Submitted:
18 January 2024
Posted:
19 January 2024
You are already at the latest version
Abstract
Chronic lymphocytic leukemia (CLL) is characterized by dysfunctional B cells. Immune checkpoint molecules such as cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), and programmed death-1 (PD-1) are upregulated in patients with CLL and may correlate with prognostic markers such as beta-2 microglobulin (B2M). The aim of this study was to evaluate the levels of immune checkpoints on B-cell subsets and to further correlate them with B2M levels in patients with CLL. We recruited 21 patients with CLL and 12 controls. B cell subsets and the levels of immune checkpoint expression were determined using conventional multi-color flowcytometry. Basal levels of B2M in patients with CLL were measured using an Enzyme-linked immunosorbent assay. Patients with CLL had increased levels of activated B cells when compared to the control group, p< 0.001. The expression of PD-1 and CTLA-4 were increased on activated B cells and memory B cells, p< 0.05. There were no associations between B2M levels and the measured immune checkpoints on B cell subsets, after adjusting for sex and age. African patients with CLL express elevated levels of PD-1 and CTLA-4 immune checkpoints on activated and memory B cell subsets. However, there is no correlation between these immune checkpoint expressions and B2M levels.
Keywords:
Chronic lymphocytic leukaemia
; immune checkpoints
; B cell subsets
; beta-2 microglobulin
; programmed death protein 1
; cytotoxic T-lymphocyte associated protein 4
1. Introduction
Chronic lymphocytic leukemia (CLL) is the most common type of leukemia among adults, with a global prevalence of about 3.5 cases per 100,000 people (1). In high-income countries, CLL accounts for more than a third of all leukemia cases (1). Notably, low-to-middle income countries have a five to ten-fold lower age-adjusted incidence rate of CLL as compared to high-income countries (2). CLL is a lymphoproliferative disorder that is characterized by functionally incompetent B cells with a distinct immunophenotype (3). However, there are divergent findings regarding the predictive significance of B cell subsets in in leukaemia (4).
Regulatory B cells (Breg) modulate T cell-driven anti-tumor immunity, promoting the expression of forkhead box protein 3 (FoxP3+) in regulatory T cells (T-regs), which dampens the innate and adaptive antitumor immune response (4, 5). These immonusuppressive mechanisms involve Bregs that acquire inhibitory ligands and signal transducer and activator of transcription 3 (STAT3) phosphorylation, and the induction of interleukin 10 (IL-10) and transforming growth factor-β (TGF-β) (4, 5). The expression of inhibitory molecules such as programmed death-1 (PD-1) or programmed death-ligand 1 (PD-L1) inhibit the anti-tumor function of Bregs (4). In patients with CLL, immune checkpoints such as T-cell immunoglobulin-3 (TIM-3), cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), and programmed death-1 (PD-1) are upregulated (6, 7). The use of immune checkpoint inhibitors (ICI) such as PD-1 antagonists, has fundamentally altered therapeutic approaches in patients with CLL (8). Targeting PD-1 has shown to improve the overall survival of patients with CLL while also reducing adverse effects (8). In patients with relapsed CLL, zanubrutinib, a Bruton’s tyrosine kinase inhibitor is associated with fewer cardiac adverse events and improved progression-free survival (9).
There has been a significant transformation in the management of patients with CLL. Advances in prognostic markers, enhanced predictive capabilities for clinical outcomes have collectively made vital impact in the management of CLL. The important adverse prognostic markers which include immunoglobulin heavy-chain variable region gene (IGHV) mutation status, zeta-chain-associated protein kinase-70 (ZAP-70) and CD38 expression, and beta 2 microglobulin (B2M) levels (10). The levels of B2M are considered as an independent marker for poor prognosis in patients with CLL (11). The serum levels of B2M are elevated in patients with CLL who at an advanced stage of the disease, and these levels decrease post-treatment (12). The primary aim of this study was to determine the levels of immune checkpoint expression on peripheral B-cell subsets in patients with CLL and to further correlate these immune checkpoints with B2M levels in patients with CLL.
2. Methods and Materials
2.1. Patient Recruitment
Patients and healthy control participants were recruited between July 2019 to May 2022 from King Edward VIII Hospital, a tertiary healthcare facility located in Durban, KwaZulu-Natal, South Africa. The study was conducted in accordance with the Declaration of Helsinki, and ethical approval was obtained from the University of KwaZulu-Natal Biomedical Research Ethics Committee (BE456/18), South Africa. Written informed consent was obtained from all study participants. In this study we excluded patients with CLL who were on treatment and only untreated patients were included along with age-matched healthy controls with no clinical signs of infection. The ethnicity of the participants was self-reported.
2.2. Sample Collection
Peripheral blood was collected from consenting participants by venepuncture into 6mL ethylenediaminetetraacetic acid (EDTA) tubes (BD Bioscience, USA). Samples were transported at room temperature from the hospital to the laboratory.
2.3. Haematological Analysis
Haematological parameters including the white blood cell count, haemoglobin and platelet count were measured using an automated Coulter AcT Diff haematology analyser (Beckman Coulter Inc., California, United States) within 1–2 hours of peripheral blood collection.
2.4. Isolation of Peripheral Blood Mononuclear Cells (PBMCs)
The peripheral blood mononuclear cells (PBMCs) were isolated from whole blood samples by density-gradient centrifugation method, using the Ficoll-Paque PLUS (Amersham, Biosciences, Uppsala, Sweden), as previously described (13). Briefly, 3 mL of Ficoll-Paque PLUS was aliquoted into a 15 mL centrifuge tube (Sigma-Aldrich, Germany) and 4 mL of whole blood was carefully layered on the Ficoll-Paque PLUS gradient and the samples were centrifuged at 400× g for 40 minutes at 20 °C. The PBMC layer was then collected and stored at −80°C.
