
Submitted:
08 February 2024
Posted:
09 February 2024
You are already at the latest version
Abstract
Thymomas (TH) are a unique group of heterogeneous tumors of the thymic epithelium. In particular, the subtypes B2 and B3 tend to be aggressive and metastatic. Radical tumor resection remains the only curative option for localized tumors, while more advanced TH require multimodal treatment. Deep sequencing analyses have failed to identify known oncogenic driver mutations in TH, with the notable exception of the GTF2I mutation, which occurs predominantly in type A and AB TH. However, there are multiple alternative non-mutational mechanisms (e.g., perturbed thymic developmental programs, metabolism, non-coding RNA networks) that control cellular behavior and tumorigenesis through deregulation of critical molecular pathways. Here, we attempted to show how the results of studies investigating such alternative mechanisms could be integrated into a current model of TH biology. This model could be used to focus ongoing research and therapeutic strategies.
Keywords:
Thymus
; thymoma
; oncogenic driver
; mutations
; chromosomal aberrations
; RNA
; methylation
; gene expression
; apoptosis
; proteomics
; metabolomics
1. Introduction
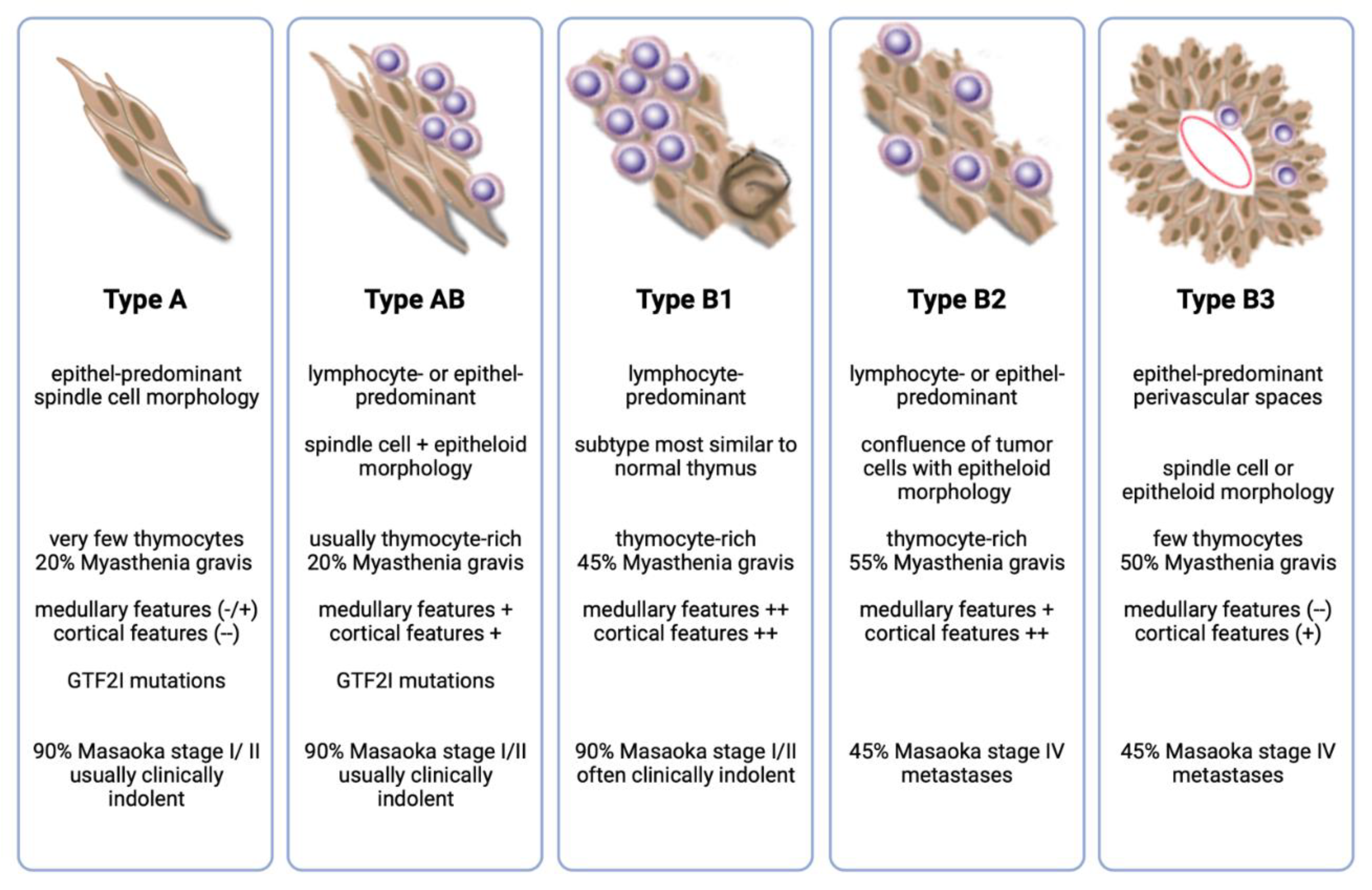
Thymomas (TH) are rare epithelial tumors of the anterior mediastinum. Their most characteristic common feature is the variable preservation of important "organotypic" features of the normal thymus, in particular, the ability to promote the maturation of T cells from immature precursors [1]. Based on the morphology of the neoplastic epithelial cells and the relative proportion of immature T cells, the World Health Organization (WHO) recognizes five main TH subtypes (A, AB, B1, B2, and B3 ) with different epidemiologic, histologic, and clinical characteristics, such as their propensity to present in advanced stages or to metastasize [2] (Figure 1).
Probably due to their ability to promote and export tumor-derived T cells, approximately 40% of TH are associated with paraneoplastic autoimmune diseases, the most common of which is myasthenia gravis (MG) [3]. These organotypic features distinguish TH from the more aggressive thymic carcinomas (TC), which have lost the ability to promote T cell maturation and are therefore not associated with MG. Another highly characteristic and fundamental difference between TH and TC is their pattern of progression, with prolonged confinement to the thoracic cavity and the virtual absence of lymph node metastasis in TH. Molecular data from The Cancer Genome Atlas (TCGA) program have confirmed that TH and TC are distinct entities [4]. Among TH, the data suggest that type A and AB are distantly related neoplasms. In contrast, type B thymomas form a different cluster that appears to represent a spectrum rather than separate entities, an observation supported by the frequent finding of more than one type B component in a given tumor. These observations are important since they point to different molecular routes in the pathogenesis of the four main thymic epithelial tumor (TET) groups. While many THs are slow-growing and relatively indolent neoplasms, type B2 and B3 THs are more aggressive and may present in advanced clinical stages with infiltration of adjacent anatomic structures or intrathoracic metastases. Advanced TH are treated by a multimodal approach with systemic chemotherapy, surgery, and radiotherapy [5]. The TCGA study has confirmed the notion that THs are among the adult tumors with the lowest mutational burden and very few recurrent gene mutations, with the notable exception of GTF2I mutations, which are preferentially found in type A and AB thymomas [4,6]. This observation may explain why, so far, there has been little benefit from molecularly targeted approaches in patients with TH, even compared to TC, where there are now a couple of therapeutic options available. However, while this may have come as a disappointment to many, a vast body of literature over the decades has helped to pave the way towards a better understanding of the driving factors behind the biology of TH. Based on current knowledge, it seems plausible to assume that TET develop along (at least) three different molecular routes: A. type A and AB thymomas with few chromosomal alterations but with a highly specific and recurrent GTF2I mutation as a fundamental driver, B. type B thymomas with increasing numbers of chromosomal alterations, but so far no known driver mutations, suggesting alternative, non-mutational mechanisms determining their behavior, and C. thymic carcinomas with multiple chromosomal alterations and several recurrent gene mutations (reviewed elsewhere in this Cancers Special Issue). Through multiple subtype-specific mechanisms (Figure 1), these alternative molecular routes appear to converge in at least 7 critical cellular pathways that can be considered the hallmarks of TH biology: 1. Wnt signaling, 2. altered receptor tyrosine kinase (RTK) signaling, 3. RAS/MAPK/ERK signaling, 4. PI3K/AKT/mTOR signaling, 5. cell cycle regulation, 6. cellular senescence, and apoptosis resistance, and 7. epigenetic regulation. This list is probably incomplete, and other mechanisms may emerge as modern bioinformatic analysis of multi-omics helps better manage the available data's complexity. In this narrative review, we will try to summarize and highlight the current knowledge about the non-mutational drivers of TH. The fact that thymomas are not only malignant neoplasms but are also uniquely capable of distorting the immune system adds another layer of complexity that is intrinsic to the biology of these tumors and cannot be ignored but is beyond the scope of this review. Immunotherapy in TH is described elsewhere in this special issue of Cancers.
Figure 2.
Non-mutational mechanisms and "hallmark" molecular pathways in thymomas. Perturbations at different levels of cellular control converge into deregulation of critical signaling pathways.
Figure 2.
Non-mutational mechanisms and "hallmark" molecular pathways in thymomas. Perturbations at different levels of cellular control converge into deregulation of critical signaling pathways.
