Submitted:
01 November 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
Casein Kinase ІІ (CK2) is one of the most versatile kinases. Its involvement in almost all cellular pathways makes it the Master Regulator of biochemical processes in a cell. This kinase is essential in regulating inflammation, cell differentiation, and cell cycle regulation. Often simultaneously. Its emerging role in senescence also indicates its function's centrality in cellular metabolism. The strategy to target this kinase to treat musculoskeletal disorders seems effective. These disorders often include a component of inflammation, dysregulated cell differentiation, and aging. This review focuses on CK2 target discovery and the diversity of its substrate interactions. We then transition toward the implication of CK2 in musculoskeletal disorders. Through a summary of current strategies for CK2 targeting with a new approach, we discuss the potential aspects that could be addressed in future drug research.
Keywords:
Casein Kinase II
; Musculoskeletal
; Osteoarthritis
; Osteoporosis
; CX-4945
; CIBG-300
; Cell Signaling
; Drug Development
; Targeted therapy
1. Introduction
Protein kinase CK2 (formerly called “Casein Kinase 2”) is a ubiquitous serine/threonine kinase. The literature describes its involvement as a lateral player in many biochemical pathways [1,2]. The ubiquitous presence and simultaneous precision of regulation make this kinase an outlier. With two homogenous regulatory β subunits and two heterogenous catalytic subunits α and α’, the kinase can identify and phosphorylate its vast range of substrates, estimated to be more than 400 today [3,4]. CK2 is ubiquitously present, expressed in almost all types of cells and is always active. Its mechanism of exerting regulation is different than other kinases which activate in response to particular stimuli [5,6,7].
The multiformity in mechanism of action of CK2 is essential for its myriad of functions. The factors which account to this property of the kinase are observed through studying the implication of CK2 in development and disease [5]. The factors are summarized here to explain the complexity CK2’s involvement in pathology. This is not an extensive list and with newer studies even more mechanisms are discovered. The kinase recognizes different types of substrates due to its β subunit which is not strictly a regulatory subunit. Requirement of the β subunit in only a part of CK2 substrate activation makes its mechanism of action enigmatic. The catalytic subunits α and α’ are almost same in structure but their involvement in catalytic activity is different. Each subunit has a set of substrates with which they interact individually. CK2 expression is under regulation of different growth factors and so is the expression of subunits. Also, the expression level of a single subunit affects expression and activity of other subunits. The localization of CK2 changes based on cell type and metabolic status. All these factors make targeting CK2 in diseases challenging [1,2,8]. However, efforts to unveil these complexities help design future therapeutics to overcome the past challenges. The implications of CK2 in diseases and disorders have been studied extensively. Expression and activity of CK2 is dysregulated in several diseases [1,2]. CK2 carries out signal transduction through – BMP signaling, Wnt signaling, Hedgehog signaling, ,PTEN/PI3K/Akt-PKB and NF-κB [9,10,11,12,13]. All these pathways have an interconnected regulation due to kinase like CK2 which functions as a “lateral player”. A pathological condition could arise if the function of the lateral player is disrupted [5].
‘Bone and joint’ diseases are traditionally described as disorders of respective physiological structures. However, the link between these musculoskeletal diseases and metabolic dysregulation was recently investigated. Bone fracture is the most common musculoskeletal disorder. The implication of diet-induced obesity in fracture healing was studied [14]. The relation between bone homeostasis and metabolic disorders in Osteoporosis (OP) was studied [15,16,17,18,19,20]. Osteoarthritis, small molecular metabolites were assessed for biomarker identification and targeting [21]. More studies linking OA to metabolic pathology are performed [22,23]. Rheumatoid arthritis (RA) is an autoimmune disorder affecting joints. The role of metabolism in RA was studied in relation to immune response [24,25]. Deviation in metabolic processes affects tissue homeostasis, and response to endocrine regulation is established [26]. The role of the inflammatory system and senescence in these diseases is also linked with metabolic dysregulation [27,28]. The role of CK2 in modulating biochemical pathways in all of these processes is very well established [29,30,31,32]. Thus, CK2 is an ideal druggable target, being a Master regulator. Correction of several different pathways is possible by targeting this kinase alone.
Targeting this kinase in musculoskeletal diseases is a lucrative strategy. Studies which explored the role of CK2 in bone differentiation and homeostasis help understand function of CK2. of CK2 was important for Osteoblastic differentiation through Ascorbic acid activation. It was activated in does dependent fashion with L-Ascorbic acid treatment and upregulated phosphorylation of transcription factor Ikaros. Ikaros is a bifunctional transcription factor for Osteoblast differentiation within the bone marrow niche[33]. Inhibition of CK2 is explored as a strategy to control loss of bone mass in relation to estrogen withdrawal [34].
The only clinically approved inhibitor of CK2 Silmitasertib (CX-4945) is a first-in-class small molecule of its kind [35]. It is an ATP-competitive inhibitor. Its use in cancer therapies is under investigation. However, like most currently available CK2 inhibitors, its mechanism of action is generalized rather than disease specific. Thus, controlling its effect on CK2 activity under different circumstances is impossible. Also, complete inhibition of catalytic activity of CK2 causes cytotoxicity. This is due to the cells' dependence on the basal level of phosphorylation of CK2 substrates for survival [7]. Cytotoxicity is not always desirable in treating diseases, especially in musculoskeletal disorders. The concept of modulating phosphorylation of a few CK2 substrates rather than broad-level blockade is emerging. This strategy will help take a pathology-tailored approach [36].
A peptide drug inhibitor of CK2 named CIGB300 is a promising candidate for the pathology-tailored approach. This synthetic peptide binds to the phsophoacceptor domain of CK2 [37]. It was discovered as an effective peptide in treating cervical cancer . The regulation of specific mediators was studied in Large Cell Lung Carcinoma (LCLC), Non-Small Cell Lung Cancer (NSCLC), advanced cervical cancer, Acute Myeloid Leukemia [38,39,40,41]. Due to its substrate-specific inhibition, it is highly effective and has mild side effects. This drug is under investigation in Phase 2 clinical trials, and studies about its effectiveness will soon be available [32]. The pathway-specific rather than global inhibitory effect of CIGB300 is thus emerging in cancer treatment.
For new drugs, the success rate of candidate molecules to move from bench to bedside is only 10%. This low rate exists despite the robustness of preclinical studies and excellent administration of clinical trials. A long processing time between drug discovery and commercial production may cause a candidate drug to be withdrawn despite being an excellent preclinical performer. Huge costs are involved with this process, and more importantly, the high failure rate delays the advances in better treatment for affected individuals. Lack of biomarker identification, off-target activity, and excess toxicity are a few of the challenges identified that can be addressed while designing newer candidates. This process can utilize advanced knowledge of the drug target's molecular interactions and biochemical activity [42].
For the development of specific inhibitors of CK2 for musculoskeletal disorders, understanding the crucial interactions of CK2 in disease pathology is essential. In this review, we discuss methods of CK2-target identification and specific functions of individual subunits. Later, we discuss interactions and related deregulation for the diseases of the musculoskeletal system. We then describe currently available inhibitors of CK2 and their mechanism of action. Finally, we discuss an example of CK2 activity modulation using synthetic peptides targeting specific interactions of CK2 within signal transduction cascade.
2. Targets of CK2
CK2 is known and studied for its role in various biological processes for nearly 60 years. Yet, its functions are discovered in the latest research, expanding the versatility of mechanisms it is known to undertake.
CK2 has two heterogenous alpha subunits, α and α’, and two homogenous β subunits. The enzyme functions as a holoenzyme, and each subunit can also work in isolation. Individual subunits of CK2 seem to play diverse roles exclusive to the holoenzyme form. The catalytic units of the kinase are acidophilic. Substrates often have acidic amino acids near the phosphorylation site. The consensus sequence on the phosphorylation target ([pS/pT]-{P}-x-[E/D] or [pS/pT]-{P}-x-pS is identified as the minimum identification sequence required for target identification [3,43]. However, due to the short length of this sequence and flexibility for amino acids in positions second and third, the frequency of occurrence of this sequence across all proteins is high. With the consensus prediction software PrositePlus, the number of eukaryotic targets comes out to be 3,000. The number of predicted targets was much higher than the actual targets [4]. Also, there is a possible overlap between other kinases for the phosphorylation of the same consensus sequence. Thus, target validation is important in determining the real substrates of CK2. It also helps target interactions where alternate kinases cannot nullify the drug's effect. Approaches like phospho-proteomics have helped determine the number of targets this kinase affects.
