Submitted:
09 December 2023
Posted:
11 December 2023
You are already at the latest version
Abstract
The molecular mechanisms underlying insect gall formation remain unclear. A major reason for the inability to identify the responsible genes is that most insect galls are produced on woody plants and therefore cannot be experimentally validated in the laboratory. To overcome these problems, we established a new galling insect model, Smicronyx madaranus. Our manipulation experiments using nail polish sealing and insecticide treatment revealed an age-dependent change in gall formation during S. madaranus; adult females and larvae are responsible for gall induction and enlargement, respectively. Furthermore, it has been suggested that substances released during oviposition and larval feeding are involved in each process. Phylogenetic analysis showed that most gall-forming weevils, including S. madaranus, belong to a single lineage, suggesting that gall-forming traits evolved in a specific lineage within Smicronyx. The efficacy of RNA interference (RNAi) in S. madaranus was confirmed by targeting multicopper oxidase 2 gene. It is expected that the mechanisms of gall formation will be elucidated by a comprehensive functional analysis of candidate genes using RNAi and the S.madaranus galling system in the near future.
Keywords:
Insect gall
; manipulation of galling insect
; field dodder
; phylogeny of the genus Smicronyx
; RNA interference
1. Introduction
Some insects induce abnormally enlarged tissues called “galls” in plants [1,2]. Galls function as shelters for inducers and protect them from natural enemies and environmental stressors. They also serve as feeding sites for gall-forming insects [3,4]. Therefore, the gall is conceivable as an “extended phenotype” of insects [5]. However, the mechanisms underlying gall formation in insects remain unclear. The phytohormones auxins and cytokinins biosynthesized in insects have been proposed to be important contributors to the process of gall formation [6,7]. In addition, certain protein effectors potentially released by insects are candidates for initiating plant gall formation [8,9,10,11]. However, the functions of potential protein effectors have only been implicated by RNA-seq and genomic analyses, and the causal relationship between candidate genes and gall formation has not been confirmed. A major reason for the inability to identify genes involved in gall formation is that most insect galls are produced on woody plants [12,13] and therefore cannot be experimentally validated in the laboratory. To overcome these problems, a model system for plants and insects that can be manipulated in the laboratory is required.
Most weevils of the genus Smicronyx feed on parasitic plants of the genus Cuscuta, and some species form galls on plants [14,15]. Smicronyx madaranus Kono, 1930 feeds on C. campestris and forms spherical fruit-like galls on it [16]. In a previous study, we established a stable maintenance system for S. madaranus and C. campestris in the laboratory as a new galling model system and investigated their gall-forming behavior, gall formation process, and histochemical and physiological features [17]. The study showed that adult females bore holes in the nodes of C. campestris with their long rostrum and lay eggs inside the holes; then, the tissues of the nodes enlarged and transformed into spherical galls (Movie S1). The larvae grow by feeding on the starch-rich tissue inside the gall and emerged in approximately 12 days. Notably, S. madaranus-induced galls greatly increase the photosynthetic activity of the holoparasitic plant C. campestris which depends on the host plant for its nutrients and shows low photosynthetic activity [17]. This is a specific feature of S. madaranus-induced galls, in contrast to the general galls formed on the leaves of various plants in which photosynthetic gene expression is suppressed [18,19,20]. This implies that general plant galls undergo a transformation from a nutrient-producing organ (source) to a nutrient-receiving organ (sink), whereas S. madaranus-induced galls undergo the opposite transformation. Therefore, it is important to elucidate the mechanism of gall formation in S. madaranus to understand the commonality and diversity among insect galls.
In this study, as a preliminary step towards elucidating the molecular mechanisms involved in gall formation, we attempted to clarify the growth stages of the weevil involved in gall induction and enlargement through manipulation experiments on S. madaranus. We also clarified the phylogenetic position of S. madaranus in relation to other gall-forming and non-gall-forming species. In addition, the efficacy of RNAi in S. madaranus as a functional gene analysis method was evaluated by targeting the multicopper oxidase 2 (MCO2) gene, a phenotypic marker gene.
2. Materials and Methods
2.1. Insect and Plant
Laboratory strain of S. madaranus were collected from Neagari Nomi City, Ishikawa, Japan. They were maintained on C. campestris parasitizing Nicotiana benthamiana in the laboratory at 28 ˚C in the long day regimen (14L10D). Cuscuta campestris (with three to five shoots, approximately 10 cm long) parasitizing Vicia faba (approximately 25 cm long) was used for some experiments, as described later. Rearing and cultivation conditions were the same as those described previously [17]. To increase survival from the pupal to adult stages, the following modifications were made: Last instar larvae emerging from the galls were collected in a petri dish S35-DC12 (Fine Plus International, Kyoto, Japan) containing vermiculite moistened with water. The petri dish was placed in a container containing a saturated saline solution to maintain a humidity of 75%. The entire container was covered with aluminum foil to keep it dark and stored at 28˚C. Individuals that metamorphosed into pupae were transferred to new petri dishes lined with paper. The petri dishes were then placed back into the container to complete the pupal-to-adult metamorphosis.
2.2. Observation of S. madaranus in the Initial Gall
Initial galls (1.5–2 mm in width, Figure 1A) appeared approximately 3 days after weevil introduction on C. campestris [17]. Twenty initial galls were collected from a laboratory-maintained colony. We then dissected the galls with forceps under a stereomicroscope M165C (Leica, Wetzlar, Germany) and observed the weevils located inside the gall.
2.3. Ovipositor Sealing Experiment
It is difficult to distinguish between the sexes of S. madaranus based on appearance. Therefore, we identified females for this experiment by observing their mating behavior as follows: Twenty young adults were kept in a small petri dish FALCON no. 35100 (60×15 mm) (Corning, NY, USA) and allowed to mate for 2 days. During this period, five shoots of C. campestris cut into 10 cm lengths were placed in the petri dish as food. Shoots were replaced daily. Individuals on the underside during mating were considered females and used in the experiment. To inhibit oviposition, white nail polish was applied with a toothpick to cover the tail ends of the females (Figure S1A) under a stereomicroscope M165C (Leica). In the control group, the same nail polish was applied to the elytra.
To determine whether the sealing treatment affected the locomotor activity, we measured and compared the distance between sealed and control individuals. One day after treatment, weevils were placed in a container made of a silicon O-ring (17.8 mm inner diameter) and cover glass (18 mm × 18 mm, Matsunami Glass Industry, Osaka, Japan), and placed on a white LED light (3.2 mW/m2) (Figure S1B). Their behavior was then recorded directly for 5 min using a Camera Module v.2.1 connected to a Raspberry Pi 3 Model B (Raspberry Pi Foundation, Cambridge, UK) (Movie S2). The video was analyzed using the original source code written in R language [22] v.4.2.1. The script code detected the points at which the weevils were located at any given moment and calculated the distance they had moved by summing the distances between the points at different times. The experiments were repeated thrice. Ten individuals from each treatment group were used for each experiment.
