
Submitted:
16 January 2024
Posted:
17 January 2024
You are already at the latest version
Abstract
Folk medicine is widely used in Angola, even for human African trypanosomiasis (sleeping sickness) in spite of the fact that the reference treatment is available for free. Aiming to validate herbal remedies in use, we have selected nine medicinal plants and assessed their antitrypanosomal ac-tivity. 122 extracts were prepared using different plant parts and solvents. 15 extracts from seven different plants exhibited in vitro activity against Trypanosoma brucei rhodesiense bloodstream forms >70% at 20 µg/ml. The dichloromethane extract of Nymphaea lotus (leaves and leaflets) and the ethanolic extract of Brasenia schreberi (leaves) had IC50 values ≤10 µg/ml. These two aquatic plants are of particular interest. They are being co-applied in the form of a decoction of leaves because they are considered by local healers as male and female of the same species, the ethnotaxon “longa dia simbi”. Bioassay-guided fractionation led to the identification of eight active molecules: gallic acid (IC50 0.5 µg/ml), methyl gallate (IC50 1.1 µg/ml), 2,3,4,6-tetragalloyl-glucopyranoside, ethyl gallate (IC50 0.5µg/ml), 1,2,3,4,6-pentagalloyl-β-glucopyranoside (IC50 20µg/ml), gossypet-in-7-O-β-glucopyranoside (IC50 5.5µg/ml), and hypolaetin-7-O-glucoside (IC50 5.7µg/ml) in B. schreberi, and 5-[(8Z,11Z,14Z)-heptadeca-8,11,14-trienyl] resorcinol (IC50 5.3µg/ml) not described to date in N. lotus. Five of these active constituents were detected in the traditional preparation. This work provides the first evidence for the ethnomedicinal use of these plants in the management of sleeping sickness in Angola.
Keywords:
ethnopharmacology
; African medicinal plant
; antiprotozoal
; trypanosomiasis
; Brasenia schreberi
; Nymphaea lotus
; Angola
1. Introduction
The extensive use of folk medicine in Africa, composed mainly of medicinal plants, is linked to cultural as well as economic reasons. This is why the WHO encourages African member states to promote and integrate traditional medical practices in their health systems [1]. In Angola, 72%1 of the population uses herbal medicines to treat various medical affections, including parasitic infections such as human African trypanosomiasis (HAT), also called sleeping sickness.
HAT is a vector-borne disease Neglected Tropical Disease (NTD) that is transmitted by the bite of infected tsetse flies (Glossina spp.). HAT is caused by two subspecies of the protozoan parasite Trypanosoma brucei: T. b. gambiense in west and central Africa including Angola is responsible for the chronic form, whereas T. b. rhodesiense prevalent in eastern Africa causes the acute form [2]. Both forms are fatal if untreated. The majority of HAT-cases are of the Gambian form (g-HAT) and 57 million people are at risk of contracting g-HAT [3]. In Angola, g-HAT is endemic in the northwestern part. It is prevalent in seven of eighteen provinces [4]. It affects mainly remote rural communities, where the health infrastructure is basic and accessibility complicated [4].
Until recently, the chemotherapy of HAT relied on only five drugs, according to disease stage and parasite subspecies. This was unsatisfactory because the clinically available drugs had limitations such as toxicity, resistance, high cost, and parenteral administration [5]. The recent approval of fexinidazole as a new oral drug for both stages of g-HAT greatly facilitates the treatment and will increase the coverage [6,7]. The current reference treatment is available for free in Angola. Nevertheless, a previous ethnobotanical study reporting the use of local herbal remedies against sleeping sickness pointed out that 40% of the infected patients had recoursed first to herbal remedies before receiving the medical reference treatment [8]. Therefore, the investigation of herbal remedies is of high practical relevance. There have been several reports on the antitrypanosomal activity of traditionally used African medicinal plants [9,10,11,12,13,14,15,16,17,18,19]; this is the first such study from Angola.
The laboratory results demonstrate that the medicinal plants in use to treat HAT possess antitrypanosomal activity. Bioassay-guided fractionation led to the identification of eight active molecules. Furthermore, the study provides evidence for the antitrypanosomal potential of a local preparation made of B. schreberi and N. lotus in the management of sleeping sickness in Angola.
2. Results and Discussion
2.1. Selection of the candidate plants
In a previous ethnobotanical study, 30 species of medicinal plants had been identified in the management of sleeping sickness in Angola [8]. We selected 9 species for further pharmaco-chemical investigation. The plants were selected based on four criteria: the Use Report (UR), the correlation between traditional reported preparation and clinical data, the quality of the narrative content, and the novelty of the plant. The selected plants are summarized in Table 1.
2.2. Screening of extracts against Trypanosoma brucei rhodesiense
A total of 122 extracts were prepared from different parts of the nine plant species. Each plant part was extracted consecutively with hexane, dichloromethane, ethanol, methanol, and water. The extracts were tested for their in vitro growth inhibition (GI) activity against Trypanosoma brucei rhodesiense bloodstream forms at a concentration of 20 µg/ml. Of the 122 extracts, 16 showed a strong activity (GI of 91% - 100%), 13 extracts a marked activity (71% - 90% GI), 14 extracts a moderate activity (51% - 70% GI), 19 extracts a weak activity (31% - 50% GI) and 60 extracts were inactive (GI <30%). A detailed description of the plant species, the parts extracted, solvent, extraction yield, and percentage of growth inhibition (GI%) is given in Supplementary Table S1.
Only one of the nine investigated plants lacked inhibitory activity, P. schweinfurthii, whereas all other plants demonstrated at least one extract with a moderate antitrypanosomal activity. Of the nine plants, seven had already been investigated for their antitrypanosomal activity, namely B. owariensis, C. febrifuga, E. abyssinica, M. charantia, N. lotus, S. latifolius, and V. madiensis [10,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39]. To the best of our knowledge, B. schreberi (Table S1, extracts ID 96, 98, 109, 110, 111) is reported for its antitrypanosomal activity for the first time here. Previous reports of the in vitro activity of C. febrifuga (trunk bark and leaves parts) [21], S. latifolius (root parts) [32], E. abyssinica (root parts) [22,23,40] and V. madiensis (leaves) [36,37] were confirmed in this preliminary screening. An in vivo study had provided promising results with a 70% methanol extract of N. lotus, reducing the parasitemia in mice infected with T. b. brucei at a dose of 100 mg/kg/day [30]. However, here the 70% methanol extract of N. lotus only showed a moderate in vitro inhibitory activity (Table S1, extract ID 89, 114). Furthermore, the antitrypanosomal activity of M. charantia was not confirmed [21]. Several reasons can account for such discrepancies, for example different extract preparations or variation in the chemical composition of the plants. The latter was reflected by the difference in inhibitory activity within the same extract type of three different varieties of N. lotus collected from three different sites at different times (see Table S1, extracts IDs 88-94, 112-118, 110-121). Thus, N. lotus methanolic extracts IDs 89 and 114 exhibited a moderate activity, in contrast to the methanolic extract ID 121, which was inactive.
15 active extracts from seven different species displayed a growth inhibition activity >70% at 20 µg/ml and were selected for further analysis (Table 2). Aside from activity, other considerations such as polarity and plant parts were also taken into account for the selection of the extracts. The selected extracts were also tested against two other trypanosomatid pathogens, Trypanosoma cruzi and Leishmania donovani, as well as the malaria parasite Plasmodium falciparum. In vitro 50% inhibitory concentrations (IC50) and selectivity indices (SI) were determined (Table 3). In general, the extracts were more active against T. b. rhodesiense and P. falciparum than against T. cruzi and L. donovani. All the extracts had selectivity indices >1 for T. b. rhodesiense and P. falciparum. However, none of the extracts exhibited a high selectivity, which is not unusual due to the heterogeneous composition of the crude extracts. Further purification and isolation of the active constituents may highly improve the selectivity, as will also become apparent here.
The root aqueous and ethanol 80% extracts of E. abyssinica (extracts IDs 46, 47, 54) showed antitrypanosomal activity and the aqueous extract (extract ID 46) exhibited the most potent IC50 value against T. b. rhodesiense with 1.8 µg/ml. This is in agreement with Freiburghaus et al. [22], who had demonstrated similar in vitro activity for the root methanolic extracts of E. abyssinica harvested at two different periods (IC50 of 3.3 and 6.8 µg/ml vs. 4.1 µg/ml for ID 47, Table 3). Due to the several phytochemical studies already realized on this plant [28,29,41,42,43], we concentrated our efforts on B. schreberi (extracts IDs 109, 110, 111) and N. lotus (extracts IDs 91, 92), which displayed IC50 values ≤10 µg/ml against T. b. rhodesiense and P. falciparum (Table 2) and whose antitrypanosomal activity had remained mostly unexplored.
B. schreberi is a floating-leaves plant originating from North America and distributed throughout Africa, Asia and Australia. It has so far not been investigated for its antitrypanosomal activity. B. schreberi is used in a traditional preparation in combination with N. lotus in the management of sleeping sickness in Angola. Both are aquatic plants, and the invasiveness of B. schreberi makes it a competitor to N. lotus in its natural environment (Figure 4). Regarding its genus, two studies investigated the antitrypanosomal activity in the Nymphaeaceae. The first is from Nigeria and reported antitrypanosomal activity of Nymphaea odorata with an IC50 value <5 µg/ml against T. b. brucei [30]. The second demonstrated in vivo antitrypanosomal potency for N. lotus [20]. However, no active molecules responsible for this activity have been described so far from this plant.
We first selected two midrange polarity extracts for further chemical investigation: the ethanolic extract (extract ID 110, underlined in Table 3) of the leaves of B. schreberi2 (IC50 = 7.1 ± 4.6 µg/ml) and the dichloromethane extract (extract ID 116, underlined in Table 3) of the leaves and leaflets of N. lotus (IC50 = 12.2 ± 4.6 µg/ml). Then we used a semi-preparative chromatography–based bioguided activity fractionation to tentatively identify the active constituents.
2.3. Isolation of active constituents from Brasenia schreberi and Nymphaea lotus
The 80% ethanol crude extract of B. schreberi leaves (extract ID 110) was first submitted to vacuum liquid chromatography (VLC) to remove the highly polar constituents (Figure S1). The VLC methanolic fraction (BS_EE80_VLC_MeOH) had demonstrated the most promising antitrypanosomal activity, with a GI value of 84.6% at 10 µg/ml (Figure S2) and was selected for fractionation. To optimize the semi-preparative fractionation, the analytical conditions were first determined by HPLC and the conditions were then geometrically transferred to the semi-preparative HPLC with a gradient transfer method [44]. The fractions were pooled according to UV and ELSD peaks (Figure 1A and 1B). In total, 21 fractions were collected and assayed against T. b. rhodesiense. Finally, 5 fractions (F3, F6, F10, F11, F12) displayed a strong activity (GI% > 91% at 10 µg/ml), markedly stronger than the VLC methanolic extract itself (Figure 1C; Figure S2).
Fractions F3 and F11 yielded two single compounds, (1) and (6). Fractions F6, F10, F12 were further purified using semi-preparative HPLC and yielded 5 minor compounds, (2 to 5) and (7) (Figure S3). NMR and high-resolution MS analysis resulted in identification of the seven active constituents, namely, gallic acid (1) [45], methyl gallate (2) [45], 2,3,4,6 tetragalloyl-glucopyranoside (3) [46] ethyl gallate (4) [47], 1,2,3,4,6 pentagalloyl-β-glucopyranoside (5) [48], gossypetin-7-O-β-glucopyranoside (6) [49], hypolaetin-7-O-glucoside (7) [50] (Figure 3, S4-11).