In this effort, Chojnowski et al. have developed a method for CK2 target validation. The co-substrate specificity of CK2 was taken into advantage for developing this technique. Identification of CK2 substrates from complex biological can be performed. The co-substrate activity of CK2 is that it can use either Adenosine triphosphate (ATP) or Guanosine triphosphate (GTP) to phosphorylate substrates. An analog of GTP - guanosine 5'-[γ-thio]triphosphate (GTPγS) was incorporated into the cell suspension. Other endogenous kinases do not use this substrate. Substrates phosphorylated with CK2 were identified by the presence of ‘PγS’ using Mass Spectrometry. Further, validation of CK2 targets with co-immunoprecipitation is possible with substrate-specific antibodies and pull-down assays. This method allows the use of versatile samples and helps capture phosphorylation reactions while the cells are still alive [5,44].
Being a highly pleiotropic kinase, it has numerous ways of regulating processes throughout the cell. One of the ways in which it functions as a pleiotropic kinase is by targeting substrates at different cellular localizations at different times during cellular processes. Targeting its kinase activity by inhibitors targeting the ATP binding site does not simultaneously alter the phosphorylation of all its targets. Changes in phosphorylation levels of CK2 targets two different CK2 inhibitors, CX4945 (Silmitasertib) and GO289, were different. Both inhibitors have very low off-target activity and have an almost overlapping set of affected CK2 phosphorylation targets. However, the extent of reduction of phosphorylation is very different from these two. Both inhibitors are structurally unrelated. Yet they bind the same site on CK2α and cause very different effects. The difference could be attributed to the ability of CK2 to identify its targets under different conditions [5,32].
For the identification of substrates that are non-redundant, CK2 inhibitor non-sensitive mutants were created. The kinase is known to phosphorylate only 20% of the substrates containing the minimal recognition motif. With the mutants and subsequent use of inhibitors, the expanse of substrates was estimated in cell lines. Studies using lysates do not consider the temporal and spatial factors for CK2-substrate interactions. Hence, these studies used the Stable Isotope Labeling Using Amino Acids in Cell culture (SILAC) approach. This allowed the study of interaction when the cells performed their normal function. The disparity was observed within popular CK2 inhibitors, with only a proportion of inhibitors showing dose-dependent inhibition of CK2 activity [45].
3. Subunits of CK2 and their diverse roles
There are many instances where individual subunits of CK2 perform distinct functions. Targeting those interactions during disease depends on individual subunit targeting rather than kinase activity inhibitors. Inhibition of a particular subunit or its depletion in cellular compartments affects the expression and activity of other subunits. These are explained using the following examples.
A coordinated regulation mechanism between the three holoenzyme subunits was observed during skeletal muscle differentiation. The mechanism involved regulating early Myogenic Regulator Genes such as MyoD and myogenin, followed by the expression of muscle-specific genes such as caveolin -3, troponin, and myomixer. The holoenzyme's individual subunits and catalytic activity also coordinated plasma membrane fusion and the shutting of embedded proteins. In individual knockouts of subunits in C2C12 cells with CRISPR/Cas9, the deletion of one subunit affected the expression of other subunits, showing their interdependence. Deletion of the CK2α subunit downregulated CK2β expression. Deletion of CK2β subunit upregulated CK2α expression. Deletion of CK2α’ did not alter the levels of the other two units. In knockouts of CK2α and CK2β, MyoD expression was reduced and abrogated, respectively, whereas the deletion of CK2α’ did not have an effect. The reduction of MyoD expression in CK2α deletion was attributed to the downregulation of CK2β in these mutants. In CK2β null mutants, the expression of this early MRF was not activated until the re-induction of the subunit in CK2β knockdown C2C12 cells already expressing low levels of MyoD. With increasing expression of CK2β in these cells, the level of MyoD increased, which was also reflected at transcription levels. This indicates the direct involvement of CK2β with other epigenetic modifiers for activating the expression of this early MRF. The level of MyoD expression was not affected by varying the catalytic activity of CK2 by pharmacological inhibition or changes in the expression of CK2α and CK2α’ subunits. During the later stages of myogenic differentiation, the levels of MyoD reduce, whereas the expression of muscle-specific genes increases in WT C2C12 cells. Here, pharmacological inhibition of kinase activity in CK2α’ knockout mutants reduced the expression of these markers, indicating the role of the CK2α subunit in the expression of muscle-specific markers. The role of CK2α’ during myotube formation was seen to be non-dispensable. With the deletion of this subunit, there was a reduction in the number and size of myotubes compared to WT. This subunit is also sufficient to rescue reduced fusogenic activity of muscle fibers [46]. The effects of kinase activity inhibition versus individual subunit knockout are depicted in Figure 1.
Also, by performing subunit knockout, the extent of the contribution of individual subunits was determined. This approach is helpful in the creation of high-specificity drugs. In most cases, the inhibition of kinase activity in a broad sense does not impact the function of single subunits. In these cases, focusing on a single subunit is critical in making the therapeutic precise and effective. In skeletal muscle development and homeostasis, a CK2α subunit is required [47].
The phosphoproteome of each subunit was studied with the help of catalytic and regulatory subunit knockout mutants using CRISPR/Cas9 technology. With the knockout of CK2α and α’ subunits, there was a substantial reduction in phosphorylation of predicted CK2 targets (18 out of 24). Expression of the CK2β subunit was reduced with this deletion. An increased rate of degradation of CK2β was a possible explanation for this observation. There was upregulation in phosphorylation in some of the non-CK2 phosphorylation targets. This could be explained by changes in the expression and activity of other kinases in the absence of CK2 catalytic subunits. For the knockout of CK2β subunits, there was a proportion of reduction of phosphorylation that was slightly lower than that of the catalytic units (9 out of 15). Here, the catalytic subunit expression was increased compared to the wild type. For some of the CK2 substrates, like CDC37 (pS13), the knockout of CK2 did not affect phosphorylation. But for the substrate AKT S129, the phosphorylation was drastically reduced to the extent it was almost absent with the knockout of either the catalytic or regulatory subunit. These differential effects on substrates shed light on the function of CK2 subunits as compensatory to each other in some cases or the strict requirements of holoenzymes in others. Changes in the phosphorylation status of predicted CK2 targets and non-targets based on sequence prediction indicated that only the presence of the consensus sequence is not sufficient or essential for target identification and phosphorylation to occur [48].
4. Inhibitors of CK2
The two most promising CK2 inhibitors include small molecule CX-4945 and peptide inhibitor CIBG-300. Both are under clinical investigation for their use in cancer treatment. There is a potential for developing CK2-targeting drugs for the treatment as well. The ATP-competitive CK2-specific small molecule inhibitor exploits the differences in this kinase's ATP-binding domain with others. The ATP-binding domain in the CK2α subunit is small compared to other kinases of this family.
Undesirable effects are seen while using CX-4945. The inhibition of Cdc2-like kinases (Clks) was due to off-target effects of CX-4945. It also affected alternating splicing of several genes, including CK2 [49]. In Cholangiocarcinoma, the drug induced methuosis and caused cell death in CK2 independent manner. These effects indicate the uncontrolled consequences of using generalized CK2 inhibitors [50]. The effectivity of CX4945 in cancer is also lower than a few other inhibitors of CK2, and it is ascribed to its unspecific action [51]. Recently a more specific molecule based on the structure of CX-4945 is getting developed [52]
The utility of CK2 inhibition as a strategy to target bone marrow stromal cells and the effect of the inhibition on other cell types generated from the same stem cell niche was studied. The effect of pharmacological inhibition of CK2 with CX-4945 on osteoblast and osteoclast viability was tested. It was found that for osteoblasts, cell viability was only slightly affected at higher concentrations. Only one-fifth of the concentration was required for osteoclast generation to reduce the viability 4. The off-target activity of this type of inhibitor is reported in literature [53]. Further studies are required for clinical translation. Other examples of ATP competitive inhibitors are included in Table 1 and Table 2, along with their effectiveness in clinical application.
