
Submitted:
27 January 2024
Posted:
31 January 2024
You are already at the latest version
Abstract
Fire blight, caused by the plant-pathogenic bacterium Erwinia amylovora, is a highly contagious and difficult-to-control disease due to the efficient dissemination, survival, and scarcity of effective control methods. Copper and antibiotics are the most used treatments but pose environmental and human health risks. Bacteriophages (phages) constitute an ecological, safe, and sustainable fire blight control alternative. The goal of this study was to search for specific E. amylovora phages from plant material, soil, and water samples in Mediterranean environments. A collection of phages able to specifically infect and lyse E. amylovora strains was generated from former fire blight-affected orchards in Eastern Spain. Following in vitro characterization, assays on immature fruit revealed that preventively applying some of the phages or their combinations delayed the onset of fire blight symptoms and reduced the disease severity, suggesting their biocontrol potential in Spain and other countries. Morphological and molecular characterization of the selected E. amylovora phages classified them as members of the class Caudoviricetes (former Myoviridae family) and genus Kolesnikvirus. This study reveals Mediterranean settings as plausible sources of E. amylovora-specific bacteriophages, and provides the first effective European phage cocktails in plant material for the development of sustainable fire blight management measures.
Keywords:
phytopathogenic bacteria
; biological control
; phages
; environmental samples
; lysis
; specificity
; pear trees
; loquats
1. Introduction
Erwinia amylovora (Burrill)[1] a bacterial species belonging to the family Erwiniaceae [2] is the etiological agent of the highly infectious fire blight disease. This pathogen affects a large number of species of the Rosaceae family, including economically important fruit trees such as apple, pear, loquat and quince, and also ornamental and wild plants [3,4]. The host shoots, leaves, flowers, fruits, and rootstock are susceptible to the disease, with infections significantly reducing fruit production and ultimately causing tree death. Under optimal conditions, fire blight outbreaks can destroy entire orchards in a single season, disrupting fruit production for years and leading to devastating economic losses in the areas where it is present [5,6,7].
The first report of fire blight dates from 1780, on the detection of the disease in the Hudson Valley, in New York State (USA). Subsequently, it spread throughout North America and, in the 20th century, reached New Zealand, Egypt and England through trading of infected plant material, and afterwards the disease spread all over Europe and the Mediterranean countries [4].
In Spain, E. amylovora was first detected in 1995, in the North of the country. Since then, some other outbreaks have been reported in Northern, Central and Eastern Spanish provinces. The current pest situation evaluated by the European and Plant Protection Organization (EPPO) is that the pathogen is present with restricted distribution, and under eradication [8]. According to European Union (EU) Reglamentation, in Spain and the rest of the EU member states, E. amylovora is considered a Protected Zone Quarantine Pest (PZQP) [9]. The most effective approach to combat fire blight disease involves an integrated management encompassing prevention measures, early detection, and disease control strategies. Agrochemicals such as antibiotics (e.g., streptomycin) and copper compounds usually provide the most effective control against E. amylovora. However, their persistent agricultural application has given rise to concerns regarding their long-term efficacy and sustainability. In the EU, agricultural use of antibiotics is prohibited, and that of copper-based compounds is highly restricted [10]. The selection of antibiotic resistant and copper tolerant strains [10,11,12,13,14], together with the negative effects of these compounds on the environment and human health, have led to a rising demand for safer and more sustainable agricultural practices [15,16,17,18,19].
The use of biological agents is a promising, environmentally friendly, and sustainable alternative to copper and antibiotics[20]. Among them, bacterial viruses or bacteriophages (phages) are natural bacterial predators that can specifically infect and destroy their bacterial target. Bacteriophage-based treatment of bacterial plant diseases has received renewed interest in the last years with several studies reporting promising control against important plant pathogenic bacterial species, including E. amylovora [21,22,23,24,25,26,27,28,29].
E. amylovora phages can be isolated from infected plants and the surrounding soil and water [30,31,32,33], although their success in biocontrol is variable [34]. A key factor determining bacteriophage treatment efficacy, apart from their lytic nature, is their ability to persist on the phyllosphere. UV light, and other environmental factors limit the success of phage application [35,36]. Some strategies to circumvent these challenges can be planning applications late in the day, working on protective formulations, and using carrier bacteria that minimize the effects of UV irradiation [37,38]. However, isolation of phages adapted to the climate conditions of the locations where these will be applied, or at least similar to them, could help maintaining high phage populations on the plant surfaces and improve their efficacy.
As a preliminary step for the design of phage-based treatments against the fire blight disease, the goal of this study was to isolate and characterize E. amylovora-specific bacteriophages from Mediterranean regions in Spain, where the disease was present. We tested the in vitro activity of different phage isolates against a collection of E. amylovora strains. Additionally, because E. amylovora is a quarantine organism in the European Union and testing products in the field or even a regular greenhouse is not possible, we assessed the efficacy of preventive and control phage treatments using immature fruit. Based on their morphology, the bacteriophages isolated in this study were myovirus. When applied preventively in immature fruits, i.e., before infection happens, the isolated phages demonstrated disease control capabilities. Our research addresses the critical need for effective and environmentally sustainable fire blight management strategies in Mediterranean and/or Mediterranean-like climate areas, where existing disease control measures are limited. The evidence provided in our study constitutes the first steps toward designing a bacteriophage-based treatment to mitigate the impact of fire blight disease in these settings. Besides, this work opens avenues for further investigation into the adaptability and efficacy of the isolated phages and phage cocktails in climatic conditions other than the Mediterranean. On top of that, the lytic phage cocktails assayed in this work constitute the first described in Europe with effective biocontrol activity against E. amylovora in plant material, which significantly adds valuable insights into the use of phage therapy in agriculture for plant disease control.
2. Materials and Methods
2.1. Bacterial Strains and General Growth Conditions
Spanish E. amylovora strains, UPN 527, IVIA 1554, IVIA 1526.6, IVIA 1614.1, IVIA 1614.2 and IVIA 1892.1, from different locations and host plant species (Table 1), were used as host strains for phage isolation and propagation. Strains were routinely cultured on the general media Luria Bertani Agar (LBA) [39] and Nutrient Agar amended with Sucrose and Yeast Extract (NASYE) [40] at 28°C for 48 h. For bacteriophage isolation and amplification, liquid versions of the previous media (LB and NBSYE) were employed, also at 28°C, with shaking at 150 r.p.m.
Isolation of E. amylovora from plant material, and soil and water environmental samples (Table 1), as well as pathogen re-isolation from inoculated fruit were performed on semi-selective CCT medium [41]. For phage host range and specificity assays, a collection of 19 E. amylovora strains (Table 1) and 22 bacterial species were used (Table 2). In this case, E. amylovora strains were grown on NASYE and the other plant pathogenic bacteria on LBA, King’s B Agar (KBA) [42] or Yeast Extract Peptone Glucose Agar (YPGA) [43], depending on the bacterial species. Pure cultures of all the strains were cryopreserved at −80 °C in 25% (v/v) glycerol. Quarantine bacteria and samples were handled in a microbiology laboratory under Biosecurity Level 2 (BSL2) conditions.
2.2. Phage Enrichment, Isolation, Purification and Amplification
Eight pear tree plots in the Valencian Community (Eastern Spain) with localized fire blight outbreaks were surveyed, and five of them sampled in late spring of 2018. Samples consisted of shoots, leaves, flowers, and fruits from symptomatic and asymptomatic trees, as well as soil from below symptomatic trees and, when possible, from environmental water nearby the affected orchards. Once collected, samples were placed into sterile bags and stored at 4°C until use.
For phage isolation, samples of 1, 5 and 10 g of each plant material type were washed with sterile distilled water for 1 h prior to be crashed with antioxidant maceration buffer [44]. Volumes from washing and crushing of plant material samples were mixed in a ratio 1:1 with NBSYE 2X medium inoculated separately with different E. amylovora strains (UPN 527, IVIA 1554, IVIA 1526.6, IVIA 1614.1, IVIA 1614.2, IVIA 1892.1 and CFBP 1430) adjusted to OD600nm of 0.4 and incubated at 28 °C with shaking. Similarly, samples of 10 g or 10 mL of soil or water, respectively, were mixed with NBSYE 2X and inoculated as described above. The E. amylovora-specific phage content in the samples was enriched similar to Gill et al.[40] for 24 and 48h at 28°C and 150 r.p.m. Cell debris from cultures showing lysis were pelleted by centrifugation (10,000 r.p.m., 10 min) and filtered through 0.22-μm pore diameter filters to obtain clean phage suspensions. E. amylovora-inoculated medium without samples were used as negative lysis control.
For phage purification, filtered lysates were analyzed by serial decimal dilutions in SM buffer (200 mM NaCl2; 10 mM MgSO4; 50 mM and Tris-HCl. pH 7.5) and plated using the double-layer agar method. For this, 0.2 mL aliquots of bacterial culture (OD600nm = 0.4) were mixed with 0.1 mL of phage filtrate dilution and 5 mL top agar (NASYE, 0.6 % agar), poured onto NASYE plates and incubated at 28 °C for 24 h.
After incubation, morphologically different plaque-forming units (PFU) were picked and purified with the respective host strain, and this process was repeated until obtaining identical plaques.
Purified phages were propagated in NBSYE medium and titered as PFU/mL by the double agar overlay assay. Pure phage suspensions were maintained for short periods at 4ºC and cryopreserved at -20 °C in 25% glycerol for longer periods.
2.3. Phage Host Range and Specificity
The host range of the isolated phages was performed by the spot test according to Born et al. [30], using a collection of Spanish and reference E. amylovora strains isolated from fire blight outbreaks in different host plants and locations, listed in Table 1. Specificity analysis of a selection of phages was also assayed by the spot test, against bacterial strains representing species other than E. amylovora, including other phytopathogenic bacteria listed in Table 2. To do so, melted top agar was mixed with 0.2 mL cultures (OD600nm = 0.4) of the tested bacterial species and poured onto solid plates. Next, 5 μL of filtered, pure phage lysates at about 108 PFU/mL were spotted onto the top agar. Spots with cultures of the E. amylovora strains used for propagation were included as positive controls. Once the phage droplets were absorbed by the medium, plates were incubated overnight at 28 °C. Areas of lysis below the phage spots were scored as positive, while those where no changes were observed with the surrounding lawn were scored as negative. Additionally, the lytic activity of the selected phages was confirmed in liquid NBSYE medium. All the assays were repeated in independent experiments.
2.4. Bacteriophage-Based Biocontrol Assays In Vitro
2.4.1. E. amylovora Biocontrol In Vitro by Single Phages.
E. amylovora population dynamics were monitored in the presence of single phages. Growth dynamics of single phages were assayed by co-inoculation of the host strain and the selected phage in microtiter plates using a spectrophotometer plate reader (Tecan Infinite MNano) for 24 h at 28 °C and shaking, performing OD600 nm readings at regular intervals. Each well contained 0.2 mL of the host culture at around 106 CFU/mL and phage at 10⁷ PFU/mL. Control wells containing only NBSYE medium, host strain and phage suspensions alone were included. At least three replicates per phage and host strain were performed in at least two independent assays.
Comparisons between the growth of E. amylovora strains alone and with individual phages were assessed during an incubation period of 23 h by calculating the Areas Under the Curve (AUC) for each treatment. Then, AUCs of the different treatments were compared by one-way Brown-Forsythe ANOVA tests (α = 0.05) designed for comparisons of groups with different SDs. Multiple comparisons between the controls and the treatments were carried out by Dunnett’s T3 post-hoc analyses (α = 0.05).
2.4.2. E. amylovora Biocontrol In Vitro by Phage Combinations
E. amylovora population dynamics were also monitored in the presence of phage cocktails or mixes. The growth dynamics of selected phage combinations or mixes (cocktails) were assayed as described above, by co-inoculation with single E. amylovora strains.
2.5. Bacteriophage-Based Biocontrol Assays Ex Vivo
2.5.1. E. amylovora Biocontrol on Immature Fruits by Single Phages
Assays on immature fruit were carried out on loquats (Eriobotrya japonica cv. Tanaka) according to [20,45]. Briefly, 2.5–3 cm diameter fruit were surface-disinfected with 2 % sodium hypochlorite, rinsed with sterile distilled water, and dried under the hood. Phage suspensions were prepared in SM buffer at 108 PFU/mL. E. amylovora suspensions were prepared in PBS at an OD600 of 0.2 (around 108 CFU/mL) using NASYE plate cultures and diluted to 106 CFU/mL. Disinfected fruit were wounded in three equidistant points with a sterile 100-μL pipette tip and used for fire blight preventive and control assays.
To test preventive treatments with bacteriophages, each wound was inoculated with 10 µL phage at 108 PFU/mL (106 PFU/wound) 24 h before inoculation with 10 µL of E. amylovora suspension at 106 CFU/mL (104 CFU/wound). For disease control (co-inoculation) experiments, 10 µL phage plus 10 µL of E. amylovora were co-inoculated in each wound. Fruits inoculated only with PBS, SM, phages or E. amylovora alone were included as controls. Phages were assayed in three replicate fruit in at least two independent experiments. Challenged fruit were incubated at 28 °C in humid chambers, as described previously [20,45]. Fruit were monitored for the onset, incidence and severity of fire blight symptoms.
