Submitted:
14 December 2023
Posted:
14 December 2023
You are already at the latest version
Abstract
The populations of clearwing moth borers in Korea have recently exploited and severe damage on pin oaks (Quercus palustris Munchh.). We conducted the field monitoring and the molecular analyses to identify them in an integrated manner. Morphological examination and molecular analyses on COI-gene based intra-and inter-specific genetic divergences (GD) were revealed that the borers were recognized as two invasive species, Sphecodoptera sheni and Paranthrenella pinoakula sp. nov.. The maximum intraspecific GD was found to be 1.9%, whereas the minimum interspecific GD was confirmed as 8.1%, with a distinct barcoding gap. Both MJ network and NJ tree were also showed that 18 haplotypes (HAPs) were detected from the 52 COI sequences. The borers were revealed total 17 HAPs: i) H1-H7 were detected from all seven regions with S. sheni; ii) Wonju and Goyang populations of S. sheni were revealed as more than three HAPs; iii) H7 was closely connected with H8 of Chinese population of S. sheni; iv) H9-H10 were detected as the other samples from Wonju population with P. pinoakula sp. n., and closely located with congeneric species. Maximum likelihood tree also revealed that P. pinoacula sp. n. nested within the congeneric species by genetically separating from the S. sheni.
Keywords:
invasive insect pests
; regional populations
; genetic divergence
; COI
; street insect pests
1. Introduction
Climate change presents a global threat to humanity [1,2]. Certain human activities, such as the expansion of worldwide trade and the increase in domestic and international travel, have led to unanticipated issues, such as the invasive species. Since 1900, 89 different invasive species have appeared in Korea, with 33 of them, or 38% of the total, being introduced in the last 19 years [3]. Moreover, extreme weather phenomena are not only spreading exotic pests but also triggering outbreaks of native insect pests in human habitats and forests. While the ecological function of urban street trees is providing natural scents and shade that act as a carbon sink in the urban areas, they also serve a special habitat for invasive forest pests.
Pin oak, Quercus palustris Munchh. (Fagaceae) was one of the most popular varieties to be planted as a domesticated street tree since the early 2000s in Korea (Figure 1). However, around 2014, signs of clearwing borer damage began to be reported from street trees of pin oaks in Korea. Our research team has been conducting nationwide monitoring for the past two years. Multiple pin oak populations planted as street trees show significant pest damage signs throughout Korea. The congruent symptoms were caused by the larvae, which bore holes in the xylem, crawled inside the tunnels to feed, and oozing milky sap with coarse reddish-brown excrement on the bark of pin oaks. The larvae attack the cambium layer of the xylem structure and create a pupal frame under barks. This leads to stunted tree growth and can cause decline. In addition, emerged adult moths leave conspicuous marks including pupal exuviae and holes on the bark that disturb the urban landscape. (Figure 2A-F). These larvae are often confused with beetle larvae due to the similarities on perforating feeding habit, large body size and milky coloration. However, the number of prolegs, development of pinaculums and other characteristics allow them to be classified as clearwing moth larvae (Figure 2G).
The clearwing moth family (Lepidoptera: Cossidae: Sesiinae) is a group of medium to large-sized that resemble adult bees or wasps, with 1,562 species and subspecies recorded worldwide [4], including many agricultural (e.g. Nokona pernix, N. purpurea, N. regalis, Synanthedon bicingulata, S. haitangvora etc.) and forestry pests (e.g. Scasiba rhynchioides, Synanthedon quercus etc.).
The study has two objectives. Firstly, we monitor and identify the insect pest species attacking pin oak street trees based on morphological and molecular analyses in Korea. Secondly, we intended the prediction of domestic routes of migration within Korea. To achieve the research objectives, we analyzed genetic diversity and haplotypes, and constructed a neighbor joining (NJ) tree in addition to planting history. Furthermore, phylogenetic analysis was used to infer the relationships between the insect pest groups.
2. Materials and Methods
2.1. Sampling
From 2022 to 2023, 41 individuals were collceted by hands using pruning shearsin seven regions (Bucheon, Goyang and Seongnam of Province Gyeonggi (GG); Chuncheon and Wonju of Prov. Gangwon (GW); Cheongju of Prov. Chungcheongbuk (CB); Daegu Metropolitan city and Gwangju Metropolitan city), planted with domestic pin oak street trees in Korea. Among the areas, the trees in Goyang and Wonju were planted in early 2000s, while the others were planted within 5 years. In Gwangju Metropolitan city, we have only confirmed the insect pest damage, but no individuals were caught. Since they were in larval or pupal stages, they were reared in the falcon tube (200ml) individually to adults maintaining at 26-27°C, 12:12 (L:D), 60% humidity in the laboratory. For morphological identification, all individuals were examined their external characters and photographed under Leica Z16 APO stereomicroscope and Dhyana 400DC (4M) sCMOS camera (TUCSEN) with Mosaic Analysis Software. Genital slide vouchers were made according to Kim et al. [5]. All specimens and slide vouchers were deposited in the laboratory of Insect Phylogenetics and Evolution, Jeonbuk National University (IPE JBNU), Korea.
2.2. Molecular protocol
Total 52 sequences were included for molecular protocol. Of them, 50 sequences were used as ingroup: i) 9 sequences of 6 species (Paranthrenella auriplena, P. cinnamoma, P. chrysophanes, P. formosicola, P. terminalia and Sphecodoptera sheni) from BOLD public data (ANICF612-10, ANICF613-10, ANICF615-10, EPNG8497-14, EPNG8498-14, GBMNE5028-21, GBMNE61298-22, GSCMB812-12, LTOLB087-08); ii) 41 sequences of 2 species (Sphecodoptera sheni and Paranthrenella pinoakula sp. n.) from field survey. Two sequences of one species (Anatrachyntis japonica: Cosmopterigidae) used as out group.
DNA was extracted from the head or leg of each specimen using Tissue Genomic DNA Isolation Kit Mini in LaboPassTm DNA Purification Kit (Cosmo Genetech Co. Ltd., Republic of Korea), following the manufacturer’s protocol. For PCR amplification for mitochondrial cytochrome oxidase subunit I (653 bp), the universal primer set, LCO 1490 (5’-GGTCAACAAATCATCATAAAGATATTGG-3’) and HCO 2198 (5’ GCTACAACATAATAAGTATCATG-3’) [6], was used in the following condition: initial denaturation at 94°C for 5 minutes, followed by 34 cycles of denaturation at 95°C for 1 minutes, annealing at 45.2°C for 1 minutes, extension at 72°C for 1 mintues, and a final extension at 72°C for 5 minutes. Amplification was performed using AccuPower PCR Premix (Bioneer, Daejeon, Republic of Korea) following the standard protocols. The PCR products were checked in 1.2% agarose gels, purified and sequenced at Macrogen, Inc. (Geumcheon-Gu, Seoul, Republic of Korea).
