
Submitted:
04 December 2023
Posted:
05 December 2023
You are already at the latest version
Abstract
This research is aimed to investigate the effects of Lactoplantibacillus plantarum (L. plantarum) on growth performance, oxidation resistance, immunity, as well as cecal microbiota in broilers. This work classed three hundred and sixty 1-day-old broilers as three groups randomly, including control (CON, basal diet), antibiotic (ANT, 75 mg kg-1 chlortetracycline added into basal diet), or probiotic group (LP, 5×108 CFU kg-1 Lactoplantibacillus plantarum HJLP-1 contained within basal diet). Animal were then fed for 42 days, and each group consisted of 15 broilers for 8 replicates. Compared with CON, L. plantarum supplementation significantly improved the average weight daily gain (AWDG) (P < 0.05) while reducing feed-to-gain ratio over the entire supplemental period (P < 0.05). Birds fed L. plantarum had marked lower serum ammonia and xanthine oxidase levels (P < 0.05) than those in ANT and CON. Significant improvements (P < 0.05) of superoxide dismutase, catalase, serum IgM and IgY contents in broilers fed L. plantarum were also observed when compared those in CON and ANT groups. Both L. plantarum and antibiotics decreased the pro-inflammatory factor IL-1β level significantly (P < 0.05), while only L. plantarum promoted the anti-inflammatory factor IL-10 level dramatically in the serum (P < 0.05) compared with CON. The L. plantarum dramatically (P < 0.05) increased acetic acid and butyric acid concentrations in the cecal contents,when compared to those in CON and ANT. Among the differences revealed by 16S rRNA analysis, L. plantarum markedly improved the community richness of cecal microbiota. At the genus level, butyric acid-producing bacteria Ruminococcus and Lachnospiraceae were found in higher relative abundance in samples of L. plantarum treated birds. In conclusion, dietary L. plantarum supplementation promoted the growth and health of broilers, likely by inducing a shift in broiler gut microbiota toward short-chain fatty acid (SCFA)-producing bacteria. Therefore, L. plantarum has tremendous potential as an alternative for antibiotics in poultry breeding.
Keywords:
Lactoplantibacillus plantarum
; broiler
; growth performance
; immune function
; cecal microbiota
1. Introduction
Broiler chickens are one of the main sources of animal protein for human consumption. To maximize the production, intensive indoor systems are employed by the commercial poultry producers [1]. However, the intensively reared broiler chickens encounter the simultaneous action of multiple stressors from the housing environment, which causes high mortality of broilers from common infectious or metabolic diseases [2]. Conventional poultry farms use antibiotics for treatment, prophylaxis, and growth promotion, which give the rise of antibiotic resistance pathogens. The emergence and spread of antibiotic resistance affect animal and human health and become a global concern [3,4]. Therefore, many countries have made efforts to limit antibiotics application into feeds. In 2020, antibiotics added to feed as growth promoters were completely banned in China.
With the ban on antibiotics, an increasing number of probiotics are being developed as an alternative to antibiotics to be added to the rations [5,6]. Suitable probiotics can improve the intestinal flora, enhance the absorption of nutrients by livestock and poultry, strengthen immunity, and reduce the impact on farm animal health and growth induced by feeding environment [7].
Lactoplantibacillus plantarum is a homofermentative, aerotolerant, Gram-positive bacterium that produces both isomers of lactic acid as the main end-product of carbohydrate fermentation [8]. It can be obtained in numerous sources, such as human feces human breast milk, fermented food products and cheese [9,10,11,12,13]. Evaluating probiotic properties of L. plantarum revealed its antioxidant and immune-enhancing effects [14]. L. plantarum is suggested to have an important effect on gastrointestinal disorders and diarrhea prevention [15,16]. In broiler chickens, feeding L. plantarum can improve growth performance, promote gut microbial homeostasis, prevent pathogen infection, and reduce the intestinal barrier injury caused by toxins [17,18,19]. However, as a considerable number of studies have shown that L. plantarum has positive effects on intestinal health, more evidence is needed to establish whether L. plantarum plays an efficient role in replacing antibiotics as growth promoter in broiler feeding. Therefore, the present work evaluated whether L. plantarum affected broilers in terms of the growth performance, oxidation resistance, immune response, intestinal microbiota, and short chain fatty acid profiles of broilers and compared with those of antibiotic growth promoters.
2. Material and Methods
2.1. Experimental Animals and Design
A total of 360 Avian chickens (one-day-old) were individually weighed and randomly assigned into three groups: (1) control group (CON) fed basal diet; (2) antibiotic group (ANT) fed the base diet supplemented with 75 mg/ kg of chlortetracycline; (3) probiotic group (LP) fed the basal diet plus 5 × 108 CFU/ kg of L. plantarum HJLP-1 supplied by Vegamax Biotechnology Co. Ltd. (Huzhou, China). Each treatment 15 birds of 8 replicates in each pen. Dietary formulations met or exceeded the daily growth needs of broiler chickens (NRC, 1994; Table 1). The feeding experiment lasted for 42 d. Animals had free access to water and feed. Continuous light, suitable temperature and air quality for white feather broiler breeding were maintained in the coop.
2.2. Growth Performance
The weights of broilers from each pen were taken on days 1, 21 and 42 for evaluating average weight daily gain (AWDG). Meanwhile, the remaining feed was weighed for calculating the average daily feed intake (ADFI) together with feed to gain ratio (F/G).
2.3. Sample Collection
Eight broilers from every group were randomized to collect samples on day 42 (one broiler from each replicate). Left -wing venous blood collection was completed in the 5-mL sterilized tubes and centrifugated at 3000× g and 4 °C for 10-min period for obtaining serum. After blood collection, the broilers were sacrificed. Cecal contents were aseptically removed. Each sample was preserved under -80 °C prior to subsequent analyses.
2.4. Measurements of Biochemical Parameters in Serum
Serum samples were screened for biochemical parameters about metabolites, antioxidants, immunoglobulin and cytokines using the corresponding diagnostic kits obtained from the Nanjing Angle Gene Bioengineering Co., Ltd. (Nanjing, China).
2.5. Assessment of Cecal Microflora Contents by 16S rRNA Sequencing
Cecal microbial genomic DNA was extracted using the QIAamp DNA Stool Mini Kit (Qiagen GmbH, Hilden, Germany). Then, 338F-806R region (V3-V4 hypervariable region, 468 base pairs) of the bacterial 16S rRNA was amplified and sequenced on Illumina MiSeq platform (Illumina, San Diego, USA). Finally, the composition and abundance of the caecum microflora were determined by alpha and beta diversity analysis.
2.6. Short Chain Fatty Acid Concentration Analysis
The concentration of cecal short chain fatty acid (SCFA) was estimated by headspace sampler gas chromatography (Agilent Technologies, USA) using the method of Thanh et al. [20]. The 0.5 g sample was dissolved in 1 mL water. After centrifugation at 12,500 g for 5 min, the supernatant was extracted and mixed with 25% phosphoric acid. The concentration of SCFAs was determined by an Agilent Technologies 7890A Network System equipped with a 30 m × 0.32 mm × 1.8 μm column (DB-624) and flame ionization detector.
