
Submitted:
17 December 2023
Posted:
18 December 2023
You are already at the latest version
Abstract
Spodoptera frugiperda is a significant economic pest of maize in Uganda. Many Ugandan maize farmers employ synthetic insecticides as their main form of control despite the negative impacts of these chemicals. The study aimed to determine the effect of synthetic, semi-synthetic and bio-pesticides on maize leaf damage, and abundance of S, frugiperda and its parasitoids in Wakiso and Lira districts. The experiments were laid out in an RCBD with six treatments in 2020 and nine treatments in 2021, replicated four times. The treatments with the biggest decrease in leaf damage and infestation were Amdocs and Roket®, followed by Nimbecidine. The biopesticides were not always more effective than the untreated control, though; their efficacy was often lower than that of synthetics, semi-synthetics, and botanical. We recovered one egg parasitoid, Telenomus remus, and seven egg and egg-larval parasitoids (Coccygidium luteum, Coccygidium sp., Cotesia icipe, Chelonus sp., Micranisa sp., Charops cf. diversipes, and Tachinidae sp. Among these, C. cf diversipes, Chelonus sp., C. luteum, C. icipe and Tachinidae were the most abundant. Parasitism was low, averaging 10% for eggs and 5.3% for larvae. Synthetic, the semi-synthetic and Nimbecidine often increased higher yield when compared with the untreated control. There were genrally low population of parasitoids, althoguh there were some indications of reduced population of parasitoids in plots treated with Amdocs and Roket®, while percent parasitism was not affected. In some instances, higher yields were even realized in untreated control. While considering the tactics to integrate, assessing the practices more compatible with biological practices will be necessary.
Keywords:
Cypermethrin
; Emamectin Benzoate
; botanical
; Metarhizium anisopliae
; Beauveria bassiana
; Nimbecidine
; profenofos
; yield
1. Introduction
Maize (Zea mays L.) is the world’s primary food, feed, income and industrial crop [1]. In Uganda, maize, cassava, banana and sweet potato are the four key staple crops [2]. Maize export earned Uganda 86.39 million USD in 2022 [3]. Despite its importance, the production of maize is affected by many biotic and abiotic (climatic and edaphic) constraints. Among the biotic factors are insect pests, which include Busseola fusca (Fuller 1901) (the maize stalk borer), Chilo partellus (Swinhoe in 1885) (spotted stalk borer), and various termite species which are recognized as key pests in Uganda [4,5]. Recently, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), commonly named fall armyworm (FAW), has established as an economic pest of maize too. Spodoptera frugiperda, a native to the Americas, was first reported in Africa in early 2016 [6], from where it spread very quickly, and by 2017, it was found in most of sub-Saharan Africa [7]. The pest was first detected in Uganda in 2016 [8] and spread to all maize-producing regions by 2018 [8]. In Uganda, it was predicted that S. frugiperda could cause up to $193 million and 11% of losses in the maize sector [9]. In addition, S. frugiperda infestation contaminates maize with aflatoxins and fumonisins [10].
Farmers use several practices to control S. frugiperda, but using insecticides remains the most common practice [11]. Using insecticides increases production costs, endangers human health, and negatively impacts the environment and beneficial insects [12]. Furthermore, extensive use of synthetic insecticides to manage S. frugiperda has led to the development of resistance in the New World [13,14,15]) and potentially in the introduced populations [16,17,18]. The evolution of resistance is a significant biosecurity issue to contend with and necessitates the search for sustainable and effective approaches. Pesticide use in farmers’ fields also influences the population of natural enemies by altering the fitness of insect pests and natural enemies. It can negatively affect the next generations [19] or cause direct mortality. For instance, Chlorfenapyr, chlorpyrifos, and spinosad reduce the longevity of Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae) females exposed to treated host eggs [20], and Cypermethrin reduced the number of ladybird beetles [21]. There is a need to develop an integrated strategy that incorporates biorationals, including botanicals and biopesticides, which are deemed essential components to address the problems associated with managing S. frugiperda using these synthetic insecticides.
In Africa, studies have demonstrated that Metarhizium anisopliae and Beauveria bassiana [22,23], and neem oil [24] have the potential to control S. frugiperda infestation in maize. Additionally, several parasitoid species have been reported to attack S. frugiperda in its native range [25] and its invasive range in Africa [26,27,28,29,30,31]. Some of these are targeted for production and release to boost S. frugiperda control in African smallholder farming systems. However, the effectiveness of the biorationals and their effects on the abundance of parasitoids has received limited attention in Africa. No one has yet conducted studies on the influence of synthetic, semi-synthetic and biorationals insecticides on the abundance of parasitoids and parasitism of S. frugiperda in Uganda. In Ghana, parasitism was reduced in some fields treated with Emamectin benzoate [32]. Similarly, Metarhizium anisopliae ICIPE 7, M. anisopliae ICIPE 41 and M. anisopliae ICIPE 78 reduced the emergence of C. icipe and parasitism of S. frugiperda by the same parasitoid in Kenya [33,34]. This is an indication that although biopesticides are considered more ecofriendly, they may have detrimental effects on natural enemies. This study, therefore, aimed to; 1) determine the abundance and damage of S. frugiperda under different classes of insecticides and 2) establish the species composition and abundance of parasitoids associated with S. frugiperda under different classes of insecticides in the field.
2. Materials and Methods
2.1. Study Sites
2.1.1. The Experiment Was Conducted at the National Crops Resources Research
Institute (NaCRRI) Namulonge and Ngetta Zonal Agricultural Research and Development Institute (ZARDI) located in Lira and Wakiso districts. These sites were chosen because they lie in different agroecological zones. The NaCRRI has a bimodal rainfall and lies at 0.52111° N of the Equator and 32.62685° E, and at an altitude of 1,200 m above sea level. It receives annual rainfall between 800 and 1,100 mm, with slightly humid conditions (average 65%) and an average annual temperature of 22°C. The soil is dark, reddish-brown, sandy loam, with a pH range of 5.5 to 6.2. Ngetta ZARDI is in Northern Uganda and lies at 2.294531° N and 32.92067° E, at an altitude of about 1077 m asl. It receives a bimodal rainfall pattern with a short period between the two seasons. The mean annual rainfall is about 1,300 mm. The average temperature of the district is 30°C.
2.1.2. Experimental Layout
The experiments were laid out in a Randomized Complete Block Design (RCBD) with six and nine treatments in two seasons of 2020 and 2021, respectively. Each year’s first and second rainy seasons are distinguished by the letters A and B, respectively. The treatments were replicated four times. The insecticides used in this experiment include the synthetic organophosphate insecticide (Roket®) (a.i. profenofos plus cypermethrin-based), Avermectin (Amdocs) (a.i. emamectin benzoate and abamectin), five biopesticides (Beauveria bassiana and Metarhizium anisopliae, Metarhizium anisopliae ICIPE 7, Metarhizium anisopliae ICIPE 78, General insecticide cocktail (comprising Beauveria bassiana, Isaria fumosoroseus, Lecanicillium lecani, Metarhizium anisopliae and Purporeocilium lilacanus) and a botanical (Nimbecidine) (a.i. Azadirachtin) (Table 1). Untreated control was also included for comparison. , Amdocs and Nimbecidine were sourced from the open market, while Beauveria bassiana, Metarhizium anisopliae and the General insecticide cocktail biopesticides were sourced from Milk Weed Biologicals, a producer and distributor of biological control agents in Uganda. Metarhizium anisopliae ICIPE 7 and Metarhizium anisopliae ICIPE 78 based biopesticides, which have been demonstrated to be effective, registered as Detain® and Achieve®, respectively in Kenya, Tanzania and Uganda, and recommended for managing FAW [22] were sourced from RealIPM (https://realipm.com/products/). Roket® was chosen because it is the most widely used insecticide for controlling S. frugiperda among farmers. Nimbecidine and other biopesticides were considered more human and environmentally-friendly bioproduct.
The Longe 10H maize was sourced from Nalweyo Seed Company (NASECO) and planted in 2020A, 2021A and 2021B. In 2020B, we used Longe 5 maize variety from pearl seeds because of seed unavailability.
The plant spacing was 75 cm by 25 cm, between and within rows, respectively. Each plot measured 6 m x 5 m. Two seeds of each variety were planted per hill and later thinned to one seedling per hill after emergence. The study was conducted under natural infestation, from planting up to harvest. Spraying with the different insecticides was done three times at 10-day intervals from 10 days after the emergence (DAE) of seedlings using a 20 L Farmate NS-20 knapsack sprayer. Di ammonium phosphate fertilizer was applied as a blanket treatment at planting (50 kg/ha) and top-dressed with Urea fertilizer 3-4 weeks after planting (50 kg/ha). Hand weeding was done thrice using a hoe and no irrigation was applied.
2.1.3. Data Collection
Data on leaf damage and FAW abundance were collected 10 days after seedling emergence (DAE), followed by 10-day intervals, up to 50 DAE. Data were collected from 20 plants randomly selected per plot. Data on damage severity was collected using a scale of (0-9) representing the different levels of damage, according to [35]. Each plant was examined after scoring for leaf damage/incidence, and the number of S. frugiperda eggs, larvae and adults were recorded. The yield data were obtained by weighing twenty dry cobs from the 20 sampled plants from which damage data were collected per plot. The fresh cobs were weighed, and their weight and grain moisture content were recorded. Moisture content was recorded using digital grain moisture meter (SATAKE, Moistex Model SS-7, Satake Eng. Co., Tokyo, Japan).
The larvae and egg masses of FAW were collected from each sampled plant and reared in the laboratory for the emergence of parasitoids. The egg masses were picked with a piece of maize leaf on which they were found and placed in Petri dishes with humid filter paper. Each larva was kept separately in a plastic vial and provided with maize leaf to sustain until FAW or parasitoid emergence in the laboratory. Leaves collected from 3-4 weeks-old untreated maize plants grown in a screenhouse were used to feed the larvae. The maize leaves were changed daily until a parasitoid adult or pupation of S. frugiperda larvae emerged. Egg masses were kept individually until hatching or parasitoid emergence. All parasitoid species that emerged from the samples were preserved in 90% ethanol for morphological and molecular identifications
2.1.4. Identification of Parasitoids
Morphological identification was achieved by comparing emerged parasitoids with those reared earlier in Uganda[36]. The identity of the parasitoids was confirmed using molecular techniques. This was achieved by extracting DNA from individual samples, running PCR, sequencing the PCR products, and using the Pregap4 and Gap4 sequence analysis programs within the Staden sequence analysis package to analyze trace files and assemble contigs [37]. All PCR products were then sent to Macrogen Europe B.V. (Amsterdam, The Netherlands) for purification and sequencing. The clean partial mtCOI sequences were compared with sequences in GenBank using Blast search program against the non-redundant (nr) DNA database [38], and where necessary, also compared to the International Barcode of Life (iBoL) database [39] to determine the species.
2.1.5. Data Analysis
Data collected were entered in Microsoft office excel (version 2019), cleaned, and exported to R-studio (version 1.4.17.17) using R version 4.2.1 (R core team, 2021). Data were summarized to obtain mean damage per replication, a total of egg batches, and larvae per 20 sampled plants. Data were analyzed by season for each location. Data on mean damage, damage incidence, number of larvae per 20 plants, and larval infestation were checked for normality using the Shapiro-Wilk test [40]. Since all of them were not normally distributed even after the Tukey power transformation, they were then analyzed using the Kruskal Wallis test in R studio using the “r companion package”. Mean separation was done using the DunnTest in Rstudio using the FSA package. Larval parasitism was calculated as a percentage of the total number of larvae collected that had been parasitized, while egg parasitism was calculated as a percentage of the total number of egg batches collected that had been parasitized. Since the number of egg batches and adults were low, they were not subjected to statistical analysis. Damage incidence was calculated as a proportion (%) of the total plants sampled that had leaf damage symptoms.
Grain yield (t/ha) was determined from field weight per plot and corrected to 13.5% moisture content as:
Grain yield (t ha-1) = (FW*0.8*(100-mc)*10,000) / (86.5*3.75*1000)
Where mc is the field moisture content of grain per plot, 0.8 is the shelling coefficient, 10,000 m2 is the area of a hectare, and 3.75 m2 is the area of the 20 plants per plot.
Crop yield loss was then calculated; as the difference between yield obtained in Amdocs treatment per location per season (yo) and actual yield (y), and potential yield, expressed as a percentage Walker (1983).
Linear regression analysis was performed to establish the relationship between yield and damage, damage incidence and number of FAW larvae per 20 plants.
3. Results
3.1. Leaf Damage Due to Spodopptera frugiperda in 2020A
Spodoptera frugiperda leaf damage and the incidence of damaged plants in 2020 are presented in Table 2 and Table 3. Spodoptera frugiperda leaf damage occurred in both locations and seaons (Table 2 and Table 3).
In 2020A, plots sprayed with Amdocs had the lowest leaf damage in both districts. This translated to over and above 50% reduction in leaf damage when compared with the untreated plots, which registered the highest level of damage. The level of damage in the remaining four treatments was similar for Roket®, B. bassiana and Nimbecidine in Lira. Also, the two biopesticides had similar damage levels. In Wakiso, plots treated with Roket® followed those treated with Amdocs, which had lower leaf damage, but leaf damage did not differ significantly between the rest of the treatments and control plots (Table 2).
In 2020B, the lowest leaf damage levels in Lira were observed in plots treated with Nimbecidine. In the same district, similar somewhat lower damage levels were observed in plots treated with Amdocs, Roket® and B. bassiana. The damage in M. anisopliae treated plot was higher and similar to that of the untreated control. In Wakiso, similar and the lowest damage levels were recorded in plots treated with Amdocs and Roket®, whilst the control registered the highest damage. Damage in the plots treated with the two biopesticides and Nimbecidine were similar (Table 2).
Generally, the percentage of damaged plants was very high (>85%) in both locations and seasons (Table 2). The percentage of damaged plants only differed significantly in 2020A in Lira, where Amdocs had a significantly lower percentage of damaged plants in 2020A while there were similarities in the other treatments and control.