2.3. Data analysis
Intra-and inter-specific pairwise genetic distance and Neighbor joining (NJ) tree were conducted by using MEGA 7.0 under Kimura 2-parameter model [7]. To know evolutionary relationships among haplotypes of the sesiid species, the median joining (MJ) network was analyzed in Network ver. 10.2.0.0. A phylogenetic tree was constructed based on the haplotypes of Sesiinae detected in this study. Maximum likelihood (ML) tree was implemented by IQ-Tree 2.1.1 [8] with best-fit model, GTR+F+G4, according to the Bayesian information criterion [9,10]. Branch support values were produced by ultrafast bootstrap pseudoreplicates (UFB) [11] and a Shimodaira-Hasegawa approximate likelihood ratio test (SH-aLRT) [12]. Every test was with each 100,000 pseudoreplicates for UFB and SH-aLRT.
3. Results
3.1. Morphological identification
Two sesiid borers were identified as two species, Sphecodoptera sheni and Paranthrenella pinoakula sp. nov., based on adult and genitalic characteristics.
3.1.1. Taxonomic accounts
- Genus Sphecodoptera Hampson, [1893]
- Sphecodoptera Hampson, [1893]: 189. Type species. Sphecia repanda Walker
- Sphecia Hampson, 1919: 80.
- Sesia Spatenka, Lastuvka, Gorbunov, Tosevski & Arita, 1993: 87
- Spherodoptera Matsumura, 1931: 1017.
- Scasiba Matsumura, 1931: 8.
- Sphecodoptera sheni (Arita & Xu, 1994)
- Sesia sheni Arita & Xu, 1994: 61. Type locality. Nanjing, Jiangsu, China
- Scasiba caryavora Xu & Arita, 1994: 2.
- Sphecodoptera sheni Kallies, Arita, Owada & Wang, 2014: 587.
Material examined. Korea. GW: 2♀, Wonju, Moosil-dong 1710, 8.VII.2020, J-K. Jung; 2♂, 1♀, ditto, 12.VII.2022, J-K Jung, gen. slide no. IPE04, IPE05/S.Kim; 5exs, Chuncheon, Woodoo-dong 1121, 12.V.2023, B. Lee. GG: 9 exs, Seongnam, Bungdang-gu, Sampyong-dong, 15.VII.2022, JK Jung; 5, Goyang, Ilsan, Juyeob-dong 152, 6.V.2023, B. Lee; 6, Bucheon, Yeokgok-dong, 3. VI. 2023, B. Lee. CB, 2, Chungju, Heungdeok-gu, 28. IV.2023, S. Kim. Daegu, 6, Jung-gu, Gukchaebosang-ro 139gil 1, 8-12.IV.2023. I. Park.
Diagnosis. This species is superficially close to Scasiba rhynchioides in having yellowish scales on thorax, but it can be easily recognized by the 1st tergite of abdomen black with a narrow and yellow posterior band and the 2nd and 3rd tergites dark brown with reddish brown centrally (Figure 3E). The male genitalia of S. sheni is also differentiated from S. rhynchioides by a finger-like processus on the saccular margin antemedially (Figure 5E). Female genitalia are also distinguished from that by the long and narrow ovate corpus bursae bearing distinct signum (Figure 5G-H).
Host Plants. Carya illinoensis (Wang.) K. Koch (Juglandaceae); Quercus suber Linnaeus, Q. variabilis Blume, Q. acutissima Carruthers and Castanea mollissima Blume (Fagaceae) [13,14]; Quercus palustris Münchh. (this study).
Distribution. Korea (GG, GW, CB, Daegu) (this study), China (Jiangsu) [14].
Remark. Of the pin oak street borers, the large-sized sesiids were identified as S. sheni in this study. This species was found in Korea for the first time since it was reported form China. According to Arita et al. [14], this species has been reported as a pest of North American native pecan trees planted as street trees in Nanjing, China. Based on investigation of number of pupal exuviae, S. sheni was higher than other sessid borer density and their distribution on tree was aggregated between ground to 1m in heights.
- Genus Paranthrenella Strand, 1916
- Paranthrenella Strand, 1916: 47. Type Species. Paranthrenella formosicola Strand
- Paranthrenella pinoakula sp. nov. Kim
Type Material. Holotype: One male, Korea, GW, Wonju, Moosil-dong 1710, 2.VII.2022, JK Jung, gen. slide no. IPE03/ S. Kim; Paratypes: two males, ditto, 8.VII.2020, JK Jung.
Diagnosis. This species is externally similar to Paranthrenella formosicola in having a long and slender abdomen (Figure 4E). However, it can be readily distinguished from the former due to the lesser development of yellow scales between the thorax and tegula (Figure 4C), as well as the abdomen dorsally (Figure 4E). Except for the 4th and 7th tergite, the presence of yellow scales on the posterior margin of each tergite is not noticeable. In contrast, P. formosicola has thickly developed yellow markings on the posterior margin of the 2nd, 4th and 6th tergites in male and almost entire tergites in female. The male genitalia is also differentiated from that by the asymmetrical valva including a ridge line and aedeagus bearing four small cornuti at apex.
Description. Male (Figure 4). Wingspan. 22.0-28.0 mm; forewing length 10.0-12.5 mm; body length 17.0-19.5 mm. Head: frons pale brown; vertex dark brown; occipital fringe yellow; scape of antenna yellow entirely, shorter than 1/2 diameter of eye; flagellum blackish brown antemedial and apical parts, tinged with yellow postmedial part dorsally, short setose at apex, yellow ventrally; labial palpus entirely yellow; 3rd segment of labial palpus 2/3 length of 2nd segment. Thorax: patagium blackish brown, mixed with pale brown laterally; tegula blackish brown with yellow scales on posterior margin; mesothorax blackish brown with two spots of yellow scales on posterior margin; metathorax blackish brown. Forewing hyaline; costal margin blackish brown; dorsal margin yellowish brown; wing base blackish brown tinged with bright yellow; bright yellow between radius vein 2 (R2) and R3; cilia yellowish brown; forewing vernation: radius 4 (R4) stalked with R5 antermedially; anterior transparent area (ATA) extended to 3/4 of forewing; posterior transparent area (PTA) extended to 4/5 of forewing; apical 1/3 exterior transparent area tinged with yellowish brown scales. Abdomen blackish brown with yellow scales noticeably at posterior margin of tergite 4 (T4) and T7.
Male genitalia (Figure 6). Tegumen-uncus complex broad; scopula androconialis well-developed, slightly shorter than the length of tegumen-uncus complex; crista gnathi lateralis large, a cup-shaped; crista gnathi medialis relatively narrow, oval. Valvae elongated, oval, asymmetrical; right valva with asymmetric ridge line from basal crista sacculi to apex, rather dense setose followed the ridge from base to 1/3 of valva; left valva without ridge. Saccus with bifurcate terminal margin, a reverse blunt fork-shaped, as long as vinculum. Aedeagus as long as valva, gradually narrow from base to apex; vesica curved at sub-apex; bearing four cornuti.
Female genitalia. Unknown.
Host Plants. Quercus palustris Münchh. (this study).
Distribution. Korea (GW) (this study).
Etymology. The species name is derived from a common name of the host plant (Quercus palustris), pinoak- plus a Latin diminutive suffix, -ula, referring to the insect pest borer of the pin-oak.
Remark. In the pin oak street insect pests, the small-sized borers were identified as new species, P. pinoakula sp. n., in this study. This species was only found from WonJu (GW) in Korea, and their densities were relatively lower compared to S. sheni.
