
Submitted:
28 November 2023
Posted:
29 November 2023
You are already at the latest version
Abstract
Shrubs are a significant component of urban vegetation residing in parks, but they experience various influences of biotic and abiotic agents, among which aphids play an important role. In this work, the effect of aphid colonization on three shrub species in urban environment was examined. Fourteen parameters have been analyzed, describing photosynthetic pigment content and gas exchange to carbon, nitrogen, and hydrogen partition. While no significant effect of colonization was found on photosynthetic pigment parameters, the effect was significant on gas exchange parameters. The strongest effect aphid colonization achieved on nitrogen partition and C/N ratio, parameters that could be suggested in further similar studies. All parameters were classified in two groups according to principal component analysis, suggesting correlation between nitrogen and carbon content, C/N ratio, measured gas exchange parameters and chlorophyll a content. The ratio between net photosynthesis and dark respiration (A/K) was in the second group, suggesting that this parameter provide additional information on the effect of aphid colonization, and deserve special attention in further studies. There were differences in reaction of examined shrub species, where the strongest reaction by gross primary productivity (GPP) achieved Hybiscus syriacus, and by nitrogen partition and C/N ratio Spirea trilobata and Cydonia japonica.
Keywords:
aphids
; shrubs
; gas exchange
; photosynthetic pigments
; CHN partition
1. Introduction
Vegetation plays an important role in urban environments. It consists of plants inhabiting green spaces such as parks, forests, private residential lawns, and other open areas [1]. These areas are among the primary supporters of urban region's sustainability, people’s wellness, and urban life quality, considered to provide social, ecological, and economic benefits [1,2]. The increased attention directed to green infrastructure (i.e., the employment of plants in cities) arises from the numerous ecosystem services they provide, which support and enrich human life [3,4]. The numerous supported or enabled vegetation-based benefits include the production of food, regulation of noise and air pollution, the urban heat island effect reduction, control of biogeochemical cycling, flood mitigation, biodiversity maintenance, rendering the rest and recreation of city residents (beneficial for physical health, socialization, stress level), as well as increasing of the metropolitan property values [1,5].
Shrubs are a significant component of urban vegetation residing in parks, urban wooded settings, gardens, scrubland, and hedges, alongside railway or road infrastructure [5]. Ornamental shrubs Hibiscus syriacus. Cydonia japonica, and Spiraea x vanhouttei are common members of urban flora in Serbia. Along with other plants of urban ecosystem they experience various changes due to biotic and natural influences [6]. Speaking of the first, these shrubs were recognized as host plants for various aphid species during previous investigations [7,8,9]. Aphids (Hemiptera, Aphididae) are important worldwide pests of more than 400 of either cultivated or self-seeding plant species, including ornamental plants [10]. According to a recent report, 374 species were found within the territory of Serbia [9]. Aphids negatively affect their host plants in several ways [11]. Being phloem feeders, they penetrate sieve elements using their stylets to ingest sugars, nitrogen compounds and other nutrients necessary also to plant growth and reproduction. Furthermore, their saliva injected during the feeding phase could exhibit a phytotoxic effect. Along with direct injuries, aphids also transmit numerous plant viruses [12]. Aphids’ infestation of ornamental plants affects vigor and aesthetic appearance, due to promoted curling, distortion, and chlorosis of leaves, hardening of the buds, and malformations and discoloration (fading, stains, necrosis) of various organs [12,13]. Finally, honeydew deposits covering the leaf cuticle are frequently occupied by sooty molds (black filamentous saprophytic ascomycetes) and may hinder the photosynthetic activity of leaves [11].
The contribution of urban shrubs in the improvement of environmental quality and human health is enabled by their ability to decrease levels of air pollution and temperature, reduce energy consumption in buildings, intercept water and reduce surface run-off, reduce the atmosphere’s carbon dioxide levels (by sequestering of carbon, leading to climate change mitigation) [6]. Furthermore, shrubs provide food and shelter for animals [14] and therefore contribute to the preservation and increase of urban biodiversity. Considering the living habits of aphids infesting urban shrubs, and possible negative consequences, it seems necessary to recognize and quantify changes in plant attributes underlying plant vigor and fitness in (often inhospitable) urban environments. The adverse effects on plant growth, metabolism, mineral nutrition, photosynthetic rate, concentration of chlorophyll, and gas exchange parameters [15,16,17,18,19,20,21] have been reported in aphid-infested plants.
The influence of plant aphids on plant growth elements (diameter and height) has not been quantified in previous research, suggesting that the growth elements are not reliable enough to determine the degree of negative impact of the aphid diet on the host plant. Plant development components are also dependent on other environmental and internal variables that might operate as "concealment" of the aphid diet's harmful influence. Thus, it appears necessary to investigate and quantify the values of fundamental physiological parameters in colonized and uncolonized plant leaves. The aim of this work was to 1) identify aphid species and characterize their colonies present in the frequently cultivated ornamental shrubs species in urban areas of Serbia, i.e., Hibiscus syriacus , Cydonia japonica and Spirea x vanhouttei, and 2) measure and compare rates of photosynthesis and respiration, and contents of photosynthetic pigments, nitrogen, carbon, and hydrogen in healthy and infested leaves. Due to the abundance and distribution of species they belong to, the number of species that colonize, complex development and lifestyle, specific morphological and anatomical features, as well as the consequences of aphid’s nutrition on the host plants, these results are valuable for entomologists and plant production experts.