Gallic acid (1) and its ester derivatives (2 to 5) are common natural polyphenols, widely present in plants and fungi. These secondary metabolites are known for a range of applications [51] and possess several activities such as antioxidant and neuroprotective [52], anti-inflammatory [53], antitumor [54,55,56,57], antibacterial [58,59,60]. Among the compounds studied, three (1, 4, and 7) had already been described from B. schreberi [61,62] as well as gossypetin, the aglycone of 6. This compound is predominantly present in the genus Hibiscus and has been isolated in many other plant species, like Drosera peltata [63] or Equisetum fluviatile [64]. However, the presence of 2, 3, 5, and 6 in the genus Brasenia is reported for the first time here.
The dichloromethane extract of the leaves and leaflets of N. lotus (extract ID 116) was fractionated by normal phase semi-preparative chromatography using the same method as described previously but in normal phase. The 62 fractions generated were combined according to their UV and ELSD peaks (Figure 2A and 2B) in eight fractions (F1-F8) and assayed against T. b. rhodesiense (Figure 2C). One active fraction (F4) demonstrated a strong activity (GI% 97.4% at 10 µg/ml). Analysis of fraction F4 revealed a single constituent structurally elucidated by NMR and high-resolution MS, and identified as a known alkenyl resorcinol (8) [65] (Figure 3). The resorcinolic lipids have been associated with plants, bacteria and fungi [66]. They are mostly found in the members of families Anacardiaceae (e.g. cashew, mango), Ginkgoaceae (e.g. Ginkgo biloba) and Graminaceae (e.g. cereals) [67]; to the best of our knowledge, occurrence in the Nymphaeaceae is reported here for the first time. Nymphaea odorata was described in a study on Nigerian medicinal plants for its activity against T. b. brucei [68]. The structures of the resorcinol (8) identified from N. lotus and the seven active constituents identified from B. schreberi (1-7) are shown in Figure 3.
2.4. Antiprotozoal activity of the identified components
Except compound (3), for which we had insufficient plant material, the identified compounds were evaluated against T. b. rhodesiense and other protozoa. Compounds (1), (2), (4) and (5) could be purchased and were assayed against T. cruzi, L. donovani and P. falciparum (Table 4). Compounds (6), (7) and (8) were assayed only against T. cruzi and L. donovani (Table 4). Ethyl gallate (4) and methyl gallate (2) had IC50 values against T. b. rhodesiense of 0.6 µg/ml and 1.1 µg/ml, respectively, as well as of 2.1 µg/ml and 3.0 µg/ml against P. falciparum. The highest antitrypanosomal activity was found for gallic acid (1) and ethyl gallate (4) with IC50 against T. b. rhodesiense of 0.5 µg/ml and 0.6 µg/ml, respectively. None of the compounds demonstrated promising activity against T. cruzi. Resorcinol alkyl (8) had an IC50 of 2.5 µg/ml against L. donovani and a moderate selectivity (SI: 5.2). The two glycosidic flavones (6) and (7) displayed similar activities across the three trypanosomatides. The glucuronate flavones were less potent than their aglycones [69], suggesting that the antitrypanosomal activity of compounds (6) and (7) could be improved by removing the glycosidic part. The gallotannin pentagalloyl glucose (5) displayed the weakest overall antiprotozoal activity.
Our findings are in agreement with the reported activity of gallic acid (1) and ethyl gallate (4) against bloodstream forms of T. b. brucei [70,71]. Gallic acid and its ester derivative (2) inhibited the sn-glycerol-3-phosphate oxidase system of T. b. brucei in vitro [72]. Another possible mechanism of action of gallic acid is via its capacity to chelate iron and deprive the parasite [73,74]. Due to the amphiphilic nature of alkyl esters (2 and 4), these compounds might disrupt the plasma membrane, leading to trypanosome death [75]. Yet another possible target is the trypanosome alternative oxidase TAO; intriguingly, T. brucei spp. aquaglyceroporin null mutants, which are resistant to the drugs melarsoprol and pentamidine, are at the same time hypersensitive to inhibitors of TAO, including octyl gallate and propyl gallate [76].
The results with amastigote T. cruzi are consistent with the previous finding that gallic acid and two of its ester derivatives (2 and 4) were inactive (IC50 >100 µM) against epimastigote T. cruzi [77]. The detected antiplasmodial activity of compounds (2) and (4) were higher than previously reported [78]. However, another study had demonstrated a strong in vitro activity against P. falciparum for methyl gallate (2) (IC50 of 2.5 ng/ml) isolated from Alectryon serratus leaves [79] and an IC50 of 1.3 ng/ml for gallic acid (1). The finding that methyl gallate (2) and ethyl gallate (4) have a higher antiplasmodial activity than gallic acid (1) itself is corroborated by a previous report [80]. The gallotannin pentagalloylglucose (5) has demonstrated several biological activities [81]. The antileishmanial activity obtained here was lower than in a previous report [82], though. The resorcinol alkyl (8) displayed an encouraging inhibitory activity when tested against axenic amastigotes of L. donovani (IC50 of 2.5 µg/ml). However, it did not demonstrate conclusive activity when tested in an intramacrophage assay (IC50 >11 µg/ml). Interestingly, an isomer of (8), 5-heptadeca-8’Z,11’Z,16-trienylresorcinol, was isolated from the mushroom Merulius incarnatus and had a similar activity against leishmania (IC50 of 3.6 µg/ml) [83] as found here. The saturation degree of the alkyl chain impacts the bioactivity, resulting in loss of activity when saturated [83] as well as the stereochemical orientation of the double bond system. In addition to the presence of unsaturation in the alkyl chain, a free phenolic hydroxyl group is required for bioactivity of resorcinol alkyls [84,85]. Besides its interesting activity against leishmania, compound (8) displayed the best activity against T. cruzi among the isolated constituents (IC50 of 9.1 µg/ml). This is in agreement with Matutino Bastos et al. (2019), who had assayed two derivates of cardol against T. cruzi trypomastigote and amastigote forms [86]. Our results, together with these previous findings, ask for further investigation on resorcinol alkyls as potential compounds against L. donovani and T. cruzi.
2.5. Active constituents in local herbal preparation
In the northern province Uíge of Angola, the ethnotaxon “Longa dia simbi” is used for the treatment of sleeping sickness in the form of a decoction. “Longa dia simbi” in the local language Kikongo means “a tray”, referring to the leaves lying as a tray on the surface of the water. “Longa dia simbi” is made of Brasenia schreberi and Nymphaea lotus. The two species are considered by local traditional healers as the same plant; B. schreberi as “female” and N. lotus as “male” (see Figure 4). To validate the potential antitrypanosomal activity of the traditional preparation, the crude extracts of the decoction of B. schreberi (the leaves) and of N. lotus (leaves and leaflets) were analyzed by Ultra High-Performance Liquid Chromatography (UHPLC-MS) to detect the presence of the previously identified active constituents.
Resorcinol alkyl, the major active component of N. lotus, was not detected in the decoction (extract ID 112, Table S1), which was to be expected given the lipophilic structure of this compound. Nevertheless, the presence of compounds (1), (4), (6) and (7) was confirmed by LC-MS and UV analysis. These four constituents can account for the observed in vitro activity (GI% value 31-50%; Table S1) of the aqueous extract of N. lotus (ID 112) against T. b. rhodensiense. The UV-MS analysis of B. schreberi decoction (extract ID 109) revealed the presence of five active components (1), (2), (5), (6) and (7). These findings confirm the first activity screening, where the decoction extract of the leaves of B. schreberi (ID 109) displayed a strong inhibitory activity against T. b. rhodesiense (GI% value >91% / see Table S1). A quantification of the identified active compounds (1), (2), (4), and (5) was realized by UHPLC-UV Single Quadrupole MS analysis using pure reference substance (Table 5).
3. Materials and Methods
3.1. Chemicals
LC/MS grade acetonitrile was obtained from VWR International, LLC, formic acid (99%) via Thommen-Furler AG from Carlo Erba Reagents. Ultrapure water was obtained from an in-house ultrapure water system from Sartorius AG. The reference compounds gallic acid (1), methyl gallate (2), and ethyl gallate (4) were purchased from Sigma-Aldrich; 1,2,3,4,6-pentagalloyl-β-glycopyranoside (5) was obtained via Lucerna-Chem from MedChem Express. The reference compounds used as positive controls for drug efficacy testing were melarsoprol (Sanofi-Aventis, received from WHO), benznidazole (Epichem Pty Ltd, received from DNDi), miltefosine (Sigma), chloroquine (Sigma), artesunate (Mepha), and podophyllotoxin (Sigma).
3.2. Plant collection and identification
The plant material has been collected in the northern province Uíge of Angola. The nine plant species were authenticated by the Center of Studies and Scientific Investigation on Botanic of the Faculty of Science from University of Agostinho Neto, Luanda, Angola. The corresponding voucher specimen were deposited at the herbarium of the Center of Studies and Scientific Investigation on Botanic (Table 1).
3.3. Extract preparation
Different types of extraction procedures were carried out on the powdered dry material. A detailed description of plant species, parts extracted, solvents, drug-solvent ratio, and extraction yields is given in Table S1.
For increasing polarity extraction, the plant material was successively extracted for 18 ± 2 h at room temperature under constant stirring with hexane, dichloromethane (DCM), methanol (MeOH), and distilled water (H2O). After filtration, the extracts were evaporated under vacuum (Büchi Rotavapor, Switzerland) and dried under nitrogen stream. The solvent-free extracts were stored at 4 °C until use.
To replicate traditional preparations, a 20-fold quantity of water in relation to plant material was used for the extraction and boiled for 15 min. The decoction was filtrated with a Büchner under vacuum or with a filter paper (Macherey-Nagel). The filtrates (AqDec) were freeze-dried and stored at 4 °C until use. Additionally, a 10% ethanolic extract (MetT) was produced by maceration at room temperature for 2 h. Filtration and drying were performed similarly as for the decoction.
For alcoholic extraction, an 80% ethanol extract (EtOH80%) was prepared by adding a ten-fold quantity of solvent in relation to plant material and extracted at room temperature for 2 h under constant agitation. Extracts were filtered through a filter paper (Machery-Nagel), concentrated on a rotavapor (Büchi, Switzerland) at 40 °C until 60 mbar, freeze-dried, and stored at 4 °C until use. In order to assess previously referenced activity of some plant species, the extraction procedure was reproduced as published (MeOH70%, AqMac, MeOH80%).
3.4. General chromatographic procedures
NMR spectroscopic data were recorded on a Bruker Avance III HD 600 MHz NMR spectrometer equipped with a QCI 5 mm Cryoprobe and a SampleJet automated sample changer (Bruker BioSpin, Rheinstetten, Germany). Chemical shifts (δ) were measured in parts per million (ppm) using the CD3OD signal as internal standard for all spectra (δH 3.31; δC 49.0), and coupling constants (J) are reported in Hz. Complete assignment was performed based on two-dimensional experiments (COSY, NOESY, HSQC and HMBC). High resolution tandem mass spectrometry (HRMS/MS) data were obtained on a Q Exactive Focus quadrupole-orbitrap mass spectrometer (Thermo Scientific, Bremen, Germany) using heated electrospray ionization (HESI-II) in the positive and negative modes. Reverse and normal phase analysis were performed on a high-performance liquid chromatography (HPLC) Agilent 1260 Infinity LC and Agilent 1100 series system, respectively, both consisting of a degasser, a mixing pump, an autosampler, and a diode array detector (DAD) (Agilent Technologies, Santa Clara, USA) connected to an evaporative light scattering detector (ELSD) Sedex LT-ELSD 85 or ELSD Sedex 55 (Sedere, Alfortville, France) to detect non-UV absorbing compounds. Fractionation of the enriched ethanolic extract of B. schreberi (BS_EE80_VLC_MeOH) and the dichloromethane extract of N. lotus (NLotus_DCM) were performed on a semi-preparative HPLC equipment (Armen modular spot prep II, Saint-Avé, France) connected to a ELSD Sedex 55 (Sedere, Alfortville, France). Sub-fractions of the VLC_MeOH extracts of B. schreberi were purified with a Shimadzu system equipped with a LC-20 A module pumps, an SPD-20 A UV/VIS detector, a 7725I Rheodyne® valve and an FRC-10 A fraction collector (Shimadzu, Kyoto, Japan).
