Submitted:
22 December 2023
Posted:
22 December 2023
You are already at the latest version
Abstract
The C-X-C motif chemokine receptor-1 (CXCR1) is a rhodopsin-like G-protein-coupled receptor, expressed on the cell surface of immune cells and tumors. CXCR1 interacts with some C-X-C chemokines, such as CXCL6, CXCL7, and CXCL8/interleukin-8, which are produced by various cells. Since CXCR1 is involved in several diseases including tumors and diabetes mellitus, drugs targeting CXCR1 has been developed. Therefore, the development of sensitive monoclonal antibodies (mAbs) for CXCR1 has been desired for the diagnosis and treatment. This study established a novel anti-mouse CXCR1 (mCXCR1) mAb; Cx1Mab-1 (rat IgG1, kappa), using the Cell-Based Immunization and Screening (CBIS) method. Cx1Mab-1 reacted with mCXCR1-overexpressed Chinese hamster ovary-K1 (CHO/mCXCR1) and mCXCR1-overexpressed LN229 glioblastoma (LN229/mCXCR1) in flow cytometry. Cx1Mab-1 demonstrated a high binding affinity for CHO/mCXCR1 and LN229/mCXCR1 with a dissociation constant of 2.6×10-9 M and 2.1×10-8 M, respectively. Furthermore, Cx1Mab-1 could detect mCXCR1 by western blot analysis. These results indicated that Cx1Mab-1 is useful for detecting mCXCR1, and provides a possibility for targeting mCXCR1-expressing cells in vivo experiments.
Keywords:
mouse CXCR1
; CBIS method
; monoclonal antibody
; flow cytometry
1. Introduction
Chemokines can be classified into four different subfamilies: C-C, C-X-C, C-X3-C, and X-C, depending on the number and position of cysteine residues in their N-terminus. C-X-C means the first two cysteine amino acids are divided by other amino acids. Chemokines play pivotal roles in immune responses, such as infiltration and migration of immune cells by binding to its cognate receptors [1,2,3].
The C-X-C motif chemokine receptor 1 (CXCR1), also referred to as C-X-C motif chemokine 8 (CXCL8) receptor/interleukin-8 (IL-8) receptor alpha, is one of the G-protein coupled receptors (GPCRs) composed of 350 amino acids with structural seven transmembrane domains [4]. CXCL6 and CXCL7 are also the chemokine ligands of CXCR1 [5]. At the nearly same time that CXCR1 was discovered, the C-X-C motif chemokine receptor 2 (CXCR2), a receptor for CXCL6, CXCL7, and IL-8, was also identified [6,7,8]. CXCR2 is also a receptor for CXCL2 and CXCL3 and has 77% amino acids homology with CXCR1 [9]. Various cells express CXCR1/CXCR2 on the cell surface, including macrophages, neutrophils, basophils, natural killer cells, mast cells, and other leukocytes [10,11]. Furthermore, CXCR1/CXCR2 expression has also been confirmed in non-immune cells, such as endothelial cells, fibroblasts, and tumor cells [5,12]. Due to their homology and ligand overlapping, the functions of CXCR1/CXCR2 have been clarified in many diseases [13].
The IL-8 plays a pivotal role in the tumor microenvironment (TME) as a factor of immunosuppressive cells’ recruitment, angiogenesis, and induction of epithelial to mesenchymal transition (EMT) [4]. Functional heterogeneity of cancer-associated fibroblasts (CAFs) is regulated by IL-8 and other CXCR1/2 ligands [14]. The normal ovarian fibroblast is converted to CAFs by IL-8 and triggers tumor growth in mice. Aberrant CXCR1/2 ligands expression in CAFs is closely related to gastric cancer progression and poor prognosis in patients [15]. In TME of glioma, IL-8+CD4+ T cells function in a complex manner with other IL-8-producing cells to govern the immunosuppressive properties [16]. IL-8 expression is positively correlated with CXCR1/2 and might be a predictive marker of poor prognosis in glioma, metastatic urothelial carcinoma, and metastatic renal cell carcinoma patients [17,18]. Furthermore, the blockade of the IL-8-CXCR1/2 axis enhances the anti-programmed cell death-1 (PD-1) therapeutic effects on glioma [19]. Thus, CXCR1/2 molecules are promising novel cancer therapeutic targets for tumors [20]. Reparixin, a non-competitive allosteric inhibitor of CXCR1/2, could prevent polymorphonuclear cell recruitment [21]. A phase II clinical trial using navarixin (SCH-527123), a selective CXCR1/2 antagonist, has been conducted in combination with anti-PD-1 mAb, pembrolizumab, in advanced solid tumors [22,23]. Navarixin blocks neutrophil recruitment and suppresses the tumor cell growth in a murine melanoma model. CXCR1/2 are attractive targets for treatment against multiple cancers.
The CXCR1/2 and their chemokine ligands regulate the immune system, which mediates the progression of diseases such as cancer. Therefore, CXCR1/2 targeting antibodies will contribute to the diagnosis and therapy. Using the Cell-Based Immunization and Screening (CBIS) method, we have developed many monoclonal antibodies (mAbs) against C-C chemokine receptors, including anti-mouse CCR3 [24], anti-mouse CCR8 [25,26,27], and anti-human CCR9 [28] mAbs. Next, we have been trying to establish the anti-C-X-C motif chemokine receptor mAbs. We recently obtained an anti-mouse CXCR3 mAb [29], an anti-mouse CXCR4 mAb [30], an anti-mouse CXCR6 mAb [31]. In this study, we have successfully developed anti-mouse CXCR1 (mCXCR1) mAbs using the CBIS method which is applicable to flow cytometry and western blot analysis.
2. Materials and Methods
2.1. Cell lines
LN229, Chinese hamster ovary (CHO)-K1, and P3X63Ag8U.1 (P3U1) cells were obtained from the American Type Culture Collection (Manassas, VA). pCMV6neo-myc-DDK vector with mCXCR1 (Accession No.: NM_178241) was purchased from OriGene Technologies, Inc. (Rockville, MD). The plasmid was transfected into the cell lines using a Neon transfection system (Thermo Fisher Scientific, Inc., Waltham, MA). Subsequently, LN229 and CHO-K1, which stably overexpressed mCXCR1 with C-terminal myc-DDK tags (hereinafter described as LN229/mCXCR1 and CHO/mCXCR1, respectively) were established using a cell sorter (SH800; Sony Corp., Tokyo, Japan), following cultivation in a medium containing 0.5 mg/mL G418 (Nacalai Tesque, Inc., Kyoto, Japan).