Allosteric inhibitors of CK2 can bind to three different sites. Most effective inhibitors bind at the α and β unit interface. This approach retains the catalytic activity of the kinase and hence only affects the phosphorylation of CK2β dependent substrates [32]. The regulatory β subunits act as docking sites for a few CK2 substrates and protect the catalytic subunits from degradation. The other two sites where allosteric inhibitors can bind are the αD pocket, the linker region in the ATP binding site [73], and the αC helix region, which is rich in glycine. Examples of these are listed in Table 1 and Table 2. Detailed descriptions of various CK2 inhibitors and strategies they undertake can be found in current literature [76]. The structure-activity relation of CK2 inhibitors can also be found in literature [74,77]. Many of these are either not highly specific or have low cell permeability. Some compounds known to inhibit CK2 do not show cytotoxicity in cancer cell lines. However, their utility in treating diseases such as the musculoskeletal system is worth investigating. In these diseases, re-directing pathways involving deregulated CK2 is more critical than causing general cytotoxicity.
The synthetic peptide CIBG-300 was discovered for its effective treatment of Cervical cancer. It was also found to be anti-angiogenic [78]. The drug is derived from a cyclic nine-mer peptide. This peptide inhibited phosphorylation of human papillomavirus type-16 E7 (HPV-16 E7) oncoprotein phosphorylation in a phage display assay. Upon conjugating with cell-penetrating peptide Tat (amino acids 48-68) to increase the cell permeability of the peptide, it inhibited phosphorylation of the HPV-16 E7 oncoprotein by CK2 and induced apoptosis in cell cultures [37]. This peptide was then studied in different types of cancer. Substrates of CK2 interacted differently with CIGB-300. It reacted more with catalytic subunits and interacted less with the regulatory subunit. Also, it reacted with the CK2α’ more than the CK2α subunit [38]. The target of this peptide was B23/nucleophosmin, a nucleolar protein where that selectively modulated processes specific genes. It affected protein synthesis, energetic metabolism, and biogenesis of ribosomes [79]. These findings have led to a change in the defining mechanism of action of CIBG-300. Previously termed as CK2 inhibitor, its specificity in targeting selected phosphosites rather than generalized inhibition is getting attention.
It is an effective strategy to target CK2 with other kinases that are either phosphorylated by it or act on the same substrate. Here, BRD4 is a kinase that, along with CK2, is highly expressed in Breast Cancer and is phosphorylated by CK2. A dual inhibitor for C2 and BRD4 was more effective than targeting CK2 alone. Similarly, PIM kinases, along with CK2, interact with MYC. Pathways involved are non-redundant. In diseases with high expression of PIM and CK2, it was more effective to target both kinases. Kinase SRPK1 is involved in angiogenesis and overexpressed in several cancers. SRPIN803 was found to inhibit SRPK1 and CK2 together. Its ophthalmic application was also studied. TNIK and DYRK1are help maintain the phosphorylation of key regulators in cancer stem cells. Compound 108600 inhibited the TNIK, DYRK1, and CK2. But there was also an allosteric change in the CK2 α subunit to inhibit holoenzyme formation [77].

Table 2.
Multitarget inhibitors of CK2.
| Name | Status for Clinical Applica-tion | Reference |
|---|---|---|
| Compound 58 | Able to overcome drug resistance in cancer treatment. Targets CK2 and BRD4. | [79,80] |
| Compound 60 | Highly selective. Reduces tumor growth and lessens cancer symptoms in vivo and in vitro, with no apparent side effects. It is considered a potential therapeutic in triple-negative breast cancer. Targets CK2 and BRD4. Has potent and balanced activity against BRD4 and CK2. | [79,80] |
| Naphtho[2,1-b:7,6-b′]difuran-2,8-dicarboxylic acid hydrate (CPA), CPB, AMR | High selectivity for CK2 and the kinase PIM. Lack of cell permeability; hence, it cannot be used clinically. | [81] |
| 8-hydroxy-4-methyl-9-nitrobenzol(g)chrome-2-one (NBC) | High selectivity for CK2 and PIM. Induces apoptosis. | [69,82] |
| 1-β-D-2′-deoxyribofuranosyl-4,5,6,7- tetrabromo-1H-benzimidazole (TDB) | High selectivity for CK2 and PIM. Extremely high selectivity indicates it has clinical potential. | [83] |
| Compound 66 | Cytotoxic against cancer cells but not healthy cells. Inhibits the proliferation of various cancer cell lines. Reduces the viability of cancer cells more effectively than CX-4945. It is membrane-permeable and targets CK2 and PIM. | [82] |
| 6-(4-Hydroxy-3-methoxybenzylidene)-5-imino-2-(trifluoromethyl)-5H-[1,3,4]thiadiazolo[3,2-a]pyrimidin-7(6H)-one (SRPIN803; CK2 inhibitor XIII). | Inhibits both CK2 and SRPK1, which causes aberrant angiogenesis. Significantly inhibits cell viability in Jurkat cell lines. In vivo studies suggest it prevents the formation of intraocular neovascularization. | [84] |
| 108600 | Inhibitory effect on CK2/TNIK/DYRK1. The inhibitory effect on CK2α’ is ten times stronger than on CK2α. Inhibits tumor growth in breast cancer cells and overcomes chemical resistance. In vitro and in vivo studies suggest it is an optimal inhibitor in clinical settings. | [85] |
Inhibitors with a higher specificity can target specific subunits or forms of the CK2 holoenzyme. Some of them, like CIBG-300, are capable of inhibiting selective phosphosites. They are tested for their use in cancer treatment as well as treatment of other diseases. These are listed in Table 3.
5. Implication of CK2 in musculoskeletal disorders
5.1. Rheumatoid arthritis: Rheumatoid arthritis (RA) is an autoimmune condition that affects the joints. CK2 is overexpressed in RA, so inhibiting the kinase is a potential therapeutic strategy. Human primary CD4+T lymphocytes isolated from patients with RA were stimulated with the CK2 inhibitor CX-4945, which caused the suppression of cellular responses in Th1 and Th17 cells compared to controls but caused Th2 cellular activity to be enhanced. In vivo, studies using collagen-induced RA were conducted by treating the mice with CX-4945 via ingestion. Mice who had received CX-4945 were found to have inhibited Th1 and Th17 cell responses and subsequently reduced RA symptoms [89,90]. Primary Fibroblast-Like Synoviocytes (FLS) from patients diagnosed with RA showed higher levels of CK2 expression and activity than primary FLS patients diagnosed with OA. Stimulation of the RA primary cells with CX-4945 suppressed the cell cycle, which CK2 regulates. The inhibition of CK2 and the ensuing suppression of the cell cycle resulted in the decreased migration and proliferation of RA-FLSs. They attenuated the symptoms of RA, such as inflammation and joint pain The suppression of CK2 activity by CX-4945 has been shown through both in vivo and in vitro studies to be a novel therapeutic approach for RA. (Figure 2, 3, 4)
5.2. Osteoarthritis (OA) is a degenerative disease of the articular cartilage that becomes increasingly common with advanced age, especially for women. Specifically, OA is caused by the loss of number and activity of chondrocytes. Induction of apoptosis in chondrocytes is observed during OA. Reduced activity of chondrocytes causes increased breakdown of the collagen matrix and structural damage of the cartilage. CK2 is important for the inhibition of apoptosis. Parathyroid hormone-related protein (PTHrP) exerts its protective activity against mitochondria-dependent pro-apoptotic activity via CK2. Its nuclear localization causes increased expression of CK2 with increased nuclear retention and activity [91]. Expression of CK2 is reduced in chondrocytes of patients diagnosed with OA, and its inhibition sensitizes chondrocytes to TNFα mediated cell death [92]. CK2 is also involved in oxidative stress response. CK2 signaling is important for activating transcription factor NF-E2-related factor 2 (Nrf2), leading to the expression of Heme oxygenase-1 (HO-1). This enzyme is activated in chondrocytes in response to peroxynitrite-induced oxidative stress [93]. Also, inhibition of catalytic activity of CK2 with 4,5,6,7-terabromo-2-azabenzimidazole (TBB), 5,6-dichlorobenzimidazole 1-β-D-ribofuranoside (DRB) induced senescence and apoptosis in chondrocytes. Overexpression of HO-1 reduced the TBB-induced senescence. Chondrocytes overexpressing HO-1 had reduced sensitivity towards TBB-induced senescence. Knockdown of CK2 led to reduced type II collagen and increased β catenin expression [94]. Chondrocytes treated with CK2 inhibitors and αB-crystallin siRNA were sensitized for apoptosis. However, αB-crystallin had a protective effect. Its expression was modulated, and cellular localization was changed after CK2 inhibition [92]. The role of αB-crystallin in aiding CK2 to prevent chondrocyte apoptosis should be explored further. Thus, CK2 protects chondrocytes from the effects of oxidative stress and apoptosis.