3.5. Fractionation and isolation of active constituents
The 80% ethanolic extract of B. schreberi leaves and the dichloromethane extract of N. lotus leaves were fractionated and purified in order to isolate eight active constituents. The ethanolic extract of B. schreberi was first subjected to a vacuum liquid chromatography (VLC) to remove very polar compounds. A 500 mL sintered-glass Büchner funnel attached to a vacuum line was packed with a C18 reverse phase Zeoprep® 40-63 μm (Lobar® Merck, Darmstadt, Germany), activated with methanol (4 x 250 mL) and equilibrated with distilled water (4 x 250 mL). The dry load composed of 3.53 g of the grinded extract mixed with the same stationary phase (1:1 w/w) was then loaded uniformly on the top of the stationary phase. The sample was eluted using water (6 x 250 mL) followed by methanol (6 x 250 mL) and washed with ethyl acetate (6 x 250 mL). The water fraction was lyophilized, while the methanol and ethyl acetate fractions were evaporated, to yield BS_EE80_VLC_H2O (2.5 g), BS_EE80_VLC_MeOH (2.6 g) and BS_EE80_VLC_EtOAc (85 mg), respectively. The optimized analytical conditions for BS_EE80_VLC_MeOH were determined by HPLC as a step gradient from 5% to 14% of B in 5 min, then 14% to 30% of B in 5 min, 30% to 60% of B in 30 min and 60% to 100% of B in 5 min held during 10 min. Then, a geometrical gradient transfer was applied from analytical to semi-preparative scale using chromatographic calculations to ensure the same selectivity. The fractionation was performed on 80 mg of the extract (BS_EE80_VLC_MeOH) on a semi-preparative HPLC system (Armen modular spot prep II, Saint-Avé, France) using an Interchim C18 column (250 × 21.2 mm, 10 μm; Interchim, Montluçon, France), with water (A) and methanol (B) containing 0.1% formic acid as mobile phase. The purification was performed using the same step gradient as the analytical conditions with a flow rate fixed at 17 mL/min. The UV detection was set at 254 nm and ELSD detection was performed under the following conditions: 40 °C, 3.1 bar N2 and gain 8. The separation led to 61 fractions combined in 21 fractions according to their UV and ELSD signal (Figure 1A, 1B). All fractions were evaporated and submitted to the in vitro growth inhibition assay (Figure 1C). Fractions F3 and F11 exhibited an activity, and compounds (1) (0.7 mg) and (6) (4 mg) were identified as major compounds of these two fractions. The fractions F6, F10 and F12, which displayed an activity but could not be identified, were further purified on a Shimadzu semi-preparative equipment using a X-bridge C18 column (250 × 10 mm, 5 μm; Waters, Milford, MA, USA), with water (A) and methanol (B) containing both 0.1% formic acid as mobile phase. The purification of F6, F10 and F12 was performed using a step gradient from 17% to 25% of B in 60 min, held during 10 min. Briefly, F6 (5.4 mg), F10 (6.5 mg) and F12 (3.6 mg) were dissolved separately in 300 μL of methanol, added to a spatula of Zeoprep C18 silica (40-63 μm) and dried gently under N2 stream. The mixture was loaded in a cartridge for dry load injection according to the method developed by Queiroz et al. [91]. The flow rate was fixed at 5 mL/min. The UV detection was set at 254 nm (F12) and 280 nm (F6, F10). The separation led respectively to 27 sub-fractions for F6, 21 for F21and 12 for F12. Sub-fractions were combined according to their UV detections (Figure S3). Using this approach, compound (2) (0.1 mg) and (3) (0.1 mg) from F6, compound (4) (0.1 mg) and (5) (0.5 mg) from F10, compound (7) (0.6 mg) from F12 were isolated.
The dichloromethane extract of N. lotus was fractionated on a semi-preparative system (Armen modular spot prep II, Saint-Avé, France) using an Interchim SIHP column (21.2 x 250 mm, 10 μm; Interchim, Montluçon, France) equipped with a Universal Guard Selectivity (UGS) SI pre-column cartridge holder (3 x 6 mm i.d., 10 µm); with hexane (A) and ethyl acetate (B) as mobile phase. The purification was performed using a linear gradient from 5% to 100% of B in 40 min, held during 10 min. The flow rate was fixed at 17 mL/min, the UV detection at 280 nm. This fractionation led to 62 fractions combined in 8 fractions according to their UV detection (Figure 2A and B). Using this approach, compound (8) (1.6 mg) was isolated from F4. The fraction was evaporated and submitted to the in vitro growth inhibition assay (Figure 2C).
3.6. UHPLC_HRMS / MS analysis
UHPLC-HRMS/MS analysis was performed for the active extracts and pure compounds using a Waters® Acquity UPLC system connected to a Q Exactive Focus mass spectrometer (Thermo Scientific, Bremen, Germany) with a heated electrospray ionization (HESI-II) in the positive and negative modes. The optimized HESI-II parameters were as follows: source voltage, 3.5 kV (pos), 3.8 kV (neg); sheath gas flow rate (N2), 55 units; auxiliary gas flow rate, 15 units; spare gas flow rate, 3.0; capillary temperature, 275 °C (pos), 320 °C (neg); S-Lens RF Level, 45. The mass analyzer was calibrated using a mixture of caffeine, methionine-arginine-phenylalanine-alanine-acetate (MRFA), sodium dodecyl sulfate, sodium taurocholate and Ultramark 1621 in an acetonitrile/methanol/water solution containing 1% formic acid by direct injection. The data-dependent MS/MS events were performed on the four most intense ions detected in full scan MS (Top 3 experiment). The MS/MS isolation window width was 1 Da, and the normalized collision energy (NCE) was set to 35 units. In data-dependent MS/MS experiments, full scans were acquired at a resolution of 35 000 FWHM (at m/z 200) and MS/MS scans at 17 500 FWHM both with a maximum injection time of 50 ms. After being acquired in a MS/MS scan, parent ions were placed in a dynamic exclusion list for 2.0 sec. Separation was achieved on an Acquity BEH C18 column (2.1 × 50 mm; 1.7 µm; Waters, Milford, MA, USA) with water (A) and acetonitrile (B) as mobile phase. The temperatures in the autosampler and in the column oven were fixed at 25 and 40 °C, respectively. Separation was performed with a linear gradient from 5% to 95% of B in 7 min, held during 1 min and then 1 min isocratic step at 5% of B for column reconditioning. Injection volume was set at 2 µL, the flow rate was fixed at 0.6 mL/min. An Acquity UPLC photodiode array detector (PDA) was used to acquire PDA spectra, which were collected from 210 to 450 nm. In positive ion mode, the di-isooctyl phthalate C24H38O4 [M + H]+ ion (m/z 391.28429) was used as an internal lock mass.
3.7. HPLC-DAD-ELSD analysis
The extracts of B. schreberi were analyzed by HPLC with DAD and ELSD detection on an Interchim C18 column (250 × 4.6 mm i.d., 10 μm; Interchim, Montluçon, France) equipped with a Nova-Pak® C18 pre-column cartridge holder (4 µm, 60 Å), using a mobile phase consisting of water (A) and methanol (B) containing both 0.1% formic acid; separation was performed with a linear gradient from 5% to 100% of B in 40 min, held during 5 min; flow rate: 1 mL/min; injection volume: 10 μL. The samples were diluted in methanol to 10 mg/mL. The UV detection was recorded at 210, 254, 280 and 366 nm. ELSD conditions: 45 °C, 3.5 bar N2 and gain 8.
The DCM extract of N. lotus was analyzed by normal phase HPLC with UV and ELSD detections on a Interchim SIHP column (250 x 4.6 mm, 10 µm; Interchim, Montluçon, France) equipped with a Universal Guard Selectivity (UGS) SI pre-column cartridge holder (3 x 6 mm i.d., 10 µm) using a mobile phase consisting of hexane (A) and ethyl acetate (B); separation was performed as described in the previous paragraph except that the samples were diluted in ethyl acetate.
3.8. NMR spectroscopic data
The recorded spectroscopic data were compared with the ones available in the literature to identify unambiguously compound (1) as gallic acid [45], (2) as methyl gallate [45], (3) as a mixture of 2 tetragalloylglucose [46], (4) as ethyl gallate [47], (5) as 1,2,3,4,6-pentagalloyl-β-glucopyranoside [48], (6) as gossypetin 7-O-glucopyranoside [49], (7) as hypolaetin-7-O-glucoside [50] and (8) as an alkenyl resorcinol [65].
3.9. Quantification of active pure compounds
The analysis was performed with an UHPLC-MS (UPLC with QDa detector, Waters) equipped with an Acquity column (BEH C18 2. 1 mm x 100 mm, 1.7 µmµL with the following parameters: mobile phase water:formic acid (1000:1 v/v) (A) and acetonitrile (B); flow rate 0.3 mL/min; column temperature 35 °C; temperature of the sample chamber 15 °C; injection volume 5 µL. The gradient used was set at 1% during 2 min, then 1-5% in 1 min, then 5-15% in 9 min held during 1 min, followed by 5 min from 15 to 95% held during 2.5 min. The analysis was carried out with the QDa detector in negative mode. The cone voltage was set to -15 V, ESI Capillary: 0.81 kV and the capillary temperature to 600 °C. The quantification was done over their respective mass traces in SIR mode (selected ion recording): 169 Da (gallic acid), 183 Da (methyl gallate), 197 Da (ethyl gallate) and 469 Da (1,2,3,4,5 pentagalloyl-β-glucopyranoside).
Gallic acid was quantified with a PDA detector at 270 nm. A standard curve was used for the quantification from 0.240 - 245.7 mg/L (R2 = 0.9998). Methyl gallate, ethyl gallate and 1,2,3,4,6 pentagalloyl-β-glucopyranoside were quantified by UHPLC-MS with a QDa detector and standard curves were established at 0.004 - 0.132 mg/L (R2 = 0.9978), 0.094 - 94.080 mg/L (R2 = 0.9999), and 0.28 – 175 mg/L (R2 = 0.9993) respectively. Each sample was filtered (0.2 µm) and prepared at 1 mg/mL in distilled water.
3.10. Antiprotozoal activity and cytotoxicity testing
Growth inhibition (GI) activity against T. b. rhodesiense STIB 900 was determined as follows: in a 96-well microtiter plate, 50 µL of HMI-9 medium supplemented with 15% heat-inactived horse serum were added to each well. 10 µL of the plant extract stock solution was added to each well. Then 50 µL of bloodstream-form trypanosomes were added, adjusted with a cell counter (CASY, Schärfe System, Germany) to 4 × 104 cells/ml. Another 50 µL of HMI-9 medium supplemented with 15% heat-inactived horse serum was added to each well of the microtiter plate. The final concentration of the tested extract was 20 µL/ml. The plate was incubated at 37 °C under a 5% CO2 atmosphere for 72 h. 10 µL of Alamar blue solution (12.5 mg resazurin dissolved in 100 ml distilled water) was added to each well and the plate incubated for another 2 to 4 h. Then, the plate was read with a Spectramax Gemini XS microplate fluorometer (Molecular Devices Corporation, Sunnyvale, CA, USA) using an excitation wavelength of 530 nm and an emission wavelength of 590 nm. Fluorescence was expressed as percentage of the untreated control. A GI >91% was considered a strong inhibitory activity, between 71%-90% a marked activity, between 51%-70% a moderate activity, between 31%-50% as a weak activity, <30% as not active. IC50 determination was performed in a similar way, but with serial dilutions of the plant extract (or pure compound) covering a range from 90 to 0.123 µg/ml. IC50 values were calculated by linear interpolation selecting values above and below the 50% inhibition mark.