2. Materials and Methods
2.1. Plant Material
Aiming to compare the physiological characteristics of aphid-infested and control (non-infested) leaves, samples of Spirea trilobata, Hibiscus syriacus, and Cydonia japonica were collected from the urban environment (campus of University of Novi Sad; 45°14'43"N 19°51'05" E). Three samples of five fully developed leaves were taken from the tip of shoots of three colonized and three non-colonized mature plants about 10 years old of each examined host species. Hibiscus syriacus was colonized by Aphis gossypii, Cydonia japonica by Aphis spiraecola, and Spiraea x vanhouttei by Aphis spiraecola. The level of infestation of all three examined species was high. All the shrubs that are being studied have shoots colonized on them, which is a trait that Aphis gossypii and Aphis spiraecola have in common. Prior to measurements and analyses, insects and their remains were gently removed from the leaf's surface. All analyses and measurements were performed in triplicate.
2.2. Leaf Photosynthetic Characteristics
The net photosynthesis (A) and dark respiration rates (K) were determined polarographically, using a Hansatech DW1 electrode (Hansatech Instruments Ltd, UK). The quantity of oxygen emitted (molO2 cm-2 h-1) indicated the level of photosynthesis, whereas the amount of oxygen absorbed (molO2cm-2h-1) determined the level of respiration. The leaf sections used for gas exchange measurements were cut from the middle of the leaf lamina of both control and infested leaves, avoiding the leaf nerves. The leaf samples were immersed in a pH 7.6 - 7.8 buffer solution containing 10 mM NaHCO3 [22]. From these, the following parameters were calculated: GPP, which is gross primary productivity (А+К); A/K – A/K ratio; A/GPP – A/GPP ratio. Following the measurement of gas-exchange parameters leaves from the same plants were used for the evaluation of photosynthetic pigments’ concentration. The concentration of chlorophyll a (Chla), chlorophyll b (Chlb), and carotenoids (Car) was measured spectrophotometrically (Beckman DU-65, CA), in absolute acetone extracts and expressed as mg g-1 dry weight. [23].
2.3. Analysis of Carbon, Nitrogen, and Hydrogen Partition
The elemental content of nitrogen (N), carbon (C), and hydrogen (H) in fully developed, oven-dried leaves of the tested species, both colonized and non-colonized, was measured using the CHN Vario EL III element analyzer (Elementar Analysensysteme GmbH, Germany). The elemental partition of the plant material was determined thermoconductometrically using a standard method.
2.4. Statistical Analysis
The data were processed by two-way factorial analysis of variance, with Species and Colonization as the main effects, based on which Tukey's Honest Significant Difference (HSD) test was performed at the significance level α=0.05. The relationship between the investigated parameters was described using the Pearson correlation coefficient, based on total means at the level of interaction Species × Colonization. This correlation matrix was the base for principal component analysis (PCA). The loadings of measured parameters with the first two principal components were used for agglomeration of measured parameters, where parameters that had their highest loading with the same principal component were regarded to be correlated and belong to the same group. Statistical analysis was performed using STATISTICA 13 software package [24].
3. Results
3.1. Analysis of Variance
According to the results of the analysis of variance (Table 1), there was no significant effect of factor Colonization nor interaction Species × Colonization on parameters describing the content of photosynthetic pigments. The only significant effect achieved factor Species on the variation of the content of chlorophyll a (Chla), the total content of chlorophyll a and b (Chla+b) and the content of carotenoids (Car). All parameters describing the partition of total nitrogen (N), carbon (C), and hydrogen (H), as well as the carbon/nitrogen ratio (C/N), were significantly dependent on factor Species. For these parameters, a significant effect of Colonization was found for all of them except for the partition of hydrogen (H), while the effect of interaction Species × Colonization was significant only for the partition of nitrogen (N) and carbon/nitrogen ratio (C/N).
The effects of factors Species and Colonization were significant for all examined physiological parameters, but the effect of interaction Species × Colonization was not significant for any of them.
According to the results of Tukey’s HSD test, there were no significant differences between colonized and non-colonized leaves in the content of photosynthetic pigments nor in parameters derived from them (Table 2). The lowest content of photosynthetic pigments was found in Cydonia japonica, and the highest in Hibiscus syriacus and Spirea trilobata. There were no significant differences between the examined species in the content of chlorophyll b (Chlb) and Chla/Chlb ratio.
All parameters describing the partition of nitrogen (N) or carbon (C) were significantly lower in colonized than in non-colonized leaves, while the difference in hydrogen partition was not significant. In total, the species differed significantly in all examined parameters describing the partition of N, C or H. However, at the level of interaction Species × Colonization, the reaction of examined species on aphid colonization was significant only in N and C/N. This suggests considerable sensitivity of these two parameters on the aphid colonization in examined shrub species.
All physiological parameters describing dark respiration and photosynthesis significantly differed between colonized and non-colonized leaves in total. In total, dark respiration (K), net photosynthesis (A) and their sum, gross primary productivity (GPP) were significantly lower in colonized leaves, while A/K and A/GPP ratios were significantly higher, suggesting higher reaction of leaves on colonization by dark respiration. In total, Hybiscus syriacus achieved the highest and Cydonia japonica the lowest values of dark respiration and net photosynthesis. However, only in Hybiscus syriacus it was found that dark respiration and GPP sum were significantly lower in colonized than in non-colonized leaves, while in other cases at the level of interaction Species × Colonization, the reaction on aphid colonization of examined species by examined physiological parameters was not statistically significant.