3.2. Leaf Damage Due to Spodoptera frugiperda in 2021A
The severity of leaf damage differed significantly between treatments in all the seasons and locations (Table 3). In 2021A, Amdocs and Roket® were superior in reducing leaf damage in both districts and seasons, although Amdocs outperformed Roket® in 2021A in Lira (Table 3). In Lira, M. anisopliae ICIPE 7 and 78, M. anisopliae and B. bassiana had similar damage levels in 2021A, whilst the highest damage was recorded in the untreated control. In 2021B, however, all the other treatments and the untreated control registered similar levels of damage.
In 2021A in Wakiso, the level of damage in plots treated with Ma ICIPE 7 and 78, and Nimbecidine were similar, and lower than those registered under the General Insecticide Cocktail, M anisopliae, B. bassiana and the untreated control. In Wakiso in 2021B, except in Amdocs and Roket® treated plots, damage severity was similar in all the other remaining treatments plus the untreated control (Table 3). In 2021, the incidence of damaged plants was high (>93%) and did not differ significantly between treatments in all the seasons in the respective districts (Table 3).
3.3. Larval Abundance and Incidence of Plants Infested with Larvae of Spodoptera frugiperda in 2020
The mean number of S. frugiperda larvae per 20 plants and infestation differed significantly between treatments in one season in Lira (P < 0.01) and all seasons in Wakiso (Table 4). In 2020A, number of larvae were lower in Amdocs treated plots, but similar in all other treatments (almost more than two-fold the number registered in Amdocs treated plots). In 2020A in Wakiso, Amdocs had the lowest larval infestation. This was followed by Roket® and Nimbecidine-treated plots, which registered almost three and four times more larvae than Amdocs, respectively. The rest of the treatments did not differ significantly. In 2020B in Wakiso, save for Amdocs and Roket® treated plots, the rest of the treatments did not differ significantly in larval numbers.
The percentage of plants infested with larvae differed significantly in 2020A and not in 2020B in Lira, (Table 4); the lowest larval infestation was in Amdocs-treated plots, whilst the rest of the treatments had a significantly higher and similar percentages of infested plants (almost two-fold and above). In Wakiso, Amdocs reduced the percent of infested plants damage more than other treatments in 2020A; this was followed by Roket® (with more than twice more infested plants when compared with Amdocs), while the rest of the treatments had similar and above three times more infested plants. In 2020B in Wakiso, only Amdocs and Roket® registered the lowest percent of infested plants; the rest of the treatments plus the control had similar levels of infested plants.
3.4. Larval Abundance and Incidence of Plants Infested with Larvae of Spodoptera frugiperda in 2021
The abundance of larvae of S. Spodoptera did not differ significantly between treatments in Lira in 2021A and B (Table 5). In Wakiso, however, the abundance of larvae differed significantly between the treatments in both seasons, when Amdocs treated plots registered the lowest larval numbers. This was followed by plots treated with Roket®. Metarihzium anisopliae ICIPE 7 and 78, and Nimbecidine also registered slightly lower numbers of larvae when compared to the untreated control in 2021A. In 2021B, except in Amdocs and Roket® treated plots, the rest of the treatments did not differ significantly in the level of larval infestation.
The percent of plants infested with larvae did not differ significantly between treatments in both seasons of 2021 in Lira (Table 5). However, there were significant differences between treatments in both seasons in Wakiso (Table 5), where Amdocs and Roket® treated plots registered significantly lower percent of infested plants; the percent of infested plants did not differ significantly between the remaining treatments.
3.5. Abundance of Spodoptera frugiperda Egg Batches and Adults
The total numbers of egg batches and adults are presented in Table 6. We recovered a total of 360 egg batches from the different locations. Although not analyzed statistically, there were some patterns in the trends of observations; we recorded more egg batches in Lira and Waksio in 2020 than in 2021. There were more egg batches in the treated plots than in the untreated plots in Lira in 2020A, and in Wakiso in both seasons of 2020 (Table 6). The untreated control had higher egg batch numbers in Lira in both 2021 seasons and in 2021B in Wakiso. There were no discernible patterns in the number of adults between treatments, but Wakiso also registered the highest number of adults on three of four seasons.
3.6. Variation in Leaf Damage Severity with the Age of Maize Plants under Different Treatments
Leaf damage severity generally increased in all treatments between 10 DAE to 20 DAE, and peaked between 20 to 40 DAE (Figure 1). Amdocs and Roket® were the most superior in reducing leaf damage at the different stages of maize growth in both locations. The only exceptions were in Lira in 2020B and 2021A. In 2020B, plots treated with Nimbecidine had the least damage throughout, while in 2021A, Nimbecidine was second to Amdocs. The biopesticides had leaf damage severity falling between those of Amdocs or Roket® and the untreated control.
3.7. Variation in the Incidence of Damaged Plants with the Age of Maize Plants under Different Treatments
The variation in the percentage of damaged plants peaked either at 10 DAE or at 20 DAE, with no clear discernible pattern between the treatments in all seasons (Figure 2).
3.8. Variation in the Abundance of Larvae of Spodoptera frugiperda with the Age of Maize Plants under Different Treatments
In Lira, larval numbers were generally higher in the untreated control and peaked at 10 or 20 DAE, except in the untreated control where it peaked at 30 DAE in both seasons of 2020 (Figure 3). In Wakiso, larval abundance peaked at 20, 30 or 40 DAE depending on the treatment and season (Figure 3); in 2020A, larval abundance peaked at 20 DAE in Amdocs and Roket® treated plots, whilst the peak in the rest of the treatments occurred at 30 DAE. A similar pattern was observed in 2020B, except in the control plot where it peaked at 30 DAE. In 2021A in Wakiso, the peaks either occurred at 30 or 40 DAE, depending on the treatment. In 2021B, the highest numbers of larvae were recorded at 20 DAE in almost all treatments except the untreated control, which peaked at 20 DAE (Figure 3).
3.9. Variation in the Percent of Infested Plants with the Age of Maize Plants under Different Treatments
Similar to the pattern in larval abundance, the percentage of infested plants generally peaked 20 or 30 DAE in both districts (Figure 4). The only exception was in Wakiso in 2021A, when percent of infested plants peaked at 40 DAE in some treatments. The percentage of plants infested with larvae was generally lower in Amdocs, Roket® and occasionally Nimbecidine-treated plots, and highest in untreated plots on most sampling occasions (Figure 4).
4. Grain Yield of Maize in Different Locations and Seasons
Grain yield was not significantly different between the treatments (P > 0.05) in all seasons in both locations (Table 7). In some instances, the untreated control out yielded some insecticide-treated plots as seen in 2020B in Lira and 2021B in Wakiso district. The overall yield gain across treatments and locations averaged 9.2%. The highest gain was 38.7%.
4.1. The Relationship between Grain Yield and Leaf Damage
The regression between grain yield and leaf damage was generally insignificant, except for 2021B in Lira, where a significant negative relationship was observed between the two variables. Although not significant, negative relationships were observed in four cases, whilst non-significant positive relationships between grain yield and leaf damage were observed in two cases (Figure 5).
4.2. The Relationship between Grain Yield and Larval Numbers
Although insignificant, the relationship between grain yield and larval numbers was negative in three of four seasons in each location. A significant negative relationship was observed between grain yield and larval abundance only in Lira in 2021B (Figure 6).
4.3. The Relationship between Grain Yield and Percentage of Infested Plants
4.4. Parasitoids Recovered from Spodoptera frugiperda Eggs and Larvae
During the study, we reared eight species of parasitoids from S. frugiperda; one from eggs (Telenomus remus) and seven from the larvae (Coccigydium spp, Cotesia icipe, Chelonus sp, Micranisa sp, Charops cf. diversipes and Tachinidae sp) (Table 8). Three other species of parasitoids (Eurytomidae sp., Parapanteles sp. and Dolicogenidea sp) that were recovered from cocoons on maize plants.
4.4.1. Parasitism of Eggs of Spodoptera frugiperda
All the recovered eggs of S. frugiperda were parasitized by T. remus. Irrespective of treatment, egg parasitism was 15.2% (in 2020A), 19.2% (2020B), 5.7% (2021A) and no parasitism in 2021B in Lira. The corresponding values for Wakiso were 21.7% (2020A), 4.4% (2020B), 13.6% (2021A), and there was no parasitism in 2021B. The variation in egg parasitism across treatments was not consistent between seasons for each location (Table 9 and Table 10).
In 2020 in Lira, the highest egg parasitism levels were recorded in the untreated control in 2020A and in plots treated with Amdocs in 2020B (Table 9). In the same year in Wakiso, the highest egg parasitism level was recorded in plots spayed with Nimbecidine in 2020A and those treated with B. bassiana in 2020B. Combined over locations and seasons, egg batch parasitism averaged 10%.
In 2021, egg parasitism was only recorded in two treatments in both locations in 2021A, whereas no parasitism was recorded in both locations in 2021B (Table 10). The highest egg parasitism was recorded in a plot treated with M. anisopliae in Lira, whilst both plots that registered egg parasitism in Wakiso had 100% egg batch parasitism.
4.4.2. Parasitism of Larvae of Spodoptera frugiperda
Larval parasitism in the different locations and seasons are presented in Table 11 and Table 12. A total of seven (7) larval parasitoid species were recovered in 2020 (Table 11) and six (6) in 2021 (Table 12).
In 2020, we recovered Charops cf. diversipes, C. luteum and C. icipe from Lira. In Wakiso, we reared seven species from S. frugiperda larvae. These included C. cf. diversipes, Chelonus sp., C. luteum, Coccigydium sp, C. icipe, Micranisa sp and a Tachinidae sp. When pooled over locations and seasons, larval parasitism averaged 5.3%. Larval parasitism was 0.7% and 1% in 2020A and 2020B, respectively in Lira, and 21.1% and 4.7% in the respective seasons in Wakiso.
In 2021, we recovered four larval parasitoid species from Lira (Chelonus sp., C. icipe, Tachinidae sp and C. cf diversipes) (Table 12) and six species from Wakiso (Chelonus sp., C. luteum, Coccygidium sp, Tachinidae sp., C. cf diversipes, and C. icipe (Table 12). Irrespective of treatments, larval parasitism was 0.7% and 3% in Lira, and 7.4% and 3.4% in Wakiso in 2021A and 2021B, respectively.
5. Discussion
We conducted this study to investigate the effectiveness of different insecticides in controlling S. frugiperda infestation and damage, and to document parasitoids and parasitism of this pest in two different locations. The results of our study have shown significant differences in leaf damage among treatments in all the seasons and locations. Comparatively, plots treated with Amdocs (Emamectin Benzoate 2% + Abamectin 1%), a semi-synthetic insecticide, had significantly lower leaf damage, damage incidence, and larval infestation. This was followed closely by the plots treated with the synthetic insecticide, Roket® (Profenofos 40% + Cypermethrin 4% EC.) Nimbecidine, a botanical, was sometimes as superior as Roket®, and at times reduced leaf damage just the same as the EPFs. Control plots generally had higher damage and larval infestation levels. The EPFs performed better or at the same level as the untreated control. Comparison between treatments (insecticides). We recovered eight species of parasitoids from S. frugiperda; Telenomus remus from eggs, and seven egg/larval or larval parasitoids Coccigydium spp, Cotesia icipe, Chelonus sp, Micranisa sp, Charops cf. diversipes and Tachinidae sp. Three other parasitoid species (Eurytomidae sp., Parapanteles sp. and Dolicogenidea sp) were reared from cocoons on maize plants. Despite the diversity of parasitoids, parasitism was generally low, averaging 10% in eggs and 5.3% for larvae. Because of the low number of parasitoids encountered no discernible association could be made between parasitoid abundance/parasitism and the treatments.
The results of this study have shown that application of insecticides, especially the synthetics, semi synthetic and a botanical reduced infestation and damage, and could explain why farmers in Uganda use insecticides as a key measure to control S. frugiperda [36]. The superiority of insecticides may be attributed to their high potency to S. frugiperda. The effectiveness of Avermectins such as Emamectin benzoate to control S. frugiperda have been reported for laboratory and field studies [32,45,46]. Their effectiveness is attributed to ovicidal and larval effects and translaminar uptake, which ensures long presence in the parenchyma tissue of sprayed plants [47], resulting in an extended protection duration. Neem on the other hand is known to have antifeedant, ovicidal, insect growth regulation, repellant, and mating disruption activity [48,49]. Our findings corroborate earlier reports where emamectin benzoate caused higher mortality of S. frugiperda than organophosphates or pyrethroids [46,50,51]. Similarly, the reported effectiveness of Nimbecidine is in tandem with earlier reports that show good effectiveness of Neem in controlling S. frugiperda [32,45,52]. Although the tested biopesticides showed some reduction in leaf damage, they were not always better than the untreated control and performed below the synthetic, semi-synthetic, and botanical pesticides. This is consistent with earlier work where moderate effectiveness of biopesticides were reported when compared to synthetics and semi-synthetic insecticides.
The diversity of parasitoids reported in this study are similar to those reported in different parts of Uganda [36], with one exception. Here we report Micranisa sp being associated with S. frugiperda in Uganda for the first time. Larval parasitoids recovered included Charops cf. diversipes, Coccygidium luteum, Coccygidium sp., Cotesia icipe, Chelonus sp, Micranisa sp and an unidentified tachinid. These are consistent with previous studies on the parasitoid complex of S. frugiperda across Africa, and specifically in Uganda [36]. The parasitoids C. luteum and C. icipe have also been recorded on S. frugiperda in other East African countries [53]. In our study, we found that Wakiso had a higher parasitoid species richness than Lira where all the seven parasitoid species were recovered from larvae, while only five species were recovered from Lira. The reasons for this are not clear, but may be attributable to several factors, including landscape factors. Among the other parasitoids recovered from maize plants, Eurytomidae sp are known are parasitoids of African stem borer, Busseola fusca [54]. Parapanteles spp are reported as parasitoids of Ledipodteran pests including Isturgia exerraria infesting rooibos tea plants in South Africa [55], and S. frugiperda in Zambia [28].