5.3. Bone fractures: Often associated with osteoporosis, bone fractures can be induced or aggravated by various conditions, often relating to a weakening of bone or a loss in bone mass. Casein Kinase Interacting Protein-1 (CKIP-1) interacts with CK2α subunit and regulates signaling between CK2α and effector molecules of BMP signaling [95,96]. The CKIP-1 knockout mice had abnormally high levels of bone mass. The effects of CKIp-1 on bone growth and repair were age-dependent, with CKIp-1 having a more substantial effect on the bone mass of 18-month-old mice and comparatively little effect on 2-month-old mice. CKIp-1 is a negative regulator of osteogenic differentiation, and age-related inflammation causes an upregulation of CKIp-1, although the exact mechanisms of this upregulation remain unclear [97,98]. Inhibition of interaction between CK2α and CKIp-1 is a possible therapeutic strategy for bone fractures.
5.4. Osteoporosis (OP) is a degenerative disease that, as of 2023, affects more than 200 million people. Incidence rate of OP is 1.7% of men and 26% of women over age 50 worldwide [99]. OP is associated with loss in bone mass and mineral density, which weakens the bones and predisposes them to fractures. Glucocorticoid-induced osteoporosis (GIO) is a form of osteoporosis in patients undergoing treatment for various inflammatory or auto-immune diseases via glucocorticoid medications. Primary cells from human patients and mice with GIO were found to have elevated levels of Casein kinase 2-interacting protein 1 (CKIP-1) and reduced levels of Smad1/5 compared to controls. In vitro, elevated expression and activity of CKIp-1 in osteoblasts inhibited Smad-dependent BMP signaling. CKIp-1 regulates BMP signaling and can recruit CK2α to the plasma membrane. Hence, its overexpression can affect the activation of BMP signaling of Osteoblasts and Osteoclasts. It increased the ubiquitination of Smad1 and suppressed BMP signaling. This affected the Differentiation of MSCs into Osteoblasts. These alterations could have contributed to lesser mineralization. Suppressing CKIp-1 interaction with CK2 could be a novel therapeutic strategy for treating GIO. It would help promote BMP signaling and thereby induce bone formation and repair [100]. A challenge in creating therapeutics for osteoporosis is maintaining the optimal balance between osteoblasts and osteoclasts, cells that create bone and cells that degrade bone, respectively. Ideally, a therapeutic for OP would increase osteoblast activity while decreasing osteoclast activity, inducing bone formation and repair.
6. Implications of CK2 in Musculoskeletal Cancers
6.1. Rhabdomyosarcoma is a cancer initiated in the skeletal muscle and can metastasize to other tissues. CK2 is found to be upregulated in rhabdomyosarcoma cells, so the inhibition of CK2 and the subsequent promotion of apoptosis among tumor cells is a primary goal of research. Research into possible therapeutics has included using the cloned human tumor cell lines JR1 and Rh30 cells. When these cells were stimulated with the CK2 inhibitor 5,6-dichlorobenzimidazole (DRB), the proliferation of pro-apoptotic signals commenced, such as cytochrome c and Smad/DIABLO, which predictably led to widespread cell death. JR1 and Rh30 cells transfected with either short hairpin RNA targeted to CK2α or kinase-inactive CK2aα/CK2α’ led to heightened sensitivity to pro-apoptotic signals, suggesting that they further suppressed CK2 activity. Inhibitors of CK2 have the potential as therapeutics to prevent the growth and metastasis of rhabdomyosarcoma cells [101]. Here, inhibition of kinase activity and knockdown of α/α’ subunit increased apoptosis (Figure 5).
6.2. Osteosarcoma is a bone malignancy originating in the osteoblasts. CK2 is overexpressed in osteosarcoma, with CK2α and CK2β being upregulated in cloned human osteosarcoma cells, compared to healthy osteoblasts and mesenchymal stem cells. The osteosarcoma cells were cultured and stimulated with the CK2 inhibitor CX-4945, which was found to suppress the growth and proliferation of the tumor cells and increase the apoptosis rate. Interestingly, the proliferation and differentiation of mesenchymal stem cells was unaffected. The siRNA-induced knockdown of the CK2α and CK2β subunits in vitro suppressed the growth and proliferation of osteosarcoma cells. In vivo studies included the xenografting of 143B osteosarcoma cells into mice, which were then treated with CX-4945. Like the in vitro findings, CX-4945 inhibited the proliferation of the tumor cells and, therefore, constrained the growth of the tumor. CX-4945 has potential as a therapeutic for osteosarcoma because its inhibition of CK2 induces the apoptosis of sarcoma cells while having little to no effect on mesenchymal stem cells [102].
7. Targeting CK2-Substrate interaction
One of the most integral ways in which CK2 regulates the function of the musculoskeletal system is through the Bone Morphogenetic Protein (BMP) Signaling Pathway. The BMP signaling pathway regulates chondrogenesis and osteogenesis and is significant in treating various musculoskeletal conditions. More specifically, CK2 interacts with the BMPRIa-BMPRII receptor complex in a manner that mimics that of the growth factor BMP2. The binding of factor BMP2 to the receptor complex causes the release of CK2 from three specific locations on the receptor. The release of CK2 initiates the subsequent phosphorylation of downstream signaling, which causes the differentiation of mesenchymal stem cells into adipocytes, osteoblasts, and chondrocytes, depending on the pathway initiated. However, initiating the signaling cascade via BMP2 induces various harmful side effects; therefore, finding a similar ligand that does not facilitate such consequences is optimal [103]. Three mimetic peptides were synthesized to initiate the BMP signaling pathway without BMP2. These peptides contained one of three CK2 phosphorylation sites on BMPRIa (SLKD, SYED, or SLYD). At the N-terminus, there is the Antennapedia Homeodomain sequence for cellular uptake. Flanking the CK2 phosphorylation motif, there is a BMPRIa homologous sequence. These peptides are named CK2.1, CK2.2, and CK2.3 based on the phosphorylation site they contain [9]. CK2.1 was found to initiate chondrogenesis both in vivo and in vitro in mouse models, suggesting it has potential as a therapeutic for OA and other conditions characterized by cartilage degradation. CK2.1 increased proteoglycan synthesis and elevated levels of collagen type II without causing chondrocyte hypertrophy, a frequent occurrence with BMP2-induced signaling[104,105]. Rabbit mesenchymal stem cells were treated with a bilayer peptide-loaded scaffold consisting of CK2.1 coated β-glycerophosphate/chitosan, which resulted in chondrogenesis and Osteoblastogenesis. This technique could be beneficial for treating articular osteochondral defects, common in OA patients, involving damage to both the cartilage and underlying bone [100]. CK2.2 induced both adipogenesis and Osteogenesis, while CK2.3 induced Osteogenesis. In treating OP and other disorders that reduce bone mass and mineral density, CK2.3 would be preferred because its use does not simultaneously induce adipogenesis, as does CK2.2 [105]. CK2.3 has been tested on mouse models in vitro and in vivo and activates ERK phosphorylation by preventing CK2 binding to the BMRPIa-BMPRII receptor complex. CK2.3 induces greater bone area, bone mass, and mineral density [103]. C2C12 cloned mouse myoblast cells were treated with the synthetic peptide CK2.3 and were found to have decreased Osteoclastogenesis, increased osteoblastogenesis, and increased mineralization compared to controls. In vivo, injections of CK2.3 via calvarial injection in mice increased bone area, density, and growth [9]. In vivo, studies of 6 to 9-day-old C57BL/6J mice injected with CK2.3 via the calvaria and 8-week-0old mice injected via the tail vein found that the mice showed increased bone formation and increased bone mineral density. At the same time, osteoclast activity appeared to be suppressed, indicating CK2.3 has the potential to alleviate the symptoms of OP by simultaneously enhancing osteoblast activity and inhibiting osteoclast activity, inducing bone growth and repair [106,107,108].
