
Submitted:
12 February 2024
Posted:
13 February 2024
You are already at the latest version
Abstract
One of the most concerning global environmental issues is the pollution of agricultural soils by heavy metals (HMs), especially cadmium (Cd), which not only affects human health by taking Cd-containing foods but also impacts the quality of rice. The soil's nitrification and denitrification processes, coupled with the release of volatile organic compounds by plants, raise substantial concerns. The understanding of three aspects of paddy soil under conditions of heavy-metal pollution and water management is currently limited. In this review, we summarized recent literature related to the deleterious effects of Cd on both soil processes related to the N cycle and rice quality, particularly aroma, in different water management practices. Under both continuous flooding (CF) and alternate wetting and drying (AWD) conditions, cadmium (Cd) has been observed to reduce both nitrification and denitrification processes. The adverse effects are more pronounced in continuous flooding (CF) compared to alternate wetting and drying (AWD). Similarly, the alteration in rice aroma is more significant in AWD than in CF. The precise modulation of volatile organic compounds (VOCs) by Cd, remains unclear based on the available literature. Nevertheless, HM accumulation is higher in AWD conditions compared to CF, leading to a detrimental impact on volatile organic compounds (VOCs). The literature concluded that AWD practices should be avoided in Cd contaminated fields to decrease accumulation and maintain the quality of rice. In the future, rhizospheric engineering and plant biotechnology can be used to decrease the transport of HMs from the soil to the plant’s edible parts.
Keywords:
Heavy metals
; volatile organic compounds
; N cycle
; Paddy soil
; Water management
Rice serves as a staple food for half of the global population, with numerous Asian and African nations, such as Madagascar and Liberia, exhibiting annual per capita rice consumption exceeding 100 kg. In Africa, the consumption of rice is going faster as compared to other commodities because of the growing urban population [2]. In 2023, about 523.1 million tons of rice were produced globally [3]. The majority of rice production comes from Asian countries, with significant contributions from Indonesia, Vietnam, Thailand, Myanmar, China, Bangladesh, India, the Philippines, and Pakistan, as reported by the FAO in 2023. Western and Eastern Asia together account for over 90% of both global rice production and consumption. China’s role in worldwide rice production stands at 41% [3]. This accounts for approximately 23% of China’s total cultivated land[3,4].
Water is a crucial element for the growth and development of rice plants. Rice grown throughout the globe by different water management, like continuous flooding among them is the one that is count for the unproductive water loss by seepage, evaporation, and percolation [5,6,7]. Recent studies showed that continuous flooding conditions are optional for high yields in short-term trails. However, few long-term field studies have been conducted which provide a little information about crop yield during water-saving practices in the fields [5]. Because freshwater is distributed unevenly in terms of both space and time, farmers strive to conserve as much rainwater as they can. Farmers utilize this water for irrigating rice fields when the soil reaches a specific dryness threshold. Currently, the following rice irrigation modes are in use throughout China: Continuous flooding, midseason flooding with intermittent drying, midseason flooding with intermittent irrigation, midseason flooding with rain-fed conditions. The most popular method is flooding-midseason drying-flooding [5].
Nitrogen (N) cycling involves both biotic and abiotic transformations of nitrogen in paddy soils. These transformations encompass processes such as ammonification, N immobilization, nitrification, denitrification, anaerobic ammonium oxidation (anammox), dissimilatory nitrate reduction to ammonium (DNRA), and nitrogen fixation. In paddy soils, the importance of nitrification and denitrification are well documented and a recent addition to this is archaeal ammonium oxidation [8,9,10]. Reports indicate that the ammonia oxidization to nitrite, carried out by microorganisms encoding the enzymes of ammonia-oxidizing bacteria [11], serves as the initial rate-limiting step in nitrification) and/or ammonia-oxidizing archaea [12,13,14]. Nitrifiers microorganisms are distributed in few functional taxa while denitrifies bacteria are broadly distributed in many taxa which are responsible for the nitrate as an alternative electron acceptor for respiration [15].
Plants interact with plants and other living communities. Therefore, they have developed a variety of strategies. One of them is the diffusion of volatile organic compounds (VOCs); these signals provoke the defense mechanism against biotic and abiotic stresses and several other environmental factors [15]. Angiosperm and gymnosperms release a number of VOCs, such as fatty acids, amino acid derivatives, phenylpropanoids/benenoids, and terpenoids, which are most common [16,17,18]. Through priming, neighbors, plants developed stress resistance due to VOCs of the stress-exposed plants [19]. This phenomenon was established between tobacco (Nicotiana attenuate) and sagebrush (Artemisia tridentata) plants [20]. The former was exposed to the VOCs of the later clipped one. The clipped sagebrush mimics insect damage and tobacco-developed herbivore resistance.
Soil acts as a seed bank of microbes, and plant roots recruit the microbes from the soil. The presence of heavy metals in the soil leads to changes in both soil biomass and microbial community structure. In the rhizosphere of contaminated plants, roots exhibit a higher prevalence of heavy metal-resistant bacteria but a lower overall microbial population compared to the rhizosphere of unpolluted plants [21,22,23,24]. Within the soil, cadmium [1] induces alterations in microbial community structure, reduces microbial carbon use efficiency, and results in an increase in the microbial C: N ratio [25]. Recent studies in metals-polluted soil have focused on the microbiome of model plants such as Arabidopsis thaliana, metal accumulating or metal tolerant and hyper-accumulating plants [26,27,28]. Both, biotic and abiotic factors shape microbial community assembly in the plant rhizosphere [29]. The ecology of the rice microbial community under metal stress, particularly Cd stress, is the least studied area. The majority of metal-contaminated soil-grown plant rhizosphere consists of metal-resistant bacteria that can maintain metabolic functions [30,31]. Thiobacillus, Pedobacter, and Geobacter have been reported as high metal resistant bacteria genera responsible for Fe (III) reduction in the Cd contaminated rice rhizosphere [32,33,34]. However, some PGPRs such as Dyella and Novosphingobium colonized the rice rhizosphere [34]. Furthermore, the role of VOCs role in soil microbial N transformation also needs to be discussed in detail during HM stress.
Nitrification, denitrification, VOCs, and plant endophytes are very important factors in terms of plant health. In this review, we will focus on Cd detrimental effects on the N transformation, VOCs of rice, and endophytes under different water management practices.
2. Different Water Management Practices for Rice Cultivation
Globally, about 4 billion people are suffering from water scarcity [35]. Furthermore, for sustainable rice production, a shortage of water is a major threat. In order to ensure food security, alternative ways such as less water for rice cultivation are very important. Therefore, in modern agriculture practices water-saving irrigation for rice cultivation must be the point of concentration in future research. Various approaches and technologies can be taken into account, such as alternate wetting and drying (AWD), reducing the outflow of unproductive water, adopting direct seeding, implementing the rice intensification system, employing the rice aerobic system, embracing the ground cover rice production system, and utilizing tensiometers for scheduled irrigation. These technologies can improve water use efficiency and significantly decrease the demand for water for the production of rice.
2.1. Continuous Flooding (CF)
Warm and waterlogged soil is used for rice cultivation. Continuous flooding (CF) is a traditional rice-growing practice in which the rice paddies are flooded by the farmers throughout the rice-growing season. Globally, 70% of the rice cultivation is carried out under CF. CF based rice cultivation is highly productive but some problems related to soil health and environment have been reported. Soil degradation, elevated methane emissions, and the substantial accumulation of harmful substances like arsenic and mercury are primarily associated with rice cultivation using chemical fertilizer-based practices [36].
Lowland rice irrigation covers about 79 million hectares worldwide, with an estimated 75% world’s rice production. The global crop irrigation area covered in Asia is approximately 56%, where 40-46% is rice cultivation [37]. About 64-83% of the irrigated area in the Southeast, 46-52% in the East, and 30-35% in the South Asia are occupied by rice cultivation. In comparison to other irrigated crops in the field, rice needs 2-3 times more water [38]. From developed 70% freshwater resources worldwide, rice irrigated receives 24-30% share.
Several factors, such as global warming, rice competition with other seasonal crops, industries, and civic requirements for water, may cause economic or physical water shortages for rice cultivation [39]. In the future, several million hectares of lowland rice irrigated systems will face water shortage by 2050 [5]. In account of all the issues related to the CF rice irrigation system, the development of water-saving irrigation system is compulsory for sustainable rice cultivation [5]. The International Rice Research Institute (IRRI) introduced alternate wetting and drying (AWD) water management technology, among other water-saving rice irrigation systems, to deal with expected water shortages in the future. AWD has got worldwide attention and several research studies are available that are becoming increasingly widespread [40].
2.2. Alternate Wetting and Drying
Developing agronomic practices that minimize water usage for irrigation while sustaining high yields is essential to support the increasing global population. To address the issue of water scarcity in rice agricultural production, alternate wetting and drying (AWD) water management practices are being adopted [36,41]. Under AWD conditions, rice fields are drained out for a certain period in which the field obtains a certain level of moisture and is then re-flooded. It has been reported that using AWD water technology saves about 23% of the water used for rice irrigation when compared to CF practices [6,38]. AWD technology has the potential to reduce greenhouse gas (GHG) emissions, particularly methane (CH4) gas [36,42]. Alternate wetting and drying conditions decreased by about 45-90% global warming potential (GWP − CH4 + N2O) compared to CF. Human activities generate about 48Gt CO2 annually, which can increase global warming. Therefore, to reduce GHG emissions from paddy fields, it is critical to promote AWD adaptation globally [43]. In the USA, for example, farmers received carbon credits under the project “Voluntary Emission Reductions in Rice Management Systems” involves farmers adopting practices aimed at minimizing greenhouse gas (GHG) emissions, with AWD being one practice [44]. On the other hand, AWD also reduced arsenic (As) accumulation in rice grains [41,45] and methylmercury concentration in soils, and lowered the consumption of energy/fuel where pumping is in practice for irrigation [36,46,47]. Many efforts have been made to promote AWD in Asia because to its benefits [48]. In China, the “mid-season” drain is widely adopted which is somewhat similar to AWD [49]. However, global AWD technology adoption is limited due to a variety of factors limiting rice productivity [36,46,50].
Since the 1990s, China has embraced water-saving irrigation methods such as thin-shallow-wet-dry irrigation [51] and alternate drying-wetting [52,53]. Soil water management strategies influence oxygen transfer, nitrogen transformation, and soil redox potential [9,10,54]. Soil microbial biomass carbon (MCB) and nitrogen (MBN) have been significantly affected by irrigation practices [55,56]. In paddy soils, MBC and MBN decreased in continuous flooding irrigation as compared to intermitted irrigation [57]. Other studies have reported high N2O emissions from paddy fields with intermittent irrigation and midseason drainage [58,59].
Soil moisture is very important during the rice grain filling stage. Grain quality and yield are both impacted by water levels during rice reproduction and the filling stage [60,61]. Both CF and AWD effectively enhanced the quality of rice grains [62]. For example, AWD improved grain head recovery, grain weight, and protein contents [63,64]. Similarly, rice kernel opaqueness, abortiveness, and chalkiness were reduced significantly in AWD [60,64]. In addition, AWD significantly increased the nutritional value of the rice grain such as high antioxidants, total tocopherols, γ-oryzanol, flavonoids, zinc and iron [65]. Furthermore, AWD significantly decreased arsenic in the rice grain [66].
2.3. Popular Methods around the Globe
Globally, rice production and greenhouse gas production are under the influence of two important factors i.e., water management and application of nitrogen fertilizer. China is one of the largest rice-producing countries; water shortage threatens rice cultivation. The most popular rice cultivation method is CF. However, due to water shortage and greenhouse gas emissions, nowadays different water-saving irrigation systems are in practice around the globe (Table 1). Among different water-saving irrigation, AWD is the most practiced technology for rice production.
However, different studies reported AWD decreased CH4 and increased N2O emissions. Moreover, while applying medium nitrogen (N), the optimal rice grain yield has been observed in the case of AWD in contrast to the conventional flooding-midseason drainage-flooding irrigation (PFD) approach. Different studies from Bangladesh, the Philippines, China, and Pakistan reported a 4.52% average increase in rice yield (t/ha) under AWD than CF. Similarly, water use efficiency (WUE) was also increased in AWD as compared to CF.
Another rice production system termed as [46] applied in different regions of the world. In ARS, rice is grown under non-saturated, non-flooded, and non-puddled soil conditions [67]. The yield obtained from ARS was higher than in upland conditions but lower than in flooded conditions. In cool temperate regions, ARS is a successful system for rice production. However, a partial aerobic rice system (PARS) is more suitable alternative for ARS in warm humid regions. There have been reports that ARS reduces rice yield over time, and it has been proposed that this technique may not be ideal for long-term rice agriculture [68]. Several factors among them are Zn, Fe, K, N, and P deficiencies, root-knot nematodes and weeds are responsible for the yield reduction in ARS [69]. However, it save about 50% of irrigation water compared to CF.

Table 1.
Different strategies used for Water-Saving irrigation of Rice Production.
| S. No | Technologies for Conserving Water in Rice Cultivation. | Reference |
|---|---|---|
| 1 |
|
[70,71,72] |
|
[68] | |
|
[73] | |
| 2 |
|
[10,74] |
|
[36,75] | |
|
[76,77] | |
|
||
|
[73] | |
|
[78] | |
|
[73] | |
|
[79] | |
|
[73] | |
|
[1] |
2.4. Heavy Metal Pollution in Paddy Soil
Cadmium [1] is considered one of the hazardous heavy metals that permeates the soil through various sources [9]. Its pollution is a severe global issue since it can build in agricultural soil and edible sections of crops, and eating those contaminated components can cause cancer in humans [10]. Various natural and anthropogenic sources like rock weathering, atmospheric deposition, urbanization, and wastewater irrigation, sewage sludge application, overuse of phosphate fertilization, mining, and smelting originate Cd contamination in the soil [80]. Geological parent material decides the background of Cd level. However, generally it has been reported a range from (0.06-1.1 mg kg-1) with a 0.01 mg kg-1 lower and 2.7 mg kg-1 higher value [80]. However, due to human activities, the Cd level (up to 14 mg kg-1) has been reported in several regions of the world. Several factors such as cultivars, crop species, and soil properties contribute to the accumulation of Cd in plants.
2.5. Sources
2.5.1. Natural Ways of Cd Origination (Geogenic)
Naturally, Cd is existing in all rocks in entire soils. Among the top 20 priority hazardous metals, Cd ranked 7th [81]. Different rocks have different Cd concentrations. For example sedimentary deposits rocks have higher Cd than igneous and metamorphic rocks [82]. Thus, the soils originating from sedimentary phosphatic rocks have more Cd due to high Cd concentration [83,84]. Where the soils develop on phosphorites, shales or carbonates then the geogenic soil Cd concentration range from 0.01-10 or 100 mg kg-1 [85,86]. Cd exhibits weak sorption to the surfaces of both organic matter and minerals in the soil. Cadmium from mineral lattices releases slowly by weathering of high-Cd parent rocks [87].
Soil Cd is hard to distinguish either via geogenic or anthropogenic origin. Generally, Cd enriched topsoil is assumed to occur via farm and atmosphere inputs. Nevertheless, this is not the case, as Cd is transported to subsoils through the biological cycle [88]. Furthermore, via vegetation, Cd in agricultural brings from subsoils to topsoil [89]. At a continental scale, the concentration of Cd in European soil is primarily influenced by geological factors rather than anthropogenic inputs, despite the widespread use of intensive fertilizers, urban wastes, and manures in agriculture [90].
