
Submitted:
01 December 2023
Posted:
04 December 2023
You are already at the latest version
Abstract
Increasing evidence of sexual dimorphism in pathophysiology of metabolic complications caused by sex steroids is under investigation. The gut microbiota represents a complex microbial ecosystem involved in energy metabolism, immune response, nutrition acquisition, and health of host organisms. Gender-specific differences in composition are present between females and males. The purpose of this study was to use cross-sex fecal microbiota transplantation (FMT) for the detection of sex-dependent metabolic, hormonal, and gut microbiota changes in female and male recipients. Healthy non-obese female and male Wistar rats were divided into donor, same-sex, and cross-sex recipient groups. After a 30-day period of FMT administration, biochemical markers (glucose and lipid metabolism) and sex hormones were measured and gut microbiota was analyzed. Cross-sex male recipients displayed significantly lower testosterone concentration compared to males that received same-sex FMT. Sex-dependent changes caused by cross-sex FMT were detected, while several bacterial taxa correlated with plasma testosterone levels. This study represents the first to study the effect of cross-sex changes in the gut microbiome concerning metabolic and hormonal changes/status in adult non-obese Wistar rats. Herein, we present the cross-sex FMT as a potential tool to modify sex-specific pathologies.
Keywords:
sex differences
; testosterone
; gut microbiota
; microbiome
; fecal microbiota transplantation
; metabolic disease
; cross-sex
1. Introduction
The prevalence of metabolic diseases such as obesity, metabolic syndrome, and type 2 diabetes is increasing worldwide [1,2,3] . There is evidence of sexual dimorphism in the metabolic complications caused by sex hormones such as estrogens and androgens [4,5,6]. Gut microbiota plays a crucial physiological role in the local and systemic immune response and energy extraction. Dysbiosis of gut microbiota appears to be involved in various diseases including metabolic disorders [7]. The overall incidence of obesity is higher in women compared to men with increasing age resulting in abdominal fat gain. Moreover, the prevalence of metabolic syndrome is also greater in women, while the incidence of type 2 diabetes (T2D) is reversed. In men, a higher prevalence of T2D occurs before puberty, while in women after menopause [8,9,10]. These differences in the development of obesity are supported by differences in fat distribution. While in men and postmenopausal women, fat is distributed in an abdominal (visceral) fat pattern, in premenopausal women it is a typical peripheral (subcutaneous) distribution pattern. Both patterns are strongly regulated and related to sex steroid changes during life in both genders [11]. The physiological changes in sex steroid concentrations after menopause lead to fat redistribution [12], while androgen deprivation in men results in increased adiposity in a visceral fat mass [13]. Testosterone supplementation in non-obese men with androgen deficiency leads to a decrease in visceral adiposity, improved glycemic control, and insulin sensitivity [14,15]. The gut microbiota represents a complex microbial ecosystem strongly related to nutrition acquisition, energy metabolism, immune response, and regulating the metabolic health of the host [16,17,18]. The role of gut microbiota in metabolic disorders has been investigated in recent years [19,20,21]. Gender-specific differences in gut microbiota composition are evident between males and females [22], however, the effect of cross-sex fecal microbiota transplantation (FMT) on these differences has not yet been elucidated. Research on this specific topic is limited, and little is known about the susceptibility of both sexes to receive a transplant or about the ability of the sex-specific transplant to colonize and rebuild the donor ecosystem. The present study hypothesizes that the cross-sex replacement of gut microbiome might change the hormonal and metabolic status in non-obese, healthy female and male rats. Therefore, obesity and metabolic markers (body weight, glycemia, cholesterol, AST, uric acid), sex hormones, and gut microbiome composition of female and male rats served as donors of FMT were assessed in a cross-sex and same-sex manner and compared with hormonal and metabolic status of FMT recipient rats. To the best of our knowledge, this paper is the first to study the effect of cross-sex changes in the gut microbiome concerning metabolic and hormonal changes/status in adult non-obese Wistar rats.
2. Results
2.1. Body Weight and Food Intake
The body weight gain in female rats increased over time (time: F(4,64)=3.839; p<0.05) in all experimental groups, while the effect of FMT was not observed (F(2,16)=0.67; p=ns) (Figure 1A). In males, the body weight did not change during the FMT administration (time: F(4,68)=1.34; p=ns; FMT: F(2,17)=0.088; p=ns, Figure 1B). Cumulative food intake in female rats was increased during the FMT administration in all experimental groups (time: F(4,36)=1476; p<0.001; FMT:(F(2,9)=38.37; p<0.001). Both the same-sex recipient and cross-sex recipient female rats consumed significantly more food during the FMT administration (day 1, day 7, day 14, day 21, day 28; all: p<0.05 for same-sex and cross-sex recipient females) in comparison to donor female rats (FIg. 1.C). Donor female rats consumed less food during the FMT administration compared to both, same-sex recipient (day 1, day 7, day 14, day 21, day 28; all: p<0.05), and cross-sex recipient female rats (day 1, day 7, day 14, day 21, day 28; all: p<0.05, Figure 1C). Cumulative food consumption in male rats increased over time (F(4,32)=368.1; p<0.001), but a significant effect of the FMT on food consumption was not detected (F(2,8)=1.067; p=ns, Figure 1D). Regarding terminal body weight, a significant effect of sex (F(1,33)=198; p<0.001) was observed: donor females displayed lower body weight compared to donor males (t(33)=6.765; p<0.001), same-sex recipient females displayed lower body weight than same-sex recipient males (t(33)=8.747; p<0.001), and cross-sex recipient females had lower body weight than cross-sex recipient males (t(33)=9.084; p<0.001, Figure 1A). FMT did not affect the terminal body weight of rats (F(2,33)=0.373; p=ns, Figure 1A).
2.2. Metabolic Parameters
Two-way ANOVA showed a main effect of sex on fasting insulin concentrations (F(1,33)=10.64; p<0.01), a quantitative index of insulin sensitivity (QUICKI; F(1,33)=14.02; p<0.001), circulating LDL (F(1,33)=8.94; p<0.01) and triacylglycerol concentrations (F(1,33)=5.142; p<0.05). Fasting glycemia, plasma total cholesterol and HDL cholesterol concentration, AST, ALT, and uric acid concentration were not affected by sex (all: p=ns). FMT did not affect any of the metabolic parameters (all: p=ns, Figure 2B–K).
2.3. Testosterone
Plasma testosterone concentrations showed sex differences with higher testosterone concentrations in males (F(1,33)=30.13; p<0.001) than females. Female donors displayed lower concentrations of testosterone compared to male donors (t(33)=3.361; p<0.01). Two-way ANOVA indicated a significant effect of 30-day-long FMT application on circulating testosterone concentration (F(2,33)=4.455; p<0.05). Same-sex recipient females displayed lower concentrations of plasma testosterone compared to same-sex recipient males (t(33)=5.089; p<0.001), while no sex differences between cross-sex females and males were detected (t(33)=1.335; p=ns). Lower testosterone concentrations were detected in cross-sex male recipients compared to same-sex male recipients (t(33)=3.738; p<0.01, Figure 3A).