3.2. Molecular analyses
3.2.1. Genetic divergence and distribution
Genetic divergences within all species included in this study are presented in Table 1. Except for four species (Paranthrenella auriplena, P. cinnamoma, P. formosicola and P. terminalis) which having only one sequence, a total of 647 comparison pairs of four species (S. sheni, P. pinoakula sp. n., P. chrysophanes and Anatrachyntis japonica) were calculated within the species. The intraspecific genetic distance (GD) averaged 0.3% and the maximum intraspecific divergence did not exceeding 1.9%. These of S. sheni and P. pinoakula sp. n. were 1.7% and 0.3% respectively.
The highest value of intraspecific GD was found in P. chrysophanes with 1.9%. The interspecies GD was analyzed with 564 comparison pairs. The minimum interspecific GD among all seven species was calculated to be 8.1% between P. auriplena and P. chrysophanes, while the maximum interspecific genetic value was 16.8% (averaging 13.8%) between P. terminalis and P. chrysophanes. The distribution of their genetic divergence is shown in Figure 7. All species analyzed in this study exhibited a distinct barcode gap between the maximum of intraspecific GD and the minimum of interspecific GD in the COI gene.
3.2.2. NJ Tree analysis
NJ tree analysis, a heuristic method based on genetic distance, showed that S. sheni and P. pinoakula sp. n., branched into clade A and B, respectively (Figure 8). In clade A, Chinese S. sheni individual was the first to branch out and followed by the Wonju (GW) population. And then, each population of Goyang (GG) and Chuncheon (GW) were branched off, followed by the populations of Bucheon (GG) and Cheongju (CB), and all diverse populations were claded together. All individuals of P. pinoakula sp. n. were nested within the congeneric species in clade B. Of them, the P. pinoakula sp. n. was also genetically close to P. formosicola which was the allied species having similar morphology.
3.2.3. Haplotype analysis
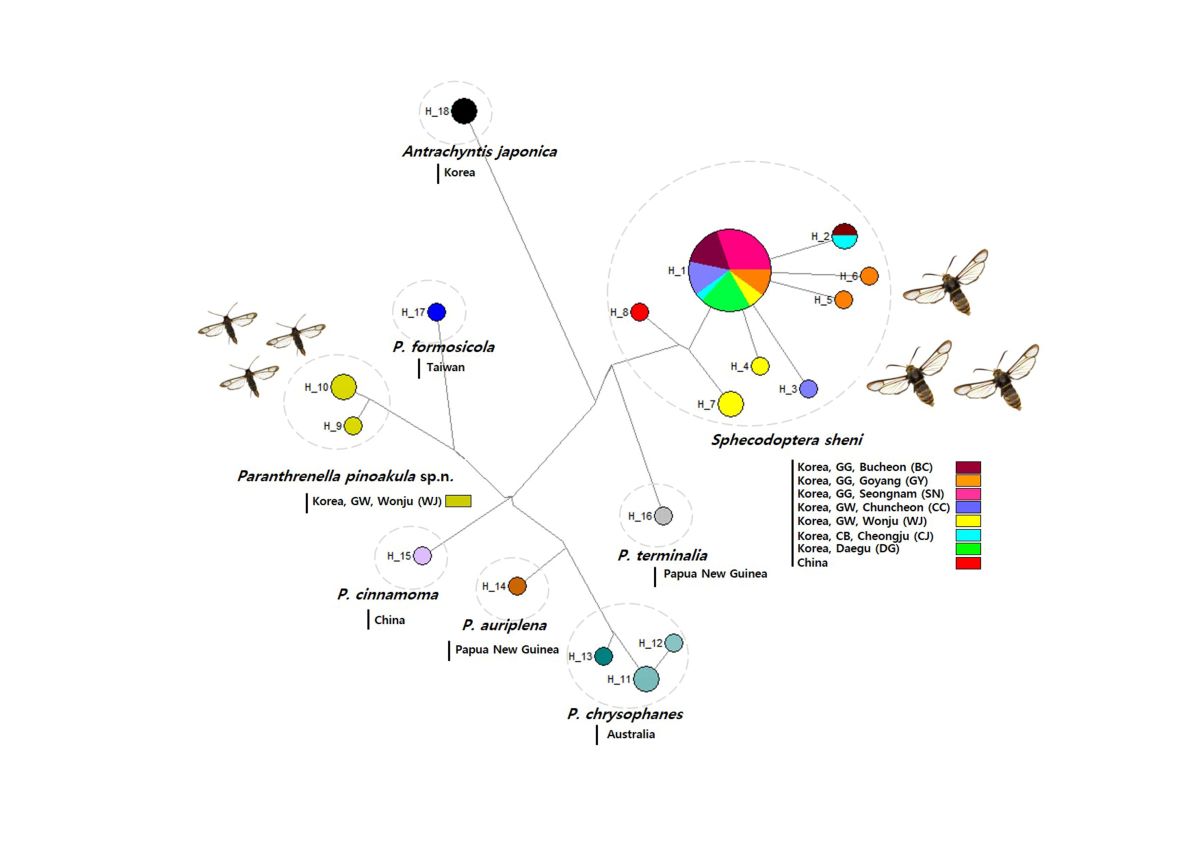
The MJ network analysis presented that eighteen haplotypes were detected from the 52 COI sequences (Figure 9). A total of 36 sequences of Korean clearwing borers were revealed total ten haplotypes. H1-H8 haplotypes of 34 sequences were recognized as Sphecodoptera sheni, and H9-H10 of three sequences were revealed as Paranthrenella pinoakula sp. n.
H1-H7 were detected from Korean samples (BC, GY, SN, CC, WJ, CJ, DG) with large-sized borers, S. sheni. WJ and GY populations of S. sheni were revealed as three haplotypes, and among them, H7 haplotype (WJ) was closely connected with H8 haplotype of Chinese S. sheni. BC and CJ populations of that werzze presented as two haplotypes, H1 and H2. CC population of that was recognized as two haplotypes, H1 and H3, whereas SN and Daegu populations were detected in only H1. H9 and H10 haplotypes were detected as the other Korean Wonju population of small-sized borers, P. pinoakula sp. n., and closely located with each congeneric species.
3.2.4. Phylogenetic tree
Although one gene is not enough to analyze a phylogenetic tree, Maximum Likelihood tree (MLtree) was constructed to show the phylogenetic relationship between the species and individual under species-level. The MLtree is derived from a statistical method based on evolutionary models including assumptions about the probability distribution of the data. As a result, all individuals of the large-sized clearwing moth borer, Sphecodoptera sheni, were separated from the other Paranthrenella spp. (Figure 10, clade A: Sh-aLRT/ UFB/ PP: 98.8/100/100). The small-sized clearwing moth borer, P. pinoakula sp. nov. was also phylogenetically claded within the genus Paranthrenella (Figure 10, clade B: Sh-aLRT/ UFB/ PP: 100/100/100). The species is closely located to the clade of P. formosicola and P. terminalia.
4. Discussion
The purpose of this study is twofold. The first objective is to identify the recent outbreak of invasive insect pests attacking pin-oaks, one of the popular street tree species planted in Korea. The pin-oak borers are actually referred to as a variety of clearwing moths, e.g. Scasiba rhynchioides (Butler), Synanthedon bicingulata (Staudinger), S. quercus (Matsumura) that are similar in appearance. In morphological identification based genitalic characters, pin-oak street tree borers identified as Sphecodoptera sheni and Paranthrenella pinoakula sp.nov..