Ovipositor-sealed and control weevils were individually placed in a box containing C. campestris parasitizing V. faba seedlings and maintained for four days. Then, 20 nodes of C. campestris were cut and collected from the box. The nodes were examined for the egg-laying holes and gall formation using a Leica M165C stereomicroscope.
2.4. Effects of the Larva on Gall Formation
Ten young adult weevils (of mixed sex) were placed in a box containing three seedlings of V. faba parasitized with C. campestris and allowed to form galls. All the weevils were removed from the boxes after 48 h. The following two insecticides were then applied to the emerged initial galls to eliminate any larvae present inside; Dantotsu water solvent insecticide (Sumitomo Chemical, Tokyo, Japan) is a neonicotinoid-based agent containing 16.0% of clothianidin; Ortolan granules (Sumitomo Chemical) is an organic phosphorus-based agent containing 50.0% of acephate. These insecticides have different modes of action; therefore, it is unlikely that both insecticides would directly affect the gall formation mechanisms in C. campestris. Both insecticides were diluted with sterile water to 1/1000 (v/v) and Squash Adhesive [final concentration: 0.1% (v/v)] (Maruwa Biochemical, Tokyo, Japan) was added to improve penetration into the gall. Two microliters of each insecticide solution were applied to the galls using a micropipette. The control group was treated with 0.1% Squash Adhesive. The galls were marked with markers. To confirm the larval status inside the galls, five days after treatment, the galls were cut with a laser blade, and the interiors were observed using a Leica M165C stereomicroscope. Twenty-four, 21, and 17 galls were examined in the control, Dantotsu, and Ortholan treatments, respectively. The effect of insecticide treatment on gall enlargement was analyzed as follows. Images of the galls were captured with a ruler every 24 h using a digital camera (Stylus TG-3 Tough; Olympus, Tokyo, Japan). Gall widths were calculated by image analysis using Fiji [23] and compared between the treatment and control groups.
2.5. Molecular Phylogenetic Analysis of S. madaranus and Its Related Species
DNA was isolated from samples (Table S1) using a conventional phenol extraction method. The purified DNA was resuspended in 20 µl of TE buffer [10mM Tris-HCl (pH 8.0), 0.1 mM EDTA]. Partial sequences of cytochrome c oxidase subunit I (COI) were amplified using KOD-FX Neo DNA polymerase (Toyobo, Osaka, Japan) with the primers LCO1490 and HCO2198 (Table S2). The PCR was carried out with the following temperature profile: 94 ℃ for 2 min, followed by 35 cycles of 98 ℃ for 10 s, 50 ℃ for 30 s, and a final extension at 68 ℃ for 30 s. The PCR products were purified using a polyethylene glycol solution (PEG6000 20% and 2.5 mM). The PCR amplicons were directly sequenced in both directions with the same primers using a BigDye Terminator v3.1 Cycle Sequencing Kit and Applied Biosystems 3500 Genetic Analyzer (Thermo Fisher Scientific, MA, USA). The resulting sequences were assembled and analyzed using the ATGC ver. 4 software (GENETYX, Tokyo, Japan).
For the molecular phylogenetic analysis, the COI sequences of four species in Japan and previously published sequences of the genus Smicronyx were included in the dataset, and Tychius capucinus and T. breviusculus were used as outgroups. The dataset included five gall-forming species (S coecus, S. guineanus, S. jungermanniae, S. madaranus, and S. smreczynskii) [16,24,25,26,27,28] and 18 species (S. albosquamosus, S. australis, S. dentirostris, S. fallax, S. gracillipes, S. japonicus, S. jordanicus, S. longitarsis, S. lutulentus, S. nebulosus, S. pauperculus, S. pseudocoecus, S. reichi, S. rubricatus, S. rufus, S. san, S. syriacus, and S. zonatus) with no description of gall formation [15,16,24,25,29,30,31,32,33,34,35]. Analyses were performed using Bayesian inference (BI), maximum-likelihood (ML) and maximum parsimony (MP) criteria. For BI, the most appropriate model of sequence evolution (TN93+G model) was selected using the MEGA X model-selection option [36]. The parameters for the selected substitution model were estimated using the data. A total of 100,000 trees were obtained (ngen = 10,000,000, sample freq = 100) using MrBayes 3.2.7 [37], and the first 25% of these trees (25,000) were considered ‘burned in’ and discarded. A consensus tree was constructed from the remaining trees based on a 50%-majority-rule. Two independent runs were performed using the same sequence-evolution model. For ML, bootstrap analysis of 1,000 replications was performed based on the same model as BI in MEGA X. Initial trees for the heuristic search were automatically obtained by applying the Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood approach. For MP, all characters were included and weighed equally, and 1,000 bootstrap replicates were performed using MEGA X. The Subtree-Pruning-Regrafting algorithm was used with search level 1, where the initial trees were obtained by the random addition of sequences (10 replicates). Gall formation traits were mapped to the resulting phylogenetic tree.
2.6. Expression Analysis and RNAi for the S. madaranus Multicopper Oxidase 2 Gene (SmMCO2)
Two candidate MCO2 genes (hereafter SmMCO2A and SmMCO2B) were identified in S. madaranus by RNA sequencing, which was performed by a collaborator (K. Bessho-Uehara, in preparation), and confirmed by Sanger sequencing (Figures S2 and S3). Samples used for gene expression analysis were prepared as follows: Weevils were collected from stages P2 to A3, as defined in this study (Figure S4). Total RNA was extracted from each sample, consisting of three mixed-sex individuals, using the NucleoSpin RNA XS Kit (Takara-Bio, Shiga, Japan). Using 300 ng of total RNA, 20 µl of cDNA was synthesized using the PrimeScriptⅡ 1st strand cDNA Synthesis Kit (Takara-Bio) and the Random 6 mer primer supplied with the kit. Quantitative PCR of the SmMCO2 genes was performed using THUNDERBIRD Next SYBR qPCR Mix (Toyobo) and Mx3005P (Agilent Technologies, Santa Clara, CA, USA) with specific primer sets, as shown in Table S2. In this study, we analyzed the expression of both SmMCO2A and SmMCO2B using a single primer set, without distinguishing between them (Table S2). The PCR temperature profile was 95℃ for 1 min, followed by 40 cycles of 95 ℃ for 5 s, 62 ℃ for 30 s, and a final extension at 72 ℃ for 30 s. Dissociation stage was performed at 95℃ for 15 s, 62℃ for 1 min, followed by a slow ramp to 95℃. Quantitative PCR and dissociation curve analyses were performed on three samples per stage using a standard curve method, as previously described [38,39].