2.4. Gut Microbiome
We analyzed the 16S rRNA gene amplicon sequencing data of the gut microbiome samples in female and male recipients that received either same-sex or cross-sex FMT, as well as the donors’ samples. Alpha diversity analysis, represented by the Shannon index, indicated no variations in microbial richness and evenness of same-sex females, cross-sex females, same-sex males, and cross-sex females (p=ns, data not shown). Similarly, the initial principal component analysis (PCA) did not show any specific clustering patterns of samples corresponding to the four recipient groups when the microbial composition was based on ASV level (Figure 3B), neither on phylum, class, order, family, and genus levels. Importantly, the female and male donor pool samples were clearly separated from each other suggesting sex-based microbiome composition differences in donor mice. To examine any minor compositional variations between groups, we performed a redundancy analysis (RDA). The RDA results revealed specific group clustering of individual samples and segregation of the four groups, suggesting a potential association between specific microbial communities and sex (R2= 0.2921, p<0.05, permutation test; Supplementary Figure S1). To obtain a deeper insight into the sex-based differences in FMT, we performed additional RDA for sub-groups: mice receiving FMT from female donors (same-sex recipient females and cross-sex recipient males), mice receiving FMT from male donors (same-sex recipient males and cross-sex recipient females), all female recipients (receiving FMT from females - same-sex females, and receiving FMT from males - cross-sex females), and all male recipients (receiving FMT from males - same-sex males, and receiving FMT from females- cross-sex males). The results suggest that the FMT was successfully transplanted in the case of male donors, showing that same-sex males and cross-sex females receiving male FMT had significantly different microbiome composition (R2= 0.3932, p<0.01), where ASVs belonging to orders Bacilli RF39, Clostridia UCG-014, Clostridia vadin BB60, Gastranaerophilales, family Lachnospiraceae, Oscillospiraceae, and genera Alistipes, Oscillibacter, Oscillibacteraceae NK4A214 group (p<0.01) were enriched in the female recipients and ASVs belonging to class Bacteroidia, order Bacteroidales (p≤ 0.001), family Lactobacillaceae and genera Lactobacillus (p≤ 0.01) were enriched in male recipients (Figure 3D). In contrast, the same-sex females and cross-sex males who received female FMT did not differ significantly by their microbiome composition (R2= 0.2065, p= ns; Figure 3C). Accordingly, the male recipients reacted differently to FMT from male and female donors (R2= 0.3000, p<0.05). The microbiome of male recipients that received FMT from males was characterized by an increased proportion of ASVs belonging to the class Bacteroidia, family Lachnospiraceae, and unidentified ASV_285 bacteria (p≤ 0.001), order Bacteroidales, genera Lachnospiraceae NK4A136 group and Eubacterium coprostanoligenes group (p<0.01). Male recipients that received FMT from females contained an increased proportion of unidentified ASV_314 bacteria (p<0.001), and ASVs belonging to orders Bacilli RF39, Clostridia UCG-014, Bacteroidales, Gastranaerophilales, and genera Streptococcus, Frisingicoccus, Family XIII AD3011 group (p<0.01; Fig.3.F). In contrast, the female recipients maintained nearly the same microbiome composition, independent of whether they received FMT from females or males (R2= 0.1854, p= ns; Figure 3E). To reveal specific ASVs that could be associated with plasma concentrations of testosterone (TST), Spearman correlation analysis was performed. Several bacterial species showed positive or negative correlations with plasma concentrations of TST. In males, the abundance of Bacteroides negatively correlated with TST levels in males (including same-sex and cross-sex males, R= -0.7055; p<0.01; Figure 3G), while in females this effect was not observed (same-sex females and cross-sex females, R= -0.429; p=ns; Figure 3K). Circulating concentrations of TST in same-sex recipient females and cross-sex recipient males negatively correlated with the proportion of Frisingicoccus (R= -0.7217; p<0.01; Figure 3H), while in female recipient groups, we did not observe any significant association between TST and Frisingicoccus abundance (R= -0.2045; p= ns; Figure 3L). The proportion of Lachnospiraceae positively correlated with plasma TST concentration in male recipient groups (R= 0.7961; p<0.01; Figure 3I), while in female recipients no significant association was observed (R= -0.2153; p= ns; Figure 3M). The proportion of a specific unidentified ASV_314 showed a negative correlation with plasma TST concentrations in male recipients (R= 0.6983; p<0.01; Figure 3J), but not in female recipients (R= -0.0445; p= ns; Figure 3N).
3. Discussion
Sexual dimorphism plays a crucial role in the pathogenesis of various metabolic disorders; therefore, the pathophysiological role of sex steroids has been studied in recent years. Sex differences are evident in the prevalence and severity of metabolic disorders, including obesity and metabolic syndrome [23,24,25,26]. The role of biological sex in the pathophysiology of metabolic disorders is still not fully understood. Interestingly, gut microbiota plays an important role in host metabolism and immunity [27]. Gut microbiota dysbiosis is associated with the development of numerous disorders. Recently, treatment based on modification of gut microbiota has been proposed and investigated in many immune, auto immune, and metabolic diseases [28,29,30,31]. Fecal microbiota transplantation (FMT) seems to be one of the tools to therapeutically shape the gut microbiota. Various components of metabolism, including energy balance and the metabolism of glucose and lipids, are controlled in a sexually dimorphic manner. In the present study, we have shown that sex differences in non-obese healthy rats are in body weight, insulin sensitivity, and plasma lipid markers (LDL, triacylglycerols). Sex differences in glucose and lipid metabolism have been previously researched [4,32]. FMT in same-sex and cross-sex recipient females and males did not affect the sex differences in the mentioned metabolic components. Although, plasma testosterone concentrations differed between the female and male donor and same-sex recipient subjects. Interestingly, there was no sex difference between cross-sex recipient female and male animals. Male