CHO-K1, P3U1, and CHO/mCXCR1 were also cultured in a Roswell Park Memorial Institute (RPMI) 1640 medium (Nacalai Tesque, Inc.) that was supplemented with 10% heat-inactivated fetal bovine serum (FBS, Thermo Fisher Scientific Inc.), 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). LN229 and LN229/mCXCR1 were cultured in a Dulbecco’s Modified Eagle Medium (DMEM) (Nacalai Tesque, Inc.) that was supplemented with 10% heat-inactivated FBS (Thermo Fisher Scientific Inc.), 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Nacalai Tesque, Inc.). Then, cells were grown in a humidified incubator, which was supplied with 5% CO2 and 95% air at 37°C.
2.2. Antibodies
The anti-mouse CXCR1/IL-8RA mAb (clone 1122A) was purchased from R&D Systems Inc. (Minneapolis, MN). An anti-DYKDDDDK mAb (clone 1E6) and an anti-β-actin mAb (clone AC-15) were purchased from FUJIFILM Wako Pure Chemical Corporation (Osaka, Japan) and Sigma-Aldrich Corp. (St. Louis, MO), respectively. The secondary Alexa Fluor 488-conjugated anti-rat IgG and Alexa Fluor 488-conjugated anti-rabbit IgG were purchased from Cell Signaling Technology, Inc. (Danvers, MA).
2.3. Production of hybridomas
For developing anti-mCXCR1 mAbs, a six-week-old female Sprague-Dawley rat (CLEA Japan, Tokyo, Japan) was immunized intraperitoneally with 1 × 109 cells of LN229/mCXCR1. The immunogen was harvested after brief exposure to 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.). We added Alhydrogel adjuvant 2% (InvivoGen, San Diego, CA) as an adjuvant in the first immunization. Three additional injections of 1 × 109 cells of LN229/mCXCR1 were performed without an adjuvant every week. We performed a final booster immunization of 1 × 109 cells of LN229/mCXCR1 intraperitoneally two days before harvesting splenocytes. We fused the harvested splenocytes with P3U1 cells using polyethylene glycol 1500 (PEG1500; Roche Diagnostics, Indianapolis, IN). Hybridoma cells were cultured in the RPMI1640 medium, supplemented as shown above. We further added hypoxanthine, aminopterin, and thymidine (HAT; Thermo Fisher Scientific, Inc.), 5% Briclone (NICB, Dublin, Ireland), and 5 μg/mL of plasmocin into the medium. The hybridoma supernatants were screened by flow cytometry using CHO/mCXCR1 and parental CHO-K1. The culture supernatants of hybridomas were filtrated and purified using Ab-Capcher Extra (ProteNova, Kagawa, Japan).
2.4. Flow cytometry
CHO-K1 and CHO/mCXCR1 cells were harvested after brief exposure to 2.5 g/L-Trypsin/ 1 mmol/L-EDTA Solution with Phenol Red (Nacalai Tesque, Inc.). CHO-K1, CHO/mCXCR1, LN229, and LN229/mCXCR1 cells were washed with 0.1% bovine serum albumin in phosphate-buffered saline (PBS) and treated with primary mAbs for 30 min at 4°C. Afterward, cells were treated with Alexa Fluor 488-conjugated anti-rat IgG (1:1000) or Alexa Fluor 488-conjugated anti-rabbit IgG (1:1000) as well, following the collection of fluorescence data, using an SA3800 Cell Analyzer (Sony Corp., Tokyo, Japan).
2.5. Determination of the binding affinity by flow cytometry
CHO/mCXCR1 and LN229/mCXCR1 were suspended in 100 μL serially diluted anti-mCXCR1 mAbs (25 to 0.006 µg/mL), after which Alexa Fluor 488-conjugated anti-rat IgG (1:200) or Alexa Fluor 488-conjugated anti-rabbit IgG (1:200) were added. Fluorescence data were subsequently collected, using an SA3800 Cell Analyzer (Sony Corp.), following calculation of the dissociation constant (KD) by fitting the binding isotherms into the built-in; one-site binding model in GraphPad PRISM 8 (GraphPad Software, Inc., La Jolla, CA).
2.6. Western blot analysis
Cell lysates (10 μg) were boiled in sodium dodecyl sulfate (SDS) sample buffer (Nacalai Tesque, Inc.), after which proteins were separated on 5%–20% polyacrylamide gels (FUJIFILM Wako Pure Chemical Corporation) and transferred to polyvinylidene difluoride membranes (Merck KGaA). After blocking with 4% skim milk (Nacalai Tesque, Inc.) in 0.05% Tween 20-containing PBS, membranes were incubated with 1 μg/mL of Cx1Mab-1, 1 μg/mL of an anti-mouse CXCR1/IL-8RA mAb (clone 1122A), 1 μg/mL of an anti-DYKDDDDK mAb (clone 1E6), or 1 μg/mL of an anti-β-actin mAb (clone AC-15). Then, they were incubated again with horseradish peroxidase-conjugated anti-rat immunoglobulins (for Cx1Mab-1; diluted 1:10000; Sigma-Aldrich Corp.), anti-mouse immunoglobulins (for anti-DYKDDDDK, and anti-β-actin; diluted 1:1000; Agilent Technologies, Inc., Santa Clara, CA), or anti-rabbit immunoglobulins (for 1122A; diluted 1:1000; Agilent Technologies, Inc.). Finally, protein bands were detected using ECL Plus Western Blotting Substrate (Thermo Fisher Scientific, Inc.) or ImmunoStar LD (FUJIFILM Wako Pure Chemical Corporation) with a Sayaca-Imager (DRC Co. Ltd., Tokyo, Japan).
3. Results
3.1. Establishment of anti-mCXCR1 mAbs
To develop anti-mCXCR1 mAbs, we employed the CBIS method using mCXCR1-overexpressed cells. Hybridoma screening was conducted using flow cytometry (Figure 1). A rat was intraperitoneally immunized with LN229/mCXCR1. Subsequently, hybridomas were seeded into 96-well plates, after which flow cytometric analysis was used to extract CHO/mCXCR1-reactive and CHO-K1-nonreactive supernatants. Afterward, we obtained CHO/mCXCR1-reactive supernatants in 78 of 958 wells (8.1%). We finally established clone Cx1Mab-1 (rat IgG1, kappa) by limiting dilution and additional screening.