2.5.2. Anthropogenic Sources of Cd Input
Human activities are the leading cause of worldwide Cd deposition in the environment, accounting for 13, 000 tons of the 30, 000 tons of Cd inclined to the environment each year [10,91]. In the agriculture soils, Cd had been introduced by various sources and its concentration is generally lower than 0.2 mg kg-1. The level of Cd in the agriculture soils increased due to long-term phosphate fertilizer application along with sewage water used for irrigation purposes [92]. Discharged wastewater, mining activities (exhaust and solid waste), the smelting sector, and atmospheric deposition, on the other hand, all contribute to Cd contamination of agricultural soil. Blast furnaces sludge and flue dust contain a relatively high content of Cd [93]. The oxidizable and residual fraction of textile dyeing plants, sludge, has been reported to consist of an average 3.72 mg kg-1 Cd [94]. In Changsha City which is the capital of Hunan Province of China, dust analysis showed that 50% Cd was insoluble and reducible fractions in PM 2.5 from re-suspension of dust, fuel combustion, vehicular emissions, and other pollution sources, speciation analysis during spring [95]. Nevertheless, the predominant source of Cd input into the soil arises from agricultural activities, such as the irrigation of wastewater. Unnecessary phosphate and organic fertilizer applications, and random disposal of untreated urban sewage sludge and garbage [96]. The issue of Cd input via phosphate fertilizer application in agriculture soils of China is controllable because naturally phosphate and Cd contents are low in the native phosphate rocks of China, which lead to import high-quality phosphate fertilizers which contain a high level of Cd [97]. The Cd content was about 7.5-156 mg kg-1, 84-144 mg kg-1, 9.5 mg kg-1, and 24.5 mg kg-1 in imported ammonium phosphate, single superphosphate, normal superphosphate, and triple-superphosphate, respectively, which exceeded the standards [97]. Atmospheric deposition of Cd in paddy soil have been reported about 65-70% [98]. Similarly, fertilizers, particularly phosphate fertilizers and biosolid contribute 10-50% each to Cd accumulation in paddy field [99]. However, in China, animal manure about 55% contributed to total soil Cd inputs [100].
2.6. Chinese Paddy Fields and Heavy Metal (HM)
In china, paddy soil Cd contamination is a widespread problem. The Cd paddy soil issue is more sever in some mining and industrial regions of China. The range of paddy soil Cd contamination range from 0.01-5.50 mg kg-1, with an average value of 0.23 mg kg-1 [101]. Hunan was reported with highest Cd (0.73 mg kg-1), whereas Guangxi and Sichuan provinces were recorded 0.70 and 0.46 mg kg-1 paddy soil Cd. However, in 1990s, the background value of Cd in Chinese paddy soil was 0.1 mg kg-1 [98]. Paddy Cd contamination across China following the trend like Northwest > South central > East > Northeast > North [98]. In China, paddy fields are mostly contaminated by Cd due to the smelting of the ores of Cu, Pb, and zinc [91,102]. Cd contents of rice grains are greatly varying. Rice grains sampled from the field and market were reported to have raised levels of Cd, up to 7.0 and 1.24 mg kg-1, respectively [102]. A study containing 100 rice grain samples in Hunan, one of the large nonferrous metals production zones, reported only 15% of rice grains meet the standard level of As, Cd and Pb [103]. Another study randomly collected 63 rice grain samples from the open markets of Jiangxi and Anhui, along with 7 samples from Guangdong., Hunan, and Jiangxi, and Cd level in about 70% of grain samples was found to be higher than the standard 0.2 mg kg-1 [104].
3. HM and Soil N Transformation
Rice plants prefer ammonium to nitrate. When both ammonium and nitrate are present, rice seedlings exhibit a faster uptake of NH4+ compared to NO3− [105]. From last half-century, due to the NH4+ preference of the rice, farmers used to cultivate rice in paddy fields, ammonium-based fertilizers like ammonium sulfate and urea are applied [105]. In addition, organic fertilizers for rice cultivation are also commonly applied in the fields, in which the organic N is converted to ammonium through a biological process called ammonification (mineralization of N). Due to the low level of oxygen or depletion in paddy soils, the rate of ammonification is slower compared with upland soils [106]. Depletion of O2 in soils not only slow down the ammonification but also limits the process of ammonium oxidation (NH4+ to NO3-) and immobilization of N (microbial assimilation of NH4+ i.e., the conversion of inorganic N to organic N), which lead to the accumulation of NH4+ in soils [107].
3.1. The Significance of Nitrification and Denitrification Processes in Paddy Soils
The most limiting nutrient for rice production is generally known as N. Denitrification in paddy soils was initially studied in order to make the efficient use of fertilizers and link the gaseous products such as NO, N2O, and N2 with the microbial process involved in the reduction of nitrate [108]. The thin oxidized layer of paddy soils is the region where nitrification occurs while denitrification takes place below the oxidized layer [109]. The loss of N from paddy soils included ammonia volatilization, nitrate leaching and nitrification [110]. The denitrification occurred just following nitrification in paddy soils. It is proposed that ammonium fertilizers should be deeply applied before waterlogging (4-5 days) in paddy fields to overcome the N loss from the paddies [111]. N transformation on a microscale in the flooded soil surface layer reported a high number of nitrifiers and denitrifiers bacteria in the above 2 cm layer than the below-reduced layer of paddy soil [11,112]. After one to two weeks of waterlogging low-molecular-weight organic molecules (acetate) accumulated in the top layer which increased the denitrifiers bacteria. The correlation of organic carbon and denitrifiers bacteria in another environment along with lake sediments and river waters also have been reported [113]. The most important factor for the controlling of denitrification is the availability of organic C since it provides electrons donor for the denitrifiers. Rice plant roots release O2 in the rhizosphere, therefore nitrification and denitrification activities were also reported for rice rhizosphere [114,115] and green algae which grown in rice fields [116], as well as in the bulk soil between the oxidized and reduced layers. The process of nitrification-denitrification in rice paddy soil may cause the loss of N from the applied fertilizer. Globally, 40% of the total greenhouse gas emission recorded from paddy soils. Different studies have been reported the little emission of N2O from the paddy soil under control condition than the upland field crops [117,118]. Denitrification is a strong activity of rice paddy soils, as a result, little emission of N2O occurred by conversion of N2O into N2 [119]. Furthermore, several studies on dissolved N2O in the rice surface and groundwater have been conducted [120]. These findings suggested that little emission of N2O gas from rice paddy soil occurred due to the N2O reducers, present in the rice paddy fields [121]. However, recent findings suggested that iron plague in the rhizosphere contribute largely to N2O emission that was previously ignored [122].
3.2. Nitrification
Nitrification is a totally microbial mediated process in which nitrate (NO3−) formation takes place by ammonium (NH4+) oxidation via nitrite (NO2−) [9,123,124] (Figure 1). This is the energy acquiring process for autotrophic nitrifiers. The process in which NO3- or NO2- formation takes place from inorganic N (NH4+) and organic N without energy acquired by microbes is known as heterotrophic nitrification and the microbes termed as heterotrophs [123].
Nitrification has been divided into two steps: the oxidation of ammonia (NH4+→NO2−) and nitrite oxidation (NO2−→NO3−). Nitrosospira spp. and Nitrosomonas spp. of Betaproteobacteria and Nitrosococcus spp. of Gamma proteobacteria, these bacteria are known as ammonia-oxidizing bacteria [11] can carry out the oxidation of ammonia. Archaea, along with bacteria can also take place in ammonia oxidation which is termed as ammonia-oxidizing archaea [12] [125,126]. Different studies have been reported a higher number of AOA than AOB in the ocean [127]. A positive correlation of AOB abundance was observed with rice paddy nitrification activity soil which indicates that AOB could be the major players of nitrification than AOA [128]. However, with the application of urea fertilization increased the abundances of both AOA and AOB in the rice rhizosphere [129,130]. Recent reports showed, comammox relative contribution to nitrification is higher than AOA and AOB in the rice paddy soil [128].
The diversity of ammonia-oxidizing bacteria [11] communities has been investigated in rice paddy soils [131]. The rice genotypes can also influence the diversity of AOB [132]. In China, rice paddy soil the Nitrosospira spp. are the most dominated AOB [133,134,135], while biochar amended rice paddy soil favored amoA gene containing AOB just related to Nitrosomonas spp., [136]. Among the other factors, fertilizer practices may change the AOB community in the soil because of the environments with high ammonia concentration (like wastewater) Nitrosomonas spp. predominates while in the soil Nitrosospira spp. is the major AOB [137]. Nitrite-oxidizing-bacteria (NOB) form the genera of Nitrobacter, Nitrospina, Nitrococcus, and Nitrospira perform the nitrite oxidation which is the second step of nitrification [138,139].
3.3. Denitrification
Denitrification is a process in which gaseous form of N (NO, N2O and N2) forms via the stepwise reduction of (NO3- and NO2-) by microbial respiration (Figure 1). Microbes utilize nitrogen oxides as electron acceptors in lieu of oxygen when operating under anaerobic conditions for respiration.
Nitrous oxide also acts as an electron acceptor but non-denitrifiers reduce N2O [140]. Two types of nitrite reductases are responsible for denitrification i.e copper-containing NirK encoded by gene nirK, and NirS cytochrome cd1-containing enzyme encoded by nirS gene [12]. These enzymes transform oxidized ammonia to NO, and N2O. Finally nitrous oxide reductase enzymes encoded by nosZ, reduces N2O into N2. Nitrite reductases (nirK and nirS) gene related community has been studied in detail [12,141]. In the rice paddy soil, the average gene copy number of nirK, nirS, and nosZ were 7.65×10-7, 3×10-7, and 8.71×10-7, respectively [9] 16S sequencing approaches in paddy soil, revealed that denitrifiers community genes are distributed in Betaprotobacteria. The phylum was further divided in to Azospira and Burkholderiales, belonging to the genus Herbaspirillum, and Rhodocyclales in the rice paddy soil under denitrifying conditions [142].
The soil type, plant genome, pH, Eh, organic matter, temperature, substrate, and fertilizers affects the response of microbes towards denitrification. For example, with increasing temperature, increases the denitrification rate in paddy soil [143,144]. Similarly, high moisture also increased denitrification [144]. However, opposite is true for N fertilizer. N fertilizers increased the nirK gene abundances and decreased nirS in paddy field [141].
3.4. Production of N2O
Nitrous oxides production occurs through different sources in the soil, incomplete denitrification can lead to the production of N2O (NO3−→NO2− →NO →N2O →N2). The lack of N2O reducing enzymes in the denitrifiers can also lead to the production of N2O [145], one such example is the denitrification activities of fungus, in which N2O is the major end product. Not only denitrifiers are the only source of N2O, but some ammonia oxidizers are also the source of N2O production either through hydroxylamine oxidation or nitrifier denitrification [146]. Both AOA and AOB have been reported for the nitrifier denitrification because they are positive for nitrite reductase gene (nirK). These microbes first perform the activity of ammonium oxidation then oxygen-depleted environment converts the product into N2O and N2 which is termed as nitrifier denitrification or the reduction of nitrite [147]. One of the intermediate products of ammonia oxidation is hydroxylamine, during the oxidation of hydroxylamine, N2O produced as a byproduct. A study reported both AOA and AOB for the N2O is production via the hydroxylamine oxidation pathway. In addition, dissimilatory nitrate reduction to ammonium (DNRA) bacteria has been reported for N2O [148]. Non-denitrifiers can also produce N2O by the reduction of nitric oxide as detoxification of toxic NO molecule. In this process, quinol-type nitric oxide reductase (qNor) has been reported instead of denitrifiers containing cytochrome c-type nitric oxide reductase (cNor) [140,149]. Chemodenitrification has also been reported in which nitrite is reduced to NO and N2O [150].
3.5. Reduction of N2O
Though different way N2O produced in the soil can be used as electron acceptors by different N2O reducer’s microbes for respiration which contains the N2O reductase (nosZ) [151,152]. The ability of N2O reduction into N2 had been reported in a phylogenetically diverse group of bacteria and archaea. This reduction of N2O is the part of denitrification but some denitrifiers lack the enzymes to reduce it into N2 gas. In addition, the reduction of N2O can also be done by some DNRA bacteria in the rice paddy soil [153]. Both culture-dependent (FSC) and culture-independent [90] analyses were used for the detection of N2O reducers in the rice paddy soil [154,155]. Herbaspirillum sp. strains were grown under the conditions which exist during rice paddy soil denitrification and were reported as the active player of N2O reduction step of the denitrification [156]. Herbaspirillum and Azospirillumcan had been reported as denitrifiers recently, depending on the environmental condition these bacteria may be acting like N fixers or N losers from the soil just like legume symbiont Bradyrhizobium which perform both processes (N fixation and denitrification) [157,158].
3.6. Effect of Cd on N Cycle in Paddy Soils
For bioavailability, different definitions exist but it can be defined as the chemical availability to the living receptors either directly or through uptake (Figure 1). The main concern during metal contamination in the soil is the bio-available portion of metal not the total metal concentration because only the bio-available portion uptake is possible for plants from the soil [159]. The most important factors which control the availability of the metals in the soil are pH and organic matter [160,161]. In the rhizosphere of the plant, heavy metals bioavailability depends on soil type, plant genotype, metals speciation, and the change in biological and physicochemical of the soil induced by roots [162,163]. Spatially and temporally rhizosphere varies which acts as a dynamic habitat [164]. The rhizosphere is the most complex hot spot which is comprised of a diverse plant-associated microbial community, responsible for the metal mobilization and immobilization in the soil either directly or indirectly [165,166]. The mobilization of metals occurs through the release of chelators or redox changes done by the microbial processes in the rhizosphere such as chelation, chemical transformation, and soil pH reduction via protonation, while the immobilization of metals occurs in the rhizosphere by biosorption or bioprecipitation via microbes [167,168]. In a recent study reported higher metals-tolerant bacteria in the rhizosphere microbial communities [169].
3.6.1. Nitrification
Soil physical and chemical properties of soil influence the microbial-mediated processes rates in the field environments [170,171,172]. Nitrification and denitrification processes are not only affected by the soil pH but a recent study reported that soil NH4+-N and C/N also responsible while changing the abundances of soil microbial communities [173]. The effect of metals on ammonia oxidation depends upon the specific metal or combination of metals in the environments like no change in the abundance of AOA was reported in mercury-spiked soil used for vegetable cultivation while Cu spiked up to 1000 mg kg-1 changed the AOA community in the grassland and arable soils [174,175]. A study has reported that Cu was more toxic to the ammonia oxidizer community as compared to Ni. For example, Cu strongly affects the AOB community in the sludge-amended soil and adaptability was more common to Zn and Ni [176]. However, general microbial community was shown more vulnerable to Cu and Cr contamination as compared to Zn and Ni as well [177]. Heavy metals pollution had a negative effect on the abundance, diversity, activity, and composition of AOA and AOB in the natural soil [10]. Another study reported a higher copy number of AOA than AOB in the rice paddy field, contaminated with heavy metals and the ratio of AOA to AOB copy number was in the range of 2.5 to 59 [175].
3.6.2. Denitrification
Denitrification is a heavy metal sensitive process and the heavy metals concentration in the soil directly proportional to its inhibition [178]. Heavy metals change the denitrifiers in the soil which reflect the inhibition of denitrification (Figure 1). A recent study reported a significant change in the composition of the denitrifiers in the metals polluted paddy soil via denaturing gradient gel electrophoresis (DGGE) [179]. In addition, in the rice paddy soil, the microbial community has been reported to adopt the metal stress by changing the abundances of certain microbial taxa. Not only the abundance but also the functional potential has been reported altered in the metals polluted paddy soil. The replacement process is the main factor for the adaptability of heavy metals of the microorganisms while soil pH variation is totally responsible for the community richness[180]. The negative effect of metals on the abundance of denitrifiers was also varied with the elements. The copy of nirK and nosZ genes sharply decreased with the spiked Cu in a 6 days lab incubation experiments but a recovery in copy number of the denitrifiers genes has been reported for long time incubation [181]. The heavy metals pollution especially the Cd decreased the abundance, diversity, and structure of both nitrifiers and denitrifiers in the rice paddy soil [182].
3.6.3. Greenhouse Gases from Paddy Fields under Metals Contamination
The application of N fertilizers is essential for rice production with high yield. However, due to the overuse of N fertilizers, the production of GHG such as nitrous oxide (N2O) is one of the major concerns [183,184]. Different studies have addressed these problems by specifying which soil types are more prone to N losses by describing many environmental and management factors linking to biological nitrogen transformation [185,186]. However, the question, how heavy metals pollution could affect the biological transformation in the soil is still on the list to answer, especially, the introduction of heavy metals through atmospheric deposition versus spiked metals into the soil which suddenly increases the heavy metals concentration in the soil [176,187]. Another issue is the linking between changes in N transformation genes abundances and heavy metals contamination in soils. It will become more difficult to address this issue in rice paddy soils where significant changes occur in the redox potential due to wet/dry cycles which affect the heavy metals bioavailability, structures of microbial community and rate of biological transformations of N [180,188].