In vitro growth inhibitory activity of the extracts and pure compounds against T. cruzi (intracellular amastigote forms grown in L6 rat myoblasts), L. donovani (axenic amastigote forms in acidic medium or intracellular amastigotes in mouse primary macrophages), and P. falciparum (erythrocytic stages in culture) was determined as described previously [87]. Cytotoxicity against L6 cells was assessed by using a similar protocol as outlined for IC50 determination with T. b. rhodesiense, except that rat skeletal myoblasts (L6 cells) were used. Reference compounds were melarsoprol for T. b. rhodesiense, benznidazole for T. cruzi, miltefosine for L. donovani, chloroquine and artesunate for P. falciparum, and podophyllotoxin for L6 cells.
4. Conclusion
Aiming to provide preliminary safety and efficacy validation of traditional herbal preparations, we investigated the cytotoxicity and antitrypanosomal activity of different extracts from medicinal plants that are being used in Angola in the treatment of sleeping sickness. After a preliminary activity screening, 15 active extracts were retained. Two extracts of two different aquatic plants, Brasenia schreberi and Nymphaea lotus, displayed IC50 values ≤ 10 µg/ml. Interestingly, these two Nymphaeales are being used in combination in a traditional preparation for the management of sleeping sickness in the northern part of Angola. While this is the first investigation of their antitrypanosomal constituents, B. schreberi and N. lotus have been investigated for several other bioactivities such as antioxidant [88] and anti-inflammatory [61,89], antibacterial [41,42,90], antialgal [43], anti-adipogenic [62] as well as cholesterol lowering [91] and inhibition of HIV-1 reverse transcriptase [92,93].
In the present study, we report on the bioactivity-guided fractionation of the dichloromethane extract of N. lotus and the VLC methanolic extract of B. schreberi with the identification of 8 active constituents (1-8). The presence of several antitrypanosomal compounds, gallic acid, methyl gallate, ethyl gallate and 1,2,3,4,6 pentagalloyl-β-glucopyranoside in the traditional preparation made of the leaves and leaflets of B. schreberi and N. lotus provides first evidence of the potential of the local preparation in the management of sleeping sickness in Angola. However, toxicity and in vivo efficacy remain to be further investigated.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Overview of the all the plants extracts, their preparation and antitrypanosomal activity, Figure S1: ELSD chromatograms for the ethanolic extract of B. schreberi before and after VLC enrichment, Figure S2: Growth inhibition activity (%) against T. b. rhodesiense of B. schreberi leave extracts at 20 and 10 µg/mL., Figure S3 : Separation of fraction F6 (chromatogram A), F10 (chromatogram B) and F12 (chromatogram C) of VLC methanolic extract of the leaves of B. schreberi, Figure S4: 1H NMR data and spectrum of compound 1 in CD3OD at 600 MHz, Figure S5: NMR data and spectra of compound 2 in CD3OD at 600 MHz, Figure S6: 1H NMR data and spectrum of compound 3 in CD3OD at 600 MHz, Figure S7: NMR data and spectra of compound 4 in CD3OD at 600 MHz, Figure S8: NMR data and spectra of compound 5 in CD3OD at 600 MHz, Figure S9: NMR data and spectra of compound 6 in DMSO-d6 at 600 MHz, Figure S10: NMR data and spectra of compound 7 in CD3OD at 600 MHz, Figure S11: NMR data and spectra of compound 8 in CD3OD at 600 MHz, References. .
Author Contributions
Conceptualization, N.V. and P.M.; methodology, N.V., P.M., M.K., J.L.W.; validation, N.V., P.M. and J.L.W.; formal analysis N.V., M.K. and P.M.; investigation, N.V., T.B., M.R., M.C., S.K.M., M.K., S.S., E.W., K.M., L.M.; writing—original draft preparation, N.V.; writing—review and editing, P.M., T.B., M.K., E.W., E.Q., J.L.W.; visualization, N.V. and T.B.; supervision, P.M.; project administration, N.V.; funding acquisition, N.V., P.M. All authors have read and agreed to the published version of the manuscript.
Funding
We gratefully acknowledge financial support by the Swiss TPH and the R. Geigy-Stiftung.
Acknowledgments
My first thanks go to the former Director of the FOMETRA in Uíge (Angola) Sr. António Kiakubua for his role as “gate opener” and his long-lasting support. Many thanks are also due to the late Coordinator of the CATEMATA, Sr.Kitoko Mayavângwa called Papa Kitoko, for helping me building trust with the traditional practitioners. Dr. Vasilisa Pedan from ZHAW for her sound advice in extract preparation and her constant availability as well as Katrin Jedrys for valuable comments and constant encouraging words and kindness. Special thanks to Sylvan Hälg from the Swiss TPH for his support in the in vitro assays. Robin Huber for his support in conducting the dry-load fractionation. A particular thank to Dr. Sergio Wittlin for giving the opportunity to test the antiplasmodial potential of the extracts.
Conflicts of Interest
The authors declare no conflict of interest.
| 1 | Percentage given by the hosting country’s speaker at the 1st National Conference of Traditional Medicine and Complementary Practices held in Luanda in August 2012. |
| 2 | It has to be clarified that in case of B. schreberi the leaves without petiole were extracted and tested, whereas for N. lotus, leaves and petiole were tested. In both cases, the plant part is referred as “leaves”. |
References
- WHO. Stratégie de l'OMS pour la médecine traditionnelle pour 2002-2005. World Health Organization.; Genève: Organisation mondiale de la Santé: 2002.
- Büscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human African trypanosomiasis. The Lancet 2017, 390, 2397-2409. [CrossRef]
- Simarro, P.P.; Cecchi, G.; Franco, J.R.; Paone, M.; Diarra, A.; Ruiz-Postigo, J.A.; Fevre, E.M.; Mattioli, R.C.; Jannin, J.G. Estimating and mapping the population at risk of sleeping sickness. PLoS Negl Trop Dis 2012, 6, e1859. [CrossRef]
- Truc, P.; Grebaut, P.; Lando, A.; Makiadi Donzoau, F.; Penchenier, L.; Herder, S.; Geiger, A.; Vatunga, G.; Josenando, T. Epidemiological aspects of the transmission of the parasites causing human African trypanosomiasis in Angola. Ann Trop Med Parasitol 2011, 105, 261-265. [CrossRef]
- Kennedy, P.G. Clinical features, diagnosis, and treatment of human African trypanosomiasis (sleeping sickness). The Lancet Neurology 2013, 12, 186-194. [CrossRef]
- Valverde Mordt, O.; Tarral, A.; Strub-Wourgaft, N. Development and Introduction of Fexinidazole into the Global Human African Trypanosomiasis Program. Am J Trop Med Hyg 2022, 106, 61-66. [CrossRef]
- Bernhard, S.; Kaiser, M.; Burri, C.; Maser, P. Fexinidazole for Human African Trypanosomiasis, the Fruit of a Successful Public-Private Partnership. Diseases 2022, 10. [CrossRef]
- Vahekeni, N.; Neto, P.M.; Kayimbo, M.K.; Maser, P.; Josenando, T.; da Costa, E.; Falquet, J.; van Eeuwijk, P. Use of herbal remedies in the management of sleeping sickness in four northern provinces of Angola. J Ethnopharmacol 2020, 256, 112382. [CrossRef]
- Gurib-Fakim, A.; Mahomoodally, M. African Flora as potential sources of medicinal plants: towards the chemotherapy of major parasitic and other infectious diseases- a review. Jordan Journal of Biological Sciences 2013. [CrossRef]
- Ibrahim, M.A.; Mohammed, A.; Isah, M.B.; Aliyu, A.B. Anti-trypanosomal activity of African medicinal plants: a review update. J Ethnopharmacol 2014, 154, 26-54. [CrossRef]
- Schmidt, T.; Khalid, S.; Romanha, A.; Alves, T.; Biavatti, M.; Brun, R.; Da Costa, F.; De Castro, S.; Ferreira, V.; de Lacerda, M. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases-part I. Curr Med Chem 2012, 19, 2128-2175. [CrossRef]
- Ogungbe, I.; Setzer, W. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases—Part III: In-silico molecular docking investigations. Molecules 2016, 21, 1389. [CrossRef]
- Mahomoodally, M.F. Traditional Medicines in Africa: An Appraisal of Ten Potent African Medicinal Plants. Evidence-Based Complementary and Alternative Medicine 2013, 2013. [CrossRef] [PubMed]
- Simoben, C.V.; Ntie-Kang, F.; Akone, S.H.; Sippl, W. Compounds from African Medicinal Plants with Activities Against Selected Parasitic Diseases: Schistosomiasis, Trypanosomiasis and Leishmaniasis. Nat Prod Bioprospect 2018, 10.1007/s13659-018-0165-y. [CrossRef]
- Nwodo, N.J.; Ibezim, A.; Ntie-Kang, F.; Adikwu, M.U.; Mbah, C.J. Anti-Trypanosomal Activity of Nigerian Plants and Their Constituents. Molecules 2015, 20, 7750-7771. [CrossRef]
- Gehrig, S.; Efferth, T. Development of drug resistance in Trypanosoma brucei rhodesiense and Trypanosoma brucei gambiense. Treatment of human African trypanosomiasis with natural products (Review). International journal of molecular medicine 2008, 22, 411-419. [CrossRef]
- Mwangi, V.I.; Mumo, R.M.; Nyachieo, A.; Onkoba, N. Herbal medicine in the treatment of poverty associated parasitic diseases: A case of sub-Saharan Africa. Journal of herbal medicine 2017, 10, 1-7. [CrossRef]
- Tullius Scotti, M.; Scotti, L.; Ishiki, H.; Fávaro Ribeiro, F.; Marques Duarte da Cruz, R.; Pedrosa de Oliveira, M.; Jaime Bezerra Mendonça, F. Natural products as a source for antileishmanial and antitrypanosomal agents. Combinatorial chemistry & high throughput screening 2016, 19, 537-553. [CrossRef]
- Plaatjie, M.; Onyiche, T.; Legoabe, L.; Ramatla, T.; Nyembe, N.; Suganuma, K.; Thekisoe, O. Medicinal plants as potential therapeutic agents for trypanosomosis: a systematic review. Adv Trad Med 2022. [CrossRef]
- Yusuf, A.B.; Iliyasu, B.; Abubakar, A.; Onyekwelu, N.A.; Igweh, A.C.; Ojiegbu, F.N.; Bot, D.Y. Preliminary evaluation for anti-trypanosomal activity of aqueous stem bark extract of Crossopteryx febrifuga in Trypanosoma congolense-infected rats. Journal of Pharmacy & Bioresources 2005, 2, 111-115. [CrossRef]