Double-stranded RNA (dsRNA) targeting the SmMCO2 genes was synthesized by in vitro transcription of PCR amplicons containing the T7 promoter using RiboMAX Large Scale RNA Production Systems (Promega, WI, USA). The PCR-generated DNA template was amplified using KOD-Plus-Neo (Toyobo) and the specific primer sets listed in Table S2. The PCR temperature profile was 95℃ for 2 min, followed by 5 cycles of 98 ℃ for 10 s, 58 ℃ for 30 s, and a final extension at 68 ℃ for 20 s. This was followed by 30 extended cycles of 98 ℃ for 10 s, 62 ℃ for 30 s, and a final extension at 68 ℃ for 30 s were performed. dsRNA targeting the enhanced green fluorescent protein (EGFP) gene was synthesized as described above and used as a control. 0.5 µg of dsRNA for each gene was injected into last instar larvae (Figure S4) using a capillary needle made from a glass capillary (Drummond Scientific Company, 1-5 µl, 90 mm long) with a needle puller PN-31 (Narishige, Tokyo, Japan). Eight days after dsRNA injection, total RNA was extracted from each individual using the Maxwell RSC instrument and Maxwell RSC simplyRNA Tissue kit (Promega). Quantitative PCR and dissociation curve analysis for the SmMCO2 genes were performed on 15 samples each from the RNAi treatment and control groups, as described above. We then observed the effect of RNAi on the body color of weevils at 12 and 24 h after adult eclosion.
2.7. Nucleotide Sequence Accession Numbers
The sequences reported here have been submitted to the DDBJ/EMBL/GenBank database under the accession numbers LC778253–LC778268 for COI, and LC789100 and LC789101 for SmMCO2A and SmMCO2B, respectively.
2.8. Statistics
Behavioral activities and gall formation numbers were compared between the ovipositor-sealed and control groups using the Wilcoxon rank sum test and Fisher’s exact test, respectively. Fisher’s exact test with Bonferroni correction was used to compare survival between the two insecticides and the controls. The pairwise Wilcoxon rank sum test with Bonferroni correction was used to assess the effect of insecticides on the width of the developed gall. One-way Analysis of Variance (ANOVA) was used to evaluate the differences in SmMCO2 gene expression levels between growth stages. Tukey’s test was used for post hoc multiple comparisons. Welch's t-test was used to evaluate the differences in SmMCO2 gene expression levels between the RNAi-treated and control groups. All statistical analyses were performed using R software [22] v.4.2.1.
3. Results and Discussion
3.1. Gall is induced by adult female, but not by larva
Our previous study showed that galls were induced by weevil oviposition after boring with its long rostrum [17]. When we dissected the initially developed galls, eggs—not larvae—were always presented inside (20/20 galls)(Figure 1B). This finding indicates that gall induction occurs before the larvae hatch. This suggests that gall induction is caused by adult saliva or eggs and the substances delivered with them.
3.2. Gall Formation is Initiated with Substances Delivered during Oviposition
Eggs of herbivorous insects have been reported to induce plant responses [40,41,42]. Eggs and oviposition fluids are commonly involved in gall formation in hymenopteran groups [43,44,45,46], and some weevil species [47,48]. To test the possibility that galls were induced by substances delivered with the eggs of S. madaranus, ovipositor sealing experiments were performed (Figure S1A). Ovipositor sealing resulted in a slight decrease in movement compared to that in the control group, but was not significantly different in all three experiments (Figure S1C). Egg-laying holes and galls were formed in the control group (Table 1). The ovipositor-sealed group exhibited fewer egg-laying holes. This may have been due to a decrease in locomotor activity (Figure S1C). Although egg-laying holes were observed, no galls formed in the ovipositor-sealed group (Table 1). These results suggest that gall induction is strongly related to eggs and substances delivered during oviposition and not to saliva released during egg-laying hole formation.
3.3. Gall Enlargement is Influenced by Larva
Our previous study showed that gall growth stopped, and gall size decreased after S. madaranus larvae escaped [17]. This suggests that gall growth is strongly influenced by the presence of weevil larvae inside the galls. To test this possibility, we eliminated larvae from the galls using insecticides and examined their influence on gall growth.
First, insecticidal efficacy was assessed five days after treatment. In the control group, surviving larvae were found in almost all the galls (Figure 2A). In contrast, only one surviving larva was found for each of the two insecticide treatments. Survival was significantly different between the control and Dantotsu treatments and between the control and Ortolan treatments. On the other hand, no significant differences were observed between the two insecticide groups (Figure 2A). These results indicated that these insecticides were sufficiently effective in killing S. madaranus from galls.
Next, we analyzed the effect of killing larvae on gall enlargement. The control galls increased in size over time. However, when the weevils were killed with either insecticide, the galls stopped enlarging (Figure 2B). These results indicate that the presence of larvae in the galls is responsible for their enlargement. Smicronyx madaranus larvae feed vigorously inside the galls, and their inner cells show active cell division, presumably in response to larval ingestion [17]. Therefore, saliva-derived substances produced by S. madaranus larvae may be involved in the enlargement of galls.
In previously reported weevil species, both the initiation and enlargement of galls were driven by either adult oviposition (e.g., Ceutorhynchus napi on Brassica napus; Rhinusa pilosa on Linaria vulgaris) [41,48] or larval feeding (e.g., Acythopeus burkhartorum on Coccinia grandis; Conotrachelus albocinereus on Parthenium hysterophorus) [49,50]. In S. madaranus, the two gall-formation processes are differentially controlled by adult oviposition and larval feeding. This suggests that gall-formation mechanisms are diverse, even within weevil species.
3.4. Phylogenetic Analysis of the Smicronyx Madaranus and its Related Species
Phylogenetic analysis of the mitochondrial COI genes was performed for 23 species of the genus Smicronyx, including S. madaranus. Smicronyx madaranus belonged to a different clade from S. dentirostris, S. japonicus, and S. rubricatus, which coexist in Japan (Figure 3). Smicronyx smreczynskii Solari, 1952 was found in the same clade as S. madaranus. S. madaranus and S. smreczynskii are similar in appearance and nature; they form spherical galls on Cuscuta plants with increased photosynthetic activity [17,51]. This result suggests that S. madaranus and S. smreczynskii are synonymous for the same species. However, a detailed analysis is required to test this possibility.
The results also showed that almost all gall-forming species (S. coecus, S. jungermanniae, S. madaranus, and S. smreczynskii) belonged to the same clade (Figure 3). This suggests that the gall-forming trait evolved in a specific lineage within Smicronyx. To date, more than 130 species have been described in the genus Smicronyx [15,25,30,33]; however, the species used in this study did not cover the entire genus. In the future, more detailed phylogenetic analyses using high-resolution genetic markers for a larger number of species are required to provide a complete picture of the evolution of gall-forming traits in this genus.
3.5. RNAi is Effective for Gene Silencing in S. madaranus
In recent years, RNA-seq and genome analyses of gall-forming insects have been performed, and several candidate genes for gall formation have been identified [9,11,52]. However, functional analysis of the genes is necessary to prove causality, that they are responsible for the gall formation. To assess the efficacy of RNAi in S. madaranus, we knocked down the multicopper oxidase 2 (SmMCO2) gene, which is involved in cuticle melanization [53]. This gene was chosen because its knockdown results in a visible change in body color, making it easy to evaluate the efficacy of RNAi.