2.5. T Cell Depletion and B Cell Isolation from Peripheral Blood Mononuclear Cells
To enrich the B cell population from the collected PBMCs, we performed T cell depletion and positively selected B cells using the BD IMag isolation system (BD Bioscience, USA). Briefly, 50 µL PBMCs were incubated with 5 µL of biotinylated human T lymphocyte enrichment cocktail (BD Biosciences, San Jose, CA, USA) for 15 minutes at room temperature. After the incubation, 50 µL of streptavidin particles were added to the T cell depleted PBMC samples and incubated for 30 minutes, at room temperature. The samples were then reconstituted into 1 mL of 3.2% sodium citrated buffer and samples were placed on the BD Imag for 8 minutes. Isolated B cells were reconstituted into 100 µL PBS and stored −80 °C.
2.6. Measurements of B Cell Subsets
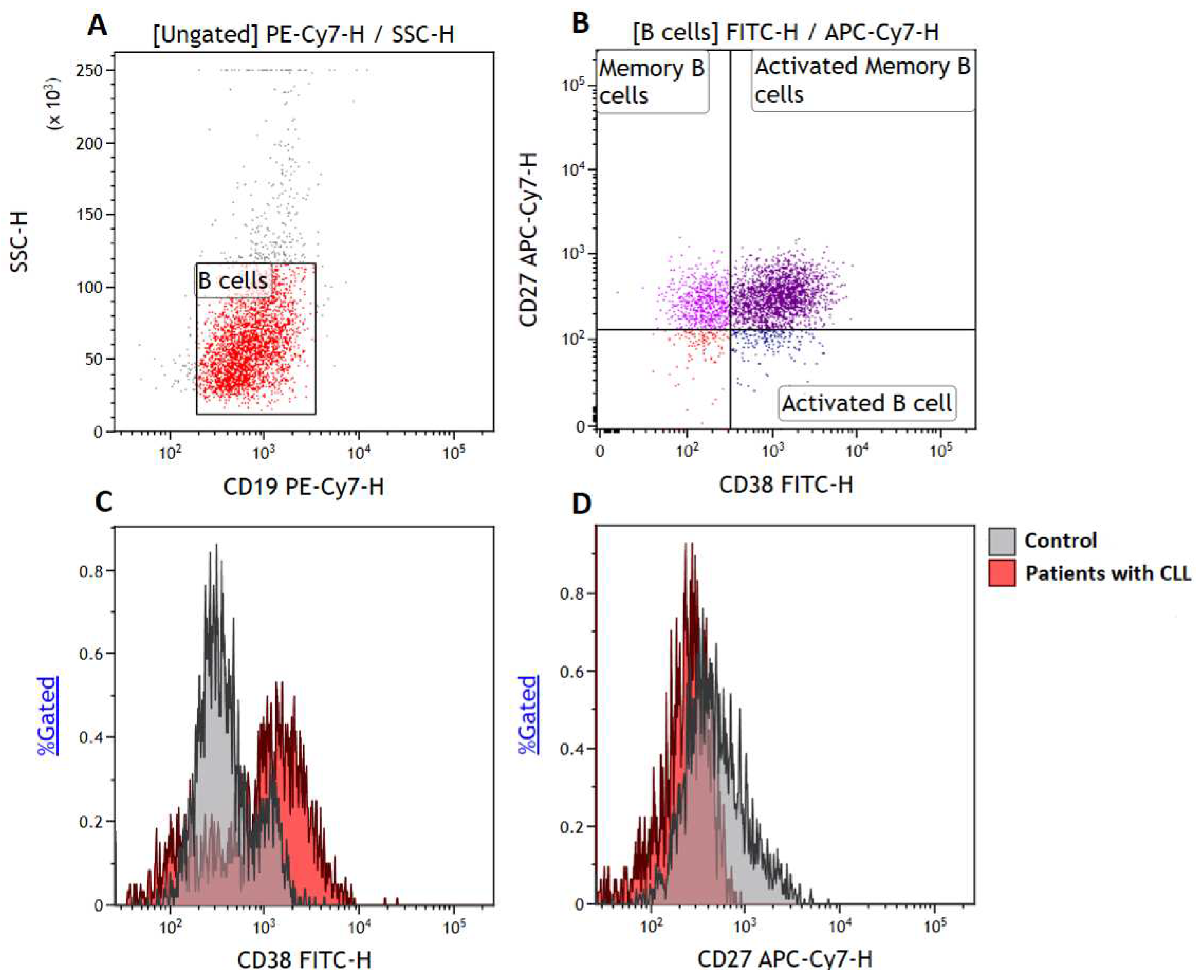
To quantify B cell subsets, we made use of a six-colour flow cytometry panel consisting of CD38-FITC, CD152-PE, CD273-APC, CD19-PE/Cy7, CD27-APC/Cy7 and CD279-BV421 (BioLegend, San Diego, CA, USA). We acquired at least 5000 B cells (CD19+ events) Figure 1A and defined memory B cells as CD19+CD27+ events (14, 15) (Figure 1B), activated B cells as CD19+CD27-CD38+ events and activated memory B cells (15) as CD19+CD27+CD38+ events (Figure 1C).
2.7. Measurements of Immune Checkpoint Levels on B Cell Subsets
To determine the levels of immune checkpoint expression on B cell subsets we measured the expression of CD279 (PD-1), CD273 (PD-L2) and CD152 (CTLA-4) on BMEM, and activated B cells. A total of 5 000 CD19+ events were acquired at a medium flow rate. All the data were acquired using the BD FACSCanto II flow cytometer (BD Biosciences, San Diego, CA, USA) and analysed using Kaluza version 1.2 (Beckman coulter, Inc Brea, CA, USA).
Figure 1.
Gating strategy. (A) B cells were gated based on side scatter (SSC) and CD19 expression, (B) illustrates the gating of activated B cells defined as CD19+CD38+CD27-, memory B cells defined as CD19+CD27+ events and activated memory B cells defined as CD19+CD27+CD38+ events, respectively. Histograms demonstrate the levels of activated B cells (C) and memory B cells (D) on the control group and patients with CLL.
Figure 1.