- Mesia, G.K.; Tona, G.L.; Nanga, T.H.; Cimanga, R.K.; Apers, S.; Cos, P.; Maes, L.; Pieters, L.; Vlietinck, A.J. Antiprotozoal and cytotoxic screening of 45 plant extracts from Democratic Republic of Congo. J Ethnopharmacol 2008, 115, 409-415. [CrossRef]
- Freiburghaus, F.; Ogwal, E.N.; Nkunya, M.H.; Kaminsky, R.; Brun, R. In vitro antitrypanosomal activity of African plants used in traditional medicine in Uganda to treat sleeping sickness. Tropical Medicine & International Health 1996, 1, 765-771. [CrossRef]
- Freiburghaus, F.; Steck, A.; Pfander, H.; Brun, R. Bioassay-guided isolation of a diastereoisomer of kolavenol from Entada abyssinica active on Trypanosoma brucei rhodesiense. Journal of ethnopharmacology 1998, 61, 179-183. [CrossRef]
- Atindehou, K.K.; Schmid, C.; Brun, R.; Koné, M.; Traore, D. Antitrypanosomal and antiplasmodial activity of medicinal plants from Côte d’Ivoire. Journal of Ethnopharmacology 2004, 90, 221-227. [CrossRef]
- Nibret, E.; Ashour, M.L.; Rubanza, C.D.; Wink, M. Screening of some Tanzanian medicinal plants for their trypanocidal and cytotoxic activities. Phytother Res 2010, 24, 945-947. [CrossRef]
- Gupta, S.; Raychaudhuri, B.; Banerjee, S.; Das, B.; Mukhopadhaya, S.; Datta, S.C. Momordicatin purified from fruits of Momordica charantia is effective to act as a potent antileishmania agent. Parasitology international 2010, 59, 192-197. [CrossRef]
- Santos, K.K.; Matias, E.F.; Sobral-Souza, C.E.; Tintino, S.R.; Morais-Braga, M.F.; Guedes, G.M.; Santos, F.A.; Sousa, A.C.; Rolon, M.; Vega, C., et al. Trypanocide, cytotoxic, and antifungal activities of Momordica charantia. Pharm Biol 2012, 50, 162-166. [CrossRef]
- García, M.; Monzote, L.; Scull, R.; Herrera, P. Activity of Cuban plants extracts against Leishmania amazonensis. ISRN pharmacology 2012, 2012. [CrossRef]
- Phillips, E.A.; Sexton, D.W.; Steverding, D. Bitter melon extract inhibits proliferation of Trypanosoma brucei bloodstream forms in vitro. Exp Parasitol 2013, 133, 353-356. [CrossRef]
- Garba, M.H.; Kabiru, A.Y.; Yusuf, A.M.; Muhammad, A.H.; Lekene, B.J.; Kabir, M.; Joseph, A. In vivo trypanocidal activity of Nymphaea lotus Linn. methanol extract against Trypanosoma brucei brucei. Asian Pacific Journal of Tropical Disease 2015, 5, 808-812. [CrossRef]
- Camacho, M.d.R.; Phillipson, J.D.; Croft, S.L.; Solis, P.N.; Marshall, S.J.; Ghazanfar, S.A. Screening of plant extracts for antiprotozoal and cytotoxic activities. Journal of Ethnopharmacology 2003, 89, 185-191. [CrossRef]
- Bizimana, N.; Tietjen, U.; Zessin, K.H.; Diallo, D.; Djibril, C.; Melzig, M.F.; Clausen, P.H. Evaluation of medicinal plants from Mali for their in vitro and in vivo trypanocidal activity. J Ethnopharmacol 2006, 103, 350-356. [CrossRef]
- Igoli, J.O.; Gray, A.I.; Clements, C.J.; Mouad, H.A. Anti-Trypanosomal Activity and cytotoxicity of some compounds and extracts from Nigerian Medicinal Plants. In Phytochemicals-Bioactivities and impact on Health, IntechOpen: 2011.
- Longdet, I.; Achemu, H.; Okanlawon, C. Potentials of Methanolic Extract of N. Latifolia Stem Bark Against T. Congolense Infection in Experimental Rats. Journal of Agricultural Sciences and Policy Research 2014.
- Olanrewaju, C.A.; Idris, H.S.; Okwute, S.K. Investigation on the trypanocidal effects of aqueous extracts of Vernonia amygdalina and Nauclea latifolia in albino rats. Researcher 2014, 6, 61-69. [CrossRef]
- Nwodo, N.; Okoye, F.; Lai, D.; Debbab, A.; Kaiser, M.; Brun, R.; Proksch, P. Evaluation of the in vitro trypanocidal activity of methylated flavonoid constituents of Vitex simplicifolia leaves. BMC Complement Altern Med 2015, 15, 82. [CrossRef]
- Nwodo, N.; Agbo, M.; Brun, R. In vitro and in vivo Antitrypanosomal studies of the leaf extract of Vitex simplicifolia. Afr. J. Pharm. Res. Dev 2012, 4, 35-40.
- Ayawa, N.G.; Ramon-Yusuf, S.B.; Wada, Y.A.; Oniye, S.J.; Shehu, D.M. Toxicity study and anti-trypanosomal activities of aqueous and methanol whole plant extracts of Brillantaisia owariensis on Trypanosoma brucei-induced infection in BALB/c mice. Clin Phytoscience 2021, 7, 39-49. [CrossRef]
- Etim, I.S.; Ugwu, T.N.; Ukachukwu, C.O.; Aroh, K.U. Preliminary Evaluation of Anti-trypanosome Impact of Methanol, Alkaloid and Flavonoid Extracts of Sarcocephalus latifolius in T. brucei Infected Mice. Int J Biochem Res Rev 2022, 10.9734/IJBCRR/2022/v31i630332. [CrossRef]
- Sempombe, J.; Mugoyela, V.; Mihale, M.J.; Zacharia, A.; Ipagala, P.; Kilulya, K.F. Preliminary in vivo antitrypanosomal activity and cytotoxicity of Entada abyssinica, Securinega virosa and Ehretia amoena. East and Central African Journal of Pharmaceutical Sciences 2014, 17, 37-43.
- Akinjogunla, O.; Adegoke, A.; Udokang, I.; Adebayo-Tayo, B. Antimicrobial potential of Nymphaea lotus (Nymphaeaceae) against wound pathogens. Journal of medicinal plants Research 2009, 3, 138-141.
- Akinjogunla, O.; Yah, C.; Eghafona, N.; Ogbemudia, F. Antibacterial activity of leave extracts of Nymphaea lotus (Nymphaeaceae) on Methicillin resistant Staphylococcus aureus (MRSA) and Vancomycin resistant Staphylococcus aureus (VRSA) isolated from clinical samples. Ann Biol Res 2010, 1, 174-184.
- Elakovich, S.D.; Wooten, J.W. An examination of the phytotoxicity of the water shield, Brasenia schreberi. Journal of chemical ecology 1987, 13, 1935-1940. [CrossRef]
- Challal, S.; Queiroz, E.F.; Debrus, B.; Kloeti, W.; Guillarme, D.; Gupta, M.P.; Wolfender, J.-L. Rational and efficient preparative isolation of natural products by MPLC-UV-ELSD based on HPLC to MPLC gradient transfer. Planta medica 2015, 81, 1636-1643. [CrossRef]
- Kamatham, S.; Kumar, N.; Gudipalli, P. Isolation and characterization of gallic acid and methyl gallate from the seed coats of Givotia rottleriformis Griff. and their anti-proliferative effect on human epidermoid carcinoma A431 cells. Toxicology Reports 2015, 2, 520-529. [CrossRef]
- Cammann, J.; Denzel, K.; Schilling, G.; Gross, G.G. Biosynthesis of gallotannins: β-glucogallin-dependent formation of 1, 2, 3, 4, 6-pentagalloylglucose by enzymatic galloylation of 1, 2, 3, 6-tetragalloylglucose. Archives of biochemistry and biophysics 1989, 273, 58-63. [CrossRef]
- Leela, V.; Saraswathy, A. Isolation and characterization of phytoconstituents from Acacia leucophloea flowers (Roxb) wild. Int Res J Pharm 2013, 4, 107-109. [CrossRef]
- Zhao, W.-H.; Gao, C.-C.; Ma, X.-F.; Bai, X.-Y.; Zhang, Y.-X.J.J.o.C.B. The isolation of 1, 2, 3, 4, 6-penta-O-galloyl-beta-D-glucose from Acer truncatum Bunge by high-speed counter-current chromatography. 2007, 850, 523-527. [CrossRef]
- Yang, C.; Shi, J.-G.; Mo, S.-Y.; Yang, Y.-C.J.J.o.A.n.p.r. Chemical constituents of Pyrrosia petiolosa. Journal of Asian Natural Products Research 2003, 5, 143-150. [CrossRef]
- Zapesochnaya, G.; Pangarova, T. Hypolaetin 7-glucoside from Caryopteris monolica. Chemistry of Natural Compounds 1973, 9, 521-521. [CrossRef]
- Choubey, S.; Varughese, L.R.; Kumar, V.; Beniwal, V. Medicinal importance of gallic acid and its ester derivatives: a patent review. Pharmaceutical patent analyst 2015, 4, 305-315. [CrossRef]
- Daglia, M.; Di Lorenzo, A.; F Nabavi, S.; S Talas, Z.; M Nabavi, S. Polyphenols: well beyond the antioxidant capacity: gallic acid and related compounds as neuroprotective agents: you are what you eat! Current Pharmaceutical Biotechnology 2014, 15, 362-372. [CrossRef]
- Kroes, B.v.; Van den Berg, A.; Van Ufford, H.Q.; Van Dijk, H.; Labadie, R. Anti-inflammatory activity of gallic acid. Planta medica 1992, 58, 499-504. [CrossRef]
- Hu, H.; Lee, H.-J.; Jiang, C.; Zhang, J.; Wang, L.; Zhao, Y.; Xiang, Q.; Lee, E.-O.; Kim, S.-H.; Lü, J. Penta-1, 2, 3, 4, 6-O-galloyl-β-d-glucose induces p53 and inhibits STAT3 in prostate cancer cells in vitro and suppresses prostate xenograft tumor growth in vivo. Molecular cancer therapeutics 2008, 7, 2681-2691. [CrossRef]
- Kant, R.; Yen, C.-H.; Hung, J.-H.; Lu, C.-K.; Tung, C.-Y.; Chang, P.-C.; Chen, Y.-H.; Tyan, Y.-C.; Chen, Y.-M.A. Induction of GNMT by 1, 2, 3, 4, 6-penta-O-galloyl-beta-D-glucopyranoside through proteasome-independent MYC downregulation in hepatocellular carcinoma. Scientific reports 2019, 9, 1968. [CrossRef]
- Locatelli, C.; Filippin-Monteiro, F.B.; Creczynski-Pasa, T.B. Alkyl esters of gallic acid as anticancer agents: A review. European journal of medicinal chemistry 2013, 60, 233-239. [CrossRef]
- Lee, H.; Lee, H.; Kwon, Y.; Lee, J.-H.; Kim, J.; Shin, M.-K.; Kim, S.-H.; Bae, H. Methyl gallate exhibits potent antitumor activities by inhibiting tumor infiltration of CD4+ CD25+ regulatory T cells. The Journal of Immunology 2010, 185, 6698-6705. [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microbial Drug Resistance 2013, 19, 256-265. [CrossRef]
- Buzzini, P.; Arapitsas, P.; Goretti, M.; Branda, E.; Turchetti, B.; Pinelli, P.; Ieri, F.; Romani, A. Antimicrobial and antiviral activity of hydrolysable tannins. Mini reviews in medicinal chemistry 2008, 8, 1179-1187. [CrossRef]
- Choi, J.G.; Mun, S.H.; Chahar, H.S.; Bharaj, P.; Kang, O.H.; Kim, S.G.; Shin, D.W.; Kwon, D.Y. Methyl gallate from Galla rhois successfully controls clinical isolates of Salmonella infection in both in vitro and in vivo systems. PLoS One 2014, 9, e102697. [CrossRef]
- Legault, J.; Perron, T.; Mshvildadze, V.; Girard-Lalancette, K.; Perron, S.; Laprise, C.; Sirois, P.; Pichette, A. Antioxidant and anti-inflammatory activities of quercetin 7-O-β-D-glucopyranoside from the leaves of Brasenia schreberi. Journal of medicinal food 2011, 14, 1127-1134. [CrossRef]
- Shimoda, H.; Nakamura, S.; Hitoe, S.; Terazawa, S.; Tanaka, J.; Matsumoto, T.; Matsuda, H. Anti-adipogenic polyphenols of water shield suppress TNF-alpha-induced cell damage and enhance expression of HAS2 and HABP2 in adiponectin. Nat. Prod. Chem. Res 2014, 2, 146.