2.7. Statistical Analysis
One-way analysis of variance (ANOVA) was performed using GraphPad Prism 8.2.1 software (GraphPad Prism Inc., USA), followed by Tukey’s method for comparing differences. Results were expressed as means with standard error of mean (SEM). Statistical significance was set at P < 0.05. Analysis of bacteria community data was conducted using the cloud majorbio platform (http://cloud.majorbio.com/).
3. Results
3.1. Growth Performance
Table 2 displays growth performance analysis. In comparison with control, both L. plantarum HJLP-1 and antibiotics treatments improved the body weight (BW) significantly (P < 0.05) during the start period (day 1 to 21). While during the finisher period (days 21 to 42), only LP group showed remarkably elevated BW (P < 0.05). Accordingly, during the start period, the birds fed L. plantarum HJLP- 1 or antibiotics had higher average weight daily gain (ADWG) than birds in CON group (P < 0.05). In the finisher period and during the whole supplementation period (days 1–42), LP group had markedly elevated ADWG compared with ANT and CON groups (P < 0.05). During the start period, supplementation with antibiotics decreased the F/G compared with the LP and CON groups (P < 0.05). Whereas in the finisher period and during the whole supplementation period, broilers fed with L. plantarum HJLP-1 had close F/G ratio to those of ANT group and obviously decreased relative to CON group (P < 0.05).
3.2. Serum Metabolite Index
Effects of L. plantarum HJLP-1 on the metabolite index in broiler serum are shown in Table 3. Although either L. plantarum HJLP-1 or antibiotic supplementation significantly decreased the serum ammonia (NH3) level (P < 0.05) relative to CON group, the serum ammonia levels of broilers fed L. plantarum HJLP-1 dramatically decreased compared with broilers fed antibiotics (P < 0.05). serum urea nitrogen (UN) and uric acid (UA) contents were not significantly different. Furthermore, birds in the LP group had lower serum xanthine oxidase (XOD) levels than birds in CON and ANT groups (p < 0.05).
3.3. Antioxidant Index in Serum
Effects of L. plantarum HJLP-1 on the antioxidant index in serum are shown in Table 4. Addition of L. plantarum HJLP-1 significantly improved the SOD and CAT levels in serum (P < 0.05). MDA and GSH-PX contents were not significantly different across diverse groups.
3.4. Immunoglobulin and Cytokines Index in Serum
As shown in Table 5, both L. plantarum HJLP-1 addition markedly elevated serum IgM and IgY contents in broilers (P < 0.05) compared with that in CON and ANT groups. Compared with CON, L. plantarum HJLP-1 supplementation reduced pro-inflammatory factor IL-1β levels significantly (P < 0.05) and promoted anti-inflammatory factor IL-10 expression dramatically (P < 0.05). Antibiotics also markedly reduced IL-1β levels (P < 0.05), but had no effect on IL-10 levels. In addition, the pro-inflammatory factor TNF-α level in the serum of broilers fed L. plantarum HJLP-1 remarkably decreased compared with broilers fed antibiotics (P < 0.05).
3.5. Microflora Structure in the Cecal Contents
As shown in Figure 1(A–E), L plantarum supplementation induced some changes in microflora structure in the cecal digesta of broilers. Venn diagram demonstrated that the unique Operational Taxonomic Units (OTUs) of the LP group were 166, much higher than those of CON (39) and ANT (24) groups (Figure 1A). According to genus relative abundance-based principal component analysis (PCA), microbiota in the LP group clearly separated from the CON and ANT groups (Figure 1B). Furthermore, L plantarum supplementation markedly improved the community richness, as evidenced by the significantly increased ACE and Chao indices, compared to other groups. (P < 0.05) (Figure 1C). Samples from ANT and LP groups had higher proportion of Firmicutes and lower ratio in Bacteroidetes than those from CON group, at the level of phylum (Figure1D). Results of linear discrimination analysis coupled with effect size (LEfSe) indicated Lactoplantibacillus plantarum supplementation mainly enriched f_Ruminococcaceae and unclassified_f_Lachnospiraceae; antibiotic supplementation enriched Alistipes and Oscillospirales, whereas Bacteroides, Bacteroidaceae and Barnesiellaceae were enriched at genus level of control broilers (Figure 1E).
3.6. Short Chain Fatty Acid Concentrations
Figure 2 displays SCFA results. Butyric acid and acetic acid within the cecal contents of broilers fed L. plantarum HJLP-1 dramatically increased (P < 0.05) compared with ANT and CON. There was no significant modification in propionic acid concentration among the three treatment groups.
3.7. Link between Cecal Bacteria Community and Short Chain Fatty Acid
Figure 3 shows the spearman correlation heatmap of the top 50 genera and short chain fatty acid. During the digestion, f_Ruminococcaceae, Alistipes and unclassified_f_Lachnospiraceae were significantly positively corelated with butyric acid production, whereas, Bacteroides had a significant negative correlation.
4. Discussion
Probiotics are widely reported to promote broiler health and development [21,22,23]. Lactoplantibacillus plantarum has also been shown to promote growth in broilers [24,25,26]. Consistent with these previous reports, in this study, supplementation with L. plantarum HJLP-1 increased the BW of broilers without influencing feed intake (FI), resulting in reduced F/G ratio (improved feed conversion) over the entire supplemental period to a level equivalent to or lower than that under antibiotics. Therefore, the increased ADG might be attributed to the improvement in nutrient utilization. These beneficial effects on growth performance indicate that L. plantarum HJLP-1 could be an effective substitute for antibiotics in poultry diets.
Serum uric acid (UA), urea nitrogen (UN), and ammonia levels have been considered as indicators to evaluate the utilization of amino acid in broilers [27]. Elevated serum indexes can cause disruptions in amino acid and nucleotide metabolism, ultimately affecting antioxidant capacity [28] and immune function of the body [29]. In this study, L. plantarum HJLP-1 supplementation reduced serum UA, UN, ammonia (P < 0.05), and XOD (P < 0.05), compared with CON and ANT. Hence, L. plantarum HJLP-1 may have a positive effect on amino acid utilization in broiler diets. XOD represents a critical enzyme responsible for UA production; The enhanced XOD activity can assist in generating UA with producing reactive oxygen species (ROS) [30].