Among all the green GHG, N2O is the most radiative GHG increasing 0.26% in the atmosphere per year [180]. Incomplete denitrification results in the production of nitrous oxide in which the last reduction step fails to reduce the N2O into N2 in the soil. It had been reported that about 60% of the total N2O globally is produced by anthropogenic sources with the major players is known as paddy fields [189,190]. China’s paddy fields produced up to 29.0 Gg of N2O emissions which are about 7-11% of total GHG emission from China’s croplands [51,191]. Recent studies have shown that in metal-polluted paddy soils, microbial biomass and fungal to bacterial ratio are declined with an increase in the concentration of heavy metals, which could lead to changes in carbon (C) cycling. Heavy-metal contamination in soil could affect the rates of microbial-mediated biogeochemical processes. For example, soil nitrification rates were suppressed by metal salts such as CdCl2 and ZnCl2, both in spiked soil samples in short-term studies and in polluted fields where metals typically accumulated at a slower rate [192,193,194]. In a study using surface wetland sediments spiked with multiple metals, total denitrification activity was inhibited by Cd (30.9%), Zn (24.9%) and Cu (18.9%) over a period of 7 days of incubation, with Cd being the strongest inhibitor[195]. Heavy-metal pollution was found to affect the activities of denitrifying bacteria and ammonia oxidizers in soil [196].
3.7. Different Forms of Cd and Its Mineralization under the Change in Redox Conditions of the Soil
Various forms of heavy metals are consistently found in the soil, each exhibiting distinct levels of toxicity, bioavailability, and solubility such as 1) insolubly precipitated with other soil, 2) The structural elements of the lattices within clay minerals, 3) Substitutable in both organic and inorganic components, and, 4) dissolved (in soil solution) [197]. The properties of soil such as pH, soil redox potentials (Eh), cation exchange capacity, and organic matter contents, Mn and Fe oxides, clay minerals, and calcium carbonate [9]. Changes in soil Eh impact the bioavailability of Cd due to alterations in redox status and variations in electron acceptors. In aerobic, oxidizing conditions, Cd commonly exists as a soluble salt and cationic form, specifically Cd2+. However, in anaerobic situations with a low redox potential, Cd tends to exist as precipitated species like CdS and CdCO3 [198]. Conversely, alterations in redox potential influence both the organic matter and mineral components of the soil [100]. Furthermore, soil redox conditions regulate the microbial community characteristics and functions [199,200].
The soil moisture regime regulates both soil Eh and biological activity. Paddy soil typically experiences saturated conditions. Heavy metals are found in solid-phase components, and the introduction of heavy metals to paddy soil leads to their transition from more labile fractions to less labile fractions [201,202]. At field capacity moisture, Cd exhibited lower stability when compared to saturated conditions [203]. The saturated regime resulted in more complete transformation of metals toward the stable fractions than the field capacity regime [203]. Soil kept dry exhibited significantly higher soluble concentrations of Cd, Cu, and Zn than the field capacity and the saturated treatment [204].
Management practices such as air drying cycles are pertinent to soil moisture conditions, influencing the exchangeable fraction of heavy metals in the soil. Air drying enhanced Cd desorption in soil and the leachability of metals from dredged canal sediment increased during drying and oxidation processes [205,206]. Drying the soil in an oven resulted in notable rises in the concentrations of dissolved copper (Cu) and nickel (Ni). This outcome can be attributed to the complexation of these metals with increased levels of humic acid and fulvic acid in the soil [207]. Air drying enhances the extractability of manganese [118], iron (Fe), copper (Cu), and zinc [91] at low-incubation moisture levels, while reducing the extractability of metals at high-incubation moisture levels [208]. In paddy soil incubated under three distinct moisture regimes, the exchangeable fraction of copper (Cu), lead (Pb), and cadmium [1] exhibited the following order: 75% field capacity > wetting–drying cycle > flooding. Conversely, the reducible fraction followed the reverse order [209,210]. Eh stands as a crucial factor influencing the mobility of metals in soil or sediment, and is closely tied to soil moisture regulation [211]. Under oxidizing conditions, there is a discernible likelihood of significant heavy metal release [212].
4. Toxic Effects of Cd on Rice
Elevated cadmium [1] concentrations have the potential to impede plant growth, reduce grain yield and quality [212]. This can result in modifications to plant tissues, and disrupt the process of photosynthesis [212], and regulating expressions of at least 36 proteins in rice [213]. Accumulation and transport of Cd occur in various organs and cell organelles, once it enters into the rice [214]. Different researchers have reported the toxic effects of Cd on plant growth [215,216]. Stunted growth and chlorosis are the most typical Cd toxicity symptoms [217]. Due to the direct and indirect interaction of the high level of Cd in the leaves, chlorosis (chlorophyll loss) takes place. The change in physiological activities of plant-like respiration, cell proliferation, and photosynthesis indirectly [217], nitrogen metabolism, plant water relationship, and mineral nutrition under direct Cd stress had been reported, which resulted in poor growth and low biomass of plants [218]. Sensitivity to drought, enzymes inactivation, and damages to membranes of plants have been well-establishing facts of Cd toxicity [219]. It stops indirectly the photosynthesis of the plant by closing the stomata due to water conductance which reduces the access of CO2 to the plant [220]. Directly, Cd prevents the photosynthesis by affecting electron transport, chloroplast organization, chlorophyll biosynthesis, and activity of Calvin Benson Bassham cycle enzymes [221,222].
The general symptoms of Cd toxicity in rice plants include growth inhibition, chlorosis of leaves and sheaths, brown root system, and wilted leaves [203,223]. Different metals-sulfide complexes make brown pigment which deposits on the surface of rice roots [224]. The production of lipid peroxides is a response to Cd toxicity in rice plants. In addition, the uptake of plant essential elements such as Fe, Zn, Mn, and Cu has also been reported to be affected by Cd toxicity in rice plants [225,226]. In addition, the light-harvesting center and photosystem II along with chlorophyll metabolism in rice leaves are affected by Cd [227].
5. HM and VOCs in Paddy Soil under DWMP
Various water management practices influence the Cd uptake in rice (Figure 2). Volatile organic compounds (VOCs) emitted by vegetation encompass a wide array of chemical molecules, predominantly comprised of isoprene, monoterpenes, and methanol [15,228]. In any case, the variety of plant-emitted VOCs depends on the plant species, the distinctive parts of the plants, or the growth conditions [229]. Plants can emit higher quantities of these molecules by directing a greater amount of carbon into stress-triggered volatile organic compounds (VOCs) when subjected to attacks by pathogens or herbivores. Additionally, exposure to various abiotic stresses such as UV radiation, ozone, drought, eutrophication, warming, and overall oxidative stress can also contribute to the increased release of these compounds [230,231,232,233]. Actually, many VOCs of the family of terpenes, such as isoprene and monoterpenes, are antioxidants and can protect cells from oxidative stress[233,234].
Under aerobic irrigation of rice, Cd uptake and isoprene emission decreased [235,236]. Isoprene (2-methyl-1, 3-butadiene) is an important plant VOC known for its plant protection role against numerous abiotic stimuli, including heat [237] and drought stress [238]. Cd decreases the activity of isoprene synthase, an enzyme responsible for conversion of dimethylallyl diphosphate (DMAPP) to isoprene [239,240]. Similarly, One class of VOCs, that Cd amplified was green leaf volatile (GLVs), which play a part in plant defense [239,241]. Furthermore, Cd contamination modulating the emission of terpenoids and expression of genes encoding terpene synthases (TPSs). TPSs is responsible for different terpenoids formation utilizing isopentenyl diphosphate (IPP) and DMAPP [239].
5.1. Volatile Terpenoids
The most abundant (more than 40,000 individual compounds) and structurally diverse group of floral volatiles are called terpenoids [242]. Terpenoids are essential for the growth and development of plants. Terpenoids are the components of abscisic acid (C15), membrane structures (sterols C30), photosynthetic pigments (carotenoids C40), involves in mitochondrial electron transport chain, and gibberellins (C20). Several plant secondary metabolites are terpenoids, such as mono-, sesqui- and diterpenes play important role in plant-plant interactions and plant-environment interaction [243].
Sesquiterpenes, Isoprenes and monoterpenes are known as volatile terpenoids and the largest class of plant volatile compounds (PVCs). They exist in almost every part of the plants including seeds, fruits, leaves, and roots. However, the predominantly PVCs are floral volatiles called lipophilic liquids. Floral volatiles are low molecular weight and high vapor pressure at ambient temperatures [243,244]. There is a link between the PVCs and plants pollinators [244,245]. They attract pollinators such as bats, birds, mammals, insects, and other animals as vectors for pollens transfer [246]. For Long range communication and evolutions of flowering plant, floral scent is an important source of communication [247]. Floral volatiles changes the behavioral response in respective pollinators. Floral volatiles upon emission guiding the pollinators towards the flowers. For example, more volatile terpenoids emitting by latifolia and Petunia axillaris, moths pollinated than the same genus Petunia integrifolia and Silene dioica a day-emitting bee-pollinated [243,248]. Cd inhibit terpenoids by interfering with enzymes responsible for terpenoid production production, such as geranylgeranyl diphosphate synthase (GGPPS), 1-deoxy-D-xylulose-5-phosphate synthase (DXS), and 1-deoxy-D-xylulose-5-phosphate reductoisomerase (DXR). In rice, Cd reported to induce oxidative stress and damage the cellular membranes. Cellular membranes are mostly composed of sterols and carotenoids i.e., terpenoids. Furthermore, Cd in rice disrupting the hormonal balance and signaling pathways responsible for terpenoids metabolism [249].
5.2. Effects of Heavy Metals on Rice Fragrance
Aromatic 2-acetyl-1-pyrroline is the main ingredient in rice fragrance (2AP). Two genes, betaine aldehyde dehydrogenase (BADH1) and BADH2, functioned in the 2AP biosynthetic pathway. The exact mechanism of Cd affecting these genes is not well studied. However, studies reported that Cd may interfere with the expression and functions of BADH1 and BADH2, limiting rice yield and quality. For example, upon exposure to Cd, the expression of BADH1 and BADH2 in rice seedlings. Similar Cd lowered the activity of enzymes encoded by these genes BADH and betaine aldehyde synthase (Figure 3) [26,250]. These enzymes are in responsible for converting the aldehyde betaine into glycine, a precursor of 2AP. Glycine is a compatible solute which protect plants from stress. The decreased activity of these enzymes lower the levels of glycine betaine and higher levels of betaine aldehyde leads to plant toxicity. Furthermore, the accumulation of 2PA accumulation due to Cd in the grain also affected the rice fragrance [250].
The duplication of the BADH2 gene in rice decreased Cd accumulation in rice root and shoots under salt and Cd stress. Duplication increased the expression and activity of BADH2, which uptake more manganese, a micronutrient and competitor with Cd for transport in rice cell [251]. Furthermore, the introduction of the BADH2 gene duplication into various rice cultivars reduced Cd accumulation without impacting yield or grain quality [251]. Similarly, 96 rice accessions were found to have 12 single nucleotide polymorphisms (SNPs) in six cadmium-related genes, comprising BADH1 and BADH2 [99]. SNPs are linked with the Cd accretion in rice grains. In rice breeding practices, SNPs might be used as molecular markers for low-Cd accumulating rice cultivars.
6. Conclusions and Future Directions
Cadmium contamination poses serious threats to rice agriculture and paddy soil N transformation. The N transformation is severely affected in AWD practices than CF. Similarly, Cd interferes with the signaling and synthesis of plant cells that are responsible for VOC production and aroma. High Cd deposition in rice cells inhibits VOC formation; nevertheless, the precise mechanism by which VOC synthesis is modulated by DWMP remains unknown. This may compromise plant defense, induced by VOCs and rice aroma as well. The issue of rice aroma, quality, and yield can be mitigated in future breeding procedures by introducing low-Cd absorption rice cultivars and utilizing water-saving technology.
Currently, Cd accumulation in grains is reduced via agronomic parameters (soil improvement, water, fertilizer, tillage management) and applying silicon and abscisic acid externally. Another strategy is microbial and phytoremediation. However, these processes are cost-effective and laborious with low efficiency. Therefore, screening and breeding low Cd accumulation rice varieties is the only choice considered economical and effective. Crossbreeding is very effective in transferring low Cd gene pool to offspring. However, molecular marker assistant breeding can reduce the screening process in breeding and save time. In addition, mutation breeding also helps to develop low Cd accumulating variety quickly. However, molecular marker assistant breeding can reduce the screening process in breeding and save time. CRISPR and hybridization technologies can also be used to develop new low-Cd accumulator rice verities. Adaptations of such techniques by the breeders may develop rice variety that can be grown in Cd-polluted paddies but will maintain the aroma and grain quality.
Funding
This work was supported by the National Natural Science Foundation of China (31970001), Science and Technology Project of Guangdong Province (2018A050506075) and the Joint Open Project from Key Laboratory for Enhancing Resource Use Efficiency of Crops in South China, Ministry of Agriculture and Rural Affairs, China and Industrial College of South China Agricultural University-Guangdong Lardmee Fertilizer Co., Ltd.