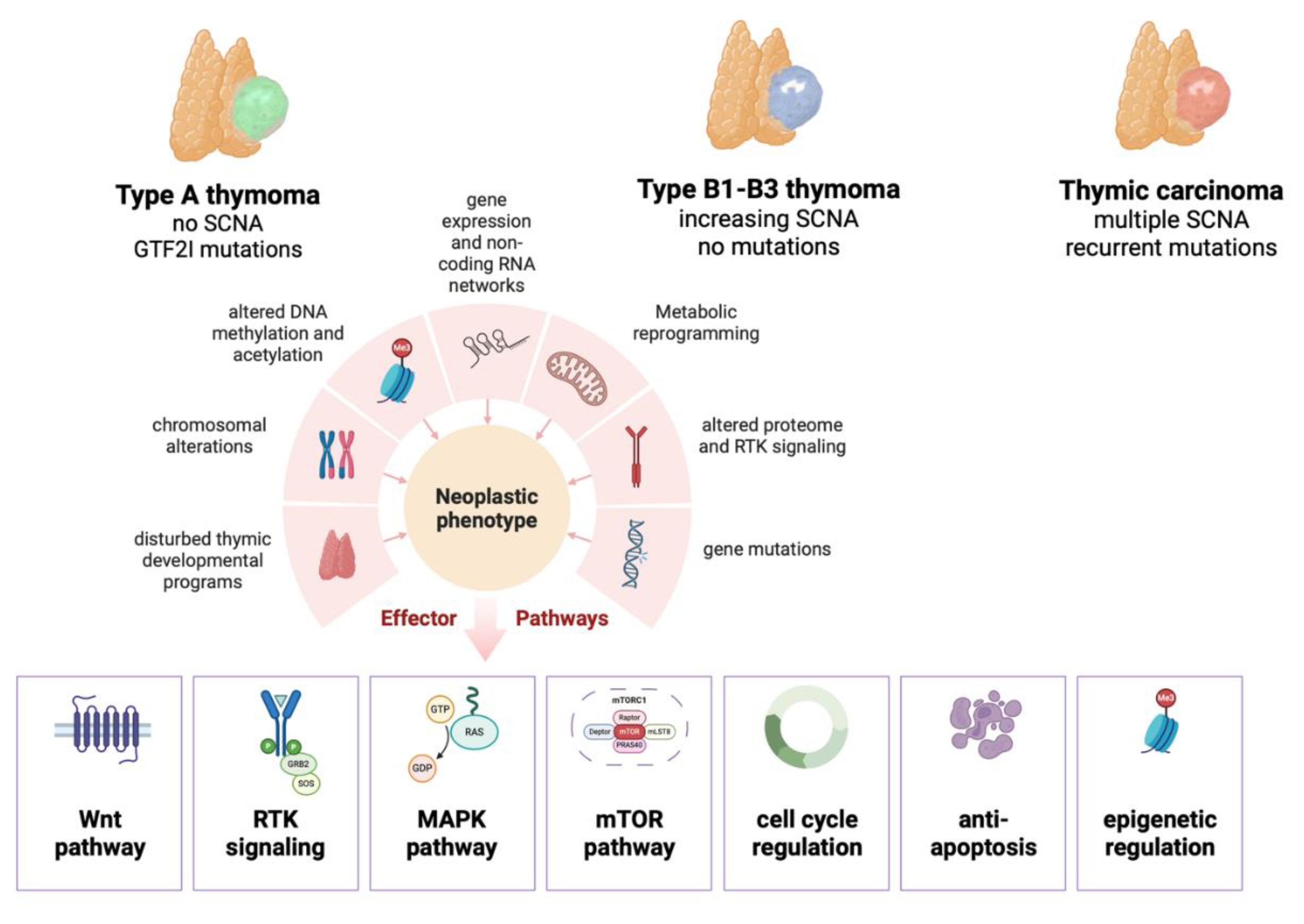
Disturbed thymic developmental programs in thymomas
Most TH contain subtype-specific mixtures of functional, albeit defective, cells with cortical and medullary differentiation, suggesting their origin from bipotent thymic progenitors [7]. The absence of the medullary autoimmune regulator gene (AIRE) in 95% of TH and recent studies in mice with mutations in GTF2I have provided experimental support for this hypothesis [8,9]. Therefore, it is to be expected that many of the programs and factors that govern the development and maintenance of the normal thymus are also important in the pathogenesis of TH. Cortical and medullary epithelial cells (cTECs and mTECs) of the normal thymus develop from a single bi-potent endodermal progenitor cell [10,11]. Recent single-cell analyses have revealed the vast complexity of TEC with at least nine distinct TEC subsets, including two cTEC and three mTEC clusters, one immature or uncommitted TEC cluster with canonical TEC identity genes (FOXN1, PAX9, SIX1), as well as three more clusters with neuroendocrine (BEX1, NEUROD1), muscle-like myoid (MYOD1, DES), and myelin-epithelial (SOX10, MPZ) cell features [12]. Major signaling pathways that play a critical role in TEC development and function include WNT, BMP, transforming growth factor beta (TGF beta), insulin-like growth factor (IGF), and fibroblast growth factor (FGF) signaling [13,14,15,16].
WNT signaling is the best studied in TH of the factors listed above. WNT signaling plays a key role in the development and maintenance of the thymus, and decreased expression of WNT proteins or increased levels of WNT inhibitors are associated with TEC senescence and thymic involution in mice and humans [17,18,19]. WNT4 is secreted from normal thymic epithelial cells and activates a signaling network via G-protein-dependent Frizzled receptors in an autocrine manner [20]. This autocrine loop is also active in TH, especially in the more aggressive subtypes [21]. Importantly, WNT signaling in TH appears to be non-canonical (i.e., β-catenin-independent) through the so-called PCP/JNK pathway [22] and may be supported by AKT and NF-kB signaling [21]. Wnt signaling also activates FOXN1 [14], a critical transcription factor that is deficient in mice and humans with the nude phenotype [23] and essential for the differentiation and maturation of all TEC [24]. FOXN1 promotes the proliferation of thymic epithelial progenitor cells and mTECs in the normal thymus [25] and in thymoma cells [26,27]. FOXN1 protein is expressed in most thymomas but partially lost in TC [28].
GTF2I is another transcription factor with a role in thymic development that is preferentially mutated in type A and AB thymomas [6]. However, a recent study using microdissected tissues reported high mutation rates of 42% also in type B thymomas [29]. In endogenous mouse models with specific expression of mutated GTF2I in FOXN1+ thymic epithelial cells, the mutation causes an incomplete block of TEC differentiation with accumulation of immature TECs and reduced thymopoietic activity [8] and leads to the formation of thymomas resembling WHO type B1 and B2 TH in aged animals [30]. The mutation consistently confers increased resistance to apoptosis, tolerance to DNA damage, and alterations in cell cycle regulation with enrichment of MYC, E2F, and G2M checkpoint target genes [8,30,31]. In addition, the mutation is associated with a growth advantage of the transformed cells through activation of glycolysis and altered lipid biosynthesis [31]. TH with GTF2I mutations showed increased expression of genes related to cell morphogenesis, receptor tyrosine kinase signaling, retinoic acid receptors, neuronal processes, and WNT and SHH signaling. Downregulated pathways included apoptosis, cell cycle, DNA damage response, hormone receptor signaling, RAS/MAPK, and TSC/mTOR [4]. GTF2I is a critical factor in the pathogenesis of the neurodevelopmental 7q11.23 microduplication autism spectrum disorder [32], where it is responsible for transcriptional dysregulation in early pluripotent neuronal stem cells by repressing critical neuronal genes through recruitment of lysine demethylase 1 (LSD1) [33]. Importantly, it has been shown in patient-derived neuronal cells that histone deacetylase inhibitors (HDACi) can reduce abnormal expression levels of GTF2I [32]. This therapeutic option that could be further pursued also in thymoma patients with GTF2I mutations.
Functional implications of chromosomal alterations in thymomas
Chromosomal aberrations are important because they were among the first molecular findings in TH [34] and are highly characteristic and reproducible, although their functional implications are still poorly understood. Their frequency increases from type A to B3 TH and correlates with the increasing aggressiveness of these subtypes [4,35]. Large-scale, whole- and arm-level sCNAs occur predominantly in tumors without gene mutations [4].
Type A showed very few arm-level copy number alterations [36]. The most common arm-level chromosomal alterations in thymomas (and in thymic squamous cell carcinomas) included losses on chromosome 6p25.1-p24.3, 6q25.3, 9p12.3, 13q and 16q, 22p12.1-p12.2, and gains on chromosome 1q31.1, 1q43, 3p12.2, 8q23.1, 9p24.3, 10q21.1, 12p12.3, 14q12, and Xq23 andXq25 [4,36]. Type B3 TH and TC showed overlapping chromosomal alterations with frequent arm-level CN gains of 1q and losses of chromosomes 6 and 13q. This finding starkly contrasts an integrative unsupervised clustering of five data platforms, which suggested that TCs are profoundly different from thymomas [4]. Petrini et al. were the first to correlate these copy number alterations with known cancer-related genes [36]. They identified BCL2 as a likely target of copy number gain at chr. 18q and CDKN2A as a likely target of loss at chr. 9p21. CDKN2A is a tumor suppressor gene that encodes for the two tumor suppressor proteins, p14 and p16 [37]. CDKN2A mutations are found in approximately 6% of TH, compared to 40% in TC [38,39]. p16 exerts its functions by inducing senescence in early transformed cells or by inducing apoptosis through TP53, the inhibition of the cyclin-dependent kinases CDK4 and CDK6, and the restoration of the retinoblastoma protein during the G1/S cell cycle phase [40,41]. Importantly, loss of p16 expression is not exclusively dependent on CN loss of CDKN2A but can also occur as a consequence of p16 promoter methylation [36,42,43] or miR-24 deregulation [44] and its loss correlates with tumor aggressiveness [36,45]. Together, these data arguably make CDKN2A/p16 one of the most relevant altered genes in thymic epithelial tumors. Another highly important finding of the study by Petrini et al. was the identification of profound deregulation of BCL2 family genes in TH, namely CN gains of BCL2 in tumors with chromosomal gains at chr. 18q or focal gene amplifications of BCL2 [36]. Frequent overexpression of BCL2 protein in TET had already been previously described [46,47,48,49]. Moreover, the same study also revealed CN gains of MCL1 [50] and BCL-XL in a substantial number of cases [36]. The fact that anti-apoptosis plays a central role in TET biology is further supported by previous observations describing overexpression of the anti-apoptotic protein BIRC3 in TC and downregulation of the pro-apoptotic protein NOXA in type B3 TH [51], and overexpression of the anti-apoptotic cellular FLICE-like inhibitory protein cFLIP [52]. Furthermore, functional studies using BH3 profiling in cell lines and tissue samples showed that B2 and B3 TH are exquisitely dependent on pro-survival factors such as MCL-1 and BCL-xL [36,53]. These observations suggest that TETs are promising candidates for clinical trials, e.g. with inhibitors of MCL-1 and BCL-xL.