Gating strategy. (A) B cells were gated based on side scatter (SSC) and CD19 expression, (B) illustrates the gating of activated B cells defined as CD19+CD38+CD27-, memory B cells defined as CD19+CD27+ events and activated memory B cells defined as CD19+CD27+CD38+ events, respectively. Histograms demonstrate the levels of activated B cells (C) and memory B cells (D) on the control group and patients with CLL.
2.8. Measurements of Serum Soluble Beta-2-Microglobulin (B2M) Levels
To measure the plasma levels of B2M, a prognostic marker in patients with CLL (16), we made use of the beta-2-microglobulin Human enzyme-linked immunosorbent assay kit (ThermoFisher Scientific, Waltham, Massachusetts, USA) according to the manufacturer’s instruction.
2.9. Sample Size Estimation
We calculated the minimum sample size of patients needed to detect a large effect size (d) of 1.14 in the expression of immune checkpoints, namely PD-1 and PD-L1 at 80% power and alpha (α) of 0.05 using GPower 3.1.94 software (Universität, Germany). To detect a large effect size between two independent means using an unpaired t-test, we required a minimum of twenty-one patients with CLL (n = 21) and twelve controls (n = 12).
2.10. Statistical Analysis
All statistical analysis was performed using GraphPad Prism version 8 software, (GraphPad Software Inc, San Diego, CA, USA) and (17). All non-parametric data were log-transformed prior to statistical analysis and reported as mean and standard deviation. An unpaired Student’s t-test was performed to compare parametric data between two groups. A repeated measures one-way ANOVA was used to compare parametric multiple data of the same group with Dunnett’s as a post hoc test for multiple comparisons. To correct for multiple comparisons, a Bonferroni-corrected critical P-value of ˂ 0.0167 was considered as statistically significant.
2.11. Data Availability
The data generated in this study are available upon request from the corresponding author.
3. Results
3.1. Patients Characteristics and Haematological Parameters
This study comprised of 21 patients with CLL and 12 healthy controls. The mean age of the patients with CLL was 62.33 ± 13.31 years and controls had a mean age of 56.58± 15.67. The included study participants were from different ethnic groups, which comprised of African (n = 30), European (n = 2), and Indian (n = 1). The study included 39.39% females and 60.61% males.
In patients with CLL, the white blood cell count was significantly increased (130.4 x 103 ± 29.71) compared to controls (5.26 × 103 ± 1.38), p = 0.0005) (Table 1). Whereas the red blood cell count and haemoglobin were significantly reduced in patients with CLL compared to age-matched control patients (p < 0.0001). There were no statistically significant differences in the platelet count in patients with CLL when compared to the control group (p = 0.1831).
3.2. Increased Levels of Activated B Cells in Patients with CLL
We evaluated the levels of B cell subsets in patients with CLL (Figure 2). Notably, patients with CLL had significantly increased levels of activated B cells (57.39 ± 8.001) compared to the control group (28.47 ± 19.01; p = 0.0002).
Furthermore, the expression of memory B cells in patients with CLL (40.95 ± 8.353) was found to be statistically comparable to that in the control group (46.45 ± 20.90; p = 0.3984), as was the expression of activated memory B cells expressing (37.31 ± 7.191) in patients with CLL compared to the control group (44.25 ± 21.34; p = 0.2956).
3.3. Elevated Levels of PD-1 Expression on Activated B-Cells and Memory B Cells in Patients with CLL
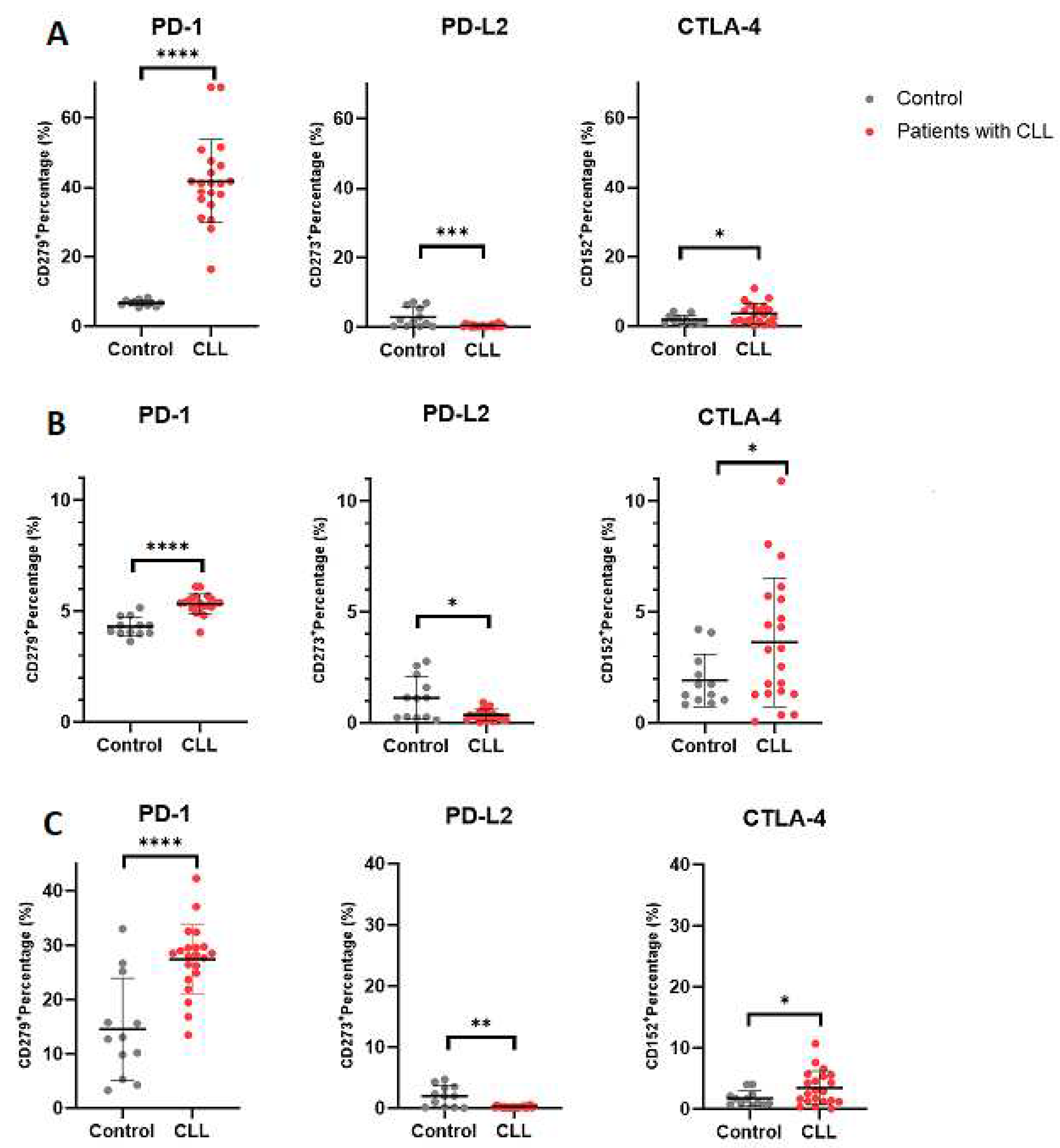
Significantly elevated PD-1 expression was observed on B cells from patients with CLL (40.20 ± 6.601) compared to the control group (6.690 ± 0.8160; p < 0.0001). Likewise, PD-1 expression was increased on activated B cells in patients with CLL (5.325 ± 0.4397) relative to the control group (4.305 ± 0.4317; p < 0.0001). Moreover, a notable increase in PD-1 expression was noted on memory B cells in patients with CLL (27.44 ± 6.358) compared to the control group (14.59 ± 9.395; p < 0.0001).