References
- Wang, H.; Zhang, Y.; Zhang, Y.; McDaniel, M.D.; Sun, L.; Su, W.; Fan, X.; Liu, S.; Xiao, X. Water-saving irrigation is a ‘win-win’management strategy in rice paddies–With both reduced greenhouse gas emissions and enhanced water use efficiency. Agricultural Water Management 2020, 228, 105889. [Google Scholar] [CrossRef]
- Mohidem, N.A.; Hashim, N.; Shamsudin, R.; Che Man, H. Rice for food security: Revisiting its production, diversity, rice milling process and nutrient content. Agriculture 2022, 12, 741. [Google Scholar] [CrossRef]
- Bin Rahman, A.R.; Zhang, J. Trends in rice research: 2030 and beyond. Food and Energy Security 2023, 12, e390. [Google Scholar] [CrossRef]
- Zhao, M.; Lin, Y.; Chen, H. Improving nutritional quality of rice for human health. Theoretical and Applied Genetics 2020, 133, 1397–1413. [Google Scholar] [CrossRef]
- Wu, X.H.; Wang, W.; Yin, C.M.; Hou, H.J.; Xie, K.J.; Xie, X.L. Water consumption, grain yield, and water productivity in response to field water management in double rice systems in China. PloS one 2017, 12, e0189280. [Google Scholar] [CrossRef]
- Arouna, A.; Dzomeku, I.K.; Shaibu, A.-G.; Nurudeen, A.R. Water Management for Sustainable Irrigation in Rice (Oryza sativa L.) Production: A Review. Agronomy 2023, 13, 1522. [Google Scholar] [CrossRef]
- Shao, G.; Cui, J.; Lu, B.; Brian, B.J.; Ding, J.; She, D. Impacts of controlled irrigation and drainage on the yield and physiological attributes of rice. Agricultural Water Management 2015, 149, 156–165. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Ul-Allah, S.; Siddique, K.H. Physiological and agronomic approaches for improving water-use efficiency in crop plants. Agricultural Water Management 2019, 219, 95–108. [Google Scholar] [CrossRef]
- Afzal, M.; Yu, M.; Tang, C.; Zhang, L.; Muhammad, N.; Zhao, H.; Feng, J.; Yu, L.; Xu, J. The negative impact of cadmium on nitrogen transformation processes in a paddy soil is greater under non-flooding than flooding conditions. Environment international 2019, 129, 451–460. [Google Scholar] [CrossRef]
- Afzal, M.; Tang, C.; Yu, M.; Muhammad, N.; Zhao, H.; Xu, J. Water regime is important to determine cadmium toxicity on rice growth and rhizospheric nitrifier communities in contaminated paddy soils. Plant and Soil 2022, 472, 609–628. [Google Scholar] [CrossRef]
- Xia, L.; Li, X.; Ma, Q.; Lam, S.K.; Wolf, B.; Kiese, R.; Butterbach-Bahl, K.; Chen, D.; Li, Z.; Yan, X. Simultaneous quantification of N2, NH3 and N2O emissions from a flooded paddy field under different N fertilization regimes. Global Change Biology 2020, 26, 2292–2303. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Ashida, N.; Kai, A.; Isobe, K.; Nishizawa, T.; Otsuka, S.; Yokota, A.; Senoo, K.; Ishii, S. Presence of Cu-Type (NirK) and cd1-Type (NirS) Nitrite Reductase Genes in the Denitrifying Bacterium Bradyrhizobium nitroreducens sp. nov. Microbes and environments 2018, 33, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhao, J.; Guo, Z.; Ma, J.; Xu, H.; Jia, Z. Differential contributions of ammonia oxidizers and nitrite oxidizers to nitrification in four paddy soils. The ISME journal 2015, 9, 1062–1075. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Yao, P.; Liu, J.; Zhao, B.; Zhang, G.; Zhao, M.; Yu, Z.; Zhang, X.-H. Diversity, abundance, and niche differentiation of ammonia-oxidizing prokaryotes in mud deposits of the eastern China marginal seas. Frontiers in Microbiology 2016, 7, 137. [Google Scholar] [CrossRef] [PubMed]
- Ninkovic, V.; Markovic, D.; Rensing, M. Plant volatiles as cues and signals in plant communication. Plant, cell & environment 2021, 44, 1030–1043. [Google Scholar]
- Ramos, S.E.; Schiestl, F.P. Rapid plant evolution driven by the interaction of pollination and herbivory. Science 2019, 364, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.-S.; Zhang, B.; Zhan, H.; Lv, Y.-L.; Jia, X.-L.; Wang, T.; Yang, N.-Y.; Lou, Y.-X.; Zhang, Z.-B.; Hu, W.-J. Phenylpropanoid derivatives are essential components of sporopollenin in vascular plants. Molecular plant 2020, 13, 1644–1653. [Google Scholar] [CrossRef] [PubMed]
- Farré-Armengol, G.; Filella, I.; Llusia, J.; Peñuelas, J. Bidirectional interaction between phyllospheric microbiotas and plant volatile emissions. Trends in Plant Science 2016, 21, 854–860. [Google Scholar] [CrossRef]
- Ninkuu, V.; Zhang, L.; Yan, J.; Fu, Z.; Yang, T.; Zeng, H. Biochemistry of terpenes and recent advances in plant protection. International Journal of Molecular Sciences 2021, 22, 5710. [Google Scholar] [CrossRef]
- Brosset, A.; Blande, J.D. Volatile-mediated plant–plant interactions: volatile organic compounds as modulators of receiver plant defence, growth, and reproduction. Journal of experimental botany 2022, 73, 511–528. [Google Scholar] [CrossRef]
- Yi, J.; Lo, L.S.H.; Liu, H.; Qian, P.-Y.; Cheng, J. Study of heavy metals and microbial communities in contaminated sediments along an urban estuary. Frontiers in Marine Science 2021, 8, 741912. [Google Scholar] [CrossRef]
- Bao, P.; Li, G.-X.; Sun, G.-X.; Xu, Y.-Y.; Meharg, A.A.; Zhu, Y.-G. The role of sulfate-reducing prokaryotes in the coupling of element biogeochemical cycling. Science of the Total Environment 2018, 613, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.; Tardy, V.; Bonnineau, C.; Billard, P.; Pesce, S.; Lyautey, E. Changes in sediment microbial diversity following chronic copper-exposure induce community copper-tolerance without increasing sensitivity to arsenic. Journal of hazardous materials 2020, 391, 122197. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.; Zhang, C.; Mathews, E.; Tang, C.; Franks, A. Microbial community dynamics in the rhizosphere of a cadmium hyper-accumulator. Scientific reports 2016, 6, 36067. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Luan, Y.; Ning, Y.; Wang, L. Effects and mechanisms of microbial remediation of heavy metals in soil: a critical review. Applied Sciences 2018, 8, 1336. [Google Scholar] [CrossRef]
- Muehe, E.M.; Weigold, P.; Adaktylou, I.J.; Planer-Friedrich, B.; Kraemer, U.; Kappler, A.; Behrens, S. Rhizosphere microbial community composition affects cadmium and zinc uptake by the metal-hyperaccumulating plant Arabidopsis halleri. Applied and environmental microbiology 2015, 81, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Visioli, G.; D’Egidio, S.; Sanangelantoni, A.M. The bacterial rhizobiome of hyperaccumulators: future perspectives based on omics analysis and advanced microscopy. Frontiers in plant science 2015, 5, 752. [Google Scholar] [CrossRef]
- Roman-Ponce, B.; REZA-VÁZQUEZ, D.M.; Gutierrez-Paredes, S.; María de Jesús, D.; Maldonado-Hernandez, J.; Bahena-Osorio, Y.; Estrada-De los Santos, P.; WANG, E.T.; VÁSQUEZ-MURRIETA, M.S. Plant growth-promoting traits in rhizobacteria of heavy metal-resistant plants and their effects on Brassica nigra seed germination. Pedosphere 2017, 27, 511–526. [Google Scholar] [CrossRef]
- Qu, Q.; Zhang, Z.; Peijnenburg, W.; Liu, W.; Lu, T.; Hu, B.; Chen, J.; Chen, J.; Lin, Z.; Qian, H. Rhizosphere microbiome assembly and its impact on plant growth. Journal of agricultural and food chemistry 2020, 68, 5024–5038. [Google Scholar] [CrossRef]
- Oves, M.; Qari, H.A.; Khan, M.S. Sinorhizobium saheli: Advancing Chromium Mitigation, Metal Adsorption, and Plant Growth Enhancement in Heavy Metal-Contaminated Environments. Journal of Plant Growth Regulation 2023, 1–20. [Google Scholar] [CrossRef]
- Lin, H.; Liu, C.; Li, B.; Dong, Y. Trifolium repens L. regulated phytoremediation of heavy metal contaminated soil by promoting soil enzyme activities and beneficial rhizosphere associated microorganisms. Journal of Hazardous Materials 2021, 402, 123829. [Google Scholar] [CrossRef]
- Li, L.; Liu, Z.; Meng, D.; Liu, X.; Li, X.; Zhang, M.; Tao, J.; Gu, Y.; Zhong, S.; Yin, H. Comparative genomic analysis reveals the distribution, organization, and evolution of metal resistance genes in the genus Acidithiobacillus. Applied and Environmental Microbiology 2019, 85, e02153-02118. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lei, X.; Qin, L.; Sun, X.; Wang, L.; Zhao, S.; Wang, M.; Chen, S. Fe (III) reduction due to low pe+ pH contributes to reducing Cd transfer within a soil-rice system. Journal of Hazardous Materials 2021, 415, 125668. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Wang, R.; Gao, X.; Wang, K.; Lin, Z.; Ge, J.; Liu, T.; Wei, S.; Chen, W.; Xie, R. Cultivar-specific response of bacterial community to cadmium contamination in the rhizosphere of rice (Oryza sativa L.). Environmental Pollution 2018, 241, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Mekonnen, M.M.; Hoekstra, A.Y. Four billion people facing severe water scarcity. Science advances 2016, 2, e1500323. [Google Scholar] [CrossRef] [PubMed]
- Mallareddy, M.; Thirumalaikumar, R.; Balasubramanian, P.; Naseeruddin, R.; Nithya, N.; Mariadoss, A.; Eazhilkrishna, N.; Choudhary, A.K.; Deiveegan, M.; Subramanian, E. Maximizing water use efficiency in rice farming: A comprehensive review of innovative irrigation management technologies. Water 2023, 15, 1802. [Google Scholar] [CrossRef]
- Kumar, D.; Ramesh, K.; Jinger, D.; Rajpoot, S.K. Effect of potassium fertilization on water productivity, irrigation water use efficiency, and grain quality under direct seeded rice-wheat cropping system. Journal of Plant Nutrition 2022, 45, 2023–2038. [Google Scholar]
- Surendran, U.; Raja, P.; Jayakumar, M.; Subramoniam, S.R. Use of efficient water saving techniques for production of rice in India under climate change scenario: A critical review. Journal of Cleaner Production 2021, 309, 127272. [Google Scholar] [CrossRef]
- Ansari, A.; Lin, Y.-P.; Lur, H.-S. Evaluating and adapting climate change impacts on rice production in Indonesia: a case study of the Keduang subwatershed, Central Java. Environments 2021, 8, 117. [Google Scholar] [CrossRef]
- Brunet-Loredo, A.; López-Belchí, M.D.; Cordero-Lara, K.; Noriega, F.; Cabeza, R.A.; Fischer, S.; Careaga, P.; Garriga, M. Assessing Grain Quality Changes in White and Black Rice under Water Deficit. Plants 2023, 12, 4091. [Google Scholar] [CrossRef]
- Linquist, B.A.; Anders, M.M.; Adviento-Borbe, M.A.; Chaney, R.L.; Nalley, L.L.; da Rosa, E.F.; van Kessel, C. Reducing greenhouse gas emissions, water use, and grain arsenic levels in rice systems. Glob Chang Biol 2015, 21, 407–417. [Google Scholar] [CrossRef]
- Echegaray-Cabrera, I.; Cruz-Villacorta, L.; Ramos-Fernández, L.; Bonilla-Cordova, M.; Heros-Aguilar, E.; Flores del Pino, L. Effect of Alternate Wetting and Drying on the Emission of Greenhouse Gases from Rice Fields on the Northern Coast of Peru. Agronomy 2024, 14, 248. [Google Scholar] [CrossRef]
- Jang, E.-K.; Lim, E.M.; Kim, J.; Kang, M.-J.; Choi, G.; Moon, J. Risk Management of Methane Reduction Clean Development Mechanism Projects in Rice Paddy Fields. Agronomy 2023, 13, 1639. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, W.; Li, S.; Zhu, K.; Hua, X.; Harrison, M.T.; Liu, K.; Yang, J.; Liu, L.; Chen, Y. Integrated management approaches enabling sustainable rice production under alternate wetting and drying irrigation. Agricultural Water Management 2023, 281, 108265. [Google Scholar] [CrossRef]
- Das, S.; Chou, M.-L.; Jean, J.-S.; Liu, C.-C.; Yang, H.-J. Water management impacts on arsenic behavior and rhizosphere bacterial communities and activities in a rice agro-ecosystem. Science of The Total Environment 2016, 542, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Pearson, K.A.; Millar, G.M.; Norton, G.J.; Price, A.H. Alternate wetting and drying in Bangladesh: Water-saving farming practice and the socioeconomic barriers to its adoption. Food and Energy Security 2018, 7, e00149. [Google Scholar] [CrossRef]
- Suhaili, W.S.H. Adoption of technology to improve self-sufficiency in paddy plantations in Brunei: Challenges and mitigation strategies for intermediate stakeholders. In Proceedings of the IOP Conference Series: Earth and Environmental Science; 2023; p. 012011. [Google Scholar]
- Mote, K.; Rao, V.P.; Ramulu, V.; Kumar, K.A.; Devi, M.U.; Sudhakara, T. Alternate wetting and drying irrigation technology for sustainable rice (Oryza sativa) production. Paddy and Water Environment 2023, 21, 551–569. [Google Scholar] [CrossRef]
- He, G.; Wang, Z.; Cui, Z. Managing irrigation water for sustainable rice production in China. Journal of Cleaner Production 2020, 245, 118928. [Google Scholar] [CrossRef]
- Haonan, Q.; Jie, W.; Shihong, Y.; Zewei, J.; Yi, X. Current status of global rice water use efficiency and water-saving irrigation technology recommendations. Journal of Agronomy and Crop Science 2023. [Google Scholar] [CrossRef]
- Yue, Q.; Ledo, A.; Cheng, K.; Albanito, F.; Lebender, U.; Sapkota, T.B.; Brentrup, F.; Stirling, C.M.; Smith, P.; Sun, J. Re-assessing nitrous oxide emissions from croplands across Mainland China. Agriculture, Ecosystems & Environment 2018, 268, 70–78. [Google Scholar]
- Zhuang, Y.; Zhang, L.; Li, S.; Liu, H.; Zhai, L.; Zhou, F.; Ye, Y.; Ruan, S.; Wen, W. Effects and potential of water-saving irrigation for rice production in China. Agricultural Water Management 2019, 217, 374–382. [Google Scholar] [CrossRef]
- Ju, Q.; Du, L.; Liu, C.; Jiang, S. Water resource management for irrigated agriculture in China: Problems and prospects. Irrigation and Drainage 2023. [Google Scholar] [CrossRef]