rats receiving FMT from female donors displayed lower plasma concentrations of testosterone compared to male recipients receiving same-sex FMT. Sex hormones, such as estrogens, progesterone, and testosterone are major determinants of differences between sexes in mammals. Females and males displayed the same sex hormone types but with different organ production, concentration in blood, and interaction with the target organ [33]. Ongoing discussion is focused on the role of the interaction of sex hormones with a composition of the gut microbiota and its function in sexual differences in many disorders. These findings suggest the important role of sex steroids in the composition of gut microbiota. It was shown that colon tissue can express molecules involved in steroidogenesis, and subsequently synthesis and metabolism of testosterone [34]. We showed that a higher abundance of Bacteroides in male rats who received same-sex, and cross-sex FMT was associated with lower plasma testosterone concentrations. It was previously shown that higher Porphyromonadaceae (phylum Bacteroidetes) abundance in gut microbiota composition was detected in control females and males following castration. Moreover, castrated adult male mice showed the same gut microbiota composition compared to adult female mice [35]. On the other hand, in both recipient female groups, we did not observe any association between Bacteroides abundance and testosterone concentration. In the present study, a higher abundance of the Lacchnospiraceae (phylum Firmicutes) in males was associated with higher plasma concentrations of testosterone. The Firmicutes can promote the synthesis of testosterone and boost the host organism [36]. In ovariectomized females comparison to control males, Kaliannan et al. observed a drop in the abundance of the Proteobacteria phylum, a decrease in the Firmicutes/Bacteroides ratio, a higher Bifidobacterium/Enterobacteriacea ratio, and an increase in the abundance of Akkermansia (phylum Verrucomicrobia) [37]. It was previously shown that the concentration of testosterone in patients with T2D was negatively associated with the abundance of Lachnospirales and Firmicutes [38]. We have shown that the higher abundance of genus Frisingicoccus (phylum Firmicutes, class Clostridia, order Lachnospirales, family Lachnospiraceae) was associated with lower plasma testosterone concentrations in male recipients. In contrast, Lachnospiraceae on the level of family positively correlated with plasma testosterone concentrations. These results suggest the importance of gut microbiota composition investigation at all species levels concerning sex steroids. Cross-sex FMT in male recipients can promote a decrease in plasma testosterone concentrations. The commensal microbiota community can potentially affect sex hormones produced by enzymatic activity in the intestine. It was shown that gut microbiota is involved in the deglucuronidation of dihydrotestosterone, and testosterone resulting in high concentrations of androgens in the host organism [39,40]. In the current study, a specific unidentified ASV_314 was detected, which was also identified by another research team [40]. This unidentified bacterial ASV_314 abundance negatively correlated with plasma testosterone concentrations in males. All identified bacterial species (Bacteroides, Lacchnospiraceae, Frisingicoccus) were associated with plasma testosterone concentrations just in male same-sex and cross-sex recipients, with clear clustering of animals according to FMT. The clustering, and association of bacterial species involved in the testosterone changes were not significant in female recipients (same-sex or cross-sex FMT recipients). These findings point out the sex difference in the ability and effectiveness of FMT between female and male recipient rats. The decline in abundance of Bacteroides, Prevotella, Desulfovibrio, Lactobacillus, and Oxalobacter genera diversity in the gut is related to the higher prevalence of chronic metabolic disorders. In both lean and obese individuals, gut dysbiosis and low richness in gut microbial abundance are associated with higher adiposity, insulin resistance, dyslipidemia, and low-grade inflammation [41]. Gender differences in gut microbiome diversity have been reported with a higher abundance of Bacteroides and Prevotella genera in male than female individuals [42]. Androgens regulate the sex-specific gut dysbiosis in obese female and male mice, resulting in a decline in glucose metabolism driven by Prevotella sp. Bacteroides massiliensis and Cupriavidus metallidurans [21]. In addition, androgen-deficient obese mice showed sex-dependent development of metabolic dysfunction linked with enriched gut diversity with Turicibacter and Lactobacillus reuteri that caused early death in male mice [43]. Moreover, estrogen treatment in high-fat diet-fed mice showed a modest impact on gut microbiome diversity, resulting in an increased abundance of Collinsella aerofaciens F in males [44]. Thus, the gut microbiota composition might be partially responsible for sex differences in sex hormones and in the prevalence and severity of metabolic diseases.
4. Materials and Methods
4.1. Animals
Female (n=19) and male (n=21) Wistar rats at the age of 10 months were purchased from Velaz (Charles River, Velaz, Prague, Czech Republic). All animals were maintained under the standard conditions (temperature 25±2 °C and humidity 55±10 %) with a 12:12 hours light-dark cycle, and ad libitum access to standard chow (KMK20, EYPY, Czech Republic) and water. Rats were group-housed (2-3 rats per cage) in polycarbonate cages (open caging system, 50×36×19 cm) with bedding (Safe Select Fine, Velaz, Prague, Czech Republic) and polycarbonate enrichment for rats - tunnels (Velaz, Prague, Czech Republic). All experimental procedures were approved by the Ethical Committee of the Institute of Molecular Biomedicine, Comenius University, Bratislava, and have been conducted following the EU Directive 2010/63/EU and Slovak legislation.
4.2. Experimental Groups
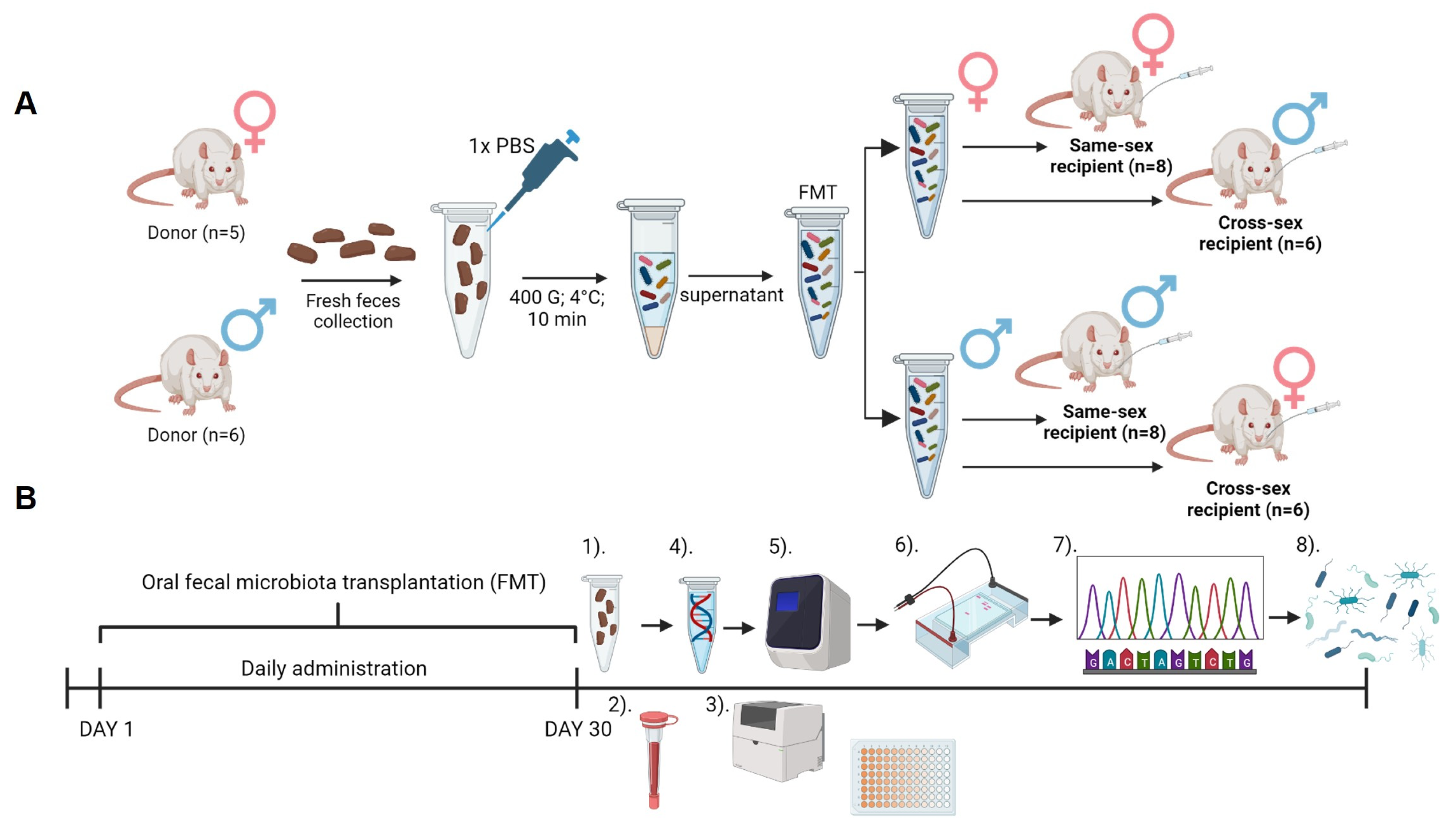
Animals were divided into experimental groups according to a received vehicle: PBS (female donor: n=5; male donor: n=6), FMT from same-sex (female same-sex recipient: n=8; male same-sex recipient: n=8), and FMT from the opposite sex (females cross-sex recipient: n=6; males cross-sex recipient: n=7). Animals from the same experimental group were housed together. Body weight and food consumption were measured weekly (Figure 4A).