The TCGA dataset [4] has extended the correlations between chromosomal alterations and potential genes of interest in commonly deleted or amplified chromosomal regions that await further experimental validation: PTPRG, FOXP1, and RYBP (3p13), CDYL (6p25.1-24.3), ARID1B, ZDHHC14 (6q25.3), NF2, CHEK2 (22p12.1-p12.2), PTGS2 (1q31.1), ACTN2 (1q43), PTPRD (9p24.3), PIK3C2G (12p12.3), and FOXG1 (14q12).
A recent publication suggests that the frequency of recurrent gene fusions in TH may have been underestimated. Based on RNA seq data, Ji et al. identified gene fusions in 36% (9/25 samples), including 1 sample with multiple gene fusions [54]. Of these, five fusions were unique to TH and included the previously reported KMT2A-MAML2 gene fusion [55] in 36% of cases, as well as HADHB-REEP1, COQ3-CGA, MCM4-SNTB1, and IFT140-ACTN4. The KMT2A gene encodes histone-lysine N-methyltransferase 2A, also known as acute lymphoblastic leukemia 1 (ALL-1), myeloid/lymphoid, or mixed-lineage leukemia 1 (MLL1). MLL1 is a positive global regulator of gene transcription with an essential role in regulating Hox genes [56]. MAML2 is a member of a family of NOTCH signaling coactivators. The KMT2A-MAML2 fusion gene has been shown to be oncogenic [57]. It disrupts NOTCH signaling and suppresses HES1 promoter activation [58]. A single published case of a thymoma with a neurotrophic tyrosine receptor kinase (NTRK) gene fusion showed a significant response to entrectinib [59].
Altered DNA methylation and acetylation in thymomas
DNA methyltransferase (DNMT) enzymes facilitate DNA methylation and significantly affect gene expression. DNMTs establish and maintain methylation patterns by adding methyl groups to cytosine residues within CpG dinucleotides. Alterations in CpG methylation contribute to cancer by causing hypermethylation of gene promoters, leading to transcriptional silencing of tumor suppressor genes and hypomethylation of intergenic and intronic sequences, inducing chromosomal instability in proto-oncogene regions. In mammalian genomes, DNMT3a and DNMT3b are responsible for de novo methylation, while DNMT1 is responsible for maintaining methylation patterns. Increased DNMT activity in cancer cells is associated with tumor aggressiveness and poor patient prognosis [60]. Numerous published studies, summarized in recent reviews [61,62], have addressed the epigenetics of TET. Most of the available literature has focused on the potential use of DNA methylation markers to discriminate between different histologic subtypes or between TH with and without myasthenia gravis or as a prognostic indicator. Studies analyzing global methylation patterns in TET have found substantial hyper- or hypomethylation of CpG sites with significant correlations with TH histologic subtype [4,63,64,65], GTF2I or HRAS mutation status, and mRNA and miRNA expression patterns [4]. Previous studies using a candidate gene approach revealed hypomethylation of several tumor suppressor genes (e.g., APC1A, CDKN2A, E-cad, FHIT, hMLH1, MGMT, RARβ, RASSF1A) or hypermethylation of DNMT1, DNMT3a, and DNMT3b [66] in early stage thymomas (TMs) compared to more aggressive or late stage TMs and thymic carcinomas (TCs) (reviewed in [61,62]). The specific silencing of these genes with a role in regulating cell cycle progression and DNA repair suggests an essential step during TH progression.
Acetylation is a covalent histone modification that occurs at evolutionarily conserved lysine residues of nucleosome complexes. Acetyl groups added to histones loosen their chemical interactions with DNA, promoting the decompaction of tightly packed chromatin. This process enhances accessibility to transcriptional machinery components, establishing permissive chromatin states for gene expression [67]. Conversely, removal of acetyl groups from histones by histone deacetylases (HDACs) generally results in transcriptional repression and silencing of tumor suppressor genes. Overexpression of class I (HDAC1-3 and 8) and class II (HDAC4-7 and 9-10) HDACs is associated with poor prognosis in TH [68]. The HDCA inhibitor belinostat has shown activity in TH in vitro [69] and clinical trials [70,71].
Altered gene expression and non-coding RNA networks in thymomas
Many studies have analyzed mRNA expression profiles in TET [4,13,72,73,74,75,76,77,78,79,80,81]. However, the results are difficult to compare due to differences in study focus, methodology, number of samples, controls (e.g., TH vs. normal thymus, TH vs. TC, TH with and without myasthenia gravis). Recent studies have shown a high concordance with the main TH subgroups [4,13]. TH with GTF2I mutations (type A and AB thymomas) were in enriched for genes related to human embryonic stem cell pluripotency and Wnt/β-catenin signaling [82], lymphocyte-rich type B1 and B2 thymomas were enriched for genes related to T-cell receptor signaling, CTLA4 signaling, and ICOS/ICOSL signaling, whereas type B3 thymomas overexpressed genes related to G protein-coupled receptor signaling, Toll-like receptor signaling regulation of epithelial-mesenchymal transition (EMT), as well as NF-κB, EGF, FAK, and telomerase signaling. Differentially regulated pathways that were identified in more than one study included cell cycle regulation [4,72,79,81], MAPK/ERK signaling [4,78], hormone signaling [4,72], Wnt signaling [78,82], and apoptosis [4,81]. In a study by Badve et al., the top pathways associated with metastasis were amino acid metabolism, steroid and glycosphingolipid biosynthesis, cell cycle checkpoint proteins, and Notch signaling [72].
Non-coding RNAs (ncRNAs) play critical roles in various biological processes, diseases and tumors, and many published studies show that their expression patterns can be used e.g. for accurate classification and prognostic prediction of TH. The role of ncRNAs in TET has been addressed in several recent excellent reviews [61,62,83] and will only be briefly highlighted here. ncRNAs are produced by transcription from various genomic regions and undergo post-transcriptional maturation and modification [84]. They can be classified into small non-coding RNAs (miRNAs and siRNAs) and long non-coding RNAs (lncRNAs), which can be further subdivided into circular and linear forms [85]. LncRNAs have emerged as important modulators of gene expression, affecting networks in different human cancers, including TET [86]. Acting at different levels of control in gene regulation, lncRNAs affect pathways related to cell fate determination in normal and pathological conditions. Circular lncRNAs (circRNAs), characterized by covalently closed structures, exhibit stability and diverse functions, such as serving as microRNA sponges, modulating RNA-binding proteins, or acting as protein scaffolds [87]. Studies have shown differential expression of circRNAs in TH [88]. Studies of lncRNA profiles in TH tissues suggest that specific lncRNA expression alterations may be associated with overall or disease-free survival in TH. Su et al. identified four lncRNAs (ADAMTS9-AS1, HSD52, LINC00968, and LINC01697) that were significantly associated with recurrence-free survival (RFS) in TET [89]. These lncRNAs stratified patients into high or low-risk groups, outperforming traditional prognostic models. In a separate study of 25 TH patients, ADAMTS9-AS1, which was associated with RFS, showed altered expression with five other lncRNAs affecting various cancer types and biological processes [54]. In particular, ADAMTS9-AS1 and LINC00324 expression correlated with patient prognosis. These lncRNAs were associated with miRNA cluster dysregulation and target gene alterations in tumorigenic signaling pathways, such as PI3K/Akt, FoxO, HIF-1, and Notch, supporting their roles in TH progression. Using data mining approaches, two groups found that upregulation of AFAP1-AS1 and downregulation of LINC00324 were associated with RFS in TET patients [54,90]. Ji et al. performed a functional annotation of the top 10 lncRNAs with altered expression in TH and identified many predicted target genes [54] that merit further functional investigation. In addition, the lncRNA RP11-424C20.2 regulates UHRF1 expression by sponging miR-378a-3p, which influences thymoma prognosis through interactions with infiltrating immune cells and modulation of the tumor immune microenvironment [91,92]. Among small ncRNAs, microRNAs (miRNAs) are prominent in post-transcriptional regulation by binding to specific mRNA targets and inducing degradation or translational inhibition [93]. miRNAs can also repress key enzymes that drive epigenetic remodeling [94]. Radovich et al. identified selective overexpression of a large microRNA cluster on chr19q13.42 (C19MC) exclusively in type A and AB TH, which was also confirmed by TCGA data [4]. Overexpression of this microRNA cluster activates the PI3K/AKT/mTOR pathway, and PI3K/AKT/mTOR inhibitors reduced cell viability in a type AB TH cell line [77]. Another miRNA cluster on chr14q32 (C14MC) is expressed in TH and downregulated in TC [95]. Integrating a large amount of TH data into a lncRNA-mRNA-miRNA regulatory network, Ji et al. predicted that overexpression of miRNA clusters activates the PI3K-Akt/FoxO/HIF-1/Rap-1 signaling pathway [54]. Data from the TCGA study [96] show that high expression of LOXL1-AS1 and low expression of miR-525-5p are associated with poor prognosis in TET. LOXL1-AS1 acts as a sponge for miR-525-5p, promoting HSPA9 expression and thereby enhancing the growth and invasion of TET cells while inhibiting apoptosis [96]. MiR-525-5p acts as a tumor suppressor by repressing HSPA9, which is upregulated in TETs and correlates with poor patient survival. Another network involves LINC00174, miR-145-5p, and target genes in thymic tumorigenesis [97]. Upregulation of LINC00174 is negatively associated with miR-145-5p [98] and suggests an oncogenic role in TETs, affecting cell growth, migration, and lipid metabolism. The lncRNA MALAT1, acting as a miR-145-5p sponge, contributes to thymic cancer development, with its downregulation leading to decreased proliferation and increased apoptosis. The combination of MALAT1 silencing and miR-145-5p overexpression has a synergistic effect [99]. Iaiza et al. found that methylation and delocalization of MALAT1 through the methyltransferase METTL3 induces c-MYC expression in aggressive TH and TC. Silencing of METTL3 combined with cisplatin or c-MYC inhibitors promotes apoptosis in TC cells [100].