Malondialdehyde, T-SOD, CAT, and GSH-Px represent major factors adopted for assessing oxidative status of the enzymatic system. As the lipid peroxidation production MDA can damage physical cell activity and alter biological membrane function, which has been frequently adopted to be the biomarker for oxidative injury. In contrast, SOD protects cells from free radicals by catalysing the conversion of superoxide into oxygen and oxygen peroxide (H2O2). Later, CAT and GSH-PX can convert the obtained hydrogen peroxide to water for protecting cells against damage due to oxygen stress [31]. In vitro studies revealed that some L. plantarum fermentate potently reduce and scavenge free radicals [32,33]. In vivo studies have shown that L. plantarum colonizes the intestinal tract, which has an important effect on protecting against free radicals. Li et al. (2021) [34] found that feeding Lactoplantibacillus plantarum KSFY06 at high concentrations can inhibit the decrease of oxidation associated enzymes and complexes in serum and liver caused by liver injury effectively. Izuddin et al. (2020) [35] reported that dietary with Lactoplantibacillus plantarum supplementation in diet of post weaning lambs can improve antioxidants levels in serum and rumen, as well as, upregulate antioxidase activities in liver and rumen barrier function. Similar to these studies, adding L. plantarum HJLP-1 in diet significantly increased serum T-SOD and CAT activities in broilers. Therefore, L. plantarum HJLP-1 promoted the antioxidant capacity of broilers mainly by scavenging free radicals.
Serum immunoglobulin, a key indicator for animal’s humoral immunity status, represents a defense mechanism against the intrusion of foreign substances into the living body. IgA, IgG (IgY), and IgM represent the critical immunoglobulins with avian species [36,37]. According to our results, supplementation with L. plantarum HJLP-1 enhanced broilers immunity through promoting serum IgM and IgY levels. Such result conformed other studies [38], where feeding a postbiotic produced by L. plantarum HJLP-1 can improve serum IgM and IgY levels in broilers. Based on the fact that cytokines have essential effects on inflammatory and immune responses, the balance of them exerts a critical effect on resisting infection. Increases in pro-inflammatory factors, like TNF-α, IL-6, and IL-1β, induce systemic inflammation reaction and tissue damage [39], while IL-10 is the anti-inflammatory and tolerogenic factor blocking pro-inflammatory factor production [40]. Some studies have reported that L. plantarum could upregulate anti-inflammatory factors while suppressing pro-inflammatory factors within the intestinal mucus of broilers challenged by toxins or pathogens [18,41]. The results of this study revealed that L. plantarum HJLP-1 also decreased serum IL-1β expression while increasing IL-10 expression, which indicated that supplementation with L. plantarum HJLP-1 might be beneficial for attenuating the systemic inflammatory response in broilers.
Numerous studies have concluded that probiotics had positive effects on health, performance, and disease resistant of the hosts, as they can regulate the gut microbial flora balance in hosts by inhibiting the proliferation of pathogenic species and improving the number of beneficial bacteria [33,42,43]. Therefore, the beneficial effects of L. plantarum HJLP-1 on broilers in this study may be attributed to its influence on the endogenous commensal microflora and changes in metabolite production by the intestinal bacteria.
SCFAs, especially butyric acid, propionate, and acetic acid represent main bacterial fermentation end products of dietary components, especially indigestible fibers in the intestine and play important roles in maintaining the health of host. Both of acetic acid and lactic acid were detected in postbiotics produced by a variety of Lactobacillus plantarum [44]. Acetic acid can contribute to environmental acidification and inhibit the growth of less acidophilic organisms (most pathogenic bacterial species) in the same ecosystem [45]. Butyric acid has been shown to have a critical effect on growth promotion [46,47], oxidative stress reduction [48], immune modulation, anti-inflammatory effects [49], and inhibition of colonization by pathogenic bacteria in broilers [50]. According to our results, the dramatically increased butyric acid and acetic acid concentrations in the cecal contents of broilers fed L. plantarum HJLP-1 maybe partly responsible for the beneficial effects of L. plantarum HJLP-1 on broilers.
Diverse microorganisms densely colonized in the gastrointestinal tract of poultry have been considered to have vital effects on host health and growth performance [51]. Our results indicate that L. plantarum HJLP-1 plays a role in modulating the gut microbiota of broilers. L. plantarum can enrich the cecal microbiota communities with unique microbiota, as reflected by significantly increased ACE, Chao richness indices, and abundance of unique OTUs in LP compared to CON and ANT. Compared to the samples of CON group, the decreased Bacteroidetes abundance while increased Firmicutes abundance observed in LP and ANT group samples consider an increase of Firmicutes/Bacteroidetes ratio at the cecal level, which showed positively relation to an increase in BW in broilers [52,53], which may be a possible reason for the growth-promoting effects of L. plantarum and antibiotics. Moreover, As indicated by the spearman correlation heatmap, Ruminococcaceae and Lachnospiraceae that enriched in LP group promote butyric acid production. The Alistip, which enriched in ANT group, also have positive correlation on butyric acid production. While the Bacteroides that enriched in control group inhibit the butyric acid production. This is probably the main reason for the dramatic increase in butyric acid by L. plantarum HJLP-1 and antibiotic.
5. Conclusion
Taken together, Lactoplantibacillus plantarum HJLP-1 supplementation could improve growth performance and promote the antioxidant capacity and immune function of broilers. These beneficial effects of L. plantarum HJLP-1 on broilers may be attributed to its influence on some specific bacterial abundance in the intestinal microbiota, resulting in a dramatic increase in butyric acid and acetic acid content. Therefore, L. plantarum HJLP-1 could be an efficient substitute for antibiotics to promote growth and health of broiler in production.
Author Contributions
Caimei Yang and Yinglei Xu designed this experiment and edited manuscript. Shuting Wang, Qing Li performed animal experiments, Shuting Wang and Ruiqiang Zhang collected samples and analyzed them, and Jie Feng guided the implementation of the experiment and revised the article. All authors read and approved the final manuscript.
Funding
This work was supported by Zhejiang Provincial Key Research and Development Program (No. 2019C02051; No. 2020C02032); Zhejiang Provincial Leading Innovation and Entrepreneurship Team Project (No.2020R01015); Key Laboratory of Animal Feed and Nutrition of Zhejiang Province (No. 202105); the Scientific Research Project of Zhejiang Provincial Education Department (No.YZ20200002) and Vegamax Green Animal Health Products Key Agricultural Enterprise Research Institute of Zhejiang Province (No.2021Y30004).
Institutional Review Board Statement
The animal care protocol was approved by the Animal Ethics Committee of Zhejiang A&F University (SYXKzhe2016-087).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Vegamax Biotechnology Co. Ltd. (Anji, Zhejiang, China) for providing the Lactoplantibacillus plantarum.
Conflicts of Interest
None of authors declare a conflict of interest.