The efficacy of E. amylovora fire blight disease control by the tested phages, as well as severity of symptoms were evaluated along 6 days after inoculation. Briefly, the disease control efficacy % (E) was determined as the E = (Ic - It / T) x 100, where Ic is the number of infected wounds in the phage-untreated E. amylovora-inoculated controls; It is the number of infected wounds in fruit treated with both E. amylovora and phages; and T is the total wound number in each assay repetition. The disease severity after 6 days of incubation was determined after classifying symptoms in 5 disease severity levels, where 0 was absence of symptoms; 1, sunken tissues, chlorinated tissue, or shade areas around the wound indicative of infection; 2, up to 2-mm-wide necrotic lesions around the wound; 3, up to 4-mm-wide necrotic lesions around the wound; 4, expanding necrosis (≥ 4 mm wide), usually reaching the peduncle and/or calyx areas of the fruit. The disease severity % (S) was calculated as S = , where i is the severity index class, Ni is frequency within the assay, T is the total number of wounds and 4 represents the maximum severity index used for classification.
2.5.2. E. amylovora Biocontrol on Immature Fruits by Phage Combinations
In this case, selected phage combinations or cocktails were used similarly as explained above, for preventive and co-inoculation experiments. The phage cocktails assayed were mix 1 (UV_Eaϕ21, UV_Eaϕ24 and UV_Eaϕ28), mix 2 (UV_Eaϕ6, UV_Eaϕ21, UV_Eaϕ24 and UV_Eaϕ28), mix 3 (UV_Eaϕ6, UV_Eaϕ21, UV_Eaϕ24, UV_Eaϕ25 and UV_Eaϕ28) and mix 4 (UV_Eaϕ6, UV_Eaϕ21, UV_Eaϕ24, UV_Eaϕ25, UV_Eaϕ27 and UV_Eaϕ28). Fruit were inoculated, incubated, monitored, sampled and processed as described above.
Statistical analysis of data obtained from biocontrol assays was performed using GraphPad Prism version9.
2.6. Phage Morphology
Phage virions from 0.22-μm-filtered freshly prepared phage lysates (about 1010 PFU/mL) obtained as above-mentioned (paragraph 2.2.) were prepared for transmission electron microscopy (TEM) as described by Biosca et al. [46]. In short, 5 μL lysates were absorbed on fresh formvar and carbon covered grids for 1 min. Excess of sample was removed and the grids were stained with 1 % of phosphotungstic acid (pH 7.0) for 1 minute and air-dried. Electron microscopic visualizations of the phage virions were performed using the TEM Hitachi HT7800 operated at 120 kV and with 20 Mpx CMOS EMSIS XAROSA digital camera, at the Central Service for Experimental Research (SCSIE) facility (Universitat de València, Burjassot, Spain). Phage virion dimensions (head diameter and tail length) were determined on micrographs with ImageJ software version 1.53m [47] in at least 10 viral particles from each phage.
2.7. Phage Molecular Characterization
2.7.1. Phage Genome Sequencing
DNA extraction from selected phages was performed according to Biosca et al. [46] Shortly, bacterial nucleic acids in filtered phage lysates were eliminated by treatment with DNase and RNase for 1 h at 37°C, followed by heat inactivation with EDTA for 10 min at 37 °C.
Phage DNA was isolated with the NucleoSpin® Plasmid isolation kit (Macherey-Nagel, Düren, Germany), following the protocol for Low-Copy plasmid isolation.
Total DNA quantification was performed using a Qubit 4TM Fluorometer (ND-2000, ThermoFisher, Wilmington, DE, USA) with dsDNA HS Assay KitTM. DNA libraries were made with Nextera XT Library Preparation Kit (Illumina, USA) following the manufacturer’s instructions. Sequencing was carried out using the Illumina MiSeq platform with 2 × 250 bp paired end sequencing (Illumina, USA). Raw data read output was subjected to quality control, trimming and de novo assembly with CLC Genomics Workbench 10.1.1 software (QIAGEN, Hilden, Germany). De novo contigs were analyzed by BLASTn and BLASTx [48].
2.7.2. Phage Genome Annotation
Open Reading Frames (ORFs) search in de novo assembly contigs was performed with Geneious Prime® 2023.2 based on nucleotide similarity with RefSeq genome NC_041978. In addition, genome annotations were confirmed by RASTtk [49], and all annotated ORFs were reviewed and refined through database searches using the BLASTp [48]
For the specific search of proteins on the newly assembled genomes, phage proteins were downloaded from NCBI (20,006 Endolysins, 1,331 Holins and 608 Depolymerases, accessible in Dec. 2023) and were compared using tBLASTn against the new assembled genomes, using Geneious Prime® 2023.2 software. All annotated genomes were deposited on NCBI database (PP079182, PP079183, PP079184, PP079185, PP079186 and PP079187).
Prediction of lytic or lysogenic lifecycle was performed with the tool PhageAI [50].
2.7.3. Phage Phylogenetic Analysis
Phylogenetic analyses were performed with the whole genomes of the six Spanish phages and a selection of 28 RefSeq viral genomes belonging to subfamily Ounavirinae original from China, Lithuania, Vietnam, Canada, South Korea, Spain and the USA.
Accurate complete genome alignment was performed with MAFFT V.7 [51]. The phylogenetic tree was reconstructed in MEGA X [52], using the maximum likelihood algorithm, supported using 500 bootstrap replicates, and selecting the best substitution model computed (GTR + G) implemented. In addition, a proteomic tree was constructed from a selection of reference genomes with the ViPTree web server [53] based on normalized tBLASTx scores. For the viral proteomic tree figure, Newick format file was downloaded from VipTree web server and visualized in Geneious Prime® 2023.2 software.
3. Results and Discussion
3.1. Isolation of Mediterranean E. amylovora-Specific Bacteriophages was Linked to the Presence of Symptomatic Plant Material
A collection of 124 bacteriophages was isolated from four out of the five analyzed plots. Most isolated phages came from plant material (79.8 %). Specifically, all the positive phage isolations came from symptomatic pear tree samples. The next main source of E. amylovora bacteriophages was soil underneath affected pear trees (16.9 % phages), followed by environmental water near the affected trees (3.2 % phages). Symptomatic plant material and the soil beneath diseased trees are frequently reported as good sources of E. amylovora bacteriophages [31,40,54].
The isolation of bacteriophages from water sources in fire blight-affected areas might indicate the presence of E. amylovora cells in the water. Under laboratory conditions, the pathogen can survive in distilled water, mineral water, rainwater and river water for variable periods depending on the incubation temperature, the presence of microbiota, the type of water, etc. [55,56,57]. However, our attempts to isolate the pathogen in the water samples where the phages were found were unsuccessful. Although the transitory presence of E. amylovora cannot be discarded, it is worth mentioning that phage concentrations are usually ten times higher than those of their bacterial target in the environment [58,59]. Another possibility, apart from a native higher survival capability, could be that the isolated phages are also natural predators of other host species present in the water. Indeed, wastewater has been reported as a source of bacteriophages against E. amylovora and other plant pathogens [60,61].
3.2. Mediterranean Bacteriophages were Active against E. amylovora Strains from Different Locations and Hosts
All the isolated bacteriophages showed lytic activity against different E. amylovora strains, including Spanish, French, Serbian and American strains isolated from pear, apple, and ornamental and wild host species of the genera Cotoneaster, Crataegus, and Pyracantha. Among the collection of bacteriophages isolated initially, 28 showed lytic activity against 58-100 % of the tested E. amylovora strains according to a spot assay (Table 3). Information about the origin of the 28 selected bacteriophages UV_Eaϕ1 to UV_Eaϕ28 (hereinafter abbreviated as ϕ1 to ϕ28) can be found in Supplementary Table S1).
Despite the high genome homogeneity of the E. amylovora species [62], bacteriophages active against a set of E. amylovora strains usually lack activity against a more or less considerable percentage of the tested strains [31,63]. These differences have been associated with factors such as the geographical origin of the host strains and their relative amylovoran production. The capacity of bacteriophages to infect different E. amylovora strains also probably correlates with differences in the host strain CRISPR system, the receptors for phage infection [63] and other unexplored resistance mechanisms in this pathogen.
Specificity tests with the selected 28 bacteriophages showed no lytic activity against bacterial species other than E. amylovora, including plant pathogens and other Gram-positive and Gram-negative species from different countries and sources (Table 3).
Assays in liquid culture confirmed the lytic activity of the 28 bacteriophages against a selection of nine Spanish E. amylovora strains. Some phages that apparently did not lyse certain E. amylovora strains in spot assays, showed high activity against the same strains in liquid medium. As an example, phages ϕ15, ϕ20, ϕ22-24, and ϕ27-28, which did not show lytic activity against E. amylovora IVIA 1892.1 in a spot assay (Table 3), were able to completely lyse populations of the same strain for 20-23 h in liquid medium (Figure 1A). Experimental method-dependent differences in the lytic activity of bacteriophages have been reported by different authors [63,64]. The reasons behind these differences can be explained by the diversity of phage resistance mechanisms and the infection steps each method favors, can detect and/or relies upon [65]. Factors affecting the host cell growth, production of exopolysaccharides, etc., may also contribute to variability in the results depending on the methodology and conditions employed.
All the selected Mediterranean phages exerted control of E. amylovora populations in liquid cultures, efficiently reducing the bacterial populations to the initial numbers for 15-23 h depending on the tested host strain and phage (Figure 1). The most sensitive strain was IVIA 1892.1, which was lysed by all the bacteriophages with barely any changes in the OD600nm values from time 0 to 20-23 h (Figure 1A). In the remaining E. amylovora strains, towards the last 3-4 h of the experiment, a slight increase of bacterial populations was observed while treated with different phages (Figure 1B and C), similar to other works [66,67]. This can be explained by the selection of phage-resistant host cell subpopulations under the assayed in vitro conditions [59,68,69,70].
Regardless of the assayed host strain and phage, the OD600nm values within the assayed period (23 h) were significantly lower (P < 0.0001) in E. amylovora cells treated with individual bacteriophages than when grown alone, as revealed by AUC comparison analysis (Figure 1).
3.3. The Designed Phage Mixes Control E. amylovora Populations Under In Vitro Conditions
In this study, four different phage mixes with combinations of bacteriophages ϕ6, ϕ21, ϕ24, ϕ25, ϕ27 and ϕ28 were designed. These phages were selected based on their origin (plant material, soil, and water) (Supplementary Table S1) and their lytic activity against E. amylovora strains from different geographical locations and hosts (Table 3). Again, those phages selected that did not appear to lyse some strains of E. amylovora by spot test, did show lytic activity against the same strains in liquid medium (Table 3). The phage composition of the four mixes were: MIX 1, phages ϕ21, ϕ24 and ϕ28; MIX 2, phages ϕ6, ϕ21, ϕ24 and ϕ28; MIX 3, phages ϕ6, ϕ21, ϕ24, ϕ25 and ϕ28; and MIX 4, phages ϕ6, ϕ21, ϕ24, ϕ25, ϕ27 and ϕ28.
Figure 2 shows the activity of the 4 phage mixes against 3 Spanish E. amylovora strains from different locations and hosts. In all cases, the phage mixes reduced E. amylovora populations to around the initial levels throughout the assayed period (P < 0.0001). Moreover, the slight increase of E. amylovora populations seen at the end of the experimental period when using individual phages (Figure 1) was either suppressed, as in the case of strain IVIA 1614.2 (Figure 1C vs Figure 2C) or reduced compared to when using many of the assayed individual phages against strain IVIA 1526.6 (Figure 1B vs Figure 2B).
The use of phage cocktails instead of individual phages has some advantages. Co-evolution of phages and pathogens in the same environments may lead to the accumulation of different phage-resistance mechanisms in the host cells. These mechanisms may target the phage adsorption to the host cell surface and other stages of the phage infection cycle [69,71]. The emergence of phage-resistant bacteria can be minimized using optimized mixtures of bacteriophages with different mechanisms of action and/or adaptation to different conditions. The range of virulence genes and degrees of environmental tolerance of the phages contained in a phage cocktail may also help compensate for each other's limitations, enhance each other's abilities and/or improve their global efficacy under varied conditions [22,72,73]. The phages in a mix might also use different host cell surface receptors and strategies to overcome bacterial defense mechanisms against viral nucleic acids, like CRISPR-Cas systems, restriction-modification systems, etc. [74]. This makes it harder for bacteria to develop multiple resistance mechanisms simultaneously, contributing to the control efficacy of the mix. Finally, cocktails with phages with different host specificities may also help broaden the range of target bacterial strains [75].
3.4. Preventive Application of Phage Mixes Increases the Disease Control Efficacy and Reduces Symptom Severity with Respect to Individual Phages in Detached Fruit Assays
Figure 3 shows the individual phage activities against the Spanish E. amylovora strains IVIA 1892.1 (Figure 3A, C and E) and IVIA 1526.6 (Figure 3B, D and F). A two-way ANOVA analysis revealed that, regardless of the assayed E. amylovora strain, both the timing of phage application (preventive vs co-inoculation) and the treatment type (with or without the different phages) had a very significant effect on the outcome (in both cases, P < 0.0001).
In experiments with the ϕ6, ϕ21, ϕ24, ϕ25, ϕ27 and ϕ28 phages and E. amylovora strain IVIA 1892.1, only phages ϕ21 and ϕ25 provided a significant control efficacy of around 56 % but, only when applied preventively (P < 0.0001) (Figure 3A). This application method not only improved the disease control efficacy percentage but, also significantly reduced the disease severity in inoculated fruits. Preventive treatments with phages ϕ21 and ϕ25, and also ϕ24 and ϕ27 revealed disease severity percentages of 11.1 % (P < 0.0001), 25.0 % (P < 0.0001), 41.7 % (P = 0.0014) and 58.3 % (P = 0.0358), compared with the 100 % observed in the fruit inoculated with E. amylovora alone (Figure 3C and E). In most cases, bacteriophage treatments were ineffective when co-inoculated with E. amylovora (Figure 3A, C and E). One exception was the treatment with ϕ27, which reduced the disease severity by around 53 % (P < 0.0043) with respect to treatments with E. amylovora alone (Figure 3C and E).