3.2. Principal Component Analysis
There were five principal components calculated by principal component analysis. Factor loadings between them and the original variables are presented in Table 3. Examined parameters (original variables) had their highest loadings with the first two principal components, which explained 86.4% of the total variance. Because the correlation between principal components is zero, it is assumed that original variables that have their highest loadings with the same principal component are also correlated between themselves. In this way, examined parameters are classified into two groups, according to the principal component with which they have their highest factor loading (Figure 1). The first group consists of Chla, Chla+b, N, C, C/N, K, A, GPP, and the second group of Chlb, Chla/b, Car, H, A/K and A/GPP. According to the results of Tukey’s test, the reaction of species by the parameters of the first group on the aphid colonization was significant in most cases, while the reaction by the parameters of the second group was not. However, in further studies, special attention should be paid to the second-group parameters because their low correlation with the first-group parameters suggests that they carry different information about the reaction of examined species on aphid colonization.
4. Discussion
The examined parameters are important indicators of the physiological condition or performance of plants and are part of the considerably complex process of reaction of plants on aphid colonization. All examined processes are mutually related and small changes of some of them can mitigate or halt the process of photosynthesis and growth.
Results obtained by the examination of photosynthesis and respiration showed some regularities in plant reaction to the attack of aphids. It was notified that examined aphid species change measured parameters. This is important because parameters by which plant reacts to aphid colonization could be used in further studies regarding differences in tolerance to the aphid attack between and within woody plant species.
In this study, the differences between Cydonia japonica and Hybiscus syriacus in Chla and Chla+b, as well as between Cydonia japonica and Spirea trilobata in Chla+b and Car were significant, but the reaction of examined species on aphid colonization by parameters describing the content of photosynthetic pigments both in total and within each of examined species was not. Thus, there were no general trends in the reaction of host plants on aphid colonization by the content of photosynthetic pigments. This is not in concordance with several studies conducted on wheat [25], Fabaceae species [26,27], sclerophyllous oak species (Quercus suber and Q. ilex) [27], and soybean [28], that, in general, suggest a decrease of the content of photosynthetic pigments in infested plants.
Also, according to [29], the ratio of contents of chlorophyll a and chlorophyll b varies between 3:1 and 5:1. In poplar varieties that ratio is higher in colonized than in non-colonized leaves [20]. In non-colonized leaves, it varied between 3.21 and 4.11, while in colonized leaves that ratio was higher and varied between 4.7 and 8.76. Chloroplast degradation is one of the most frequent reactions of plants experiencing aphid feeding, which is indicated by the number of chlorotic spots on leaves [30]. By injection of saliva in plant tissue and especially in conducting vessels, aphids can influence all parts of the plant. The findings of research by Cichocka et al. indicate that aphid saliva can cause significant losses in grain production in plants for up to decades [31].
Lack of significance in traits describing photosynthetic pigment content between colonized and non-colonized plants could be caused by deviations within treatments at the level of interaction Species × Colonization, resulting in relatively high residual variation and low precision in detecting significant differences between the treatments. This could be caused by the fact that this study was conducted in field trials, where the start of colonization was not known. However, it could also indicate the tolerance of examined plants to the pest, as described by [28] and [25]. Thus, it seems that traits describing the content of photosynthetic pigments and those derived from them are not suitable for precise analysis of the effects of aphid colonization in examined species, especially if the infestation is mild, or plants are tolerant to the aphid attack.
However, in total, there was a significant decrement in net photosynthesis, dark respiration, and gross primary productivity, as well as an increase in A/K and A/GPP ratios in colonized plants of examined species. The decrement of K and GPP was particularly clear in Hybiscus syriacus. The decrement in net photosynthesis and dark respiration in infested leaves is in concordance with the findings of some other studies [31,20], suggesting that changes in traits describing gas exchange related to photosynthesis are more precise and more likely to be detected than in traits describing the content of photosynthetic pigments.
The influence of aphids depends on the variety, plant age, and developmental stage of the aphid species. The high number of aphids influences the physiological state of the plant, and it is well known that the process of photosynthesis is especially sensitive to such influences. In colonized plants, a decrement of photosynthesis intensity of 10 – 20% was recorded, depending on the aphid species and its host, which is in concordance with the results of Miles [32], as well as Shahzad et al. [25], who found decreased both in photosynthetic rate and grain yield in several wheat cultivars. At the same time, the decrease of dark respiration was even more intensive varying from 26.5 in Spirea trilobata to 34.4% in colonized plants of Hybiscus syriacus. The fact that the decrease in dark respiration is more intense than the decrease in net respiration is confirmed by the increment of A/K and A/GPP ratios, suggesting that net photosynthesis is favoured against dark respiration by plants during aphid infestation.
By feeding on plants, the aphids take nutrients of phloem sap that are necessary for their growth and development. In our study, aphids caused a significant decrement in nitrogen content and an increase in C/N ratio in all examined species, in total and within the species at the level of interaction Species × Colonization. In total, the partition of carbon was found to be lower in colonized than in non-colonized leaves, while there were no significant differences in hydrogen partition. However, differences in the partition of carbon between non-colonized and colonized leaves were not significant within examined species at the level of interaction Species × Colonization. These data emphasize the importance of nitrogen content and C/N ratio in the evaluation of the reaction of plants to aphid colonization. Both N and C belong to the same PCA group as C/N. However, according to the results of Tukey’s HSD test, the difference in C/N seems to be more related to differences in N, than in C. Indeed, the same is confirmed by factorial loadings with principal components, where the rest of the variance of C/N and N that is not explained by the first principal component is more explained by the second principal component, while that of C is more explained by the third principal component.