- Zhang, Z.; Furman, A. Soil redox dynamics under dynamic hydrologic regimes-A review. Science of The Total Environment 2021, 763, 143026. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Zhang, J.; Yu, Y.; Ma, Q.; Kong, Y.; Pan, W.; Wu, L.; Jin, Q. Alternate wetting–drying enhances soil nitrogen availability by altering organic nitrogen partitioning in rice-microbe system. Geoderma 2022, 424, 115993. [Google Scholar] [CrossRef]
- Maharjan, M.; Sanaullah, M.; Razavi, B.S.; Kuzyakov, Y. Effect of land use and management practices on microbial biomass and enzyme activities in subtropical top-and sub-soils. Applied Soil Ecology 2017, 113, 22–28. [Google Scholar] [CrossRef]
- Soremi, P.A. CHANGES IN MICROBIAL BIOMASS CARBON, NITROGEN AND GRAIN YIELD OF LOWLAND RICE VARIETIES IN RESPONSE TO ALTERNATE WET AND DRY WATER REGIME IN THE INLAND VALLEY OF DERIVED SAVANNA. Journal of Agricultural Sciences (Belgrade) 2019, 64. [Google Scholar]
- Liu, X.; Zhou, T.; Liu, Y.; Zhang, X.; Li, L.; Pan, G. Effect of mid-season drainage on CH4 and N2O emission and grain yield in rice ecosystem: A meta-analysis. Agricultural Water Management 2019, 213, 1028–1035. [Google Scholar] [CrossRef]
- Liao, B.; Wu, X.; Yu, Y.; Luo, S.; Hu, R.; Lu, G. Effects of mild alternate wetting and drying irrigation and mid-season drainage on CH4 and N2O emissions in rice cultivation. Science of The Total Environment 2020, 698, 134212. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, M.; Li, X.; Dai, Q.; Xu, K.; Guo, B.; Hu, Y.; Wang, W.; Huo, Z. Effects of Soil Types and Irrigation Modes on Rice Root Morphophysiological Traits and Grain Quality. Agronomy 2021, 11, 120. [Google Scholar] [CrossRef]
- Yang, X.; Wang, B.; Chen, L.; Li, P.; Cao, C. The different influences of drought stress at the flowering stage on rice physiological traits, grain yield, and quality. Scientific Reports 2019, 9, 3742. [Google Scholar] [CrossRef]
- Norton, G.J.; Shafaei, M.; Travis, A.J.; Deacon, C.M.; Danku, J.; Pond, D.; Cochrane, N.; Lockhart, K.; Salt, D.; Zhang, H.; et al. Impact of alternate wetting and drying on rice physiology, grain production, and grain quality. Field Crops Research 2017, 205, 1–13. [Google Scholar] [CrossRef]
- Maneepitak, S.; Ullah, H.; Paothong, K.; Kachenchart, B.; Datta, A.; Shrestha, R.P. Effect of water and rice straw management practices on yield and water productivity of irrigated lowland rice in the Central Plain of Thailand. Agricultural Water Management 2019, 211, 89–97. [Google Scholar] [CrossRef]
- Ishfaq, M.; Akbar, N.; Zulfiqar, U.; Ali, N.; Ahmad, M.; Anjum, S.A.; Farooq, M. Influence of water management techniques on milling recovery, grain quality and mercury uptake in different rice production systems. Agricultural Water Management 2021, 243, 106500. [Google Scholar] [CrossRef]
- Colombo, R.; Moretto, G.; Barberis, M.; Frosi, I.; Papetti, A. Rice Byproduct Compounds: From Green Extraction to Antioxidant Properties. Antioxidants 2024, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Carrijo, D.R.; Nakayama, Y.; Linquist, B.A.; Green, P.G.; Parikh, S.J. Impact of Alternate Wetting and Drying Irrigation on Arsenic Uptake and Speciation in Flooded Rice Systems. Agriculture, Ecosystems & Environment 2019, 272, 188–198. [Google Scholar] [CrossRef]
- Mahapatra, B.; Bhupenchandra, I.; Devi, S.; Kumar, A.; Chongtham, S.; Singh, R.; Babu, S.; Bora, S.; Devi, E.; Verma, G. Aerobic Rice and its significant perspective for sustainable crop production. Indian Journal of Agronomy 2021, 66, 383–392. [Google Scholar] [CrossRef]
- Parthasarathi, T.; Vanitha, K.; Mohandass, S.; Vered, E. Evaluation of drip irrigation system for water productivity and yield of rice. Agronomy Journal 2018, 110, 2378–2389. [Google Scholar] [CrossRef]
- Ghosh, A.; Singh, O.; Berliner, J.; Pokhare, S. System of crop rotation: a prospective strategy alleviating grain yield penalty in sustainable aerobic rice production. International Journal of Plant Production 2021, 15, 577–587. [Google Scholar] [CrossRef]
- Samoy-Pascual, K.; Lampayan, R.M.; Remocal, A.T.; Orge, R.F.; Tokida, T.; Mizoguchi, M. Optimizing the lateral dripline spacing of drip-irrigated aerobic rice to increase water productivity and profitability under the water-limited condition. Field Crops Research 2022, 287, 108669. [Google Scholar] [CrossRef]
- Masseroni, D.; Ricart, S.; De Cartagena, F.R.; Monserrat, J.; Gonçalves, J.M.; De Lima, I.; Facchi, A.; Sali, G.; Gandolfi, C. Prospects for improving gravity-fed surface irrigation systems in Mediterranean European contexts. Water 2017, 9, 20. [Google Scholar] [CrossRef]
- Kaur, M.; Sharma, K. Rice Productivity and Water Use Efficiency under Different Irrigation Management System in North-Western India. Indian Journal of Extension Education 2022, 58, 65–68. [Google Scholar] [CrossRef]
- Alvarenga, P.; Fernández-Rodríguez, D.; Abades, D.P.; Rato-Nunes, J.M.; Albarrán, Á.; López-Piñeiro, A. Combined use of olive mill waste compost and sprinkler irrigation to decrease the risk of As and Cd accumulation in rice grain. Science of the Total Environment 2022, 835, 155488. [Google Scholar] [CrossRef] [PubMed]
- Mubangizi, A.; Wanyama, J.; Kiggundu, N.; Nakawuka, P. Assessing Suitability of Irrigation Scheduling Decision Support Systems for Lowland Rice Farmers in Sub-Saharan Africa—A Review. Agricultural Sciences 2023, 14, 219–239. [Google Scholar] [CrossRef]
- Islam, S.F.-u.; Sander, B.O.; Quilty, J.R.; de Neergaard, A.; van Groenigen, J.W.; Jensen, L.S. Mitigation of greenhouse gas emissions and reduced irrigation water use in rice production through water-saving irrigation scheduling, reduced tillage and fertiliser application strategies. Science of The Total Environment 2020, 739, 140215. [Google Scholar] [CrossRef] [PubMed]
- Mlangeni, A.T. Methylation of arsenic in rice: Mechanisms, factors, and mitigation strategies. Toxicology Reports 2023, 11, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Tang, S.; Pan, W.; Xu, M.; Liu, X.; Ni, L.; Mao, X.; Sun, T.; Fu, H.; Han, K. Long-term application of controlled-release fertilizer enhances rice production and soil quality under non-flooded plastic film mulching cultivation conditions. Agriculture, Ecosystems & Environment 2023, 358, 108720. [Google Scholar]
- Ghulamahdi, M.; Sulistyono, E.; Lubis, I.; Sastro, Y. Response of rice peat humic acid ameliorant. saturated,. soil. culture.(SSC) within tidal. swamps. In Proceedings of the IOP Conference Series: Earth and Environmental Science; 2023; p. 012009. [Google Scholar]
- Isnawan, B.; Hariyono, H.; Gumilar, A. Selection of Intermittent Irrigation to Increase Growth and Yield of Some Local Rice Varieties (Oryza sativa L.) in the Rainy Season. Jurnal Ilmiah Multidisiplin Indonesia (JIM-ID) 2023, 2, 55–62. [Google Scholar] [CrossRef]
- Huang, Y.; Mubeen, S.; Yang, Z.; Wang, J. Cadmium contamination in agricultural soils and crops. In Theories and Methods for Minimizing Cadmium Pollution in Crops: Case Studies on Water Spinach; Springer, 2022; pp. 1–30. [Google Scholar]
- Shahid, M.; Dumat, C.; Khalid, S.; Niazi, N.K.; Antunes, P.M. Cadmium bioavailability, uptake, toxicity and detoxification in soil-plant system. Reviews of Environmental Contamination and Toxicology 2017, 241, 73–137. [Google Scholar] [PubMed]
- Mar, S.S.; Okazaki, M. Investigation of Cd contents in several phosphate rocks used for the production of fertilizer. Microchemical Journal 2012, 104, 17–21. [Google Scholar] [CrossRef]
- Faridullah, F.; Umar, M.; Alam, A.; Sabir, M.A.; Khan, D. Assessment of heavy metals concentration in phosphate rock deposits, Hazara basin, Lesser Himalaya Pakistan. Geosciences Journal 2017, 21, 743–752. [Google Scholar] [CrossRef]
- Boumaza, B.; Kechiched, R.; Chekushina, T.V.; Benabdeslam, N.; Senouci, K.; Merzeg, F.A.; Rezgui, W.; Rebouh, N.Y.; Harizi, K. Geochemical distribution and environmental assessment of potentially toxic elements in farmland soils, sediments, and tailings from phosphate industrial area (NE Algeria). Journal of Hazardous Materials 2024, 465, 133110. [Google Scholar] [CrossRef]
- Canty, M.J.; Scanlon, A.; Collins, D.M.; McGrath, G.; Clegg, T.A.; Lane, E.; Sheridan, M.K.; More, S.J. Cadmium and other heavy metal concentrations in bovine kidneys in the Republic of Ireland. Science of The Total Environment 2014, 485-486, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Jyoti, V.; Saini-Eidukat, B.; Hopkins, D.; DeSutter, T. Naturally elevated metal contents of soils in northeastern North Dakota, USA, with a focus on cadmium. J Soils Sediments 2015, 15, 1571–1583. [Google Scholar] [CrossRef]
- Robson, T.C.; Braungardt, C.B.; Rieuwerts, J.; Worsfold, P. Cadmium contamination of agricultural soils and crops resulting from sphalerite weathering. Environmental Pollution 2014, 184, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Reimann, C.; Fabian, K.; Flem, B. Cadmium enrichment in topsoil: Separating diffuse contamination from biosphere-circulation signals. Science of The Total Environment 2019, 651, 1344–1355. [Google Scholar] [CrossRef] [PubMed]
- Imseng, M.; Wiggenhauser, M.; Keller, A.; Müller, M.; Rehkämper, M.; Murphy, K.; Kreissig, K.; Frossard, E.; Wilcke, W.; Bigalke, M. Fate of Cd in Agricultural Soils: A Stable Isotope Approach to Anthropogenic Impact, Soil Formation, and Soil-Plant Cycling. Environ Sci Technol 2018, 52, 1919–1928. [Google Scholar] [CrossRef] [PubMed]
- Birke, M.; Reimann, C.; Rauch, U.; Ladenberger, A.; Demetriades, A.; Jähne-Klingberg, F.; Oorts, K.; Gosar, M.; Dinelli, E.; Halamić, J. GEMAS: Cadmium distribution and its sources in agricultural and grazing land soil of Europe — Original data versus clr-transformed data. Journal of Geochemical Exploration 2017, 173, 13–30. [Google Scholar] [CrossRef]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environmental and Experimental Botany 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Fu, Y.; Li, F.; Guo, S.; Zhao, M. Cadmium concentration and its typical input and output fluxes in agricultural soil downstream of a heavy metal sewage irrigation area. Journal of Hazardous Materials 2021, 412, 125203. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, S.; Sher, F.; Chen, J.; Xin, Y.; You, Z.; Wen, L.; Hu, M.; Qiu, G. A review on recycling and reutilization of blast furnace dust as a secondary resource. Journal of Sustainable Metallurgy 2021, 7, 340–357. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, S.; Ai, F.; Jin, L.; Zhu, N.; Meng, X.-Z. Identification of industrial sewage sludge based on heavy metal profiles: a case study of printing and dyeing industry. Environmental Science and Pollution Research 2022, 1–10. [Google Scholar] [CrossRef]
- Zhai, Y.; Liu, X.; Chen, H.; Xu, B.; Zhu, L.; Li, C.; Zeng, G. Source identification and potential ecological risk assessment of heavy metals in PM2.5 from Changsha. Science of The Total Environment 2014, 493, 109–115. [Google Scholar] [CrossRef]
- Khan, Z.I.; Haider, R.; Ahmad, K.; Alrefaei, A.F.; Mehmood, N.; Memona, H.; Akhtar, S.; Ugulu, I. The Effects of Irrigation with Diverse Wastewater Sources on Heavy Metal Accumulation in Kinnow and Grapefruit Samples and Health Risks from Consumption. Water 2023, 15, 3480. [Google Scholar] [CrossRef]
- Tang, X.; Li, Q.; Wu, M.; Lin, L.; Scholz, M. Review of remediation practices regarding cadmium-enriched farmland soil with particular reference to China. Journal of environmental management 2016, 181, 646–662. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.-H.; Zhu, J.; Liu, S.-H.; Pan, S.-F.; Ji, X.-H. Input and output of cadmium (Cd) for paddy soil in central south China: fluxes, mass balance, and model predictions. Environmental Science and Pollution Research 2020, 27, 21847–21858. [Google Scholar] [CrossRef]
- Li, W.; Xu, F.; Cai, T.; Zhao, W.; Lin, J.; Huang, J.; Wang, L.; Bian, J.; Fu, J.; Ouyang, L. The Relationship between Cadmium-Related Gene Sequence Variations in Rice and Cadmium Accumulation. Agronomy 2023, 13, 800. [Google Scholar] [CrossRef]
- Luo, J.; de Klein, C.A.M.; Ledgard, S.F.; Saggar, S. Management options to reduce nitrous oxide emissions from intensively grazed pastures: A review. Agriculture, Ecosystems & Environment 2010, 136, 282–291. [Google Scholar] [CrossRef]
- Liu, X.; Tian, G.; Jiang, D.; Zhang, C.; Kong, L. Cadmium (Cd) distribution and contamination in Chinese paddy soils on national scale. Environmental Science and Pollution Research 2016, 23, 17941–17952. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-Y.; Liu, C.; Zhu, J.; Li, F.; Deng, D.-M.; Wang, Q.; Liu, C. Cadmium availability in rice paddy fields from a mining area: The effects of soil properties highlighting iron fractions and pH value. Environmental Pollution 2016, 209, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.N.; Lei, M.; Sun, G.; Huang, Q.; Lu, Y.; Deacon, C.; Meharg, A.A.; Zhu, Y.G. Occurrence and partitioning of cadmium, arsenic and lead in mine impacted paddy rice: Hunan, China. Environ Sci Technol 2009, 43, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Y.; Li, L.Q.; Pan, G.X. [Variation of Cd, Zn and Se contents of polished rice and the potential health risk for subsistence-diet farmers from typical areas of South China]. Huan Jing Ke Xue 2009, 30, 2792–2797. [Google Scholar]
- Jia, L.; Xie, Y.; Wang, Z.; Luo, L.; Zhang, C.; Pélissier, P.M.; Parizot, B.; Qi, W.; Zhang, J.; Hu, Z.; et al. Rice plants respond to ammonium stress by adopting a helical root growth pattern. Plant J 2020, 104, 1023–1037. [Google Scholar] [CrossRef]