4.3. Fecal Microbiota Transplantation and Recipient Sample Collection
For the preparation of the FMT solution, freshly collected feces were used. Fresh fecal boluses were placed into 15 ml falcon tubes under sterile conditions from every donor animal separately. Feces samples were weighed and 1,5 g from every bolus was pooled. Pooled feces were dissolved in 1X PBS and immediately centrifuged (400 g, 4 °C, 10 min). The supernatant was collected and used as FMT. For the administration of FMT, oral gavage was used in a volume of 100 ul per animal [45,46]. Animals received FMT daily for 30 days (Fig.1.A). Feces samples from recipients were collected individually at the end of the experiment under sterile conditions and immediately frozen at -80 °C (Figure 4B)
4.4. Blood Analysis
Following a 30-day-long FMT, blood collection was performed. Before blood collection animals were fasted overnight. Fasting glycemia was measured using a glucometer (Accu Chek Performa, Roche Slovakia, s.r.o Diabetes Care, Bratislava, Slovakia). Blood was collected from the tail vein to the EDTA collection tube (K3 EDTA, Microvette, Sarstedt, Nümbrecht, Germany). Blood was centrifuged for 10 min at 1600 g and 4°C and plasma supernatant was stored at -20°C until further analysis. Besides measuring the concentration of fasting insulin (Rat Insulin ELISA kit, Mercodia, Uppsala, Sweden), plasma samples were used to measure circulating concentrations of triglycerides (TAG), total cholesterol (CHOL), high-density lipoproteins (HDL), low-density lipoproteins (LDL), aspartate aminotransferase (AST), and alanine aminotransferase (ALT) using Biolis 24i Premium analyzer (Tokyo Boeki Machinery, Tokyo, Japan). Plasmatic testosterone was measured using the ELISA method (DRG Diagnostic, Marburg, Germany) (Figure 4B). From fasting glycemia and fasting insulin concentrations, the quantitative index of insulin sensitivity (QUICKI) was calculated [47].
4.5. Microbiome Analysis
Bacterial DNA was extracted using a commercial kit (QIAamp Fast DNA Stool Mini kit, Qiagen, Hilden, Germany) following the manufacturer's instructions. The DNA quality and quantity control was verified by spectrophotometric determination (Qubit dsDNA HS Assay Kit, Thermo Fisher Scientific Inc.). Variable V4 region of the 16S rRNA gene was amplified with the modified 515F forward primer 5´ GTGYCAGCMGCCGCGGTAA 3´, and the 806R reverse primer 5´GGACTACNVGGGTWTCTAAT 3´ [48,49], also used in the protocol for paired-end 16S rRNA sequencing on the Illumina platform by the Earth´s Microbiome Project [50]. The PCR mixture included the following: 10 µl of Ruby Taq Master Mix polymerase 2x (Jena Bioscience), 5 µl of extracted bacterial DNA, and 0,5 µl of each primer (200-400 nM). The total reaction volume was 20 ul. The PCR conditions were: 95 °C for an initial 5 min, followed by 28 cycles of 95 °C for 30 sec, 50 °C for 45 sec, and 72 °C for 60 sec. The quality and quantity of amplified DNA were analyzed using 1,5% gel electrophoresis with 2µl of Gel Loading Dye (New England BioLabs, Inc.) and 100 bp DNA Ladder (New England BioLabs, Inc.) (Fig.4.B). The samples were pair-end sequenced on the MiniSeq Sequencing System of the Illumina platform (ID MN01227) at the Research Resources Center (MC937), University of Illinois in Chicago, IL. Sequenced raw reads could be found with the project accession PRJEB70472 in ENA.
4.6. Data Processing and Results Analysis
The quality of paired-end Illumina sequences was checked using FastQC v.0.11.9 [51] and MultiQC v.1.13 [52]. Primer sequences were removed by Cutadapt v3.5100 [53] using the linked behavior, an overlap of 10 nucleotides, removing reads of length zero, and discarding untrimmed sequences. Afterwards, the sequence processing was continued using DADA2 v1.22.0101 [54] which is the core of the pipeline. This package is used for trimming sequence ends to remove low-quality ends, as well as, for ASVs generation and chimera removal. Sequence quality profiles were inspected again using FastQC v.0.11.9 [51] and MultiQC v.1.13 [52], and quality trimming was carried out accordingly. The ‘truncLen’ parameter was set at 132 bp for the R1 and R2 files. Filter out of low-quality R1 and R2 reads was performed using a max expected error of 2 and 5 respectively, with the rest of the default parameters. Paired-end reads were merged in order to reconstruct the amplicon sequence variants (ASVs) using the DADA2 core sample inference algorithm with the pool inference behavior. Subsequently, chimeric sequences were removed using the consensus method of the DADA2 package. Finally, 16S rDNA V4 gene sequences were classified by the IDTAXA classifier of the DECIPHER v2.22108 [55] package with a 50% confidence using SILVA SSU r138 (modified) database implemented in the IDTAXA classifier (Figure 4B).
4.7. Statistical Analysis
For the statistical evaluation and visualization of body weight gain, food consumption, and biochemical parameters, GraphPad Prism version 8.0.1 (GraphPad Software, Inc., CA, USA) was used. Two-way ANOVA was performed with independent factors: sex and fecal microbiota transplantation (FMT) with Bofenroni post-hoc test. For the statistical evaluation of microbiome data, RStudio version 4.2.1 served as the primary software tool. Initially, sequences with less than one count were removed and data normalized. Shannon index, principal component analysis (PCA), and redundancy analysis (RDA) were obtained using vegan, and ggplot2 libraries. RDA was done with a 1000 permutations test. Spearman correlations of the bacterial species with testosterone concentrations were analyzed in the JASP software, and visualized in GraphPad Prism 8.0.1 (GraphPad Software, Inc., CA, USA).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Redundancy analysis of the gut microbiota - all recipients.
Author Contributions
Conceptualization, A.F. and P.B.; methodology, P.C.; formal analysis, V.B.; investigation, M.D and V.B..; data curation, D.P.S.; writing—original draft preparation, A.F. and P.B.; writing—review and editing, R.G.; supervision, P.C.; project administration, P.C. and R.G.; funding acquisition, M.D. and R.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Slovak Research and Development Agency under the contract no. APVV-21-0370 and by the Ministry of Education, Science, Research and Sport of the Slovak Republic under the contract no. VEGA 1/0649/21 and VEGA 1/0398/24, and by the grants CDEIGENT/2021/008 and CIACIF/2022/070 from Generalitat Valenciana.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee of Institute of Molecular Biomedicine (protocol code 4/10/2021 approved on 4th October 2021).