Parasitism of field collected eggs and larvae were low, averaging 10% for egg batches and 5.3% for the larvae. These suggest that the levels of parasitism are still low, but nevertheless demonstrate an important role that parasitoids can play in the Integrated management of S. frugiperda. The levels of parasitism of eggs of S. frugiperda by T. remus in our study is less than the figures reported in other countries: 69.3% (in Kenya) and 58.5% (in Tanzania) [56] and 34.4% in Cameroon [57] on maize, and 78 and 64% in laboratory and fields of sorghum in Niger, respectively [29]. Similarly, the overall larval parasitism reported in this study is lower than what was reported earlier in Uganda [36] where parasitism averaged 9.2%. In Ghana, parasitism of S. frugiperda larvae ranged from 0% to 35.6% [27]. In the previous study, we did single sampling in several locations (almost 20) while the present study was conducted in only two locations. Thus, location differences may account for the differences between parasitism levels. The parasitism level reported in this study is lower than the 9.5% reported in Mozambique [58]. Among the parasitoids, Ch. cf diversipes, Chelonus sp, C. luteum, C. icipe and Tachinidae sp were more abundant, especially in Wakiso, and are candidates for mass production and release.
Our interest was also to compare parasitism of S. frugiperda under different treatments. Although the abundance of parasitoids was low or at times absent, our results of 2020A in Wakiso had generally higher number of parasitoids, which can provide some preliminary observations. Plots treated with Amdocs and Roket® recorded lower abundance and richness of parasitoids, suggesting that these treatments could potentially have negative effects on the abundance of the parasitoids. The reduction in abundance could be attributed to an indirect reduction in the abundance of the host, or detrimental effects on specific species of parasitoids. It is evident from these results that C. cf diversipes and C. luteum were not recovered from Amdocs treated plots in 2020A in Wakiso, while Chelonus sp was not recovered from Roket® treated plots. Ironically, however, percent parasitism was generally lowest in the untreated control, lending credence to the observation that percent parasitism by certain parasitoids is not affected by the different treatments. The lowest parasitism of S. frugiperda larvae by C. icipe were observed in plots treated with M. anisopliae in 2020A in Wakiso, suggesting that this EPF could have a negative effect on the parasitoid. Whereas M. anisopliae ICIPE 7 and 78 were reported to reduce parasitism by C. icipe [33,34], we did not observe this because of the low abundance of parasitoids in the seasons when we used the two products. Further observations will thus be needed to conclude on the effects of the two EPFs on parasitism of S. frugiperda by the different parasitoids.
Grain yield was generally higher in insecticide treated plots than in the untreated plots, going up to 38.7% yield advantage over the untreated control. This is evidence that the reduction in leaf damage led to higher yields as had earlier been reported by some authors such as [45]. Although this was the case, our data also showed that the untreated control at times had higher yields when compared with the treated plots despite succumbing to high damage levels. This is consistent with previous report that the relationship between foliar damage by S. frugiperda and grain yield does not usually result in significant yield reduction, due to the compensation effect [59,60]. Severe losses, however, occur when the whorl is destroyed, because of reduced photosynthetic area and thus compromised grain yield [61]. One of the reasons for the recovery is good rainfall that enhances crop vigor.
6. Conclusions
Our findings have demonstrated that synthetic and the semi pesticides reduce S. frugiperda damage and larval abundance more than biopesticides., and often result in higher yield when compared with the untreated control. The botanical Nimbecidine was also good in reducing damage and larval numbers. We recovered one egg parasitoid and seven larval parasitoids, which were directly associated with S. frugiperda. Because of the low population of parasitoids, we were unable to concretely pin the effect of treatments on their abundance and parasitism. Nevertheless, we recovered many parasitoids in one season, and the results appear to show that application of Amdocs and Roket® reduced the population of parasitoids, but not percent parasitism. It may thus be possible that a reduction in the number of parasitoids resulted because both a decrease in the number of hosts and direct effects of the pesticides.
While pesticide application reduces infestation and damage, yield did not necessarily differ significantly when there is high rainfall. Thus, it will be prudent to recommend an Integrated Pest Management Strategy, which incorporates good agronomic practices such as early planting, fertilizer application, proper weeding, and scouting and applying pesticides based on infestation/damage thresholds. Unfortunately, we do not have thresholds that are recommended for maize farmers in Uganda. However, as recommended by Nboyine et [62], about two sprays targeting VE – V5 and V6 – V12 stages of maize can be adequate to control S. frugiperda when infestation occurs. While considering the tactics to integrate, it will be necessary to assess the practices that are more compatible with biological practices. We recommend more studies on the natural enemies of S. frugiperda in Uganda, and their interaction with other control practices.
Author Contributions
Conceptualization, M.H.O.; methodology, M.H.O., S.A.A, D.K; formal analysis, M.H.O., G.O., A.L.A., D.K; investigation, M.H.O., A.L.A., G.O., S.A.A.; resources, M.H.O.; writing—original draft preparation, M.H.O., G.O., A.L.A., S.A.A., D.K., G.H., K.S.A., S.N; writing—review and editing, M.H.O., G.O., S.A.A., D.K., A.L.A., S.N., G.H., K.S.A., S.S; visualization, G.O., A.L.A.; supervision, M.H.O., S.A.A.; project administration, M.H.O., S.N., S.S; funding acquisition, M.H.O., S.N., S.S. All authors have read and agreed to the published version of the manuscript.
Funding
The authors gratefully acknowledge the financial support for this research provided by The Agriculture Cluster Development Project (Project ID No. P145037), USAID/OFDA and USAID/BHA through the project titled “Reinforcing and Expanding the Community-Based Fall Armyworm Spodoptera frugiperda (Smith) Monitoring, Forecasting for Early Warning and Timely Management to Protect Food Security and Improve Livelihoods of Vulnerable Communities - CBFAMFEW II” grant number 720FDA20IO00133, and the European Union (EU) funded project “Integrated pest management strategy to counter the threat of invasive fall armyworm to food security and Eastern Africa (FAW-IPM)” GRANT NUMBER OOD/2018402-634.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful to Onen Denish Oyaro, Peter Wasswa, Derick Oyirwoth, George Osenduru, and Florence Aryenyo, for assistance in the laboratory and field. Tonny Amodoi is acknowledged for data entry.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B.M. Global maize production, consumption and trade: Trends and R&D implications. Food Secur. 2022, 14, 1295–1319. [Google Scholar] [CrossRef]
- FAO FAOSTAT Available online: http://www.fao.org/faostat.
- BoU Composition of exports available online: https://www.bou.or.ug/bouwebsite/Statistics/.
- Matama-Kauma, T.; Schulthess, F.; Ogwang, J.A.; Mueke, J.M.; Omwega, C.O. Distribution and relative importance of Lepidopteran cereal stemborers and their parasitoids in Uganda. Phytoparasitica 2007, 35, 27–36. [Google Scholar] [CrossRef]
- Sekamatte, M. Options for integrated management of termites (Isoptera: Termitidae) in smallholder maize-based cropping systems in Uganda, Makerere University, 2001.
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in west and central Africa. PLoS One 2016, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, A.P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Day, R.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. 2017.
- Otim, M.H.; Tay, W.T.; Walsh, T.K.; Kanyesigye, D.; Adumo, S.; Abongosi, J.; Ochen, S.; Sserumaga, J.; Alibu, S.; Abalo, G.; et al. Detection of sister-species in invasive populations of the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) from Uganda. PLoS One 2018, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda J.E. Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in eastern Zimbabwe; Elsevier Ltd, 2019; Vol. 120;
- Dowd, P.F. Insect management to facilitate preharvest mycotoxin management. J. Toxicol. - Toxin Rev. 2003, 22, 327–350. [Google Scholar] [CrossRef]
- Kansiime, M.K.; Mugambi, I.; Rwomushana, I.; Nunda, W.; Lamontagne-Godwin, J.; Rware, H.; Phiri, N.A.; Chipabika, G.; Ndlovu, M.; Day, R. Farmer perception of fall armyworm (Spodoptera frugiperda J.E. Smith) and farm-level management practices in Zambia. Pest Manag. Sci. 2019, 75, 2840–2850. [Google Scholar] [CrossRef]
- Bateman, M.L.; Day, R.K.; Luke, B.; Edgington, S.; Kuhlmann, U.; Cock, M.J.W. Assessment of potential biopesticide options for managing fall armyworm (Spodoptera frugiperda) in Africa. J. Appl. Entomol. 2018, 142, 805–819. [Google Scholar] [CrossRef]
- Yu, S.J. Insecticide resistance in the fall armyworm, Spodoptera frugiperda (J. E. Smith). Pestic. Biochem. Physiol. 1991, 39, 84–91. [Google Scholar] [CrossRef]
- Carvalho, R.A.; Omoto, C.; Field, L.M.; Williamson, M.S.; Bass, C. Investigating the molecular mechanisms of organophosphate and pyrethroid resistance in the fall armyworm Spodoptera frugiperda. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Gutirrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Terán-Santofimio, H.; Rodriguez-Maciel, J.C.; Difonzo, C. Field-evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 2019, 112, 792–802. [Google Scholar] [CrossRef]
- Boaventura, D.; Martin, M.; Pozzebon, A.; Mota-Sanchez, D.; Nauen, R. Monitoring of target-site mutations conferring insecticide resistance in Spodoptera frugiperda. Insects 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, B.; Zheng, W.; Liu, C.; Zhang, D.; Zhao, S.; Li, Z.; Xu, P.; Wilson, K.; Withers, A.; et al. Genetic structure and insecticide resistance characteristics of fall armyworm populations invading China. Mol. Ecol. Resour. 2020, 20, 1682–1696. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Zhang, J.; Shen, H.; Wang, X.; Padovan, A.; Walsh, T.K.; Tay, W.T.; Gordon, K.H.J.; James, W.; Czepak, C.; et al. Whole-genome sequencing to detect mutations associated with resistance to insecticides and Bt proteins in Spodoptera frugiperda. Insect Sci. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Arnemann, J.A.A.; James, W.J.J.; Walsh, T.K.K.; Guedes, J.V.C.V.C.; Smagghe, G.; Castiglioni, E.; Tay, W.T.T. Mitochondrial DNA COI characterization of Helicoverpa armigera (Lepidoptera: Noctuidae) from Paraguay and Uruguay. Genet. Mol. Res. 2016, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.R.; Carvalho, G.A.; Moura, A.P.; Couto, M.H. .; Maia, J.B. Impact of insecticides used to control Spodoptera frugiperda (J.E. Smith) in corn on survival, sex ratio, and reproduction of Trichogramma pretiosum riley offspring. Chil. J. Agric. Res. 2013, 73, 10–11. [Google Scholar] [CrossRef]
- Ullah, M.; Ullah, F.; Khan, M.A.; Ahmad, S.; Jamil, M.; Sardar, S.; Tariq, K.; Ahmed, N. Efficacy of various natural plant extracts and the synthetic insecticide cypermethrin 25EC against Leucinodes orbonalis and their impact on natural enemies in brinjal crop. Int. J. Trop. Insect Sci. 2022, 42, 173–182. [Google Scholar] [CrossRef]
- Akutse, K.S.; Kimemia, J.W.; Ekesi, S.; Khamis, F.M.; Ombura, O.L.; Subramanian, S. Ovicidal effects of entomopathogenic fungal isolates on the invasive fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Appl. Entomol. 2019, 143, 626–634. [Google Scholar] [CrossRef]
- Ngangambe, M.H.; Mwatawala, M.W. Effects of entomopathogenic fungi (EPFs) and cropping systems on parasitoids of fall armyworm (Spodoptera frugiperda) on maize in eastern central, Tanzania. Biocontrol Sci. Technol. 2020, 30, 418–430. [Google Scholar] [CrossRef]
- Sisay, B.; Tefera, T.; Wakgari, M.; Ayalew, G.; Mendesil, E. The efficacy of selected synthetic insecticides and botanicals against fall armyworm, Spodoptera frugiperda, in Maize. Insects 2019, 10. [Google Scholar] [CrossRef]
- Molina-Ochoa, J.; Carpenter, J.E.; Heinrichs, E.A.; Foster, J.E. Parasitoids and parasites of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas and Caribbean basin: An inventory. Florida Entomol. 2003, 86, 254–289. [Google Scholar] [CrossRef]
- Abang, A.F.; Nanga, S.N.; Kuate, A.F.; Kouebou, C.; Suh, C.; Masso, C.; Saethre, M.G.; Mokpokpo Fiaboe, K.K. Natural enemies of fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in different agro-ecologies. Insects 2021, 12, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Day, R.; et al. Parasitoid complex of fall armyworm, Spodoptera frugiperda, in Ghana and Benin. Insects 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Durocher-Granger, L.; Mfune, T.; Musesha, M.; Lowry, A.; Reynolds, K.; Buddie, A.; Cafà, G.; Offord, L.; Chipabika, G.; Dicke, M.; et al. Factors influencing the occurrence of fall armyworm parasitoids in Zambia. J. Pest Sci. (2004). 2020. [Google Scholar] [CrossRef]
- Laminou, S.A.; Ba, M.N.; Karimoune, L.; Doumma, A.; Muniappan, R. Parasitism of locally recruited egg parasitoids of the fall armyworm in Africa. Insects 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kenis, M.; du Plessis, H.; Van den Berg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus remus, a candidate parasitoid for the biological control of Spodoptera frugiperda in Africa, is already present on the continent. Insects 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Amadou, L.; Baoua, I.; Malick, N.B.; Karimoune, L.; Muniappan, R. Native parasitoids recruited by the invasive fall armyworm in Niger. Indian J. Entomol. 2018, 80, 1253–1254. [Google Scholar] [CrossRef]
- Agboyi, L.K.; Nboyine, J.A.; Asamani, E.; Beseh, P.; Badii, B.K.; Kenis, M.; Babendreier, D. Comparative effects of biopesticides on fall armyworm management and larval parasitism rates in northern Ghana. J. Pest Sci. (2004). 2023. [Google Scholar] [CrossRef]
- Munywoki, J.; Omosa, L.K.; Subramanian, S.; Mfuti, D.K.; Njeru, E.M.; Nchiozem-Ngnitedem, V.-A.; Akutse, K.S. Laboratory and field performance of Metarhizium anisopliae isolate ICIPE 41 for sustainable control of the invasive fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). Agronomy 2022, 12, 2636. [Google Scholar] [CrossRef]
- Chepkemoi, J.; Fening, K.O.; Ambele, F.C.; Munywoki, J.; Akutse, K.S. Direct and indirect infection effects of four potent fungal isolates on the survival and performance of fall armyworm larval parasitoid Cotesia icipe. Sustain. 2023, 15. [Google Scholar] [CrossRef]
- Davis FM, W.W. Visual rating scales for screening whorl- stage corn for resistance to fall armyworm. Mississippi Agricultural & Forestry Experiment Station, Technical Bulletin 186, Mississippi State University, MS39762, USA. 1992. [Google Scholar] [CrossRef]
- Otim, M.H.; Aropet, S.A.; Opio, M.; Kanyesigye, D.; Opolot, H.N.; Tay, W.T. Parasitoid distribution and parasitism of the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in different maize producing regions of Uganda. Insects 2021, 12, 1–20. [Google Scholar] [CrossRef]
- Andrews, S. Sequence assembly using the Staden package (Babraham Bioinformatics). 2008, 3, 1–31.