Such close relation of the partition of two of three examined basic elements: nitrogen and carbon, as well as the C/N ratio with the aphid colonization, could be of considerable importance for further research in plant reaction on aphid infestation.
According to Dong et al. [33], amino acid metabolism is upregulated in leaf of citrus plants by psyllid feeding, the leaves were thicker, but the partition of nitrogen was not significantly different than in control leaves. According to Sadras et al. [34], the C/N ratio is closely related to plants' suitability to herbivores, where high C/N reduces the fitness of insects on host plants. This parameter is more closely associated with photosynthetic protein content than the concentration of amino acids in sap. They found a negative correlation between the number of Rhopalosiphum padi on wheat leaves and the molar concentration of sugars in the stem, stressing that besides lowering of nitrogen content, another important defending mechanism of the plant is the increment of content of labile carbohydrates which can lead to osmotic stress in aphids. Both mechanisms, low nitrogen content and increment of phloem sap carbohydrates’ concentration, could contribute to the decrement of nitrogen content and of C/N ratio in colonized leaves that were found in our study. In the course of evolution, many insects developed mechanisms for improved uptake of nitrogen from plant tissues, considering the fact that, regarding the insects’ needs, these tissues have a relatively low content of this element [35,36]. Thus, it seems that nitrogen partition and C/N could be directly linked with the reaction of the plant to the aphid colonization, which is in concordance with the significant difference in these parameters between colonized and non-colonized leaves that were found in our study. However, Hawkins et al. [37] presented data that suggest the inhibitory effect of aphid colonization on N and P uptake in some herbaceous leguminous species. In cowpea and pea, the N and P percentage of plant dry weight in infested plants of several herbaceous leguminous plant species did not differ from control plants, but because the control plants were greater, the absolute amounts of N and P in the were higher than in infested plants. The opposite was found in broad beans. As such, a decrease in N could represent a defense mechanism the plant uses to combat the aphid infestation as well as the outcome of the aphids' feeding reducing the plant's ability to consume nitrogen.
Based on the acquired results and previous studies, we assume that N and C/N, two parameters that are relatively easy to determine, would be important in the fast assessment of plant species responses to aphids and other sap-sucking insects. As a result, future research on this topic should give close attention to these criteria.
Author Contributions
L.P.P., N.N., B.K. and S.O. designed the study, analysed, and interpreted the results; L.P.P., M.D. and L.K., collected samples; L.P.P. determined aphid species; L.P.P., N.N., and S.O. performed the laboratory experiment; L.P.P., B.K., and N.N. performed the statistical analysis and interpreted the results; L.P.P., B.K., N.N., M.D. and V.V. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the Ministry of Science, Technological Development, and Innovation of the Republic of Serbia (Contract No. 451-03-47/2023-01/200197).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Endsley, K.A. Remote sensing of socio-ecological dynamics in urban neighbourhoods. In Comprehensive Remote Sensing; Liang, S., Ed.; Elsevier Inc.: The Netherland, 2018; Volume 9, pp. 90–105. [Google Scholar] [CrossRef]
- De Carvalho, R.M.; Szlafsztein, C.F. Urban vegetation loss and ecosystem services: The influence on climate regulation and noise and air pollution. Environ. Pollut. 2019, 245, 844–852. [Google Scholar] [CrossRef]
- Stroud, S. , Peacock, J., Hassall, C. (2022): Vegetation-based ecosystem service delivery in urban landscapes: A systematic review. Basic Appl. Ecol. 2022, 61, 82–101. [Google Scholar] [CrossRef]
- Vastag, E.; Kesić, L.; Karaklić, V.; Zorić, M.; Vuksanović, V.; Stojnić, S. Physiological performance of sweetgum (Liquidambar styraciflua L.) and Norway Maple (Acer platanoides L.) under drought conditions in urban environment. Topola/Poplar 2019, 204, 17–27. [Google Scholar]