The Spanish E. amylovora strain IVIA 1526.6 was less virulent than the strain IVIA 1892.1 (Figure 3) but, overall, we observed similar trends as with both strains. Preventive phage application provided an overall better result in controlling the incidence and the severity of symptoms than when co-inoculated with E. amylovora (P < 0.0001). In this case, treatments with phages ϕ6, ϕ24, and ϕ21 conferred 100.0 %, 89.0 %, and 66.7 % disease control efficacy, respectively, when applied 24 h before E. amylovora inoculation. However, only the results with phages ϕ6 and ϕ24 were statistically significant (P ≤ 0.0273) (Figure 3B). Regarding the disease severity, preventive phage application avoided symptom development (phages ϕ6, ϕ25, ϕ27 and ϕ28) and/or provided significant symptom reduction (phages ϕ21 and ϕ24), all these effects being statistically significant (P ≤ 0.0107) (Figure 3D and F). Co-inoculation treatments with E. amylovora IVIA 1526.6 had no significant effects on disease control efficacy (Figure 3B). Although all the co-inoculated fruit showed clear reductions in disease severity, none of the observed effects resulted in statistical significance (P ≥ 0.1231) (Figure 3D and F).
The performance of the bacteriophages improved when mixed in cocktails (Figure 4). Like the application of individual phages, the co-inoculation of the phage mixes plus E. amylovora did not significantly improve the disease control efficacy (Figure 4A). However, preventive use of the bacteriophage mixes 2, containing phages ϕ6, ϕ21, ϕ24, and ϕ28, and 3, containing the latter phages plus ϕ25, provided a significant increase in the disease control efficacy of around 56 % (P ≤ 0.0136). The most significant effects of the phage mixes were linked to the reduction of symptom severity (Figure 4 B and C). In this case, both the preventive application and co-inoculation of the phage mix with the pathogen significantly reduced the disease severity. However, the best results were obtained after the preventive application of the phage mix (Figure 4B and C). For instance, the bacteriophage mixes reduced symptom severity from 83.3 % in the fruit inoculated with E. amylovora IVIA 1892.1 alone, to 11.1 % - 38.9 % when applied 24 h before E. amylovora inoculation (P ≤ 0.0004), and to 22.2 % - 47.2 % when co-inoculated with the pathogen (P ≤ 0.0040).
To explore the efficacy of the E. amylovora phage cocktails against E. amylovora strains from countries other than Spain, their performance against the French E. amylovora reference strain CFBP 1430 was also tested (Figure 5). The assays in liquid medium showed that all the phage mixes were able to limit E. amylovora growth to OD600nm values under those of the untreated controls (P < 0.0001) (Figure 5A). Among the assayed phage cocktails, mixes 2-4 kept E. amylovora populations at levels close to the initials throughout the assay. However, as discussed above, mix 1 delayed the emergence of a subpopulation of phage-resistant host cells for about 7 h, which grew to OD600nm values that were half of those reached by the controls towards the end of the experiment. These observations do not necessarily guarantee poor outcomes regarding the phage mix biocontrol activity. Development of phage-resistance in other bacterial pathogens has been shown to trigger physiological changes in the resistant bacteria with trade-offs in virulence, growth rates and other fitness-related traits [76,77]. The delayed occurrence of phage-resistant cells and lower pathogen cell numbers in the presence of the phages might also influence the host cell overall ability to thrive in the plant tissues and cause infections. Linked to this, despite the results observed in vitro (Figure 5A), the assay with detached fruit revealed that the phage mix 1 was as effective or even more in controlling the disease than the other three mixes, reaching control efficacy values of 66.7 % (P < 0.0001) (Figure 5A), and reducing the symptom severity to around 13.9 % compared to the 100 % developed by the pathogen inoculated alone (P < 0.0001) when applied preventively (Figure 5C and D).
The other E. amylovora phage mixes also affected the onset of symptoms, symptom development and disease severity when assayed in detached fruits. Out of the three remaining phage mixes (mixes 2-4), only mix 2 achieved a significant disease control efficacy of around 33 % when applied preventively (P = 0.0132) (Figure 5B).
None of the mixes provided effective control when co-inoculated with the pathogen. After being applied preventively, mixes 2-4 reduced symptom severity to 30.3 % - 52.8 % (P ≤ 0.0009). However, unlike mix 1, these mixes also achieved a significant reduction of the symptom severity when co-inoculated with the pathogen, reaching percentages of 58.3 % - 69.4 % compared to the 100 % observed in the controls (P ≤ 0.0428) (Figure 5C and D).
Overall, assays on immature fruit revealed that preventive application of some of the Mediterranean E. amylovora phages or their combinations not only delayed the onset of fire blight symptoms but also reduced the disease severity, suggesting their biocontrol potential in Spain and other countries. Given that, so far, there are no registered phage cocktails available in EU countries for plant pathogenic bacteria, including E. amylovora [77], this study is the first to design European phage cocktails against the fire blight pathogen.
3.5. Mediterranean E. amylovora Phages are Myovirus
The six bacteriophages observed by transmission electron microscopy showed similar morphologies, with icosahedral heads between 60-78 nm in diameter, collar and long, contractile tails between 93-102 nm (Figure 6). Phage ϕ6 (Figure 6A) has a capsid of 61.11 ± 5.06 nm and a tail of 93.02 ± 3.31 nm. The capsid of phage ϕ21 (Figure 6B) measures 59.82 ± 3.98 nm and the tail 94.56 ± 7.45 nm. Phage ϕ24 (Figure 6C), presents a capsid of 69.28 ± 5.32 nm and a tail of 94.22 ± 4.53 nm in length. Phage ϕ25 (Figure 6D) has a capsid of 63.85 ± 5.00 nm and a tail of 94.23 ± 9.23 nm. Phage ϕ27 (Figure 6E) displays a capsid of 63.46 ± 4.62 nm and a tail of 101.92 ± 4.62 nm. Finally, phage ϕ28 (Figure 6F) is the largest, with a capsid of 78.63 ± 7.41 nm in diameter and tails of 102.77 ± 5.4 nm in length.
Following the classification model by Tolstoy et al. [78], distinctive features of the myovirus (former Myoviridae family) include contractile, more or less rigid and long (80-455 nm) tails separated from the head by a collar. According to measurements and micrographs of the bacteriophages visualized with the transmission electron microscope (Figure 6), the six Mediterranean E. amylovora phages could be morphologically identified as myovirus, present class Caudoviricetes in accordance to recent taxonomy update of the ICTV bacterial viruses subcommittee [79].
3.6. Mediterranean E. amylovora Bacteriophages are Members of the Genus Kolesnikvirus
The sequencing of the genomes from phages ϕ6, ϕ21, ϕ24, ϕ25, ϕ27 and ϕ28 (hereinafter referred to vEam_PM_6, vEam_PM_21, vEam_S_24, vEam_W_25, vEam_PM_27 and vEam_W_28, respectively, where PM stands for plant material, S stands for soil and W stands for water), all of them isolated in the Mediterranean settings, resulted in the assembly of six genomes ranging in size from 84,686 to 84,690 nt, with a consistent GC content of 43.41 % in all cases. Annotation of the Open Reading Frames (ORFs) in these six genomes revealed a consistent coding of 117 in all cases. Additionally, analyses searching for other conserved domains of interest indicated that they do not encode any new endolysins, holins, or depolymerases, based on the data available in December 2023. PhageAI classified the six Mediterranean E. amylovora bacteriophages as virulent or lytic phages with a predicted value of 100 %.
BLASTn analysis performed with the six new E. amylovora phage genomes revealed a striking similarity of 99.8 % between the six phages selected for this study and the bacteriophage species Kolesnikvirus M-M7 (subfamily Ounavirinae).
The similarity of these selected phages to other 28 RefSeq genomes within their respective genera was further evaluated through phylogenetic analysis. The outcomes consistently demonstrated that the newly isolated phages clustered effectively within the Kolesnikvirus genus, specifically within the Kolesnikvirus M7 species-cluster. Furthermore, the analysis successfully highlighted the appropriate clustering of species within the Moglevirus, Suspvirus, and Felixounavirus genera (Figure 7).
Given that the results observed in the analysis of the proteomic tree revealed that the new phages described in this study belong to the Kolesnikvirus genus, a comparative analysis was conducted with all complete genomes of the two species described in this genus (Kolesnikvirus Ea214 (KEa214) and Kolesnikvirus M7 (KM7)) (Figure 8A).
The results confirmed that the new Mediterranean phages of E. amylovora described in this study cluster with isolates from the KM7 species. Interestingly, in this analysis, it can be observed that phages with E. amylovora as a specific host only belong to the clade of KM7 species, while the clade of the KEa214 species groups isolates capable of infecting both E. amylovora and other bacteria such as Salmonella enterica and Hafnia alvei. These results reinforce those obtained in the host range assays (Table 3) and suggest that isolates from the KM7 species may be more specific to E. amylovora than those of the KEa214 species. Therefore, these isolates could potentially be a suitable and safe tool to control E. amylovora without altering the natural microbiome of the host plant and the surrounding environment.
Comparative analyses of complete genomes revealed that the six Mediterranean phages encode up to 117 ORFs, of which 47 have known function, consistent with the vB Eam_MM7 isolate (NC041978). The number of polymorphisms between the Mediterranean phages and the reference genome NC041978 ranged from 398 to 418 nucleotides. These polymorphisms were predominantly located in intergenic regions and in some hypothetical proteins (Figure 8B), where mutations did not involve changes in the polarity and/or hydrophobicity of amino acids which could make changes in the protein structure encode.
Finally, in this analysis, based on complete genomes, another noteworthy result was observed: the new E. amylovora phage isolates from Mediterranean settings show a significant similarity to other E. amylovora phages isolated in Belarus, France, Switzerland and South Korea. This finding suggests that phage species within this clade, in addition to potentially being specific to E. amylovora, exhibit a low degree of genetic divergence, similar to what is observed in the pangenome of E. amylovora. This genetic similarity suggests that the new Mediterranean E. amylovora phages could be of interest in the application of phage therapy for preventive control of E. amylovora in Spain and other countries.
4. Conclusions
This study sets up preliminary steps towards designing bacteriophage cocktails effective against the fire blight disease in the field, these being the first cocktails of European phages of E. amylovora with successful biocontrol activity in plant material, although further work would be still required. Different approaches can be explored in the future to improve the designed cocktails and their activity against all the E. amylovora strains. Increasing the starting phage concentrations, optimizing the application timing, and repeating phage treatments several times during the infection period would potentially reduce the observed differences in the phage mix efficacy against different E. amylovora strains. Another factor to consider in the optimization of designed bacteriophage cocktails is the persistence of phages in the environment. Thus, studies on the survival ability of E. amylovora phages and the factors involved would be needed, as well as development of methods to improve phage application, such as microencapsulation using carrier bacteria, including in the formulation protective agents against environmental challenges [21,80,81].
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Table S1: Origin of a selection of 28 Mediterranean bacteriophages with lytic activity against Erwinia amylovora (Ea).
Author Contributions
Conceptualization, E.G.B.; methodology, E.G.B., R.D.S., F.M.; À.F.-S., R.V., J.F.C.-S. and B.Á.; software, R.D.S., F.M., J.F.C.-S and B.Á.; validation, E.G.B., R.D.S., F.M. and B.Á.; formal analysis, E.G.B., R.D.S., F.M., J.F.C.-S. and B.Á.; investigation, E.G.B., R.D.S., F.M., À.F.-S., R.V., J.F.C.-S. and B.Á.; resources, E.G.B.; writing—original draft preparation, E.G.B., R.D.S., F.M. and B.Á.; writing—review and editing, E.G.B., R.D.S., F.M., À.F.-S., R.V., J.F.C.-S. and B.Á.; supervision, E.G.B.; project administration, E.G.B.; funding acquisition, E.G.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MCIN/AEI/10.13039/501100011033, the Spanish Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), and “ERDF A way of making Europe”, grant number RTA2015-00087-C02.
Data Availability Statement
The genome sequences of the six Mediterranean Erwinia amylovora bacteriophages (vEam_PM_6, vEam_PM_21, vEam_S_24, vEam_W_25, vEam_PM_27 and vEam_W_28) were deposited at GenBank (PP079182, PP079183, PP079184, PP079185, PP079186 and PP079187).