SmMCO2 expression was low at P2 (early pupal stage) and P3 (mid-pupal stage) (Figure 4A and Figure S4). The expression level of SmMCO2 increased rapidly from P4 (late pupal stage), when the black spotted pattern appeared on the wings. The expression then continued to increase until A2 (mid-adult stage), when the head capsule and thorax were melanized. After the A2 stage, SmMCO2 expression tended to decrease as the whole body became black. We injected dsRNA of SmMCO2 into the last instar larvae, and observed the effect on the gene expression and the body color blacking. dsRNA treatment effectively decreased SmMCO2 expression (Figure 4B). SmMCO2-RNAi individuals did not show any black spot patterns on their wings at the A1 stage (Figure 4C). The body color of the SmMCO2-RNAi group remained light brown, even after the whole body of the control group darkened. These results indicate that RNAi is effective for functional gene analysis in S. madaranus. In this study, SmMCO2 was identified as a gene involved in the darkening of body color and the formation of a characteristic spotted pattern on the elytra of S. madaranus as described in the coloration of other insects [53].
4. Conclusion and Perspectives
Gall formation is an extended phenotype in which insects reprogram plant tissue development and switch to suitable shelter and food for themselves [4,5]. Therefore, it is attractive in various fields of study, ranging from fundamental biology to applied biology. However, because many galls form on woody plants [12,13], it is difficult to conduct experimental studies in a laboratory. Therefore, insect models are required to better understand the mechanisms of gall formation. Smicronyx madaranus is an excellent model that can be maintained throughout the year and subjected to various manipulations in the laboratory, which will accelerate research on the mechanisms of gall formation.
Our manipulation experiments revealed that there is an age-dependent role change in gall formation in S. madaranus; adult females and larvae are responsible for gall induction and enlargement (Table 1, Figure 1 and Figure 2). Substances released during oviposition and larval feeding are expected to be involved in each process. Phytohormones and protein elicitors released by oviposition and feeding have been suggested to play important roles in gall formation [6,7,8,10,11,41,44,48,54,55]. In the future, it will be necessary to identify candidate genes using RNA-seq analysis at each developmental stage of S. madaranus to understand the insect-derived factors involved in gall formation.
Our phylogenetic analysis showed that almost all gall-forming weevils, including S. madaranus, belong to a single lineage within the genus Smicronyx (Figure 3), suggesting that the gall-forming trait evolved in a specific lineage within Smicronyx. Gall-forming weevils are thought to have evolved from seed or fruit feeders [14]. The ability to produce galls, a fruit-like food source, is considered a highly adaptive evolution, as host plants produce flowers only at limited times of the season. Recent molecular biology studies have shown that the expression of flower and fruit development genes is significantly upregulated in the developing galls of many plant species [18,20,56]. Similar mechanisms may be involved in the development of galls in C. campestris. Further experimental studies using RNA-seq are required. Comparative genomic analyses between closely related gall-forming and non-gall-forming species will be useful for elucidating the molecular evolution of Smicronyx weevils that enable gall formation.
In this study, we showed that RNAi was effective in adult S. madaranus through an experiment targeting the MCO2 gene. It is expected that the mechanisms of gall induction in adult females can be elucidated through a comprehensive functional analysis of candidate genes using RNAi. However, the injection of dsRNA is difficult for larvae living in the gall. Therefore, other techniques are needed for the functional analysis of the genes expressed in larvae. Maternal RNAi [57], feeding-RNAi via the host plant [58], and genome editing [59] will be useful for this purpose.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplemental Movie Legend. Movie S1: Observation of gall formation using time-lapse photography. Movie S2: Digital video tracking of S. madaranus locomotor activity. Figure S1: Effects of sealed ovipositors on behavioral activity. Figure S2: Alignment of putative amino acid sequences of SmMCO2A and SmMCO2B. Figure S3: Molecular phylogenetic analysis of SmMCO2 and related genes based on deduced amino acid sequences. Figure S4: Growth stage and body color changes in S. madaranus.
Author Contributions
Conceptualization, Tsutomu Tsuchida; Formal analysis, Ryo Ushima, Ryoma Sugimoto, Yota Sano, Hinako Ogi, Ryuichiro Ino, Keisuke Shimada and Tsutomu Tsuchida; Funding acquisition, Tsutomu Tsuchida; Investigation, Ryo Ushima, Ryoma Sugimoto, Yota Sano, Hinako Ogi, Ryuichiro Ino and Tsutomu Tsuchida; Methodology, Keisuke Shimada and Tsutomu Tsuchida; Project administration, Tsutomu Tsuchida; Resources, Hiroshi Hayakawa; Supervision, Tsutomu Tsuchida; Visualization, Ryo Ushima and Tsutomu Tsuchida; Writing – original draft, Ryo Ushima, Ryoma Sugimoto, Yota Sano, Hinako Ogi, Ryuichiro Ino , Keisuke Shimada and Tsutomu Tsuchida; Writing – review & editing, Tsutomu Tsuchida.
Funding
This study was supported by JSPS KAKENHI (Grant Number JP21H02203).
Data Availability Statement
This article has no additional data.
Acknowledgments
We thank K. Bessho-Uehara and T. Wakasugi for useful comments, A. Yokomichi and K. Kawabe for technical assistance, H. Kawase, T. Wakasugi, R. Murakami and Y. Tsutsumiuchi, for sampling of Smicronyx.
Conflicts of Interest
The authors declare no competing interests, financial or otherwise.
References
- Redfern, M. Redfern, M. Plant galls; Collins, 2011.
- Shorthouse, J.D.; Rohfritsch, O. Biology of Insect-induced galls; Oxford University Press: New York, 1992. [Google Scholar]
- Price, P.; Fernandes, G.; Waring, G. Adaptive nature of insect galls. Environ. Entomol. 1987, 16, 15–24. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K. The adaptive significance of insect gall morphology. TRENDS in Ecology and Evolution 2003, 18, 512–522. [Google Scholar] [CrossRef]
- Dawkins, R. The extended phenotype; Oxford University Press: Oxford, UK, 1982. [Google Scholar]
- Tooker, J.F.; Helms, A.M. Phytohormone dynamics associated with gall insects, and their potential role in the evolution of the gall-inducing habit. J. Chem. Ecol. 2014, 40, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Yokokura, J.; Ito, T.; Arai, R.; Yokoyama, C.; Toshima, H.; Nagata, S.; Asami, T.; Suzuki, Y. Biosynthetic pathway of the phytohormone auxin in insects and screening of its inhibitors. Insect Biochem. Mol. Biol. 2014, 53, 66–72. [Google Scholar] [CrossRef]
- Giron, D.; Huguet, E.; Stone, G.N.; Body, M. Insect-induced effects on plants and possible effectors used by galling and leaf-mining insects to manipulate their host-plant. J. Insect Physiol. 2016, 84, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Escalante, L.N.; Chen, H.; Benatti, T.R.; Qu, J.; Chellapilla, S.; Waterhouse, R.M.; Wheeler, D.; Andersson, M.N.; Bao, R.; et al. A massive expansion of effector genes underlies gall-formation in the wheat pest Mayetiola destructor. Curr. Biol. 2015, 25, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Cambier, S.; Ginis, O.; Moreau, S.J.M.; Gayral, P.; Hearn, J.; Stone, G.N.; Giron, D.; Huguet, E.; Drezen, J.M. Gall wasp transcriptomes unravel potential effectors involved in molecular dialogues with oak and rose. Front. Physiol. 2019, 10, 926. [Google Scholar] [CrossRef]
- Korgaonkar, A.; Han, C.; Lemire, A.L.; Siwanowicz, I.; Bennouna, D.; Kopec, R.E.; Andolfatto, P.; Shigenobu, S.; Stern, D.L. A novel family of secreted insect proteins linked to plant gall development. Curr. Biol. 2021, 31, 1836–1849. [Google Scholar] [CrossRef]
- Gonçalves-Alvim, S.J.; Fernandes, G.W. Biodiversity of galling insects: historical, community and habitat effects in four neotropical savannas. Biodivers. Conserv. 2001, 10, 79–98. [Google Scholar] [CrossRef]
- Veldtman, R.; McGeoch, M. Gall-forming insect species richness along a non-scleromorphic vegetation rainfall gradient in South Africa: The importance of plant community composition. Austral. Ecol. 2003, 28, 1–13. [Google Scholar] [CrossRef]
- Korotyaev, B.; Konstantinov, A.; S.W, L.; Mandelshtam, M.; Volkovitsh, M. Gall-inducing Coleoptera, 2005; pp. 239–271.