- Jones, L.; Anderson, S.; Læssøe, J.; Banzhaf, E.; Jensen, A.; Bird, D.N.; Miller, J.; Hutchins, M.G.; Yang, J.; Garrett, J.; Taylor, T.; Wheeler, B.W.; Lovell, R.; Fletcher, D.; Qu, Y.; Vieno, M.; Zandersen, M. A typology for urban Green Infrastructure to guide multifunctional planning of nature-based solutions. Nature-Based Solutions 2022, 2, 100041. [Google Scholar] [CrossRef]
- Khan, N.; Jhariya, M.K.; Yadav, D.K.; Banerjee, A. Structure, diversity and ecological function of shrub species in an urban setup of Sarguja, Chhattisgarh, India. Environ Sci Pollut Res Int. 2020, 27, 5418–5432. [Google Scholar] [CrossRef] [PubMed]
- Poljaković-Pajnik, L.; Stojanović, D.; Drekić, M.; Pilipović, A.; Vasić, V. Climate change and invasive insect species in forests, urban areas and nurseries in Serbia. In Proceedings of the Water in Forests International Conference of KASZÓ-LIFE, Croatia, Serbia, 29–30 May 2018. [Google Scholar]
- Ilić Milošević, M.; Žikić, V.; Milenković, D.; Stanković, S.S.; Olivera Petrović-Obradović, O. Diversity of aphids (Homoptera: Aphididae) in Southeastern Serbia. Biologica Nyssana 2019, 10, 49–57. [Google Scholar] [CrossRef]
- Borbély, C.; György, Z.; Jacobsen, S.K.; Musa, F.; Ouředníčková, J.; Sigsgaard, L.; Skalsky, M.; Marko, V. First records of the invasive aphid species, Aphis spiraecola, in Kosovo, Slovakia, the Czech Republic, the United Kingdom and Denmark. Plant Protect. Sci. 2021, 57, 70–74. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide, 2nd ed.; John Wiley and Sons Ltd.: Chichester, UK, 2000. [Google Scholar]
- Dedryver, C.-A.; Le Ralec, A.; Fabre, F. The conflicting relationships between aphids and men: A review of aphid damage and control strategies. C. R. Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef]
- Coeur d’Acier, A.; Hidalgo, N.P.; Petrović-Obradović, O. Aphids (Hemiptera, Aphididae). Chapter 9.2. BioRisk. 2010, 4, 435–474. [Google Scholar] [CrossRef]
- Szpeiner, A. Aphididae (Hemiptera) on ornamental plants in Córdoba (Argentina). Rev. Soc. Entomol. Argent. 2008, 67, 49–56. [Google Scholar]
- Gleditsch, J.M.; Carlo, T.A. Fruit quantity of invasive shrubs predicts the abundance of common native avian frugivores in central Pennsylvania. Divers. Distrib. 2011, 17, 244–253. [Google Scholar] [CrossRef]
- Cuperus, G.W.; Radcliffe, E.B.; Barnes, D.K.; Marten, G.C. Economic injury levels and economic thresholds for pea aphid, Acyrthosiphon pisum (Harris), on alfalfa. Crop Prot. 1982, 1, 453–463. [Google Scholar] [CrossRef]
- Burd, J.D.; Elliott, N.C. Changes in Chlorophyll a Fluorescence Induction Kinetics in Cereals Infested with Russian Wheat Aphid (Homopetra: Aphididea). J. Econ. Entomol. 1996, 89, 1332–1337. [Google Scholar] [CrossRef]
- Telang, A.; Sandström, J.; Dyreson, E.; Moran, N.A. Feeding damage by Diuraphis noxia results in a nutritionally enhanced phloem diet. Entomol. Exp. Appl. 1999, 91, 403–412. [Google Scholar] [CrossRef]
- Ni, X.; Quisenberry, S.S.; Heng-Moss, T.; Markwell, J.; Higley, L.; Baxendale, F.; Sarath, G.; Klucas, R. Dynamic change in photosynthetic pigments and chlorophyll degradation elicited by cereal aphid feeding. Entomol. Exp. Appl. 2002, 105, 43–53. [Google Scholar] [CrossRef]
- Wang, T. , Quisenberry, S.S., Ni, X., Tolmay, V. Aphid (Hemiptera: Aphididae) resistance in wheat near-isogenic lines. J. Econ. Entomol. 2004, 97, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Poljaković-Pajnik, L.; Petrović-Obradović, O.; Drekić, M.; Orlović, S.; Vasić, V.; Kovačević, B.; Kacprzyk, M.; Nikolić, N. Aphid feeding effects on physiological parameters of poplar cultivars. Proceedings of rge VII International Scientific Agriculture Symposium “Agrosym 2016”, Jahorina, Bosnia and Herzegovina, 6–9 October 2016; pp. 2848–2855. [Google Scholar]
- Hurej, M.; Werf, W.V.D. The influence of black bean aphid, Aphis fabae Scop., and its honeydew on the photosynthesis of sugar beet. Ann. Appl. Biol. 1993, 122, 189–200. [Google Scholar] [CrossRef]
- Gonzalez, L.; Bolaño, C.; Pelissier, F. Use of oxygen electrode in measurements of photosynthesis and respiration. In Handbook of Plant Ecophysiology Techniques; Regiosa Roger, M.J., Ed.; Springer: Dordrecht, Holand, 2001; pp. 141–153. [Google Scholar] [CrossRef]
- Wettstein, D. Chlorophyll-letate und submikroskopische Formwechsel der Plastiden. Exp. Cell. Res. 1957, 12, 427–433. [Google Scholar] [CrossRef] [PubMed]
- TIBCO Software Inc. (2020). Data science work-bench, version 14. Web site. Available online: http://www.tibco.com/products/data-science.