Acknowledgments
Authors would like to thank for the Grant RTA2015-00087-C02 funded by MCIN/AEI/10.13039/501100011033, the Spanish Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), and “ERDF A way of making Europe”. We also thank Central Service for Experimental Research (SCSIE) from the Universitat de València (UV) (Valencia, Spain) service technicians for their qualified support in microscopy, particularly to M.T. Mínguez (SCSIE, UV) for her expert assistance. The authors would also like to thank María M. López and Ana Palacio Bielsa from the Instituto Valenciano de Investigaciones Agrarias (IVIA) (Moncada, Valencia, Spain) and the Centro de Investigación y Tecnología Agroalimentaria de Aragón (CITA), respectively, for IVIA and CITA strains. Rosa Vázquez was the recipient of a Collaboration Grant number 23CO1/010009 from Spanish Ministerio de Educación, Formación Profesional y Deportes. Félix Morán is grateful for the possibility of a short-term postdoctoral stay in the BACPLANT group (reference GIUV2015-219) at the Universitat de València, which has allowed him to participate in the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Winslow, C.-E.A.; Broadhurst, J.; Buchanan, R.E.; Krumwiede, C.; Rogers, L.A.; Smith, G.H. The Families and Genera of the Bacteria: Final Report of the Committee of the Society of American Bacteriologists on Characterization and Classification of Bacterial Types. J Bacteriol 1920, 5, 191–229. [Google Scholar] [CrossRef] [PubMed]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-Based Phylogeny and Taxonomy of the “Enterobacteriales”: Proposal for Enterobacterales Ord. Nov. Divided into the Families Enterobacteriaceae, Erwiniaceae Fam. Nov., Pectobacteriaceae Fam. Nov., Yersiniaceae Fam. Nov., Hafniaceae Fam. Nov., Morganellaceae Fam. Nov., and Budviciaceae Fam. Nov. Int J Syst Evol Microbiol 2016, 66, 5575–5599. [Google Scholar] [CrossRef] [PubMed]
- Momol, M.T.; Aldwinckle, H.S. Genetic Diversity and Host Range of Erwinia amylovora. In Fire blight: the disease and its causative agent, Erwinia amylovora.; CABI Publishing, 2000; pp. 55–72.
- Van Der Zwet, T.; Orolaza-Halbrendt, N.; Zeller, W.; American Phytopathological Society. Fire Blight : History, Biology, and Management. 2012, 421.
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S. V.; Machado, M.A.; et al. Top 10 Plant Pathogenic Bacteria in Molecular Plant Pathology. Mol Plant Pathol 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Gusberti, M.; Klemm, U.; Meier, M.S.; Maurhofer, M.; Hunger-Glaser, I. Fire Blight Control: The Struggle Goes On. A Comparison of Different Fire Blight Control Methods in Switzerland with Respect to Biosafety, Efficacy and Durability. Int J Environ Res Public Health 2015, 12, 11422. [Google Scholar] [CrossRef]
- Aćimović, S.G.; Santander, R.D.; Meredith, C.L.; Pavlović, Ž.M. Fire Blight Rootstock Infections Causing Apple Tree Death: A Case Study in High-Density Apple Orchards with Erwinia amylovora Strain Characterization. Frontiers in Horticulture 2023, 2, 1082204. [Google Scholar] [CrossRef]
- EPPO Erwinia amylovora EPPO Global Database. Available online: https://gd.eppo.int/taxon/ERWIAM (accessed on 29 December 2023).
- Anonymous. Commission Implementing Regulation (EU) 2019/2072 of 28 November 2019 Establishing Uniform Conditions for the Implementation of Regulation (EU) 2016/2031 of the European Parliament and the Council, as Regards Protective Measures against Pests of Plants, and Repealing Commission Regulation (EC) No 690/2008 and Amending Commission Implementing Regulation (EU) 2018/2019. 2019, 1–279.
- Tancos, K.A.; Borejsza-Wysocka, E.; Kuehne, S.; Breth, D.; Cox, K.D. Fire Blight Symptomatic Shoots and the Presence of Erwinia amylovora in Asymptomatic Apple Budwood. Plant Dis 2017, 101, 186–191. [Google Scholar] [CrossRef]
- Förster, H.; McGhee, G.C.; Sundin, G.W.; Adaskaveg, J.E. Characterization of Streptomycin Resistance in Isolates of Erwinia amylovora in California. Phytopathology 2015, 105, 1302–1310. [Google Scholar] [CrossRef]
- Sundin, G.W.; Wang, N. Antibiotic Resistance in Plant-Pathogenic Bacteria. Annu Rev Phytopathol 2018, 56, 161–180. [Google Scholar] [CrossRef]
- Sholberg, P.L.; Bedford, K.E.; Haag, P.; Randall, P. Survey of Erwinia amylovora Isolates from British Columbia for Resistance to Bactericides and Virulence on Apple. Canadian Journal of Plant Pathology 2001, 23, 60–67. [Google Scholar] [CrossRef]
- Al-Daoude, A.; Arabi, M.I.E.; Ammouneh, H. Studying Erwinia amylovora Isolates from Syria for Copper Resistance and Streptomycin Sensitivity. Journal of Plant Pathology 2009, 91, 203–205. [Google Scholar]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Köhl, J.; Jones, J.B.; Aubertot, J.N. Thirteen Decades of Antimicrobial Copper Compounds Applied in Agriculture. A Review. Agronomy for Sustainable Development 2018 38:3 2018, 38, 1–18. [Google Scholar] [CrossRef]
- Taylor, P.; Reeder, R. Antibiotic Use on Crops in Low and Middle-Income Countries Based on Recommendations Made by Agricultural Advisors. CABI Agriculture and Bioscience 2020, 1, 1–14. [Google Scholar] [CrossRef]
- Imran, M.; Das, K.R.; Naik, M.M. Co-Selection of Multi-Antibiotic Resistance in Bacterial Pathogens in Metal and Microplastic Contaminated Environments: An Emerging Health Threat. Chemosphere 2019, 215, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Kering, K.K.; Kibii, B.J.; Wei, H. Biocontrol of Phytobacteria with Bacteriophage Cocktails. Pest Manag Sci 2019, 75, 1775–1781. [Google Scholar] [CrossRef] [PubMed]
- Coelho, F.C.; Squitti, R.; Ventriglia, M.; Cerchiaro, G.; Daher, J.P.; Rocha, J.G.; Rongioletti, M.C.A.; Moonen, A.C. Agricultural Use of Copper and Its Link to Alzheimer’s Disease. Biomolecules 2020, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Herrero, G.; Álvarez, B.; Santander, R.D.; Biosca, E.G. Screening for Novel Beneficial Environmental Bacteria for an Antagonism-Based Erwinia amylovora Biological Control. Microorganisms 2023, Vol. 11, Page 1795 2023, 11, 1795. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.B.; Svircev, A.M.; Obradović, A.Ž. Crop Use of Bacteriophages. Bacteriophages 2018, 1–18. [Google Scholar] [CrossRef]
- Balogh, B.; Jones, J.; Iriarte, F.; Momol, M. Phage Therapy for Plant Disease Control. Curr Pharm Biotechnol 2010, 11, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Doffkay, Z.; Dömötör, D.; Kovács, T.; Rákhely, G. Bacteriophage Therapy against Plant, Animal and Human Pathogens. Acta Biologica Szegediensis 2015, 291–302. [Google Scholar]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and Bacterial Plant Diseases. Front Microbiol 2017, 8, 212667. [Google Scholar] [CrossRef]
- Álvarez, B.; Biosca, E.G. Bacteriophage-Based Bacterial Wilt Biocontrol for an Environmentally Sustainable Agriculture. Front Plant Sci 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Svircev, A.; Roach, D.; Castle, A. Framing the Future with Bacteriophages in Agriculture. Viruses 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Vu, N.T.; Oh, C.S. Bacteriophage Usage for Bacterial Disease Management and Diagnosis in Plants. Plant Pathol J 2020, 36, 204. [Google Scholar] [CrossRef] [PubMed]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The Future of Phage Biocontrol in Integrated Plant Protection for Sustainable Crop Production. Curr Opin Biotechnol 2021, 68, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Fortuna, K.J.; Holtappels, D.; Venneman, J.; Baeyen, S.; Vallino, M.; Verwilt, P.; Rediers, H.; De Coninck, B.; Maes, M.; Van Vaerenbergh, J.; et al. Back to the Roots: Agrobacterium-Specific Phages Show Potential to Disinfect Nutrient Solution from Hydroponic Greenhouses. Appl Environ Microbiol 2023, 89. [Google Scholar] [CrossRef] [PubMed]
- Born, Y.; Fieseler, L.; Marazzi, J.; Lurz, R.; Duffy, B.; Loessner, M.J. Novel Virulent and Broad-Host-Range Erwinia amylovora Bacteriophages Reveal a High Degree of Mosaicism and a Relationship to Enterobacteriaceae Phages. Appl Environ Microbiol 2011, 77, 5945–5954. [Google Scholar] [CrossRef] [PubMed]
- Boulé, J.; Sholberg, P.L.; Lehman, S.M.; O’Gorman, D.T.; Svircev, A.M. Isolation and Characterization of Eight Bacteriophages Infecting Erwinia amylovora and Their Potential as Biological Control Agents in British Columbia, Canada. Canadian Journal of Plant Pathology 2011, 33, 308–317. [Google Scholar] [CrossRef]
- Nagy, J.K.; Király, L.; Schwarczinger, I. Phage Therapy for Plant Disease Control with a Focus on Fire Blight. Cent Eur J Biol 2012, 7, 1–12. [Google Scholar] [CrossRef]
- Park, J.; Kim, B.; Song, S.; Lee, Y.W.; Roh, E. Isolation of Nine Bacteriophages Shown Effective against Erwinia amylovora in Korea. Plant Pathol J 2022, 38, 248–253. [Google Scholar] [CrossRef]
- DuPont, S.T.; Cox, K.; Johnson, K.; Peter, K.; Smith, T.; Munir, M.; Baro, A. Evaluation of Biopesticides for the Control of Erwinia amylovora in Apple and Pear. Journal of Plant Pathology 2023, 1–13. [Google Scholar] [CrossRef]
- Iriarte, F.B.; Balogh, B.; Momol, M.T.; Smith, L.M.; Wilson, M.; Jones, J.B. Factors Affecting Survival of Bacteriophage on Tomato Leaf Surfaces. Appl Environ Microbiol 2007, 73, 1704–1711. [Google Scholar] [CrossRef] [PubMed]
- Gayder, S.; Kammerecker, S.; Fieseler, L. Biological Control of the Fire Blight Pathogen Erwinia amylovora Using Bacteriophages. Journal of Plant Pathology 2023 2023, 1–17. [Google Scholar] [CrossRef]
- Jones, J.B.; Vallad, G.E.; Iriarte, F.B.; Obradović, A.; Wernsing, M.H.; Jackson, L.E.; Balogh, B.; Hong, J.C.; Momol, M.T. Considerations for Using Bacteriophages for Plant Disease Control. Bacteriophage 2012, 2, e23857. [Google Scholar] [CrossRef] [PubMed]
- Gayder, S.; Parcey, M.; Nesbitt, D.; Castle, A.J.; Svircev, A.M. Population Dynamics between Erwinia amylovora, Pantoea agglomerans and Bacteriophages: Exploiting Synergy and Competition to Improve Phage Cocktail Efficacy. Microorganisms 2020, Vol. 8, Page 1449 2020, 8, 1449. [Google Scholar] [CrossRef] [PubMed]
- Bertani, G. Studies on Lysogenesis. I. The Mode of Phage Liberation by Lysogenic Escherichia coli. J Bacteriol 1951, 62, 293–300. [Google Scholar] [CrossRef]
- Gill, J.J.; Svircev, A.M.; Smith, R.; Castle, A.J. Bacteriophages of Erwinia amylovora. Appl Environ Microbiol 2003, 69, 2133–2138. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, C.; Klos, E. New Medium for Detecting Erwinia amylovora and Its Use in Epidemiological Studies. Phytopathology 1984, 74, 1342. [Google Scholar] [CrossRef]
- King, E.O.; Ward, M.K.; Raney, D.E. Two Simple Media for the Demonstration of Pyocyanin and Fluorescin. J Lab Clin Med 1954, 44, 301–307. [Google Scholar] [PubMed]
- Lelliott, R.; Stead, D. Methods for the Diagnosis of Bacterial Diseases of Plants. 1987.
- EPPO PM 7/20 (3) Erwinia amylovora. EPPO Bulletin 2022, 52, 198–224. [CrossRef]
- Santander, R.D.; Biosca, E.G. Erwinia amylovora Psychrotrophic Adaptations: Evidence of Pathogenic Potential and Survival at Temperate and Low Environmental Temperatures. PeerJ 2017, 2017. [Google Scholar] [CrossRef]
- Biosca, E.G.; Català-Senent, J.F.; Figàs-Segura, À.; Bertolini, E.; López, M.M.; Álvarez, B. Genomic Analysis of the First European Bacteriophages with Depolymerase Activity and Biocontrol Efficacy against the Phytopathogen Ralstonia solanacearum. Viruses 2021, 13, 2539. [Google Scholar] [CrossRef] [PubMed]
- Rasband, W. ImageJ 1.53m 2022. Available at: https://ij.imjoy.io/ [Accessed December 29, 2023].