- Li, L.; Wu, T.; Li, Y.; Hu, X.; Wang, Z.; Liu, J.; Qin, W.; Ashraf, U. Deep fertilization improves rice productivity and reduces ammonia emissions from rice fields in China; a meta-analysis. Field Crops Research 2022, 289, 108704. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Sun, R.; Hu, S.; Qiao, Z.; Wang, S.; Mi, X. NH4+-N/NO3−-N ratio controlling nitrogen transformation accompanied with NO2−-N accumulation in the oxic-anoxic transition zone. Environmental Research 2020, 189, 109962. [Google Scholar] [CrossRef]
- Yoon, S.; Song, B.; Phillips, R.L.; Chang, J.; Song, M.J. Ecological and physiological implications of nitrogen oxide reduction pathways on greenhouse gas emissions in agroecosystems. FEMS microbiology ecology 2019, 95, fiz066. [Google Scholar] [CrossRef]
- Tan, X.; Shao, D.; Gu, W. Effects of temperature and soil moisture on gross nitrification and denitrification rates of a Chinese lowland paddy field soil. Paddy and Water Environment 2018, 16, 687–698. [Google Scholar] [CrossRef]
- Yi, L.; Sheng, R.; Wei, W.; Zhu, B.; Zhang, W. Differential contributions of electron donors to denitrification in the flooding-drying process of a paddy soil. Applied Soil Ecology 2022, 177, 104527. [Google Scholar] [CrossRef]
- Chen, L.; Liu, X.; Hua, Z.; Xue, H.; Mei, S.; Wang, P.; Wang, S. Comparison of nitrogen loss weight in ammonia volatilization, runoff, and leaching between common and slow-release fertilizer in paddy field. Water, Air, & Soil Pollution 2021, 232, 1–11. [Google Scholar]
- Han, B.; Mo, L.-Y.; Fang, Y.-T.; Di, H.J.; Wang, J.-T.; Shen, J.-P.; Zhang, L.-M. Rates and microbial communities of denitrification and anammox across coastal tidal flat lands and inland paddy soils in East China. Applied Soil Ecology 2021, 157, 103768. [Google Scholar] [CrossRef]
- Jia, B.; Niu, Z.; Wu, Y.; Kuzyakov, Y.; Li, X.G. Waterlogging increases organic carbon decomposition in grassland soils. Soil Biology and Biochemistry 2020, 148, 107927. [Google Scholar] [CrossRef]
- Wang, N.; Zhao, Y.-H.; Yu, J.-G.; Xue, L.-H.; Li, H.-B.; Yang, L.-Z. Roles of bulk and rhizosphere denitrifying bacteria in denitrification from paddy soils under straw return condition. Journal of Soils and Sediments 2021, 21, 2179–2191. [Google Scholar] [CrossRef]
- Ding, L.-J.; Cui, H.-L.; Nie, S.-A.; Long, X.-E.; Duan, G.-L.; Zhu, Y.-G. Microbiomes inhabiting rice roots and rhizosphere. FEMS microbiology ecology 2019, 95, fiz040. [Google Scholar] [CrossRef] [PubMed]
- Malyan, S.K.; Bhatia, A.; Tomer, R.; Harit, R.C.; Jain, N.; Bhowmik, A.; Kaushik, R. Mitigation of yield-scaled greenhouse gas emissions from irrigated rice through Azolla, Blue-green algae, and plant growth–promoting bacteria. Environmental Science and Pollution Research 2021, 28, 51425–51439. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, X.; Wang, Y.; Zhu, B. A three-year experiment of annual methane and nitrous oxide emissions from the subtropical permanently flooded rice paddy fields of China: Emission factor, temperature sensitivity and fertilizer nitrogen effect. Agricultural and Forest Meteorology 2018, 250, 299–307. [Google Scholar] [CrossRef]
- Chidthaisong, A.; Cha-un, N.; Rossopa, B.; Buddaboon, C.; Kunuthai, C.; Sriphirom, P.; Towprayoon, S.; Tokida, T.; Padre, A.T.; Minamikawa, K. Evaluating the effects of alternate wetting and drying (AWD) on methane and nitrous oxide emissions from a paddy field in Thailand. Soil Science and Plant Nutrition 2018, 64, 31–38. [Google Scholar] [CrossRef]
- Ranatunga, T.; Hiramatsu, K.; Onishi, T.; Ishiguro, Y. Process of denitrification in flooded rice soils. Reviews in Agricultural Science 2018, 6, 21–33. [Google Scholar] [CrossRef]
- Jurado, A.; Borges, A.V.; Brouyère, S. Dynamics and emissions of N2O in groundwater: A review. Science of the Total Environment 2017, 584, 207–218. [Google Scholar] [CrossRef]
- Li, J.; Wang, S.; Luo, J.; Zhang, L.; Wu, Z.; Lindsey, S. Effects of biochar and 3, 4-dimethylpyrazole phosphate (DMPP) on soil ammonia-oxidizing bacteria and nos ZN 2 O reducers in the mitigation of N 2 O emissions from paddy soils. Journal of Soils and Sediments 2021, 21, 1089–1098. [Google Scholar] [CrossRef]
- He, M.; Tian, R.; Vancov, T.; Ma, F.; Fang, Y.; Liang, X. Response of N2O emission and denitrifying genes to iron (II) supplement in root zone and bulk region during wetting-drying alternation in paddy soil. Applied Soil Ecology 2024, 194, 105193. [Google Scholar] [CrossRef]
- Martikainen, P.J. Heterotrophic nitrification–An eternal mystery in the nitrogen cycle. Soil Biology and Biochemistry 2022, 168, 108611. [Google Scholar] [CrossRef]
- Nardi, P.; Laanbroek, H.J.; Nicol, G.W.; Renella, G.; Cardinale, M.; Pietramellara, G.; Weckwerth, W.; Trinchera, A.; Ghatak, A.; Nannipieri, P. Biological nitrification inhibition in the rhizosphere: determining interactions and impact on microbially mediated processes and potential applications. FEMS Microbiology Reviews 2020, 44, 874–908. [Google Scholar] [CrossRef]
- Jung, M.-Y.; Sedlacek, C.J.; Kits, K.D.; Mueller, A.J.; Rhee, S.-K.; Hink, L.; Nicol, G.W.; Bayer, B.; Lehtovirta-Morley, L.; Wright, C. Ammonia-oxidizing archaea possess a wide range of cellular ammonia affinities. The ISME journal 2022, 16, 272–283. [Google Scholar] [CrossRef]
- Mukhtar, H.; Lin, Y.-P.; Anthony, J. Ammonia oxidizing archaea and bacteria in east Asian paddy soils—A mini review. Environments 2017, 4, 84. [Google Scholar] [CrossRef]
- Wang, J.; Kan, J.; Zhang, X.; Xia, Z.; Zhang, X.; Qian, G.; Miao, Y.; Leng, X.; Sun, J. Archaea dominate the ammonia-oxidizing community in deep-sea sediments of the Eastern Indian Ocean—from the equator to the Bay of Bengal. Frontiers in microbiology 2017, 8, 415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Sun, H.-F.; Kang, Y.-C.; Jiang, R.; Wang, Y.-R.; Zhang, R.-Y.; Zhang, S.-N.; Li, Y.-Y.; Li, P.; Yang, F. Comammox plays a functionally important role in the nitrification of rice paddy soil with different nitrogen fertilization levels. Applied Soil Ecology 2024, 193, 105120. [Google Scholar] [CrossRef]
- Tang, H.; Xiao, X.; Li, C.; Cheng, K.; Pan, X.; Li, W. Effects of rhizosphere and long-term fertilisation practices on the activity and community structure of ammonia oxidisers under double-cropping rice field. Acta Agriculturae Scandinavica, Section B—Soil & Plant Science 2019, 69, 356–368. [Google Scholar]
- Wang, J.; Ni, L.; Song, Y.; Rhodes, G.; Li, J.; Huang, Q.; Shen, Q. Dynamic response of ammonia-oxidizers to four fertilization regimes across a wheat-rice rotation system. Frontiers in microbiology 2017, 8, 630. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ye, J.; Perez, P.G.; Huang, D.-F. Abundance and diversity of ammonia-oxidizing bacteria in rhizosphere and bulk paddy soil under different duration of organic management. African Journal of Microbiology Research 2011, 5, 5560–5568. [Google Scholar]
- Chen, S.; He, M.; Zhao, C.; Wang, W.; Zhu, Q.; Dan, X.; He, X.; Meng, L.; Zhang, S.; Cai, Z. Rice genotype affects nitrification inhibition in the rhizosphere. Plant and Soil 2022, 481, 35–48. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Y.; He, Y.; Liu, H.; Dumont, M.G.; Brookes, P.C.; Xu, J. Nitrosospira cluster 3-like bacterial ammonia oxidizers and Nitrospira-like nitrite oxidizers dominate nitrification activity in acidic terrace paddy soils. Soil Biology and Biochemistry 2019, 131, 229–237. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, Y.; Wright, A.L.; Shi, X.; Jiang, X. Different ammonia oxidizers are responsible for nitrification in two neutral paddy soils. Soil and Tillage Research 2019, 195, 104433. [Google Scholar] [CrossRef]
- Liu, H.; Li, J.; Zhao, Y.; Xie, K.; Tang, X.; Wang, S.; Li, Z.; Liao, Y.; Xu, J.; Di, H. Ammonia oxidizers and nitrite-oxidizing bacteria respond differently to long-term manure application in four paddy soils of south of China. Science of the Total Environment 2018, 633, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Ding, W.; Liu, D.; He, T.; Yoo, G.; Yuan, J.; Chen, Z.; Fan, J. Wheat straw-derived biochar amendment stimulated N2O emissions from rice paddy soils by regulating the amoA genes of ammonia-oxidizing bacteria. Soil Biology and Biochemistry 2017, 113, 89–98. [Google Scholar] [CrossRef]
- Duan, R.; Long, X.-E.; Tang, Y.-f.; Wen, J.; Su, S.; Bai, L.; Liu, R.; Zeng, X. Effects of different fertilizer application methods on the community of nitrifiers and denitrifiers in a paddy soil. Journal of soils and sediments 2018, 18, 24–38. [Google Scholar] [CrossRef]
- Su, Z.; Liu, T.; Guo, J.; Zheng, M. Nitrite oxidation in wastewater treatment: microbial adaptation and suppression challenges. Environmental Science & Technology 2023, 57, 12557–12570. [Google Scholar]
- Patil, P.M.; Parthasarathy, A.K.; Matkar, A.R.; Mahamuni-Badiger, P.; Hwang, S.; Gurav, R.; Dhanavade, M.J. Nitrite-oxidizing Bacteria: Cultivation, Growth Physiology, and Chemotaxonomy. 2023. [Google Scholar]
- Shan, J.; Sanford, R.A.; Chee-Sanford, J.; Ooi, S.K.; Löffler, F.E.; Konstantinidis, K.T.; Yang, W.H. Beyond denitrification: the role of microbial diversity in controlling nitrous oxide reduction and soil nitrous oxide emissions. Global change biology 2021, 27, 2669–2683. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, J.; Jiang, Y.; Hu, Y.; Zhang, M.; Zeng, Z. Response of bacteria harboring nirS and nirK genes to different N fertilization rates in an alkaline northern Chinese soil. European Journal of Soil Biology 2017, 82, 1–9. [Google Scholar] [CrossRef]
- Chen, Z.; Gao, X.; Xia, T.; Su, Y.; Liu, J.; Yu, L.; Wang, D.; Xu, S. Community Structure and Diversity of nirS-type Denitrifying Bacteria in Paddy Soils. 2018. [Google Scholar]
- Shan, J.; Yang, P.; Shang, X.; Rahman, M.M.; Yan, X. Anaerobic ammonium oxidation and denitrification in a paddy soil as affected by temperature, pH, organic carbon, and substrates. Biology and Fertility of Soils 2018, 54, 341–348. [Google Scholar] [CrossRef]
- Tan, X.; Shao, D.; Gu, W. Effects of temperature and soil moisture on gross nitrification and denitrification rates of a Chinese lowland paddy field soil. Paddy and Water Environment 2018, 16, 687–698. [Google Scholar] [CrossRef]
- Thakur, I.S.; Medhi, K. Nitrification and denitrification processes for mitigation of nitrous oxide from waste water treatment plants for biovalorization: Challenges and opportunities. Bioresource technology 2019, 282, 502–513. [Google Scholar] [CrossRef]
- Prosser, J.I.; Hink, L.; Gubry-Rangin, C.; Nicol, G.W. Nitrous oxide production by ammonia oxidizers: physiological diversity, niche differentiation and potential mitigation strategies. Global Change Biology 2020, 26, 103–118. [Google Scholar] [CrossRef]
- He, T.; Xie, D.; Ni, J.; Li, Z.; Li, Z. Nitrous oxide produced directly from ammonium, nitrate and nitrite during nitrification and denitrification. Journal of hazardous materials 2020, 388, 122114. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; De Vos, P.; Willems, A. Influence of nitrate and nitrite concentration on N2O production via dissimilatory nitrate/nitrite reduction to ammonium in Bacillus paralicheniformis LMG 6934. MicrobiologyOpen 2018, 7, e00592. [Google Scholar] [CrossRef] [PubMed]
- Roco, C.A.; Bergaust, L.L.; Bakken, L.R.; Yavitt, J.B.; Shapleigh, J.P. Modularity of nitrogen-oxide reducing soil bacteria: linking phenotype to genotype. Environmental Microbiology 2017, 19, 2507–2519. [Google Scholar] [CrossRef]
- Otte, J.M.; Blackwell, N.; Ruser, R.; Kappler, A.; Kleindienst, S.; Schmidt, C. N2O formation by nitrite-induced (chemo) denitrification in coastal marine sediment. Scientific Reports 2019, 9, 10691. [Google Scholar] [CrossRef] [PubMed]
- Conthe, M.; Wittorf, L.; Kuenen, J.G.; Kleerebezem, R.; van Loosdrecht, M.C.; Hallin, S. Life on N2O: deciphering the ecophysiology of N2O respiring bacterial communities in a continuous culture. The ISME journal 2018, 12, 1142–1153. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-y.; LU, S.-e.; XIANG, Q.-j.; YU, X.-m.; Ke, Z.; ZHANG, X.-p.; TU, S.-h.; GU, Y.-f. Responses of N2O reductase gene (nosZ)-denitrifier communities to long-term fertilization follow a depth pattern in calcareous purplish paddy soil. Journal of integrative agriculture 2017, 16, 2597–2611. [Google Scholar] [CrossRef]
- Pandey, A.; Suter, H.; He, J.-Z.; Hu, H.-W.; Chen, D. Nitrogen addition decreases dissimilatory nitrate reduction to ammonium in rice paddies. Applied and Environmental Microbiology 2018, 84, e00870–00818. [Google Scholar] [CrossRef]
- Liu, T.; Qin, S.; Pang, Y.; Yao, J.; Zhao, X.; Clough, T.; Wrage-Mönnig, N.; Zhou, S. Rice root Fe plaque enhances paddy soil N2O emissions via Fe (II) oxidation-coupled denitrification. Soil Biology and Biochemistry 2019, 139, 107610. [Google Scholar] [CrossRef]
- Xiang, H.; Hong, Y.; Wu, J.; Wang, Y.; Ye, F.; Ye, J.; Lu, J.; Long, A. Denitrification contributes to N2O emission in paddy soils. Frontiers in Microbiology 2023, 14, 1218207. [Google Scholar] [CrossRef]
- Farooq, M.S.; Uzair, M.; Maqbool, Z.; Fiaz, S.; Yousuf, M.; Yang, S.H.; Khan, M.R. Improving nitrogen use efficiency in aerobic rice based on insights into the ecophysiology of archaeal and bacterial ammonia oxidizers. Frontiers in Plant Science 2022, 13, 913204. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, X.; Zhang, Q. Effects of combined biochar and organic fertilizer on nitrous oxide fluxes and the related nitrifier and denitrifier communities in a saline-alkali soil. Science of the total environment 2019, 686, 199–211. [Google Scholar] [CrossRef] [PubMed]
- TENORIO, J.I.R. WHEAT CULTIVAR INFLUENCE OVER NITROGEN-FIXING BACTERIAL COMMUNITIES IN AN ANDISOL AND A NOVEL PGPB TRACKING METHOD FOR AZOSPIRILLUM SPP. BASED ON CRISPR LOCI-TARGETED PCR. UNIVERSIDAD DE LA FRONTERA, 2020.