3.4. Elevated Levels of CTLA-4 on Activated and Memory B Cells in Patients with CLL
A significant increase in CTLA-4 expression was detected on B cells in patients with CLL (3.630 ± 2.897) compared to the control group (1.919 ± 1.184; p = 0.0241). Similarly, CTLA-4 expression was elevated on activated B cells in patients with CLL (3.631 ± 2.896) relative to the control group (1.919 ± 1.184; p = 0.0240). Notably, CTLA-4 expression on memory B cells in patients with CLL (3.439 ± 2.767) was higher compared to the control group (1.755 ± 1.205; p = 0.0221).
3.5. Decreased Levels of PD-L2 Expression on Activated and Memory B Cells in Patients with CLL
Remarkably, a significant decrease in PD-L2 expression was significantly decreased on B cells in patients with CLL (0.3967 ± 0.3343) compared to the control group (2.870 ± 2.800; p = 0.0003). A similar decrease in PD-L2 expression was observed on activated B cells in patients with CLL (0.3624 ± 0.2514) compared to the control group (1.137 ± 0.9655; p = 0.0186). Additionally, PD-L2 expression on memory B cells in patients with CLL (0.2005 ± 0.1297) was reduced compared to the control group (1.956 ± 1.706; p = 0.0044).
Figure 3.
Immune checkpoint expression on B Cell subsets. Figure 3 depict the expression levels of PD-1, PD-L2 and CTLA-4 on (A) normal B cells, (B) activated B cells and (C) memory B cells, respectively. The data is presented as the mean ± standard deviation (SD).
Figure 3.
Immune checkpoint expression on B Cell subsets. Figure 3 depict the expression levels of PD-1, PD-L2 and CTLA-4 on (A) normal B cells, (B) activated B cells and (C) memory B cells, respectively. The data is presented as the mean ± standard deviation (SD).
3.6. Analysis of Immune Checkpoint (PD-1 and CTLA-4) Secretion among B Cell Subsets in Patients with CLL
In order to elucidate which B cell subsets were associated with heightened immune checkpoint secretion, we conducted a comparative assessment of immune checkpoint expression within various B cell subsets in patients with CLL (Figure 4). PD-1 levels were significantly elevated on activated B cells when compared to memory B cells (0.5877 ± 0.04654, p < 0.0001). Similarly, CTLA-4 expression was found to be elevated activated when comparable to memory B cells (0.1924 ± 0.04848, p = 0.0039).
3.7. β2 Microglobulin Levels Are not Associated with the Expression of PD-1 or CTLA-4 on B Cell Subsets
The levels of B2M are independently associated with disease progression in patients with CLL (16). In our multivariable regression model, no association between B2M levels and PD-1 on B cells (β = 0.008, SE = 0.011, p = 0.524), PD-1 on activated B cells (β = 0.008, SE = 0.011, p = 0.525), PD-1 on memory B cells (β = 0.0008, SE = 0.004, p = 0.905), CTLA-4 on B cells (β = 0.004, SE = 0.002, p = 0.426), CTLA-4 on activated B cells (β = 0.004, SE = 0.002, p = 0.419), CTLA-4 on memory B cells (β = 0.004, SE = 0.002, p = 0.419) in patients with CLL (supplementary 1).
4. Discussion
The primary aim of this study was to determine the levels of immune checkpoint expression on the peripheral B-cell subsets in patients with CLL and to further correlate these immune checkpoints with B2M levels, a confirmed independent prognostic maker in patients with CLL (16). In this study, we determined the baseline expression levels of B cell subsets in patients with CLL. Patients with CLL had a higher activated B cell fraction (CD38+ B cells) which has already been described in patients with CLL (18). In our study, the activated B cell had CD38 phenotype, which is one of the prognostic markers for patients with CLL (19, 20). Notably, the activated B cell profile is not restricted to peripheral circulation (21) and the expression of CD38 is associated with an increased proliferation index (22, 23). The increased levels of CD38+ activated B cells in patients with CLL may indicate patients with poor survival rate and response to therapy (18-20). Moreover, in our study, patients with CLL expressed higher CTLA-4 and PD-1 immune checkpoint proteins on activated B cell subset. The levels of PD-1 and CTLA-4 were higher on activated B cells when compared to the other B cell subsets.