- Shahzad, M.W.; Ghani, H.; Ayyub, M.; Ali, Q.; Ahmad, H.M.; Faisal, M.; Ali, A.; Qasim, M.U. Performance of some wheat cultivars against aphid and its damage on yield and photosynthesis. J. Glob. Innov. Agric. Soc. Sci. 2019, 7, 105–109. [Google Scholar] [CrossRef]
- Goławska, S.; Krzyżanowski, R.; Łukasik, I. Relationship between aphid infestation and chlorophyll content in Fabaceae species. Acta Biol. Crac. Ser. Bot. 2010, 52/2, 76–80. [Google Scholar] [CrossRef]
- Cárdenas, A.M.; Gallardo, P. Relationship between insect damage and chlorophyll content in Mediterranean oak species. Appl. Ecol. Environ. Res. 2016, 14, 477–491. [Google Scholar] [CrossRef]
- Diaz-Montano, J.; Reese, J.; William Schapaugh, W.; Campbell, L. Chlorophyll Loss Caused by Soybean Aphid (Hemiptera: Aphididae) Feeding on Soybean. J. Econ. Entomol. 2007, 100, 1657–1662. [Google Scholar] [CrossRef]
- Orlović, S.; Guzina, V. , Krstić, B.; Merkulov. Lj. Genetic variability in anatomical, physiological and growth characteristics of hybrid poplar (Populus x euramericana Dode (Guinier)) and eastern cottonwood (Populus deltoides Bartr.) clones. Silvae Genet. 1998, 47, 183–190. [Google Scholar]
- Miles, P.W. Feeding process of Aphidoidea in relation to effects on their food plants. In Aphids: their Biology, Natural Enemies and Control; Minks, A.K., Harrewijn, P., Eds.; Elsevier: Amsterdam, Holand, 1987; Volume 2A, pp. 321–339. [Google Scholar]
- Cichocka, E.; Goszczyński, W.; Chacińska, M. The effect of aphids on host plants. I. Effect on photosynthesis, respiration and transpiration. Aphids and Other Homopterous Insects 1992, 3, 59–64. [Google Scholar]
- Miles, P.W. Insect secretions in plants. Annu. Rev. Phitopathol. 1968, 6, 137–164. [Google Scholar] [CrossRef]
- Dong, Z.H.; Liu, X.D.; Srivastava, A.K.; Tan, Q.L.; Low, W.; Yan, X.; Wu, S.W.; Sun, X.C.; Hu, C.X. Foliar nutrition alleviate citrus plants from Asian citrus psyllid feeding by affecting leaf structure and secondary metabolism. Sci. Hortic. 2023, 309, 111667. [Google Scholar] [CrossRef]
- Sadras, V.; Vázquez, C.; Garzo, E.; Moreno, A.; Medina, S.; Taylor, J.; Fereres, A. The role of plant labile carbohydrates and nitrogen on wheat-aphid relations. Sci. Rep. 2021, 11, 12529. [Google Scholar] [CrossRef] [PubMed]
- Slansky, F., Jr. Insect Nutritional Ecology as a Basis for Studying Host Plant Resistance. Fla. Entomol. 1990, 73, 359–378. [Google Scholar] [CrossRef]
- Slansky, F., Jr. Nutritional ecology: the fundamental quest for nutrients. In Caterpillars. Ecological and Evolutionary Constraints on Foraging; Stamp, N.E., Casey, T.M., Eds. Chapman and Hall: New York, USA, 1993; pp. 29–91. [Google Scholar]
- Hawkins, C.D.B.; Whitecross, M.I.; Aston, M.J. Interactions between aphid infestation and plant growth and uptake of nitrogen and phosphorus by three leguminous host plants. Can. J. Bot. 1986, 64, 2362–2367. [Google Scholar] [CrossRef]
Figure 1.
Factor loadings for the first two principal components.
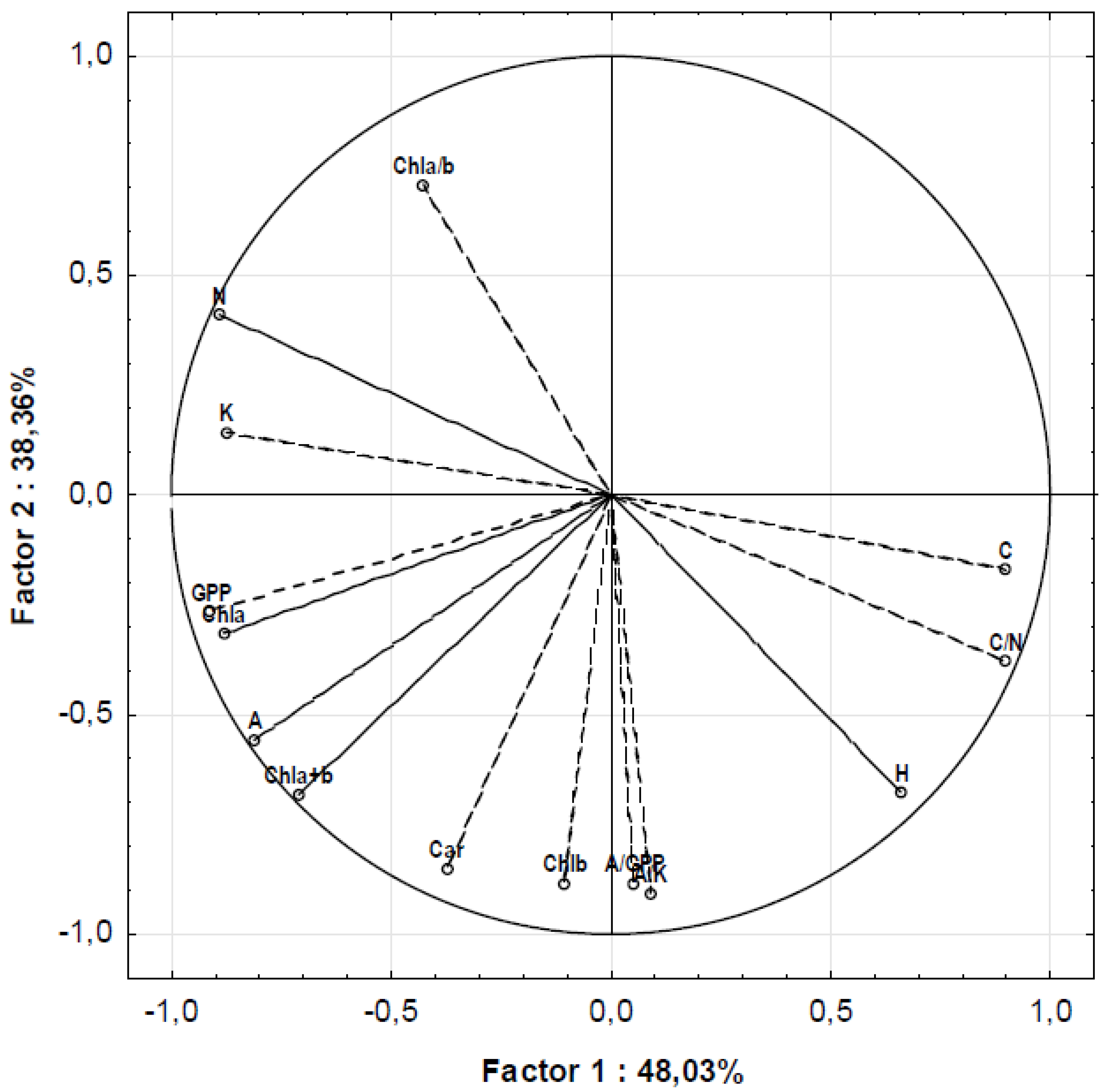

Table 1.