Metabolic reprogramming in thymomas
The metabolome of TH and TC is an evolving field and remains understudied. Circumstantial evidence that TH, especially the aggressive types, show profound changes in their metabolism comes from nuclear medicine and positron emission tomography (PET) studies using fluorodeoxyglucose. It has been shown that high preoperative glucose uptake [101,102] and high serum levels of lactate dehydrogenase (LDH) [103] can be used to predict histologic subtypes and prognosis in thymoma patients. These observations were consistent with a metabolic study in TH showing high lactic acid and glutamine levels and activation of the proline/arginine, glycolysis, and glutathione pathways, suggesting increased glycolysis and glutaminolysis [104]. Two other studies concluded that TH contain two main metabolic subgroups [105,106]. Zhang et al. identified alterations in neutral lipid biosynthesis and phospholipid metabolism, particularly the lacto and neolacto series in glycosphingolipid biosynthesis, as a central metabolic pathway associated with TH progression [105]. They identified two key enzymes of this pathway, B3GNT5, and ST3GAL6, which showed prognostic correlation with the overall survival of TH patients. Glycosphingolipids are a family of essential cell membrane molecules involved in cell‒cell recognition and signal transduction [107]. Changes in their glycosylation are associated with stem cell differentiation and various cancer-related processes, such as cell proliferation and metastasis [108]. Overexpression of B3GNT5, an essential enzyme in producing lactate and lactate-series glycosphingolipids, was associated with poor outcomes in TH patients. The other enzyme, ST3GAL6, is involved synthesizing glycolipid substrates and is altered in various cancers. Consistent with findings in lung cancer [109], where downregulation of ST3GAL6 activates EGFR/MAPK signaling and stimulates the expression of matrix metalloproteinase 2 and 9, low levels predicted poor prognosis in TH patients. Tang et al. [106] identified two distinct metabolic patterns in TH, with different metabolic scores correlating with clinical outcomes. High metabolic scores were associated with worse survival and immunosuppressive status. The study also described differential activation of the glycosphingolipid metabolism and arginine biosynthesis. Interestingly, a recent proteomic study has described overexpression of the rate-limiting enzyme for arginine biosynthesis, argininosuccinate synthase 1 (ASS1), in type B2 and B3 compared to type A, AB, and B1 TH [110]. The upregulation of ASS1 in malignant TH contrasts with many other tumors (e.g., hepatocellular carcinoma or mesothelioma), which downregulate ASS1 and consequently become dependent on extracellular arginine [111]. In addition, asparagine synthetase (ASNS), an enzyme involved in amino acid metabolism that requires glutamine to convert aspartic acid to asparagine, was upregulated in TH and correlated with reduced patient survival [106].
Proteomics and altered tyrosine kinase signaling
The proteome of TH is still a relatively unexplored area of research. A number of publications have analyzed the thymoma proteome in an unbiased manner using either tissue lysates [4,110,112,113,114,115] or serum [116,117,118,119].
Most of these studies have focused on differential protein profiles between TH subtypes or between normal thymus and TH and have typically found significant differences based on hundreds of differentially expressed proteins, some of which have also been proposed in a diagnostic context for refined TH classification [110]. Reverse phase protein array (RPPA) data from the TCGA dataset were used to identify significant pathways with differential expression between TH subtypes and identified cell cycle, apoptosis, EMT, RAS/MAPK, hormone signaling, breast reactive, and core reactive pathway [4]. Altered receptor tyrosine kinase (RTK) signaling in TH has been the focus of considerable interest over the past decades but often based on small series using immunohistochemical techniques or RT-PCR. Many of these studies have identified significant differences between TH subtypes, and overexpression of RTKs was often associated with more aggressive behavior or advanced tumor stage. The best-studied examples are the EGFR (positive in 43 - 100% of TH) [120,121,122,123,124], IGF1R (positive in 43 - 100% of TH) [125,126,127,128], VEGFR 1-3 [129,130,131] and their ligands VEGFA, VEGFC and VEGFD [131]. TH are consistently negative for KIT [132,133,134], ALK [135], HER2 [136,137], and MET [137]. However, overexpression of a protein does not necessarily prove its functional relevance, e.g., as a predictive biomarker for therapeutic response. Using phospho-RTK arrays, Küffer et al. studied the activation of 44 RTKs in 37 type B2 and B3 thymomas [138]. The EGFR was by far the most prevalent active RTK (74% of cases). However, there was a striking dichotomy between primary and metastatic tumors: all metastatic TH were negative for EGFR, and 57% of these tumors showed TYRO3/Dtk activation instead. The most frequently activated other RTKs in TH were (in descending order) FGFR2 (30%), VEGFR3 (24%), TRKC (24%), Tie2, TRKB and EphA6 (21%), VEGFR1 (19%), EphB6, FGFR4, FLT3, INSR, VEGFR2 (16% each) and ERBB3. Interestingly, this study also showed activation of RTKs that are not normally overexpressed in TH, namely ERBB2 (11%) and KIT (8%). Similarly, a recent study [139] described KITLG overexpression preferentially in type A and AB thymomas, which are consistently KIT negative by immunohistochemistry. KITLG overexpression was associated with highly specific changes in mRNA and miRNA expression profiles, upregulation of DNMT3B and downregulation of DNMT1, DNMT3A, and DNMT3L, as well as altered methylation patterns. The major upregulated pathway in TH upon KITLG overexpression was MAPK signaling.
Conclusions and future directions
Despite their heterogeneity and the lack of driver mutations in most TH, key principles and molecular features are emerging that may help to improve our understanding of TH biology and open up opportunities for targeted treatments. The TCGA data have shown that the heterogeneity of TH at the molecular level can be reduced to two main categories, namely type A and AB thymomas and type B thymomas. While it may still be relevant to investigate the exact mechanisms that differ, for example, between type B1 and B2 thymomas, the main research effort should probably focus on elucidating the molecular pathogenesis of the two main subgroups. Type A/AB thymomas carry no or few chromosomal alterations but a recurrent GTF2I mutation as the basic driver. Type B thymomas harbor a variable number of chromosomal alterations but no known driver mutations. In both settings, additional non-mutational mechanisms shape the behavior of the cell by altering critical "hallmark" pathways such as cell cycle regulation or resistance to apoptosis.
References
- Buckley, C.; Douek, D.; Newsom-Davis, J.; Vincent, A.; Willcox, N. Mature, Long-Lived Cd4+ and Cd8+ T Cells Are Generated by the Thymoma in Myasthenia Gravis. Ann. Neurol. 2001, 50, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Marx, A., F. Detterbeck, E.M. Marom, P. Ströbel, and A. Rajan. "Tumors of the Thymus: Introduction." In Who Classification of Tumours. Thoracic Tumours., edited by A. Marx and J.K.C. Chan. Lyon: International Agency for Research on Cancer, 2021.
- Marx, A.; Porubsky, S.; Belharazem, D.; Saruhan-Direskeneli, G.; Schalke, B.; Ströbel, P.; Weis, C.-A. Thymoma related myasthenia gravis in humans and potential animal models. Exp. Neurol. 2015, 270, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Radovich, M.; Pickering, C.R.; Felau, I.; Ha, G.; Zhang, H.; Jo, H.; Hoadley, K.A.; Anur, P.; Zhang, J.; McLellan, M.; et al. The Integrated Genomic Landscape of Thymic Epithelial Tumors. Cancer Cell 2018, 33, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Venuta, F.; A Rendina, E.; O Pescarmona, E.; De Giacomo, T.; Vegna, M.L.; Fazi, P.; Flaishman, I.; Guarino, E.; Ricci, C. Multimodality Treatment of Thymoma: A Prospective Study. Ann. Thorac. Surg. 1997, 64, 1585–1592. [Google Scholar] [CrossRef]
- Petrini, I.; Meltzer, P.S.; Kim, I.-K.; Lucchi, M.; Park, K.-S.; Fontanini, G.; Gao, J.; A Zucali, P.; Calabrese, F.; Favaretto, A.; et al. A specific missense mutation in GTF2I occurs at high frequency in thymic epithelial tumors. Nat. Genet. 2014, 46, 844–849. [Google Scholar] [CrossRef]