The relevance of PD-1 expression on CD4+ T cells has been well described in patients with CLL with an advanced stage (Rai III/IV) (24). To date the exhaustion levels of B cell subsets in patients with CLL has not been investigated. Immune system dysfunction is a hallmark of CLL, which primarily affects humoral immunity and is characterised by increased susceptibility to autoimmune disorders and secondary malignancies (25, 26).
Immune checkpoint proteins such as PD-1, PD-L1, PD-L2 and CTLA-4 are expressed on activated B cells (27, 28). The function of these immune checkpoints on B cells have not been fully evaluated. However, the binding of PD-L1 or PD-L2 to PD-1 receptors on T cells is known to induce phosphorylation of ITIM (Immunoreceptor Tyrosine-Based Inhibitory Motif) and ITSM (Immunoreceptor Tyrosine-Based Switch Motif) motifs within the PD-1 receptor (29). This phosphorylation leads to the recruitment of phosphatases like Src Homology 2 Domain-Containing Protein Tyrosine Phosphatase 2 and Src Homology 2 Domain-Containing Protein Tyrosine Phosphatase 1 (29, 30). Consequently, these phosphatases dephosphorylate key signaling molecules downstream of the T cell receptor (TCR), such as PI3K (Phosphoinositide 3-Kinase) and AKT (Protein Kinase B) (31-33). The overall effect of PD-1 binding is to inhibit the proliferation of B cells, leading to immune suppression and tumor immune evasion (34).
In our study, we further observed a significant increase in PD-1 levels on B cells and memory B cells among patients with CLL. Furthermore, PD-L2 expression was lower across the B cell subsets in patients with CLL. These findings underscore the complex and dynamic interplay between immune checkpoint molecules and distinct B cell populations within the CLL microenvironment. PD-1 function in T cells is widely studied, and is associated with T cell exhaustion in patients with CLL (35). However, its role on B cell is not known. In our study, we showed that PD-1 is expressed strongly on activated B cells. Thibult et al., (34) showed that PD-1 and its ligands (PD-L1 and PD-L2) are key regulatory proteins of B-cell activation and entry into the germinal centre. Moreover, the study demonstrated that PD-1 is the inhibitor of the Toll like receptor (TLR)-ligand-mediated activation of B cells. The increased expression of PD-1 may inhibit B cell differentiation into antibody secreting cells. PD-1 has a higher affinity for PD-L1 than PD-L2 (34, 36), this may explain the significant decrease in PD-L2 expression in patients with CLL.
In this study we observed a significant increase in CTLA-4 levels on B cells and memory B cells among patients with CLL. CTLA-4 suppresses humoral response to T cell-dependent and independent antigen (37). In addition, elevated CTLA-4 expression on B cells is associated with disease progression in patients with CLL (38). Moreover, the elevated expression of this immune checkpoint in peripheral blood inhibit B cell activation and proliferation (39). Consistent to this study, our study found increased levels of CTLA-4 expression in B cell subsets. Therefore, the use of immune checkpoint inhibitors targeting CTLA-4 in B cells may be beneficial in patients with CLL.
In this study, we determined the relationship between immune checkpoints on B cell subsets and B2M. There was no correlation between B2M levels and the expression of immune checkpoint molecules on B cell subsets. The elevated B2M levels are associated with an advanced CLL stage (40). In CLL, B2M levels can be elevated due to increased turnover of CLL cells or other factors. The elevated B2M is associated with advanced CLL stage (40). While immune checkpoints are dysregulated in CLL, B2M levels do not directly correlate with the expression of these checkpoints in B cell subsets. This suggests that the expression of these immune checkpoints in B cell subsets do not directly influence B2M levels.
5. Conclusion
African patients with CLL exhibits increased levels of PD-1 and CTLA-4 on B cell subsets in patients with CLL. However, the expression of these immune checkpoint molecules on B cell subsets do not correlate with B2M levels.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Age, sex adjusted multivariable regression analysis of immune checkpoints on B cell subsets with B2M.
Author Contributions
Conceptualization, A.N., B.B.N. and P.V.D.; methodology, A.N., B.B.N. and Z.A.M.; formal analysis, A.N. and B.B.N.; investigation, A.N., B.B.N. and P.V.D.; data curation, A.N., B.B.N. and Z.A.M.; writing—original draft preparation, A.N.; writing—review & editing, A.N., B.B.N. and P.V.D.; visualization, A.N., B.B.N. and P.V.D.; supervision, B.B.N. and P.V.D.; project administration, A.N. All authors have read and agreed to the published version of the manuscript.
Funding
No specific grant was obtained for this research from governmental, private, or nonprofit funding organizations.
Institutional Review Board Statement
The ethical approval for this study was granted by the University of KwaZulu-Natal Biomedical Research Ethics Committee (study approval no. BE456/18).
Informed Consent Statement
Written informed consent was obtained from all study participants.
Data Availability Statement
Data are contained within the article and supplementary materials.