- Aistova, E.V.; Bezborodov, V.G. Weevils belonging to the Genus Smicronyx Schönherr, 1843 (Coleoptera, Curculionidae) affecting dodders (Cuscuta Linnaeus, 1753) in the Russian Far East. Russ. J. Biol. Invasions 2017, 8, 184–188. [Google Scholar] [CrossRef]
- Morimoto, K.; Kojima, H. Weevils of the genus Smicronyx in Japan (Coleoptera: Curculionidae). Entomol. Rev. Jpn. 2007, 62, 1–9. [Google Scholar]
- Murakami, R.; Ushima, R.; Sugimoto, R.; Tamaoki, D.; Karahara, I.; Hanba, Y.; Wakasugi, T.; Tsuchida, T. A new galling insect model enhances photosynthetic activity in an obligate holoparasitic plant. Sci. Rep. 2021, 11, 13013. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Kimura, S.; Sakamoto, T.; Okamoto, A.; Nakayama, T.; Matsuura, T.; Ikeda, Y.; Takeda, S.; Suzuki, Y.; Ohshima, I.; et al. Reprogramming of the developmental program of Rhus javanica during initial stage of gall induction by Schlechtendalia chinensis. Front. Plant Sci. 2020, 11, 471. [Google Scholar] [CrossRef] [PubMed]
- Nabity, P.D.; Haus, M.J.; Berenbaum, M.R.; DeLucia, E.H. Leaf-galling phylloxera on grapes reprograms host metabolism and morphology. Proc. Natl. Acad. Sci. USA 2013, 110, 16663–16668. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Yoza, M.; Amano, T.; Ohshima, I.; Hirano, T.; Sato, M.H.; Sakamoto, T.; Kimura, S. Comparative transcriptome analysis of galls from four different host plants suggests the molecular mechanism of gall development. PLoS One 2019, 14, e0223686. [Google Scholar] [CrossRef] [PubMed]
- Fukatsu, T. Acetone preservation: a practical technique for molecular analysis. Mol. Ecol. 1999, 8, 1935–1945. [Google Scholar] [CrossRef]
- Ihaka, R.; Gentleman, R. R: A Language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Dieckmann, L. Beiträge zur Insektenfauna der DDR: Coleoptera - Curculionidae (Erirhinae). Beiträge zur Entomologie = Contributions to Entomology 1986, 36, 119–181. [Google Scholar]
- Haran, J. A review of the genus Smicronyx Schoenherr (Coleoptera, Curculionidae, Curculioninae) in tropical Africa. Zootaxa 2018, 4508, 267–287. [Google Scholar] [CrossRef] [PubMed]
- Dinelli, G.; Bonetti, A.; Tibiletti, E. Photosynthetic and accessory pigments in Cuscuta-Campestris Yuncker and some host species. Weed Res. 1993, 33, 253–260. [Google Scholar] [CrossRef]
- Haran, J. A new record of Smicronyx smreczynskii (Solari, 1952) on Cuscuta scandens in the South-West of France. SNUDEBILLER: Studies on taxonomy, biology and ecology of Curculionoidea 2014, 15, 4. [Google Scholar]
- Heijerman, T.; Alders, K. Smicronyx smreczynskii, een nieuwe snuitkever voor de nederlandse fauna (Coleoptera: Curculionidae). Nederlandse faunistische Mededelingen 2000, 12, 11–13. [Google Scholar]
- Morimoto, K.; Matoba, I. Addition to the genus Smicronyx of Japan (Coleoptera: Curculionidae). Entomol. Rev. Jpn 2009, 64, 67–72. [Google Scholar]
- Haran, J.; SchÜTte, A.; Friedman, A.-L.-L. A review of Smicronyx Schoenherr (Coleoptera, Curculionidae) of Israel, with description of two new species. Zootaxa 2017, 4237, 17–40. [Google Scholar] [CrossRef] [PubMed]
- Benedikt, S.; Krátk´y, J.i.; Schön, K. Nové a potvrzené druhy nosatců (Coleoptera: Curculionoidea) pro Českou republiku a Slovensko. Západočeské entomologické listy 2016 (in Czech), 7, 25-31.
- Mifsud, D.; Colonnelli, E. The Curculionoidea of the Maltese Islands (Central Mediterranean) (Coleoptera). Bulletin of the Entomological Society of Malta 2010, 3, 55–143. [Google Scholar]
- Haran, J. The Smicronychini of southern Africa (Coleoptera, Curculionidae): Review of the tribe and description of 12 new species. Eur. J. Taxon. 2021, 735, 34–73. [Google Scholar] [CrossRef]
- Dhileepan, K.; Madigan, B.; Vitelli, M.; McFadyen, R.; Webster, K.; Trevino, M. A New Initiative in the Biological Control of Parthenium. Proc. 11th Aust. Weeds Conf. 1996, 309–312. [Google Scholar]
- Anderson, D.M. The weevil genus Smicronyx in america north of Mexico (Coleoptera: Cureulionidae). One hundred and fifty-seven figures and one plate. Proceedings of the United States National Museum 1962, 113, 185–372. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Koga, R.; Fujiwara, A.; Fukatsu, T. Phenotypic effect of “Candidatus Rickettsiella viridis,” a facultative symbiont of the pea aphid (Acyrthosiphon pisum), and its interaction with a coexisting symbiont. Appl. Environ. Microbiol. 2014, 80, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, A.; Meng, X.-Y.; Kamagata, Y.; Tsuchida, T. Subcellular niche segregation of co-obligate symbionts in whiteflies. Microbiol. Spectr. 2023, 11, e04684–22. [Google Scholar] [CrossRef] [PubMed]
- Reymond, P. Perception, signaling and molecular basis of oviposition-mediated plant responses. Planta 2013, 238, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Hilker, M.; Rohfritsch, O.; Meiners, T. The plant's response towards insect egg deposition. In Chemoecology of insect eggs and egg deposition Hilker, M., Meiners, T., Eds.; Blackwell publishing, 2008; pp. 205–233.