3.2. Flow cytometry
Flow cytometric analysis was conducted using Cx1Mab-1 and another anti-mCXCR1/IL-8RA mAb (clone 1122A from R&D Systems Inc.) against CHO-K1, CHO/mCXCR1, LN229, and LN229/mCXCR1. Results showed that Cx1Mab-1 and 1122A recognized CHO/mCXCR1 dose-dependently, but not parental CHO-K1 cells (Figure 2). Both mAbs also recognized LN229/mCXCR1 dose-dependently, but not parental LN229 cells (Figure 3). Both Cx1Mab-1 and 1122A reacted with CHO/mCXCR1 and LN229/mCXCR1 even at 0.01 μg/mL concentration (Figure 2 and Figure 3), indicating that both mAbs are highly sensitive.
3.3. Determination of the binding affinity of Cx1Mab-1
The binding affinity of Cx1Mab-1 and 1122A were assessed with CHO/mCXCR1 and LN229/mCXCR1 using flow cytometry. Results showed that the KD of Cx1Mab-1 for CHO/mCXCR1 and LN229/mCXCR1 were 2.6×10-9 M and 2.1×10-8 M, respectively (Figure 4). In contrast, the KD of 1122A for CHO/mCXCR1 and LN229/mCXCR1 were 2.7 × 10−9 M and 2.8 × 10−8 M, respectively (Figure 4). These results indicate that Cx1Mab-1 possesses a moderate affinity for exogenously overexpressed-mCXCR1 in CHO-K1 and LN229 cells.
3.4. Western blot analysis
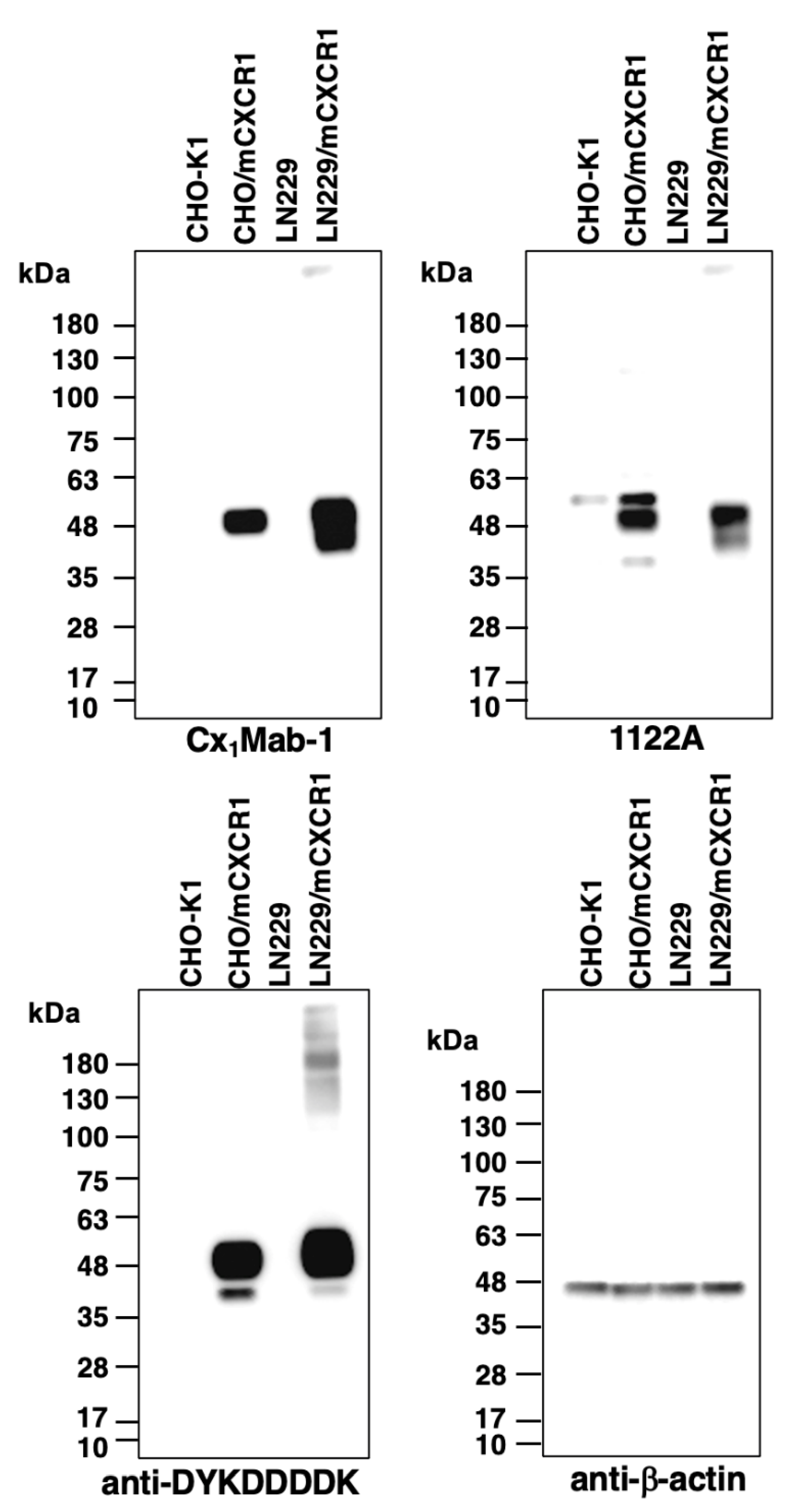
Western blot analysis was performed to further assess the availability of Cx1Mab-1. Lysates of CHO-K1, CHO/mCXCR1, LN229, and LN229/mCXCR1 cells were probed. As demonstrated in Figure 5, Cx1Mab-1 detected the ~48-kDa band of mCXCR1 in lysates from CHO/mCXCR1 and LN229/mCXCR1 cells, whereas this band was not present in lysates from CHO-K1 and LN229 cells, indicating that Cx1Mab-1 specifically detects mCXCR1-overexpressed cells.
4. Discussion
The TME is composed of various cells including cancer cells, immune cells, and stromal cells [32]. Chemokines and their receptors expressed in these cells modulate a complex TME [23,32]. Neutrophil is one of the major mediators of inflammation. IL-8 secreted from CD4+ T cells recruited CXCR1/2-expressed neutrophils to the inflammation site [33]. The Neutrophils promote inflammation by generating reactive oxygen species (ROS) [19,34]. In this study, we established Cx1Mab-1 which recognizes mCXCR1 with high sensitivity (Figure 2 and Figure 3). Therefore, Cx1Mab-1 would contribute to detecting and isolating the mCXCR1-positive lymphocytes.