8. Discussion
The evolution of small molecular inhibitors of CK2 is focused on improving specificity and reducing general toxicity. Improved cell permeability is also an essential factor. The specificity concerns inhibiting kinase activity of CK2 catalytic activity, and the inhibitors are primarily tested for their anti-oncogenic activity. These inhibitors are commonly used in combination with targeted therapy. However, their use in other diseases is not common. Also, targeting the kinase activity is not always the most effective method. The entire phosphor-kinome of CK2 is still undergoing experimental validation. Targeting of redundant interactions and false positive substrates hence stands as a possibility. The peptide drugs target the identified interaction between CK2 and BMPRIa. These are also seen to be highly cell-permeable. More investigation into their tissue-specific drug delivery and toxicity needs to be done. Evidence of their ability to drive the differentiation of mesenchymal stem cells into different lineages opens many possibilities. Drugs highly specific to kinase substrate interaction can potentially act as a molecular engineering tool. It can direct the molecular pathways towards the desired lineage. Targeting the pleiotropic kinase CK2 in this manner is an emerging prospect for designing a new generation of drugs.
The evolution of small molecular inhibitors of CK2 is focused on improving specificity and reducing general toxicity. Improved cell permeability is also an essential factor. The specificity concerns inhibiting kinase activity of CK2 catalytic activity, and the inhibitors are primarily tested for their anti-oncogenic activity. These inhibitors are commonly used in combination with targeted therapy. However, their use in other diseases is not common. Also, targeting the kinase activity is not always the most effective method. The entire phosphor-kinome of CK2 is still undergoing experimental validation. Targeting of redundant interactions and false positive substrates hence stands as a possibility. The peptide drugs target the identified interaction between CK2 and BMPRIa. These are also seen to be highly cell-permeable. More investigation into their tissue-specific drug delivery and toxicity needs to be done. Evidence of their ability to drive the differentiation of mesenchymal stem cells into different lineages opens many possibilities. Drugs highly specific to kinase substrate interaction can potentially act as a molecular engineering tool. It can direct the molecular pathways towards the desired lineage. Targeting the pleiotropic kinase CK2 in this manner is an emerging prospect for designing a new generation of drugs.
Author Contributions
Conceptualization, V.P. and A.N.; methodology, V.P.; software, V.P.; validation, A.N., V.P. and K.D.; formal analysis, V.P.; investigation, V.P.; resources, A.N.; data curation, V.P., K.D.; writing—original draft preparation, V.P.; writing—review and editing, A.N., V. P., K.D.; visualization, V.P.; supervision, A.N.; project administration, A.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not Applicable
Informed Consent Statement
Not Applicable
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors would like to acknowledge all members of the Nohe lab, the University of Delaware, and the Delaware Biotechnology Institute for their continued support during this project. The authors thank Dr. Daniel Halloran and Kelechi Chukwuocha for their immense help and valuable suggestions in writing this review.
Conflicts of Interest
Declare conflicts of interest or state “The authors declare no conflict of interest.” Authors must identify and declare any personal circumstances or interest that may be perceived as inappropriately influencing the representation or interpretation of reported research results. Any role of the funders in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; or in the decision to publish the results must be declared in this section. If there is no role, please state “The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Halloran, D.; Pandit, V.; Nohe, A. The Role of Protein Kinase CK2 in Development and Disease Progression: A Critical Review. J Dev Biol 2022, 10. [Google Scholar] [CrossRef]
- Borgo, C.; D’Amore, C.; Sarno, S.; Salvi, M.; Ruzzene, M. Protein Kinase CK2: A Potential Therapeutic Target for Diverse Human Diseases. Signal Transduct Target Ther 05 17, 6, 183. [CrossRef] [PubMed]
- Meggio, F.; Pinna, L.A. One-Thousand-and-One Substrates of Protein Kinase CK2? FASEB J 2003, 17, 349–368. [Google Scholar] [CrossRef] [PubMed]
- Franchin, C.; Borgo, C.; Zaramella, S.; Cesaro, L.; Arrigoni, G.; Salvi, M.; Pinna, L.A. Exploring the CK2 Paradox: Restless, Dangerous, Dispensable. Pharmaceuticals (Basel) 2017, 10. [Google Scholar] [CrossRef]
- Borgo, C.; D’Amore, C.; Cesaro, L.; Sarno, S.; Pinna, L.A.; Ruzzene, M.; Salvi, M. How Can a Traffic Light Properly Work If It Is Always Green? The Paradox of CK2 Signaling. Crit Rev Biochem Mol Biol 2021, 56, 321–359. [Google Scholar] [CrossRef]
- Pinna, L.A. Protein Kinase CK2: A Challenge to Canons. J Cell Sci 2002, 115, 3873–3878. [Google Scholar] [CrossRef] [PubMed]
- Pinna, L.A. Protein Kinase CK2. The International Journal of Biochemistry & Cell Biology 1997, 29, 551–554. [Google Scholar] [CrossRef]
- Chua, M.; Ortega, C.; Sheikh, A.; Lee, M.; Abdul-Rassoul, H.; Hartshorn, K.; Dominguez, I. CK2 in Cancer: Cellular and Biochemical Mechanisms and Potential Therapeutic Target. Pharmaceuticals 2017, 10, 18. [Google Scholar] [CrossRef]
- Bragdon, B.; Thinakaran, S.; Moseychuk, O.; Gurski, L.; Bonor, J.; Price, C.; Wang, L.; Beamer, W.G.; Nohe, A. Casein Kinase 2 Regulates in Vivo Bone Formation through Its Interaction with Bone Morphogenetic Protein Receptor Type Ia. Bone 2011, 49, 944–954. [Google Scholar] [CrossRef]
- Jia, H.; Liu, Y.; Xia, R.; Tong, C.; Yue, T.; Jiang, J.; Jia, J. Casein Kinase 2 Promotes Hedgehog Signaling by Regulating Both Smoothened and Cubitus Interruptus. Journal of Biological Chemistry 2010, 285, 37218–37226. [Google Scholar] [CrossRef]