- Sayers, E.W.; Beck, J.; Brister, J.R.; Bolton, E.E.; Canese, K.; Comeau, D.C.; Funk, K.; Ketter, A.; Kim, S.; Kimchi, A.; et al. Database resources of the National Center for Biotechnology information. Nucleic Acids Res. 2020, 48, D9–D16. [Google Scholar] [CrossRef] [PubMed]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of life data system: Barcoding. Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Telfer, A.; Young, M.; Quinn, J.; Perez, K.; Sobel, C.; Sones, J.; Levesque-Beaudin, V.; Derbyshire, R.; Fernandez-Triana, J.; Rougerie, R.; et al. Biodiversity inventories in high gear: DNA barcoding facilitates a rapid biotic survey of a temperate nature reserve. Biodivers. Data J. 2015, 3, e6313. [Google Scholar] [CrossRef]
- Alex Smith, M.; Fernández-Triana, J.L.; Eveleigh, E.; Gómez, J.; Guclu, C.; Hallwachs, W.; Hebert, P.D.N.; Hrcek, J.; Huber, J.T.; Janzen, D.; et al. DNA barcoding and the taxonomy of microgastrinae wasps (Hymenoptera, Braconidae): Impacts after 8 years and nearly 20,000 sequences. Mol. Ecol. Resour. 2013, 13, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Hrcek, J. , Miller, S.E., Quicke, D.L. and Smith, M.A. Molecular detection of trophic links in a complex insect host-parasitoid food web. Mol Ecol Resour 2011, 11, 786–794. [Google Scholar] [CrossRef]
- Jousselin, E.; Van Noort, S.; Berry, V.; Rasplus, J.; Rønsted, N.; Erasmus, J.C.; Greeff, J.M. one fig to bind them all: host conservatism in a fig wasp community unraveled by cospeciation analyses among pollinating and nonpollinating Fig wasps. Evolution (N. Y). 2008, 62, 1777–1797. [Google Scholar] [CrossRef] [PubMed]
- Babendreier, D.; Koku Agboyi, L.; Beseh, P.; Osae, M.; Nboyine, J.; Ofori, S.E.K.; Frimpong, J.O.; Attuquaye Clottey, V.; Kenis, M. The efficacy of alternative, environmentally friendly plant protection measures for control of fall armyworm, Spodoptera frugiperda, in maize. Insects 2020, 11, 240. [Google Scholar] [CrossRef]
- Deshmukh, S.; Pavithra, H.B.; Kalleshwaraswamy, C.M.; Shivanna, B.K.; Maruthi, M.S.; Mota-Sanchez, D. Field efficacy of insecticides for management of invasive fall armyworm, Spodoptera frugiperda (J.E. Smith) (lepidoptera: Noctuidae) on maize in India. Florida Entomol. 2020, 103, 221. [Google Scholar] [CrossRef]
- Jansson, R.K.; Brown, R.; Cartwright, B.; Cox, D.; Dunbar, D.M.; Dybas, R.A.; Eckel, C.; Lasota, J.A.; Mookerjee, P.K.; Norton, J.A.; et al. Emamectin Benzoate: A novel Avermectin derivative for control of Lepidopterous pests. In Proceedings of the 3rd international workshop: The management of diamondback moth and other crucifer pests; 1997; pp. 171–177. [Google Scholar]
- Williams, L.A.D.; Mansingh, A. The insecticidal and acaricidal actions of compounds from Azadirachta indica (A. Juss.) and their use in tropical pest management. Integr. Pest Manag. Rev. 1996, 1, 133–145. [Google Scholar] [CrossRef]
- Chaudhary, S.; Kanwar, R.K.; Sehgal, A.; Cahill, D.M.; Barrow, C.J.; Sehgal, R.; Kanwar, J.R. Progress on Azadirachta indica based biopesticides in replacing synthetic toxic pesticides. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Ahmed, K.S.; Idrees, A.; Majeed, M.Z.; Majeed, M.I.; Shehzad, M.Z.; Ullah, M.I.; Afzal, A.; Li, J. Synergized toxicity of promising plant extracts and synthetic chemicals against fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) in Pakistan. Agronomy 2022, 12, 1289. [Google Scholar] [CrossRef]
- Idrees, A.; Qadir, Z.A.; Afzal, A.; Ranran, Q.; Li, J. Laboratory efficacy of selected synthetic insecticides against second instar invasive fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) Larvae. PLoS One 2022, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tulashie, S.K.; Adjei, F.; Abraham, J.; Addo, E. Potential of neem extracts as natural insecticide against fall armyworm (Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae). Case Stud. Chem. Environ. Eng. 2021, 4, 100130. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Wakgari, M.; Ayalew, G.; Tefera, T. First report of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), natural enemies from Africa. J. Appl. Entomol. 2018, 142, 800–804. [Google Scholar] [CrossRef]
- Kfir, R. Parasitoids of the African stem borer, Busseola fusca (Lepidoptera: Noctuidae), in South Africa. Bull. Entomol. Res. 1995, 85, 369–377. [Google Scholar] [CrossRef]
- Valerio, A.A.; Whitefield, J.B.; Kole, M. Parapanteles rooibos, n. Sp. (Hymenoptera: Braconidae: Microgastrinae): The first record of the genus from the African continent. Zootaxa 2005, 855, 1. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Mendesil, E.; Likhayo, P.; Ayalew, G.; Mohamed, S.; Subramanian, S.; Tefera, T. Fall armyworm, Spodoptera frugiperda infestations in East Africa: Assessment of damage and parasitism. Insects 2019, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Abang, A.F.; Fotso Kuate, A.; Nanga Nanga, S.; Okomo Esi, R.M.; Ndemah, R.; Masso, C.; Fiaboe, K.K.M.; Hanna, R. Spatio-temporal partitioning and sharing of parasitoids by fall armyworm and maize stemborers in Cameroon. J. Appl. Entomol. 2020, 1–10. [Google Scholar] [CrossRef]
- Caniço, A.; Mexia, A.; Santos, L. First report of native parasitoids of fall armyworm Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in Mozambique. Insects 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Osae, M.Y.; Frimpong, J.O.; Sintim, J.O.; Offei, B.K.; Marri, D.; Ofori, S.E.K. Evaluation of different rates of Ampligo insecticide against fall armyworm (Spodoptera frugiperda (JE Smith); Lepidoptera: Noctuidae) in the coastal savannah agroecological zone of Ghana. Adv. Agric. 2022, 2022. [Google Scholar] [CrossRef]
- Hruska, A.J. Fall armyworm (Spodoptera frugiperda) management by smallholders. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 0–3. [Google Scholar] [CrossRef]
- Kebede, M.; Shimalis, T. Outbreak, distribution and management of fall armyworm, Spodoptera frugiperda J. E. Smith in Africa: The status and prospects. 2019, 1–16.
- Nboyine, J.A.; Asamani, E.; Agboyi, L.K.; Yahaya, I.; Kusi, F.; Adazebra, G.; Badii, B.K. assessment of the optimal frequency of insecticide sprays required to manage fall armyworm (Spodoptera frugiperda J. E Smith) in maize (Zea mays L.) in northern Ghana. CABI Agric. Biosci. 2022, 7, 1–11. [Google Scholar] [CrossRef]
Figure 1.
Mean leaf damage under different treatments over the days after seedling emergence.
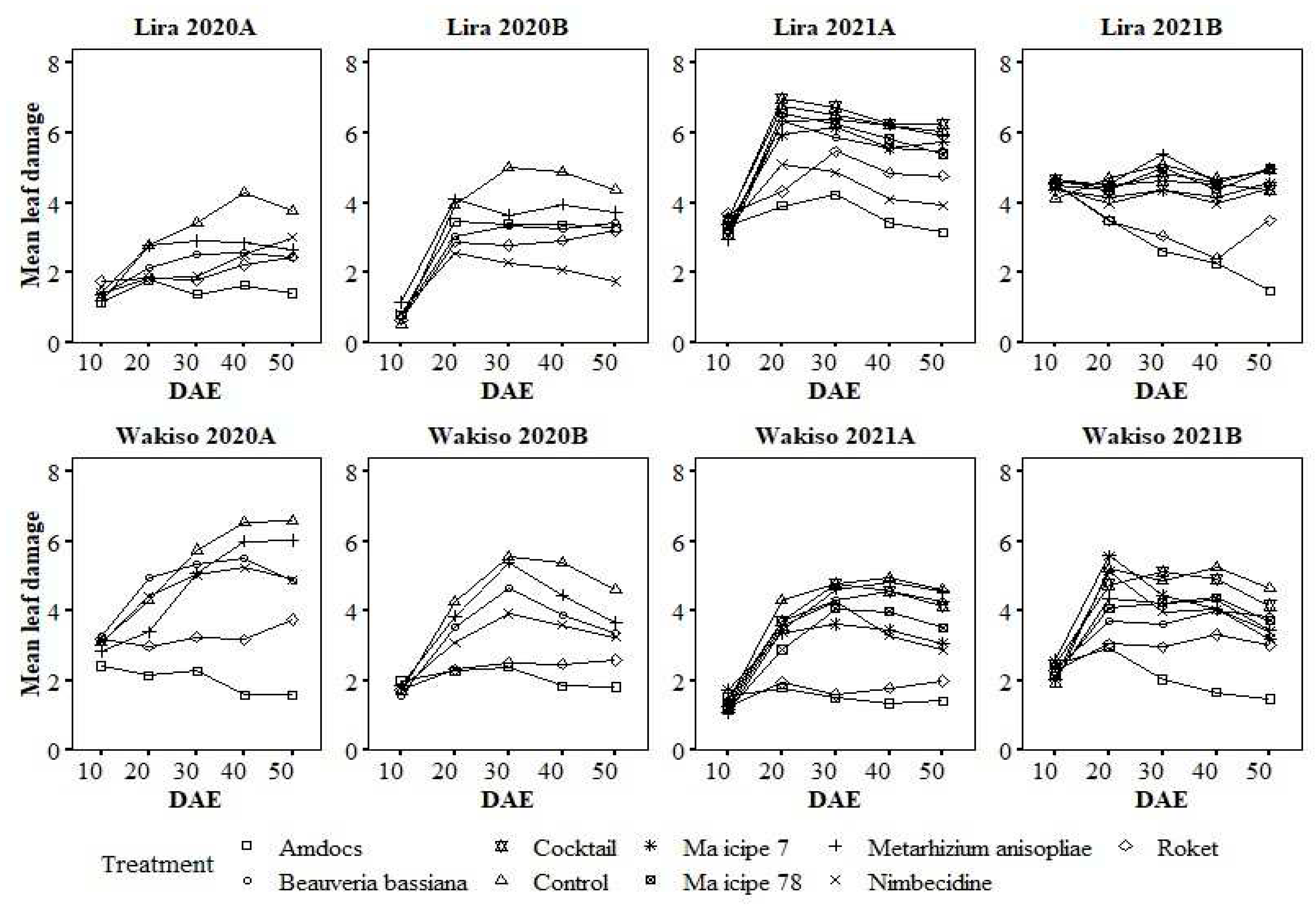
Figure 2.
Mean incidence of damaged maize plants under different treatments over the days after seedling emergence.
Figure 2.
Mean incidence of damaged maize plants under different treatments over the days after seedling emergence.
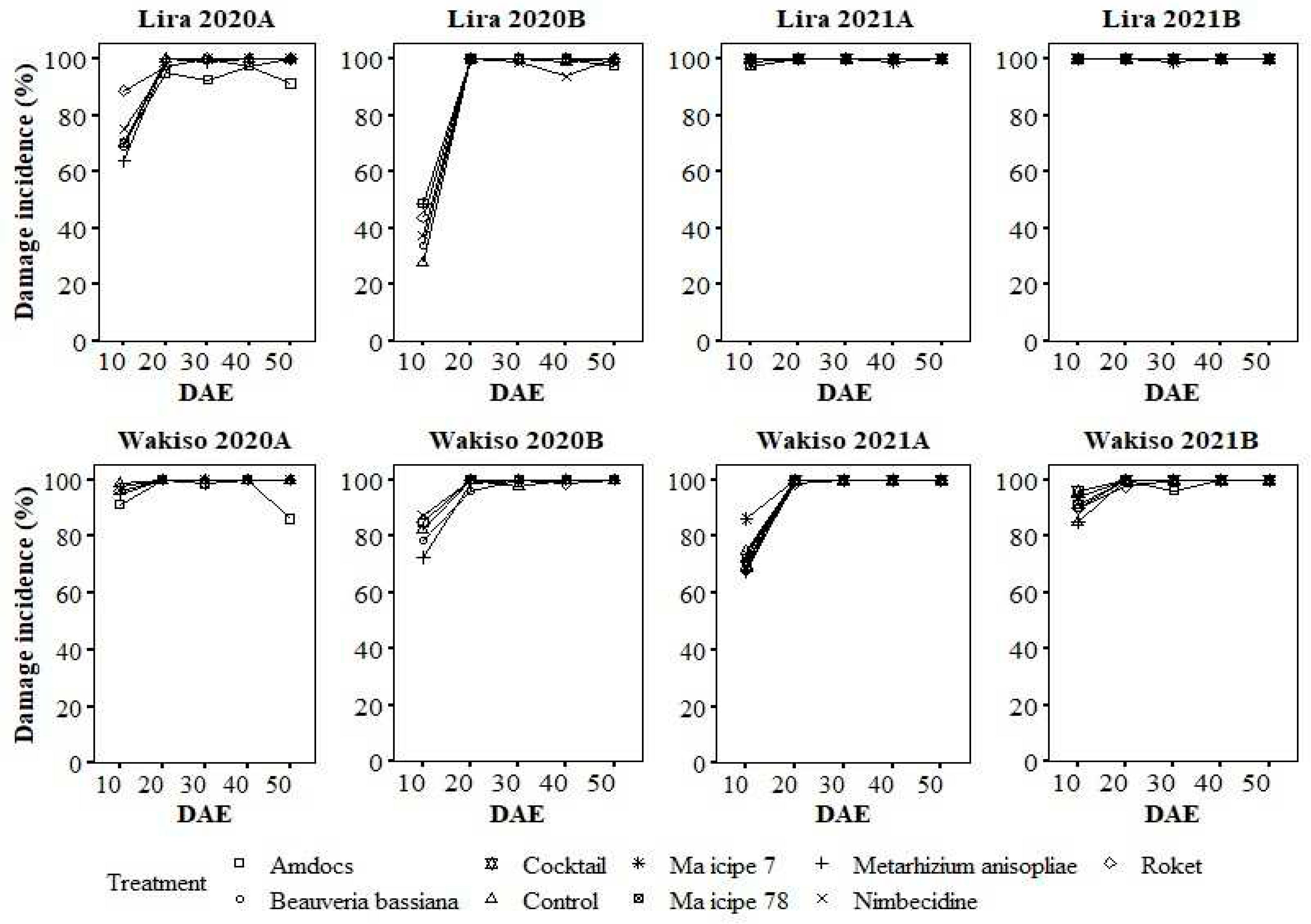
Figure 3.