Tumor-associated macrophage (TAM) plays a vital role as a tumor immune regulator in TME [35]. TAM produces IL-8 which attenuates cancer immunity by suppressing the infiltration of CD8+ T cells and enhances programmed cell death-1 ligand-1 (PD-L1) expression in TAM itself [36]. Blocking IL-8 in TAM by neutralizing antibody suppresses epithelial ovarian cancer migration [37]. Furthermore, CXCR1/2-positive neutrophils and myeloid-derived suppressor cells in cancer patients also play an important role in immune suppression by obstructing the contact between CD8+ T cells and the target tumor cells [19]. Therefore, it is interesting to investigate whether Cx1Mab-1 interferes with the IL-8-CXCR1 interaction. Furthermore, the identification of epitope is essential to know the property of Cx1Mab-1.
Therapeutic drugs that target CXCR1/2 have been developed for cancers [12]. SCH527123 is an orally available allosteric inhibitor of CXCR1/2 that can inhibit the neutrophil activity [22,38]. Reparixin selectively blocks CXCR1 which is expressed in human breast cancer stem cells (CSCs) in preclinical models [39]. Furthermore, reparixin significantly decreases tumor growth in combination with paclitaxel or docetaxel compared to chemotherapy alone [39,40]. Anti-CXCR1 mAbs also have the potential to affect CSCs in vitro [39]. Furthermore, the blockers for CXCR1/2 potentiate the efficacy of immune checkpoint inhibitors (ICIs) through suppression of neutrophils and myeloid-derived suppressor cells [19,41,42]. Numerous clinical trials demonstrate the promise of combination therapies treating CXCR1/2-targeted drugs and ICIs [4,43]. We have previously enhanced antibody-dependent cellular cytotoxicity (ADCC) activity complement-dependent cytotoxicity (CDC) by modifying mAb isotypes and defucosylation in mAbs [44,45,46]. Since Cx1Mab-1 is rat IgG1 which has no ADCC and CDC activities, it will be converted into a mouse IgG2a version to verify the effect of depletion of CXCR1+ immunosuppressive cells in preclinical models.
References
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. Febs j 2018, 285, 2944–2971. [Google Scholar] [CrossRef] [PubMed]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat Rev Immunol 2017, 17, 559–572. [Google Scholar] [CrossRef]
- White, G.E.; Iqbal, A.J.; Greaves, D.R. CC chemokine receptors and chronic inflammation--therapeutic opportunities and pharmacological challenges. Pharmacol Rev 2013, 65, 47–89. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.J.; Li, Y.B.; Yang, L.X.; Cheng, H.J.; Liu, X.; Chen, H. Roles of the CXCL8-CXCR1/2 Axis in the Tumor Microenvironment and Immunotherapy. Molecules 2021, 27. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.C.; Garcia, C.C.; Teixeira, M.M.; Amaral, F.A. The CXCL8/IL-8 chemokine family and its receptors in inflammatory diseases. Expert Rev Clin Immunol 2014, 10, 593–619. [Google Scholar] [CrossRef]
- Murphy, P.M.; Tiffany, H.L. Cloning of complementary DNA encoding a functional human interleukin-8 receptor. Science 1991, 253, 1280–1283. [Google Scholar] [CrossRef]
- Holmes, W.E.; Lee, J.; Kuang, W.J.; Rice, G.C.; Wood, W.I. Structure and functional expression of a human interleukin-8 receptor. Science 1991, 253, 1278–1280. [Google Scholar] [CrossRef]
- Wang, W.; Deng, Z.; Liu, G.; Yang, J.; Zhou, W.; Zhang, C.; Shen, W.; Zhang, Y. Platelet-derived extracellular vesicles promote the migration and invasion of rheumatoid arthritis fibroblast-like synoviocytes via CXCR2 signaling. Exp Ther Med 2021, 22, 1120. [Google Scholar] [CrossRef]
- Bi, H.; Zhang, Y.; Wang, S.; Fang, W.; He, W.; Yin, L.; Xue, Y.; Cheng, Z.; Yang, M.; Shen, J. Interleukin-8 promotes cell migration via CXCR1 and CXCR2 in liver cancer. Oncol Lett 2019, 18, 4176–4184. [Google Scholar] [CrossRef]
- Chuntharapai, A.; Lee, J.; Hébert, C.A.; Kim, K.J. Monoclonal antibodies detect different distribution patterns of IL-8 receptor A and IL-8 receptor B on human peripheral blood leukocytes. J Immunol 1994, 153, 5682–5688. [Google Scholar] [CrossRef] [PubMed]
- Tavares, L.P.; Garcia, C.C.; Machado, M.G.; Queiroz-Junior, C.M.; Barthelemy, A.; Trottein, F.; Siqueira, M.M.; Brandolini, L.; Allegretti, M.; Machado, A.M.; et al. CXCR1/2 Antagonism Is Protective during Influenza and Post-Influenza Pneumococcal Infection. Front Immunol 2017, 8, 1799. [Google Scholar] [CrossRef]
- Sitaru, S.; Budke, A.; Bertini, R.; Sperandio, M. Therapeutic inhibition of CXCR1/2: Where do we stand? Intern Emerg Med 2023, 18, 1647–1664. [Google Scholar] [CrossRef]
- Fu, X.; Wang, Q.; Du, H.; Hao, H. CXCL8 and the peritoneal metastasis of ovarian and gastric cancer. Front Immunol 2023, 14, 1159061. [Google Scholar] [CrossRef]