- Ströbel, P.; Hartmann, E.; Rosenwald, A.; Kalla, J.; Ott, G.; Friedel, G.; Schalke, B.; Kasahara, M.; Tomaru, U.; Marx, A. Corticomedullary differentiation and maturational arrest in thymomas. Histopathology 2013, 64, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Giorgetti, O.B.; Nusser, A.; Boehm, T. Human thymoma-associated mutation of the GTF2I transcription factor impairs thymic epithelial progenitor differentiation in mice. Commun. Biol. 2022, 5, 1–12. [Google Scholar] [CrossRef]
- Ströbel, P.; Murumägi, A.; Klein, R.; Luster, M.; Lahti, M.; Krohn, K.; Schalke, B.; Nix, W.; Gold, R.; Rieckmann, P.; et al. Deficiency of the autoimmune regulator AIRE in thymomas is insufficient to elicit autoimmune polyendocrinopathy syndrome type 1 (APS-1). J. Pathol. 2007, 211, 563–571. [Google Scholar] [CrossRef]
- Bleul, C.C.; Corbeaux, T.; Reuter, A.; Fisch, P.; Mönting, J.S.; Boehm, T. Formation of a functional thymus initiated by a postnatal epithelial progenitor cell. Nature 2006, 441, 992–996. [Google Scholar] [CrossRef]
- Rossi, S.W.; Jenkinson, W.E.; Anderson, G.; Jenkinson, E.J. Clonal analysis reveals a common progenitor for thymic cortical and medullary epithelium. Nature 2006, 441, 988–991. [Google Scholar] [CrossRef]
- Bautista, J.L.; Cramer, N.T.; Miller, C.N.; Chavez, J.; Berrios, D.I.; Byrnes, L.E.; Germino, J.; Ntranos, V.; Sneddon, J.B.; Burt, T.D.; et al. Single-cell transcriptional profiling of human thymic stroma uncovers novel cellular heterogeneity in the thymic medulla. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lepletier, A.; Hun, M.L.; Hammett, M.V.; Wong, K.; Naeem, H.; Hedger, M.; Loveland, K.; Chidgey, A.P. Interplay between Follistatin, Activin A, and BMP4 Signaling Regulates Postnatal Thymic Epithelial Progenitor Cell Differentiation during Aging. Cell Rep. 2019, 27, 3887–3901. [Google Scholar] [CrossRef] [PubMed]
- Balciunaite, G.; Keller, M.P.; Balciunaite, E.; Piali, L.; Zuklys, S.; Mathieu, Y.D.; Gill, J.; Boyd, R.; Sussman, D.J.; Hollander, G.A. Wnt Glycoproteins Regulate the Expression of Foxn1, the Gene Defective in Nude Mice. Nat. Immunol. 2002, 3, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.; Patel, S.R.; Mishina, Y.; Manley, N.R. Evidence for an early role for BMP4 signaling in thymus and parathyroid morphogenesis. Dev. Biol. 2010, 339, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Bleul, C.C.; Boehm, T. BMP Signaling Is Required for Normal Thymus Development. J. Immunol. 2005, 175, 5213–5221. [Google Scholar] [CrossRef]
- Wei, T.; Zhang, N.; Guo, Z.; Chi, F.; Song, Y.; Zhu, X. Wnt4 signaling is associated with the decrease of proliferation and increase of apoptosis during age-related thymic involution. Mol. Med. Rep. 2015, 12, 7568–7576. [Google Scholar] [CrossRef] [PubMed]
- Heinonen, K.M.; Vanegas, J.R.; Brochu, S.; Shan, J.; Vainio, S.J.; Perreault, C. Wnt4 regulates thymic cellularity through the expansion of thymic epithelial cells and early thymic progenitors. Blood 2011, 118, 5163–5173. [Google Scholar] [CrossRef] [PubMed]
- Varecza, Z.; Kvell, K.; Talabér, G.; Miskei, G.; Csongei, V.; Bartis, D.; Anderson, G.; Jenkinson, E.J.; Pongracz, J.E. Multiple suppression pathways of canonical Wnt signalling control thymic epithelial senescence. Mech. Ageing Dev. 2011, 132, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Kvell, K.; Varecza, Z.; Bartis, D.; Hesse, S.; Parnell, S.; Anderson, G.; Jenkinson, E.J.; Pongracz, J.E. Wnt4 and LAP2alpha as Pacemakers of Thymic Epithelial Senescence. PLoS ONE 2010, 5, e10701. [Google Scholar] [CrossRef]
- Zhang, X.; Schalke, B.; Kvell, K.; Kriegsmann, K.; Kriegsmann, M.; Graeter, T.; Preissler, G.; Ott, G.; Kurz, K.; Bulut, E.; et al. WNT4 overexpression and secretion in thymic epithelial tumors drive an autocrine loop in tumor cells in vitro. Front. Oncol. 2022, 12, 920871. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, P.; Tang, P.; Lv, P.; Li, X.; Wang, Y.; Lv, Y.; Liu, Y. Wnt4 overexpression promotes thymoma development through a JNK-mediated planar cell polarity-like pathway. Oncol. Lett. 2017, 15, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Nehls, M.; Kyewski, B.; Messerle, M.; Waldschütz, R.; Schüddekopf, K.; Smith, A.J.H.; Boehm, T. Two Genetically Separable Steps in the Differentiation of Thymic Epithelium. Science 1996, 272, 886–889. [Google Scholar] [CrossRef]
- Corbeaux, T.; Hess, I.; Swann, J.B.; Kanzler, B.; Haas-Assenbaum, A.; Boehm, T. Thymopoiesis in mice depends on a Foxn1 -positive thymic epithelial cell lineage. Proc. Natl. Acad. Sci. USA 2010, 107, 16613–16618. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wachsmuth, L.P.; Xiao, S.; Condie, B.G.; Manley, N.R. Foxn1 Overexpression Promotes Thymic Epithelial Progenitor Cell Proliferation and Mtec Maintenance, but Does Not Prevent Thymic Involution. Development 2023, 150, 8. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, P.; Liu, Y.; Lv, P.; Wang, Y.; Chen, Y. In vitro study of the effect of small interfering ribonucleic acid on the expression of FOXN1 and B cell-attracting chemokine 1 in thymoma cell lines. Thorac. Cancer 2015, 6, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, X.; Liu, Y.; Wang, Y.; Wang, H.; Lu, C.; Zhang, P. Decreased Wnt4 expression inhibits thymoma development through downregulation of FoxN1. J. Thorac. Dis. 2017, 9, 1574–1583. [Google Scholar] [CrossRef]
- Nonaka, D.; Henley, J.D.; Chiriboga, L.; Yee, H. Diagnostic Utility of Thymic Epithelial Markers CD205 (DEC205) and Foxn1 in Thymic Epithelial Neoplasms. Am. J. Surg. Pathol. 2007, 31, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Goto, T.; Hirotsu, Y.; Yokoyama, Y.; Nakagomi, T.; Otake, S.; Amemiya, K.; Oyama, T.; Mochizuki, H.; Omata, M. Primary Driver Mutations in GTF2I Specific to the Development of Thymomas. Cancers 2020, 12, 2032. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Kim, I.-K.; Bian, J.; Polyzos, A.; Di Giammartino, D.C.; Zhang, Y.-W.; Luo, J.; Hernandez, M.O.; Kedei, N.; Cam, M.; et al. A Knock-In Mouse Model of Thymoma With the GTF2I L424H Mutation. J. Thorac. Oncol. 2022, 17, 1375–1386. [Google Scholar] [CrossRef]
- Kim, I.-K.; Rao, G.; Zhao, X.; Fan, R.; Avantaggiati, M.L.; Wang, Y.; Zhang, Y.-W.; Giaccone, G. Mutant GTF2I induces cell transformation and metabolic alterations in thymic epithelial cells. Cell Death Differ. 2020, 27, 2263–2279. [Google Scholar] [CrossRef]
- Cavallo, F.; Troglio, F.; Faga, G.; Fancelli, D.; Shyti, R.; Trattaro, S.; Zanella, M.; G. D'Agostino; Hughes, J.M.; Cera, M.R.; Pasi, M.; Gabriele, M.; Lazzarin, M.; Mihailovich, M.; Kooy, F.; Rosa, A.; Mercurio, C.; Varasi, M.; Testa, G. "High-Throughput Screening Identifies Histone Deacetylase Inhibitors That Modulate Gtf2i Expression in 7q11.23 Microduplication Autism Spectrum Disorder Patient-Derived Cortical Neurons. Mol. Autism. 2020, 11, 88. [Google Scholar] [PubMed]
- Adamo, A.; Atashpaz, S.; Germain, P.-L.; Zanella, M.; D'Agostino, G.; Albertin, V.; Chenoweth, J.; Micale, L.; Fusco, C.; Unger, C.; et al. 7q11.23 dosage-dependent dysregulation in human pluripotent stem cells affects transcriptional programs in disease-relevant lineages. Nat. Genet. 2014, 47, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Zettl, A.; Ströbel, P.; Wagner, K.; Katzenberger, T.; Ott, G.; Rosenwald, A.; Peters, K.; Krein, A.; Semik, M.; Müller-Hermelink, H.-K.; et al. Recurrent Genetic Aberrations in Thymoma and Thymic Carcinoma. Am. J. Pathol. 2000, 157, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Petrini, I.; Meltzer, P.S.; A Zucali, P.; Luo, J.; Lee, C.; Santoro, A.; Lee, H.S.; Killian, K.J.; Wang, Y.; Tsokos, M. Copy number aberrations of BCL2 and CDKN2A/B identified by array-CGH in thymic epithelial tumors. Cell Death Dis. 2012, 3, e351. [Google Scholar] [CrossRef]
- Copy Number Aberrations of Bcl2 and Cdkn2a/B Identified by Array-Cgh in Thymic Epithelial Tumors. Cell Death Dis. 2012, 3, e351. [CrossRef] [PubMed]
- Sherr, C.J. The INK4a/ARF network in tumour suppression. Nat. Rev. Mol. Cell Biol. 2001, 2, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Girard, N.; Basse, C.; Schrock, A.; Ramkissoon, S.; Killian, K.; Ross, J.S. Comprehensive Genomic Profiling of 274 Thymic Epithelial Tumors Unveils Oncogenic Pathways and Predictive Biomarkers. Oncol. 2022, 27, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Kurokawa, K.; Shukuya, T.; Greenstein, R.; Kaplan, B.; Wakelee, H.; Ross, J.; Miura, K.; Furuta, K.; Kato, S.; Suh, J.; et al. Genomic characterization of thymic epithelial tumors in a real-world dataset. ESMO Open 2023, 8, 101627. [Google Scholar] [CrossRef] [PubMed]
- Rubin, S.M. Deciphering the retinoblastoma protein phosphorylation code. Trends Biochem. Sci. 2013, 38, 12–19. [Google Scholar] [CrossRef]