Acknowledgments
The authors would like to thank the King Edward Regional hospital staff and participants of this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- DeSantis, C.E.; Miller, K.D.; Dale, W.; Mohile, S.G.; Cohen, H.J.; Leach, C.R.; et al. Cancer statistics for adults aged 85 years and older. 2019, 69, 452–467. [Google Scholar]
- Yang, S.; Varghese, A.M.; Sood, N.; Chiattone, C.; Akinola, N.O.; Huang, X.; et al. Ethnic and geographic diversity of chronic lymphocytic leukaemia. Leukemia 2021, 35, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Lanasa, M.C.J.H. The American Society of Hematology Education Program Book. Novel insights into the biology of CLL 2010, 2010, 70–76. [Google Scholar]
- Gupta, S.L.; Khan, N.; Basu, S.; Soni, V. B-cell-based immunotherapy: A promising new alternative. Vaccines 2022, 10, 879. [Google Scholar] [CrossRef]
- Schwartz, M.; Zhang, Y.; Rosenblatt, J.D. B cell regulation of the anti-tumor response and role in carcinogenesis. Journal for immunotherapy of cancer 2016, 4, 1–15. [Google Scholar] [CrossRef]
- Palma, M.; Gentilcore, G.; Heimersson, K.; Mozaffari, F.; Näsman-Glaser, B.; Young, E. T cells in chronic lymphocytic leukemia display dysregulated expression of immune checkpoints and activation markers. 2017; 102, 562–572. [Google Scholar]
- Zhang, X.; Zhang, H.; Chen, L.; Feng, Z.; Gao, L.; Li, Q.J.C. TIGIT expression is upregulated in T cells and causes T cell dysfunction independent of PD-1 and Tim-3 in adult B lineage acute lymphoblastic leukemia. 2019, 344, 103958. [Google Scholar] [CrossRef]
- Ding, W.; LaPlant, B.R.; Call, T.G.; Parikh, S.A.; Leis, J.F.; He, R.; et al. Pembrolizumab in patients with, C.L.L.; Richter transformation or with relapsed, C.L.L. Blood. The Journal of the American Society of Hematology 2017, 129, 3419–3427. [Google Scholar]
- Brown, J.R.; Eichhorst, B.; Hillmen, P.; Jurczak, W.; Kaźmierczak, M.; Lamanna, N.; et al. Zanubrutinib or ibrutinib in relapsed or refractory chronic lymphocytic leukemia. New England Journal of Medicine 2023, 388, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Dal Bo, M.; Bulian, P.; Bomben, R.; Zucchetto, A.; Rossi, F.; Pozzo, F.; et al. CD49d prevails over the novel recurrent mutations as independent prognosticator of overall survival in chronic lymphocytic leukemia. Leukemia 2016, 30, 2011–2018. [Google Scholar] [CrossRef]
- Wierda, W. New prognostic factors in chronic lymphocytic leukemia. Clinical Advances in Hematology & Oncology: H&O 2009, 7, 32–33. [Google Scholar]
- Al-Rekabi, A.N.; Alwan, A.F.; Alobaidi, N.K. Assessment of beta-2 microglobulin and CD49d in patients with chronic lymphocytic leukemia pre-and posttherapy. Iraqi Journal of Hematology 2020, 9, 155–159. [Google Scholar]
- Jaatinen, T.; Laine, J. Isolation of Mononuclear Cells from Human Cord Blood by Ficoll-Paque Density Gradient. Current Protocols in Stem Cell Biology 2007, 1, 2A.1.–2A14. [Google Scholar] [CrossRef]
- Axelsson, S.; Magnuson, A.; Lange, A.; Alshamari, A.; Hörnquist, E.H.; Hultgren, O. A combination of the activation marker CD86 and the immune checkpoint marker B and T lymphocyte attenuator (BTLA) indicates a putative permissive activation state of B cell subtypes in healthy blood donors independent of age and sex. BMC Immunology 2020, 21, 14. [Google Scholar] [CrossRef]
- Joscelyn, J.; Ochoa-Repáraz, J.; Kasper, L. Principles of Immunotherapy. Clinical Neuroimmunology; Springer, 2020; pp. 17–42. [Google Scholar]
- Mkhwanazi, Z.A.; Nyambuya, T.M.; Mfusi, S.A.; Nkambule, B.B. Prognostic markers in patients with chronic lymphocytic leukaemia on targeted therapy, chemoimmunotherapy with anti-CD20 monoclonal antibody: a systematic review and meta-analysis of prognostic factors. BMC cancer 2022, 22, 1218. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. A Language and Environment for Statistical Computing 2021. Available online: https://www.r-project.org/.
- Damle, R.N.; Temburni, S.; Calissano, C.; Yancopoulos, S.; Banapour, T.; Sison, C.; et al. CD38 expression labels an activated subset within chronic lymphocytic leukemia clones enriched in proliferating B cells. Blood. The Journal of the American Society of Hematology 2007, 110, 3352–3359. [Google Scholar]
- Gentile, M.; Morabito, F.; Del Poeta, G.; Mauro, F.R.; Reda, G.; Sportoletti, P.; et al. Survival risk score for real-life relapsed/refractory chronic lymphocytic leukemia patients receiving ibrutinib. A campus CLL study. Leukemia 2021, 35, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Amaya-Chanaga, C.I.; Rassenti, L.Z. Biomarkers in chronic lymphocytic leukemia: Clinical applications and prognostic markers. Best Pract Res Clin Haematol 2016, 29, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Soma, L.A.; Craig, F.E.; Swerdlow, S.H. The proliferation center microenvironment and prognostic markers in chronic lymphocytic leukemia/small lymphocytic lymphoma. Human pathology 2006, 37, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Khoudoleeva, O.; Gretsov, E.; Barteneva, N.; Vorobjev, I. Proliferative index and expression of CD38, Zap-70, and CD25 in different lymphoid compartments of chronic lymphocytic leukemia patients. Pathology and Laboratory Medicine International 2011, 7–16. [Google Scholar]