- Chen, Y.; McAndrews, K.M.; Kalluri, R. Clinical and therapeutic relevance of cancer-associated fibroblasts. Nat Rev Clin Oncol 2021, 18, 792–804. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Yamamoto, Y.; Sakamoto, N.; Shimomura, I.; Kogure, A.; Kumazaki, M.; Yokoi, A.; Yashiro, M.; Kiyono, T.; Yanagihara, K.; et al. Cancer extracellular vesicles contribute to stromal heterogeneity by inducing chemokines in cancer-associated fibroblasts. Oncogene 2019, 38, 5566–5579. [Google Scholar] [CrossRef] [PubMed]
- Teijeira, A.; Garasa, S.; Ochoa, M.C.; Villalba, M.; Olivera, I.; Cirella, A.; Eguren-Santamaria, I.; Berraondo, P.; Schalper, K.A.; de Andrea, C.E.; et al. IL8, Neutrophils, and NETs in a Collusion against Cancer Immunity and Immunotherapy. Clin Cancer Res 2021, 27, 2383–2393. [Google Scholar] [CrossRef] [PubMed]
- Schalper, K.A.; Carleton, M.; Zhou, M.; Chen, T.; Feng, Y.; Huang, S.P.; Walsh, A.M.; Baxi, V.; Pandya, D.; Baradet, T.; et al. Elevated serum interleukin-8 is associated with enhanced intratumor neutrophils and reduced clinical benefit of immune-checkpoint inhibitors. Nat Med 2020, 26, 688–692. [Google Scholar] [CrossRef]
- Yuen, K.C.; Liu, L.F.; Gupta, V.; Madireddi, S.; Keerthivasan, S.; Li, C.; Rishipathak, D.; Williams, P.; Kadel, E.E., 3rd; Koeppen, H.; et al. High systemic and tumor-associated IL-8 correlates with reduced clinical benefit of PD-L1 blockade. Nat Med 2020, 26, 693–698. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, Q.; Tan, L.; Wu, X.; Huang, R.; Zuo, Y.; Chen, L.; Yang, J.; Zhang, Z.X.; Ruan, W.; et al. Neutralizing IL-8 potentiates immune checkpoint blockade efficacy for glioma. Cancer Cell 2023, 41, 693–710. [Google Scholar] [CrossRef]
- Ruffini, P.A. The CXCL8-CXCR1/2 Axis as a Therapeutic Target in Breast Cancer Stem-Like Cells. Front Oncol 2019, 9, 40. [Google Scholar] [CrossRef]
- Bertini, R.; Allegretti, M.; Bizzarri, C.; Moriconi, A.; Locati, M.; Zampella, G.; Cervellera, M.N.; Di Cioccio, V.; Cesta, M.C.; Galliera, E.; et al. Noncompetitive allosteric inhibitors of the inflammatory chemokine receptors CXCR1 and CXCR2: Prevention of reperfusion injury. Proc Natl Acad Sci U S A 2004, 101, 11791–11796. [Google Scholar] [CrossRef] [PubMed]
- Chapman, R.W.; Minnicozzi, M.; Celly, C.S.; Phillips, J.E.; Kung, T.T.; Hipkin, R.W.; Fan, X.; Rindgen, D.; Deno, G.; Bond, R.; et al. A novel, orally active CXCR1/2 receptor antagonist, Sch527123, inhibits neutrophil recruitment, mucus production, and goblet cell hyperplasia in animal models of pulmonary inflammation. J Pharmacol Exp Ther 2007, 322, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sadanandam, A.; Nannuru, K.C.; Varney, M.L.; Mayer-Ezell, R.; Bond, R.; Singh, R.K. Small-molecule antagonists for CXCR2 and CXCR1 inhibit human melanoma growth by decreasing tumor cell proliferation, survival, and angiogenesis. Clin Cancer Res 2009, 15, 2380–2386. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Tanaka, T.; Sano, M.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 3 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Sano, M.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 8 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Saito, M.; Asano, T.; Tanaka, T.; Kitamura, K.; Kudo, Y.; Kaneko, M.K.; Kato, Y. C(8)Mab-3: An Anti-Mouse CCR8 Monoclonal Antibody for Immunocytochemistry. Monoclon Antib Immunodiagn Immunother 2022, 41, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of an Anti-Mouse CCR8 Monoclonal Antibody (C(8)Mab-1) for Flow Cytometry and Immunocytochemistry. Monoclon Antib Immunodiagn Immunother 2022, 41, 333–338. [Google Scholar] [CrossRef]
- Nanamiya, R.; Takei, J.; Asano, T.; Tanaka, T.; Sano, M.; Nakamura, T.; Yanaka, M.; Hosono, H.; Kaneko, M.K.; Kato, Y. Development of Anti-Human CC Chemokine Receptor 9 Monoclonal Antibodies for Flow Cytometry. Monoclon Antib Immunodiagn Immunother 2021, 40, 101–106. [Google Scholar] [CrossRef]
- Ouchida, T.; Isoda, Y.; Tanaka, T.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Cx<sub>3</sub>Mab-4: A Novel Anti-mouse CXCR3 Monoclonal Antibody for Flow Cytometry. Preprints 2023. [Google Scholar] [CrossRef]
- Ouchida, T.S., H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Cx4Mab-1: A Novel Anti-mouse CXCR4 Monoclonal Antibody for Flow Cytometry. Preprints 2023 2023, 2023110501. [Google Scholar] [CrossRef]
- Kitamura, K.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Cx(6)Mab-1: A Novel Anti-Mouse CXCR6 Monoclonal Antibody Established by N-Terminal Peptide Immunization. Monoclon Antib Immunodiagn Immunother 2022, 41, 133–141. [Google Scholar] [CrossRef]
- Borlongan, M.C.; Saha, D.; Wang, H. Tumor Microenvironment: A Niche for Cancer Stem Cell Immunotherapy. Stem Cell Rev Rep 2023. [Google Scholar] [CrossRef]