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs. Nucleic Acids Res 1997, 25. [Google Scholar] [CrossRef] [PubMed]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A Modular and Extensible Implementation of the RAST Algorithm for Building Custom Annotation Pipelines and Annotating Batches of Genomes. 2015. [CrossRef]
- Tynecki, P.; Guziński, A.; Kazimierczak, J.; Jadczuk, M.; Dastych, J.; Onisko, A. PhageAI - Bacteriophage Life Cycle Recognition with Machine Learning and Natural Language Processing. bioRxiv 2020, 2020.07.11.198606. [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol Biol Evol 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The Viral Proteomic Tree Server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef] [PubMed]
- Besarab, N. V.; Akhremchuk, A.E.; Zlatohurska, M.A.; Romaniuk, L. V.; Valentovich, L.N.; Tovkach, F.I.; Lagonenko, A.L.; Evtushenkov, A.N. Isolation and Characterization of Hena1 – a Novel Erwinia amylovora Bacteriophage. FEMS Microbiol Lett 2020, 367, 70. [Google Scholar] [CrossRef] [PubMed]
- Biosca, E.G.; Santander, R.D.; Ordax, M.; Marco-Noales, E.; López, M.M. Erwinia amylovora Survives in Natural Water. Acta Hortic 2008, 793, 83–88. [Google Scholar] [CrossRef]
- Santander, R.D.; Català-Senent, J.F.; Marco-Noales, E.; Biosca, E.G. In Planta Recovery of Erwinia amylovora Viable but Nonculturable Cells. Trees - Structure and Function 2012, 26, 75–82. [Google Scholar] [CrossRef]
- Santander, R.D.; Oliver, J.D.; Biosca, E.G. Cellular, Physiological, and Molecular Adaptive Responses of Erwinia amylovora to Starvation. FEMS Microbiol Ecol 2014, 88, 258–271. [Google Scholar] [CrossRef]
- Bertolini, E.; Figàs-Segura, À.; Álvarez, B.; Biosca, E.G. Development of TaqMan Real-Time PCR Protocols for Simultaneous Detection and Quantification of the Bacterial Pathogen Ralstonia solanacearum and Their Specific Lytic Bacteriophages. Viruses 2023, Vol. 15, Page 841 2023, 15, 841. [Google Scholar] [CrossRef]
- Álvarez, B.; López, M.M.; Biosca, E.G. Influence of Native Microbiota on Survival of Ralstonia solanacearum Phylotype II in River Water Microcosms. Appl Environ Microbiol 2007, 73, 7210–7217. [Google Scholar] [CrossRef] [PubMed]
- Alič, Š.; Naglič, T.; Tušek-Žnidarič, M.; Ravnikar, M.; Rački, N.; Peterka, M.; Dreo, T. Newly Isolated Bacteriophages from the Podoviridae, Siphoviridae, and Myoviridae Families Have Variable Effects on Putative Novel Dickeyas spp. Front Microbiol 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Sabri, M.; El Handi, K.; Valentini, F.; De Stradis, A.; Achbani, E.H.; Benkirane, R.; Resch, G.; Elbeaino, T. Identification and Characterization of Erwinia Phage IT22: A New Bacteriophage-Based Biocontrol against Erwinia amylovora. Viruses 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Cui, Z.; Wang, J.; Childs, K.L.; Sundin, G.W.; Cooley, D.R.; Yang, C.H.; Garofalo, E.; Eaton, A.; Huntley, R.B.; et al. Comparative Genomics of Spiraeoideae-Infecting Erwinia amylovora Strains Provides Novel Insight to Genetic Diversity and Identifies the Genetic Basis of a Low-Virulence Strain. Mol Plant Pathol 2018, 19, 1652–1666. [Google Scholar] [CrossRef] [PubMed]
- Gayder, S.; Parcey, M.; Castle, A.J.; Svircev, A.M. Host Range of Bacteriophages Against a World-Wide Collection of Erwinia amylovora Determined Using a Quantitative PCR Assay. Viruses 2019, Vol. 11, Page 910 2019, 11, 910. [Google Scholar] [CrossRef]
- Mirzaei, M.K.; Nilsson, A.S. Isolation of Phages for Phage Therapy: A Comparison of Spot Tests and Efficiency of Plating Analyses for Determination of Host Range and Efficacy. PLoS One 2015, 10, e0118557. [Google Scholar] [CrossRef]
- Ross, A.; Ward, S.; Hyman, P. More Is Better: Selecting for Broad Host Range Bacteriophages. Front Microbiol 2016, 7. [Google Scholar] [CrossRef]
- Pereira, C.; Moreirinha, C.; Lewicka, M.; Almeida, P.; Clemente, C.; Romalde, J.L.; Nunes, M.L.; Almeida, A. Characterization and in vitro Evaluation of New Bacteriophages for the Biocontrol of Escherichia coli. Virus Res 2017, 227, 171–182. [Google Scholar] [CrossRef]
- Wójcicki, M.; Świder, O.; Średnicka, P.; Shymialevich, D.; Ilczuk, T.; Koperski, Ł.; Cieślak, H.; Sokołowska, B.; Juszczuk-Kubiak, E. Newly Isolated Virulent Salmophages for Biocontrol of Multidrug-Resistant Salmonella in Ready-to-Eat Plant-Based Food. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Luria, S.E.; Delbrück, M. Mutations of Bacteria from Virus Sensitivity to Virus Resistance. Genetics 1943, 28, 491–511. [Google Scholar] [CrossRef]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, B.; López, M.M.; Biosca, E.G. Biocontrol of the Major Plant Pathogen Ralstonia solanacearum in Irrigation Water and Host Plants by Novel Waterborne Lytic Bacteriophages. Front Microbiol 2019, 10, 492073. [Google Scholar] [CrossRef] [PubMed]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage Resistance Mechanisms. Nat Rev Microbiol 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Schmerer, M.; Molineux, I.J.; Bull, J.J. Synergy as a Rationale for Phage Therapy Using Phage Cocktails. PeerJ 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Naknaen, A.; Samernate, T.; Wannasrichan, W.; Surachat, K.; Nonejuie, P.; Chaikeeratisak, V. Combination of Genetically Diverse Pseudomonas Phages Enhances the Cocktail Efficiency against Bacteria. Sci Rep 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Molina-Quiroz, R.C.; Camilli, A.; Silva-Valenzuela, C.A. Role of Bacteriophages in the Evolution of Pathogenic Vibrios and Lessons for Phage Therapy. Adv Exp Med Biol 2023, 1404, 149–173. [Google Scholar] [CrossRef] [PubMed]
- Mapes, A.C.; Trautner, B.W.; Liao, K.S.; Ramig, R.F. Development of Expanded Host Range Phage Active on Biofilms of Multi-Drug Resistant Pseudomonas aeruginosa. Bacteriophage 2016, 6, e1096995. [Google Scholar] [CrossRef]
- Castledine, M.; Padfield, D.; Sierocinski, P.; Pascual, J.S.; Hughes, A.; Mäkinen, L.; Friman, V.P.; Pirnay, J.P.; Merabishvili, M.; De Vos, D.; et al. Parallel Evolution of Pseudomonas aeruginosa Phage Resistance and Virulence Loss in Response to Phage Treatment in vivo and in vitro. Elife 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- García, P.; Tabla, R.; Anany, H.; Bastias, R.; Brøndsted, L.; Casado, S.; Cifuentes, P.; Deaton, J.; Denes, T.G.; Islam, M.A.; et al. ECOPHAGE: Combating Antimicrobial Resistance Using Bacteriophages for Eco-Sustainable Agriculture and Food Systems. Viruses 2023, Vol. 15, Page 2224 2023, 15, 2224. [Google Scholar] [CrossRef]
- Tolstoy, I.; Kropinski, A.M.; Brister, J.R. Bacteriophage Taxonomy: An Evolving Discipline. Methods in Molecular Biology 2018, 1693, 57–71. [Google Scholar]
- Turner, D.; Shkoporov, A.N.; Lood, C.; Millard, A.D.; Dutilh, B.E.; Alfenas-Zerbini, P.; van Zyl, L.J.; Aziz, R.K.; Oksanen, H.M.; Poranen, M.M.; et al. Abolishment of Morphology-Based Taxa and Change to Binomial Species Names: 2022 Taxonomy Update of the ICTV Bacterial Viruses Subcommittee. Arch Virol 2023, 168, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Frampton, R.A.; Pitman, A.R.; Fineran, P.C. Advances in Bacteriophage-Mediated Control of Plant Pathogens. Int J Microbiol 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Ly-Chatain, M.H. The Factors Affecting Effectiveness of Treatment in Phages Therapy. Front Microbiol 2014, 5. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Erwinia amylovora-phage interactions in liquid NBSYE medium. Graphs show the growth of Spanish E. amylovora strains (A) IVIA 1892.1, (B) IVIA 1526.6, and (C) IVIA 1614.2 alone (squares) and in co-culture with Mediterranean E. amylovora phages ϕ1 to ϕ28 (circles), assayed individually. Data points are average values of triplicate experiments, and error bars show the standard deviation (SD). The P values in each chart show differences between the areas under the curve (AUC) of the controls (E. amylovora grown alone) and treatments (E. amylovora plus individual phages) based on a Brown-Forsythe ANOVA test.
Figure 1.
Erwinia amylovora-phage interactions in liquid NBSYE medium. Graphs show the growth of Spanish E. amylovora strains (A) IVIA 1892.1, (B) IVIA 1526.6, and (C) IVIA 1614.2 alone (squares) and in co-culture with Mediterranean E. amylovora phages ϕ1 to ϕ28 (circles), assayed individually. Data points are average values of triplicate experiments, and error bars show the standard deviation (SD). The P values in each chart show differences between the areas under the curve (AUC) of the controls (E. amylovora grown alone) and treatments (E. amylovora plus individual phages) based on a Brown-Forsythe ANOVA test.
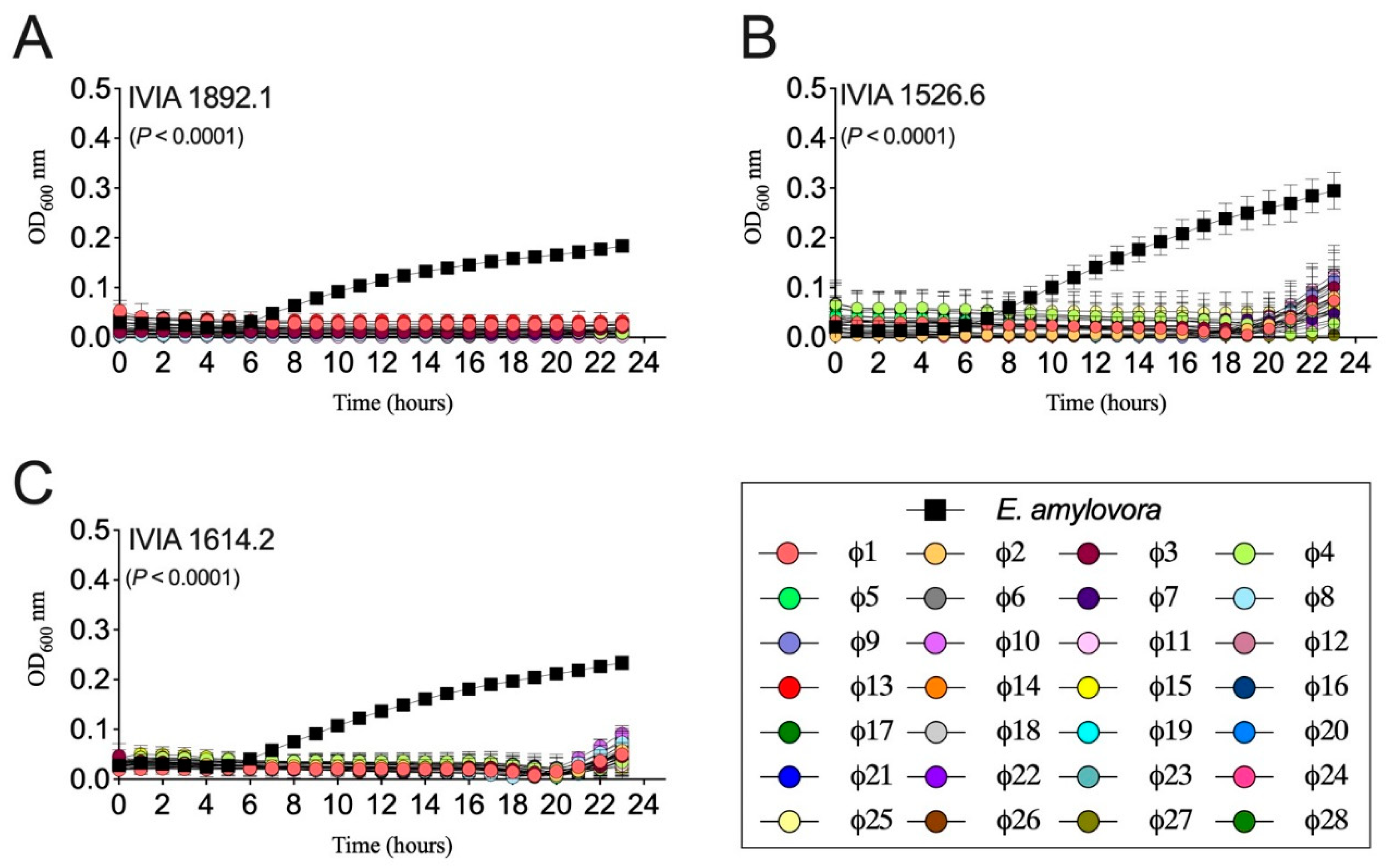
Figure 2.
Erwinia amylovora-phage interactions in liquid NBSYE medium. Graphs show the growth of Spanish E. amylovora strains (A) IVIA 1892.1, (B) IVIA 1526.6, and (C) IVIA 1614.2 alone (black symbols) and with the Mediterranean E. amylovora phage mixes (colored symbols). Data points are average values of sextuplicate assays, and error bars show the standard deviation (SD). The P values in each chart show differences between the areas under the curve (AUC) of the controls (E. amylovora grown alone) and treatments (E. amylovora plus phage mixes) based on a Brown-Forsythe ANOVA test.
Figure 2.
Erwinia amylovora-phage interactions in liquid NBSYE medium. Graphs show the growth of Spanish E. amylovora strains (A) IVIA 1892.1, (B) IVIA 1526.6, and (C) IVIA 1614.2 alone (black symbols) and with the Mediterranean E. amylovora phage mixes (colored symbols). Data points are average values of sextuplicate assays, and error bars show the standard deviation (SD). The P values in each chart show differences between the areas under the curve (AUC) of the controls (E. amylovora grown alone) and treatments (E. amylovora plus phage mixes) based on a Brown-Forsythe ANOVA test.