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Apovian, C.M. Obesity: definition, comorbidities, causes, and burden. Am. J. Manag. Care 2016, 22, s176–185. [Google Scholar] [PubMed]
- Jaacks, L.M.; Siegel, K.R.; Gujral, U.P.; Narayan, K.M.V. Type 2 diabetes: A 21st century epidemic. Best. Pract. Res. Cl. En. 2016, 30, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Sex differences in metabolic homeostasis, diabetes, and obesity. Biol. Sex. Differ. 2015, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F. Sex differences in energy metabolism: natural selection, mechanisms and consequences. Nat. Rev. Nephrol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Varlamov, O.; Bethea, C.L.; Roberts, C.T., Jr. Sex-specific differences in lipid and glucose metabolism. Front. Endocrinol. (Lausanne) 2014, 5, 241. [Google Scholar] [CrossRef] [PubMed]
- Hur, K.Y.; Lee, M.S. Gut Microbiota and Metabolic Disorders. Diabetes Metab. J. 2015, 39, 198–203. [Google Scholar] [CrossRef]
- Bredella, M.A. Sex Differences in Body Composition. Adv. Exp. Med. Biol. 2017, 1043, 9–27. [Google Scholar] [CrossRef]
- He, X.; Li, Z.; Tang, X.; Zhang, L.; Wang, L.; He, Y.; Jin, T.; Yuan, D. Age- and sex-related differences in body composition in healthy subjects aged 18 to 82 years. Medicine 2018, 97, e11152. [Google Scholar] [CrossRef]
- Wells, J.C.K. Sexual dimorphism of body composition. Best Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 415–430. [Google Scholar] [CrossRef]
- Santos-Marcos, J.A.; Mora-Ortiz, M.; Tena-Sempere, M.; Lopez-Miranda, J.; Camargo, A. Interaction between gut microbiota and sex hormones and their relation to sexual dimorphism in metabolic diseases. Biology of Sex. Differences 2023, 14. [Google Scholar]
- Lovejoy, J.C.; Champagne, C.M.; de Jonge, L.; Xie, H.; Smith, S.R. Increased visceral fat and decreased energy expenditure during the menopausal transition. Int. J. Obes. 2008, 32, 949–958. [Google Scholar] [CrossRef] [PubMed]
- van Londen, G.J.; Levy, M.E.; Perera, S.; Nelson, J.B.; Greenspan, S.L. Body composition changes during androgen deprivation therapy for prostate cancer: A 2-year prospective study. Crit. Rev. Oncol. Hemat 2008, 68, 172–177. [Google Scholar] [CrossRef]
- Allan, C.A.; Strauss, B.J.; Burger, H.G.; Forbes, E.A.; McLachlan, R.I. Testosterone therapy prevents gain in visceral adipose tissue and loss of skeletal muscle in nonobese aging men. J. Clin. Endocrinol. Metab. 2008, 93, 139–146. [Google Scholar] [CrossRef]
- Chi, J.T.; Lin, P.H.; Tolstikov, V.; Oyekunle, T.; Chen, E.Y.; Bussberg, V.; Greenwood, B.; Sarangarajan, R.; Narain, N.R.; Kiebish, M.A.; Freedland, S.J. Metabolomic effects of androgen deprivation therapy treatment for prostate cancer. Cancer Med. 2020, 9, 3691–3702. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, M.; van der Poorten, D. The gut microbiome. Aust. Fam. Physician 2017, 46, 206–211. [Google Scholar] [PubMed]
- Heintz-Buschart, A.; Wilmes, P. Human Gut Microbiome: Function Matters. Trends Microbiol. 2018, 26, 563–574. [Google Scholar] [CrossRef]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Gao, A.B.; Su, J.L.; Liu, R.X.; Zhao, S.Q.; Li, W.; Xu, X.Q.; Li, D.J.; Shi, J.; Gu, B.; Zhang, J.; et al. Sexual dimorphism in glucose metabolism is shaped by androgen-driven gut microbiome. Nat. Commun. 2021, 12. [Google Scholar]
- Shobeiri, P.; Kalantari, A.; Teixeira, A.L.; Rezaei, N. Shedding light on biological sex differences and microbiota-gut-brain axis: a comprehensive review of its roles in neuropsychiatric disorders. Biol. Sex. Differ. 2022, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ko, Y.; Kwak, C.; Yim, E.S. Gender differences in metabolic syndrome components among the Korean 66-year-old population with metabolic syndrome. Bmc Geriatr. 2016, 16. [Google Scholar]
- Wang, W.Y.; Li, C.H.; Wu, Y.S.; Chien, W.C.; Wang, K.Y.; Tzeng, W.C. Gender Differences in the Prevalence of Metabolic Syndrome Among Taiwanese Air Force Personnel A Population-Based Study. J. Cardiovasc. Nurs. 2020, 35, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Chen, Z.P.; Baudier, R.L.; Krousel-Wood, M.; Anderson, A.H.; Fonseca, V.A.; Mauvais-Jarvis, F. Sex Differences in the Progression of Metabolic Risk Factors in Diabetes Development. Jama Netw. Open 2022, 5. [Google Scholar]
- Ramezankhani, A.; Azizi, F.; Hadaegh, F. Gender differences in changes in metabolic syndrome status and its components and risk of cardiovascular disease: a longitudinal cohort study. Cardiovasc. Diabetol. 2022, 21. [Google Scholar]
- Cheng, H.Y.; Ning, M.X.; Chen, D.K.; Ma, W.T. Interactions Between the Gut Microbiota and the Host Innate Immune Response Against Pathogens. Front. Immunol. 2019, 10. [Google Scholar]
- Aggeletopoulou, I.; Konstantakis, C.; Assimakopoulos, S.F.; Triantos, C. The role of the gut microbiota in the treatment of inflammatory bowel diseases. Microb. Pathog. 2019, 137, 103774. [Google Scholar] [CrossRef] [PubMed]
- Haak, B.W.; Prescott, H.C.; Wiersinga, W.J. Therapeutic Potential of the Gut Microbiota in the Prevention and Treatment of Sepsis. Front. Immunol. 2018, 9, 2042. [Google Scholar] [CrossRef]
- Fong, W.N.; Li, Q.; Yu, J. Gut microbiota modulation: a novel strategy for prevention and treatment of colorectal cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Aggarwal, P.; Costa, R.G.E.; Cole, A.M.; Trinchieri, G. Targeting the gut microbiota for cancer therapy. Nat. Rev. Cancer 2022, 22, 703–722. [Google Scholar] [CrossRef] [PubMed]
- Strack, C.; Behrens, G.; Sag, S.; Mohr, M.; Zeller, J.; Lahmann, C.; Hubauer, U.; Loew, T.; Maier, L.; Fischer, M.; Baessler, A. Gender differences in cardiometabolic health and disease in a cross-sectional observational obesity study. Biol. Sex. Differ. 2022, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Valeri, F.; Endres, K. How biological sex of the host shapes its gut microbiota. Front. Neuroendocrin 2021, 61. [Google Scholar]
- Diviccaro, S.; Giatti, S.; Borgo, F.; Falvo, E.; Caruso, D.; Garcia-Segura, L.M.; Melcangi, R.C. Steroidogenic machinery in the adult rat colon. J. Steroid Biochem. 2020, 203. [Google Scholar]
- Org, E.; Mehrabian, M.; Parks, B.W.; Shipkova, P.; Liu, X.Q.; Drake, T.A.; Lusis, A.J. Sex differences and hormonal effects on gut microbiota composition in mice. Gut Microbes 2016, 7, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Fujita, K.; Motooka, D.; Hatano, K.; Hata, J.; Nishimoto, M.; Banno, E.; Takezawa, K.; Fukuhara, S.; Kiuchi, H.; et al. Firmicutes in Gut Microbiota Correlate with Blood Testosterone Levels in Elderly Men. World J. Mens. Health 2022, 40, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Kaliannan, K.; Robertson, R.C.; Murphy, K.; Stanton, C.; Kang, C.; Wang, B.; Hao, L.; Bhan, A.K.; Kang, J.X. Estrogen-mediated gut microbiome alterations influence sexual dimorphism in metabolic syndrome in mice. Microbiome 2018, 6, 205. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Cao, R.Y.; Liu, L.A.; Lv, Y.Y.; Qi, X.Y.; Yuan, Z.S.; Fan, X.D.; Yu, C.X.; Guan, Q.B. Correlation Between Gut Microbiota and Testosterone in Male Patients With Type 2 Diabetes Mellitus. Front. Endocrinol. 2022, 13. [Google Scholar]
- Colldén, H.; Landin, A.; Wallenius, V.; Elebring, E.; Fändriks, L.; Nilsson, M.E.; Ryberg, H.; Poutanen, M.; Sjögren, K.; Vandenput, L.; Ohlsson, C. The gut microbiota is a major regulator of androgen metabolism in intestinal contents. Am. J. Physiol. -Endocrinol. Metab. 2019, 317, E1182–E1192. [Google Scholar] [CrossRef] [PubMed]
- EU622779.1, G. Uncultured bacterium clone H-243(3) 16S ribosomal RNA gene, complete sequence. Available online: (accessed on.