- Jiang, Y.-Z.; Ma, D.; Suo, C.; Shi, J.; Xue, M.; Hu, X.; Xiao, Y.; Yu, K.-D.; Liu, Y.-R.; Yu, Y.; et al. Genomic and Transcriptomic Landscape of Triple-Negative Breast Cancers: Subtypes and Treatment Strategies. Cancer Cell 2019, 35, 428–440. [Google Scholar] [CrossRef]
- Hirose, Y.; Kondo, K.; Takizawa, H.; Nagao, T.; Nakagawa, Y.; Fujino, H.; Toba, H.; Kenzaki, K.; Sakiyama, S.; Tangoku, A. Aberrant methylation of tumour-related genes in thymic epithelial tumours. Lung Cancer 2009, 64, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Hirabayashi, H.; Fujii, Y.; Sakaguchi, M.; Tanaka, H.; Yoon, H.E.; Komoto, Y.; Inoue, M.; Miyoshi, S.; Matsuda, H. P16ink4, Prb, P53 and Cyclin D1 Expression and Hypermethylation of Cdkn2 Gene in Thymoma and Thymic Carcinoma. Int. J. Cancer 1997, 73, 639–644. [Google Scholar] [CrossRef]
- Lal, A.; Kim, H.H.; Abdelmohsen, K.; Kuwano, Y.; Pullmann, R.; Srikantan, S.; Subrahmanyam, R.; Martindale, J.L.; Yang, X.; Ahmed, F.; et al. p16INK4a Translation Suppressed by miR-24. PLoS ONE 2008, 3, e1864. [Google Scholar] [CrossRef] [PubMed]
- Aesif, S.W.; Aubry, M.C.; Yi, E.S.; Kloft-Nelson, S.M.; Jenkins, S.M.; Spears, G.M.; Greipp, P.T.; Sukov, W.R.; Roden, A.C. Loss of p16 INK4A Expression and Homozygous CDKN2A Deletion Are Associated with Worse Outcome and Younger Age in Thymic Carcinomas. J. Thorac. Oncol. 2017, 12, 860–871. [Google Scholar] [CrossRef]
- Stefanaki, K.; Rontogianni, C. H. Kouvidou, S. Bolioti. Expression of P53, Mdm2, P21/Waf1 and Bcl-2 Proteins in Thymomas. Histopathology 1997, 30, 549–55. [Google Scholar] [CrossRef] [PubMed]
- Pan, C. C.; Chen, L. S. Wang. Expression of Apoptosis-Related Markers and Her-2/Neu in Thymic Epithelial Tumours. Histopathology 2003, 43, 165–72. [Google Scholar] [CrossRef]
- Khoury, T.; Arshad, A.; Bogner, P.; Ramnath, N.; Zhang, S.; Chandrasekhar, R.; Wilding, G.; Alrawi, S.; Tan, D. Apoptosis-Related (Survivin, Bcl-2), Tumor Suppressor Gene (p53), Proliferation (Ki-67), and Non-Receptor Tyrosine Kinase (Src) Markers Expression and Correlation With Clinicopathologic Variables in 60 Thymic Neoplasms. Chest 2009, 136, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Lozada-Nur, F. HIV: common early oral markers. J. Calif. Dent. Assoc. 1989, 17, 36–41. [Google Scholar]
- Chen, F.; Yan, J.; Chang, K.; Lai, W.; Chen, R.; Jin, Y. Immunohistochemical localization of Mcl-1 and bcl-2 proteins in thymic epithelial tumours. Histopathology 1996, 29, 541–547. [Google Scholar] [CrossRef]
- Huang, B.; Belharazem, D.; Li, L.; Kneitz, S.; Schnabel, P.A.; Rieker, R.J.; Körner, D.; Nix, W.; Schalke, B.; Müller-Hermelink, H.K.; et al. Anti-Apoptotic Signature in Thymic Squamous Cell Carcinomas – Functional Relevance of Anti-Apoptotic BIRC3 Expression in the Thymic Carcinoma Cell Line 1889c. Front. Oncol. 2013, 3. [Google Scholar] [CrossRef]
- Belharazem, D., A. Grass, C. Paul, M. Vitacolonna, B. Schalke, R. J. Rieker, D. Korner, P. Jungebluth, K. Simon-Keller, P. Hohenberger, E. M. Roessner, K. Wiebe, T. Grater, T. Kyriss, G. Ott, P. Geserick, M. Leverkus, P. Strobel, and A. Marx. Increased Cflip Expression in Thymic Epithelial Tumors Blocks Autophagy Via Nf-Kappab Signalling. Oncotarget 2017, 8, 89580–94. [Google Scholar] [PubMed]
- Muller, D.; Mazzeo, R.; Koch. Functional Apoptosis Profiling Identifies Mcl-1 and Bcl-Xl as Prognostic Markers and Therapeutic Targets in Advanced Thymomas and Thymic Carcinomas. BMC Med. 2021, 19, 300. [Google Scholar] [CrossRef] [PubMed]
- Ji, G.; Ren, R.; Fang, X. Identification and Characterization of Non-Coding RNAs in Thymoma. Med. Sci. Monit. 2021, 27, e929727–1. [Google Scholar] [CrossRef] [PubMed]
- Massoth, L. R., Y. P. Hung, D. Dias-Santagata, M. Onozato, N. Shah, E. Severson, D. Duncan, B. J. Gillespie, N. F. Williams, J. S. Ross, J. A. Vergilio, S. K. Harkins, K. Glomski, V. Nardi, L. R. Zukerberg, R. P. Hasserjian, A. Louissaint, Jr., and E. A. Williams. Pan-Cancer Landscape Analysis Reveals Recurrent Kmt2a-Maml2 Gene Fusion in Aggressive Histologic Subtypes of Thymoma. JCO Precis. Oncol. 2020, 4. [Google Scholar]
- Milne, T.A.; Briggs, S.D.; Brock, H.W.; Martin, M.E.; Gibbs, D.; Allis, C.D.; Hess, J.L. MLL Targets SET Domain Methyltransferase Activity to Hox Gene Promoters. Mol. Cell 2002, 10, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Wächter, K.; Kowarz, E.; Marschalek, R. Functional characterisation of different MLL fusion proteins by using inducible Sleeping Beauty vectors. Cancer Lett. 2014, 352, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, N.; Suzukawa, K.; Shimizu, S.; Shinagawa, A.; Takei, N.; Taki, T.; Hayashi, Y.; Kojima, H.; Kawakami, Y.; Nagasawa, T. Identification of a novel fusion gene MLL-MAML2 in secondary acute myelogenous leukemia and myelodysplastic syndrome with inv(11)(q21q23). Genes Chromosom. Cancer 2007, 46, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Salame, H.; Mckey, R.; Ballout, M.; Saad, W. The First Reported Case of Neurotrophic Tyrosine Receptor Kinase Fusion-Positive Thymoma Treated Successfully With Entrectinib. Cureus 2021, 13, e20588. [Google Scholar] [CrossRef]
- Chen, C.; Yin, N.; Yin, B.; Lu, Q. DNA methylation in thoracic neoplasms. Cancer Lett. 2011, 301, 7–16. [Google Scholar] [CrossRef]
- Nicolì, V.; Coppedè, F. Epigenetics of Thymic Epithelial Tumors. Cancers 2023, 15, 360. [Google Scholar] [CrossRef]
- Psilopatis, I.; Pergaris, A.; Vrettou, K.; Theocharis, S.; Troungos, C. Thymic Epithelial Neoplasms: Focusing on the Epigenetic Alterations. Int. J. Mol. Sci. 2022, 23, 4045. [Google Scholar] [CrossRef]
- Gaiser, T.; Hirsch, D.; Porth, I.; Sahm, F.; Ströbel, P.; von Deimling, A.; Marx, A. DNA-Methylation Analysis as a Tool for Thymoma Classification. Cancers 2022, 14, 5876. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Meng, Y.; Niu, Y.; Li, S.; Liu, H.; He, J.; Zhang, Y.; Liang, N.; Liu, L.; Mao, X.; et al. Genome-wide DNA methylation profile of thymomas and potential epigenetic regulation of thymoma subtypes. Oncol. Rep. 2019, 41, 2762–2774. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yuan, Y.; Xiao, H.; Dai, J.; Ye, Y.; Zhang, Q.; Zhang, Z.; Jiang, Y.; Luo, J.; Hu, J.; et al. Discovery and validation of DNA methylation markers for overall survival prognosis in patients with thymic epithelial tumors. Clin. Epigenetics 2019, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yin, B.; Wei, Q.; Li, D.; Hu, J.; Yu, F.; Lu, Q. Aberrant DNA Methylation in Thymic Epithelial Tumors. Cancer Investig. 2009, 27, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Shvedunova, M.; Akhtar, A. Modulation of cellular processes by histone and non-histone protein acetylation. Nat. Rev. Mol. Cell Biol. 2022, 23, 329–349. [Google Scholar] [CrossRef] [PubMed]
- Palamaris, K.; Tzimou, L.-M.; Levidou, G.; Masaoutis, C.; Theochari, I.; Rontogianni, D.; Theocharis, S. Histone Deacetylases (HDACs): Promising Biomarkers and Potential Therapeutic Targets in Thymic Epithelial Tumors. Int. J. Mol. Sci. 2023, 24, 4263. [Google Scholar] [CrossRef] [PubMed]
- Arjonen, A.; Mäkelä, R.; Härmä, V.; Rintanen, N.; Kuopio, T.; Kononen, J.; Rantala, J.K. Image-based ex vivo drug screen to assess targeted therapies in recurrent thymoma. Lung Cancer 2020, 145, 27–32. [Google Scholar] [CrossRef]
- Giaccone, G.; Rajan, A.; Berman, A.; Kelly, R.J.; Szabo, E.; Lopez-Chavez, A.; Trepel, J.; Lee, M.-J.; Cao, L.; Espinoza-Delgado, I.; et al. Phase II Study of Belinostat in Patients With Recurrent or Refractory Advanced Thymic Epithelial Tumors. J. Clin. Oncol. 2011, 29, 2052–2059. [Google Scholar] [CrossRef]
- Thomas, A.; Rajan, E.; Szabo, Y. Tomita, C. A. Carter. A Phase I/Ii Trial of Belinostat in Combination with Cisplatin, Doxorubicin, and Cyclophosphamide in Thymic Epithelial Tumors: A Clinical and Translational Study. Clin. Cancer Res. 2014, 20, 5392–402. [Google Scholar] [CrossRef]
- Badve, S.; Goswami, C.; Gökmen–Polar, Y.; Nelson, R.P.; Henley, J.; Miller, N.; Zaheer, N.A.; Sledge, G.W.; Li, L.; Kesler, K.A.; et al. Molecular Analysis of Thymoma. PLoS ONE 2012, 7, e42669. [Google Scholar] [CrossRef] [PubMed]
- Gokmen-Polar, Y., R. W. Cook, C. P. Goswami, J. Wilkinson, D. Maetzold, J. F. Stone, K. M. Oelschlager, I. T. Vladislav, K. L. Shirar, K. A. Kesler, P. J. Loehrer, Sr., and S. Badve. A Gene Signature to Determine Metastatic Behavior in Thymomas. PLoS ONE 2013, 8, e66047. [Google Scholar]
- Girard, N.; Shen, R.; Guo, T.; Zakowski, M.F.; Heguy, A.; Riely, G.J.; Huang, J.; Lau, C.; Lash, A.E.; Ladanyi, M.; Viale, A.; Antonescu, C.R.; Travis, W.D.; Rusch, V.W.; Kris, M.G.; Pao, W. Comprehensive Genomic Analysis Reveals Clinically Relevant Molecular Distinctions between Thymic Carcinomas and Thymomas." Clin. Cancer Res. 2009, 15, 6790–9.