- Patten, P.E.; Buggins, A.G.; Richards, J.; Wotherspoon, A.; Salisbury, J.; Mufti, G.J.; et al. CD38 expression in chronic lymphocytic leukemia is regulated by the tumor microenvironment. Blood. The Journal of the American Society of Hematology 2008, 111, 5173–5181. [Google Scholar]
- Rusak, M.; Eljaszewicz, A.; Bołkun, Ł.; Łuksza, E.; Łapuć, I.; Piszcz, J.; et al. Prognostic significance of PD-1 expression on peripheral blood CD4+ T cells in patients with newly diagnosed chronic lymphocytic leukemia. Pol Arch Med Wewn 2015, 125, 553–539. [Google Scholar] [CrossRef] [PubMed]
- Hallek, M.; Shanafelt, T.D.; Eichhorst, B. Chronic lymphocytic leukaemia. Lancet 2018, 391, 1524–1537. [Google Scholar] [CrossRef] [PubMed]
- Forconi, F.; Moss, P. Perturbation of the normal immune system in patients with CLL. Blood 2015, 126, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, X.; Ma, Z.; Wang, C.; Yang, Q.; Byrne-Steele, M.; et al. CTLA-4 expression by B-1a B cells is essential for immune tolerance. Nat Commun 2021, 12, 525. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Akiba, H.; Iwai, H.; Matsuda, H.; Aoki, M.; Tanno, Y.; et al. Expression of programmed death 1 ligands by murine T cells and APC. J Immunol 2002, 169, 5538–5545. [Google Scholar] [CrossRef]
- Riley, J.L. PD-1 signaling in primary T cells. Immunological reviews 2009, 229, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, U. SHP-1 and SHP-2 in T cells: two phosphatases functioning at many levels. Immunological reviews 2009, 228, 342–359. [Google Scholar] [CrossRef]
- Boussiotis, V.A.; Chatterjee, P.; Li, L. Biochemical signaling of PD-1 on T cells and its functional implications. Cancer journal (Sudbury, Mass) 2014, 20, 265. [Google Scholar] [CrossRef]
- Patsoukis, N.; Wang, Q.; Strauss, L.; Boussiotis, V.A. Revisiting the PD-1 pathway. Science advances 2020, 6, eabd2712. [Google Scholar] [CrossRef]
- Chemnitz, J.M.; Parry, R.V.; Nichols, K.E.; June, C.H.; Riley, J.L. SHP-1 and SHP-2 associate with immunoreceptor tyrosine-based switch motif of programmed death 1 upon primary human T cell stimulation, but only receptor ligation prevents T cell activation. The Journal of Immunology 2004, 173, 945–954. [Google Scholar] [CrossRef]
- Thibult, M.L.; Mamessier, E.; Gertner-Dardenne, J.; Pastor, S.; Just-Landi, S.; Xerri, L.; et al. PD-1 is a novel regulator of human B-cell activation. Int Immunol 2013, 25, 129–137. [Google Scholar] [CrossRef]
- Riches, J.C.; Davies, J.K.; McClanahan, F.; Fatah, R.; Iqbal, S.; Agrawal, S.; et al. T cells from CLL patients exhibit features of T-cell exhaustion but retain capacity for cytokine production. Blood. The Journal of the American Society of Hematology 2013, 121, 1612–1621. [Google Scholar]
- Ghiotto, M.; Gauthier, L.; Serriari, N.; Pastor, S.; Truneh, A.; Nunès, J.A.; et al. PD-L1 and PD-L2 differ in their molecular mechanisms of interaction with PD-1. International immunology 2010, 22, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Linsley, P.S.; Wallace, P.M.; Johnson, J.; Gibson, M.G.; Greene, J.L.; Ledbetter, J.A.; et al. Immunosuppression in vivo by a soluble form of the CTLA-4 T cell activation molecule. Science 1992, 257, 792–795. [Google Scholar] [CrossRef] [PubMed]
- Kosmaczewska, A.; Ciszak, L.; Suwalska, K.; Wolowiec, D.; Frydecka, I. CTLA-4 overexpression in CD19+/CD5+ cells correlates with the level of cell cycle regulators and disease progression in B-CLL patients. Leukemia 2005, 19, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Karabon, L.; Partyka, A.; Ciszak, L.; Pawlak-Adamska, E.; Tomkiewicz, A.; Bojarska-Junak, A.; et al. Abnormal expression of BTLA and CTLA-4 immune checkpoint molecules in chronic lymphocytic leukemia patients. Journal of Immunology Research 2020, 2020. [Google Scholar] [CrossRef]
- Montillo, M.; Hamblin, T.; Hallek, M.; Montserrat, E.; Morra, E. Chronic lymphocytic leukemia: novel prognostic factors and their relevance for risk-adapted therapeutic strategies. Haematologica 2005, 90, 391–399. [Google Scholar]
Figure 2.
B Cell Subsets in patients with CLL. (A–C) depict the levels of activated B cells, memory B cells and activated memory B cells in patients with CLL compared to healthy controls. The data is presented as the mean ± standard deviation (SD).
Figure 2.
B Cell Subsets in patients with CLL. (A–C) depict the levels of activated B cells, memory B cells and activated memory B cells in patients with CLL compared to healthy controls. The data is presented as the mean ± standard deviation (SD).
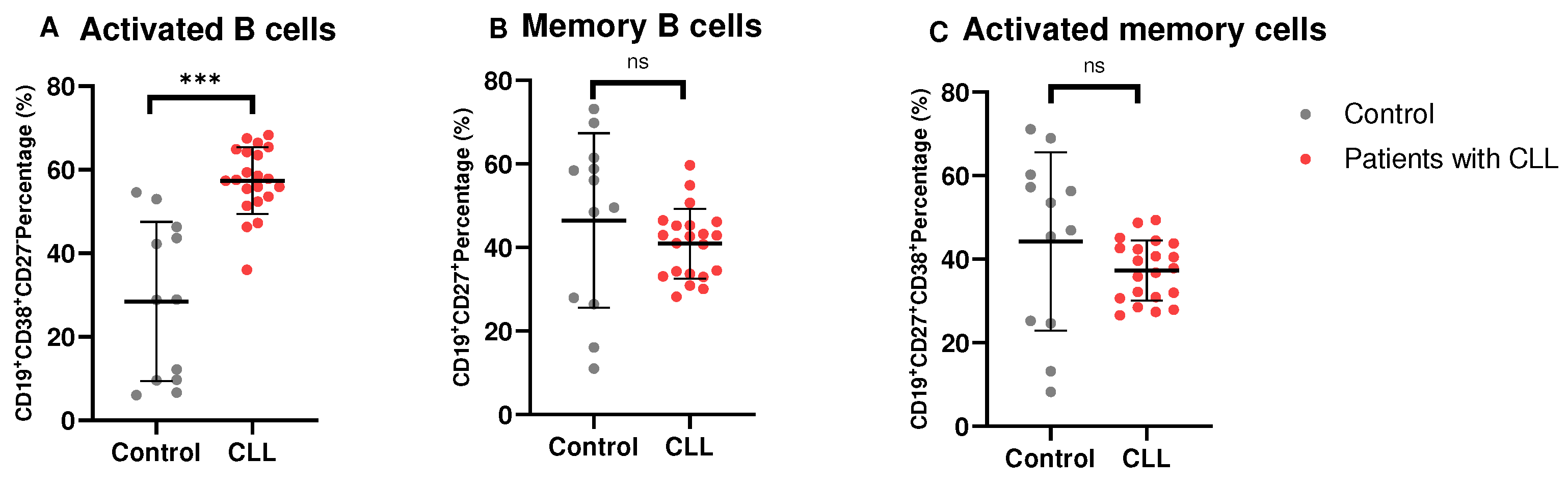
Figure 4.