We unexpectedly confirmed that there are two invasive species and that they are distinguished by body size. COI gene-based molecular identification also supported the two clearwing borers showing a distinct barcode gap within and between species. Haplotype analysis suggests that S. sheni may expand its distribution from China. The outbreak was centered in WJ (GW) and GY (GG) in having each three haplotypes, to neighboring areas of CC (GW), BC (GG) and CJ (CB) with each two haplotypes. Relatively, both SN (GG) and Daegu populations are detected to H1 haplotype, inferring a recent outbreak. On the other hand, for P. pinoakula sp. nov., only the WJ (GW) population was confirmed. In addition, our phylogenetic analysis (MLtree) shows that the two borer groups are phylogenetically distant, indicating that they may be occurred by host preference of the species rather than the occurrence of phylogenetically related groups.
Author Contributions
Conceptualization, S.K.; methodology, S.K. and J.J.; software, S.K.; formal analysis, S.K.; resources, S.K., J.J., I.P., B.L. and Y.K.; data curation, S.K.; writing—original draft preparation, S.K.; writing—review and editing, S.K., J.J., I.P., B.L. and Y.K.; visualization, S.K.; supervision, S.K.; project administration, S.K.; funding acquisition, S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This paper was supported by research funds for newly appointed professors of Jeonbuk National University in 2021. This work was also supported by Korean Institute of Planning and Evaluation for Technology in Food, Agriculture and Forestry (IPET) through Agriculture, Food and Rural Affairs Convergence Technologies Program for Educating Creative Global Leader Program (or Project), funded by Ministry of Agriculture, Food and Rural affairs (MAFRA) (no.32100103). Additionally, this work was supported by a grant from the National Institute of Biological Resources (NIBR), funded by the Ministry of Environment (MOE) of the Republic of Korea (NIBR202304203) and R&D Program for Forest Science Technology (Project No. 2019151D10-2323-0301) provided by Korea Forest Service (Korea Forestry Promotion Institute).
Acknowledgments
We are many thanks to Jinwook Kim, Jinseong Park, In-Won Jeong, Byeong Jong Lee and Seul Ki Son for their fieldworks.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; Økland, B. Complex responses of global insect pests to climate warming. Frontiers in Ecology and the Environment 2020, 18, 141–150. [Google Scholar] [CrossRef]
- World Economic Forum. Global risks report 2023. 18th Edition. Insight report; World Economic forum: Switzerland. 2023; p. 98.
- Pühringer, F; Kallies, A. Provisional checklist of the Sesiidae of the world (Lepidoptera: Ditrysia). Mitteilungen der Entomologischen Arbeisgemeinschaft Salzkammergut 2004, 4, 1-85. [CrossRef]
- Pühringer, F; Kallies, A. Provisional checklist of the Sesiidae of the world (Lepidoptera: Ditrysia). Mitteilungen der Entomologischen Arbeisgemeinschaft Salzkammergut 2004, 4, 1-85.
- Kim, S.; Lee, W.; Lee, S. Estimation of a new molecular marker of the genus Stathmopoda (Lepidoptera: Stathmopodidae): Comparing EF1a and COI sequences. Journal of Asia-Pacific Entomology, 2017, 20, 269–280. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 1994, 3, 294–299. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Molecu lar Biology and Evolution 2016, 33(7), 1870–1874. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmid, H.A.; Chernomor, O.; Dominik, S.; Michael, D.W.; Arndt, V.H.; Robert, L. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Molecular Biology and Evolution, 2020, 37 (5), 1530–1534. [Google Scholar] [CrossRef]
- Chernomor, O.; Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Systematic Biology 2016, 65 (6), 997–1008. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.; Jermiin, L.S. ModelFinder: fast model selection for accurate phylogenetic estimates. Nature methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: improving the ultrafast bootstrap approximation. Molecular Biology and Evolution, 2018; 35, 518–522. [Google Scholar] [CrossRef]
- Anisimova, M.; Gil, M.; Dufayard, J.F.; Dessimoz, C.; Gascuel, O. Survey of branch support methods demonstrates accuracy, power, and robustness of fast likelihood-based approximation schemes. Systematic Biology 2011, 60, 685–699. [Google Scholar] [CrossRef]
- Shen, B.Y. Primary study of Aegeria molybdoceps Hampson. Plant protection, 1988, 3, 23–25. [Google Scholar]
- Arita, Y; Xu, Z-G.; Liu, Y-Q. A new Sesia (Lepidoptera, Sesiidae), clearwing borer on Pecan from Nanjing, China. Tinea 1994, 14 (1), 61-64.
Figure 1.
Panoramic view of a pin oak (Quercus palustris Munchh.) street trees. (A) Daegu metropolitan city; (B), Ilsan-gu, Goyang-si, Prov. Gyeonngi.
Figure 1.
Panoramic view of a pin oak (Quercus palustris Munchh.) street trees. (A) Daegu metropolitan city; (B), Ilsan-gu, Goyang-si, Prov. Gyeonngi.

Figure 2.
diverse damage on pin oak street trees by clearwing moths. (A) reddish-brown frass by larva; (B) milky sap damage from cavity in xylem made by larva; (C) xylem cambium fed by larva; (D) pupal frame in cambium; (E) left distinctive marks on the bark after emergence; (F) zoom in exuviae after larva emergence; (G) clearwing moth larva.
Figure 2.
diverse damage on pin oak street trees by clearwing moths. (A) reddish-brown frass by larva; (B) milky sap damage from cavity in xylem made by larva; (C) xylem cambium fed by larva; (D) pupal frame in cambium; (E) left distinctive marks on the bark after emergence; (F) zoom in exuviae after larva emergence; (G) clearwing moth larva.

Figure 3.
Adult of Sphecodoptera sheni. (A) Head; (B) flagellum of antenna; (Ba) apical part of flagellum; (C) entire view; (D) wings; (E) abdomen; (F) hind leg; (G) tarsal claw of hind tarsus; scale bar indicates wingspan.
Figure 3.
Adult of Sphecodoptera sheni. (A) Head; (B) flagellum of antenna; (Ba) apical part of flagellum; (C) entire view; (D) wings; (E) abdomen; (F) hind leg; (G) tarsal claw of hind tarsus; scale bar indicates wingspan.

Figure 4.
Adult of Paranthrenella pinoakula sp. nov. (A) Head; (B) flagellum of antenna; (Ba) apical part of flagellum; (C) entire view; (D) wings; (E) abdomen; (F) hind leg; (G) tarsal claw of hind tarsus; scale bar indicates wingspan.
Figure 4.
Adult of Paranthrenella pinoakula sp. nov. (A) Head; (B) flagellum of antenna; (Ba) apical part of flagellum; (C) entire view; (D) wings; (E) abdomen; (F) hind leg; (G) tarsal claw of hind tarsus; scale bar indicates wingspan.

Figure 5.
Genitalia of Sphecodoptera sheni. (A) Male genitalia; (B) aedeagus; (C) apical part of aedeagus including cornutus; (D) uncus; (E) a finger-like processus on the saccular margin of valva; (F) saccus; (G) Female genitalia; (H) signum in corpus bursae; scale bar: 0.25 mm.
Figure 5.