References
- Robins, A. and Phillips, CJC. International approaches to the welfare of meat chickens. Worlds Poult. Sci. J. 2011, 67, 351–369. [Google Scholar] [CrossRef]
- Bellet, C.; Rushton, J. World food security, globalisation and animal farming: unlocking dominant paradigms of animal health science. Rev. Sci. Tech. 2019, 38, 383–393. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Joint FAO/OIE/WHO Expert Workshop on Non-Human Antimicrobial Usage and Antimicrobial Resistance: Scientific Assessment; World Health Organization: Geneva, Switzerland, 2003; No. WHO/CDS/CPE/ZFK/2004.7. [Google Scholar]
- Aidara-Kane, A.; Angulo, F.J.; Conly, J.M.; Minato, Y.; Silbergeld, E.K.; McEwen, S.A.; Collignon, P.J.; WHO Guideline Development Group. World Health Organization (WHO) guidelines on use of medically important antimicrobials in food-producing animals. Antimicrob Resist Infect Control. 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Alagawany, M.; Abd El-Hack, M.E.; Farag, M.R.; Sachan, S.; Karthik, K.; Dhama, K. The use of probiotics as eco-friendly alternatives for antibiotics in poultry nutrition. Environ. Sci. Pollut. R. 2018, 25, 10611–10618. [Google Scholar] [CrossRef] [PubMed]
- Mingmongkolchai, S.; Panbangred, W. Bacillus probiotics: an alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- Al-Khalaifah, H.S. Benefits of probiotics and/or prebiotics for antibiotic-reduced poultry. Poult. Sci. 2018, 97, 3807–3815. [Google Scholar] [CrossRef]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus plantarum and Its Probiotic and Food Potentialities. Probiotics Antimicrob Proteins. 2017, 9(2), 111–122. [Google Scholar] [CrossRef] [PubMed]
- Bosch, M.; Rodriguez, M.; Garcia, F.; Fernandez, E.; Fuentes, M.C.; Cune, J. Probiotic properties of Lactobacillus planta-rum CECT 7315 and CECT 7316 isolated from faeces of healthy children. Lett. Appl. Microbiol. 2012, 54, 240–246. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Zhang, L.; Zhao, Y.; Niu, C.; Yang, Z.; Li, S. Potential probiotic characterization of Lactobacillus plan-tarum strains isolated from Inner Mongolia “Hurood” cheese. J. Microbiol. Biotechnol. 2014, 24, 225–235. [Google Scholar] [CrossRef]
- Gupta, A.; Tiwari, S.K. Probiotic Potential of Lactoplantibacillus plantarum LD1 Isolated from Batter of Dosa, a South Indian Fermented Food. Probiotics Antimicrob. Proteins. 2014, 6, 73–81. [Google Scholar] [CrossRef]
- Huang, R.; Tao, X.; Wan, C.; Li, S.; Xu, H.; Xu, F.; Shah, N.P.; Wei, H. In vitro probiotic characteristics of Lactoplantibacillus plantarum ZDY 2013 and its modulatory effect on gut microbiota of mice. J. Dairy Sci. 2015, 98, 5850–5861. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, F.; Wan, C.; Xiong, Y.; Shah, N.P.; Wei, H.; Tao, X. Evaluation of probiotic properties of Lactobacillus plantarum WLPL04 isolated from human breast milk. J. Dairy Sci. 2016, 99, 1736–1746. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Lee, J.E.; Lim, S.M.; Kim, Y.J.; Lee, N.K.; Paik, H.D. Antioxidant and immune-enhancing effects of probiotic Lactobacillus plantarum 200655 isolated from kimchi. Food Sci. Biotechnol. 2019, 28, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Duary, R.K.; Bhausaheb, M.A.; Batish, V.K.; Grover, S. Anti-inflammatory and immunomodulatory efficacy of indigenous probiotic Lactoplantibacillus plantarum Lp91 in colitis mouse model. Mol. Biol. Rep. 2012, 39, 4765–4775. [Google Scholar] [CrossRef] [PubMed]
- Satish Kumar, C.S.; Kondal Reddy, K.; Reddy, A.G.; Vinoth, A.; Ch, S.R.; Boobalan, G.; Rao, G.S. Protective effect of Lactobacillus plantarum 21, a probiotic on trinitrobenzenesulfonic acid-induced ulcerative colitis in rats. Int. Immunopharmacol. 2015, 25, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Peng, Q.; Jia, H.M.; Zeng, X.F.; Zhu, J.L.; Hou, C.L.; Liu, X.T.; Yang, F.J.; Qiao, S.Y. Prevention of Escherichia coli infection in broiler chickens with Lactobacillus plantarum B1. Poult. Sci. 2017, 96, 2576–2586. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.R.; Liu, Y.L.; Duan, Y.L.; Wang, F.Y.; Guo, F.S.; Yan, F.; Yang, X.J.; Yang, X. Intestinal toxicity of deoxynivalenol is limited by supplementation with Lactobacillus plantarum JM113 and consequentially altered gut microbiota in broiler chickens. J. Anim. Sci. Biotechno. 2018, 9, 74. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Zeng, D.; Zhang, Y.; Ni, X.Q.; Wang, J.; Jian, P.; Zhou, Y.; Li, Y.; Yin, Z.Q.; Pan, K.C.; Jing, B. Lactobacillus plantarum BS22 promotes gut microbial homeostasis in broiler chickens exposed to aflatoxin B1. J. Anim. Physiol. Anim. Nutr. (Berl). 2018, 102, e449–e459. [Google Scholar] [CrossRef] [PubMed]
- Thanh, N.T.; Loh, T.C.; Foo, H.L.; Hair-bejo, M.; Azhar, B.K. Effects of feeding metabolite combinations produced by Lactobacillus plantarumon growth performance, faecal microbial population, small intestine villus height and faecal volatile fatty acids in broilers. Poult. Sci. 2009, 50, 298–306. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, Q. Effect of probiotic on growth performance and digestive enzyme activity of Arbor Acres broilers. Res. Vet. Sci. 2010, 89, 163–167. [Google Scholar] [CrossRef]
- Salim, H.M.; Kang, H.K.; Akter, N.; Kim, D.W.; Kim, J.H.; Kim, M.J.; Na, L.P.; Jong, H.B.; Choi, H.C.; Suh, O.S.; Kim, W.K. Supplementation of direct-fed microbials as an alternative to antibiotic on growth performance, immune response, cecal microbial population, and ileal morphology of broiler chickens. Poult. Sci. 2013, 92, 2084–2090. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, L.; Zhan, X.; Zeng, X.; Zhou, L.; Cao, G.; Chen, A.; Yang, C. Effects of dietary supplementation of probiotic, Clostridium butyricum, on growth performance, immune response, intestinal barrier function, and digestive enzyme activity in broiler chickens challenged with Escherichia coli K88. J. Anim. Sci. Biotechnol. 2016, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Zeng, X.F.; Zhu, J.L.; Wang, S.; Liu, X.T.; Hou, C.L.; Thacker, P.A.; Qiao, S.Y. Effects of dietary Lactobacillus plantarum B1 on growth performance, intestinal microbiota, and short chain fatty acid profiles in broiler chickens. Poult. Sci. 2016, 95, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Deepthi, B.V.; Somashekaraiah, R.; Poornachandra Rao, K.; Deepa, N.; Dharanesha, N.K.; Girish, K.S.; Sreenivasa, M.Y. Lactoplantibacillus plantarum MYS6 Ameliorates Fumonisin B1-Induced Hepatorenal Damage in Broilers. Front. Microbiol. 2017, 8, 2317. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Ma, C.; Sun, Z.; Wang, L.; Huang, S.; Su, X.; Xu, J.; Zhang, H. Feed-additive probiotics accelerate yet antibiotics delay intestinal microbiota maturation in broiler chicken. Microbiome. 2017, 5, 91. [Google Scholar] [CrossRef] [PubMed]