- Hammond, M.E.; Lapointe, G.R.; Feucht, P.H.; Hilt, S.; Gallegos, C.A.; Gordon, C.A.; Giedlin, M.A.; Mullenbach, G.; Tekamp-Olson, P. IL-8 induces neutrophil chemotaxis predominantly via type I IL-8 receptors. J Immunol 1995, 155, 1428–1433. [Google Scholar] [CrossRef]
- Thai, T.D.; Chuenchom, C.; Donsa, W.; Faksri, K.; Sripa, B.; Edwards, S.W.; Salao, K. Helicobacter pylori extract induces purified neutrophils to produce reactive oxygen species only in the presence of plasma. Biomed Rep 2023, 19, 89. [Google Scholar] [CrossRef] [PubMed]
- Qin, R.; Ren, W.; Ya, G.; Wang, B.; He, J.; Ren, S.; Jiang, L.; Zhao, S. Role of chemokines in the crosstalk between tumor and tumor-associated macrophages. Clin Exp Med 2023, 23, 1359–1373. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; He, H.; Liu, H.; Li, R.; Chen, Y.; Qi, Y.; Jiang, Q.; Chen, L.; Zhang, P.; Zhang, H.; et al. Tumour-associated macrophages-derived CXCL8 determines immune evasion through autonomous PD-L1 expression in gastric cancer. Gut 2019, 68, 1764–1773. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ma, Z.; Wang, R.; Wang, Y.; Wang, S.; Cheng, Z.; Xu, H.; Jin, X.; Li, W.; Wang, X. Thrombin facilitates invasion of ovarian cancer along peritoneum by inducing monocyte differentiation toward tumor-associated macrophage-like cells. Cancer Immunol Immunother 2010, 59, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Gonsiorek, W.; Fan, X.; Hesk, D.; Fossetta, J.; Qiu, H.; Jakway, J.; Billah, M.; Dwyer, M.; Chao, J.; Deno, G.; et al. Pharmacological characterization of Sch527123, a potent allosteric CXCR1/CXCR2 antagonist. J Pharmacol Exp Ther 2007, 322, 477–485. [Google Scholar] [CrossRef]
- Ginestier, C.; Liu, S.; Diebel, M.E.; Korkaya, H.; Luo, M.; Brown, M.; Wicinski, J.; Cabaud, O.; Charafe-Jauffret, E.; Birnbaum, D.; et al. CXCR1 blockade selectively targets human breast cancer stem cells in vitro and in xenografts. J Clin Invest 2010, 120, 485–497. [Google Scholar] [CrossRef]
- Brandolini, L.; Cristiano, L.; Fidoamore, A.; De Pizzol, M.; Di Giacomo, E.; Florio, T.M.; Confalone, G.; Galante, A.; Cinque, B.; Benedetti, E.; et al. Targeting CXCR1 on breast cancer stem cells: Signaling pathways and clinical application modelling. Oncotarget 2015, 6, 43375–43394. [Google Scholar] [CrossRef]
- Gulhati, P.; Schalck, A.; Jiang, S.; Shang, X.; Wu, C.J.; Hou, P.; Ruiz, S.H.; Soto, L.S.; Parra, E.; Ying, H.; et al. Targeting T cell checkpoints 41BB and LAG3 and myeloid cell CXCR1/CXCR2 results in antitumor immunity and durable response in pancreatic cancer. Nat Cancer 2023, 4, 62–80. [Google Scholar] [CrossRef]
- Teijeira, Á.; Garasa, S.; Gato, M.; Alfaro, C.; Migueliz, I.; Cirella, A.; de Andrea, C.; Ochoa, M.C.; Otano, I.; Etxeberria, I.; et al. CXCR1 and CXCR2 Chemokine Receptor Agonists Produced by Tumors Induce Neutrophil Extracellular Traps that Interfere with Immune Cytotoxicity. Immunity 2020, 52, 856–871. [Google Scholar] [CrossRef] [PubMed]
- Kargl, J.; Zhu, X.; Zhang, H.; Yang, G.H.Y.; Friesen, T.J.; Shipley, M.; Maeda, D.Y.; Zebala, J.A.; McKay-Fleisch, J.; Meredith, G.; et al. Neutrophil content predicts lymphocyte depletion and anti-PD1 treatment failure in NSCLC. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Suzuki, H.; Ohishi, T.; Asano, T.; Tanaka, T.; Yanaka, M.; Nakamura, T.; Yoshikawa, T.; Kawada, M.; Kaneko, M.K.; et al. Antitumor activities of a defucosylated anti-EpCAM monoclonal antibody in colorectal carcinoma xenograft models. Int J Mol Med 2023, 51. [Google Scholar] [CrossRef] [PubMed]
- Tateyama, N.; Nanamiya, R.; Ohishi, T.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Saito, M.; Asano, T.; Tanaka, T.; et al. Defucosylated Anti-Epidermal Growth Factor Receptor Monoclonal Antibody 134-mG(2a)-f Exerts Antitumor Activities in Mouse Xenograft Models of Dog Epidermal Growth Factor Receptor-Overexpressed Cells. Monoclon Antib Immunodiagn Immunother 2021, 40, 177–183. [Google Scholar] [CrossRef]
- Tanaka, T.; Suzuki, H.; Ohishi, T.; Kaneko, M.K.; Kato, Y. Antitumor activities against breast cancers by an afucosylated anti-HER2 monoclonal antibody H(2) Mab-77-mG(2a) -f. Cancer Sci 2023. [Google Scholar] [CrossRef]
Figure 1.
A schematic procedure of anti-mCXCR1 mAbs production. The procedure of Cell-Based Immunization and Screening (CBIS). LN229/mCXCR1 cells were immunized into an Sprague-Dawley rat using intraperitoneal injection. The culture supernatants of hybridoma were screened by flow cytometry. Cx1Mab-1 was established by limiting dilution.
Figure 1.
A schematic procedure of anti-mCXCR1 mAbs production. The procedure of Cell-Based Immunization and Screening (CBIS). LN229/mCXCR1 cells were immunized into an Sprague-Dawley rat using intraperitoneal injection. The culture supernatants of hybridoma were screened by flow cytometry. Cx1Mab-1 was established by limiting dilution.
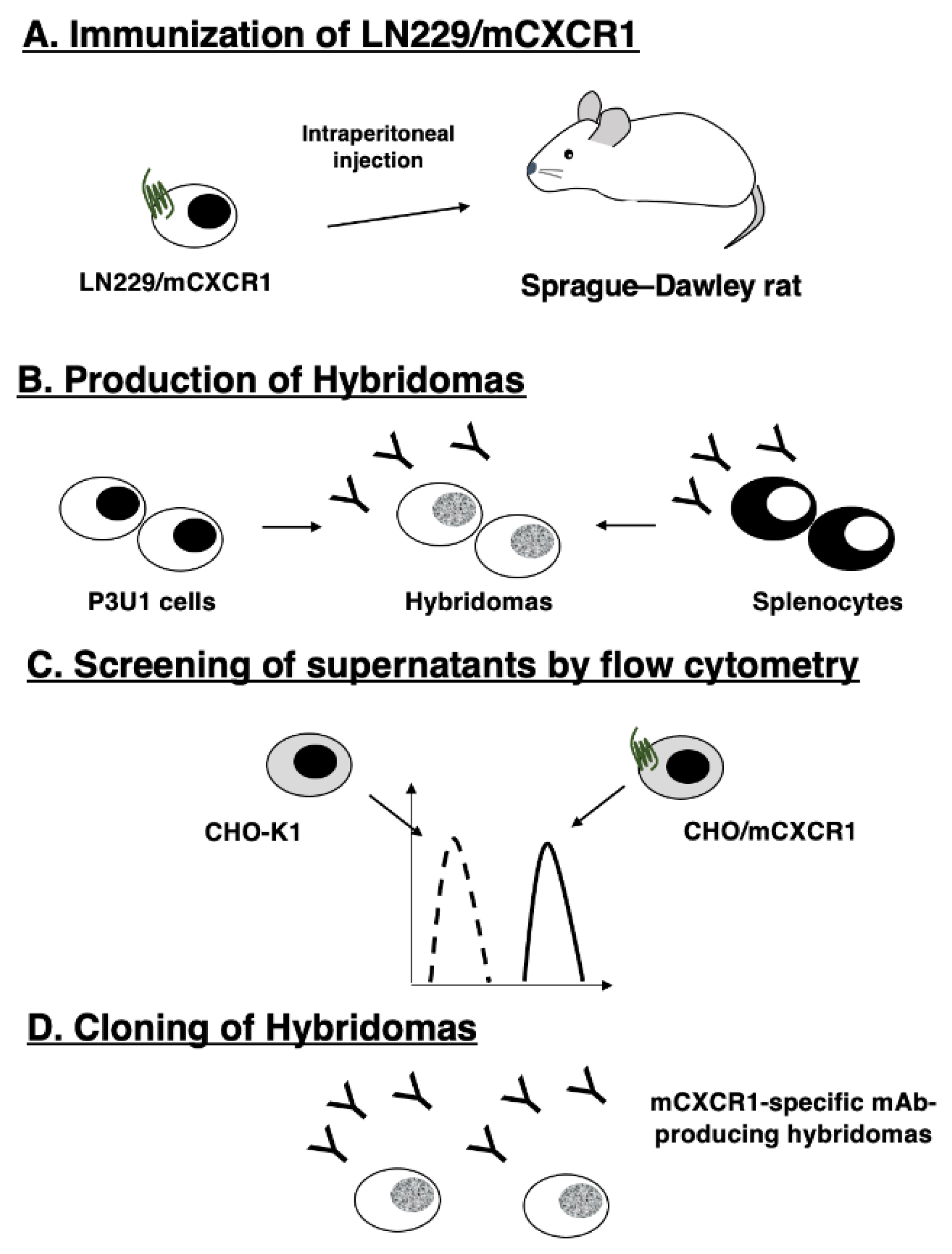
Figure 2.