- Dominguez, I.; Sonenshein, G.E.; Seldin, D.C. Protein Kinase CK2 in Health and Disease: CK2 and Its Role in Wnt and NF-κB Signaling: Linking Development and Cancer. Cell. Mol. Life Sci. 2009, 66, 1850–1857. [Google Scholar] [CrossRef]
- Torres, J.; Pulido, R. The Tumor Suppressor PTEN Is Phosphorylated by the Protein Kinase CK2 at Its C Terminus. Journal of Biological Chemistry 2001, 276, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Qin, H.; Frank, S.J.; Deng, L.; Litchfield, D.W.; Tefferi, A.; Pardanani, A.; Lin, F.-T.; Li, J.; Sha, B.; et al. A CK2-Dependent Mechanism for Activation of the JAK-STAT Signaling Pathway. Blood 2011, 118, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Khajuria, D.K.; Reider, I.; Kamal, F.; Norbury, C.C.; Elbarbary, R.A. Distinct Defects in Early Innate and Late Adaptive Immune Responses Typify Impaired Fracture Healing in Diet-Induced Obesity. Front Immunol 2023, 14, 1250309. [Google Scholar] [CrossRef]
- Xi, Y.; Shen, J.; Li, X.; Bao, Y.; Zhao, T.; Li, B.; Zhang, X.; Wang, J.; Gao, J.; Xie, Z.; et al. Regulatory Effects of Quercetin on Bone Homeostasis: Research Updates and Future Perspectives. Am J Chin Med 2023, 1–18. [Google Scholar] [CrossRef]
- Robin, F.; Chappard, D.; Leroyer, P.; Latour, C.; Mabilleau, G.; Monbet, V.; Cavey, T.; Horeau, M.; Derbré, F.; Roth, M.P.; et al. Differences in Bone Microarchitecture between Genetic and Secondary Iron-Overload Mouse Models Suggest a Role for Hepcidin Deficiency in Iron-Related Osteoporosis. FASEB J 2023, 37, e23245. [Google Scholar] [CrossRef]
- Alghadir, A.H.; Gabr, S.A.; Iqbal, A. Hand Grip Strength, Vitamin D Status, and Diets as Predictors of Bone Health in 6-12 Years Old School Children. BMC Musculoskelet Disord 2023, 24, 830. [Google Scholar] [CrossRef]
- Ribeiro, M.S.P.; Venturini, L.G.R.; Speck-Hernandez, C.A.; Alabarse, P.V.G.; Xavier, T.; Taira, T.M.; Rodrigues, L.F.D.; Cunha, F.Q.; Fukada, S.Y. AMPKα1 Negatively Regulates Osteoclastogenesis and Mitigate Pathological Bone Loss. J Biol Chem 2023, 105379. [Google Scholar] [CrossRef]
- Zhu, Z.; Liu, M.; Zhang, Y.; Wu, J.; Gao, M.; Lei, T.; Huang, F.; Chen, H.; Wu, M. Risk Factors for the Comorbidity of Osteoporosis/Osteopenia and Kidney Stones: A Cross-Sectional Study. Arch Osteoporos 2023, 18, 128. [Google Scholar] [CrossRef]
- Yoon, H.; Sung, E.; Kang, J.H.; Kim, C.H.; Shin, H.; Yoo, E.; Kim, M.; Lee, M.Y.; Shin, S. Association between Body Fat and Bone Mineral Density in Korean Adults: A Cohort Study. Sci Rep 2023, 13, 17462. [Google Scholar] [CrossRef]
- Liao, Z.; Han, X.; Wang, Y.; Shi, J.; Zhang, Y.; Zhao, H.; Zhang, L.; Jiang, M.; Liu, M. Differential Metabolites in Osteoarthritis: A Systematic Review and Meta-Analysis. Nutrients 2023, 15. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Rayman, M.P.; Gualillo, O.; Sellam, J.; van der Kraan, P.; Fearon, U. The Role of Metabolism in the Pathogenesis of Osteoarthritis. Nat Rev Rheumatol 2017, 13, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Jeong, S.; Kim, H.; Kang, D.; Lee, J.; Kang, S.B.; Kim, J.H. Disease-Modifying Therapeutic Strategies in Osteoarthritis: Current Status and Future Directions. Exp Mol Med 2021, 53, 1689–1696. [Google Scholar] [CrossRef] [PubMed]
- Comertpay, B.; Gov, E. Immune Cell-Specific and Common Molecular Signatures in Rheumatoid Arthritis through Molecular Network Approaches. Biosystems 2023, 234, 105063. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, S.; Cao, N.; Wang, Q.; Liu, Y.; Xu, Q.; Zhang, L.; Sun, C.; Xiao, X.; Yao, J. Intestinal Flora, Intestinal Metabolism, and Intestinal Immunity Changes in Complete Freud’s Adjuvant-Rheumatoid Arthritis C57BL/6 Mice. Int Immunopharmacol 2023, 125, 111090. [Google Scholar] [CrossRef]
- Rea, I.M.; Gibson, D.S.; McGilligan, V.; McNerlan, S.E.; Alexander, H.D.; Ross, O.A. Age and Age-Related Diseases: Role of Inflammation Triggers and Cytokines. Front Immunol 2018, 9, 586. [Google Scholar] [CrossRef] [PubMed]
- McGarry, T.; Biniecka, M.; Veale, D.J.; Fearon, U. Hypoxia, Oxidative Stress and Inflammation. Free Radic Biol Med 2018, 125, 15–24. [Google Scholar] [CrossRef]
- Song, J.; Bae, Y.S. CK2 Down-Regulation Increases the Expression of Senescence-Associated Secretory Phenotype Factors through NF-κB Activation. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Pagano, M.A.; Andrzejewska, M.; Ruzzene, M.; Sarno, S.; Cesaro, L.; Bain, J.; Elliott, M.; Meggio, F.; Kazimierczuk, Z.; Pinna, L.A. Optimization of Protein Kinase CK2 Inhibitors Derived from 4,5,6,7-Tetrabromobenzimidazole. J Med Chem 2004, 47, 6239–6247. [Google Scholar] [CrossRef]
- Sarno, S.; Pinna, L.A. Protein Kinase CK2 as a Druggable Target. Mol Biosyst 2008, 4, 889–894. [Google Scholar] [CrossRef]
- Trembley, J.H.; Chen, Z.; Unger, G.; Slaton, J.; Kren, B.T.; Van Waes, C.; Ahmed, K. Emergence of Protein Kinase CK2 as a Key Target in Cancer Therapy. Biofactors 2010, 36, 187–195. [Google Scholar] [CrossRef]
- Borgo, C.; Ruzzene, M. Protein Kinase CK2 Inhibition as a Pharmacological Strategy. Adv Protein Chem Struct Biol 2021, 124, 23–46. [Google Scholar] [CrossRef]
- Son, E.; Do, H.; Joo, H.-M.; Pyo, S. Induction of Alkaline Phosphatase Activity by L-Ascorbic Acid in Human Osteoblastic Cells: A Potential Role for CK2 and Ikaros. Nutrition 2007, 23, 745–753. [Google Scholar] [CrossRef]
- McCarty, M.F.; Lewis Lujan, L.; Iloki Assanga, S. Targeting Sirt1, AMPK, Nrf2, CK2, and Soluble Guanylate Cyclase with Nutraceuticals: A Practical Strategy for Preserving Bone Mass. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Pierre, F.; Chua, P.C.; O’Brien, S.E.; Siddiqui-Jain, A.; Bourbon, P.; Haddach, M.; Michaux, J.; Nagasawa, J.; Schwaebe, M.K.; Stefan, E.; et al. Discovery and SAR of 5-(3-Chlorophenylamino)Benzo[c][2,6]Naphthyridine-8-Carboxylic Acid (CX-4945), the First Clinical Stage Inhibitor of Protein Kinase CK2 for the Treatment of Cancer. J Med Chem 2011, 54, 635–654. [Google Scholar] [CrossRef]