- Donsbough, A.L.; Powell, S.; Waguespack, A.; Bidner, T.D.; Southern, L.L. Uric acid, urea, and ammonia concentrations in serum and uric acid concentration in excreta as indicators of amino acid utilization in diets for broilers. Poult. Sci. 2010, 89, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Chen, G.; Liu, H. Antioxidative Categorization of Twenty Amino Acids Based on Experimental Evaluation. Molecules. 2017, 22, 2066. [Google Scholar] [CrossRef] [PubMed]
- Perl, A. Review: Metabolic Control of Immune System Activation in Rheumatic Diseases. Arthritis Rheumatol. 2017, 69, 2259–2270. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Wang, L.; Cheng, S.; Zhou, Y.; Ma, L. Protective Effects of alpha-Lipoic Acid on Vascular Oxidative Stress in Rats with Hyperuricemia. Curr. Med. Sci. 2019, 39, 920–928. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef]
- Tang, W.; Xing, Z.Q.; Li, C.; Wang, J.J.; Wang, Y.P. Molecular mechanisms and in vitro antioxidant effects of Lactobacillus plantarum MA2. Food Chem. 2017, 221, 1642–1649. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Cao, G.T.; Ferket, P.R.; Liu, T.T.; Zhou, L.; Zhang, L.; Xiao, Y.P.; Chen, A.G. Effects of probiotic, Clostridium butyricum, on growth performance, immune function, and cecal microflora in broiler chickens. Poult. Sci. 2012, 91, 2121–2129. [Google Scholar] [CrossRef]
- Li, C.; Si, J.; Tan, F.; Park, K.Y.; Zhao, X. Lactobacillus plantarum KSFY06 Prevents Inflammatory Response and Oxidative Stress in Acute Liver Injury Induced by D-Gal/LPS in Mice. Drug Des. Devel. Ther. 2021, 15, 37–50. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A. Dietary Postbiotic Lactobacillus plantarum Improves Serum and Ruminal Antioxidant Activity and Upregulates Hepatic Antioxidant Enzymes and Ruminal Barrier Function in Post-Weaning Lambs. Antioxidants (Basel). 2020, 9, 250. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Zhen, W.R.; Geng, Y.Q.; Wang, Z.; Guo, Y.M. Effects of dietary Enterococcus faecium NCIMB 11181 supplementation on growth performance and cellular and humoral immune responses in broiler chickens. Poult. Sci. 2019, 98, 150–163. [Google Scholar] [CrossRef]
- Zhu, F.; Zhang, B.; Li, J.; Zhu, L. Effects of fermented feed on growth performance, immune response, and antioxidant capacity in laying hen chicks and the underlying molecular mechanism involving nuclear factor-kappaB. Poult. Sci. 2020, 99, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Humam, A.M.; Loh, T.C.; Foo, H.L.; Samsudin, A.A.; Mustapha, N.M.; Zulkifli, I.; Izuddin, W.I. Effects of Feeding Different Postbiotics Produced by Lactobacillus plantarum on Growth Performance, Carcass Yield, Intestinal Morphology, Gut Microbiota Composition, Immune Status, and Growth Gene Expression in Broilers under Heat Stress. Animals (Basel). 2019, 9, 644. [Google Scholar] [CrossRef]
- Kim, E.Y.; Moudgil., K.D. Immunomodulation of autoimmune arthritis by pro-inflammatory cytokines. Cytokine. 2017, 98, 87–96. [Google Scholar] [CrossRef]
- Sabat, R.; Grutz, G.; Warszawska, K.; Kirsch, S.; Witte, E.; Wolk, K.; Geginat, J. Biology of interleukin-10. Cytokine Growth Factor Rev. 2010, 21, 331–344. [Google Scholar] [CrossRef]
- Xu, T.; Chen, Y.; Yu, L.; Wang, J.; Huang, M.; Zhu, N. Effects of Lactobacillus plantarum on intestinal integrity and immune responses of egg-laying chickens infected with Clostridium perfringens under the free-range or the specific pathogen free environment. BMC Vet. Res. 2020, 16, 47. [Google Scholar] [CrossRef]
- Shin, D.; Chang, S.Y.; Bogere, P.; Won, K.; Choi, J.Y.; Choi, Y.J.; Lee, H.K.; Hur, J.; Park, B.Y.; Kim, Y.; Heo, J. Beneficial roles of probiotics on the modulation of gut microbiota and immune response in pigs. PLoS One. 2019, 14, e0220843. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.; Das, R.; Oak, S.; Mishra, P. Probiotics (Direct-Fed Microbials) in Poultry Nutrition and Their Effects on Nutrient Utilization, Growth and Laying Performance, and Gut Health: A Systematic Review. Animals (Basel). 2020, 10, 1863. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Foo, H.L.; Loh, T.C.; Lim, E.T.C.; Abdul Mutalib, N.E. Comparative Studies of Inhibitory and Antioxidant Activities, and Organic Acids Compositions of Postbiotics Produced by Probiotic Lactiplantibacillus plantarum Strains Isolated From Malaysian Foods. Front Vet Sci. 2021, 7, 602280. [Google Scholar] [CrossRef]
- Pessione, E. Lactic acid bacteria contribution to gut microbiota complexity: lights and shadows. Front Cell Infect Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef] [PubMed]
- Bedford, A.; Yu, H.; Squires, E.J.; Leeson, S.; Gong, J. Effects of supplementation level and feeding schedule of butyric acid glycerides on the growth performance and carcass composition of broiler chickens. Poult. Sci. 2017, 96, 3221–3228. [Google Scholar] [CrossRef] [PubMed]
- Aljumaah, M.R.; Alkhulaifi, M.M.; Abudabos, A.M.; Alabdullatifb, A.; El-Mubarak, A.H.; Al Suliman, A.R.; Stanley, D. Organic acid blend supplementation increases butyric acid and acetic acid production in Salmonella enterica serovar Typhimurium challenged broilers. PLoS One. 2020, 15, e0232831. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, Y.; Lu, C.; Ahmad, H.; Zhang, H.; He, J.; Zhang, L.; Wang, T. Influence of Butyric acid Loaded Clinoptilolite Dietary Supplementation on Growth Performance, Development of Intestine and Antioxidant Capacity in Broiler Chickens. Plos One. 2016, 11, e0154410. [Google Scholar] [CrossRef]
- Zhang, W.H.; Jiang, Y.; Zhu, Q.F.; Gao, F.; Dai, S.F.; Chen, J.; Zhou, G.H. Sodium butyric acid maintains growth performance by regulating the immune response in broiler chickens. Br. Poult. Sci. 2011, 52, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Ji, J.; Qu, H.; Wang, J.; Shu, D.M.; Wang, Y.; Liu, T.F.; Li, Y.; Luo., C.L. Effects of sodium butyric acid on intestinal health and gut microbiota composition during intestinal inflammation progression in broilers. Poult. Sci. 2019, 98, 4449–4456. [Google Scholar] [CrossRef]
- Gong, Y.; Yang, H.; Wang, X.; Xia, W.; Lv, W.; Xiao, Y.; Zou, X. Early intervention with cecal fermentation brothregulates the colonization and development of gut microbiota inbroiler chickens. Front. Microbiol. 2019, 10, 1422. [Google Scholar] [CrossRef]
- Salaheen, S.; Kim, S.W.; Haley, B.J.; Van Kessel, J.A.S.; Biswas, D. Alternative Growth Promoters Modulate Broiler Gut Microbiome and Enhance Body Weight Gain. Front. Microbiol. 2017, 8, 2088. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Cheng, Y.; Li, Y.; Li, X.; Zhou, Z.; Shi, D.; Li, Z.; Xiao, Y. Preliminary Study on the Effect of Bacillus amyloliquefaciens TL on Cecal Bacterial Community Structure of Broiler Chickens. Biomed. Res. Int. 2019, 5431354. [Google Scholar] [CrossRef]
Figure 1.