- Braunberger, C.; Zehl, M.; Conrad, J.; Fischer, S.; Adhami, H.-R.; Beifuss, U.; Krenn, L.J.J.o.C.B. LC–NMR, NMR, and LC–MS identification and LC–DAD quantification of flavonoids and ellagic acid derivatives in Drosera peltata. 2013, 932, 111-116. [CrossRef]
- Veit, M.; Beckert, C.; Höhne, C.; Bauer, K.; Geiger, H.J.P. Interspecific and intraspecific variation of phenolics in the genus Equisetum subgenus Equisetum. 1995, 38, 881-891. [CrossRef]
- Barrow, R.; Capon, R. Alkyl and alkenyl resorcinols from an Australian marine sponge, Haliclona Sp (Haplosclerida: Haliclonidae). Australian Journal of Chemistry 1991, 44, 1393-1405. [CrossRef]
- Kozubek, A.; Tyman, J.H. Resorcinolic lipids, the natural non-isoprenoid phenolic amphiphiles and their biological activity. Chemical reviews 1999, 99, 1-26. [CrossRef]
- Kozubek, A.; Zarnowski, R.; Stasiuk, M.; Gubernator, J. Natural amphiphilic phenols as bioactive compounds. Cellular and Molecular Biology Letters 2001, 6, 351-355.
- Oguntoye, S.O.; Bello, O.M.; Fasinu, P.S.; Khan, I.A.; Ali, Z.; Khan, S.I.; Usman, L.A. Evaluation of Selected Nigerian Medicinal Plants for in vitro Antiprotozoal Activity. The Natural Products Journal 2018, 8, 175-184. [CrossRef]
- Tasdemir, D.; Kaiser, M.; Brun, R.; Yardley, V.; Schmidt, T.J.; Tosun, F.; Rüedi, P.J.A.a.; chemotherapy. Antitrypanosomal and antileishmanial activities of flavonoids and their analogues: in vitro, in vivo, structure-activity relationship, and quantitative structure-activity relationship studies. Antimibrobial Agents and Chemotherapy 2006, 50, 1352-1364. [CrossRef]
- Koide, T.; Nose, M.; Inoue, M.; Ogihara, Y.; Yabu, Y.; Ohta, N. Trypanocidal effects of gallic acid and related compounds. Planta medica 1998, 64, 27-30. [CrossRef]
- Nose, M.; Koide, T.; Morikawa, K.; Inoue, M.; Ogihara, Y.; Yabu, Y.; Ohta, N. Formation of reactive oxygen intermediates might be involved in the trypanocidal activity of gallic acid. Biological and Pharmaceutical Bulletin 1998, 21, 583-587. [CrossRef]
- Grady, R.W.; Bienen, E.J.; Clarkson Jr, A.B.J.M.; parasitology, b. Esters of 3, 4-dihydroxybenzoic acid, highly effective inhibitors of the sn-glycerol-3-phosphate oxidase of Trypanosoma brucei brucei. Molecular and Biochemical Parasitology 1986, 21, 55-63. [CrossRef]
- Amisigo, C.M.; Antwi, C.A.; Adjimani, J.P.; Gwira, T.M.J.P.o. In vitro anti-trypanosomal effects of selected phenolic acids on Trypanosoma brucei. PLoS One 2019, 14, e0216078. [CrossRef]
- Andjelković, M.; Van Camp, J.; De Meulenaer, B.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R.J.F.C. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food Chemistry 2006, 98, 23-31. [CrossRef]
- Wink, M. Medicinal plants: a source of anti-parasitic secondary metabolites. Molecules 2012, 17, 12771-12791. [CrossRef]
- Jeacock, L.; Baker, N.; Wiedemar, N.; Maser, P.; Horn, D. Aquaglyceroporin-null trypanosomes display glycerol transport defects and respiratory-inhibitor sensitivity. PLoS Pathog 2017, 13, e1006307. [CrossRef]
- Andréo, R.; Regasini, L.O.; Petrônio, M.S.; Chiari-Andréo, B.G.; Tansini, A.; Silva, D.H.S.; Cicarelli, R.M.B. Toxicity and loss of mitochondrial membrane potential induced by alkyl gallates in trypanosoma cruzi. International scholarly research notices 2015, 2015: 924670. [CrossRef]
- Calderon, A.I.; Romero, L.I.; Ortega-Barria, E.; Brun, R.; Correa A, M.D.; Gupta, M.P. Evaluation of Larvicidal and in Vitro. Antiparasitic Activities of Plants in a Biodiversity Plot in the Altos de Campana National Park, Panama. Pharmaceutical Biology 2006, 44, 487-498. [CrossRef]
- Khasanah, U.; WidyaWaruyanti, A.; Hafid, A.F.; Tanjung, M.J.P.r. Antiplasmodial activity of isolated polyphenols from Alectryon serratus leaves against 3D7 Plasmodium falciparum. Pharmacognosy Research 2017, 9, S57. [CrossRef]
- Arsianti, A.; Astuti, H.; Simadibrata, D.M.; Adyasa, Z.M.; Amartya, D.; Bahtiar, A.; Tanimoto, H.; Kakiuchi, K.J.O.J.o.C. Synthesis and in Vitro Antimalarial Activity of Alkyl Esters Gallate as a Growth Inhibitors of Plasmodium Falciparum. Oriental Journal of Chemistry 2018, 34, 655. [CrossRef]
- Torres-Leon, C.; Ventura-Sobrevilla, J.; Serna-Cock, L.; Ascacio-Valdes, J.A.; Contreras-Esquivel, J.; Aguilar, C.N. Pentagalloylglucose (PGG): a valuable phenolic compound with functional properties. Journal of functional foods 2017, 37, 176-189. [CrossRef]
- Kolodziej, H.; Kayser, O.; Kiderlen, A.; Ito, H.; Hatano, T.; Yoshida, T.; Foo, L. Antileishmanial activity of hydrolyzable tannins and their modulatory effects on nitric oxide and tumour necrosis factor-α release in macrophages in vitro. Planta medica 2001, 67, 825-832. [CrossRef]
- Jin, W.; Zjawiony, J.K.J.J.o.N.p. 5-Alkylresorcinols from Merulius i ncarnatus. Journal of Natural Products 2006, 69, 704-706. [CrossRef]
- Jiménez-Romero, C.; Torres-Mendoza, D.; González, L.D.U.; Ortega-Barría, E.; McPhail, K.L.; Gerwick, W.H.; Cubilla-Rios, L.J.J.o.n.p. Hydroxyalkenylresorcinols from Stylogyne turbacensis. Journal of Natural Products 2007, 70, 1249-1252. [CrossRef]
- Belmonte-Reche, E.; Martínez-García, M.; Peñalver, P.; Gomez-Perez, V.; Lucas, R.; Gamarro, F.; Pérez-Victoria, J.M.; Morales, J.C.J.E.j.o.m.c. Tyrosol and hydroxytyrosol derivatives as antitrypanosomal and antileishmanial agents. European Journal of Medicinal Chemistry 2016, 119, 132-140. [CrossRef]
- Matutino Bastos, T.; Mannochio Russo, H.; Silvio Moretti, N.; Schenkman, S.; Marcourt, L.; Gupta, M.P.; Wolfender, J.L.; Ferreira Queiroz, E.; Botelho Pereira Soares, M. Chemical Constituents of Anacardium occidentale as Inhibitors of Trypanosoma cruzi Sirtuins. Molecules 2019, 24. [CrossRef]
- Mahmoud, A.B.; Maser, P.; Kaiser, M.; Hamburger, M.; Khalid, S. Mining Sudanese Medicinal Plants for Antiprotozoal Agents. Front Pharmacol 2020, 11, 865. [CrossRef]
- Oyeyemi, I.T.; Akanni, O.O.; Adaramoye, O.A.; Bakare, A.A. Methanol extract of Nymphaea lotus ameliorates carbon tetrachloride-induced chronic liver injury in rats via inhibition of oxidative stress. Journal of basic and clinical physiology and pharmacology 2017, 28, 43-50. [CrossRef]
- Xiao, H.; Cai, X.; Fan, Y.; Luo, A. Antioxidant activity of water-soluble polysaccharides from Brasenia schreberi. Pharmacognosy magazine 2016, 12, 193. [CrossRef]
- Adelakun, K.M.; Mustapha, M.K.; Muazu, M.M.; Omotayo, O.L.; Olaoye, O. Phytochemical screening and antibacterial activities of crude extract of Nymphaea lotus (water lily) against fish pathogens. Journal of Biomedical Sciences 2015, 2, 38-42. [CrossRef]
- Kim, H.; Wang, Q.; Shoemaker, C.F.; Zhong, F.; Bartley, G.E.; Yokoyama, W.H. Polysaccharide gel coating of the leaves of Brasenia schreberi lowers plasma cholesterol in hamsters. Journal of traditional and complementary medicine 2015, 5, 56-61. [CrossRef]
- Hisayoshi, T.; Shinomura, M.; Konishi, A.; Tanaka, J.; Shimoda, H.; Hata, K.; Takahashi, S.; Yasukawa, K.J.J.o.B.M. Inhibition of HIV-1 reverse transcriptase activity by Brasenia schreberi (Junsai) components. Journal of Biological Macromolecules 2014, 14, 59-65. [CrossRef]
- Hisayoshi, T.; Shinomura, M.; Yokokawa, K.; Kuze, I.; Konishi, A.; Kawaji, K.; Kodama, E.N.; Hata, K.; Takahashi, S.; Nirasawa, S., et al. Inhibition of the DNA polymerase and RNase H activities of HIV-1 reverse transcriptase and HIV-1 replication by Brasenia schreberi (Junsai) and Petasites japonicus (Fuki) components. J Nat Med 2015, 69, 432-440. [CrossRef]
Figure 1.
Semi-preparative HPLC chromatogram of the enriched methanolic extract of B. schreberi leaves with the collected fractions (F1 to F21) and the seven constituents 1 to 7. The separation of the components was detected by UV (A) and evaporative light scattering detectors (ELSD, B). The fractions were pooled according to UV and ELSD peaks. (C) Inhibitory activity of the VLC methanolic fractions against T. b. rhodesiense at 10 µg/ml. Five fractions (F3, F6, F10, F11, F12; green) displayed a strong activity (GI >91%). BS_EE80_VLC_MeOH: enriched VLC methanolic extract of the ethanolic extract of B. schreberi (80%). Control: melarsoprol at 0.072 µg/ml.
Figure 1.