F-test from two-way factorial analysis of variance for examined parameters.
| Source of variation | Chla | 1 | Chlb | Chla+b | Chla/b | Car | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species (A) | 5.286 | * | 3.590 | 7.167 | ** | 2.883 | 12.341 | ** | ||
| Colonization (B) | 0.978 | 0.135 | 0.249 | 0.141 | 2.479 | |||||
| Interaction A×B | 2.281 | 0.680 | 1.205 | 0.762 | 2.920 | |||||
| N | C | H | C/N | |||||||
| Species (A) | 2330.300 | ** | 295.800 | ** | 72.900 | ** | 17159.280 | ** | ||
| Colonization (B) | 251.000 | ** | 5.300 | * | 0.000 | 1215.895 | ** | |||
| Interaction A×B | 21.800 | ** | 1.700 | 2.400 | 202.505 | ** | ||||
| K | A | GPP | A/K | A/GPP | ||||||
| Species (A) | 22.747 | ** | 22.772 | ** | 26.742 | ** | 13.823 | ** | 15.478 | ** |
| Colonization (B) | 41.592 | ** | 8.354 | * | 25.434 | ** | 5.951 | * | 8.319 | * |
| Interaction A×B | 2.257 | 0.813 | 1.168 | 0.362 | 1.062 |
1 Labels of traits: Chla – content of chlorophyll a; Chlb – content of chlorophyll b; Chla+b - content of chlorophyll a+b, Chla/b – Chla/Chlb ratio; Car – content of carotenoids; N – partition of nitrogen; C – partition of carbon; H – partition of hydrogen; C/N – C/N ratio; K – dark respiration; A – net photosynthesis; GPP – gross primary productivity; A/K – A/K ratio; A/GPP – A/GPP ratio.
Table 2.
Tukey-s HSD-test for examined parameters in three shrub species with and without aphid colonization.
Table 2.
Tukey-s HSD-test for examined parameters in three shrub species with and without aphid colonization.
| Species | Colonization | Chla | 1 | Chlb | Chla+b | Chla/b | Car | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Spirea trilobata | 1.225 | ab | 0.694 | a | 1.919 | a | 2.130 | a | 0.672 | a | |
| Hybiscus syriacus | 1.430 | a | 0.460 | a | 1.890 | a | 3.440 | a | 0.490 | ab | |
| Cydonia japonica | 0.926 | b | 0.313 | a | 1.239 | b | 2.996 | a | 0.322 | b | |
| Non-colonized | 1.256 | a | 0.467 | a | 1.724 | a | 2.770 | a | 0.449 | a | |
| Colonized | 1.130 | a | 0.511 | a | 1.641 | a | 2.940 | a | 0.540 | a | |
| Spirea trilobata | Non-colonized | 1.418 | ab | 0.583 | a | 2.002 | a | 2.441 | a | 0.808 | a |
| Spirea trilobata | Colonized | 1.031 | ab | 0.805 | a | 1.836 | a | 1.820 | a | 0.536 | ab |
| Hybiscus syriacus | Non-colonized | 1.549 | a | 0.515 | a | 2.064 | a | 3.153 | a | 0.524 | ab |
| Hybiscus syriacus | Colonized | 1.310 | ab | 0.405 | a | 1.715 | a | 3.727 | a | 0.456 | b |
| Cydonia japonica | Non-colonized | 0.801 | b | 0.304 | a | 1.106 | a | 2.718 | a | 0.288 | b |
| Cydonia japonica | Colonized | 1.050 | ab | 0.321 | a | 1.371 | a | 3.274 | a | 0.355 | b |
| N | C | H | C/N | ||||||||
| Spirea trilobata | 2.912 | c | 47.768 | b | 6.871 | a | 16.421 | a | |||
| Hybiscus syriacus | 4.331 | a | 41.801 | c | 6.331 | c | 9.658 | c | |||
| Cydonia japonica | 3.049 | b | 48.764 | a | 6.656 | b | 16.072 | b | |||
| Non-colonized | 3.579 | a | 46.401 | a | 6.622 | a | 13.465 | b | |||
| Colonized | 3.282 | b | 45.821 | b | 6.617 | a | 14.636 | a | |||
| Spirea trilobata | Non-colonized | 3.006 | d | 47.855 | b | 6.905 | a | 15.921 | b | ||
| Spirea trilobata | Colonized | 2.818 | e | 47.682 | b | 6.836 | a | 16.922 | a | ||
| Hybiscus syriacus | Non-colonized | 4.447 | a | 41.967 | c | 6.358 | c | 9.437 | e | ||
| Hybiscus syriacus | Colonized | 4.215 | b | 41.634 | c | 6.305 | c | 9.878 | d | ||