Analysis of the composition of the fecal microbiota. (A) The Venn diagram summarizing the numbers of common and unique observed taxonomic units (OTUs) in the microflora community in the cecum contents of broiler chicken. (B) Principal component analysis (PCA). (C) Shannon, Simpson, ACE, and chao indices reflecting alpha diversity. (D) Microbiota composition at the phylum level. (E) Histogram of LDA scores for taxonomic biomarkers identified by LEfSe. LDA scores (log 10) > 4 indicate enriched taxa in cases. Significance was determined using one-way ANOVA.* represents P < 0.05, ** represents P < 0.01, and *** represents P < 0.001.
Figure 1.
Analysis of the composition of the fecal microbiota. (A) The Venn diagram summarizing the numbers of common and unique observed taxonomic units (OTUs) in the microflora community in the cecum contents of broiler chicken. (B) Principal component analysis (PCA). (C) Shannon, Simpson, ACE, and chao indices reflecting alpha diversity. (D) Microbiota composition at the phylum level. (E) Histogram of LDA scores for taxonomic biomarkers identified by LEfSe. LDA scores (log 10) > 4 indicate enriched taxa in cases. Significance was determined using one-way ANOVA.* represents P < 0.05, ** represents P < 0.01, and *** represents P < 0.001.
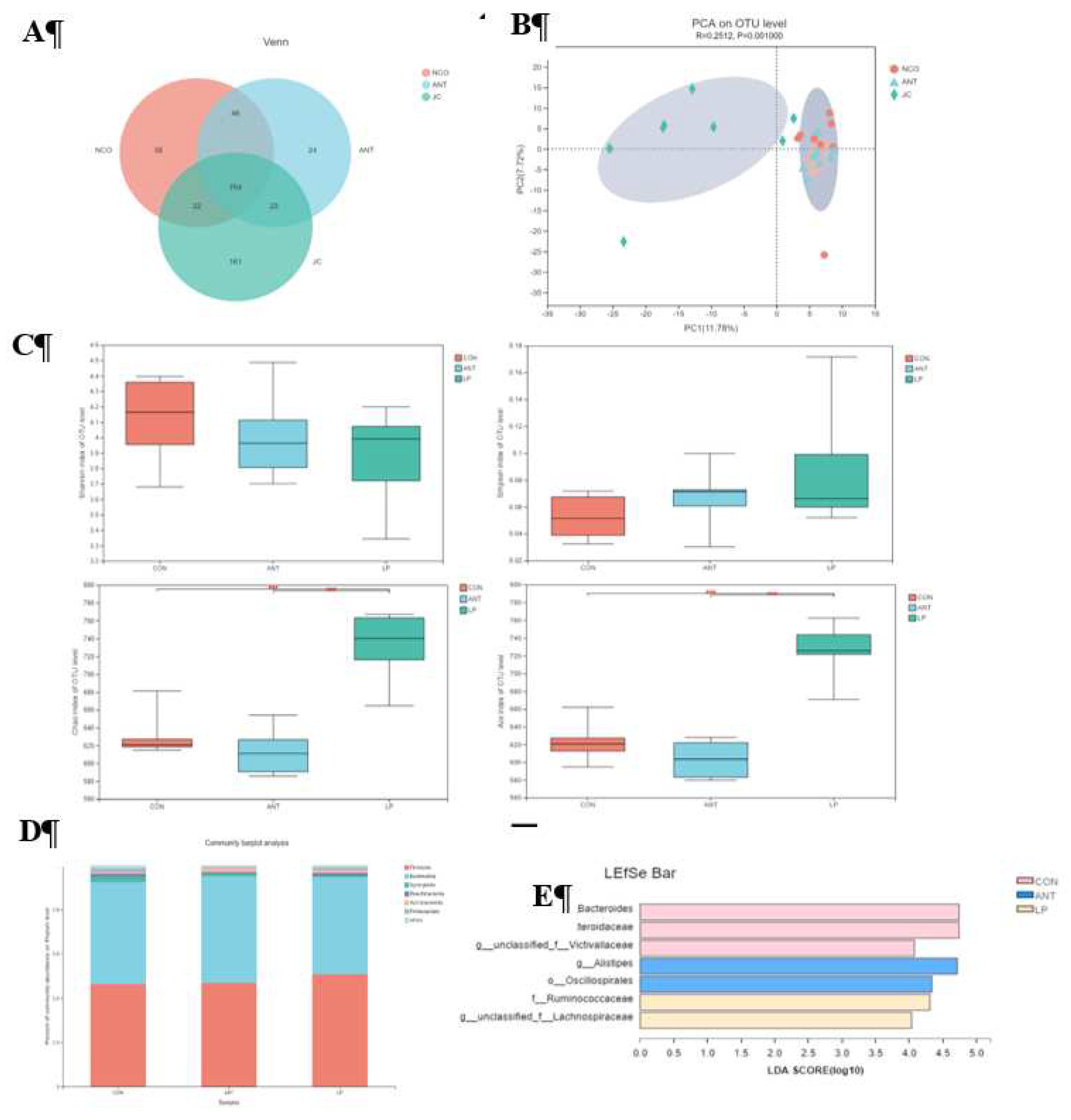
Figure 2.