- Hilker, M.; Fatouros, N.E. Plant responses to insect egg deposition. Annu. Rev. Entomol. 2015, 60, 493–515. [Google Scholar] [CrossRef] [PubMed]
- Dreger-Jauffret, F.; Shorthouse, J.D. Diversity of gall-inducing insects and their galls. In Biology of insect-induced galls Shorthouse, J.D., Rohfritsch, O., Eds.; Oxford University Press: New York, 1992; pp. 8–33. [Google Scholar]
- Oates, C.N.; Denby, K.J.; Myburg, A.A.; Slippers, B.; Naidoo, S. Insect egg-induced physiological changes and transcriptional reprogramming leading to gall formation. Plant Cell Environ. 2021, 44, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Rey, L.A. Developmental morphology of two types of Hymenopterous galls. In Biology of insect-induced galls Shorthouse, J.D., Rohfritsch, O., Eds.; Oxford University Press: New York, 1992; pp. 87–101. [Google Scholar]
- Leggo, J.J.; Shorthouse, J.D. Development of stem galls induced by Diplolepis triforma (Hymenoptera: Cynipidae) on Rosa acicularis (Rosaceae). Can. Entomol. 2006, 138, 661–680. [Google Scholar] [CrossRef]
- Rohfritsch, O. Patterns in gall development. In Biology of insect-induced galls Shorthouse, J.D., Rohfritsch, O., Eds.; Oxford University Press: New York, 1992; pp. 60–86. [Google Scholar]
- Barnewall, E.C.; De Clerck-Floate, R.A. A preliminary histological investigation of gall induction in an unconventional galling system. Arthropod Plant Interact. 2012, 6, 449–459. [Google Scholar] [CrossRef]
- Raman, A.; Cruz, Z.T.; Muniappan, R.; Reddy, G.V.P. Biology and host specificity of gall-inducing Acythopeus burkhartorum (Coleoptera: Curculionidae), a biological-control agent for the invasive weed Coccinia grandis (Cucurbitaceae) in Guam and Saipan. Tijdschr. Entomol. 2007, 150, 181–191. [Google Scholar] [CrossRef]
- Florentine, S.; Raman, A.; Dhileepan, K. Response of the weed Parthenium hysterophorus (Asteraceae) to the stem gall-inducing weevil Conotrachelus albocinereus (Coleoptera: Curculionidae). Entomol. Gen. 2002, 26, 195–206. [Google Scholar] [CrossRef]
- Anikin, V.V.; Nikelshparg, M.I.; Nikelshparg, E.I.; Konyukhov, I.V. Photosynthetic activity of the dodder Cuscuta campestris (Convolvulaceae) in case of plant inhabitation by the gallformed weevil Smicronyx smreczynskii (Coleoptera, Curculionidae). Chemistry. Biology. Ecology 2017, 17, 42–47. [Google Scholar]
- Gobbo, E.; Lartillot, N.; Hearn, J.; Stone, G.N.; Abe, Y.; Wheat, C.W.; Ide, T.; Ronquist, F. From inquilines to gall inducers: Genomic signature of a life-style transition in Synergus gall wasps. Genome Biol. Evol. 2020, 12, 2060–2073. [Google Scholar] [CrossRef] [PubMed]
- Futahashi, R.; Koshikawa, S.; Okude, G.; Osanai-Futahashi, M. Diversity of melanin synthesis genes in insects. Adv. In Insect Phys. 2022, 62, 339–376. [Google Scholar]
- Bartlett, L.; Connor, E.F. Exogenous phytohormones and the induction of plant galls by insects. Arthropod Plant Interact. 2014, 8, 339–348. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Tanaka, H.; Hasegawa, M.; Tokuda, M.; Asami, T.; Suzuki, Y. Phytohormones and willow gall induction by a gall-inducing sawfly. New Phytol. 2012, 196, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.C.; Edger, P.P.; Body, M.J.A.; Appel, H.M. A galling insect activates plant reproductive programs during gall development. Sci. Rep. 2019, 9, 1833. [Google Scholar] [CrossRef]
- Nishide, Y.; Kageyama, D.; Hatakeyama, M.; Yokoi, K.; Jouraku, A.; Tanaka, H.; Koga, R.; Futahashi, R.; Fukatsu, T. Diversity and function of multicopper oxidase genes in the stinkbug Plautia stali. Sci. Rep. 2020, 10, 3464. [Google Scholar] [CrossRef]
- Pitino, M.; Coleman, A.D.; Maffei, M.E.; Ridout, C.J.; Hogenhout, S.A. Silencing of aphid genes by dsRNA feeding from plants. PLoS One 2011, 6, e25709. [Google Scholar] [CrossRef]
- Shirai, Y.; Piulachs, M.-D.; Belles, X.; Daimon, T. DIPA-CRISPR is a simple and accessible method for insect gene editing. Cell Rep. Methods 2022, 2, 100215. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
An initial gall (A) and an egg in the initial gall (B).

Figure 2.
Effects of insecticides on weevil larvae and gall growth. (A) Larval status within treated galls. White background, alive; dark grey, undetected; black, dead. Numbers in each column indicate number of individuals in each state. Different letters (a and b) above columns indicate statistically significant differences (P<0.05, Fisher’s exact test with Bonferroni correction). (B) Effects of insecticides on gall growth. Means ± standard deviations are shown. Sample size is given in parentheses. Different letters (a and b) indicate statistically significant differences on the same day (P<0.05, pairwise Wilcoxon rank sum test with Bonferroni correction). ns means no significant difference.
Figure 2.
Effects of insecticides on weevil larvae and gall growth. (A) Larval status within treated galls. White background, alive; dark grey, undetected; black, dead. Numbers in each column indicate number of individuals in each state. Different letters (a and b) above columns indicate statistically significant differences (P<0.05, Fisher’s exact test with Bonferroni correction). (B) Effects of insecticides on gall growth. Means ± standard deviations are shown. Sample size is given in parentheses. Different letters (a and b) indicate statistically significant differences on the same day (P<0.05, pairwise Wilcoxon rank sum test with Bonferroni correction). ns means no significant difference.