Genitalia of Sphecodoptera sheni. (A) Male genitalia; (B) aedeagus; (C) apical part of aedeagus including cornutus; (D) uncus; (E) a finger-like processus on the saccular margin of valva; (F) saccus; (G) Female genitalia; (H) signum in corpus bursae; scale bar: 0.25 mm.
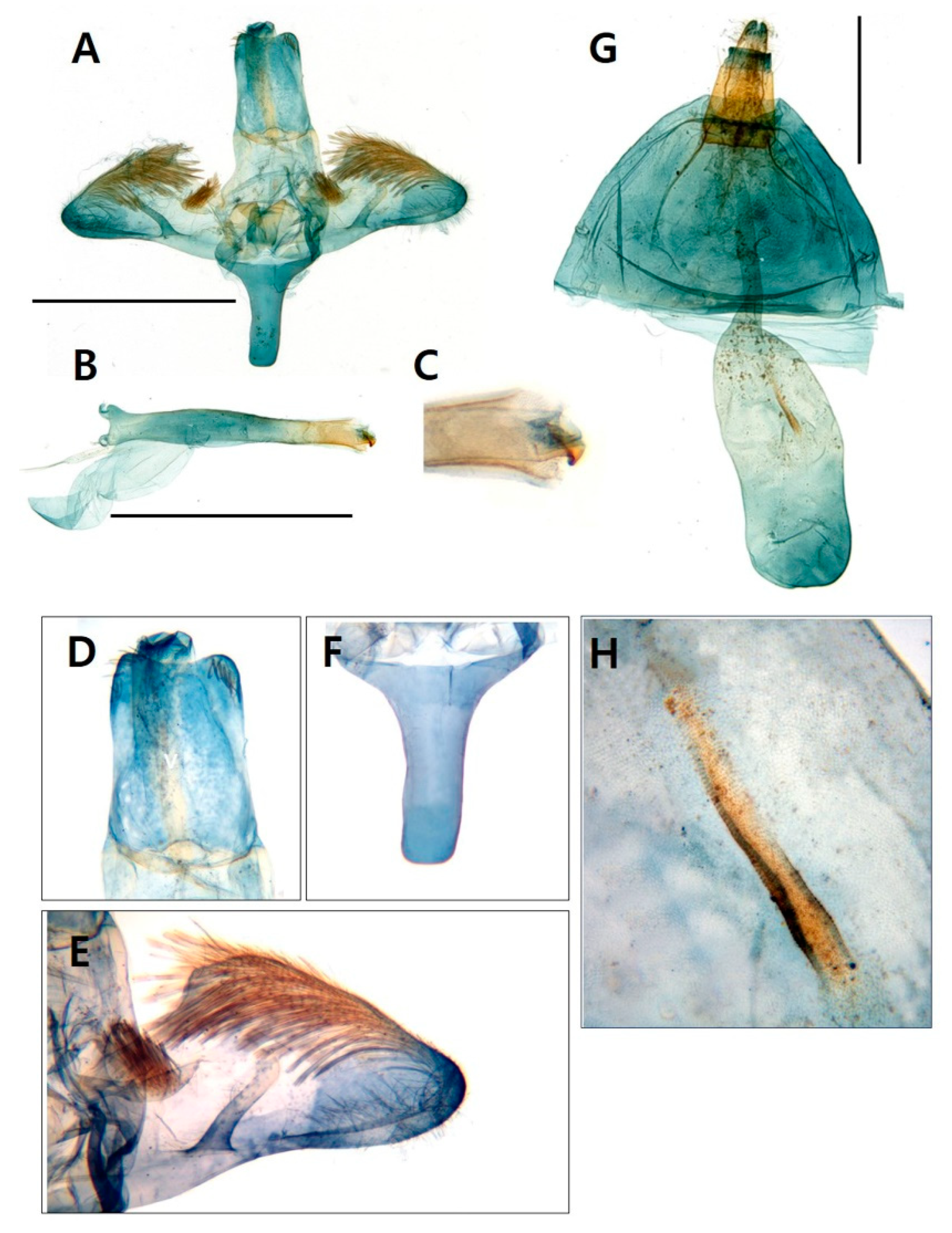
Figure 6.
Genitalia of Paranthrenella pinoakula sp. nov. (A) Male genitalia; (B) aedeagus; (C) apical part of aedeagus including cornute; (D) scopula androconialis; (E) medial gnathos flap; (F) right valva with a ridge line; (G) right valva; (H) saccus; (I) sternum I-VII segments; (J) sternum VIII segmenbbt; scale bar: 0.25 mm.
Figure 6.
Genitalia of Paranthrenella pinoakula sp. nov. (A) Male genitalia; (B) aedeagus; (C) apical part of aedeagus including cornute; (D) scopula androconialis; (E) medial gnathos flap; (F) right valva with a ridge line; (G) right valva; (H) saccus; (I) sternum I-VII segments; (J) sternum VIII segmenbbt; scale bar: 0.25 mm.
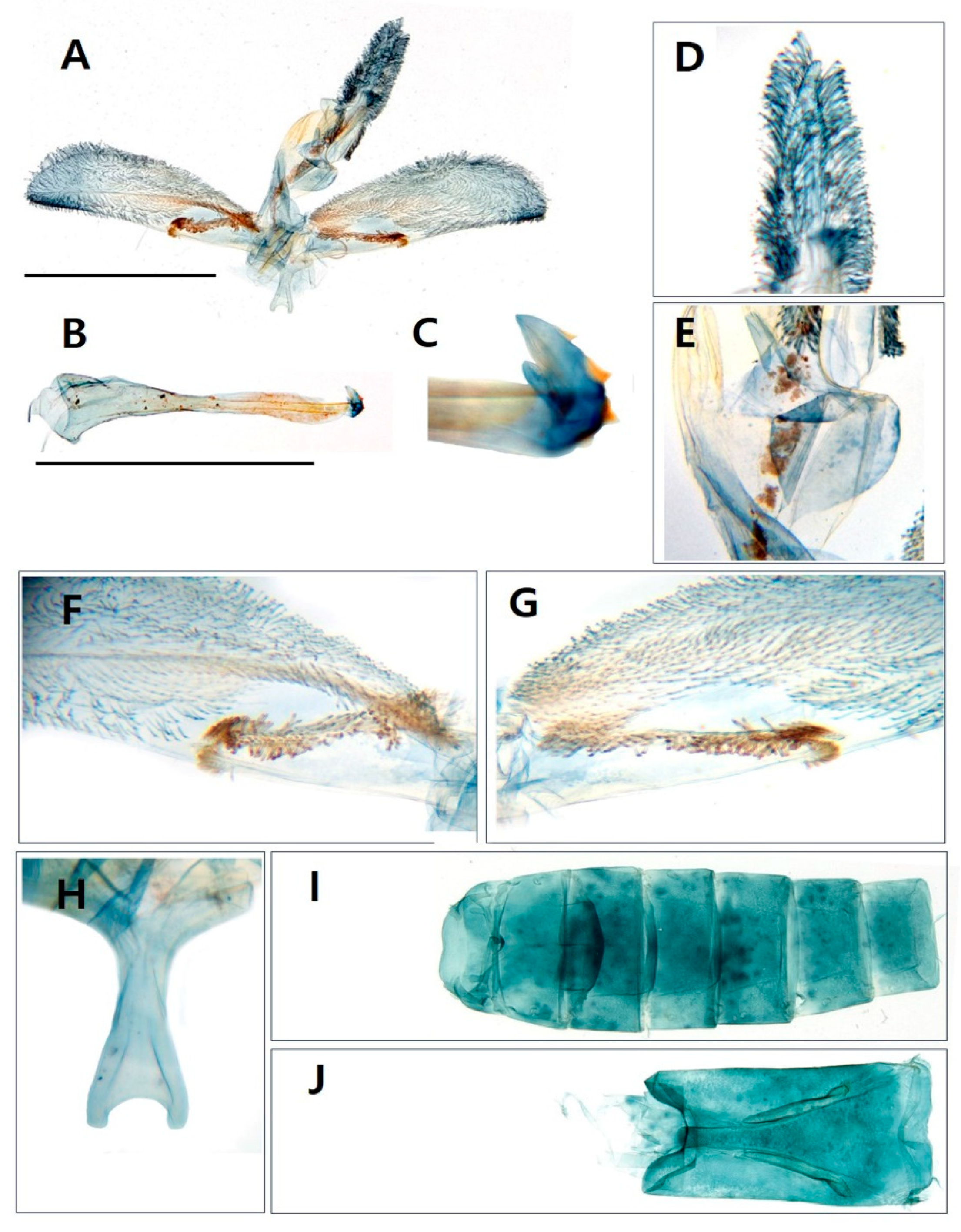
Figure 7.