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Mueller, S.; Saunier, K.; Hanisch, C.; Norin, E.; Alm, L.; Midtvedt, T.; Cresci, A.; Silvi, S.; Orpianesi, C.; Verdenelli, M.C.; et al. Differences in fecal microbiota in different European study populations in relation to age, gender, and country: a cross-sectional study. Appl. Environ. Microbiol. 2006, 72, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Hanada, K.; Minami, Y.; Kitakaze, T.; Ogata, Y.; Tokumoto, H.; Sato, T.; Kato, S.; Inui, H.; Yamaji, R. Role of gut microbiota in sex- and diet-dependent metabolic disorders that lead to early mortality of androgen receptor-deficient male mice. Am. J. Physiol-Endoc M. 2020, 318, E525–E537. [Google Scholar] [CrossRef] [PubMed]
- Hases, L.; Stepanauskaite, L.; Birgersson, M.; Brusselaers, N.; Schuppe-Koistinen, I.; Archer, A.; Engstrand, L.; Williams, C. High-fat diet and estrogen modulate the gut microbiota in a sex-dependent manner in mice. Commun. Biol. 2023, 6, 20. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.J.; Guo, Y.; Zhang, S.R.; Chen, Z.H.; Wu, K.Q.; Liu, Q.; Liu, K.J.; Wen, L.Z.; Wei, Y.L.; Wang, B.; Chen, D.F. Fecal Microbiota Transplantation Can Alleviate Gastrointestinal Transit in Rats with High-Fat Diet-Induced Obesity via Regulation of Serotonin Biosynthesis. Biomed. Res. Int. 2018, 2018. [Google Scholar]
- Di Luccia, B.; Crescenzo, R.; Mazzoli, A.; Cigliano, L.; Venditti, P.; Walser, J.C.; Widmer, A.; Baccigalupi, L.; Ricca, E.; Iossa, S. Rescue of Fructose-Induced Metabolic Syndrome by Antibiotics or Faecal Transplantation in a Rat Model of Obesity. PLoS ONE 2015, 10, e0134893. [Google Scholar] [CrossRef]
- Chen, H.; Sullivan, G.; Yue, L.Q.; Katz, A.; Quon, M.J. QUICKI is a useful index of insulin sensitivity in subjects with hypertension. Am. J. Physiol-Endoc M. 2003, 284, E804–E812. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef]
- Gilbert, J.A.; Jansson, J.K.; Knight, R. The Earth Microbiome project: successes and aspirations. Bmc Biol. 2014, 12. [Google Scholar]
- Andrews. FastQC: a quality control tool for high throughput sequence data. Available online: (accessed on.
- Ewels, P.; Magnusson, M.; Lundin, S.; Kaller, M. MultiQC: summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. 2011 2011, 17, 3. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Wright. Using DECIPHER v2.0 to Analyze Big Biological Sequence Data in R. The R. J. 2016, 8, 352. [Google Scholar] [CrossRef]
Figure 1.
Body weight change in female (A), and male (B) donor and recipient animals, and food consumption in female (C), and male (D) donor and recipient animals. Data are presented as mean ±SEM. *- differences between same-sex recipient female rats and donors; +- differences between cross-sex female rats and donors; *, P< 0.05; ** , P< 0.01; ***, P< 0.001; +, P< 0.05; ++ , P<0.01; +++, P< 0.001.
Figure 1.
Body weight change in female (A), and male (B) donor and recipient animals, and food consumption in female (C), and male (D) donor and recipient animals. Data are presented as mean ±SEM. *- differences between same-sex recipient female rats and donors; +- differences between cross-sex female rats and donors; *, P< 0.05; ** , P< 0.01; ***, P< 0.001; +, P< 0.05; ++ , P<0.01; +++, P< 0.001.
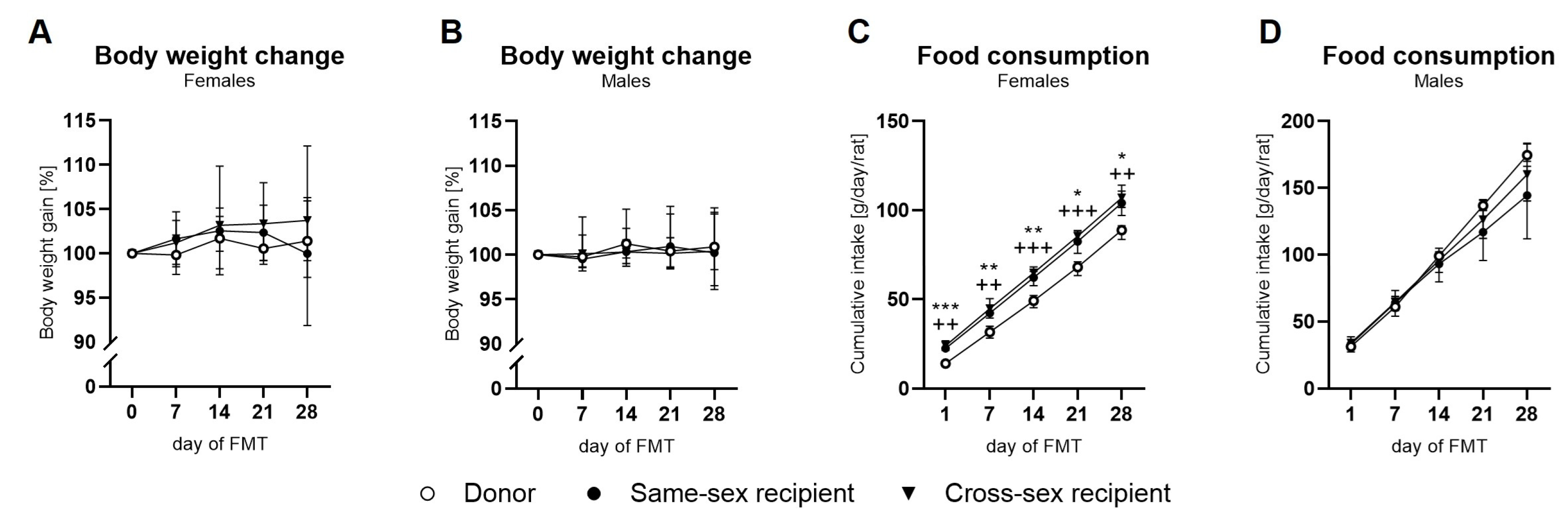
Figure 2.