Flow cytometric analysis of anti-mCXCR1 mAbs against CHO/mCXCR1 and CHO-K1. CHO/mCXCR1 (A) and CHO-K1 cells (B) were treated with 0.01–10 µg/mL of Cx1Mab-1 and 1122A, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG and Alexa Fluor 488-conjugated anti-rabbit IgG, respectively.
Figure 2.
Flow cytometric analysis of anti-mCXCR1 mAbs against CHO/mCXCR1 and CHO-K1. CHO/mCXCR1 (A) and CHO-K1 cells (B) were treated with 0.01–10 µg/mL of Cx1Mab-1 and 1122A, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG and Alexa Fluor 488-conjugated anti-rabbit IgG, respectively.
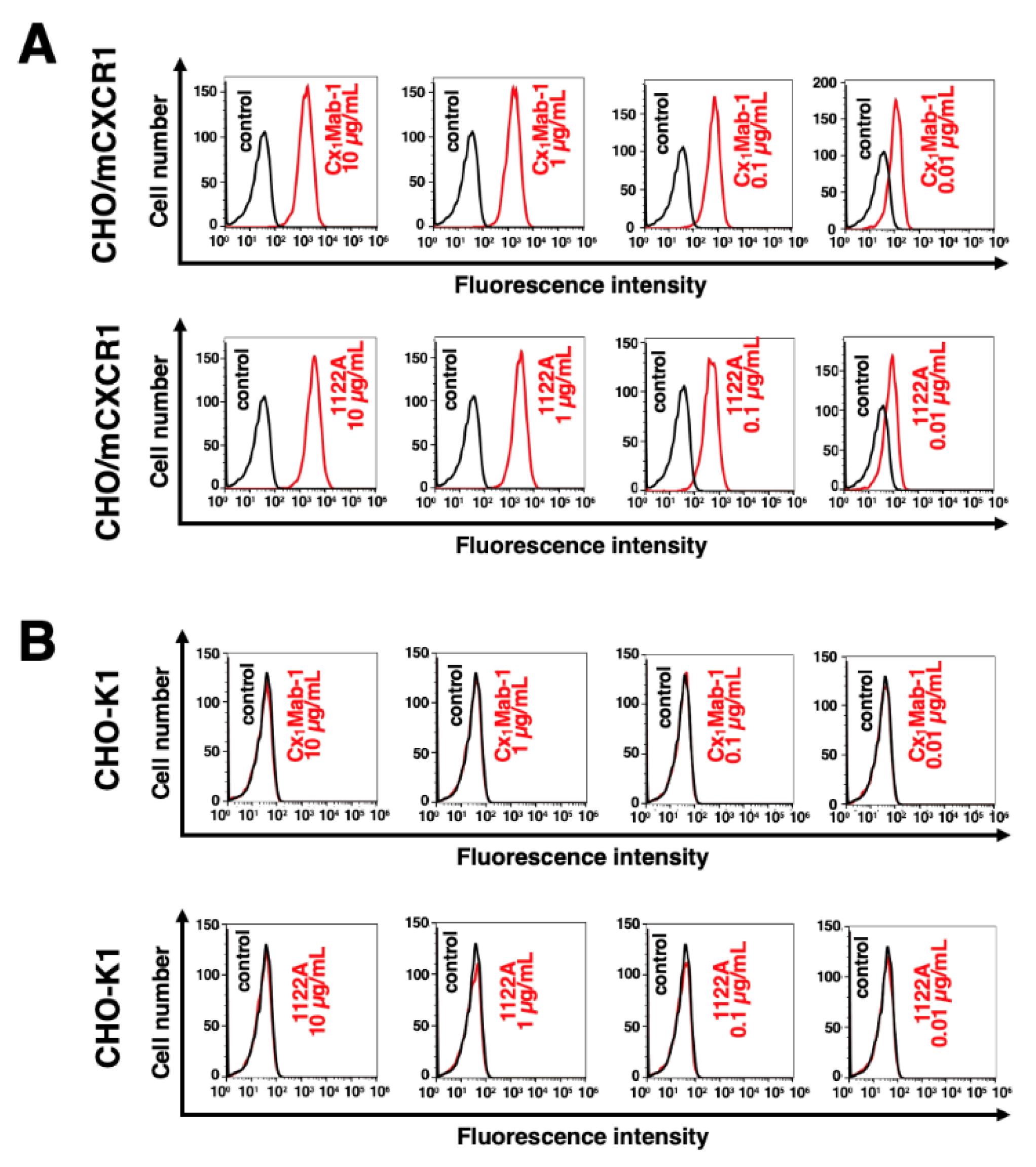
Figure 3.
Flow cytometric analysis of anti-mCXCR1 mAbs against LN229/mCXCR1 and LN229. LN229 cells (A), and LN229/mCXCR1 cells (B) were treated with 0.01–10 µg/mL of Cx1Mab-1 and 1122A, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG and Alexa Fluor 488-conjugated anti-rabbit IgG, respectively.