Distribution of genetic divergences based on the Kimura-2 parameter model for COI sequences according to taxonomic levels. 647 comparison pairs within the species and 564 comparison pairs between the species were calculated.
Figure 7.
Distribution of genetic divergences based on the Kimura-2 parameter model for COI sequences according to taxonomic levels. 647 comparison pairs within the species and 564 comparison pairs between the species were calculated.
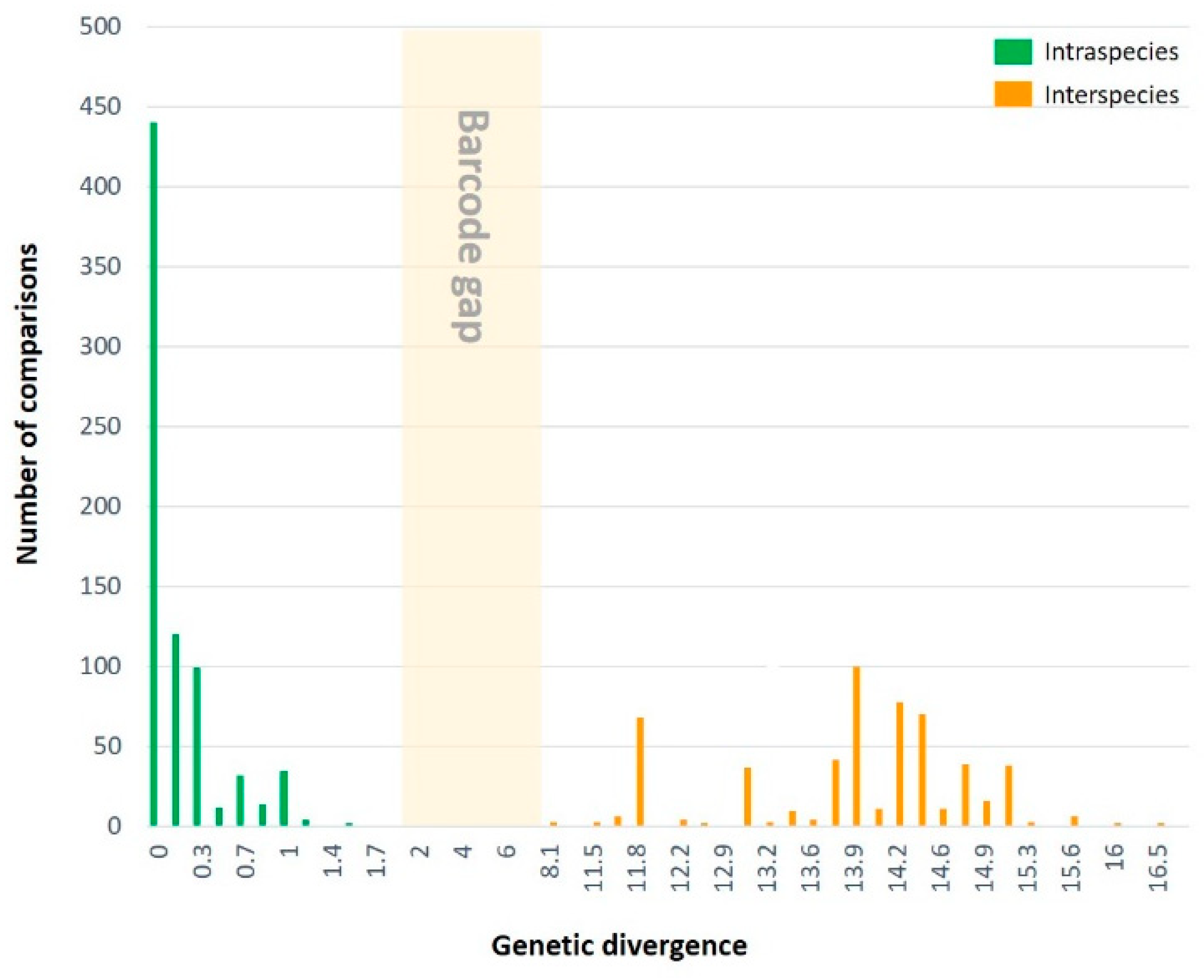
Figure 8.
Neighbor joining tree of COI gene of Sesiid species. Numbers under the branches are bootstrap percentages (%).
Figure 8.
Neighbor joining tree of COI gene of Sesiid species. Numbers under the branches are bootstrap percentages (%).