Biochemical analysis of plasma: (A) Body weight; (B) Fasting glycemia; (C) Fasting insulin concentrations; (D) Quantitative index of insulin sensitivity (QUICKI); (E) Total cholesterol concentration; (F) Concentration of triacylglycerols; (G) HDL cholesterol concentrations; (H) LDL cholesterol concentrations; (I) AST concentration; (J) ALT concentration; (K) uric acid concentration. Data are presented as mean ± SEM. *-sex difference; #- the difference between donor, same-sex, and cross-sex rats in individual sex groups; *, P< 0.05; ***, P<0.001; #, P< 0.05; HDL- high-density lipoproteins, LDL- low-density lipoproteins, AST- aspartate aminotransferase, ALT- alanine aminotransferase.
Figure 2.
Biochemical analysis of plasma: (A) Body weight; (B) Fasting glycemia; (C) Fasting insulin concentrations; (D) Quantitative index of insulin sensitivity (QUICKI); (E) Total cholesterol concentration; (F) Concentration of triacylglycerols; (G) HDL cholesterol concentrations; (H) LDL cholesterol concentrations; (I) AST concentration; (J) ALT concentration; (K) uric acid concentration. Data are presented as mean ± SEM. *-sex difference; #- the difference between donor, same-sex, and cross-sex rats in individual sex groups; *, P< 0.05; ***, P<0.001; #, P< 0.05; HDL- high-density lipoproteins, LDL- low-density lipoproteins, AST- aspartate aminotransferase, ALT- alanine aminotransferase.
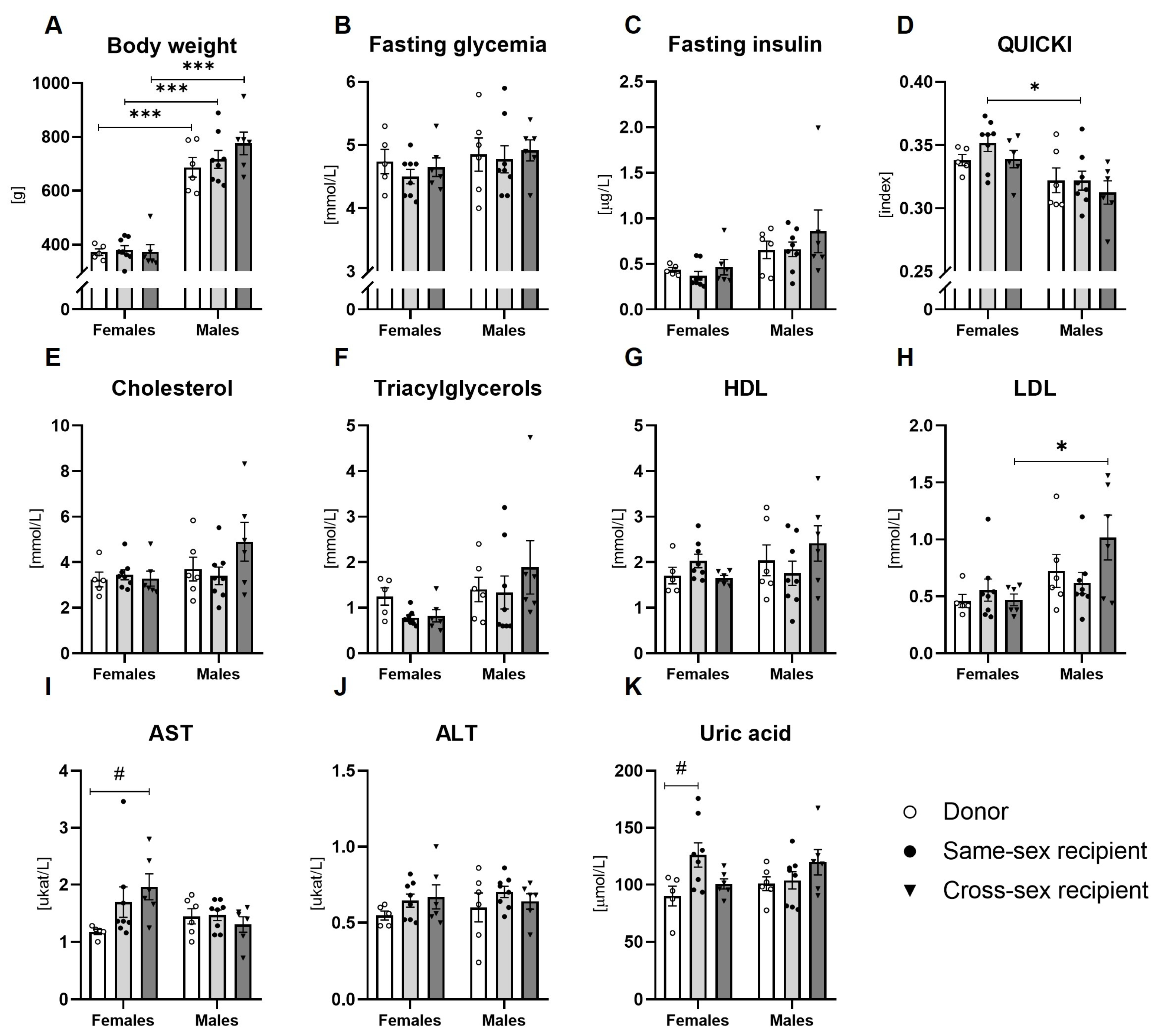
Figure 3.
Plasma testosterone concentrations and gut microbiota composition analysis: (A) Plasma testosterone concentrations; (B) Principal component analysis of the gut microbiota - all recipients; (C)- Redundancy analysis of all female recipients according to different FMT donor sex (same-sex females, cross-sex females); (D) Redundancy analysis of all male recipients according to different FMT donor sex (same-sex males, cross-sex males); (E) Redundancy analysis of all mice receiving FMT from female donors (same-sex females, cross-sex males); (F) Redundancy analysis of all mice receiving FMT from male donors (same-sex males, cross-sex females); (G) Correlation between plasma testosterone concentration and Bacteroides abundance in recipient males; (H) Correlation between plasma testosterone concentration and Frisingicoccus abundance in recipient males; (I) Correlation between plasma testosterone concentration and Lachnospiraceae abundance in recipient males; (J) Correlation between plasma testosterone concentrations and Bacteria N/A (ASV_314) in recipient males; (K) Correlation between plasma testosterone concentration and Bacteroides abundance in recipient females; (L) Correlation between plasma testosterone concentration and Frisingicoccus abundance in recipient females; (M) Correlation between plasma testosterone concentration and Lachnospiraceae abundance in recipient females; (N) Correlation between plasma testosterone concentrations and Bacteria N/A (ASV_314) in recipient females. Data are presented as mean ± SEM. *-sex difference; #- the difference between donor, same-sex, and cross-sex rats in individual sex groups; **, P< 0.01; ***, P<0.001; ##, P< 0.01.