Effects of Lactobacillus plantarum on SCFAs in broilers. Values are presented as mean ± SEM. Individual broiler was regarded as the experimental unit. n = 8 per treatment. Different superscripts differ significantly (p < 0.05). Abbreviations: CON represents broilers fed a basal diet; ANT represents the broilers fed a basal diet supplemented with 75 mg/kg chlortetracycline; LP represents the broilers fed a basal diet and Lactobacillus plantarumon.
Figure 2.
Effects of Lactobacillus plantarum on SCFAs in broilers. Values are presented as mean ± SEM. Individual broiler was regarded as the experimental unit. n = 8 per treatment. Different superscripts differ significantly (p < 0.05). Abbreviations: CON represents broilers fed a basal diet; ANT represents the broilers fed a basal diet supplemented with 75 mg/kg chlortetracycline; LP represents the broilers fed a basal diet and Lactobacillus plantarumon.
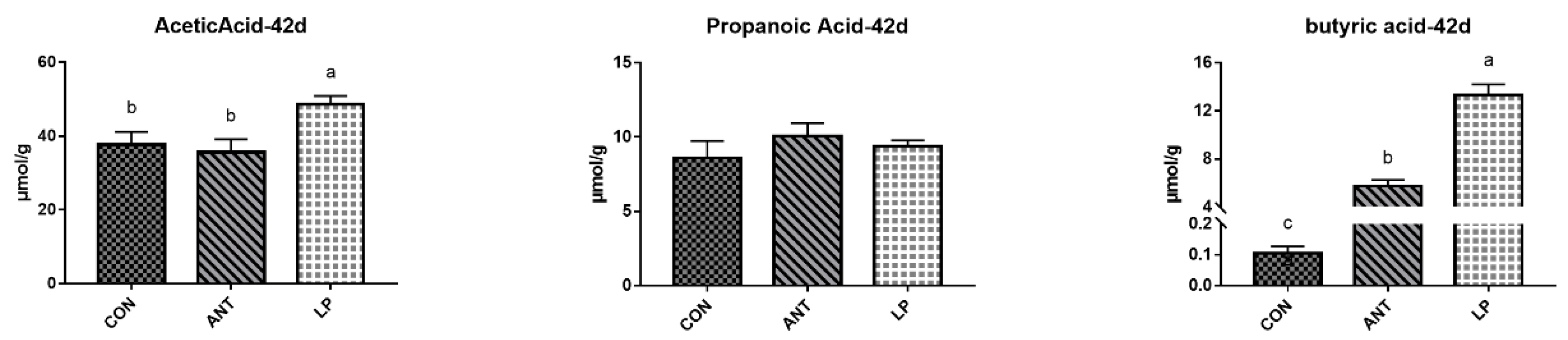
Figure 3.
Correlation heatmap of the top 50 genera and SCFAs in broilers. Significance was determined using one-way ANOVA. * represents P < 0.05, ** represents P < 0.01, and *** represents P < 0.001.
Figure 3.
Correlation heatmap of the top 50 genera and SCFAs in broilers. Significance was determined using one-way ANOVA. * represents P < 0.05, ** represents P < 0.01, and *** represents P < 0.001.
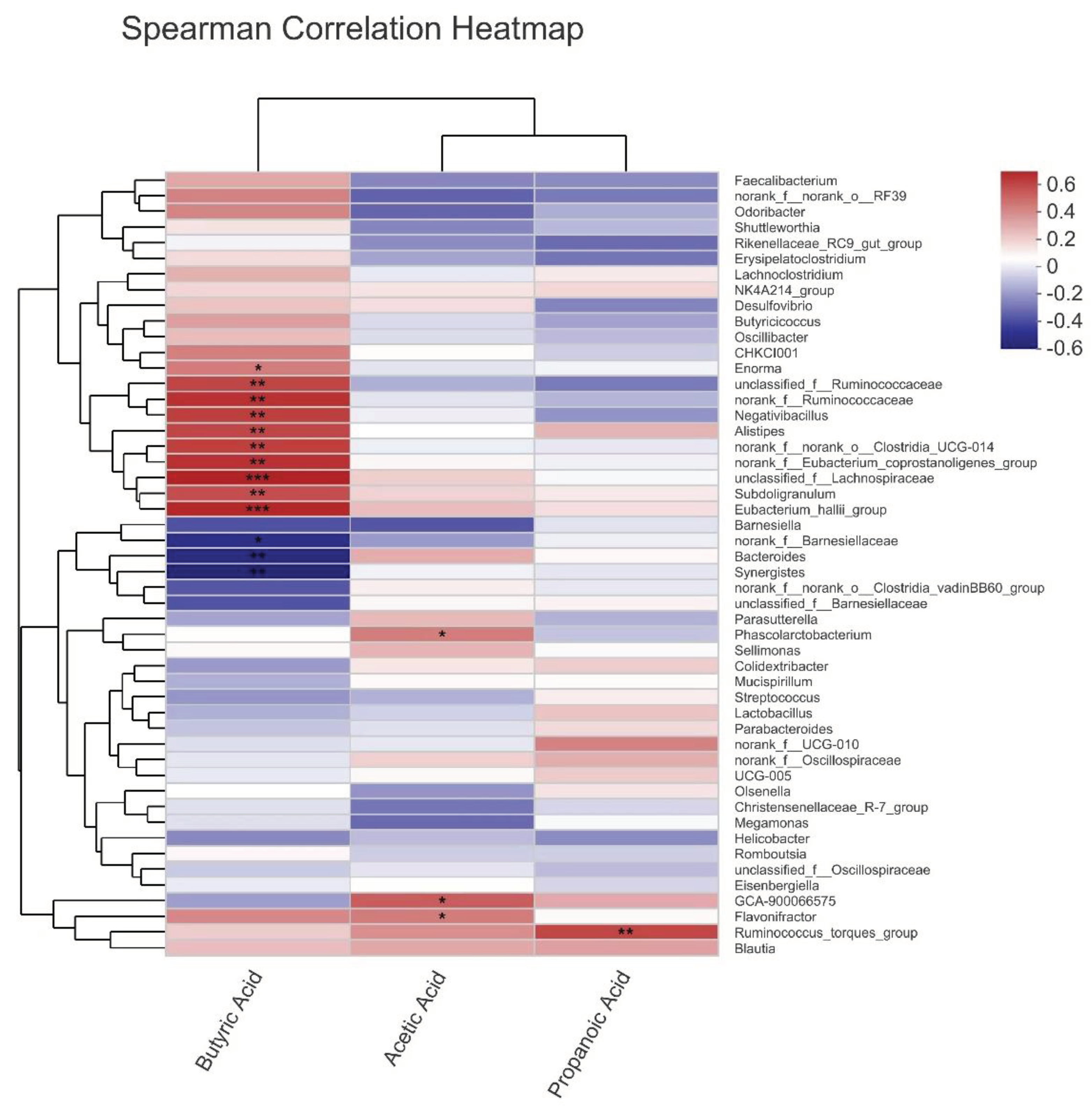

Table 1.