- Zanin, S.; Sandre, M.; Cozza, G.; Ottaviani, D.; Marin, O.; Pinna, L.A.; Ruzzene, M. Chimeric Peptides as Modulators of CK2-Dependent Signaling: Mechanism of Action and off-Target Effects. Biochim Biophys Acta 2015, 1854, 1694–1707. [Google Scholar] [CrossRef] [PubMed]
- Perea, S.E.; Baladrón, I.; Valenzuela, C.; Perera, Y. CIGB-300: A Peptide-Based Drug That Impairs the Protein Kinase CK2-Mediated Phosphorylation. Semin Oncol 2018, 45, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Pérez, G.V.; Rosales, M.; Ramón, A.C.; Rodríguez-Ulloa, A.; Besada, V.; González, L.J.; Aguilar, D.; Vázquez-Blomquist, D.; Falcón, V.; Caballero, E.; et al. CIGB-300 Anticancer Peptide Differentially Interacts with CK2 Subunits and Regulates Specific Signaling Mediators in a Highly Sensitive Large Cell Lung Carcinoma Cell Model. Biomedicines 2022, 11. [Google Scholar] [CrossRef]
- Cirigliano, S.M.; Díaz Bessone, M.I.; Berardi, D.E.; Flumian, C.; Bal de Kier Joffé, E.D.; Perea, S.E.; Farina, H.G.; Todaro, L.B.; Urtreger, A.J. The Synthetic Peptide CIGB-300 Modulates CK2-Dependent Signaling Pathways Affecting the Survival and Chemoresistance of Non-Small Cell Lung Cancer Cell Lines. Cancer Cell Int 2017, 17, 42. [Google Scholar] [CrossRef]
- Sarduy, M.R.; García, I.; Coca, M.A.; Perera, A.; Torres, L.A.; Valenzuela, C.M.; Baladrón, I.; Solares, M.; Reyes, V.; Hernández, I.; et al. Optimizing CIGB-300 Intralesional Delivery in Locally Advanced Cervical Cancer. Br J Cancer 2015, 112, 1636–1643. [Google Scholar] [CrossRef]
- Vázquez-Blomquist, D.; Ramón, A.C.; Rosales, M.; Pérez, G.V.; Rosales, A.; Palenzuela, D.; Perera, Y.; Perea, S.E. Gene Expression Profiling Unveils the Temporal Dynamics of CIGB-300-Regulated Transcriptome in AML Cell Lines. BMC Genomics 2023, 24, 373. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Gao, W.; Hu, H.; Zhou, S. Why 90% of Clinical Drug Development Fails and How to Improve It? Acta Pharm Sin B 2022, 12, 3049–3062. [Google Scholar] [CrossRef] [PubMed]
- Mauro Salvi; Sarno, S.; Cesaro, L.; Nakamura, H.; Pinna, L.A. Extraordinary Pleiotropy of Protein Kinase CK2 Revealed by Weblogo Phosphoproteome Analysis. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 2009, 1793, 847–859. [CrossRef]
- Chojnowski, J.E.; McMillan, E.A.; Strochlic, T.I. Identification of Novel CK2 Kinase Substrates Using a Versatile Biochemical Approach. Journal of visualized experiments 2019. [Google Scholar] [CrossRef]
- Gyenis, L.; Menyhart, D.; Cruise, E.S.; Jurcic, K.; Roffey, S.E.; Chai, D.B.; Trifoi, F.; Fess, S.R.; Desormeaux, P.J.; Núñez de Villavicencio Díaz, T.; et al. Chemical Genetic Validation of CSNK2 Substrates Using an Inhibitor-Resistant Mutant in Combination with Triple SILAC Quantitative Phosphoproteomics. Front Mol Biosci 2022, 9, 909711. [Google Scholar] [CrossRef]
- Salizzato, V.; Zanin, S.; Borgo, C.; Lidron, E.; Salvi, M.; Rizzuto, R.; Pallafacchina, G.; Donella-Deana, A. Protein Kinase CK2 Subunits Exert Specific and Coordinated Functions in Skeletal Muscle Differentiation and Fusogenic Activity. FASEB J 10AD, 33, 10648–10667. [CrossRef]
- Merholz, M.; Jian, Y.; Wimberg, J.; Gessler, L.; Hashemolhosseini, S. In Skeletal Muscle Fibers, Protein Kinase Subunit CSNK2A1/CK2α Is Required for Proper Muscle Homeostasis and Structure and Function of Neuromuscular Junctions. Cells 2022, 11. [Google Scholar] [CrossRef]
- Franchin, C.; Borgo, C.; Cesaro, L.; Zaramella, S.; Vilardell, J.; Salvi, M.; Arrigoni, G.; Pinna, L.A. Re-Evaluation of Protein Kinase CK2 Pleiotropy: New Insights Provided by a Phosphoproteomics Analysis of CK2 Knockout Cells. Cell Mol Life Sci 2018, 75, 2011–2026. [Google Scholar] [CrossRef]
- Kim, H.; Choi, K.; Kang, H.; Lee, S.Y.; Chi, S.W.; Lee, M.S.; Song, J.; Im, D.; Choi, Y.; Cho, S. Identification of a Novel Function of CX-4945 as a Splicing Regulator. PLoS One 2014, 9, e94978. [Google Scholar] [CrossRef]
- Lertsuwan, J.; Lertsuwan, K.; Sawasdichai, A.; Tasnawijitwong, N.; Lee, K.Y.; Kitchen, P.; Afford, S.; Gaston, K.; Jayaraman, P.S.; Satayavivad, J. CX-4945 Induces Methuosis in Cholangiocarcinoma Cell Lines by a CK2-Independent Mechanism. Cancers (Basel) 2018, 10. [Google Scholar] [CrossRef]
- Salvi, M.; Borgo, C.; Pinna, L.A.; Ruzzene, M. Targeting CK2 in Cancer: A Valuable Strategy or a Waste of Time? Cell Death Discov 2021, 7, 325. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Xu, G.; Gou, S. Novel CK2-Specific Pt(II) Compound Reverses Cisplatin-Induced Resistance by Inhibiting Cancer Cell Stemness and Suppressing DNA Damage Repair in Non-Small Cell Lung Cancer Treatments. J Med Chem 2021, 64, 4163–4178. [Google Scholar] [CrossRef] [PubMed]
- Grygier, P.; Pustelny, K.; Nowak, J.; Golik, P.; Popowicz, G.M.; Plettenburg, O.; Dubin, G.; Menezes, F.; Czarna, A. Silmitasertib (CX-4945), a Clinically Used CK2-Kinase Inhibitor with Additional Effects on GSK3β and DYRK1A Kinases: A Structural Perspective. J Med Chem 2023, 66, 4009–4024. [Google Scholar] [CrossRef]
- Siddiqui-Jain, A.; Drygin, D.; Streiner, N.; Chua, P.; Pierre, F.; O’Brien, S.E.; Bliesath, J.; Omori, M.; Huser, N.; Ho, C.; et al. CX-4945, an Orally Bioavailable Selective Inhibitor of Protein Kinase CK2, Inhibits Prosurvival and Angiogenic Signaling and Exhibits Antitumor Efficacy. Cancer Res 2010, 70, 10288–10298. [Google Scholar] [CrossRef]
- Ferguson, A.D.; Sheth, P.R.; Basso, A.D.; Paliwal, S.; Gray, K.; Fischmann, T.O.; Le, H.V. Structural Basis of CX-4945 Binding to Human Protein Kinase CK2. FEBS Lett 2011, 585, 104–110. [Google Scholar] [CrossRef]
- Pagano, M.A.; Meggio, F.; Ruzzene, M.; Andrzejewska, M.; Kazimierczuk, Z.; Pinna, L.A. 2-Dimethylamino-4,5,6,7-Tetrabromo-1H-Benzimidazole: A Novel Powerful and Selective Inhibitor of Protein Kinase CK2. Biochem Biophys Res Commun 2004, 321, 1040–1044. [Google Scholar] [CrossRef]
- Yim, H.; Lee, Y.H.; Lee, C.H.; Lee, S.K. Emodin, an Anthraquinone Derivative Isolated from the Rhizomes of Rheum Palmatum, Selectively Inhibits the Activity of Casein Kinase II as a Competitive Inhibitor. Planta Med 1999, 65, 9–13. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, S.L.; Dou, W.; Zhang, S.; Chen, J.H.; Shen, Y.; Shen, J.H.; Leng, Y. Emodin, a Natural Product, Selectively Inhibits 11beta-Hydroxysteroid Dehydrogenase Type 1 and Ameliorates Metabolic Disorder in Diet-Induced Obese Mice. Br J Pharmacol 2010, 161, 113–126. [Google Scholar] [CrossRef]