Variation in the number of Spodoptera frugiperda larvae under different treatments over the different growth stages (days after seedling emergence).
Figure 3.
Variation in the number of Spodoptera frugiperda larvae under different treatments over the different growth stages (days after seedling emergence).
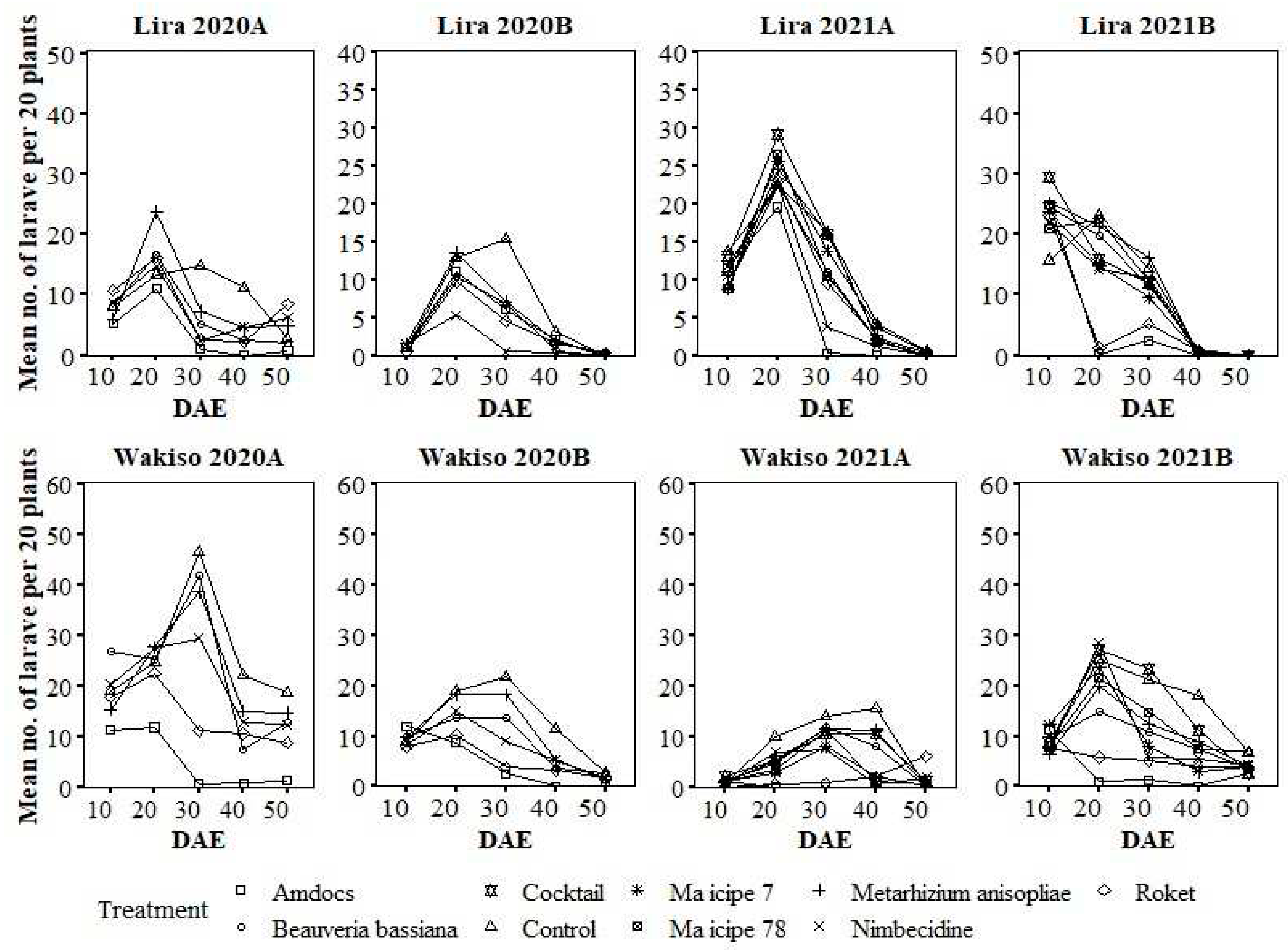
Figure 4.
Variation in the percentage of maize plants infested with larvae of Spodoptera frugiperda under different treatments over the days after seedling emergence.
Figure 4.
Variation in the percentage of maize plants infested with larvae of Spodoptera frugiperda under different treatments over the days after seedling emergence.
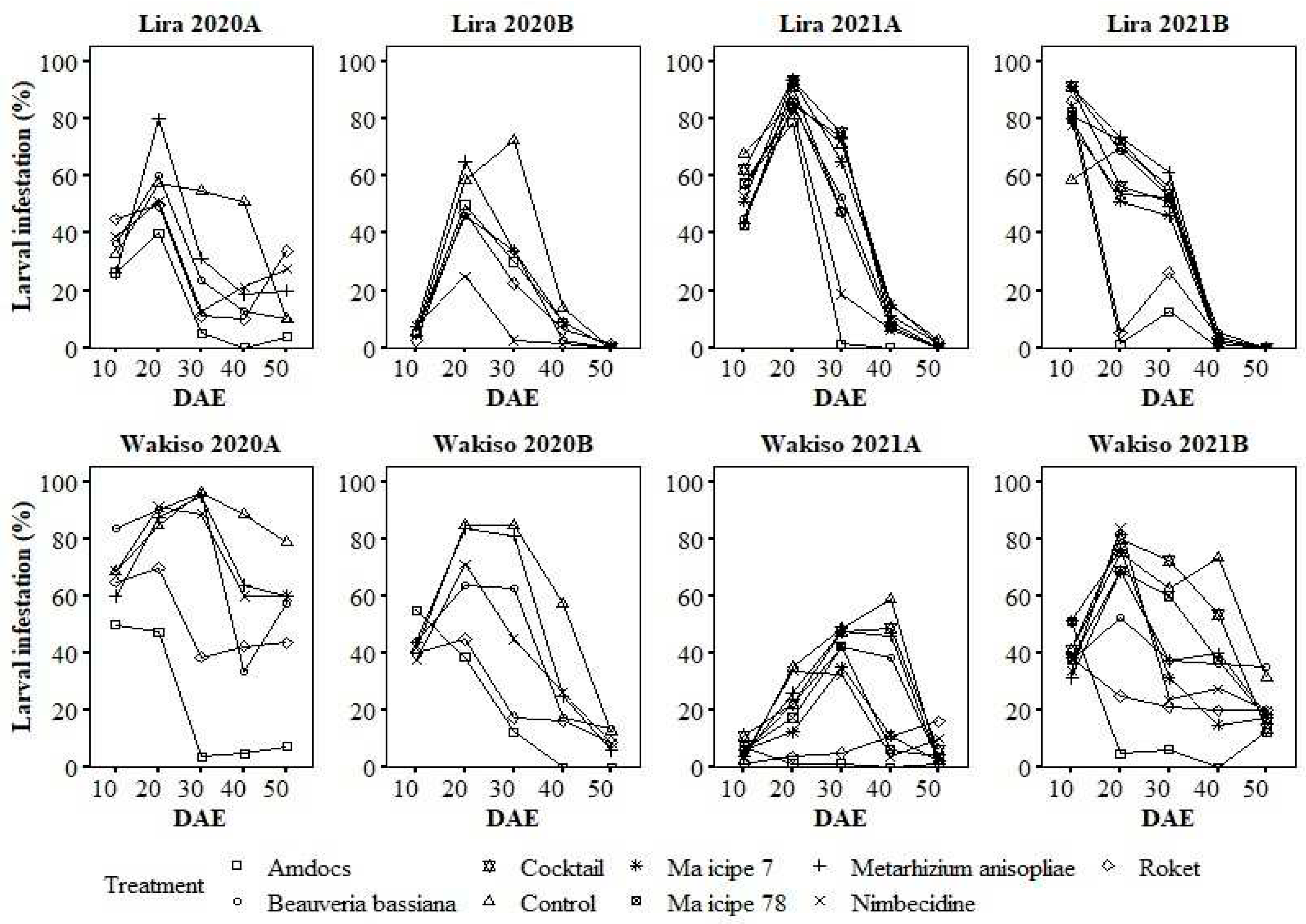
Figure 5.
Relationship between mean leaf damage score and maize grain yield.
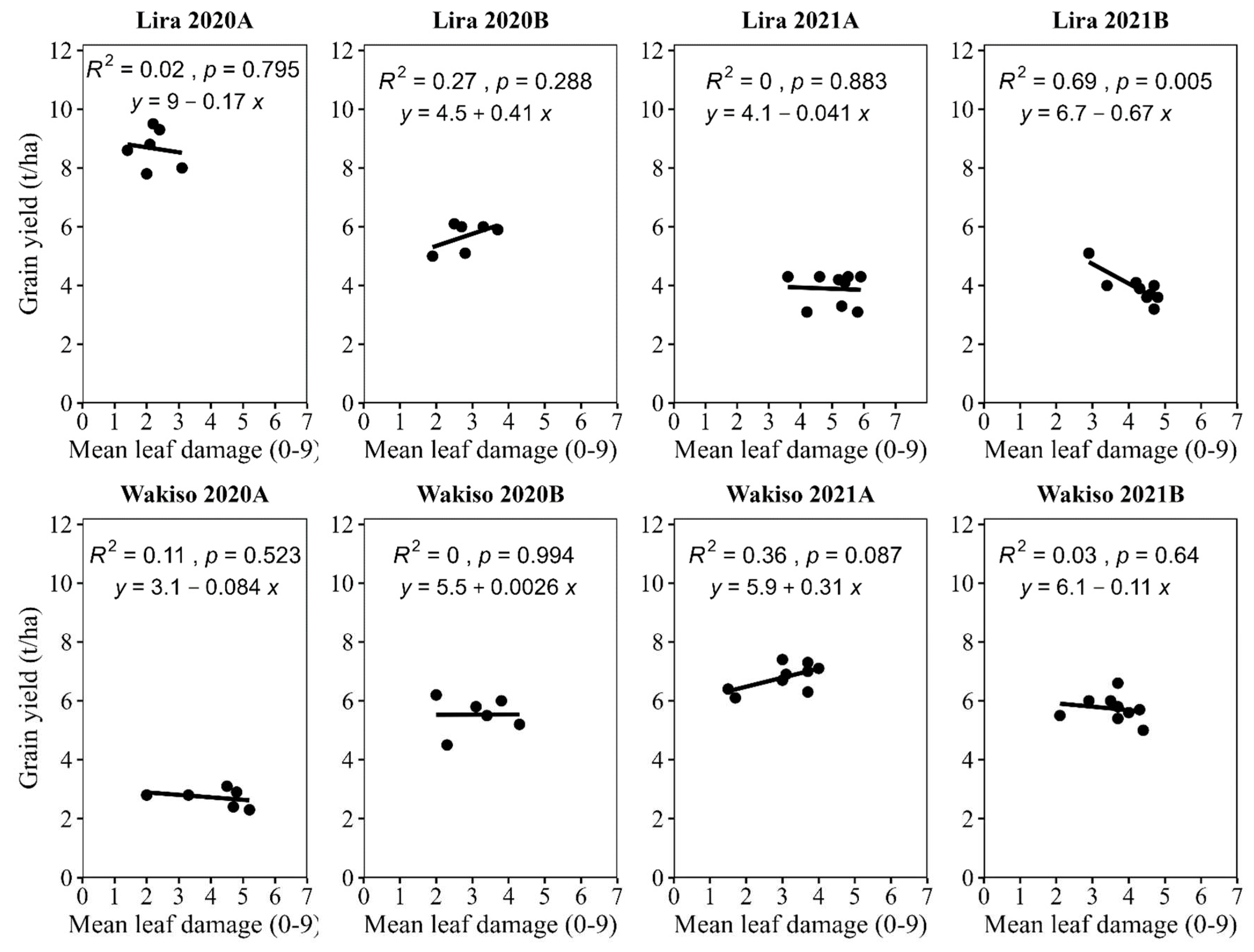
Figure 6.
Relationship between mean numbers of larvae per 20 plants with maize grain yield.