Semi-preparative HPLC chromatogram of the enriched methanolic extract of B. schreberi leaves with the collected fractions (F1 to F21) and the seven constituents 1 to 7. The separation of the components was detected by UV (A) and evaporative light scattering detectors (ELSD, B). The fractions were pooled according to UV and ELSD peaks. (C) Inhibitory activity of the VLC methanolic fractions against T. b. rhodesiense at 10 µg/ml. Five fractions (F3, F6, F10, F11, F12; green) displayed a strong activity (GI >91%). BS_EE80_VLC_MeOH: enriched VLC methanolic extract of the ethanolic extract of B. schreberi (80%). Control: melarsoprol at 0.072 µg/ml.
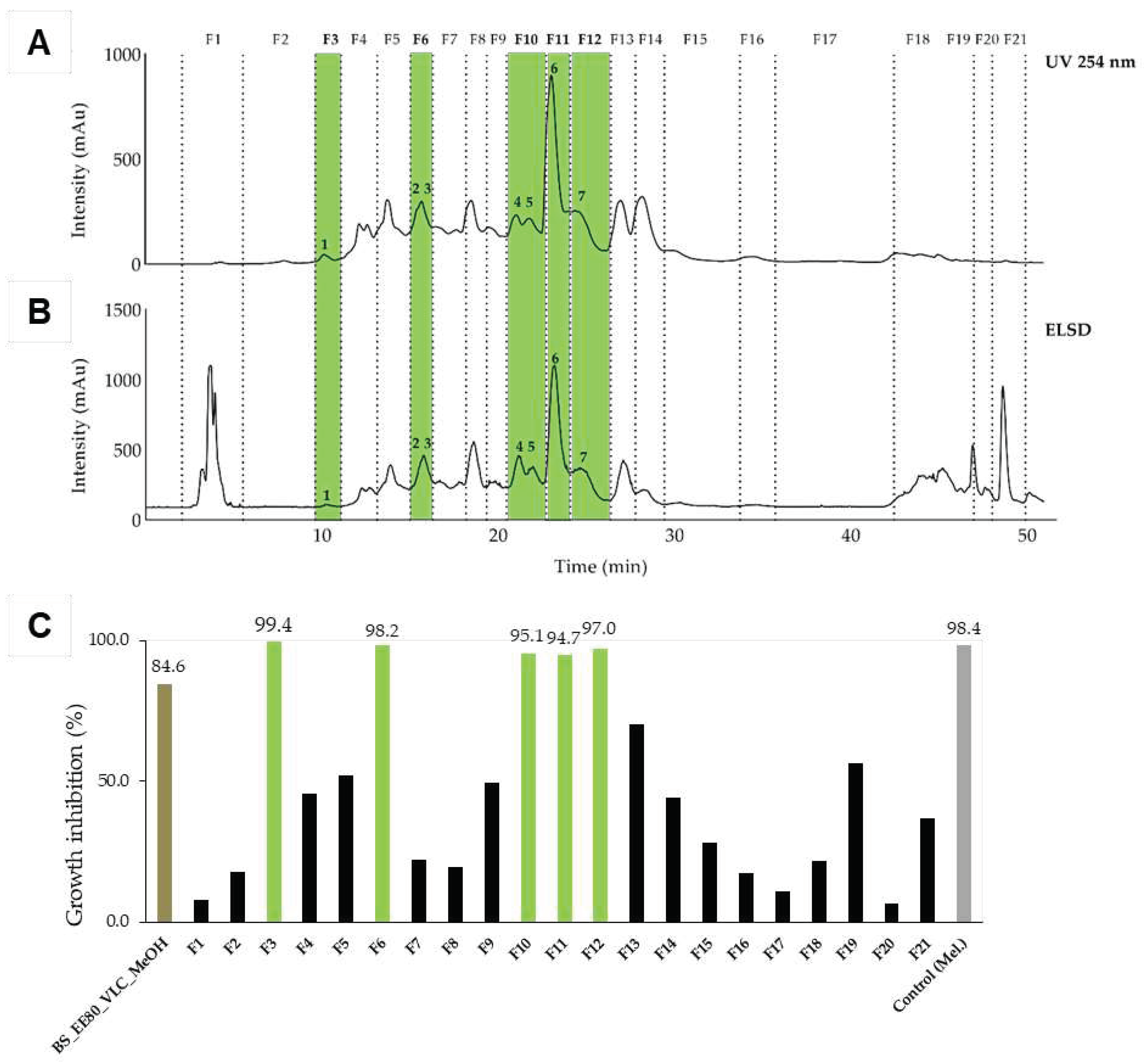
Figure 2.
Semi-preparative HPLC chromatogram of the dichloromethane extract of N. lotus highlighting the collected fractions F1 to F8 and the active constituent 8. The separation of the components was detected by UV (A) and evaporative light scattering detectors (ELSD, B). (C) Inhibitory activity against T. b. rhodesiense at 10 µg/ml of the fractions. Only one fraction (F4, green) displayed a strong activity (97%). Legend: NLotus_DCM: dichloromethane extract of N. lotus. Control: melarsoprol at 0.072 µg/ml.
Figure 2.
Semi-preparative HPLC chromatogram of the dichloromethane extract of N. lotus highlighting the collected fractions F1 to F8 and the active constituent 8. The separation of the components was detected by UV (A) and evaporative light scattering detectors (ELSD, B). (C) Inhibitory activity against T. b. rhodesiense at 10 µg/ml of the fractions. Only one fraction (F4, green) displayed a strong activity (97%). Legend: NLotus_DCM: dichloromethane extract of N. lotus. Control: melarsoprol at 0.072 µg/ml.
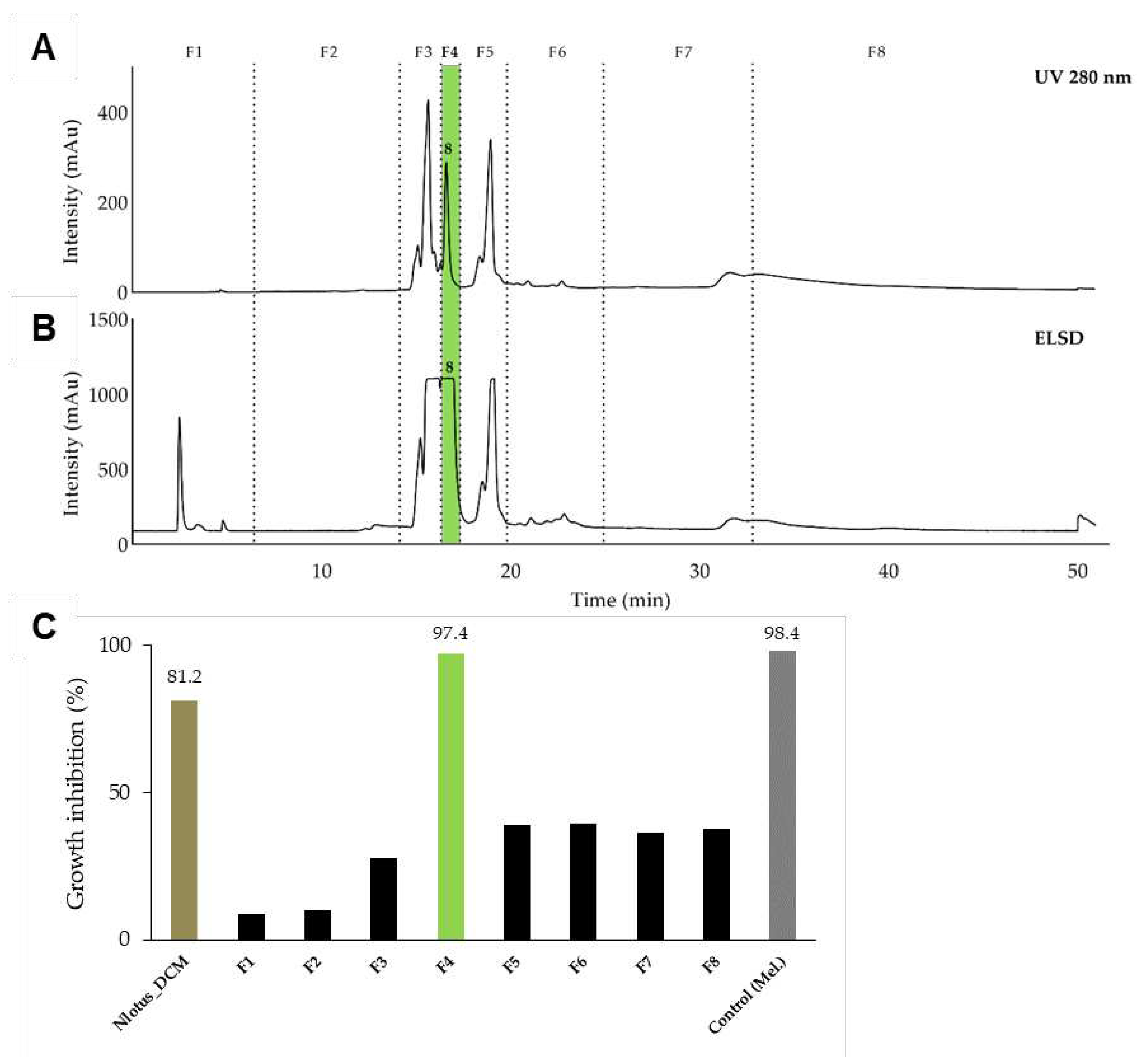
Figure 3.
Structure of the identified compounds: gallic acid (1), methyl gallate (2), 2,3,4,6 tetragalloyl-glucopyranoside (3), ethyl gallate (4), 1,2,3,4,6 pentagalloyl-β-glucopyranoside (5), gossypetin-7-O-β-glucopyranoside (6), hypolaetin-7-O-glucoside (7), 5-[(8Z,11Z,14Z)-heptadeca-8,11,14-trienyl] resorcinol (8).
Figure 3.
Structure of the identified compounds: gallic acid (1), methyl gallate (2), 2,3,4,6 tetragalloyl-glucopyranoside (3), ethyl gallate (4), 1,2,3,4,6 pentagalloyl-β-glucopyranoside (5), gossypetin-7-O-β-glucopyranoside (6), hypolaetin-7-O-glucoside (7), 5-[(8Z,11Z,14Z)-heptadeca-8,11,14-trienyl] resorcinol (8).