Immune checkpoint expression on B cell subsets. (A) illustrates the expression of PD-1 on activated (CD19+CD38+ cells) and memory B cells (CD19+CD27+ cells). (B) illustrates the levels of CTLA-4 expression on activated B cells and memory B cells. All data are presented as the mean ± standard deviation (SD).
Figure 4.
Immune checkpoint expression on B cell subsets. (A) illustrates the expression of PD-1 on activated (CD19+CD38+ cells) and memory B cells (CD19+CD27+ cells). (B) illustrates the levels of CTLA-4 expression on activated B cells and memory B cells. All data are presented as the mean ± standard deviation (SD).
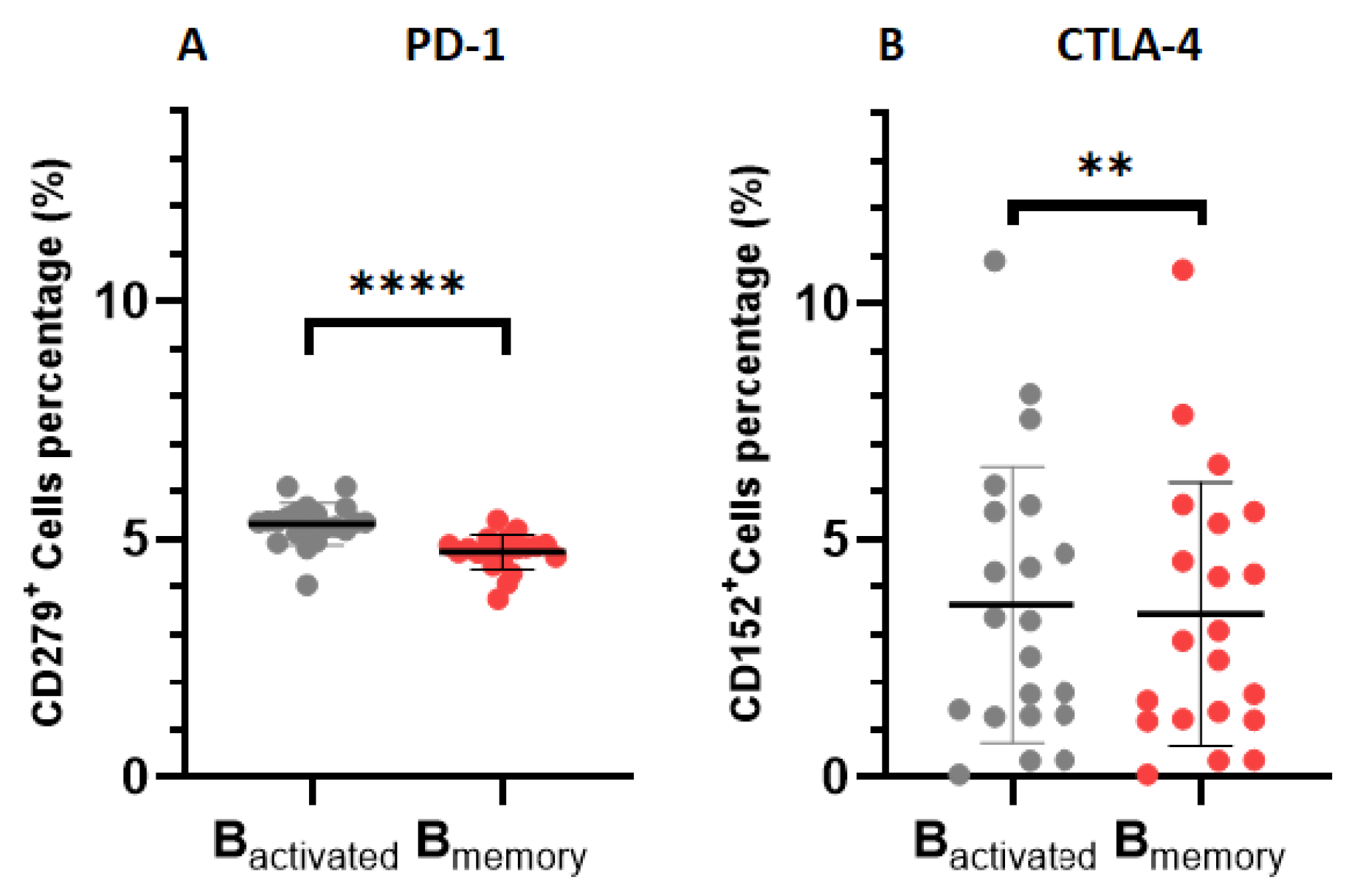

Table 1.
The baseline characteristics and haematological parameters of the participants.
| Control (n = 12) |
Patients with CLL (21) |
p-Value | |
|---|---|---|---|
| Age (Years) | 56.58± 15.67 | 62.33± 13.31 | 0.2714 |
| Male, n (%) | 58.33 | 61.9 | |
| Female, n (%) | 41.67 | 38.1 | |
| White blood cell count (103 µL) | 5.26± 1.38 | 130.4± 29.71 | 0.0005 |
| Red blood cell (106 µL) | 4.74± 0.94 | 2.10± 0.84 | <0.0001 |
| Haemoglobin (g/dL) | 14.13± 3.81 | 8.19± 2.30 | <0.0001 |
| Platelets (103 µL) | 210.4± 73.14 | 157.5± 141.9 | 0.1831 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