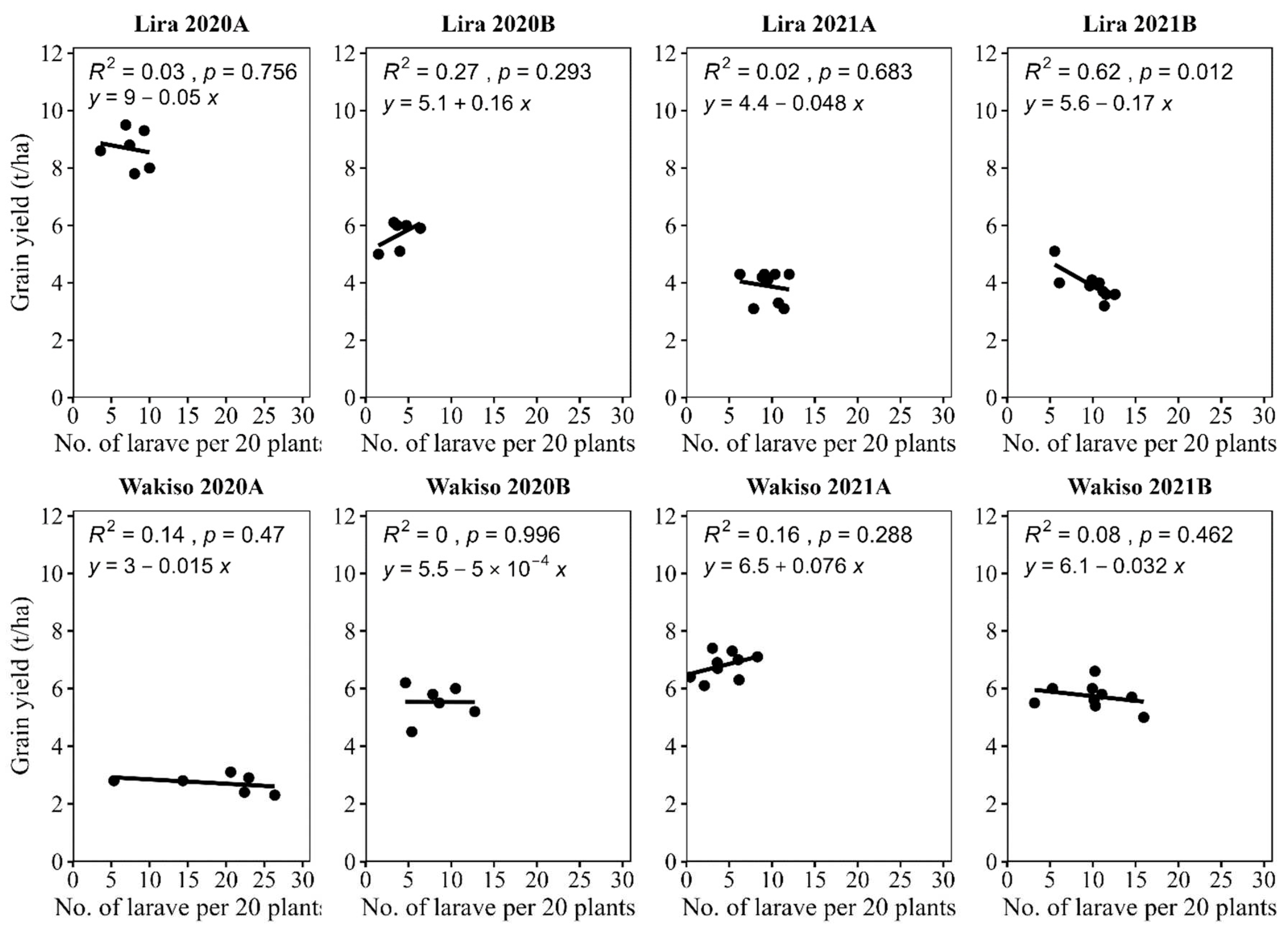
Figure 7.
Relationship between larval infestations with maize grain yield.
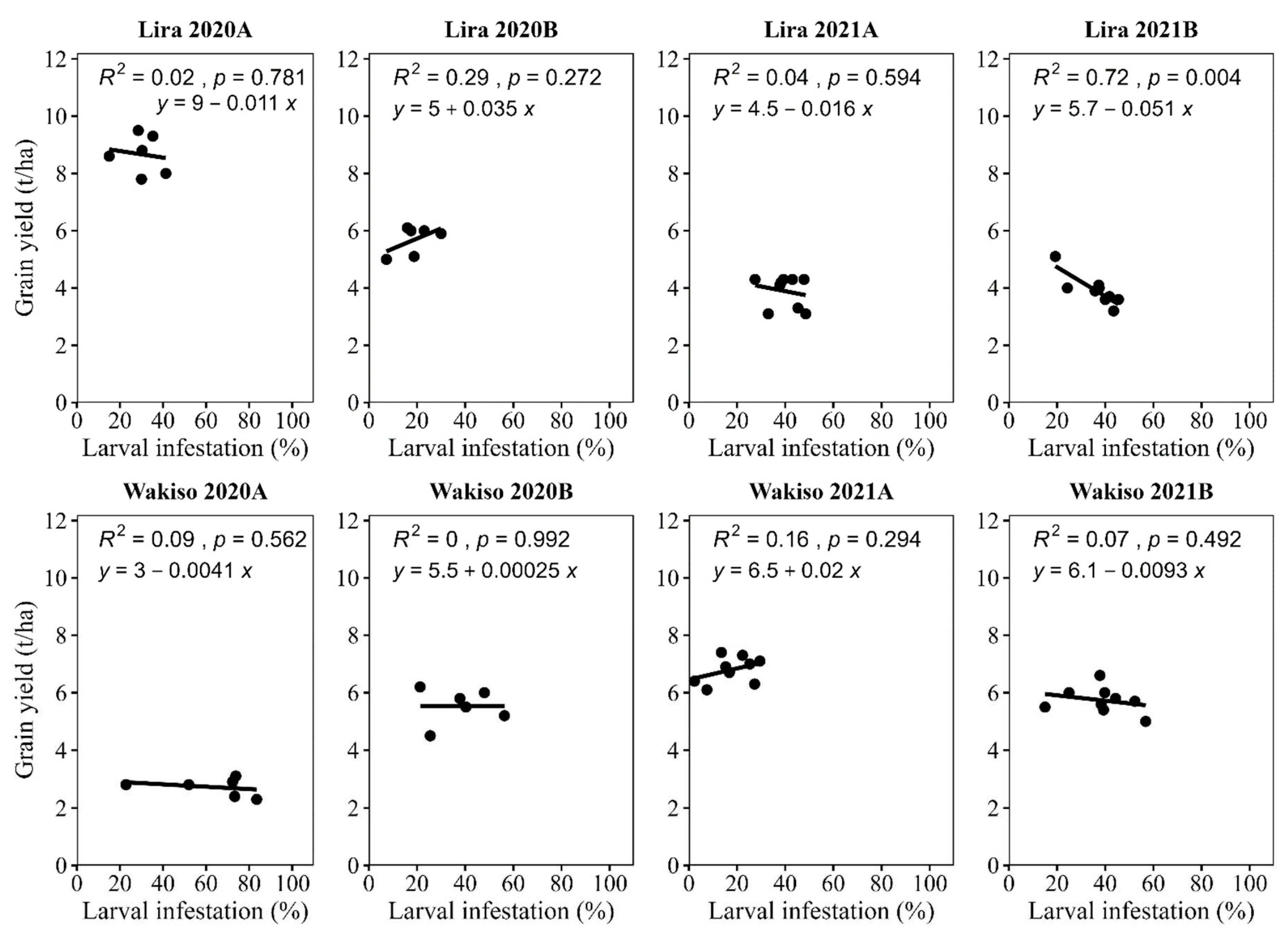

Table 1.
List of insecticides/treatments used in the study.
| Treatments | Category | Active ingredients | Application rate (per 20L) |
|---|---|---|---|
| Untreated control | - | - | |
| Beauveria bassiana (Bb) | Biopesticide | Beauveria bassiana spores | 20 g |
| General Insecticide Cocktail (GIC) | Biopesticide | Combination of biopesticides | 20 g |
| Metarhizium anisopliae (Ma) | Biopesticide | Metarhizium anisopliae spores | 20 g |
| Metarhizium anisopliae ICIPE 7 (Ma ICIPE 7) | Biopesticide | Metarhizium anisopliae strain ICIPE 7 | 20 ml |
| Metarhizium anisopliae ICIPE 78 (Ma ICIPE 78) | Biopesticide | Metarhizium anisopliae strain ICIPE 78 | 20 ml |
| Nimbecidine | Botanical | Azadirachtin 0.03% EC | 120 ml |
| Roket® | Synthetic insecticide | Profenofos 40% + Cypermethrin 4% EC | 30 ml |
| Amdocs | Semi-synthetic insecticide | Emamectin benzoate 2% + Abamectin 1% | 35 ml |
Table 2.
Mean leaf damage score and incidence of damaged maize plants under different treatments in different locations and seasons of 2020.
Table 2.
Mean leaf damage score and incidence of damaged maize plants under different treatments in different locations and seasons of 2020.
| 2020 | |||||
| Location | Treatments | FAW Leaf damage score | Damage incidence (%) | ||
| 2020A | 2020B | 2020A | 2020B | ||
| Lira | Amdocs | 1.4 ± 0.104d | 2.8 ± 0.521bc | 89.3 ± 2.931b | 89.3 ± 4.789 |
| Roket® | 2 .0 ± 0.138c | 2.5 ± 0.462cd | 97.3 ± 1.427a | 88.8 ± 5.836 | |
| Beauveria bassiana | 2.2 ± 0.222bc | 2.7 ± 0.516bc | 93.3 ± 3.105a | 86.5 ± 6.284 | |
| Metarhizium anisopliae | 2.4 ± 0.318ab | 3.3 ± 0.551ab | 92.5 ± 3.508a | 89.8 ± 4.953 | |
| Nimbecidine | 2.1 ± 0.274c | 1.9 ± 0.297d | 94.5 ± 2.922a | 86.0 ± 5.799 | |
| Control | 3.1 ± 0.485a | 3.7 ± 0.829a | 94.0 ± 3.453a | 85.3 ± 6.732 | |
| Mean | 2.2 | 2.8 | 93.5 | 87.6 | |
| SE | 0.1418 | 0.2317 | 2.0741 | 4.4713 | |
| X2 | 38.24 | 28.89 | 11.93 | 1.24 | |
| P value | <0.001 | <0.001 | 0.03577 | 0.9406 | |
| Wakiso | Amdocs | 2.0 ± 0.173c | 2.1 ± 0.116c | 95.3 ± 2.392 | 97.0 ± 2.334 |
| Roket® | 3.3 ± 0.126b | 2.3 ± 0.154c | 99.0 ± 0.585 | 96.8 ± 1.508 | |
| Beauveria bassiana | 4.8 ± 0.392a | 3.4 ± 0.514ab | 99.5 ± 0.344 | 95.0 ± 2.460 | |
| Metarhizium anisopliae | 4.7 ± 0.657a | 3.8 ± 0.580ab | 98.8 ± 0.714 | 94.3 ± 4.505 | |
| Nimbecidine | 4.5 ± 0.378a | 3.1 ± 0.363b | 99.3 ± 0.547 | 97.5 ± 2.251 | |
| Control | 5.2 ± 0.674a | 4.3 ± 0.697a | 99.8 ± 0.250 | 96.0 ± 2.422 | |
| Mean | 4.1 | 3.2 | 98.6 | 96.1 | |
| SE | 0.2666 | 0.2238 | 0.5582 | 1.3911 | |
| X2 | 62.29 | 40.98 | 9.02 | 2.79 | |
| P value | <0.001 | <0.001 | 0.1084 | 0.7327 | |
Table 3.
Mean leaf damage score, and incidence of damage maize under different treatments in Lira and Wakiso during the long and short rainy season of 2021.
Table 3.
Mean leaf damage score, and incidence of damage maize under different treatments in Lira and Wakiso during the long and short rainy season of 2021.
| 2021 | |||||
| Location | Treatments | FAW Leaf damage score | Damage incidence (%) | ||
| 2021A | 2021B | 2021A | 2021B | ||
| Lira | Amdocs | 3.6 ± 0.203d | 2.9 ± 0.529c | 100 ± 0.000 | 100 ± 0.000 |
| Roket® | 4.6 ± 0.299c | 3.4 ± 0.348bc | 100 ± 0.000 | 100 ± 0.000 | |
| General Insecticide Cocktail | 5.9 ± 0.623a | 4.5 ± 0.055a | 100 ± 0.000 | 100 ± 0.000 | |
| Metarhizium anisopliae ICIPE 7 | 5.3 ± 0.505b | 4.3 ± 0.077a | 99.8 ± 0.250 | 99.8 ± 0.250 | |
| Metarhizium anisopliae ICIPE 78 | 5.4 ± 0.620b | 4.6 ± 0.144a | 99.5 ± 0.500 | 100 ± 0.000 | |
| Metarhizium anisopliae | 5.6 ± 0.658ab | 4.8 ± 0.163a | 99.8 ± 0.250 | 100 ± 0.000 | |
| Beauveria bassiana | 5.2 ± 0.549b | 4.7 ± 0.101a | 99.8 ± 0.250 | 100 ± 0.000 | |
| Nimbecidine | 4.2 ± 0.326cd | 4.2 ± 0.101ab | 100 ± 0.000 | 100 ± 0.000 | |
| Control | 5.8 ± 0.586ab | 4.7 ± 0.163a | 100 ± 0.000 | 100 ± 0.000 | |
| Mean | 5.1 | 4.2 | 99.9 | 100.0 | |
| se | 0.1897 | 0.1190 | 0.0713 | 0.0278 | |
| X2 | 59.66 | 62.12 | 5.09 | 8.0 | |
| P value | <0.001 | <0.001 | 0.7484 | 0.4335 | |
| Wakiso | Amdocs | 1.5 ± 0.075d | 2.1 ± 0.269c | 94.5 ± 3.097 | 98.5 ± 0.896 |
| Roket® | 1.7 ± 0.139d | 2.9 ± 0.174bc | 93.5 ± 3.366 | 97.5 ± 1.333 | |
| General Insecticide Cocktail | 3.7 ± 0.624ab | 4.3 ± 0.485a | 94.0 ± 3.004 | 99.3 ± 0.547 | |
| Metarhizium anisopliae ICIPE 7 | 3.0 ± 0.344c | 4.0 ± 0.517a | 97.3 ± 1.559 | 98.8 ± 1.018 | |
| Metarhizium anisopliae ICIPE 78 | 3.1 ± 0.530bc | 3.7 ± 0.409ab | 94.3 ± 2.816 | 98.3 ± 1.271 | |
| Metarhizium anisopliae | 3.7 ± 0.701ab | 3.7 ± 0.442ab | 93.5 ± 3.346 | 97.0 ± 2.334 | |
| Beauveria bassiana | 3.7 ± 0.569abc | 3.5 ± 0.299ab | 93.5 ± 3.405 | 98.0 ± 1.376 | |
| Nimbecidine | 3.0 ± 0.512c | 3.7 ± 0.495ab | 94.8 ± 3.023 | 98.8 ± 0.801 | |
| Control | 4.0 ± 0.664a | 4.4 ± 0.628a | 95.0 ± 2.487 | 97.0 ± 1.598 | |
| Mean | 3.0 | 3.6 | 94.5 | 98.1 | |
| SE | 0.1987 | 0.1641 | 1.6969 | 0.5914 | |
| X2 | 61.55 | 53.49 | 0.93 | 3.28 | |
| P value | <0.001 | <0.001 | 0.9986 | 0.9159 | |
Table 4.