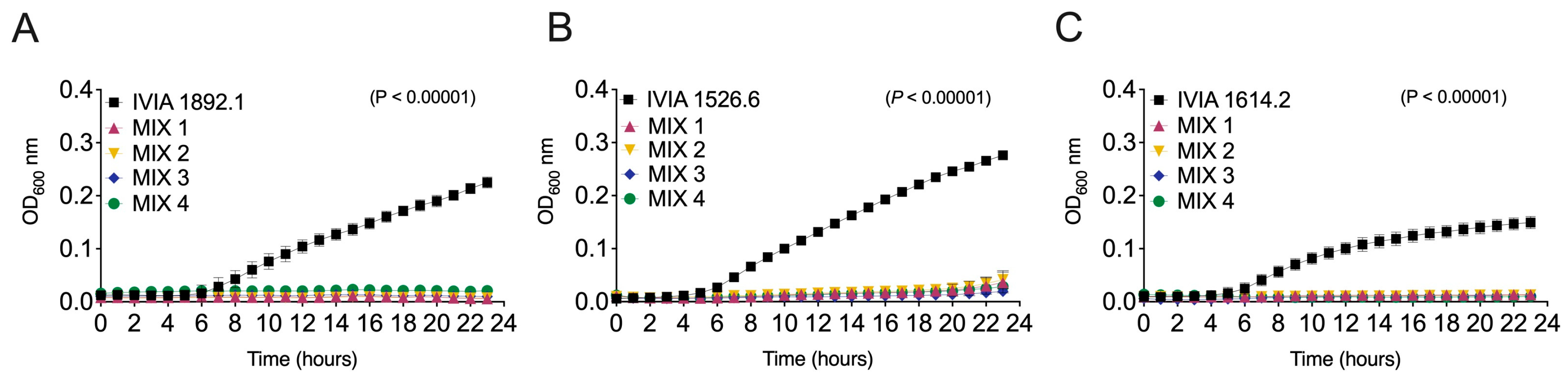
Figure 3.
Biocontrol activity of a selection of six individual Mediterranean Erwinia amylovora phages against E. amylovora strains IVIA 1892.1 (left; A, C and E) and IVIA 1526.6 (right; B, E and F) in detached, green loquat fruit. In preventive treatments, the selected phages were applied individually 24 h before E. amylovora inoculation. In co-inoculation assays, both E. amylovora and individual phages were applied simultaneously. (A and B) Disease Control Efficacy (%), (C and D) Disease Severity (%), and (E and F) representative images of inoculated fruit after a 6-day incubation period at 28°C. Columns represent average values of triplicate assays, and error bars depict the standard deviation (SD). Asterisks show significant differences among the fruits inoculated with E. amylovora alone, those treated with the pathogen plus each phage, and the negative controls (NC), assessed by Dunnett’s multiple comparison tests (α = 0.05); ns stands for not significant.
Figure 3.
Biocontrol activity of a selection of six individual Mediterranean Erwinia amylovora phages against E. amylovora strains IVIA 1892.1 (left; A, C and E) and IVIA 1526.6 (right; B, E and F) in detached, green loquat fruit. In preventive treatments, the selected phages were applied individually 24 h before E. amylovora inoculation. In co-inoculation assays, both E. amylovora and individual phages were applied simultaneously. (A and B) Disease Control Efficacy (%), (C and D) Disease Severity (%), and (E and F) representative images of inoculated fruit after a 6-day incubation period at 28°C. Columns represent average values of triplicate assays, and error bars depict the standard deviation (SD). Asterisks show significant differences among the fruits inoculated with E. amylovora alone, those treated with the pathogen plus each phage, and the negative controls (NC), assessed by Dunnett’s multiple comparison tests (α = 0.05); ns stands for not significant.
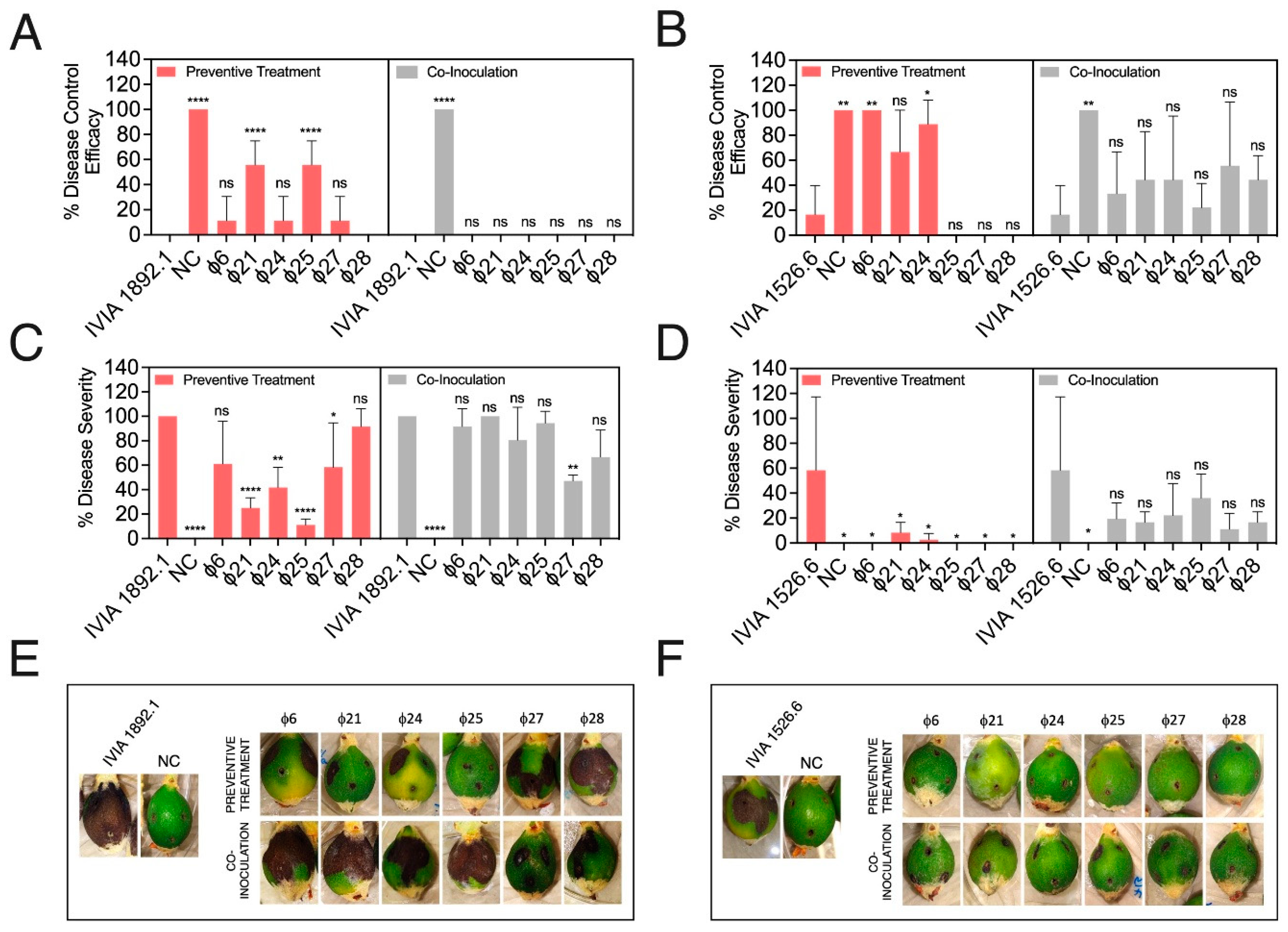
Figure 4.
Biocontrol activity of a selection of four Mediterranean Erwinia amylovora phage cocktails (MIX 1-4) against E. amylovora IVIA 1892.1 in detached, green loquat fruit. In preventive treatments, the phage mixes were applied 24 h before E. amylovora inoculation. In co-inoculation assays, both E. amylovora and the phage mix were applied simultaneously. (A and B) Disease Control Efficacy (%) and Disease Severity (%), respectively. Columns represent average values of triplicate assays, and error bars depict the standard deviation (SD). Asterisks show significant differences among the fruits inoculated with E. amylovora alone, those treated with the pathogen plus the indicated phage mix, and the negative controls (NC), assessed by Dunnett’s multiple comparison tests (α = 0.05); ns stands for not significant. (C) Representative images of inoculated fruits at the end of the assays (6 days at 28 °C).
Figure 4.
Biocontrol activity of a selection of four Mediterranean Erwinia amylovora phage cocktails (MIX 1-4) against E. amylovora IVIA 1892.1 in detached, green loquat fruit. In preventive treatments, the phage mixes were applied 24 h before E. amylovora inoculation. In co-inoculation assays, both E. amylovora and the phage mix were applied simultaneously. (A and B) Disease Control Efficacy (%) and Disease Severity (%), respectively. Columns represent average values of triplicate assays, and error bars depict the standard deviation (SD). Asterisks show significant differences among the fruits inoculated with E. amylovora alone, those treated with the pathogen plus the indicated phage mix, and the negative controls (NC), assessed by Dunnett’s multiple comparison tests (α = 0.05); ns stands for not significant. (C) Representative images of inoculated fruits at the end of the assays (6 days at 28 °C).
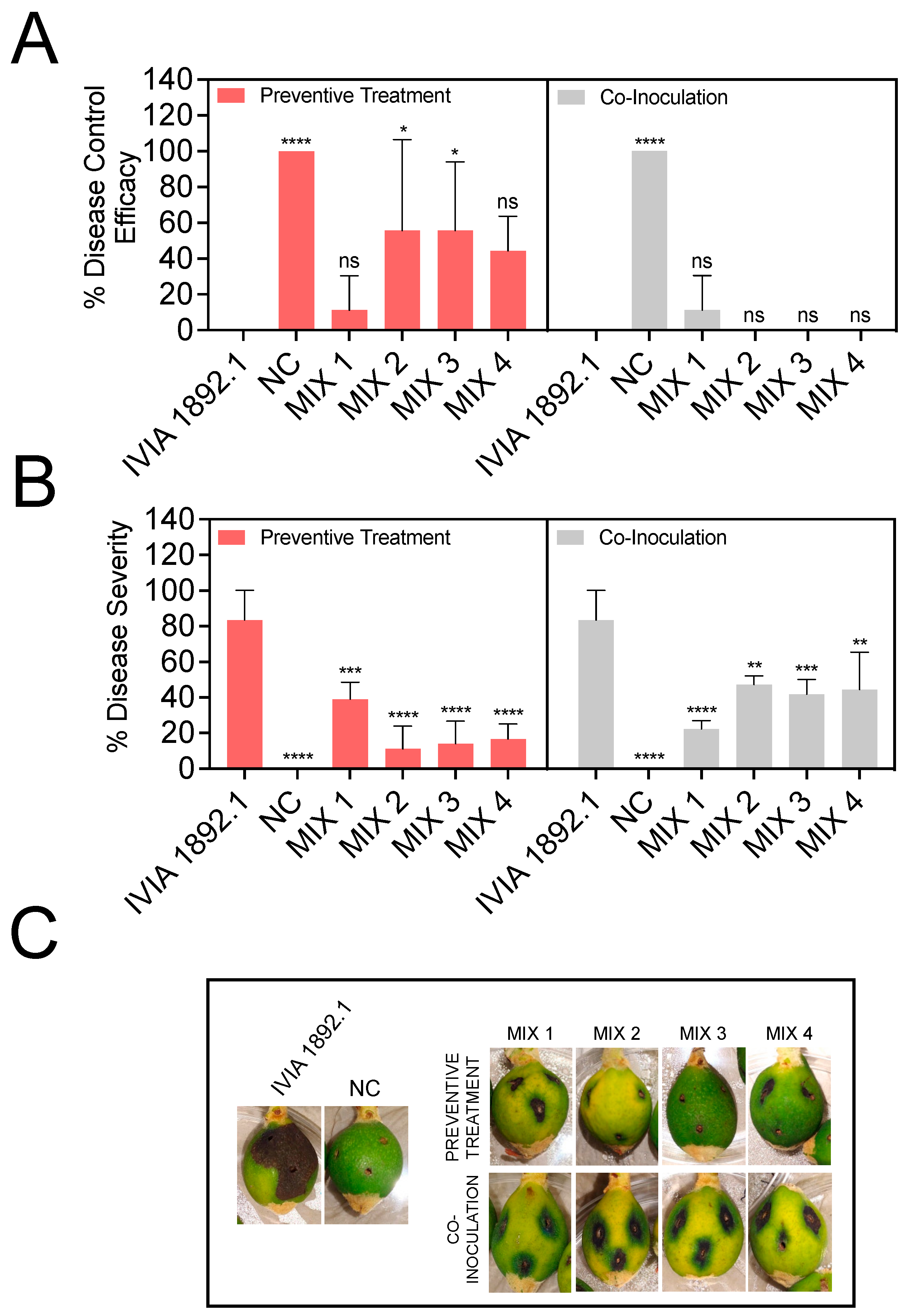
Figure 5.