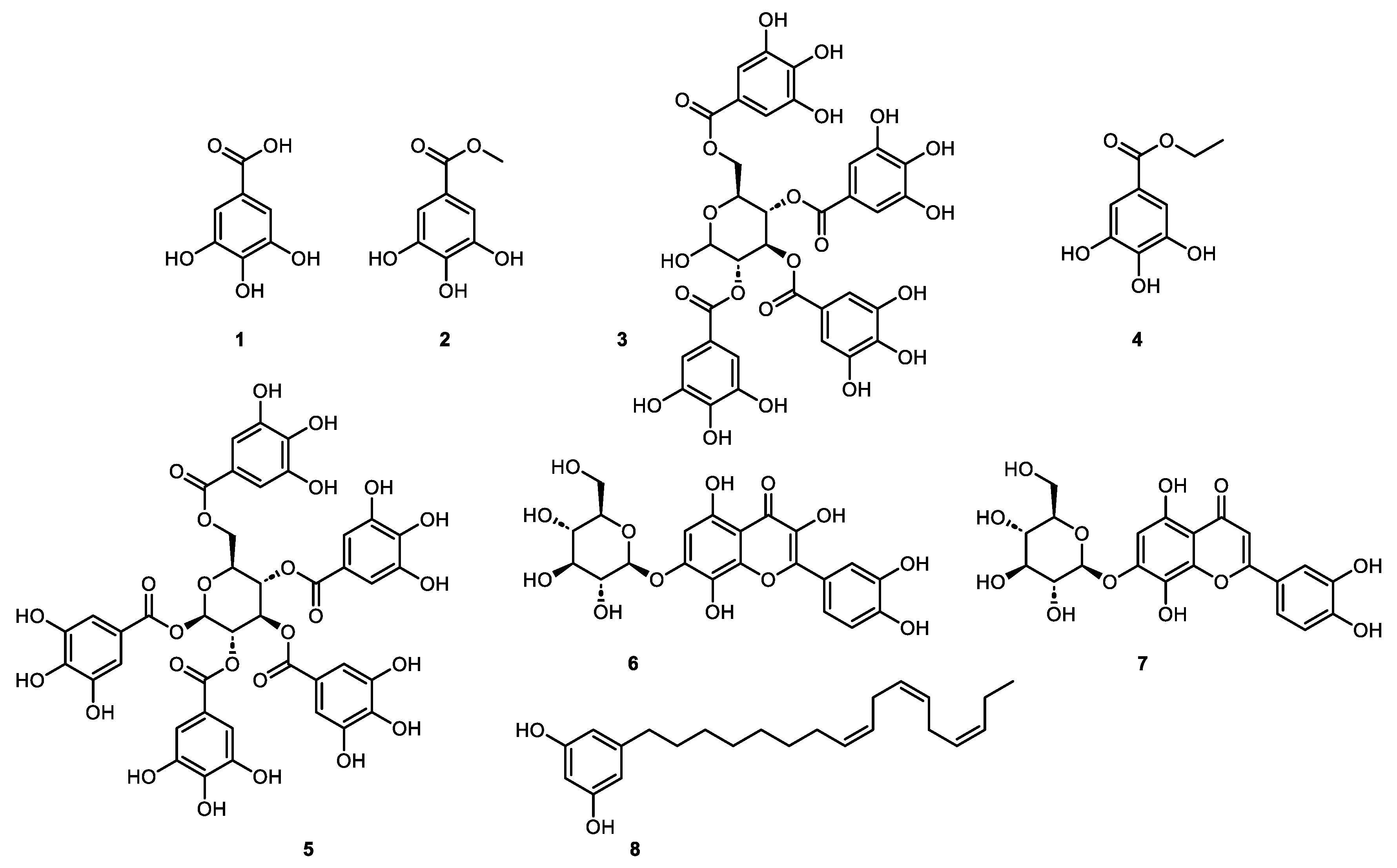
Figure 4.
Brasenia schreberi (left) and Nymphaea lotus (right) in their natural environment in Angola, province of Uíge. Insets show the leaves (B, B. schreberi left and N. lotus right) and flowers (A, B. schreberi "female"; C, N. lotus "male"). The two species are collected together, prepared as a decoction, and administered in the management of sleeping sickness.
Figure 4.
Brasenia schreberi (left) and Nymphaea lotus (right) in their natural environment in Angola, province of Uíge. Insets show the leaves (B, B. schreberi left and N. lotus right) and flowers (A, B. schreberi "female"; C, N. lotus "male"). The two species are collected together, prepared as a decoction, and administered in the management of sleeping sickness.


Table 1.
Medicinal plants from Angola analyzed in this study. Collection number of the National Botanical Center in Luanda, Angola. n.d.: not determined.
Table 1.
Medicinal plants from Angola analyzed in this study. Collection number of the National Botanical Center in Luanda, Angola. n.d.: not determined.
| Plant | Family | Collection number |
|---|---|---|
| Brillantaisia owariensis | Acanthaceae | 7925 |
| Brasenia schreberi | Cabombaceae | n.d. |
| Palisota schweinfurthii | Commelinaceae | 894 |
| Momordica charantia | Cucurbitaceae | 8591 |
| Entada abyssinica | Fabaceae | 3468 |
| Vitex madiensis | Lamiaceae | 7186 |
| Nymphaea lotus | Nymphaeaceae | 2513 |
| Crossopteryx febrifuga | Rubiaceae | 8212 |
| Sarcocephalus latifolius | Rubiaceae | 8231 |
Table 2.
The 15 most promising extracts and their activity against T. b. rhodesiense. GI, growth inhibition; Ri, rhizomes; R, roots; AeP, aerial parts; Rb, root barks; L, leaves; EtOH, ethanol; MeOH, methanol; DCM, dichloromethane.
Table 2.
The 15 most promising extracts and their activity against T. b. rhodesiense. GI, growth inhibition; Ri, rhizomes; R, roots; AeP, aerial parts; Rb, root barks; L, leaves; EtOH, ethanol; MeOH, methanol; DCM, dichloromethane.
| Extract ID | Plant name | Plant part | Solvent | GI (%) 1 |
|---|---|---|---|---|
| 46 | E. abyssinica | Ri | Aqueous | 103 |
| 47 | E. abyssinica | Ri | EtOH 80% | 101 |
| 91 | N. lotus | AeP | Hexane | 98 |
| 54 | E. abyssinica | Rb | EtOH 80% | 98 |
| 109 | B. schreberi | L | Aqueous | 99 |
| 110 | B. schreberi | L | EtOH 80% | 96 |
| 111 | B. schreberi | L | MeOH 70% | 96 |
| 92 | N. lotus | AeP | DCM | 74 |
| 115 | N. lotus | AeP | Hexane | 96 |
| 116 | N. lotus | AeP | DCM | 81 |
| 69 | V. madiensis | R | Hexane | 79 |
| 20 | C. febrifuga | L | Hexane | 85 |
| 28 | V. madiensis | L | Hexane | 96 |
| 64 | M. charantia | AeP | DCM | 72 |
| 35 | B. owariensis | L | Hexane | 96 |
1 measured at 20 µg/ml, mean of three independent replicates.
Table 3.
Antiprotozoal activities of the 15 selected active extracts, ranked by decreasing activity against T. b. rhodesiense. IC50 value and the selectivity index (SI), defined. Antitrypanosomal data and SI index represent the mean of three independent determinations and antiplasmodial data of two independent values. The IC50 values are in µg/ml. n.d: not determined. Ri = rhizomes; Rb = Root barks; AeP = aerial parts; L = leaves; Wp = leaves and stems.
Table 3.
Antiprotozoal activities of the 15 selected active extracts, ranked by decreasing activity against T. b. rhodesiense. IC50 value and the selectivity index (SI), defined. Antitrypanosomal data and SI index represent the mean of three independent determinations and antiplasmodial data of two independent values. The IC50 values are in µg/ml. n.d: not determined. Ri = rhizomes; Rb = Root barks; AeP = aerial parts; L = leaves; Wp = leaves and stems.
| Extract ID | Plantplant part | T. brucei | T. cruzi | L. donovani | P. falciparum | L6 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| IC50 | SI 1 | IC50 | SI | IC50 | SI | IC50 | SI | IC50 | ||
| 46 | E. abyssinicaRi | 1.8 | 4.5 | 14.0 | 0.6 | 29.9 | 0.3 | 6.5 | 1.2 | 6.3 |
| 47 | E. abyssinicaRi | 4.1 | 4.0 | 16.1 | 1.0 | 43.4 | 0.4 | 12.7 | 1.3 | 16.3 |
| 91 | N. lotusLi | 4.8 | 5.8 | 36.8 | 0.8 | 44.2 | 0.6 | 10.3 | 2.7 | 32.9 |
| 54 | E. abyssinicaRb | 5.1 | 3.6 | 26.4 | 0.7 | 45.8 | 0.4 | 10.4 | 1.8 | 16.0 |
| 109 | B. schreberiL | 5.9 | 2.9 | 26.7 | 0.6 | 53.0 | 0.3 | 3.5 | 4.9 | 33.8 |
| 110 | B. schreberiL | 7.1 | 4.3 | 61.5 | 0.5 | 48.1 | 0.6 | 8.1 | 3.8 | 33.8 |
| 111 | B. schreberiL | 7.9 | 4.0 | 65.9 | 0.5 | 42.4 | 0.7 | 7.5 | 4.2 | 36.0 |
| 92 | N. lotusL | 9.8 | 3.8 | 56.7 | 0.7 | 14.5 | 2.5 | 6.3 | 5.9 | 42.4 |
| 115 | N. lotusL | 11.9 | 2.5 | 45.5 | 0.6 | 20.1 | 1.5 | 14.7 | 2.0 | 34.5 |
| 116 | N. lotusL | 12.2 | 3.6 | 56.3 | 0.8 | 17.7 | 2.5 | 7.9 | 5.5 | 49.7 |
| 69 | V. madiensisR | 12.8 | 2.2 | 53.0 | 0.5 | 11.7 | 2.4 | 20.7 | 1.4 | 41.9 |
| 20 | C. febrifugaL | 13.1 | 3.5 | 64.1 | 0.7 | 46.9 | 1.0 | 21.2 | 2.2 | 47.0 |
| 28 | V. madiensisL | 13.6 | 1.7 | 42.2 | 0.6 | 23.2 | 1.0 | 23.9 | 1.0 | 22.8 |
| 64 | M. charantiaWp | 30.5 | 1.1 | 48.1 | 0.7 | 25.5 | 1.3 | 8.7 | 3.9 | 26.0 |
| 35 | B. owariensisL | 40.2 | 1.2 | 55.9 | 0.9 | 62.1 | 0.8 | >50 | n.d | 48.2 |
1 Selectivity Index, defined as the IC50 towards mammalian L6 cells divided by the IC50 towards the parasite.
Table 4.
Antiprotozoal activity of the active compounds identified from N. lotus (extract ID 116, Table 2) and B. schreberi (extract ID 110, Table 2). IC50 values are in µg/ml and represent the mean of two independent experiments.
| T. brucei | T. cruzi | L. donovani | P. falciparum | L6 | |||||
|---|---|---|---|---|---|---|---|---|---|
| IC50 | SI | IC50 | SI | IC50 | SI | IC50 | SI | IC50 | |
| Gallic acid (1) | 0.5 | 34 | 66 | 0.2 | 56 | 0.3 | >10 | n.d. | 16 |
| Methyl gallate (2) | 1.1 | 15 | 16 | 1.0 | 8.5 | 1.9 | 2.1 | 7.8 | 16 |
| Ethyl gallate (4) | 0.6 | 25 | 16 | 0.9 | 6.8 | 2.2 | 3.0 | 4.9 | 15 |
| Pentagalloyl-β-glucopyranoside (5) | 20.0 | 1.0 | 44 | 0.5 | 15 | 1.4 | 6.7 | 3.1 | 21 |
| Gossypetin-7-O-β-glucopyranoside (6) | 5.5 | 1.6 | 12 | 0.8 | 53 | 0.2 | n.d. | n.d. | 8.9 |
| Hypolaetin-7-O-glucoside (7) | 5.7 | 3.2 | 49 | 0.4 | 52 | 0.4 | n.d. | n.d. | 19 |
| Resorcinol-alkyl (8) | 5.3 | 2.5 | 9.1 | 1.4 | 2.5 | 5.2 | n.d. | n.d. | 13 |
Table 5.
Quantification of the main constituents in the decoctions of B. schreberi and of N. lotus. The values are in relation to the dried raw plant material (mg/g) and to the dry extract (mg/g).
Table 5.
Quantification of the main constituents in the decoctions of B. schreberi and of N. lotus. The values are in relation to the dried raw plant material (mg/g) and to the dry extract (mg/g).
| Active component | B. schreberi decoction | N. lotus decoction | ||
|---|---|---|---|---|
| Raw material | Extract | Raw material | Extract | |
| Gallic acid (1) | 8.8 | 50 | 5.6 | 22 |
| Methyl gallate (2) | 0.007 | 0.04 | 0.005 | 0.022 |
| Ethyl gallate (4) | n.d. | <19 ppm | n.d. | <19 ppm |
| Pentagalloyl-β-glucopyranoside (5) | 0.39 | 2.3 | 0.09 | 0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