Number of Spodoptera frugiperda larvae and percentage of plants infested under different treatments Lira and Wakiso during the long and short rainy season of 2020.
Table 4.
Number of Spodoptera frugiperda larvae and percentage of plants infested under different treatments Lira and Wakiso during the long and short rainy season of 2020.
| 2020 | |||||
| Location | Treatments | Number of larvae per 20 plants | Percentage larval infestation (%) | ||
| 2020A | 2020B | 2020A | 2020B | ||
| Lira | Amdocs | 3.6 ± 1.381b | 4.0 ± 1.154 | 15.0 ± 7.756b | 18.8 ± 9.337 |
| Roket® | 8.1 ± 1.644a | 3.3 ± 0.853 | 30.0 ± 8.339a | 16.0 ± 8.745 | |
| Beauveria bassiana | 6.9 ± 1.423a | 3.7 ± 1.093 | 28.5 ± 9.146a | 17.5 ± 9.429 | |
| Metarhizium anisopliae | 9.3 ± 2.123a | 4.8 ± 1.252 | 35.3 ± 11.411a | 23.0 ± 11.941 | |
| Nimbecidine | 7.4 ± 1.257a | 1.5 ± 0.759 | 30.3 ± 6.771a | 7.30 ± 4.617 | |
| Control | 10 ± 1.629a | 6.4 ± 1.546 | 41.3 ± 8.962a | 30.0 ± 14.869 | |
| Mean | 7.5 | 3.9 | 30.0 | 18.8 | |
| SE | 0.667 | 0.475 | 3.6042 | 4.0374 | |
| X2 | 15.78 | 9.99 | 15.30 | 10.02 | |
| P value | 0.007517 | 0.07536 | 0.009151 | 0.07458 | |
| Wakiso | Amdocs | 5.4 ± 1.570c | 4.7 ± 1.326c | 22.8 ± 10.639c | 21.3 ± 11.011c |
| Roket® | 14.4 ± 2.173b | 5.4 ± 1.022bc | 52.0 ± 6.430b | 25.5 ± 7.144bc | |
| Beauveria bassiana | 23.0 ± 3.064a | 8.6 ± 1.491ab | 72.3 ± 11.669a | 40.3 ± 10.676ab | |
| Metarhizium anisopliae | 22.4 ± 2.630ba | 10.5 ± 1.781ab | 73.3 ± 7.475a | 48.0 ± 15.287ab | |
| Nimbecidine | 20.6 ± 2.332ab | 7.9 ± 1.270ab | 73.8 ± 6.835a | 37.8 ± 10.364ab | |
| Control | 26.4 ± 2.728a | 12.8 ± 1.915a | 83.5 ± 4.650a | 56.3 ± 13.773a | |
| Mean | 18.7 | 8.3 | 62.9 | 38.2 | |
| SE | 1.173 | 0.651 | 4.8702 | 4.8796 | |
| X2 | 40.77 | 17.10 | 43.94 | 17.79 | |
| P value | <0.0001 | 0.004319 | <0.0001 | 0.003216 | |
Table 5.
Number of Spodoptera frugiperda larvae and percentage of maize infested under different treatments Lira and Wakiso during the long and short rainy season of 2021.
Table 5.
Number of Spodoptera frugiperda larvae and percentage of maize infested under different treatments Lira and Wakiso during the long and short rainy season of 2021.
| 2021 | |||||
| Location | Treatments | Number of larvae per 20 plants* | Percentage larval infestation (%) | ||
| 2021A | 2021B | 2021A | 2021B | ||
| Lira | Amdocs | 6.3 ± 1.975 | 5.6 ± 2.272 | 27.5 ± 16.924 | 19.3 ± 15.986 |
| Roket® | 9.1 ± 2.130 | 6.1 ± 2.045 | 39.3 ± 15.928 | 24.3 ± 16.166 | |
| General Insecticidal Cocktail | 12 .0± 2.589 | 11.6 ± 2.722 | 47.8 ± 18.677 | 40.0 ± 17.491 | |
| Metarhizium anisopliae ICIPE 7 | 10.8 ± 2.269 | 9.7 ± 2.151 | 45.3 ± 16.777 | 35.8 ± 15.455 | |
| Metarhizium anisopliae ICIPE 78 | 9.5 ± 2.359 | 11.2 ± 2.330 | 37.8 ± 16.310 | 41.8 ± 17.368 | |
| Metarhizium anisopliae | 10.4 ± 2.310 | 12.6 ± 2.595 | 43.0 ± 16.525 | 45.5 ± 18.931 | |
| Beauveria bassiana | 8.9 ± 2.017 | 11.4 ± 2.437 | 38.3 ± 15.439 | 43.5 ± 17.851 | |
| Nimbecidine | 7.9 ± 2.128 | 9.9 ± 2.176 | 33.0 ± 15.903 | 37.3 ± 15.362 | |
| Control | 11.4 ± 2.130 | 10.8 ± 2.669 | 48.5 ± 16.646 | 37.5 ± 14.984 | |
| Mean | 9.6 | 9.9 | 40.0 | 36.1 | |
| SE | 0.734 | 0.797 | 5.0985 | 5.1790 | |
| X2 | 7.92 | 6.54 | 7.59 | 7.15 | |
| P value | 0.4418 | 0.5868 | 0.4746 | 0.5204 | |
| Wakiso | Amdocs | 0.5 ± 0.170d | 3.2 ± 0.978c | 2.3 ± 1.335d | 15.0 ± 9.279c |
| Roket® | 2.1 ± 1.183cd | 5.3 ± 0.692bc | 7.5 ± 2.739cd | 25.0 ± 3.558bc | |
| General Insecticidal Cocktail | 6.2 ± 1.265a | 14.6 ± 2.567a | 27.3 ± 8.922a | 52.3 ± 11.802a | |
| Metarhizium anisopliae ICIPE 7 | 3.1 ± 1.072bc | 10.2 ± 1.889ab | 13.5 ± 5.665bc | 38.3 ± 11.474ab | |
| Metarhizium anisopliae ICIPE 78 | 3.6 ± 1.150abc | 11.1 ± 1.690ab | 15.3 ± 7.218abc | 44.3 ± 9.097ab | |
| Metarhizium anisopliae | 6.1 ± 1.434ab | 10.3 ± 1.857ab | 25.3 ± 9.790ab | 39.3 ± 8.241ab | |
| Beauveria bassiana | 5.4 ± 1.326ab | 10.0 ± 1.141ab | 22.3 ± 8.276ab | 39.8 ± 3.221ab | |
| Nimbecidine | 3.7 ± 1.069abc | 10.3 ± 2.362abc | 16.8 ± 6.785abc | 37.8 ± 11.722abc | |
| Control | 8.3 ± 2.016ab | 16.0 ± 2.534a | 29.5 ± 11.65ab | 56.8 ± 8.791a | |
| Mean | 4.3 | 10.1 | 17.7 | 38.7 | |
| SE | 0.447 | 0.668 | 2.6362 | 3.2852 | |
| X2 | 31.41 | 37.27 | 30.83 | 37.41 | |
| P value | 0.000119 | <0.0001 | 0.0001504 | <0.0001 | |
Table 6.
Total number of egg batches and adults of Spodoptera frugiperda under different treatments in Lira and Wakiso during the long and short rainy seasons of 2020 and 2021.
Table 6.
Total number of egg batches and adults of Spodoptera frugiperda under different treatments in Lira and Wakiso during the long and short rainy seasons of 2020 and 2021.
| Location | Insecticide treatment | Number of egg batches | Number of adults* | |||||||
| 2020A | 2020B | 2021A | 2021B | 2020A | 2020B | 2021A | 2021B | |||
| Lira | Amdocs | 12.0 | 3.0 | 5.0 | 2.0 | 1.0 | 1.0 | 0.0 | 3.0 | |
| Roket® | 11.0 | 2.0 | 0.0 | 9.0 | 1.0 | 0.0 | 1.0 | 1.0 | ||
| Beauveria bassiana | 19.0 | 2.0 | 3.0 | 5.0 | 3.0 | 1.0 | 1.0 | 2.0 | ||
| Metarhizium anisopliae | 11.0 | 6.0 | 2.0 | 6.0 | 0.0 | 0.0 | 1.0 | 0.0 | ||
| Metarhizium anisopliae ICIPE 7 | 4.0 | 4.0 | 1.0 | 0.0 | ||||||
| Metarhizium anisopliae ICIPE 78 | 5.0 | 4.0 | 0.0 | 2.0 | ||||||
| General Insecticidal Cocktail | 6.0 | 0.0 | 1.0 | 0.0 | ||||||
| Nimbecidine | 11.0 | 6.0 | 4.0 | 4.0 | 0.0 | 0.0 | 0.0 | 2.0 | ||
| Control | 2.0 | 7.0 | 6.0 | 8.0 | 2.0 | 0.0 | 0.0 | 1.0 | ||
| Total | 66 | 26 | 35 | 42 | 0 | 7 | 2 | 5 | ||
| Wakiso | Amdocs | 19.0 | 14.0 | 2.0 | 0.0 | 2.0 | 1.0 | 2.0 | 0.0 | |
| Roket® | 16.0 | 7.0 | 1.0 | 6.0 | 5.0 | 1.0 | 0.0 | 2.0 | ||
| Beauveria bassiana | 14.0 | 6.0 | 7.0 | 3.0 | 3.0 | 2.0 | 0.0 | 1.0 | ||
| Metarhizium anisopliae | 15.0 | 8.0 | 2.0 | 6.0 | 4.0 | 0.0 | 4.0 | 3.0 | ||
| Metarhizium anisopliae ICIPE 7 | 1.0 | 1.0 | 2.0 | 0.0 | ||||||
| Metarhizium anisopliae ICIPE 78 | 1.0 | 3.0 | 0.0 | 5.0 | ||||||
| General Insecticidal Cocktail | 4.0 | 2.0 | 1.0 | 0.0 | ||||||
| Nimbecidine | 17.0 | 7.0 | 2.0 | 4.0 | 3.0 | 0.0 | 0.0 | 2.0 | ||
| Control | 11.0 | 3.0 | 2.0 | 7.0 | 2.0 | 0.0 | 0.0 | 1.0 | ||
| Total | 92 | 45 | 22 | 32 | 19 | 4 | 9 | 14 | ||
| Overall total | 158 | 71 | 57 | 74 | 26 | 6 | 14 | 25 | ||
Table 7.
Grain yield and yield losses under different treatments Lira and Wakiso during the long and short rainy seasons 2020 and 2021.
Table 7.
Grain yield and yield losses under different treatments Lira and Wakiso during the long and short rainy seasons 2020 and 2021.
| Location | Treatments | Grain yield (t/ha) | Yield advantage over control (%) | ||||||
| 2020A | 2020B | 2021A | 2021B | 2020A | 2020B | 2021A | 2021B | ||
| Lira | Amdocs | 8.6 ± 0.627 | 5.1 ± 0.206 | 4.3 ± 0.370 | 5.1 ± 0.328 | 7.5 | -13.6 | 38.7 | 27.5 |
| Roket® | 7.8 ± 0.392 | 6.1 ± 0.136 | 4.3 ± 0.385 | 4.0 ± 0.683 | -2.5 | 3.4 | 38.7 | 0.0 | |
| General Insecticidal Cocktail | 4.3 ± 0.398 | 3.6 ± 0.142 | 38.7 | -10.0 | |||||
| Metarhizium anisopliae ICIPE 7 | 3.3 ± 0.095 | 3.9 ± 0.718 | 6.5 | -2.5 | |||||
| Metarhizium anisopliae ICIPE 78 | 4.1 ± 0.116 | 3.7 ± 0.206 | 32.3 | -7.5 | |||||
| Metarhizium anisopliae | 9.3 ± 0.544 | 6.0 ± 0.499 | 4.3 ± 0.608 | 3.6 ± 0.606 | 16.3 | 1.7 | 38.7 | -10.0 | |
| Beauveria bassiana | 9.5 ± 0.460 | 6.0 ± 0.389 | 4.2 ± 0.501 | 3.2 ± 0.674 | 18.8 | 1.7 | 35.5 | -20.0 | |
| Nimbecidine | 8.8 ± 0.660 | 5.0 ± 0.238 | 3.1 ± 0.531 | 4.1 ± 0.287 | 10.0 | -15.3 | 0.0 | 2.5 | |
| Control | 8.0 ± 0.420 | 5.9 ± 0.296 | 3.1 ± 0.303 | 4.0 ± 0.466 | |||||
| Mean | 8.67 | 5.68 | 3.88 | 3.906 | |||||
| SE | 1.110 | 0.404 | 0.649 | 1.010 | |||||
| Lsd | 2.519 | 1.519 | 2.0302 | 2.532 | |||||
| %cv | 12.17 | 11.182 | 20.769 | 25.729 | |||||
| P value | 0.202 | 0.085 | 0.125 | 0.442 | |||||
| Wakiso | Amdocs | 2.8 ± 0.966 | 6.2 ± 0.496 | 6.4 ± 0.595 | 5.5 ± 0.391 | 21.7 | 19.2 | -9.9 | 10.0 |
| Roket® | 2.8 ± 0.501 | 4.5 ± 0.539 | 6.1 ± 0.659 | 6.0 ± 0.850 | 21.7 | -13.5 | -14.1 | 20.0 | |
| General Insecticidal Cocktail | 6.3 ± 0.616 | 5.7 ± 0.458 | -11.3 | 14.0 | |||||
| Metarhizium anisopliae ICIPE 7 | 7.4 ± 0.638 | 5.6 ± 0.448 | 4.2 | 12.0 | |||||
| Metarhizium anisopliae ICIPE 78 | 6.9 ± 0.861 | 5.8 ± 0.670 | -2.8 | 16.0 | |||||
| Metarhizium anisopliae | 2.4 ± 0.526 | 6.0 ± 0.507 | 7.0 ± 0.290 | 5.4 ± 1.098 | 4.3 | 15.4 | -1.4 | 8.0 | |
| Beauveria bassiana | 2.9 ± 0.873 | 5.5 ± 0.825 | 7.3 ± 0.392 | 6.0 ± 0.264 | 26.1 | 5.8 | 2.8 | 20.0 | |
| Nimbecidine | 3.1 ± 0.533 | 5.8 ± 0.891 | 6.7 ± 0.866 | 6.6 ± 0.272 | 34.8 | 11.5 | -5.6 | 32.0 | |
| Control | 2.3 ± 0.415 | 5.2 ± 1.124 | 7.1 ± 0.550 | 5.0 ± 0.997 | |||||
| Mean | 2.70 | 5.515 | 6.81 | 5.732 | |||||
| SE | 1.787 | 2.355 | 1.603 | 1.812 | |||||
| Lsd | 3.195 | 3.668 | 3.190 | 3.391 | |||||
| %cv | 49.36 | 27.825 | 18.591 | 23.482 | |||||
| P value | 0.933 | 0.652 | 0.823 | 0.881 | |||||
Table 8.