Performance of four Mediterranean Erwinia amylovora phage cocktails (MIX 1-4) against the French E. amylovora reference strain CFBP 1430. (A) Effect of the phage mixes in E. amylovora growth in liquid medium. Each data point shows the average of six replicates, and the error bars denote the standard deviation (SD). The P value indicates the significance of differences between the AUCs obtained by E. amylovora grown alone (black symbols) and in co-culture with the phage mixes (colored symbols), assessed by a one-way Brown-Forsythe ANOVA test. (B-D) Biocontrol activity of the phage mix in detached, green loquat fruits after 6 days at 28 °C. In preventive treatments, the phage mixes were applied 24 h before E. amylovora inoculation. In co-inoculation assays, both E. amylovora and phage mix were applied simultaneously. (B and C) Disease Control Efficacy (%) and Disease Severity (%), respectively. Columns show average results of an experiment performed in triplicate, and the error bars are the SD. Asterisks indicate statistically significant differences among the fruits inoculated with E. amylovora alone, those treated with the pathogen plus the indicated phage mix, and the negative controls (NC), assessed by Dunnett’s post-hoc tests (α = 0.05); ns stands for not significant. (D) Representative images of inoculated fruits at the end of the assayed period.
Figure 5.
Performance of four Mediterranean Erwinia amylovora phage cocktails (MIX 1-4) against the French E. amylovora reference strain CFBP 1430. (A) Effect of the phage mixes in E. amylovora growth in liquid medium. Each data point shows the average of six replicates, and the error bars denote the standard deviation (SD). The P value indicates the significance of differences between the AUCs obtained by E. amylovora grown alone (black symbols) and in co-culture with the phage mixes (colored symbols), assessed by a one-way Brown-Forsythe ANOVA test. (B-D) Biocontrol activity of the phage mix in detached, green loquat fruits after 6 days at 28 °C. In preventive treatments, the phage mixes were applied 24 h before E. amylovora inoculation. In co-inoculation assays, both E. amylovora and phage mix were applied simultaneously. (B and C) Disease Control Efficacy (%) and Disease Severity (%), respectively. Columns show average results of an experiment performed in triplicate, and the error bars are the SD. Asterisks indicate statistically significant differences among the fruits inoculated with E. amylovora alone, those treated with the pathogen plus the indicated phage mix, and the negative controls (NC), assessed by Dunnett’s post-hoc tests (α = 0.05); ns stands for not significant. (D) Representative images of inoculated fruits at the end of the assayed period.
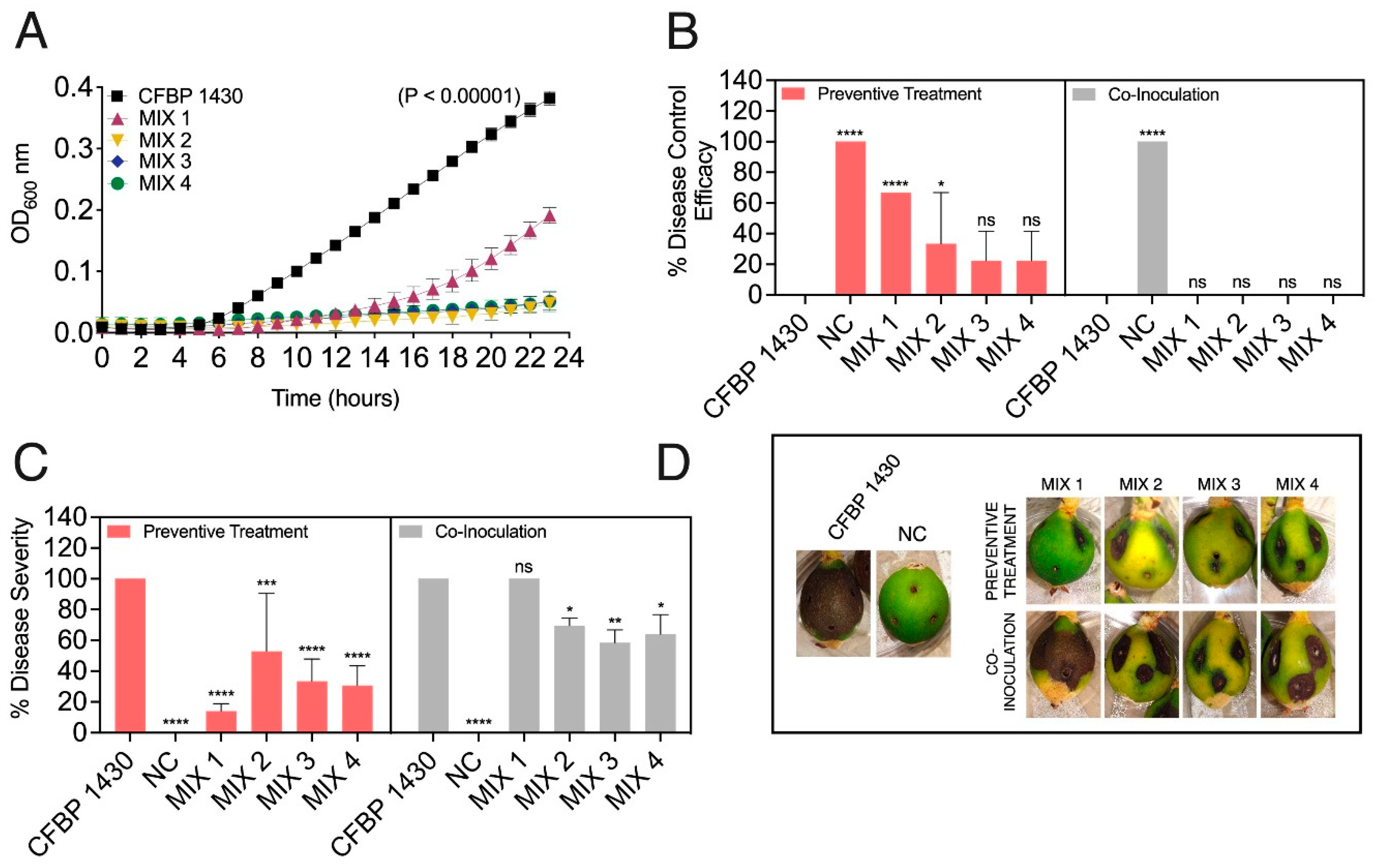
Figure 6.
Transmission electron micrographs displaying the morphology of negatively stained Mediterranean Erwinia amylovora bacteriophages. (A) Phage ϕ6 magnified x33,000; (B) Phage ϕ21 magnified x50,000; (C) Phage ϕ24 magnified x45,000; (D) Phage ϕ25 magnified x33,000; (E) Phage ϕ27 magnified x33,000; and (F) Phage ϕ28 magnified x22,000. Scale bars represent 100 nm in (A, B, C and E), and 200 nm in (D and F).
Figure 6.
Transmission electron micrographs displaying the morphology of negatively stained Mediterranean Erwinia amylovora bacteriophages. (A) Phage ϕ6 magnified x33,000; (B) Phage ϕ21 magnified x50,000; (C) Phage ϕ24 magnified x45,000; (D) Phage ϕ25 magnified x33,000; (E) Phage ϕ27 magnified x33,000; and (F) Phage ϕ28 magnified x22,000. Scale bars represent 100 nm in (A, B, C and E), and 200 nm in (D and F).
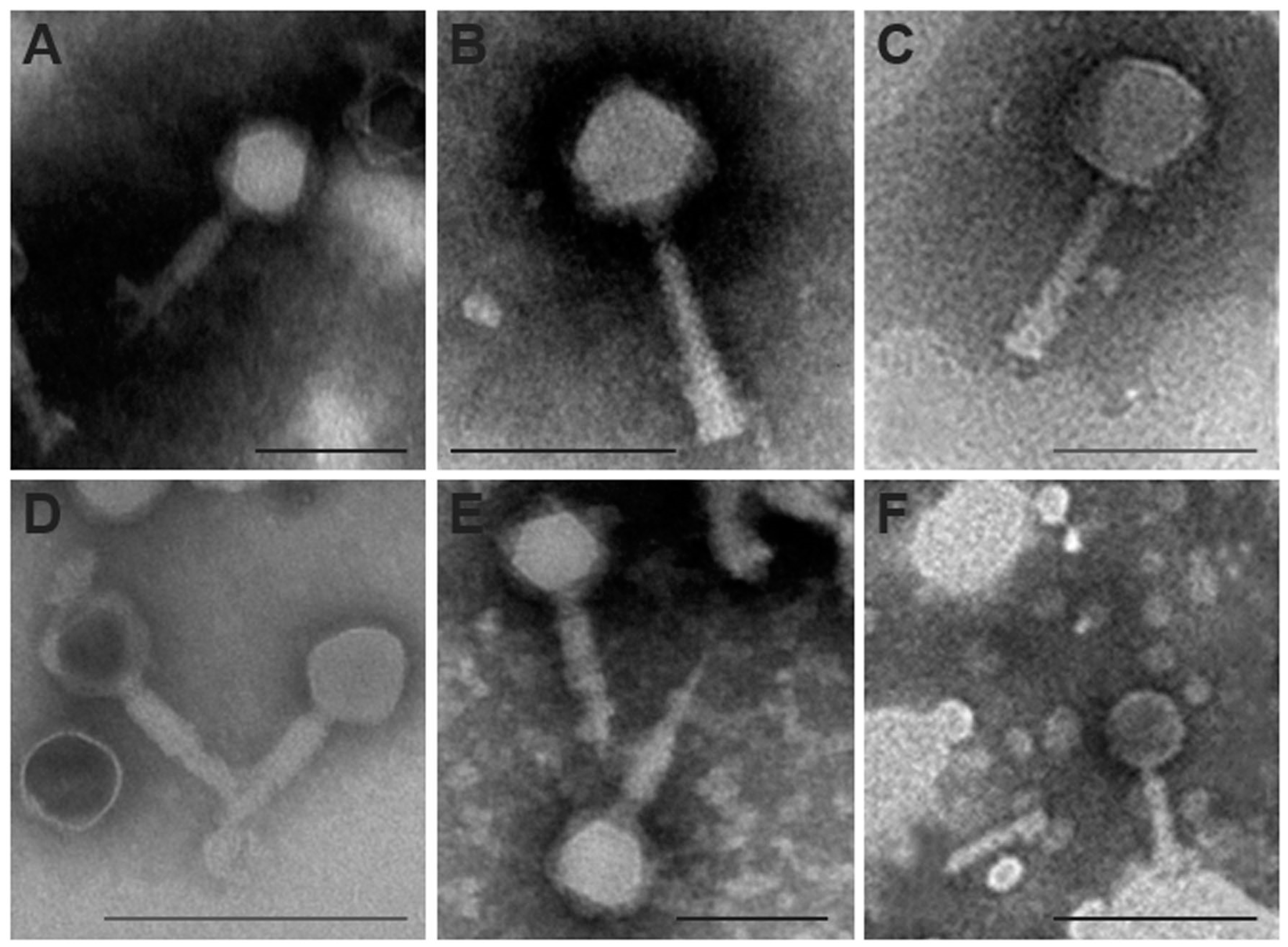
Figure 7.
Phylogenetic proteomic tree of Mediterranean Erwinia amylovora bacteriophages (vEam_PM_6, vEam_PM_21, vEam_S_24, vEam_W_25, vEam_PM_27 and vEam_W_28) with a selection of closely related phages from other sources and continents, using the ViPTree server. The phage genus is indicated by the following colors: Koleniskvirus (purple), Suspvirus (blue), Mooglevirus (black) and Felixounavirus (green).
Figure 7.
Phylogenetic proteomic tree of Mediterranean Erwinia amylovora bacteriophages (vEam_PM_6, vEam_PM_21, vEam_S_24, vEam_W_25, vEam_PM_27 and vEam_W_28) with a selection of closely related phages from other sources and continents, using the ViPTree server. The phage genus is indicated by the following colors: Koleniskvirus (purple), Suspvirus (blue), Mooglevirus (black) and Felixounavirus (green).