Figure 3.
Flow cytometric analysis of anti-mCXCR1 mAbs against LN229/mCXCR1 and LN229. LN229 cells (A), and LN229/mCXCR1 cells (B) were treated with 0.01–10 µg/mL of Cx1Mab-1 and 1122A, followed by treatment with Alexa Fluor 488-conjugated anti-rat IgG and Alexa Fluor 488-conjugated anti-rabbit IgG, respectively.
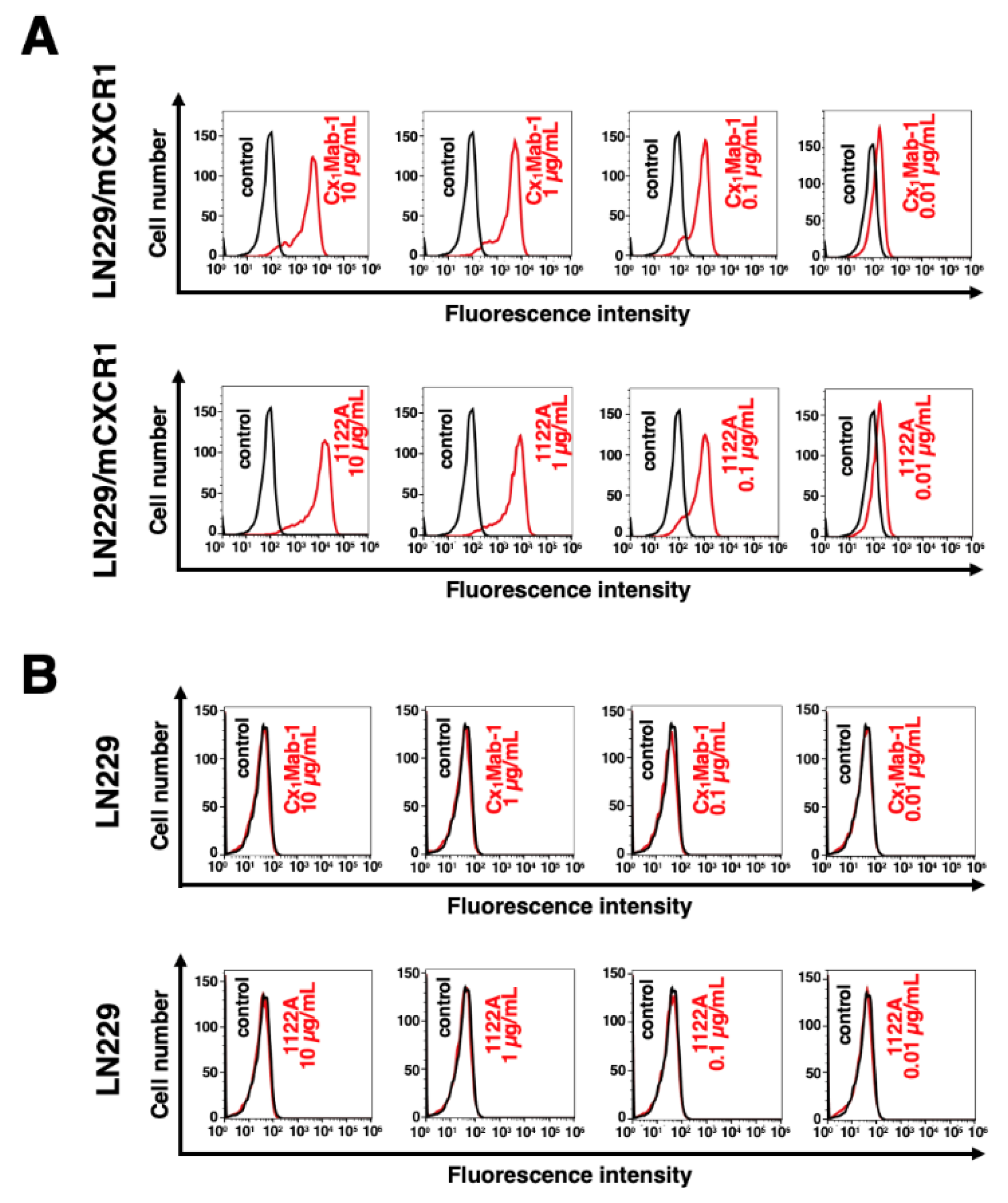
Figure 4.
The determination of the binding affinity of anti-mCXCR1 mAbs. CHO/mCXCR1 (A) or LN229/mCXCR1 (B) cells were suspended in 100 µL serially diluted Cx1Mab-1 or 1122A (25 to 0.006 µg/mL). Then, cells were treated with Alexa Fluor 488-conjugated anti-rat IgG or Alexa Fluor 488-conjugated anti-rabbit IgG. Fluorescence data were subsequently collected using a SA3800 Cell Analyzer, following the calculation of the dissociation constant (KD) by GraphPad PRISM 8.
Figure 4.
The determination of the binding affinity of anti-mCXCR1 mAbs. CHO/mCXCR1 (A) or LN229/mCXCR1 (B) cells were suspended in 100 µL serially diluted Cx1Mab-1 or 1122A (25 to 0.006 µg/mL). Then, cells were treated with Alexa Fluor 488-conjugated anti-rat IgG or Alexa Fluor 488-conjugated anti-rabbit IgG. Fluorescence data were subsequently collected using a SA3800 Cell Analyzer, following the calculation of the dissociation constant (KD) by GraphPad PRISM 8.
Figure 5.
Western blot analysis using anti-mCXCR1 mAbs. Cell lysates of CHO-K1, CHO/mCXCR1, LN229, and LN229/mCXCR1 cells were electrophoresed and proteins were transferred onto polyvinylidene difluoride membranes. After blocking, the membranes were incubated with 1 μg/mL of Cx1Mab-1, 1122A, anti-DYKDDDDK (1E6), or anti-β-actin mAb (AC-15), and then incubated with peroxidase-conjugated anti-rat IgG or anti-mouse IgG.
Figure 5.
Western blot analysis using anti-mCXCR1 mAbs. Cell lysates of CHO-K1, CHO/mCXCR1, LN229, and LN229/mCXCR1 cells were electrophoresed and proteins were transferred onto polyvinylidene difluoride membranes. After blocking, the membranes were incubated with 1 μg/mL of Cx1Mab-1, 1122A, anti-DYKDDDDK (1E6), or anti-β-actin mAb (AC-15), and then incubated with peroxidase-conjugated anti-rat IgG or anti-mouse IgG.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