| Cydonia japonica | Non-colonized | 3.284 | c | 49.381 | a | 6.602 | b | 15.037 | c | ||
| Cydonia japonica | Colonized | 2.815 | e | 48.147 | ab | 6.710 | ab | 17.107 | a | ||
| K | A | GPP | A/K | A/GPP | |||||||
| Spirea trilobata | 5.822 | b | 10.184 | a | 16.007 | b | 1.755 | a | 0.634 | a | |
| Hybiscus syriacus | 8.171 | a | 10.461 | a | 18.632 | a | 1.320 | b | 0.566 | b | |
| Cydonia japonica | 5.428 | b | 6.207 | b | 11.635 | c | 1.173 | b | 0.537 | b | |
| Non-colonized | 7.632 | a | 9.783 | a | 17.414 | a | 1.301 | b | 0.558 | b | |
| Colonized | 5.316 | b | 8.118 | b | 13.434 | b | 1.531 | a | 0.600 | a | |
| Spirea trilobata | Non-colonized | 6.711 | b | 11.368 | a | 18.079 | ab | 1.696 | ab | 0.628 | a |
| Spirea trilobata | Colonized | 4.934 | bc | 9.000 | abc | 13.934 | bcd | 1.814 | a | 0.640 | a |
| Hybiscus syriacus | Non-colonized | 9.868 | a | 11.447 | a | 21.316 | a | 1.169 | bc | 0.538 | bc |
| Hybiscus syriacus | Colonized | 6.474 | bc | 9.474 | ab | 15.947 | bc | 1.472 | abc | 0.595 | ab |
| Cydonia japonica | Non-colonized | 6.316 | bc | 6.533 | bc | 12.849 | cd | 1.039 | c | 0.509 | c |
| Cydonia japonica | Colonized | 4.539 | c | 5.882 | c | 10.421 | d | 1.307 | abc | 0.566 | abc |
1 Labels of traits: Chla – content of chlorophyll a [mg g-1 FW]; Chlb – content of chlorophyll b [mg g-1 FW]; Chla+b - content of chlorophyll a+b [mg g-1 FW], Chla/b – Chla/Chlb ratio; Car – content of carotenoids [mg g-1 FW]; N – partition of nitrogen [%]; C – partition of carbon [%]; H – partition of hydrogen [%]; C/N – C/N ratio; K – respiration [molO2 cm-2 h-1]; A – net photosynthesis [molO2 cm-2 h-1]; GPP – gross primary productivity [molO2 cm-2 h-1]; A/K – A/K ratio; A/GPP – A/GPP ratio.
Table 3.
Factor loadings for five principal components.
| Original variable a) |
Principal component b) | ||||
|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | |
| Chla | -0.881 | -0.313 | -0.042 | -0.312 | -0.162 |
| Chlb | -0.105 | -0.883 | 0.014 | 0.444 | -0.108 |
| Chla+b | -0.711 | -0.680 | -0.024 | -0.009 | -0.176 |
| Chla/b | -0.426 | 0.708 | -0.424 | -0.368 | -0.038 |
| Car | -0.373 | -0.851 | 0.105 | -0.330 | 0.133 |
| N | -0.892 | 0.411 | -0.066 | 0.157 | 0.082 |
| C | 0.898 | -0.171 | 0.375 | -0.133 | 0.071 |
| H | 0.662 | -0.678 | 0.238 | -0.212 | -0.032 |
| C/N | 0.899 | -0.377 | 0.126 | -0.145 | -0.114 |
| K | -0.875 | 0.143 | 0.461 | 0.027 | -0.025 |
| A | -0.810 | -0.557 | 0.143 | -0.034 | 0.115 |
| GPP | -0.911 | -0.267 | 0.309 | -0.007 | 0.057 |
| A/K | 0.089 | -0.907 | -0.398 | 0.059 | 0.081 |
| A/GPP | 0.053 | -0.883 | -0.461 | -0.013 | 0.064 |
| Eigenvalue | 6.725 | 5.370 | 1.111 | 0.653 | 0.141 |
| % of the total variance | 48.034 | 38.358 | 7.934 | 4.664 | 1.010 |
|
Cumulative Eigenvalue |
6.725 | 12.095 | 13.206 | 13.859 | 14.000 |
|
Cumulative % of the total variance |
48.034 | 86.392 | 94.326 | 98.990 | 100.000 |
a) Labels of traits: Chla – content of chlorophyll a; Chlb – content of chlorophyll b; Chla+b - content of chlorophyll a+b, Chla/b – Chla/Chlb ratio; Car – content of carotenoids; N – partition of nitrogen; C – partition of carbon; H – partition of hydrogen; C/N – C/N ratio; K – respiration; A – net photosynthesis; GPP – gross primary productivity; A/K – A/K ratio; A/GPP – A/GPP ratio. b) The highest loadings of original variables are bolded
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