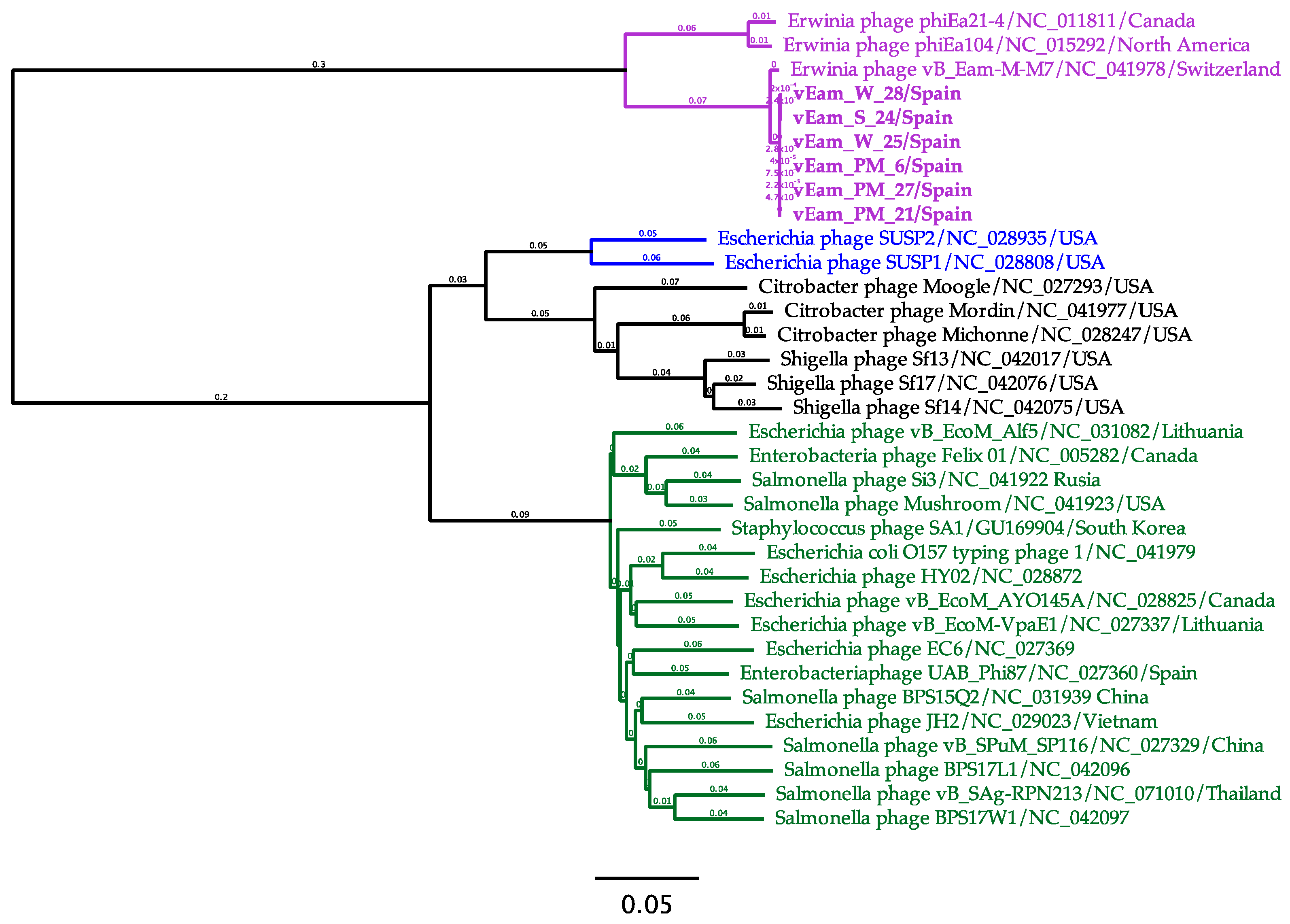
Figure 8.
Comparative analysis carried out with the complete genomes of the six Mediterranean Erwinia amylovora phages and the two species within the genus Kolesnikvirus. (A) Maximum likelihood phylogenetic tree, including Mediterranean E. amylovora phages, constructed by MEGA X using the best substitution model, General Time Reversible with G (0,20) + I (43,8 % sites) parameter, with 32 complete phage genomes from Kolenskivirus species: Kolesnikvirus M7 (Clade A) and Kolesnikvirus Ea214 (Clade B). Phage isolate name, accession numbers, host and origin are indicated. The scale bar shows the number of substitutions per site. Bootstrap percentages (500 replicates) are indicated on branches. (B) Genome map of the Mediterranean E. amylovora phage vEam_PM_21 compared by BLASTn (features by percent identity are indicated) with genome species Kolesnikvirus Ea214 (vB EamM EAP4 (OR185440)) and Kolesnikvirus M7 (vB Eam_MM7 (NC041978)). Open Reading Frames (ORFs) are indicated in grey color and GC Skew score in green and purple colors.
Figure 8.
Comparative analysis carried out with the complete genomes of the six Mediterranean Erwinia amylovora phages and the two species within the genus Kolesnikvirus. (A) Maximum likelihood phylogenetic tree, including Mediterranean E. amylovora phages, constructed by MEGA X using the best substitution model, General Time Reversible with G (0,20) + I (43,8 % sites) parameter, with 32 complete phage genomes from Kolenskivirus species: Kolesnikvirus M7 (Clade A) and Kolesnikvirus Ea214 (Clade B). Phage isolate name, accession numbers, host and origin are indicated. The scale bar shows the number of substitutions per site. Bootstrap percentages (500 replicates) are indicated on branches. (B) Genome map of the Mediterranean E. amylovora phage vEam_PM_21 compared by BLASTn (features by percent identity are indicated) with genome species Kolesnikvirus Ea214 (vB EamM EAP4 (OR185440)) and Kolesnikvirus M7 (vB Eam_MM7 (NC041978)). Open Reading Frames (ORFs) are indicated in grey color and GC Skew score in green and purple colors.


Table 1.
Erwinia amylovora strains used.
| E. amylovora strain code | Host | Geographical origin | Year |
|---|---|---|---|
| Spanish | |||
| UPN1 527 | Malus x domestica | Navarra | 1997 |
| IVIA2 1526.6 | Cotoneaster sp. | Guipúzcoa | 1996 |
| IVIA 1554 | Crataegus sp. | Segovia | 1996 |
| IVIA 1596 | Pyrus sp. | Spain | 1996 |
| IVIA 1614.1 | Pyracantha sp. | Spain | 1996 |
| IVIA 1614.2 | Cotoneaster sp. | Segovia | 1996 |
| IVIA 1626.6 | M. domestica | Navarra | 1996 |
| IVIA 1892.1 | Pyrus sp. | Guadalajara | 1998 |
| UV3 P3P2AA1 | Pyrus communis | Valencia* | 2018 |
| UV P4P2AA1 | P. communis | Valencia | 2018 |
| UV P2P4TA1 | P. communis | Valencia | 2018 |
| UV P2P4TA2.1 | P. communis | Valencia | 2018 |
| UV P2EP4Tex | P. communis | Valencia | 2018 |
| UV P3P4TA1 | P. communis | Valencia | 2018 |
| UV P3P4TA2 | P. communis | Valencia | 2018 |
| International | |||
| CFBP4 1430 | Crataegus oxyacantha | France | 1972 |
| NCPPB 5 311 | P. communis | Canada | 1952 |
| Ea273 | Malus sp. | USA | 1971 |
| CGJ2 | Malus sp. | Serbia | 2003 |
1UPN: Universidad Pública de Navarra; 2IVIA: Instituto Valenciano de Investigaciones Agrarias; 3UV: Universitat de València; 4CFBP: Collection Française de Bactèries Phytopathogènes; 5NCPPB: National Collection of Plant Pathogenic Bacteria; * Former fire-blight affected zones in the province of Valencia.
Table 2.
Bacterial species, including plant pathogens, used in phage specificity assays.
| Bacterial species | Strain code | Host | Geographical origin |
|---|---|---|---|
| Clavibacter michiganensis | IVIA1 873 | Solanum lycopersicum | Spain |
| Dickeya sp. | IVIA 4830 | S. lycopersicum | Spain |
| Pectobacterium atrosepticum | IVIA 3447 | S. tuberosum | Spain |
| P. carotovorum | IVIA 3902 | S. lycopersicum | Spain |
| Pseudomonas savastanoi pv. savastanoi | IVIA 1628.3 | Olea europaea | Spain |
| Rhizobium radiobacter | C58 | Prunus avium | USA |
| R. rhizogenes | K84 | Non-pathogenic | Australia |
| Ralstonia solanacearum | IVIA 1670 | S. tuberosum | Spain |
| Xanthomonas arboricola pv. pruni | CITA2 33 | Prunus amygdalus | Spain |
| X. vesicatoria | CECT3 792 | Unknown | Israel |
| Aeromonas hydrophila | CECT 5173 | Freshwater | France |
| Alcaligenes faecalis | CECT 928 | Unknown | Unknown |
| Bacillus cereus | CECT 495 | Chicken and turkey manure | Unknown |
| Enterococcus faecalis | CECT 481 | Unknown | Unknown |
| Escherichia coli | CECT 101 | Unknown | United Kingdom |
| Klebsiella pneumonia | CECT 143 | Unknown | United States |
| Kocuria rhizophila | CECT 241 | Soil | Unknown |
| Pseudomonas fluorescens | CECT 378 | Pre-filter tanks, town water works | United Kingdom |
| Proteus hauseri | CECT 484 | Unknown | Unknown |
| Salmonella enterica subsp. enterica | CECT 443 | Human food poisoning | United Kingdom |
| Serratia marcescens | CECT 159 | Unknown | Unknown |
| Staphylococcus aureus | CECT 4013 | Bovine mammary gland | Unknown |
1IVIA: Instituto Valenciano de Investigaciones Agrarias; 2CITA: Centro de Investigación y Tecnología Agroalimentaria de Aragón; 3CECT: Colección Española de Cultivos Tipo (Spanish Type Culture Collection).
Table 3.
Host range of a selection of 28 Mediterranean bacteriophages against a collection of 19 Erwinia amylovora strains and 22 strains of other bacterial species, assessed by the spot test.
Table 3.
Host range of a selection of 28 Mediterranean bacteriophages against a collection of 19 Erwinia amylovora strains and 22 strains of other bacterial species, assessed by the spot test.
| Host strain code | UV E. amylovora bacteriophages | |||||||||||||||||||||||||||
| ϕ1 | ϕ2 | ϕ3 | ϕ4 | ϕ5 | ϕ6 | ϕ7 | ϕ8 | ϕ9 | ϕ10 | ϕ11 | ϕ12 | ϕ13 | ϕ14 | ϕ15 | ϕ16 | ϕ17 | ϕ18 | ϕ19 | ϕ20 | ϕ21 | ϕ22 | ϕ23 | ϕ24 | ϕ25 | ϕ26 | ϕ27 | ϕ28 | |
| E. amylovora UPN 527 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| E. amylovora IVIA 1526.6 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| E. amylovora IVIA 1554 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| E. amylovora IVIA 1596 | - | - | + | - | - | + | + | + | W | - | - | W | W | - | - | - | W | W | - | W | + | W | - | -/+ | W | + | -/+ | -/+ |
| E. amylovora IVIA 1614.1 | + | + | W | + | + | + | + | + | + | - | + | + | + | + | + | - | W | + | + | + | + | + | + | + | + | W | + | + |
| E. amylovora IVIA 1614.2 | + | + | + | - | + | + | + | + | + | + | + | + | W | + | + | + | + | + | + | + | + | + | + | + | + | W | -/+ | + |
| E. amylovora IVIA 1626.6 | W | W | - | W | W | W | - | - | - | W | - | - | W | - | - | W | W | - | - | - | -/+ | - | - | -/+ | -/+ | W | W | -/+ |
| E. amylovora IVIA 1892.1 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | - | + | + | + | - | - | + | - | - | -/+ | + | + | -/+ | -/+ |
| E. amylovora UV P3P2AA1 | W | - | W | W | - | + | + | + | + | - | - | - | - | W | - | W | W | W | W | W | + | - | - | W | + | + | W | W |
| E. amylovora UV P4P2AA1 | + | - | + | + | + | + | + | + | + | + | - | - | + | + | - | + | + | - | + | + | + | - | + | + | + | + | + | + |
| E. amylovora UV P2P4TA1 | + | - | + | - | + | + | + | + | + | + | - | - | + | + | - | + | + | - | + | - | + | + | + | -/+ | + | + | -/+ | -/+ |
| E. amylovora UV P2P4TA2.1 | + | + | W | + | + | W | W | W | W | W | - | - | + | + | W | - | - | - | - | - | W | W | - | -/+ | + | W | + | -/+ |
| E. amylovora UV P2exP4T | + | + | + | + | + | + | + | + | + | + | - | - | + | + | - | + | + | - | W | W | + | - | W | W | + | + | + | W |
| E. amylovora UV P3P4TA1 | + | + | - | + | + | W | W | W | - | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | W | + | + |
| E. amylovora UV P3P4TA2 | + | + | W | + | + | W | W | W | W | + | + | + | + | W | + | + | + | + | + | + | W | + | + | + | W | + | + | + |
| E. amylovora CFBP 1430 | - | - | - | W | - | W | - | W | - | W | + | + | - | + | + | W | W | + | + | + | -/+ | + | + | + | -/+ | - | -/+ | W |
| E. amylovora NCPPB 311 | W | W | + | W | W | + | + | + | + | + | + | + | + | + | + | + | W | + | W | W | + | W | W | W | + | + | W | W |
| E. amylovora GJ-2 | + | + | + | + | + | + | + | + | W | W | W | W | + | + | W | W | W | W | W | W | + | W | W | W | + | W | + | W |
| E. amylovora 273 | - | - | - | - | - | W | - | - | - | W | - | W | - | - | W | W | - | W | W | W | W | W | - | W | - | W | - | - |
| C. michiganensis IVIA 873 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| Dickeya sp. IVIA 4830 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| P. atrosepticum IVIA 3447 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| P. carotovorum IVIA 3902 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| P. savastanoi IVIA 1628.3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| R. radiobacter C58 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| R. rhizogenes K84 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| R. solanacearum IVIA 1670 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| X. arboricola pv. pruni CITA 33 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| X. vesicatoria CECT 792 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| A. hydrophila CECT 5173 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| A. faecalis CECT 928 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| B. cereus CECT 495 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| E. faecalis CECT 481 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| E. coli CECT 101 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| K. pneumoniae CECT 143 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| K. rhizophila CECT 241 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| P. fluorescens CECT 378 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| P. hauseri CECT 484 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| S. enterica CECT 443 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| S. marcescens CECT 159 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| S. aureus CECT 4013 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| E. amylovora strains lysed (%) | 84 | 68 | 79 | 79 | 79 | 100 | 84 | 90 | 79 | 74 | 58 | 68 | 84 | 84 | 63 | 84 | 90 | 74 | 80 | 79 | 90 | 74 | 68 | 74 | 84 | 95 | 68 | 68 |
| Non-E. amylovora species lysed (%) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
+ lysis by spot test on solid medium; - no lysis; W weak lysis; -/+ no lysis by spot test on solid medium/lysis in liquid medium with selected phages.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



