; Mak, P.; Tchoudakova, A.; et al. Different catalytic properties and inhibitor responses of the goldfish brain and ovary aromatase isozymes. Gen. Comp. Endocrinol. 2001, 123, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.K.; Chiang, E.F.; Hsiao, P.H.; et al. Phylogeny, expression and enzyme activity of zebrafish cyp19 (P450 aromatase) genes. J. Steroid Biochem. Mol. Biol. 2001, 79, 299–303. [Google Scholar] [CrossRef]
- Kishida, M.; Callard, G. V. Distinct cytochrome P450 aromatase isoforms in zebrafish (Danio rerio) brain and ovary are differentially programmed and estrogen regulated during early development. Endocrinology 2001. [Google Scholar]
- Gelinas, D.; Pitoc, G.A.; Callard, G.V. Isolation of a goldfish brain cytochrome P450 aromatase cDNA: mRNA expression during the seasonal cycle and after steroid treatment. Mol. Cell. Endocrinol. 1998, 138, 81–93. [Google Scholar] [CrossRef]
- Tchoudakova, A.; Callard, G.V. Identification of multiple CYP19 genes encoding different cytochrome P450 aromatase isozymes in brain and ovary. Endocrinology 1998, 139, 2179–2189. [Google Scholar] [CrossRef] [PubMed]
- Conley, A.J.; Corbin, C.J.; Hughes, A.L. Adaptive evolution of mammalian aromatases: lessons from Suiformes. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2009, 311, 346–357. [Google Scholar] [CrossRef]
- Corbin, C.J.; Hughes, A.L.; Heffelfinger, J.R.; et al. Evolution of suiform aromatases: ancestral duplication with conservation of tissue-specific expression in the collared peccary (Pecari tayassu). J. Mol. Evol. 2007, 65, 403–412. [Google Scholar] [CrossRef]
- Amarneh, B.; Corbin, C.J.; Peterson, J.A.; et al. Functional domains of human aromatase cytochrome P450 characterized by linear alignment and site-directed mutagenesis. Mol. Endocrinol. 1993, 12, 1617. [Google Scholar]
- Shimozawa, O.; Sakaguchi, M.; Ogawa, H.; et al. Core glycosylation of cytochrome P-450(arom). Evidence for localization of N terminus of microsomal cytochrome P-450 in the lumen. J. Biol. Chem. 1993, 268, 21399–21402. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