- Sasaki, H.; Ide, N.; Fukai, I.; Kiriyama, M.; Yamakawa, Y.; Fujii, Y. Gene expression analysis of human thymoma correlates with tumor stage. Int. J. Cancer 2002, 101, 342–347. [Google Scholar] [CrossRef]
- Liang, N.; Liu, L.; Huang, C.; Liu, H.; Guo, C.; Li, J.; Wang, W.; Li, N.; Lin, R.; Wang, T.; et al. Transcriptomic and Mutational Analysis Discovering Distinct Molecular Characteristics Among Chinese Thymic Epithelial Tumor Patients. Front. Oncol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Radovich, M.; Solzak, J.P.; A Hancock, B.; Conces, M.L.; Atale, R.; Porter, R.F.; Zhu, J.; Glasscock, J.; A Kesler, K.; Badve, S.S.; et al. A large microRNA cluster on chromosome 19 is a transcriptional hallmark of WHO type A and AB thymomas. Br. J. Cancer 2016, 114, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, P.; Zhao, J.; Yang, L.; Wang, W. Identification of Molecular Characteristics and New Prognostic Targets for Thymoma by Multiomics Analysis. BioMed Res. Int. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Wang, X.; Lin, P.; Li, Y.; Chen, G.; Yang, H.; He, Y.; Li, Q.; Liu, R. Identification of potential agents for thymoma by integrated analyses of differentially expressed tumour-associated genes and molecular docking experiments. Exp. Ther. Med. 2019, 18, 2001–2014. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Wang, S.; Zhang, J.; Yan, Y.; Wang, C.; Yang, C.; Guan, Z.; Wang, C. Alteration in gene expression profiles of thymoma: Genetic differences and potential novel targets. Thorac. Cancer 2019, 10, 1129–1135. [Google Scholar] [CrossRef]
- Yu, L.; Ke, J.; Du, X.; Yu, Z.; Gao, D. Genetic characterization of thymoma. Sci. Rep. 2019, 9, 2369. [Google Scholar] [CrossRef]
- Lee, H. S. , Jang, H.J.; Shah, R.; Yoon, D.; Hamaji, M.; Wald, O.; Lee, J.S.; Sugarbaker, D.J.; Burt, B.M. Genomic Analysis of Thymic Epithelial Tumors Identifies Novel Subtypes Associated with Distinct Clinical Features. Clin Cancer Res 2017, 23, 4855–64. [Google Scholar] [CrossRef] [PubMed]
- Iaiza, A.; Tito, C.; Ganci, F.; Sacconi, A.; Gallo, E.; Masciarelli, S.; Fontemaggi, G.; Fatica, A.; Melis, E.; Petrozza, V.; et al. Long Non-Coding RNAs in the Cell Fate Determination of Neoplastic Thymic Epithelial Cells. Front. Immunol. 2022, 13, 867181. [Google Scholar] [CrossRef] [PubMed]
- Grillone, K.; Riillo, C.; Scionti, F.; Rocca, R.; Tradigo, G.; Guzzi, P.H.; Alcaro, S.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. Non-coding RNAs in cancer: platforms and strategies for investigating the genomic “dark matter”. J. Exp. Clin. Cancer Res. 2020, 39, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2018, 18, 5–18. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Luo, X.; Li, H.; Zhang, L.; Su, F.; Hou, S.; Yin, J.; Zhang, W.; Zou, L. Identification of differentially expressed circular RNAs associated with thymoma. Thorac. Cancer 2021, 12, 1312–1319. [Google Scholar] [CrossRef]
- Su, Y.; Chen, Y.; Tian, Z.; Lu, C.; Chen, L.; Ma, X. lncRNAs classifier to accurately predict the recurrence of thymic epithelial tumors. Thorac. Cancer 2020, 11, 1773–1783. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Jin, S.; Pan, X.; Wang, G.; Ye, L.; Tao, H.; Wen, H.; Liu, Y.; Xie, Q. Identification of Long Non-Coding RNAs for Predicting Prognosis Among Patients with Thymoma. Clin. Lab. 2018, 64, 1193–1198. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y.; Song, H. A Disparate Role of Rp11-424c20.2/Uhrf1 Axis through Control of Tumor Immune Escape in Liver Hepatocellular Carcinoma and Thymoma. Aging 2019, 11, 6422–39. [Google Scholar] [CrossRef]
- Obata, Y.; Furusawa, Y.; A Endo, T.; Sharif, J.; Takahashi, D.; Atarashi, K.; Nakayama, M.; Onawa, S.; Fujimura, Y.; Takahashi, M.; et al. The epigenetic regulator Uhrf1 facilitates the proliferation and maturation of colonic regulatory T cells. Nat. Immunol. 2014, 15, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Lu, T. X.; Rothenberg, M.E. Microrna. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Malumbres, M. miRNAs and cancer: An epigenetics view. Mol. Asp. Med. 2012, 34, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Enkner, F.; Pichlhöfer, B.; Zaharie, A.T.; Krunic, M.; Holper, T.M.; Janik, S.; Moser, B.; Schlangen, K.; Neudert, B.; Walter, K.; et al. Molecular Profiling of Thymoma and Thymic Carcinoma: Genetic Differences and Potential Novel Therapeutic Targets. Pathol. Oncol. Res. 2016, 23, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, H.; Zhang, X.; Ma, H. Loxl1-As1 Promotes Thymoma and Thymic Carcinoma Progression by Regulating Mir-525-5p-Hspa9. Oncol. Rep. 2021, 45. [Google Scholar] [CrossRef]
- Tito, C.; Ganci, F.; Sacconi, A.; Masciarelli, S.; Fontemaggi, G.; Pulito, C.; Gallo, E.; Laquintana, V.; Iaiza, A.; De Angelis, L.; et al. LINC00174 is a novel prognostic factor in thymic epithelial tumors involved in cell migration and lipid metabolism. Cell Death Dis. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ganci, F.; Vico, C.; Korita, E.; Sacconi, A.; Gallo, E.; Mori, F.; Cambria, A.; Russo, E.; Anile, M.; Vitolo, D.; et al. MicroRNA expression profiling of thymic epithelial tumors. Lung Cancer 2014, 85, 197–204. [Google Scholar] [CrossRef]
- Tan, S., and J. Chen. Si-Malat1 Attenuates Thymic Cancer Cell Proliferation and Promotes Apoptosis Via the Mir-145-5p/Hmga2 Pathway. Oncol. Lett. 2021, 22, 585. [Google Scholar] [CrossRef] [PubMed]
- Iaiza, A.; Tito, C.; Ianniello, Z.; Ganci, F.; Laquintana, V.; Gallo, E.; Sacconi, A.; Masciarelli, S.; De Angelis, L.; Aversa, S.; et al. METTL3-dependent MALAT1 delocalization drives c-Myc induction in thymic epithelial tumors. Clin. Epigenetics 2021, 13, 1–15. [Google Scholar] [CrossRef]
- Damron, B. The Relationship of Maximum or Intermediate Coccidiostat Levels to Broiler Chick Water Intake. Poult. Sci. 1994, 73, 33–36. [Google Scholar] [CrossRef]
- Miller, J.W.; Faubert, B.M.; Mathews, T.P.; Waters, J.K.; DeBerardinis, R.J.; Kernstine, K.H. Metabolic signatures of thymomas: potential biomarkers and treatment targets. Eur. J. Cardio-Thoracic Surg. 2023, 65. [Google Scholar] [CrossRef] [PubMed]
- Ying, J.; Huang, Y.; Ye, X.; Zhang, Y.; Yao, Q.; Wang, J.; Yang, X.; Yu, C.; Guo, Y.; Zhang, X.; et al. Comprehensive study of clinicopathological and immune cell infiltration and lactate dehydrogenase expression in patients with thymic epithelial tumours. Int. Immunopharmacol. 2024, 126, 111205. [Google Scholar] [CrossRef] [PubMed]
- Alwahsh, M.; Knitsch, R. Marchan, J. Lambert. Metabolic Profiling of Thymic Epithelial Tumors Hints to a Strong Warburg Effect, Glutaminolysis and Precarious Redox Homeostasis as Potential Therapeutic Targets. Cancers 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zeng, B.; Zhu, H.; Ma, R.; Yuan, P.; Chen, Z.; Su, C.; Liu, Z.; Yao, X.; Lawrence, A.; et al. Role of glycosphingolipid biosynthesis coregulators in malignant progression of thymoma. Int. J. Biol. Sci. 2023, 19, 4442–4456. [Google Scholar] [CrossRef]
- Tang, E.; Zhou, Y.; Liu, S.; Zhang, Z.; Zhang, R.; Huang, D.; Gao, T.; Zhang, T.; Xu, G. Metabolomic and Transcriptomic Profiling Identified Significant Genes in Thymic Epithelial Tumor. Metabolites 2022, 12, 567. [Google Scholar] [CrossRef]
- Regina Todeschini, A., and S.I. Hakomori. Functional Role of Glycosphingolipids and Gangliosides in Control of Cell Adhesion, Motility, and Growth, through Glycosynaptic Microdomains. Biochim. Biophys. Acta 2008, 1780, 421–33. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S. I. "Glycosynaptic Microdomains Controlling Tumor Cell Phenotype through Alteration of Cell Growth, Adhesion, and Motility. " FEBS Lett. 2010, 584, 1901–6. [Google Scholar] [CrossRef]
- Li, J., Y. Long. Comprehensive Landscape of the St3gal Family Reveals the Significance of St3gal6-As1/St3gal6 Axis on Egfr Signaling in Lung Adenocarcinoma Cell Invasion. Front. Cell Dev. Biol. 2022, 10, 931132. [Google Scholar] [CrossRef] [PubMed]