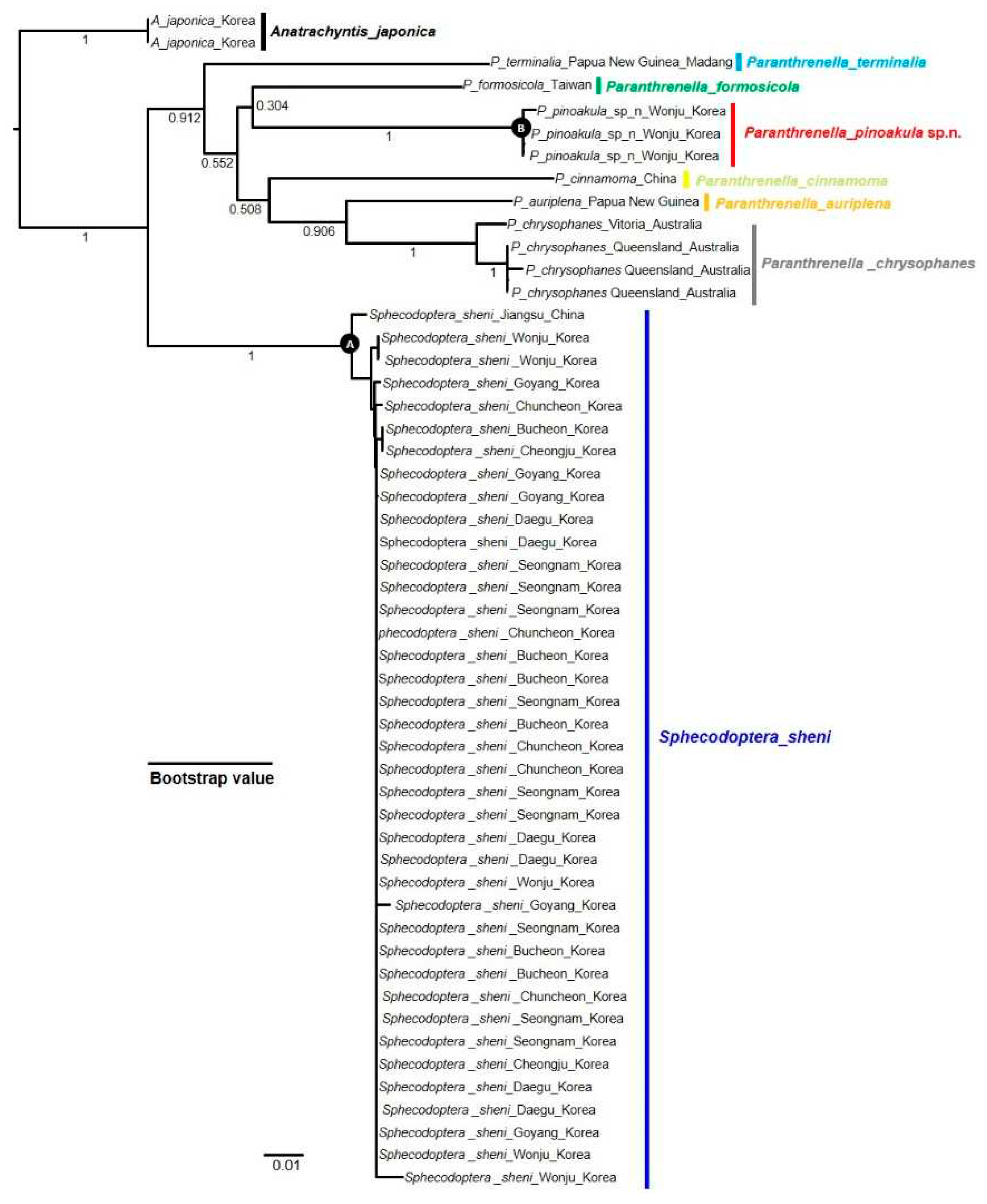
Figure 9.
Median joining network about 18 haplotypes of the 52 COI sequences. The pie size is proportional to the haplotype frequency. A dotted circle means same species.
Figure 9.
Median joining network about 18 haplotypes of the 52 COI sequences. The pie size is proportional to the haplotype frequency. A dotted circle means same species.
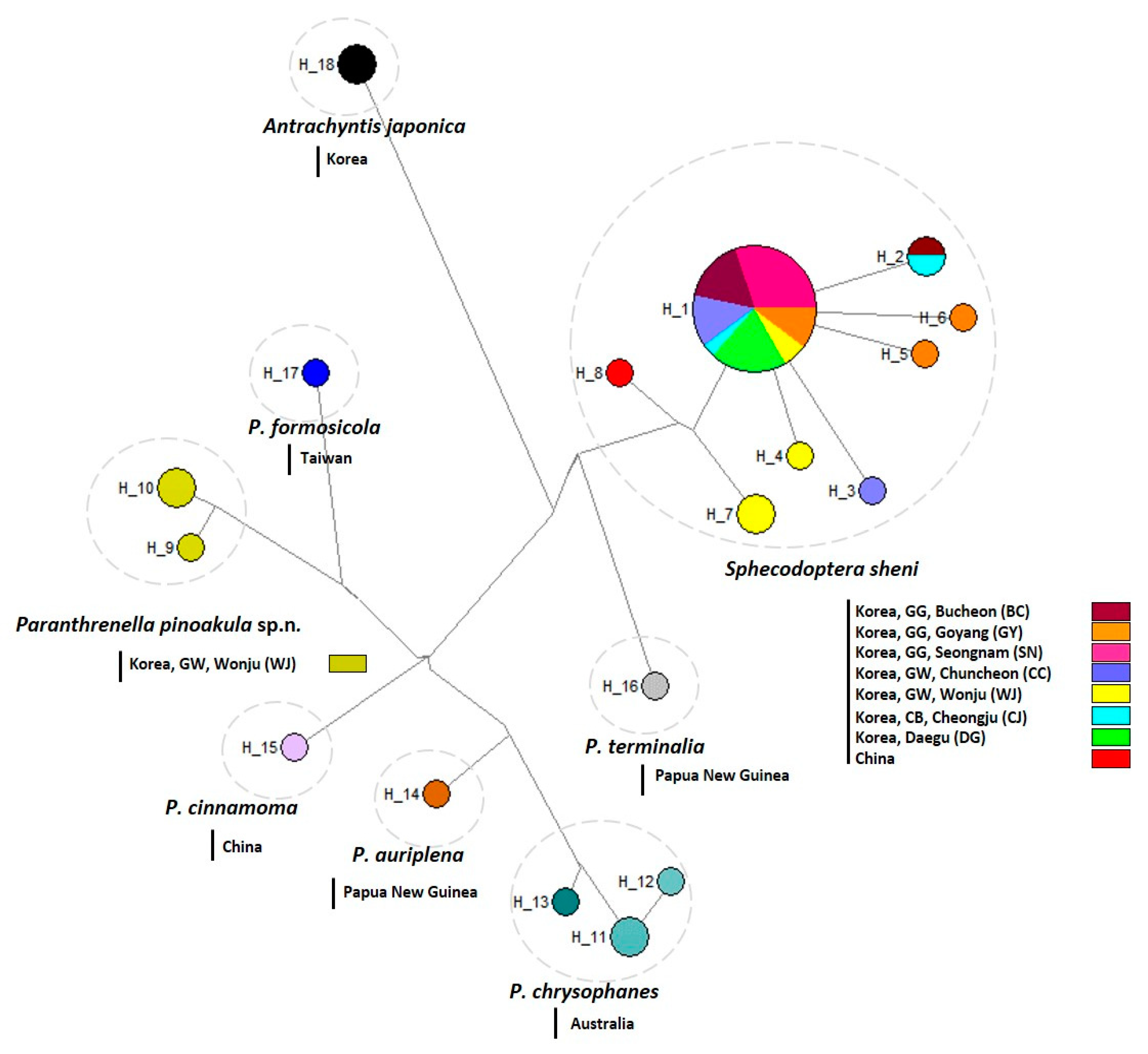
Figure 10.
Maximum likelihood tree of Sesiid species. Each node marked with three types of supporting value: Shimodaira-Hasegawa approximate likelihood ratio test (SH-aLRT): upper left; Ultrafast bootstrap (UFB) pseudoreplicates: upper right; Maximum likelihood (ML) bootstrap: below the branch.
Figure 10.
Maximum likelihood tree of Sesiid species. Each node marked with three types of supporting value: Shimodaira-Hasegawa approximate likelihood ratio test (SH-aLRT): upper left; Ultrafast bootstrap (UFB) pseudoreplicates: upper right; Maximum likelihood (ML) bootstrap: below the branch.