- Masocha, B.L.; Dikinya, O.; Moseki, B. Bioavailability and contamination levels of Zn, Pb, and Cd in sandy-loam soils, Botswana. Environmental Earth Sciences 2022, 81, 171. [Google Scholar] [CrossRef]
- Kwiatkowska-Malina, J. Functions of organic matter in polluted soils: The effect of organic amendments on phytoavailability of heavy metals. Applied Soil Ecology 2018, 123, 542–545. [Google Scholar] [CrossRef]
- Ondrasek, G.; Begić, H.B.; Zovko, M.; Filipović, L.; Meriño-Gergichevich, C.; Savić, R.; Rengel, Z. Biogeochemistry of soil organic matter in agroecosystems & environmental implications. Science of the total environment 2019, 658, 1559–1573. [Google Scholar] [PubMed]
- Shahid, M.; Shamshad, S.; Rafiq, M.; Khalid, S.; Bibi, I.; Niazi, N.K.; Dumat, C.; Rashid, M.I. Chromium speciation, bioavailability, uptake, toxicity and detoxification in soil-plant system: A review. Chemosphere 2017, 178, 513–533. [Google Scholar] [CrossRef] [PubMed]
- Podar, D.; Maathuis, F.J. The role of roots and rhizosphere in providing tolerance to toxic metals and metalloids. Plant, cell & environment 2022, 45, 719–736. [Google Scholar]
- Kuzyakov, Y.; Razavi, B.S. Rhizosphere size and shape: Temporal dynamics and spatial stationarity. Soil Biology and Biochemistry 2019, 135, 343–360. [Google Scholar] [CrossRef]
- Agarwal, P.; Vibhandik, R.; Agrahari, R.; Daverey, A.; Rani, R. Role of Root Exudates on the Soil Microbial Diversity and Biogeochemistry of Heavy Metals. Applied Biochemistry and Biotechnology 2023, 1–21. [Google Scholar] [CrossRef]
- Kaur, H.; Garg, N. Microbial community in soil-plant systems: Role in heavy metal (loid) detoxification and sustainable agriculture. In Rhizosphere Engineering; Elsevier, 2022; pp. 471–498. [Google Scholar]
- Jing, R.; Kjellerup, B.V. Biogeochemical cycling of metals impacting by microbial mobilization and immobilization. Journal of Environmental Sciences 2018, 66, 146–154. [Google Scholar] [CrossRef]
- Shan, B.; Hao, R.; Xu, H.; Li, J.; Li, Y.; Xu, X.; Zhang, J. A review on mechanism of biomineralization using microbial-induced precipitation for immobilizing lead ions. Environmental Science and Pollution Research 2021, 28, 30486–30498. [Google Scholar] [CrossRef]
- Xiao, Y.; Chen, L.; Teng, K.; Ma, J.; Xiang, S.; Jiang, L.; Liu, G.; Yang, B.; Fang, J. Potential roles of the rhizospheric bacterial community in assisting Miscanthus floridulus in remediating multi-metal (loid) s contaminated soils. Environmental Research 2023, 227, 115749. [Google Scholar] [CrossRef]
- García-Palacios, P.; Crowther, T.W.; Dacal, M.; Hartley, I.P.; Reinsch, S.; Rinnan, R.; Rousk, J.; van den Hoogen, J.; Ye, J.-S.; Bradford, M.A. Evidence for large microbial-mediated losses of soil carbon under anthropogenic warming. Nature Reviews Earth & Environment 2021, 2, 507–517. [Google Scholar]
- Zhang, G.; Bai, J.; Zhai, Y.; Jia, J.; Zhao, Q.; Wang, W.; Hu, X. Microbial diversity and functions in saline soils: A review from a biogeochemical perspective. Journal of Advanced Research 2023. [Google Scholar] [CrossRef]
- Abdu, N.; Abdullahi, A.A.; Abdulkadir, A. Heavy metals and soil microbes. Environmental chemistry letters 2017, 15, 65–84. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, H.; Zhang, Y.; Fang, X.; Zhong, X.; Lv, J. Contribution of ammonia-oxidizing archaea and bacteria to nitrogen transformation in a soil fertilized with urea and organic amendments. Scientific Reports 2023, 13, 20722. [Google Scholar] [CrossRef]
- Fait, G.; Broos, K.; Zrna, S.; Lombi, E.; Hamon, R. Tolerance of nitrifying bacteria to copper and nickel. Environ Toxicol Chem 2006, 25, 2000–2005. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Ding, Y.; Zheng, J.; Zhou, T.; Pan, G.; Crowley, D.; Li, L.; Zheng, J.; Zhang, X.; et al. Abundance, Composition and Activity of Ammonia Oxidizer and Denitrifier Communities in Metal Polluted Rice Paddies from South China. PLOS ONE 2014, 9, e102000. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xue, C.; Yu, S.; Li, F. Variations of abundance and community structure of ammonia oxidizers and nitrification activity in two paddy soils polluted by heavy metals. Geomicrobiology Journal 2019, 36, 1–10. [Google Scholar] [CrossRef]
- Li, X.; Meng, D.; Li, J.; Yin, H.; Liu, H.; Liu, X.; Cheng, C.; Xiao, Y.; Liu, Z.; Yan, M. Response of soil microbial communities and microbial interactions to long-term heavy metal contamination. Environmental Pollution 2017, 231, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Chen, C.; Ke, T.; Wang, M.; Sima, M.; Huang, S. Long-term metal pollution shifts microbial functional profiles of nitrification and denitrification in agricultural soils. Science of The Total Environment 2022, 830, 154732. [Google Scholar] [CrossRef]
- Liu, Y.; Shen, K.; Wu, Y.; Wang, G. Abundance and structure composition of nirK and nosZ genes as well as denitrifying activity in heavy metal-polluted paddy soils. Geomicrobiology Journal 2018, 35, 100–107. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Zhou, H.; Li, L.; Zheng, J.; Zhang, X.; Zheng, J.; Pan, G. Abundance, composition and activity of denitrifier communities in metal polluted paddy soils. Scientific Reports 2016, 6, 19086. [Google Scholar] [CrossRef] [PubMed]
- Ruyters, S.; Mertens, J.; T’Seyen, I.; Springael, D.; Smolders, E. Dynamics of the nitrous oxide reducing community during adaptation to Zn stress in soil. Soil Biology and Biochemistry 2010, 42, 1581–1587. [Google Scholar] [CrossRef]
- Tang, S.; Rao, Y.; Huang, S.; Xu, Y.; Zeng, K.; Liang, X.; Ling, Q.; Liu, K.; Ma, J.; Yu, F. Impact of environmental factors on the ammonia-oxidizing and denitrifying microbial community and functional genes along soil profiles from different ecologically degraded areas in the Siding mine. Journal of Environmental Management 2023, 326, 116641. [Google Scholar] [CrossRef] [PubMed]
- Chai, R.; Ye, X.; Ma, C.; Wang, Q.; Tu, R.; Zhang, L.; Gao, H. Greenhouse gas emissions from synthetic nitrogen manufacture and fertilization for main upland crops in China. Carbon balance and management 2019, 14, 1–10. [Google Scholar] [CrossRef]
- Kahrl, F.; Li, Y.; Su, Y.; Tennigkeit, T.; Wilkes, A.; Xu, J. Greenhouse gas emissions from nitrogen fertilizer use in China. Environmental Science & Policy 2010, 13, 688–694. [Google Scholar] [CrossRef]
- Robertson, G.P.; Groffman, P.M. Chapter 14 - Nitrogen transformations. In Soil Microbiology, Ecology and Biochemistry (Fifth Edition); Paul, E.A., Frey, S.D., Eds.; Elsevier, 2024; pp. 407–438. [Google Scholar]
- Kuypers, M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nature Reviews Microbiology 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- NIE, Z.-j.; QIN, S.-y.; LIU, H.-e.; ZHAO, P.; WU, X.-t.; GAO, W.; LI, C.; ZHANG, W.-w.; SUI, F.-q. Effects of combined application of nitrogen and zinc on winter wheat yield and soil enzyme activities related to nitrogen transformation. Journal of Plant Nutrition and Fertilizers 2020, 26, 431–441. [Google Scholar]
- Zhang, Y.; Xu, B.; Han, J.; Shi, L. Effects of drying-rewetting cycles on ferrous iron-involved denitrification in paddy soils. Water 2021, 13, 3212. [Google Scholar] [CrossRef]
- Kim, G.W.; Kim, P.J.; Khan, M.I.; Lee, S.-J. Effect of rice planting on nitrous oxide (N2O) emission under different levels of nitrogen fertilization. Agronomy 2021, 11, 217. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, S.; Wang, X.; Liu, C.; Zhu, G. Contribution of Ammonium-Induced Nitrifier Denitrification to N2O in Paddy Fields. Environmental Science & Technology 2023, 57, 2970–2980. [Google Scholar]
- Zou, J.; Huang, Y.; Zheng, X.; Wang, Y. Quantifying direct N2O emissions in paddy fields during rice growing season in mainland China: Dependence on water regime. Atmospheric Environment 2007, 41, 8030–8042. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, J.; Zhao, B.; Yan, P.; Zhou, G.; Xin, X. Effects of straw amendment and moisture on microbial communities in Chinese fluvo-aquic soil. Journal of Soils and Sediments 2014, 14, 1829–1840. [Google Scholar] [CrossRef]
- Qin, C.; Yuan, X.; Xiong, T.; Tan, Y.Z.; Wang, H. Physicochemical properties, metal availability and bacterial community structure in heavy metal-polluted soil remediated by montmorillonite-based amendments. Chemosphere 2020, 261, 128010. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, P.; Bellitürk, K.; Görres, J.H. Phytoremediation of heavy metal-contaminated soil by switchgrass: A comparative study utilizing different composts and coir fiber on pollution remediation, plant productivity, and nutrient leaching. International Journal of Environmental Research and Public Health 2019, 16, 1261. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Hesham Ael, L.; Qiao, M.; Rehman, S.; He, J.Z. Effects of Cd and Pb on soil microbial community structure and activities. Environ Sci Pollut Res Int 2010, 17, 288–296. [Google Scholar] [CrossRef]
- Liu, J.; Cao, W.; Jiang, H.; Cui, J.; Shi, C.; Qiao, X.; Zhao, J.; Si, W. Impact of heavy metal pollution on ammonia oxidizers in soils in the vicinity of a Tailings Dam, Baotou, China. Bulletin of environmental contamination and toxicology 2018, 101, 110–116. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Balgobind, A.; Pillay, B. Bioavailability of heavy metals in soil: impact on microbial biodegradation of organic compounds and possible improvement strategies. Int J Mol Sci 2013, 14, 10197–10228. [Google Scholar] [CrossRef]
- Ke, Y.; Si, S.; Zhang, Z.; Geng, P.; Shen, Y.; Wang, J.; Zhu, X. Synergistic passivation performance of cadmium pollution by biochar combined with sulfate reducing bacteria. Environmental Technology & Innovation 2023, 32, 103356. [Google Scholar]
- Meng, D.; Li, J.; Liu, T.; Liu, Y.; Yan, M.; Hu, J.; Li, X.; Liu, X.; Liang, Y.; Liu, H. Effects of redox potential on soil cadmium solubility: insight into microbial community. Journal of Environmental Sciences 2019, 75, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Flandes, S.; González, B.; Ulloa, O. Redox traits characterize the organization of global microbial communities. Proceedings of the National Academy of Sciences 2019, 116, 3630–3635. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, W.; Li, J.; Song, Y.; Hua, M.; Li, W.; Wen, Y.; Li, T.; He, X. Assessing the fractionation and bioavailability of heavy metals in soil–rice system and the associated health risk. Environmental Geochemistry and Health 2021, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Wu, L.; Luo, Y.; Christie, P. Effects of organic matter fraction and compositional changes on distribution of cadmium and zinc in long-term polluted paddy soils. Environmental Pollution 2018, 232, 514–522. [Google Scholar] [CrossRef]
- Hussain, B.; Ashraf, M.N.; Abbas, A.; Li, J.; Farooq, M. Cadmium stress in paddy fields: effects of soil conditions and remediation strategies. Science of The Total Environment 2021, 754, 142188. [Google Scholar] [CrossRef]
- Łukowski, A.; Dec, D. Influence of Zn, Cd, and Cu fractions on enzymatic activity of arable soils. Environmental monitoring and assessment 2018, 190, 1–12. [Google Scholar] [CrossRef]
- Haynes, R.J.; Zhou, Y.-F. Chapter Five - Retention of heavy metals by dredged sediments and their management following land application. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press, 2022; Volume 171, pp. 191–254. [Google Scholar]
- Hiller, E.; Šutriepka, M. Effect of drying on the sorption and desorption of copper in bottom sediments from water reservoirs and geochemical partitioning of heavy metals and arsenic. J Hydrol Hydromech 2008, 55, 45–58. [Google Scholar]
- Koopmans, G.F.; Groenenberg, J.E. Effects of soil oven-drying on concentrations and speciation of trace metals and dissolved organic matter in soil solution extracts of sandy soils. Geoderma 2011, 161, 147–158. [Google Scholar] [CrossRef]
- Xu JunZeng, X.J.; Lv YuPing, L.Y.; Yang ShiHong, Y.S.; Wei Qi, W.Q.; Qiao ZhenFang, Q.Z. Water saving irrigation improves the solubility and bioavailability of zinc in rice paddy. 2015. [Google Scholar]
- Li, J.; Xu, Y. Effects of clay combined with moisture management on Cd immobilization and fertility index of polluted rice field. Ecotoxicology and Environmental Safety 2018, 158, 182–186. [Google Scholar] [CrossRef]
- Xu, J.; Peng, S.; Qiao, Z.; Yang, S.; Gao, X. Binding forms and availability of Cd and Cr in paddy soil under non-flooding controlled irrigation. Paddy and water environment 2014, 12, 213–222. [Google Scholar] [CrossRef]
- Frohne, T.; Rinklebe, J.; Diaz-Bone, R.A.; Du Laing, G. Controlled variation of redox conditions in a floodplain soil: Impact on metal mobilization and biomethylation of arsenic and antimony. Geoderma 2011, 160, 414–424. [Google Scholar] [CrossRef]
- Arao, T.; Kawasaki, A.; Baba, K.; Mori, S.; Matsumoto, S. Effects of water management on cadmium and arsenic accumulation and dimethylarsinic acid concentrations in Japanese rice. Environmental Science & Technology 2009, 43, 9361–9367. [Google Scholar]
- Lee, K.; Bae, D.W.; Kim, S.H.; Han, H.J.; Liu, X.; Park, H.C.; Lim, C.O.; Lee, S.Y.; Chung, W.S. Comparative proteomic analysis of the short-term responses of rice roots and leaves to cadmium. Journal of Plant Physiology 2010, 167, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, Y.; Yu, L.; Yang, M.; Zou, X.; Yin, C.; Lin, Y. Research advances in cadmium uptake, transport and resistance in rice (Oryza sativa L.). Cells 2022, 11, 569. [Google Scholar] [CrossRef] [PubMed]
- Sterckeman, T.; Thomine, S. Mechanisms of cadmium accumulation in plants. Critical Reviews in Plant Sciences 2020, 39, 322–359. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Adrees, M.; Rizvi, H.; Zia-ur-Rehman, M.; Hannan, F.; Qayyum, M.F.; Hafeez, F.; Ok, Y.S. Cadmium stress in rice: toxic effects, tolerance mechanisms, and management: a critical review. Environmental Science and Pollution Research 2016, 23, 17859–17879. [Google Scholar] [CrossRef]
- Shaari, N.; Tajudin, M.; Khandaker, M.; Majrashi, A.; Alenazi, M.; Abdullahi, U.; Mohd, K. Cadmium toxicity symptoms and uptake mechanism in plants: a review. Brazilian Journal of Biology 2022, 84, e252143. [Google Scholar] [CrossRef]
- Sarwar, N.; Saifullah; Malhi, S.S.; Zia, M.H.; Naeem, A.; Bibi, S.; Farid, G. Role of mineral nutrition in minimizing cadmium accumulation by plants. Journal of the Science of Food and Agriculture 2010, 90, 925–937. [Google Scholar] [CrossRef]
- Waheed, A.; Haxim, Y.; Islam, W.; Ahmad, M.; Ali, S.; Wen, X.; Khan, K.A.; Ghramh, H.A.; Zhang, Z.; Zhang, D. Impact of Cadmium Stress on Growth and Physio-Biochemical Attributes of Eruca sativa Mill. Plants (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Engineer, C.B.; Hashimoto-Sugimoto, M.; Negi, J.; Israelsson-Nordström, M.; Azoulay-Shemer, T.; Rappel, W.J.; Iba, K.; Schroeder, J.I. CO2 Sensing and CO2 Regulation of Stomatal Conductance: Advances and Open Questions. Trends Plant Sci 2016, 21, 16–30. [Google Scholar] [CrossRef]
- Sharma, S.; Joshi, J.; Kataria, S.; Verma, S.K.; Chatterjee, S.; Jain, M.; Pathak, K.; Rastogi, A.; Brestic, M. Regulation of the Calvin cycle under abiotic stresses: An overview. Plant life under changing environment 2020, 681–717. [Google Scholar]
- Dikšaitytė, A.; Kniuipytė, I.; Žaltauskaitė, J.; Abdel-Maksoud, M.A.; Asard, H.; AbdElgawad, H. Enhanced Cd phytoextraction by rapeseed under future climate as a consequence of higher sensitivity of HMA genes and better photosynthetic performance. Science of The Total Environment 2024, 908, 168164. [Google Scholar] [CrossRef]
- Guo, L.; Chen, A.; He, N.; Yang, D.; Liu, M. Exogenous silicon alleviates cadmium toxicity in rice seedlings in relation to Cd distribution and ultrastructure changes. Journal of soils and sediments 2018, 18, 1691–1700. [Google Scholar] [CrossRef]
- Wang, J.; Li, D.; Lu, Q.; Zhang, Y.; Xu, H.; Wang, X.; Li, Y. Effect of water-driven changes in rice rhizosphere on Cd lability in three soils with different pH. Journal of Environmental Sciences 2020, 87, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, S.; Bao, Q.; Chu, Y.; Sun, H.; Huang, Y. Jasmonic acid alleviates cadmium toxicity through regulating the antioxidant response and enhancing the chelation of cadmium in rice (Oryza sativa L.). Environmental Pollution 2022, 304, 119178. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Rehman, A.; Shahzad, B.; Tanveer, M.; Saud, S.; Kamran, M.; Ihtisham, M.; Khan, S.U.; Turan, V.; ur Rahman, M.H. Rice responses and tolerance to metal/metalloid toxicity. In Advances in rice research for abiotic stress tolerance; Elsevier, 2019; pp. 299–312. [Google Scholar]
- Yotsova, E.K.; Dobrikova, A.G.; Stefanov, M.A.; Kouzmanova, M.; Apostolova, E.L. Improvement of the rice photosynthetic apparatus defence under cadmium stress modulated by salicylic acid supply to roots. Theoretical and Experimental Plant Physiology 2018, 30, 57–70. [Google Scholar] [CrossRef]
- Ninkovic, V.; Markovic, D.; Dahlin, I. Decoding neighbour volatiles in preparation for future competition and implications for tritrophic interactions. Perspectives in Plant Ecology, Evolution and Systematics 2016, 23, 11–17. [Google Scholar] [CrossRef]
- Abbas, F.; O’Neill Rothenberg, D.; Zhou, Y.; Ke, Y.; Wang, H.C. Volatile organic compounds as mediators of plant communication and adaptation to climate change. Physiologia Plantarum 2022, 174, e13840. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Staudt, M. BVOCs and global change. Trends in Plant Science 2010, 15, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.M.; Blande, J.D.; Souza, S.R.; Nerg, A.M.; Holopainen, J.K. Plant volatile organic compounds (VOCs) in ozone (O3) polluted atmospheres: the ecological effects. J Chem Ecol 2010, 36, 22–34. [Google Scholar] [CrossRef]
- Loreto, F.; Schnitzler, J.P. Abiotic stresses and induced BVOCs. Trends Plant Sci 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Lin, T.; Zhu, G.; He, W.; Xie, J.; Li, S.; Han, S.; Li, S.; Yang, C.; Liu, Y.; Zhu, T. Soil cadmium stress reduced host plant odor selection and oviposition preference of leaf herbivores through the changes in leaf volatile emissions. Science of The Total Environment 2022, 814, 152728. [Google Scholar] [CrossRef] [PubMed]
- Bibbiani, S.; Colzi, I.; Taiti, C.; Nissim, W.G.; Papini, A.; Mancuso, S.; Gonnelli, C. Smelling the metal: volatile organic compound emission under Zn excess in the mint Tetradenia riparia. Plant Science 2018, 271, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Huang, J.; Ouyang, Y.; Wu, L.; Song, J.; Wang, S.; Li, Z.; Han, C.; Zhou, L.; Huang, Y.; et al. Water management affects arsenic and cadmium accumulation in different rice cultivars. Environ Geochem Health 2013, 35, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.; Zhao, J.; Chen, W.; Lu, L.; Chen, J.; Lin, Z.; Qiao, Y.; Lin, H.; Tian, S. Sulfur and water management mediated iron plaque and rhizosphere microorganisms reduced cadmium accumulation in rice. Journal of Soils and Sediments 2023, 1–14. [Google Scholar] [CrossRef]
- Sasaki, K.; Saito, T.; Lämsä, M.; Oksman-Caldentey, K.M.; Suzuki, M.; Ohyama, K.; Muranaka, T.; Ohara, K.; Yazaki, K. Plants utilize isoprene emission as a thermotolerance mechanism. Plant Cell Physiol 2007, 48, 1254–1262. [Google Scholar] [CrossRef] [PubMed]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus × acerifolia plants during Mediterranean summers. New Phytol 2015, 207, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant volatile organic compounds evolution: Transcriptional regulation, epigenetics and polyploidy. International Journal of Molecular Sciences 2020, 21, 8956. [Google Scholar] [CrossRef] [PubMed]
- Delory, B.M.; Delaplace, P.; Fauconnier, M.-L.; du Jardin, P. Root-emitted volatile organic compounds: can they mediate belowground plant-plant interactions? Plant and Soil 2016, 402, 1–26. [Google Scholar] [CrossRef]
- Duc, N.H.; Vo, H.T.; van Doan, C.; Hamow, K.A.; Le, K.H.; Posta, K. Volatile organic compounds shape belowground plant–fungi interactions. Frontiers in Plant Science 2022, 13, 1046685. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: from biosynthesis to function. Plant Cell Environ 2014, 37, 1936–1949. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytologist 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Noel, J.P.; Dudareva, N. Biosynthesis of plant volatiles: nature’s diversity and ingenuity. Science 2006, 311, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T.; Halitschke, R.; Paschold, A.; von Dahl, C.C.; Preston, C.A. Volatile signaling in plant-plant interactions: “talking trees” in the genomics era. Science 2006, 311, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms. Molecules 2017, 22, 1148. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusia, J.; Peñuelas, J. Floral volatile organic compounds: Between attraction and deterrence of visitors under global change. Perspectives in Plant Ecology, Evolution and Systematics 2013, 15, 56–67. [Google Scholar] [CrossRef]
- Waelti, M.O.; Page, P.A.; Widmer, A.; Schiestl, F.P. How to be an attractive male: floral dimorphism and attractiveness to pollinators in a dioecious plant. BMC Evolutionary Biology 2009, 9, 190. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Adrees, M.; Rizvi, H.; Zia-ur-Rehman, M.; Hannan, F.; Qayyum, M.F.; Hafeez, F.; Ok, Y.S. Cadmium stress in rice: toxic effects, tolerance mechanisms, and management: a critical review. Environmental Science and Pollution Research 2016, 23, 17859–17879. [Google Scholar] [CrossRef]
- Chen, S.; Yang, Y.; Shi, W.; Ji, Q.; He, F.; Zhang, Z.; Cheng, Z.; Liu, X.; Xu, M. Badh2, encoding betaine aldehyde dehydrogenase, inhibits the biosynthesis of 2-acetyl-1-pyrroline, a major component in rice fragrance. Plant Cell 2008, 20, 1850–1861. [Google Scholar] [CrossRef]
- Yu, E.; Wang, W.; Yamaji, N.; Fukuoka, S.; Che, J.; Ueno, D.; Ando, T.; Deng, F.; Hori, K.; Yano, M.; et al. Duplication of a manganese/cadmium transporter gene reduces cadmium accumulation in rice grain. Nature Food 2022, 3, 597–607. [Google Scholar] [CrossRef]
Figure 1.