- Bremmer, F.; Bohnenberger, H.; Findeisen, P.; Welter, S.; von Hammerstein-Equord, A.; Hinterthaner, M.; Müller, D.; Küffer, S.; Okada, S.; Marx, A.; et al. Proteomic analysis identifies argininosuccinate synthetase 1 and special AT-rich sequence binding protein 1 as reliable markers for the immunohistochemical distinction between WHO types A and B3 thymomas. Histopathology 2023, 83, 607–616. [Google Scholar] [CrossRef]
- Delage, B.; Fennell, D.A.; Nicholson, L.; McNeish, I.; Lemoine, N.R.; Crook, T.; Szlosarek, P.W. Arginine deprivation and argininosuccinate synthetase expression in the treatment of cancer. Int. J. Cancer 2010, 126, 2762–2772. [Google Scholar] [CrossRef]
- Zhao, T.; Wu, J.; Liu, X.; Zhang, L.; Chen, G.; Lu, H. Diagnosis of thymic epithelial tumor subtypes by a quantitative proteomic approach. Analyst 2018, 143, 2491–2500. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Branson, O.E.; Shilo, K.; Hitchcock, C.L.; Freitas, M.A. Proteomic Signatures of Thymomas. PLoS ONE 2016, 11, e0166494. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.-L.; Fang, W.-T.; Feng, J.; Zhang, J.; Yang, X.-H.; Gu, Z.-T.; Zhu, L.; Sha, H.-F. Proteome analysis and tissue array for profiling protein markers associated with type B thymoma subclassification. . 2012, 125, 2811–2818. [Google Scholar] [PubMed]
- Lai, L.-C.; Sun, Q.-L.; Chen, Y.-A.; Hsiao, Y.-W.; Lu, T.-P.; Tsai, M.-H.; Zhu, L.; Chuang, E.Y.; Fang, W. Using proteomic profiling to characterize protein signatures of different thymoma subtypes. BMC Cancer 2019, 19, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Liu, P.; Qi, G. Exploring Potential Biomarkers of Early Thymoma based on Serum Proteomics. Protein Pept. Lett. 2024, 31, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qi, G.; Liu, Y. Proteomics analysis of serum from thymoma patients. Sci. Rep. 2023, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Qi, G.; Dong, H.; Liu, Z.; Ma, M.; Liu, P. Identification of Potential Serum Protein Biomarkers in Thymoma with Myasthenia Gravis After Docetaxel Treatment. Neurol. Ther. 2023, 12, 559–570. [Google Scholar] [CrossRef]
- Tong, T.; Zhang, J.; Jia, L.; Liang, P.; Wang, N. Integrated proteomics and metabolomics analysis reveals hubs protein and network alterations in myasthenia gravis. Aging 2022, 14, 5417–5426. [Google Scholar] [CrossRef]
- Hayashi, Y., N. Ishii. Thymoma: Tumour Type Related to Expression of Epidermal Growth Factor (Egf), Egf-Receptor, P53, V-Erb B and Ras P21. Virchows Arch. 1995, 426, 43–50. [Google Scholar] [CrossRef]
- Ionescu, D.N.; Sasatomi, E.; Cieply, K.; Nola, M.; Dacic, S. Protein expression and gene amplification of epidermal growth factor receptor in thymomas. Cancer 2005, 103, 630–636. [Google Scholar] [CrossRef]
- E Gilhus, N.; Jones, M.; Turley, H.; Gatter, K.C.; Nagvekar, N.; Newsom-Davis, J.; Willcox, N. Oncogene proteins and proliferation antigens in thymomas: increased expression of epidermal growth factor receptor and Ki67 antigen. J. Clin. Pathol. 1995, 48, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Pescarmona, E.; Pisacane, A.; Pignatelli, E.; Baroni, C. Expression of epidermal and nerve growth factor receptors in human thymus and thymomas. Histopathology 1993, 23, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Henley, J.; Koukoulis, G.; Loehrer, P. Epidermal growth factor receptor expression in invasive thymoma. J. Cancer Res. Clin. Oncol. 2002, 128, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Remon, J.; Abedallaa, N.; Taranchon-Clermont, E.; Bluthgen, V.; Lindsay, C.; Besse, B.; de Montpréville, V.T. CD52, CD22, CD26, EG5 and IGF-1R expression in thymic malignancies. Lung Cancer 2017, 108, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Girard, N.; Teruya-Feldstein, J.; Payabyab, E.C.; Riely, G.J.; Rusch, V.W.; Kris, M.G.; Zakowski, M.F. Insulin-Like Growth Factor-1 Receptor Expression in Thymic Malignancies. J. Thorac. Oncol. 2010, 5, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Zucali, P.A.; Petrini, I.; Lorenzi, E.; Merino, M.; Cao, L.; Di Tommaso, L.; Lee, H.S.; Incarbone, M.; Walter, B.A.; Simonelli, M.; et al. Insulin-like growth factor-1 receptor and phosphorylated AKT-serine 473 expression in 132 resected thymomas and thymic carcinomas. Cancer 2010, 116, 4686–4695. [Google Scholar] [CrossRef] [PubMed]
- Festenstein, H. MOLECULAR FEATURES OF THE H-2 CLASS I AND Qa ANTIGENS EXPRESSED ON GROSS VIRUS INDUCED AKR LEUKAEMIAS. Int. J. Immunogenetics 1989, 16, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Avci, N., G. Cecener. Molecular Markers for Patients with Thymic Malignancies: Not Feasible at Present? Asian Pac. J. Cancer Prev. 2014, 15, 3457–60. [Google Scholar] [CrossRef] [PubMed]
- Pfister, F.; Hussain, H.; Belharazem, D.; Busch, S.; Simon-Keller, K.; Becker, D.; Pfister, E.; Rieker, R.; Ströbel, P.; Marx, A. Vascular architecture as a diagnostic marker for differentiation of World Health Organization thymoma subtypes and thymic carcinoma. Histopathology 2017, 70, 693–703. [Google Scholar] [CrossRef]
- Lattanzio, R.; La Sorda, R.; Facciolo, F.; Sioletic, S.; Lauriola, L.; Martucci, R.; Gallo, E.; Palmieri, G.; Evoli, A.; Alessandrini, G.; et al. Thymic epithelial tumors express vascular endothelial growth factors and their receptors as potential targets of antiangiogenic therapy: A tissue micro array-based multicenter study. Lung Cancer 2014, 85, 191–196. [Google Scholar] [CrossRef]
- Angirekula, M.; Chang, S.Y.; Jenkins, S.M.; Greipp, P.T.; Sukov, W.R.; Marks, R.S.; Olivier, K.R.; Cassivi, S.D.; Roden, A.C. CD117, BAP1, MTAP, and TdT Is a Useful Immunohistochemical Panel to Distinguish Thymoma from Thymic Carcinoma. Cancers 2022, 14, 2299. [Google Scholar] [CrossRef] [PubMed]
- Kornstein, M.J.; Rosai, J. CD5 Labeling of Thymic Carcinomas and Other Nonlymphoid Neoplasms. Am. J. Clin. Pathol. 1998, 109, 722–726. [Google Scholar] [CrossRef]
- Jeong, J.-H.; Pyo, J.-S.; Kim, N.-Y.; Kang, D.-W. Diagnostic Roles of Immunohistochemistry in Thymic Tumors: Differentiation between Thymic Carcinoma and Thymoma. Diagnostics 2020, 10, 460. [Google Scholar] [CrossRef] [PubMed]
- Tiseo, M.; Damato, A.; Longo, L.; Barbieri, F.; Bertolini, F.; Stefani, A.; Migaldi, M.; Gnetti, L.; Camisa, R.; Bordi, P.; et al. Analysis of a panel of druggable gene mutations and of ALK and PD-L1 expression in a series of thymic epithelial tumors (TETs). Lung Cancer 2017, 104, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Aisner, S. C., S. Dahlberg. Epidermal Growth Factor Receptor, C-Kit, and Her2/Neu Immunostaining in Advanced or Recurrent Thymic Epithelial Neoplasms Staged According to the 2004 World Health Organization in Patients Treated with Octreotide and Prednisone: An Eastern Cooperative Oncology Group Study. J. Thorac. Oncol. 2010, 5, 885–92. [Google Scholar] [PubMed]
- Mimae, T.; Tsuta, K.; Kondo, T.; Nitta, H.; Grogan, T.M.; Okada, M.; Asamura, H.; Tsuda, H. Protein expression and gene copy number changes of receptor tyrosine kinase in thymomas and thymic carcinomas. Ann. Oncol. 2012, 23, 3129–3137. [Google Scholar] [CrossRef] [PubMed]
- Küffer, S.; Grabowski, J.; Okada, S.; Sojka, N.; Welter, S.; von Hammerstein-Equord, A.; Hinterthaner, M.; Cordes, L.; von Hahn, X.; Müller, D.; et al. Phosphoproteomic Analysis Identifies TYRO3 as a Mediator of Sunitinib Resistance in Metastatic Thymomas. Cancers 2022, 14, 4762. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, S.; Wang, Y.; Chen, Y.; Zhang, P.; Liu, Y.; Zhang, H.; Tao, Z.; Xiong, K. High expression of KITLG is a new hallmark activating the MAPK pathway in type A and AB thymoma. Thorac. Cancer 2020, 11, 1944–1954. [Google Scholar] [CrossRef]
Figure 1.
Summary of important histological and clinical features of the main thymoma subtypes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