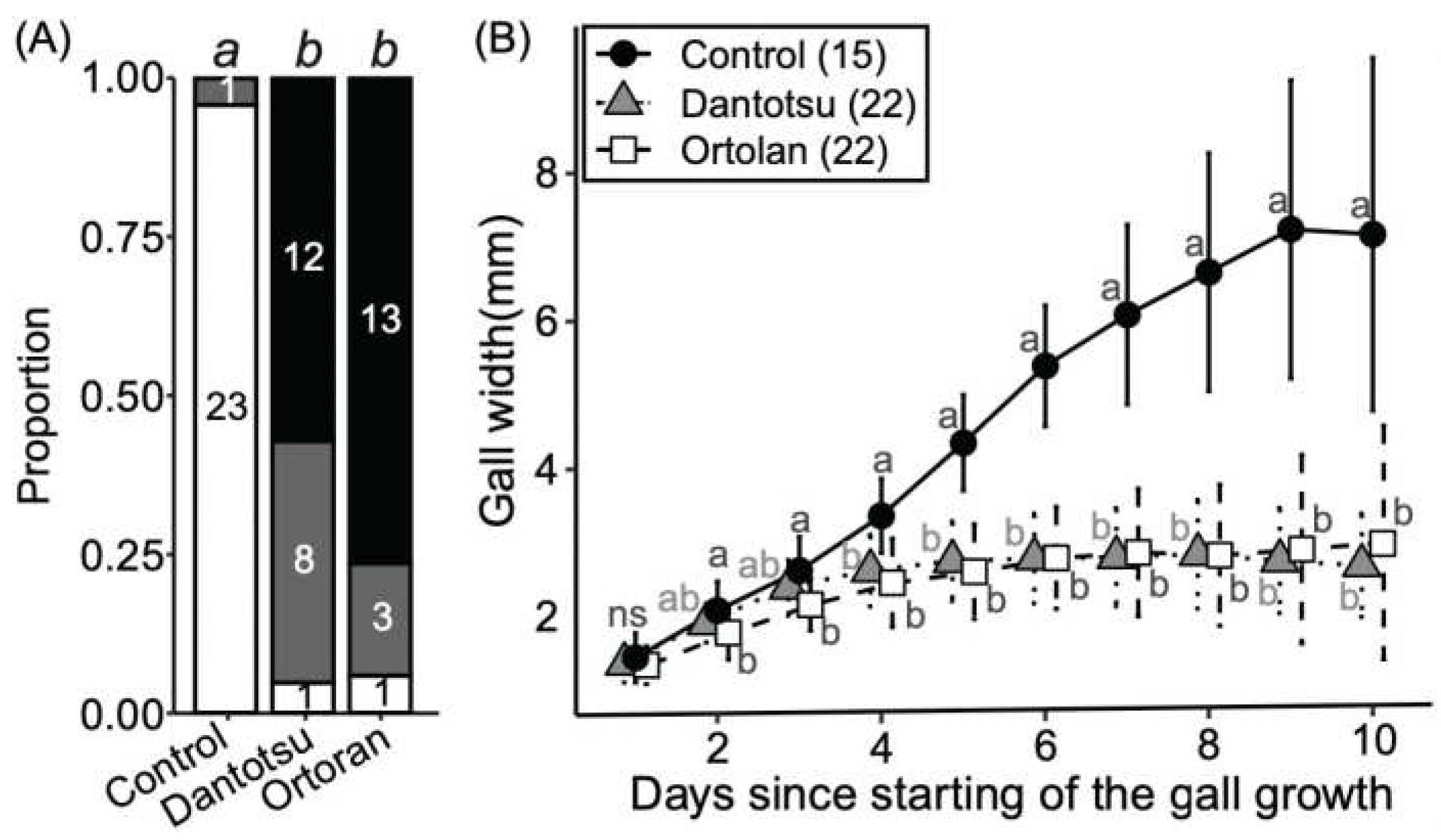
Figure 3.
Molecular phylogeny and gall-forming trait in the genus Smicronyx. The topology and branch lengths shown were obtained by Bayesian inference methods. Scale bar indicates 0.5 substitutions per site. The Bayesian posterior probabilities/the maximum likelihood bootstrap value/the maximum parsimony bootstrap value are shown below the branches to indicate the level of support for each node (only values ≥ 50% are shown), respectively. Sequences obtained in this study are in bold type. Labels of the sequences correspond to the codes in Table S1. Filled circles in the taxa represent the gall-forming species. Sequence accession numbers are in brackets.
Figure 3.
Molecular phylogeny and gall-forming trait in the genus Smicronyx. The topology and branch lengths shown were obtained by Bayesian inference methods. Scale bar indicates 0.5 substitutions per site. The Bayesian posterior probabilities/the maximum likelihood bootstrap value/the maximum parsimony bootstrap value are shown below the branches to indicate the level of support for each node (only values ≥ 50% are shown), respectively. Sequences obtained in this study are in bold type. Labels of the sequences correspond to the codes in Table S1. Filled circles in the taxa represent the gall-forming species. Sequence accession numbers are in brackets.
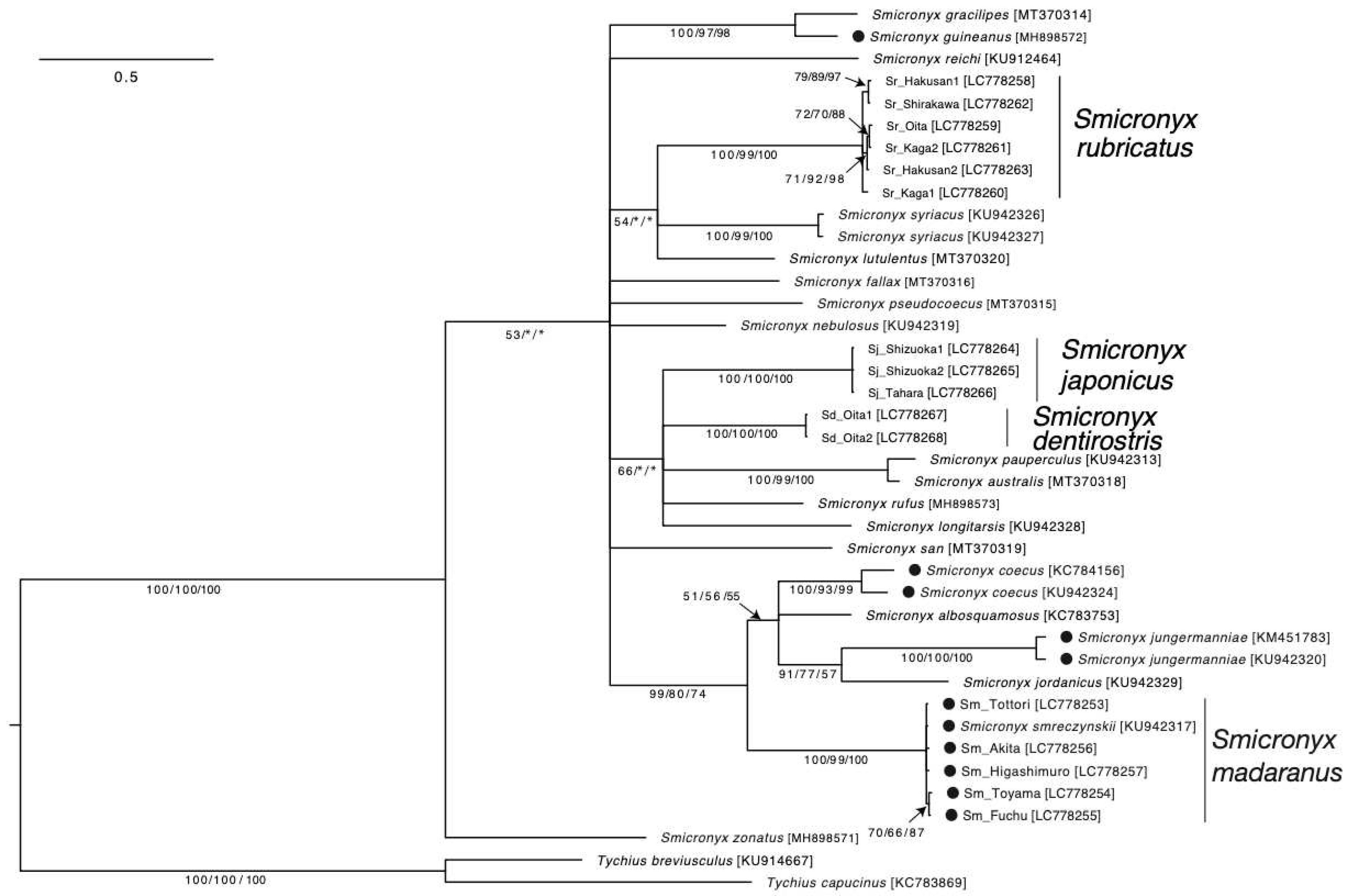
Figure 4.