Raw material composition and nutritional level of basic dietary (air-dry basis).
| Items | 1-21d | 22-42d |
|---|---|---|
| Ingredients(%) | ||
| Corn | 61.80 | 65.60 |
| Soybean meal | 22.50 | 17.55 |
| Extruded soybean | 8.45 | 10.00 |
| Import fish meal | 3.00 | 3.00 |
| CaHPO4 | 1.66 | 1.45 |
| Limestone | 1.10 | 1.00 |
| NaCl | 0.32 | 0.30 |
| DL- methionine | 0.16 | 0.10 |
| L- lysine | 0.01 | |
| Premix1 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 |
| Nutrition levels | ||
| Metabolizable energy (MJ/kg) | 12.45 | 12.70 |
| Crude protein | 21.00 | 19.20 |
| Lysine | 1.15 | 0.95 |
| Methionine | 0.54 | 0.44 |
| Calcium | 0.99 | 0.89 |
| Available phosphorus | 0.53 | 0.49 |
1 Premix is provided for feed per kg: VA 1500 IU, VB1 1.5 mg, VB6 3.0 mg, VB12 0.01 mg, VD3 200 IU, VE 10 IU, VK 0.5 mg, Biotin 0.15 mg, D-pantothenic acid 10 mg, Folic acid 0.5 mg, Nicotinic acid 30 mg, Trace elements Cu, Fe, Zn, Mn, Se, I are 8 mg, 80 mg, 40 mg, 60 mg, 0.15 mg, 0.18 mg respectively.
Table 2.
Effects of dietary supplementation of L. plantarum on growth performance of broilers.
| Items | SEM | P-Value | |||
|---|---|---|---|---|---|
| CON | ANT | LP | |||
| BW,g | |||||
| 1 d | 38.29 | 39.52 | 38.32 | 0.401 | 0.361 |
| 21 d | 732.08b | 778.24a | 800.13a | 8.945 | 0.001 |
| 42 d | 1875.86b | 1926.47b | 2124.36a | 40.63 | 0.019 |
| ADG (g/d) | |||||
| 1-21 d | 33.54b | 35.26a | 36.31a | 0.361 | 0.001 |
| 22-42 d | 55.48 b | 56.47b | 64.85 a | 1.764 | 0.048 |
| 1-42 d | 44.51b | 45.87b | 52.57a | 1.205 | 0.005 |
| ADFI,g | |||||
| 1-21 d | 54.02b | 54.42ab | 59.84a | 0.935 | 0.008 |
| 22-42 d | 118.19 | 111.66 | 118.63 | 3.432 | 0.677 |
| 1-42 d | 80.00 | 78.62 | 86.05 | 1.516 | 0.098 |
| F:G | |||||
| 1-21 d | 1.61ab | 1.54b | 1.64a | 0.017 | 0.036 |
| 22-42 d | 2.13a | 1.98ab | 1.83b | 0.046 | 0.023 |
| 1-42 d | 1.80a | 1.72ab | 1.70b | 0.018 | 0.067 |
In the same row with different superscripts differ significantly (p < 0.05). Pen was regarded as the experimental unit. n = 8 per treatment. Abbreviations: CON represents broilers fed a basal diet; ANT represents the broilers fed a basal diet supplemented with 75 mg/kg chlortetracycline; LP represents the broilers fed a basal diet and L. plantarumon. F: G means feed-to-gain ratio.
Table 3.
Effects of L. plantarum on metabolite index in serum of broilers.
| Items | SEM | P-Value | |||
|---|---|---|---|---|---|
| CON | ANT | LP | |||
| NH3(µmol/L) | 10.83a | 9.03b | 6.54c | 0.490 | 0.001 |
| BUN (mmol/L) | 0.31 | 0.29 | 0.23 | 0.016 | 0.136 |
| UA (µmol/L) | 147.84 | 129.88 | 116.41 | 6.584 | 0.148 |
| XOD (U/L) | 3.83a | 4.30a | 3.10b | 0.168 | 0.005 |
In the same row with different superscripts differ significantly (p < 0.05). Individual broiler was regarded as the experimental unit. n = 8 per treatment. Abbreviations: CON represents broilers fed a basal diet; ANT represents the broilers fed a basal diet supplemented with 75 mg/kg chlortetracycline; LP represents the broilers fed a basal diet and Lactobacillus plantarumon; BUN means blood urea nitrogen; UA means urea acid; XOD means xanthine oxidase.
Table 4.
Effects of L.plantarum on antioxidant index in serum of broilers.
| Items | SEM | P-Value | |||
|---|---|---|---|---|---|
| CON | ANT | LP | |||
| GSH-PX (U/mL) | 6. 90 | 7.83 | 8.37 | 0.591 | 0.617 |
| SOD (U/mL) | 12.28b | 12.96ab | 13.22a | 0.155 | 0.026 |
| CAT (U/mL) | 9.98b | 10.79ab | 11.80a | 0.304 | 0.039 |
| MDA (U/mL) | 8.12 | 7.56 | 7.91 | 0.199 | 0.534 |
In the same row with different superscripts differ significantly (p < 0.05). Individual broiler was regarded as the experimental unit. n = 8 per treatment. Abbreviations: CON represents broilers fed a basal diet; ANT represents the broilers fed a basal diet supplemented with 75 mg/kg chlortetracycline; LP represents the broilers fed a basal diet and Lactobacillus plantarumon; GSH-PX means glutathione peroxidase; SOD means superoxide dismutase; CAT means catalase; MDA means malonaldehyde.
Table 5.
Effects of L.plantarum on serum immunoglobulins and immune response cytokines of broilers.
| Items | SEM | P-Value | |||
|---|---|---|---|---|---|
| CON | ANT | LP | |||
| IgA ng/mL | 168.11 | 177.93 | 176.81 | 4.327 | 0.626 |
| IgY ng/mL | 1.49c | 1.79b | 2.07a | 0.067 | 0.001 |
| IgM µg/mL | 3.09c | 3.61b | 4.26a | 0.143 | 0.001 |
| IL-1βpg/mL | 94.19a | 83.02b | 83.02b | 1.900 | 0.011 |
| IL-6pg/mL | 394.04 | 393.77 | 387.22 | 4.480 | 0.801 |
| IL-10pg/mL | 17.43b | 20.82b | 28.43a | 1.272 | 0.001 |
| TNF-αpg/L | 38.74ab | 40.23a | 37.35b | 0.568 | 0.114 |
| INF-β pg/mL | 150.78 | 149.55 | 145.68 | 4.573 | 0.905 |
In the same row with different superscripts differ significantly (p < 0.05). Individual broiler was regarded as the experimental unit. n = 8 per treatment. Abbreviations: CON represents broilers fed a basal diet; ANT represents the broilers fed a basal diet supplemented with 75 mg/kg chlortetracycline; LP represents the broilers fed a basal diet and Lactobacillus plantarumon.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