Paddy soil N transformation under different water management practices and Cd contamination.
Figure 1.
Paddy soil N transformation under different water management practices and Cd contamination.
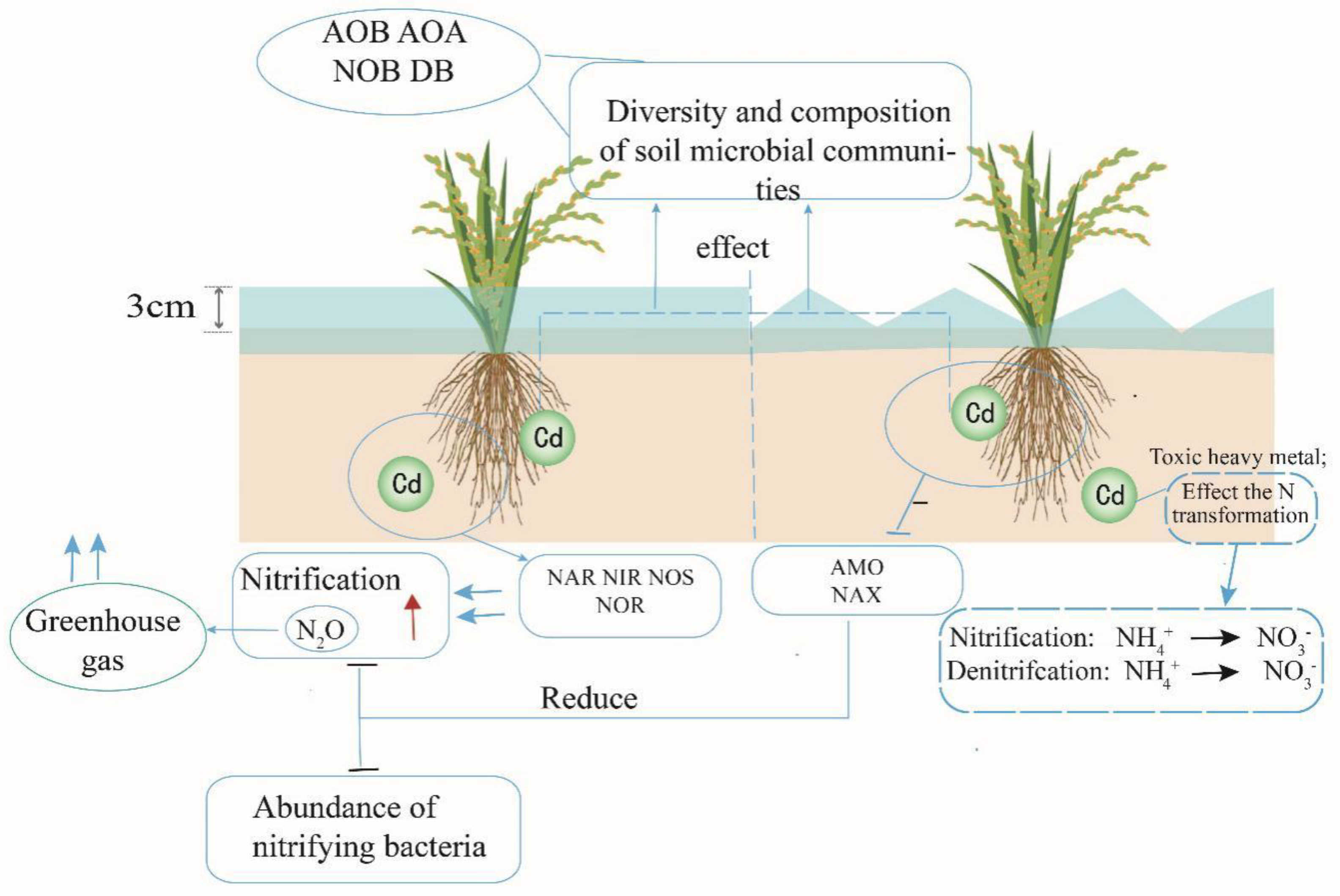
Figure 2.
Effects of SWMP on rice VOCs under Cd contamination.
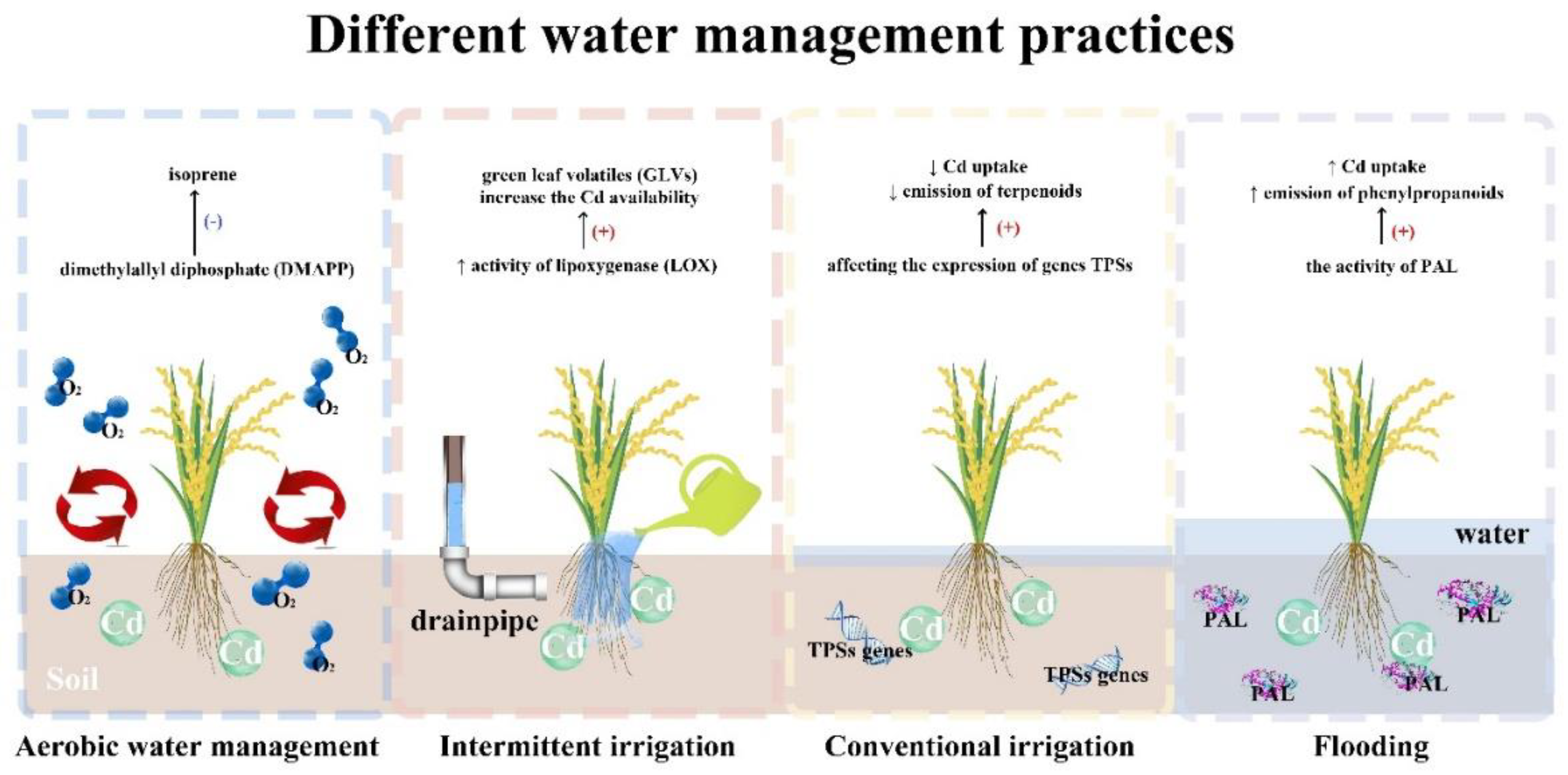
Figure 3.
The effect of Cd on rice genes responsible for fragrance.
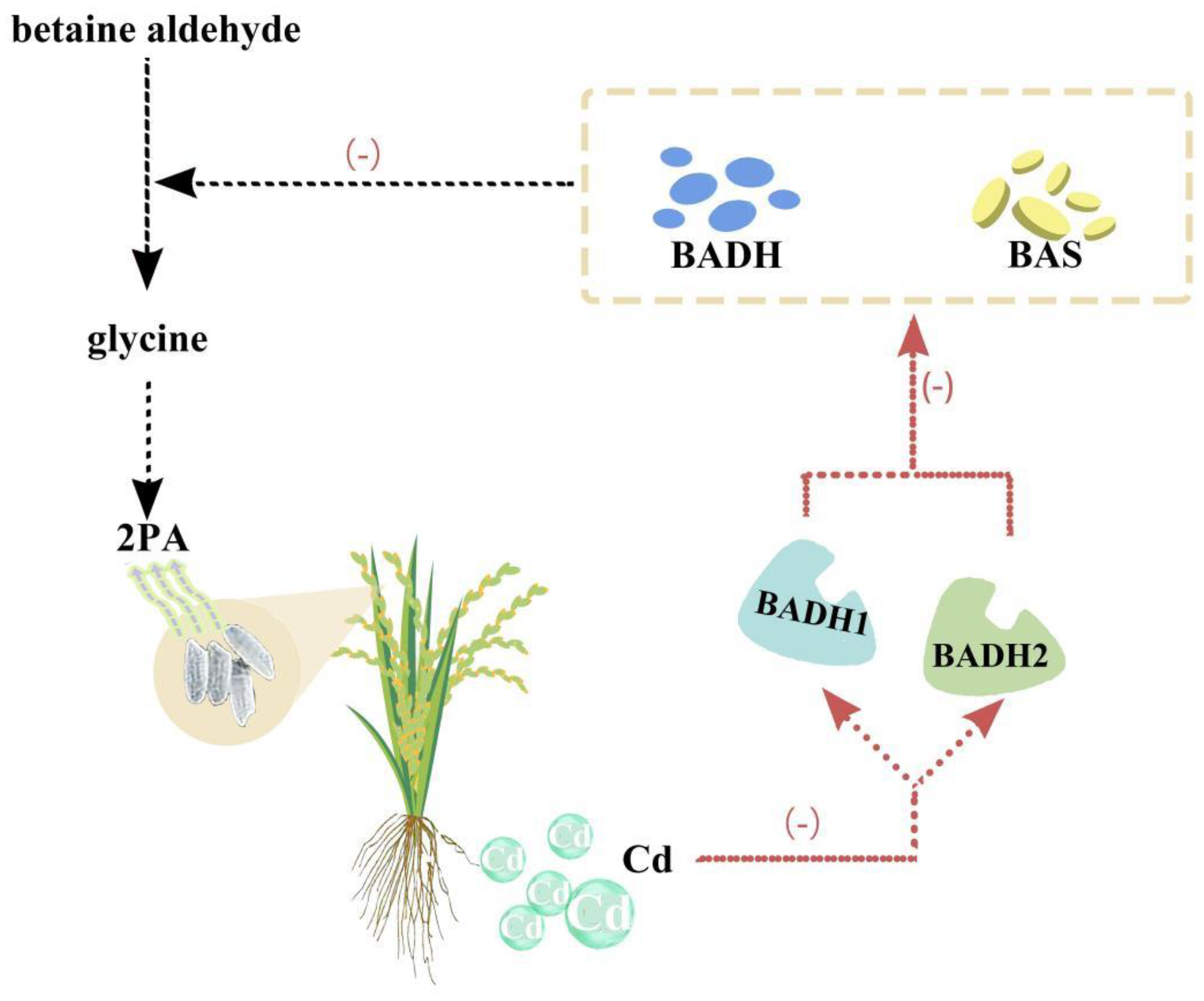
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