SmMCO2 expression and body color melanization. (A) Variation of SmMCO2 expression at different growth stages. Box plots show the distribution of the normalized SmMCO2 expression level. Different letters (a and b) indicate statistically significant differences on the same day (P<0.05, pairwise Wilcoxon rank sum test with Bonferroni correction). Growth stages (P2 to A3) correspond to Figure S4. (B) Effect of RNAi on the normalized SmMCO2 expression level. The numbers in parenthesis are sample sizes. The median expression level of the control group was designated as 1.0. Statistically significant differences were evaluated by Welch's t-test. In (A) and (B), y axes indicate SmMCO2 gene copies/EF1α gene copy. Each open circle indicates the value per sample. (C) Effect of RNAi for SmMCO2 on body color. Bars, 1 mm.
Figure 4.
SmMCO2 expression and body color melanization. (A) Variation of SmMCO2 expression at different growth stages. Box plots show the distribution of the normalized SmMCO2 expression level. Different letters (a and b) indicate statistically significant differences on the same day (P<0.05, pairwise Wilcoxon rank sum test with Bonferroni correction). Growth stages (P2 to A3) correspond to Figure S4. (B) Effect of RNAi on the normalized SmMCO2 expression level. The numbers in parenthesis are sample sizes. The median expression level of the control group was designated as 1.0. Statistically significant differences were evaluated by Welch's t-test. In (A) and (B), y axes indicate SmMCO2 gene copies/EF1α gene copy. Each open circle indicates the value per sample. (C) Effect of RNAi for SmMCO2 on body color. Bars, 1 mm.
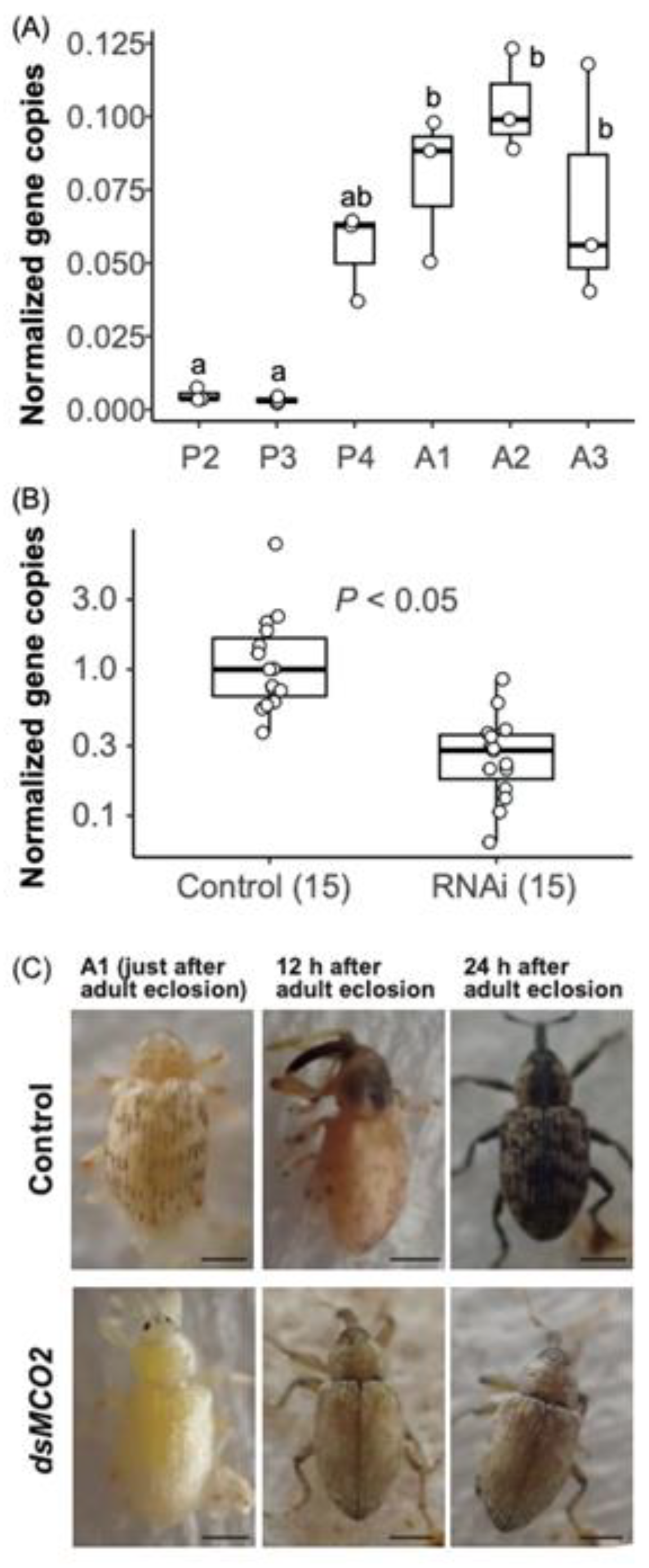

Table 1.
Effects of ovipositor sealing on gall formation.
| Treatment | na | Egg-laying holeb | Gallc |
|---|---|---|---|
| Control | 12 | 12.42 ± 5.14 | 8.67 ± 3.55 |
| Sealed | 12 | 4.17 ± 2.25 *** | 0 *** |
a Number of individuals examined. b Number of egg-laying hole formed in the nodes per an individual. c Number of galls formed by an individual. Statistical significance was calculated using Fisher's exact test (***, P <0.001).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