Natural enemies (predators and parasitoids) species recovered from eggs and larvae of Spodoptera frugiperda collected from maize fields in Uganda, host stage attacked and their sequence identity as compared with publicly available sequences from GenBank and iBoL entries.
Table 8.
Natural enemies (predators and parasitoids) species recovered from eggs and larvae of Spodoptera frugiperda collected from maize fields in Uganda, host stage attacked and their sequence identity as compared with publicly available sequences from GenBank and iBoL entries.
| Order and Family | Species | Location | Host Stage Attacked |
Species with the Closest Nucleotide Sequence Match |
Percent Identity, and Reference GenBank Accession Number and iBoL Entries |
|---|---|---|---|---|---|
|
Hymenoptera: Platygastridae |
Telenomus remus Dixon | Wakiso and Lira | Eggs | Telenomus remus | 100% (ON923739.1) [30] |
| Hymenoptera: Eurytomidae | Eurytomidae sp. | Wakiso | Egg/Larval | Eurytoma asphodeli | 87.16% KT623736.1 [41] |
| Hymenoptera: Braconidae | Coccygidium luteum | Wakiso and Lira | Larvae | Coccygidium luteum | 99.64% MT784187 [36] |
| Coccygidium sp | Wakiso and Lira | Larvae | Coccygidium sp | ||
| Cotesia icipe | Wakiso and Lira | Larvae | Cotesia icipe | 100% MN900735.1 [27], 100% MT780217 [36] | |
| Parapanteles sp. | Wakiso | Larvae | Parapanteles athamasae | 100% HM397613.1 [42] | |
| Chelonus sp. | Wakiso and Lira | Egg/Larval | Chelonus insularis | 97.42% XM_035078068 | |
| Dolichogenidea sp. | Wakiso | Larval | Dolichogenidea sp. | 93.28% JF271344.1 [43] | |
| Hymenoptera: Pteromalidae | Micranisa sp | Wakiso | Larvae | Micranisa sp | 87.34% MK530760.1 [44] |
|
Hymenoptera: Ichneumonidae |
Charops cf. diversipes | Wakiso and Lira | Larvae | Charops cf. diversipes | 100% (MT784182.1), 100% (MT784181.1), 100% (MT784179.1) [36], 100% (MT784183.1) [27] |
| Diptera: Tachinidae | Tachinidae sp. | Wakiso and Lira | Larvae/pupae | Tachinidae sp. | 99.35% (MT784176.1) [36] |
Table 9.
Total number of egg batches collected, total number of Telenomus remus recovered, and parasitism of Spodoptera frugiperda egg batches under different treatments in 2020.
Table 9.
Total number of egg batches collected, total number of Telenomus remus recovered, and parasitism of Spodoptera frugiperda egg batches under different treatments in 2020.
| Treatment | No. of Egg batches collected | No. of parasitoids |
No. of Egg batches parasitized | Overall parasitism (%) |
||||
|---|---|---|---|---|---|---|---|---|
| 2020A | 2020B | 2020A | 2020B | 2020A | 2020B | 2020A | 2020B | |
| Lira | ||||||||
| Amdocs | 12 | 3 | 113 | 20 | 6 | 2 | 50.0 | 66.7 |
| Roket® | 11 | 2 | 65 | 0 | 1 | 0 | 9.1 | 0.0 |
| Beauveria bassiana | 19 | 2 | 0 | 12 | 0 | 1 | 0.0 | 50.0 |
| Metarhizium anisopliae | 11 | 6 | 16 | 23 | 1 | 1 | 9.1 | 16.7 |
| Nimbecidine | 11 | 6 | 0 | 30 | 0 | 1 | 0.0 | 16.7 |
| Control | 2 | 7 | 14 | 0 | 2 | 0 | 100.0 | 0.0 |
| Wakiso | ||||||||
| Amdocs | 19 | 14 | 77 | 0 | 3 | 0 | 15.8 | 0.0 |
| Roket® | 16 | 7 | 32 | 0 | 3 | 0 | 18.8 | 0.0 |
| Beauveria bassiana | 14 | 6 | 140 | 0 | 5 | 0 | 35.7 | 0.0 |
| Metarhizium anisopliae | 15 | 8 | 22 | 25 | 2 | 2 | 13.3 | 25.0 |
| Nimbecidine | 17 | 7 | 103 | 0 | 5 | 0 | 29.4 | 0.0 |
| Control | 11 | 3 | 63 | 0 | 2 | 0 | 18.2 | 0.0 |
Table 10.
Total number of egg batches collected, total number of Telenomus remus recovered, and parasitism of Spodoptera frugiperda egg batches under different treatments in 2021.
Table 10.
Total number of egg batches collected, total number of Telenomus remus recovered, and parasitism of Spodoptera frugiperda egg batches under different treatments in 2021.
| Treatment | No. of Egg batches collected | No. of parasitoids | No. of Egg batches parasitized | Overall parasitism (%) | ||||
|---|---|---|---|---|---|---|---|---|
| 2021A | 2021B | 2021A | 2021B | 2021A | 2021B | 2021A | 2021B | |
| Lira | ||||||||
| Amdocs | 5 | 2 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Roket® | 0 | 9 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Beauveria bassiana | 3 | 5 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Metarhizium anisopliae | 2 | 6 | 10 | 0 | 1 | 0 | 50.0 | 0.0 |
| Metarhizium anisopliae icipe 7 | 4 | 4 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Metarhizium anisopliae icipe 78 | 5 | 4 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| General Insecticidal Cocktail | 6 | 0 | 30 | 0 | 1 | 0 | 16.7 | 0.0 |
| Nimbecidine | 4 | 4 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Control | 6 | 8 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Wakiso | ||||||||
| Amdocs | 2 | 0 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Roket® | 1 | 6 | 20 | 0 | 1 | 0 | 100.0 | 0.0 |
| Beauveria bassiana | 7 | 3 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Metarhizium anisopliae | 2 | 6 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Metarhizium anisopliae icipe 7 | 1 | 1 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Metarhizium anisopliae icipe 78 | 1 | 3 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| General Insecticidal Cocktail | 4 | 2 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Nimbecidine | 2 | 4 | 0 | 0 | 0 | 0 | 0.0 | 0.0 |
| Control | 2 | 7 | 22 | 0 | 2 | 0 | 100.0 | 0.0 |
Table 11.
Total number of larvae collected, total number of larval parasitoids recovered, and parasitism of Spodoptera frugiperda larvae under different treatments Lira and Wakiso during the long and short rainy season of 2020.
Table 11.
Total number of larvae collected, total number of larval parasitoids recovered, and parasitism of Spodoptera frugiperda larvae under different treatments Lira and Wakiso during the long and short rainy season of 2020.
| Treatment and district | No. of larvae collected | No. of parasitoids |
Overall parasitism %) |
2020A | 2020B | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020A | 2020B | 2020A | 2020B | 2020A | 2020B | Chd | Chs | Col | Cos | Coi | Mic | Tas | Chd | Col | Cos | Coi | ||
| Lira | ||||||||||||||||||
| Amdocs | 4 | 9 | 1 | 0 | 25 | 0 | 25 | |||||||||||
| Beauveria bassiana | 14 | 47 | 0 | 0 | 0 | 0 | ||||||||||||
| Control | 50 | 52 | 0 | 2 | 0 | 3.8 | 1.9 | 1.9 | ||||||||||
| Metarhizium anisopliae | 31 | 48 | 0 | 0 | 0 | 0 | ||||||||||||
| Nimbecidine | 23 | 28 | 0 | 0 | 0 | 0 | ||||||||||||
| Roket® | 18 | 19 | 0 | 0 | 0 | 0 | ||||||||||||
| Wakiso | ||||||||||||||||||
| Amdocs | 7 | 6 | 3 | 0 | 42.9 | 0 | 14.3 | 14.3 | 14.3 | |||||||||
| Beauveria bassiana | 51 | 35 | 29 | 4 | 27.4 | 11.5 | 3.9 | 7.8 | 9.8 | 3.9 | 2.0 | 8.6 | 2.9 | |||||
| Control | 94 | 46 | 13 | 4 | 17.1 | 8.7 | 1.1 | 1.1 | 3.2 | 9.6 | 2.1 | 2.2 | 4.3 | 2.2 | ||||
| Metarhizium anisopliae | 52 | 40 | 12 | 0 | 23.0 | 0 | 3.8 | 11.5 | 5.8 | 1.9 | ||||||||
| Nimbecidine | 58 | 28 | 13 | 0 | 22.3 | 0 | 3.4 | 8.6 | 3.4 | 6.9 | ||||||||
| Roket® | 21 | 12 | 5 | 0 | 23.8 | 0 | 9.5 | 4.8 | 9.5 | |||||||||
Chd = Charops cf. diversipes; Chs; Chelonus sp; Col = Coccygidium luteum; Cos = Coccygidium sp; Coi = Cotesia icipe; Mic = Micranisa sp; Tas = Tachinidae sp;.
Table 12.
Total larvae collected, number of larval parasitoids recovered, and parasitism of Spodoptera frugiperda larvae under different treatments in Lira and Wakiso during the long and short rainy season in 2021.
Table 12.
Total larvae collected, number of larval parasitoids recovered, and parasitism of Spodoptera frugiperda larvae under different treatments in Lira and Wakiso during the long and short rainy season in 2021.
| District and Treatment |
No. of FAW larvae collected |
No. of parasitoids | Parasitism (%) | 2021A | 2021B | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 21A | 21B | 21A | 21B | 21A | 21B | Chs | Col | Cos | Coi | Tas | Chd | Chs | Col | Coi | Tas | |
| Lira | ||||||||||||||||
| Amdocs | 22 | 28 | 1 | 0 | 3.6 | 3.6 | ||||||||||
| Roket® | 43 | 67 | 3 | 0 | 4.5 | 1.5 | 3 | |||||||||
| Beauveria bassiana | 43 | 110 | 1 | 3 | 2.3 | 2.7 | 2.3 | 2.7 | ||||||||
| Metarhizium anisopliae | 62 | 107 | 1 | 3 | 1.6 | 2.7 | 1.6 | 0.9 | 1.8 | |||||||
| Metarhizium anisopliae ICIPE 7 | 44 | 88 | 4 | 0 | 4.5 | 4.5 | ||||||||||
| Metarhizium anisopliae ICIPE 78 | 51 | 124 | 3 | 0 | 2.4 | 2.4 | ||||||||||
| General Insecticidal Cocktail | 49 | 113 | 4 | 0 | 3.6 | 0.9 | 2.7 | |||||||||
| Nimbecidine | 28 | 81 | 2 | 0 | 2.4 | 2.4 | ||||||||||
| Control | 62 | 98 | 1 | 2 | 1.6 | 2 | 1.6 | 1 | 1 | |||||||
| Wakiso | ||||||||||||||||
| Amdocs | 11 | 25 | 0 | 0 | 0 | 0 | ||||||||||
| Roket® | 13 | 33 | 0 | 1 | 0 | 3.0 | 3.0 | |||||||||
| Beauveria bassiana | 37 | 44 | 1 | 2 | 2.7 | 6.8 | 2.7 | 4.5 | 2.3 | |||||||
| Metarhizium anisopliae | 38 | 47 | 5 | 3 | 13.1 | 4.5 | 2.6 | 7.9 | 2.6 | 4.5 | ||||||
| Metarhizium anisopliae ICIPE 7 | 29 | 46 | 0 | 1 | 0 | 2.2 | 2.2 | |||||||||
| Metarhizium anisopliae ICIPE 78 | 16 | 59 | 1 | 2 | 6.3 | 3.4 | 6.3 | 3.4 | ||||||||
| General Insecticidal Cocktail | 34 | 55 | 3 | 0 | 8.7 | 0 | 2.9 | 2.9 | 2.9 | |||||||
| Nimbecidine | 37 | 53 | 1 | 1 | 16.2 | 1.9 | 8.1 | 8.1 | 1.9 | |||||||
| Control | 40 | 58 | 3 | 4 | 7.5 | 6.8 | 7.5 | 3.4 | 1.7 | 1.7 | ||||||
Chs= Chelonus sp; Col = Coccygidium luteum; Cos = Coccygidium sp; Coi = Cotesia icipe; Tas = Tachinidae sp; Chd = Charops cf. diversipes;.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