Figure 3.
Plasma testosterone concentrations and gut microbiota composition analysis: (A) Plasma testosterone concentrations; (B) Principal component analysis of the gut microbiota - all recipients; (C)- Redundancy analysis of all female recipients according to different FMT donor sex (same-sex females, cross-sex females); (D) Redundancy analysis of all male recipients according to different FMT donor sex (same-sex males, cross-sex males); (E) Redundancy analysis of all mice receiving FMT from female donors (same-sex females, cross-sex males); (F) Redundancy analysis of all mice receiving FMT from male donors (same-sex males, cross-sex females); (G) Correlation between plasma testosterone concentration and Bacteroides abundance in recipient males; (H) Correlation between plasma testosterone concentration and Frisingicoccus abundance in recipient males; (I) Correlation between plasma testosterone concentration and Lachnospiraceae abundance in recipient males; (J) Correlation between plasma testosterone concentrations and Bacteria N/A (ASV_314) in recipient males; (K) Correlation between plasma testosterone concentration and Bacteroides abundance in recipient females; (L) Correlation between plasma testosterone concentration and Frisingicoccus abundance in recipient females; (M) Correlation between plasma testosterone concentration and Lachnospiraceae abundance in recipient females; (N) Correlation between plasma testosterone concentrations and Bacteria N/A (ASV_314) in recipient females. Data are presented as mean ± SEM. *-sex difference; #- the difference between donor, same-sex, and cross-sex rats in individual sex groups; **, P< 0.01; ***, P<0.001; ##, P< 0.01.
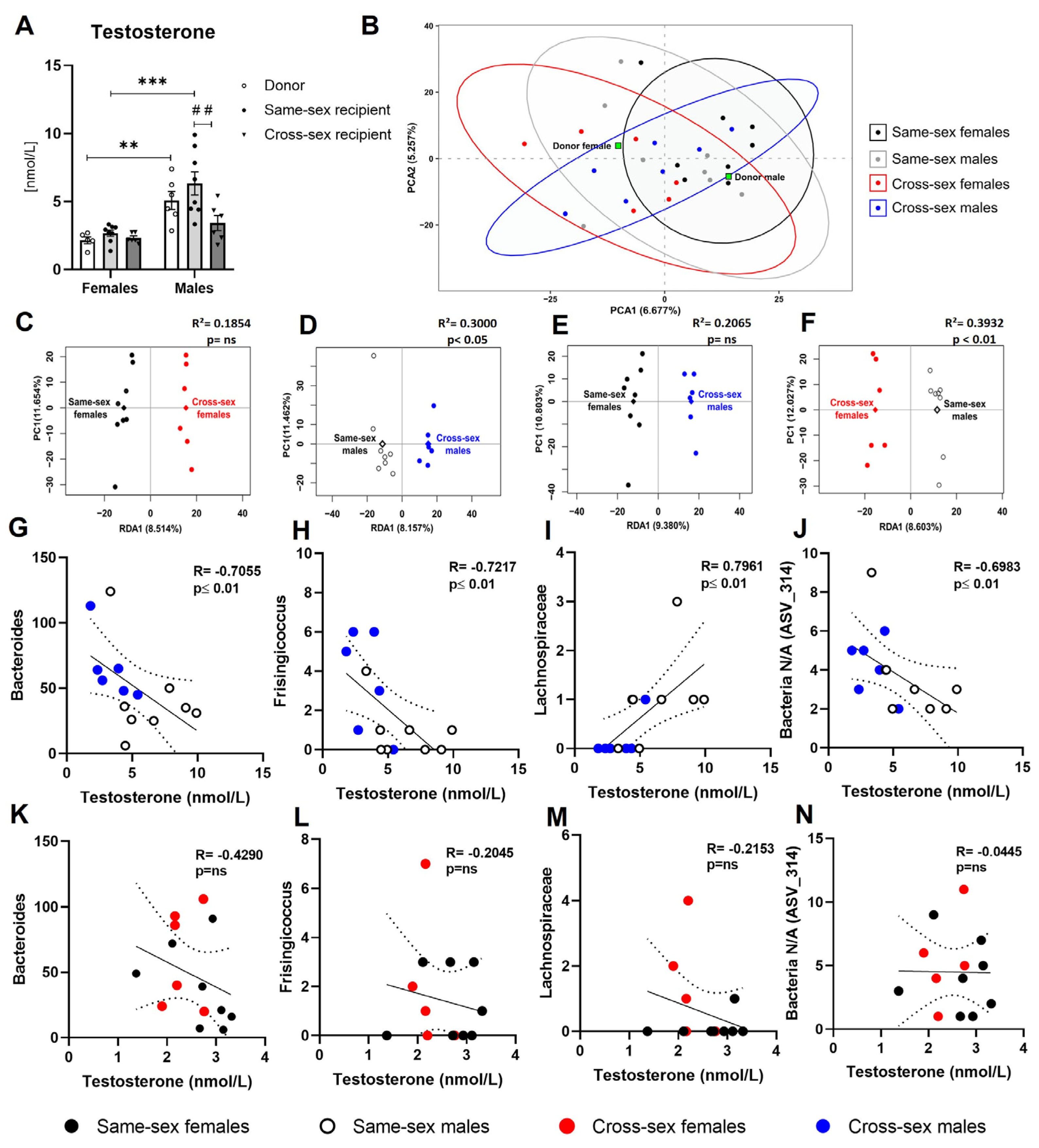
Figure 4.
Experimental design: (A)- Preparation of fecal microbiota transplantation (FMT) and experimental groups. (B)- Timeline of the experiment- 1)- Recipient sample collection; 2)- Blood collection from the tail vein and plasma preparation; 3)- Biochemical analysis of plasma samples; 4)- Isolation of bacterial DNA from stool samples; 5)- DNA analysis and quantity control, and amplification of 16S rRNA V4 regions; 6)- Endpoint Ruby Taq electrophoresis; 7) Sample sequencing using MiSeq instrument; 8)- Data processing. Created with BioRender.com.
Figure 4.
Experimental design: (A)- Preparation of fecal microbiota transplantation (FMT) and experimental groups. (B)- Timeline of the experiment- 1)- Recipient sample collection; 2)- Blood collection from the tail vein and plasma preparation; 3)- Biochemical analysis of plasma samples; 4)- Isolation of bacterial DNA from stool samples; 5)- DNA analysis and quantity control, and amplification of 16S rRNA V4 regions; 6)- Endpoint Ruby Taq electrophoresis; 7) Sample sequencing using MiSeq instrument; 8)- Data processing. Created with BioRender.com.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