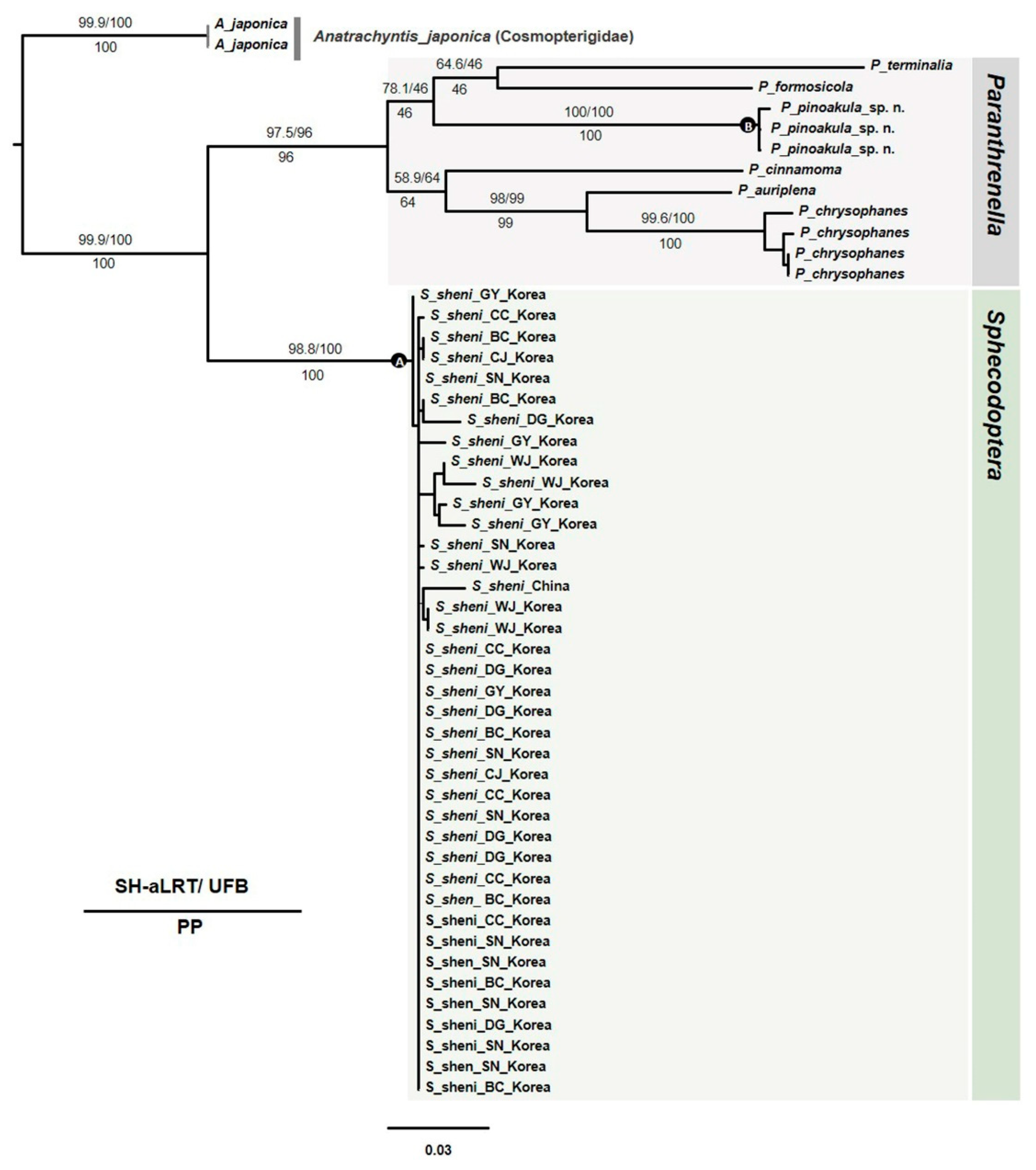

Table 1.
Genetic divergences within all species included in this study.
| 1. Different taxonomic levels | ||||
| Taxonomic level (No. of Comparison Pairs) | K2P Pairwase distances | |||
| Maximum | Minimum | Average | ||
| Within species (647) | 1.9 | 0.0 | 0.3 | |
| Between species (564) | 16.8 | 8.1 | 13.8 | |
| 2. Intraspecific genetic divergence | ||||
| Species | Comparison pairs | Intraspecific genetic divergence | ||
| (CP) | Maximum | Minimum | Average | |
| Sphecodoptera sheni | 637 | 1.7 | 0.0 | 0.2 |
| Paranthrenella pinoakulasp.n. | 3 | 0.3 | 0.0 | 0.2 |
| Paranthrenella auriplena | - | - | - | - |
| Paranthrenella cinnamoma | - | - | - | - |
| Paranthrenella chrysophanes | 6 | 1.9 | 0.0 | 0.9 |
| Paranthrenella formosicola | - | - | - | - |
| Paranthrenella terminalis | - | - | - | - |
| Anatrachyntis japonica | 1 | 0.0 | 0.0 | 0.0 |
| 3. Interspecific genetic divergence | ||||
| species | Sphecodoptera sheni | P. pinoakulasp. n. | P. auriplena | P. cinnamoma |
| Sphecodoptera sheni | CP=114, 14.6(14.2-15.3) | CP=38, 14.2(13.9-14.9) | CP=38, 15.1(14.9-15.8) | |
| Paranthrenella pinoakulasp.n. | CP=3, 13.9(13.9-13.9) | CP=3, 13.0(13.0-13.0) | ||
| Paranthrenella auriplena | CP=1, 11.5(11.5-11.5) | |||
| Paranthrenella cinnamoma | ||||
| Paranthrenella chrysophanes | ||||
| Paranthrenella formosicola | ||||
| Paranthrenella terminalis | ||||
| Anatrachyntis japonica | CP=78, 11.8(11.5-12.5) | CP=6, 14.9(14.9-14.9) | CP=2, 13.7(13.7-13.7) | CP=2, 15.6(15.6-15.6) |
| species | P. chrysophanes | P. formosicola | P. terminalis | |
| Sphecodoptera sheni | CP=152, 14.0(13.7-14.9) | CP=38, 13.1(12.9-13.7) | CP=38, 13.7(13.4-14.4) | |
| Paranthrenella pinoakulasp.n. | CP=12, 13.4(13.0-13.7) | CP=3, 11.8(11.7-12.0) | CP=3, 14.3(14.2-14.4) | |
| Paranthrenella auriplena | CP=4, 8.2(8.1-8.4) | CP=1, 13.2(13.2-13.2) | CP=1, 14.9(14.9-14.9) | |
| Paranthrenella cinnamoma | CP=4, 138(13.7-13.9) | CP=1 12.2(12.2-12.2) | CP=1, 14.4(14.4-14.4) | |
| Paranthrenella chrysophanes | CP=4, 14.4(14.2-14.6) | CP=4, 16.5(16.3-16.8) | ||
| Paranthrenella formosicola | CP=1, 12.2(12.2-12.2) | |||
| Paranthrenella terminalis | ||||
| Anatrachyntis japonica | CP=8, 15.6(15.3-16.0) | CP=2, 14.9(14.9-14.9) | CP=2, 14.1(14.1-14.1) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