- Sarno, S.; Moro, S.; Meggio, F.; Zagotto, G.; Dal Ben, D.; Ghisellini, P.; Battistutta, R.; Zanotti, G.; Pinna, L.A. Toward the Rational Design of Protein Kinase Casein Kinase-2 Inhibitors. Pharmacol Ther 2002, 93, 159–168. [Google Scholar] [CrossRef]
- Cozza, G.; Mazzorana, M.; Papinutto, E.; Bain, J.; Elliott, M.; di Maira, G.; Gianoncelli, A.; Pagano, M.A.; Sarno, S.; Ruzzene, M.; et al. Quinalizarin as a Potent, Selective and Cell-Permeable Inhibitor of Protein Kinase CK2. Biochem J 2009, 421, 387–395. [Google Scholar] [CrossRef]
- Bullock, A.N.; Debreczeni, J.E.; Fedorov, O.Y.; Nelson, A.; Marsden, B.D.; Knapp, S. Structural Basis of Inhibitor Specificity of the Human Protooncogene Proviral Insertion Site in Moloney Murine Leukemia Virus (PIM-1) Kinase. J Med Chem 2005, 48, 7604–7614. [Google Scholar] [CrossRef] [PubMed]
- Mathison, C.J.N.; Chianelli, D.; Rucker, P.V.; Nelson, J.; Roland, J.; Huang, Z.; Yang, Y.; Jiang, J.; Xie, Y.F.; Epple, R.; et al. Efficacy and Tolerability of Pyrazolo[1,5-. ACS Med Chem Lett 2020, 11, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int J Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef]
- Licciardello, M.P.; Workman, P. A New Chemical Probe Challenges the Broad Cancer Essentiality of CK2. Trends Pharmacol Sci 2021, 42, 313–315. [Google Scholar] [CrossRef]
- Wells, C.I.; Drewry, D.H.; Pickett, J.E.; Tjaden, A.; Krämer, A.; Müller, S.; Gyenis, L.; Menyhart, D.; Litchfield, D.W.; Knapp, S.; et al. Development of a Potent and Selective Chemical Probe for the Pleiotropic Kinase CK2. Cell Chem Biol 2021, 28, 546–558.e10. [Google Scholar] [CrossRef]
- Yao, K.; Youn, H.; Gao, X.; Huang, B.; Zhou, F.; Li, B.; Han, H. Casein Kinase 2 Inhibition Attenuates Androgen Receptor Function and Cell Proliferation in Prostate Cancer Cells. Prostate 2012, 72, 1423–1430. [Google Scholar] [CrossRef]
- Golub, A.G.; Bdzhola, V.G.; Kyshenia, Y.V.; Sapelkin, V.M.; Prykhod’ko, A.O.; Kukharenko, O.P.; Ostrynska, O.V.; Yarmoluk, S.M. Structure-Based Discovery of Novel Flavonol Inhibitors of Human Protein Kinase CK2. Mol Cell Biochem 2011, 356, 107–115. [Google Scholar] [CrossRef]
- Guerra, B.; Issinger, O.G. Protein Kinase CK2 in Human Diseases. Curr Med Chem 2008, 15, 1870–1886. [Google Scholar] [CrossRef] [PubMed]
- Meggio, F.; Pagano, M.A.; Moro, S.; Zagotto, G.; Ruzzene, M.; Sarno, S.; Cozza, G.; Bain, J.; Elliott, M.; Deana, A.D.; et al. Inhibition of Protein Kinase CK2 by Condensed Polyphenolic Derivatives. An in Vitro and in Vivo Study. Biochemistry 2004, 43, 12931–12936. [Google Scholar] [CrossRef]
- Chilin, A.; Battistutta, R.; Bortolato, A.; Cozza, G.; Zanatta, S.; Poletto, G.; Mazzorana, M.; Zagotto, G.; Uriarte, E.; Guiotto, A.; et al. Coumarin as Attractive Casein Kinase 2 (CK2) Inhibitor Scaffold: An Integrate Approach to Elucidate the Putative Binding Motif and Explain Structure-Activity Relationships. J Med Chem 2008, 51, 752–759. [Google Scholar] [CrossRef]
- Lindenblatt, D.; Applegate, V.; Nickelsen, A.; Klußmann, M.; Neundorf, I.; Götz, C.; Jose, J.; Niefind, K. Molecular Plasticity of Crystalline CK2α’ Leads to KN2, a Bivalent Inhibitor of Protein Kinase CK2 with Extraordinary Selectivity. J Med Chem 2022, 65, 1302–1312. [Google Scholar] [CrossRef]
- Brear, P.; De Fusco, C.; Hadje Georgiou, K.; Francis-Newton, N.J.; Stubbs, C.J.; Sore, H.F.; Venkitaraman, A.R.; Abell, C.; Spring, D.R.; Hyvönen, M. Specific Inhibition of CK2α from an Anchor Outside the Active Site. Chem Sci 2016, 7, 6839–6845. [Google Scholar] [CrossRef]
- Iegre, J.; Brear, P.; De Fusco, C.; Yoshida, M.; Mitchell, S.L.; Rossmann, M.; Carro, L.; Sore, H.F.; Hyvönen, M.; Spring, D.R. Second-Generation CK2α Inhibitors Targeting the αD Pocket. Chem Sci 2018, 9, 3041–3049. [Google Scholar] [CrossRef]
- Chen, X.; Li, C.; Wang, D.; Chen, Y.; Zhang, N. Recent Advances in the Discovery of CK2 Allosteric Inhibitors: From Traditional Screening to Structure-Based Design. Molecules 2020, 25. [Google Scholar] [CrossRef]
- Oshima, T.; Niwa, Y.; Kuwata, K.; Srivastava, A.; Hyoda, T.; Tsuchiya, Y.; Kumagai, M.; Tsuyuguchi, M.; Tamaru, T.; Sugiyama, A.; et al. Cell-Based Screen Identifies a New Potent and Highly Selective CK2 Inhibitor for Modulation of Circadian Rhythms and Cancer Cell Growth. Sci Adv 2019, 5, eaau9060. [Google Scholar] [CrossRef]
- Borgo, C.; Ruzzene, M. Protein Kinase CK2 Inhibition as a Pharmacological Strategy. Adv Protein Chem Struct Biol 2021, 124, 23–46. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Wang, J.; Zhou, Z.; Cao, S.; Zhang, J. Strategies of Targeting CK2 in Drug Discovery: Challenges, Opportunities, and Emerging Prospects. J. Med. Chem. 2023, 66, 2257–2281. [Google Scholar] [CrossRef]
- Farina, H.G.; Benavent Acero, F.; Perera, Y.; Rodríguez, A.; Perea, S.E.; Castro, B.A.; Gomez, R.; Alonso, D.F.; Gomez, D.E. CIGB-300, a Proapoptotic Peptide, Inhibits Angiogenesis in Vitro and in Vivo. Exp Cell Res 2011, 317, 1677–1688. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, P.; Zou, L.; Chen, J.; Yang, C.; He, G.; Liu, B.; Liu, J.; Chiang, C.M.; Wang, G.; et al. Discovery of Novel Dual-Target Inhibitor of Bromodomain-Containing Protein 4/Casein Kinase 2 Inducing Apoptosis and Autophagy-Associated Cell Death for Triple-Negative Breast Cancer Therapy. J Med Chem 2021, 64, 18025–18053. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, Y.; Wang, J.; Zhou, Z.; Cao, S.; Zhang, J. Strategies of Targeting CK2 in Drug Discovery: Challenges, Opportunities, and Emerging Prospects. J Med Chem 2023, 66, 2257–2281. [Google Scholar] [CrossRef] [PubMed]
- López-Ramos, M.; Prudent, R.; Moucadel, V.; Sautel, C.F.; Barette, C.; Lafanechère, L.; Mouawad, L.; Grierson, D.; Schmidt, F.; Florent, J.C.; et al. New Potent Dual Inhibitors of CK2 and Pim Kinases: Discovery and Structural Insights. FASEB J 2010, 24, 3171–3185. [Google Scholar] [CrossRef]
- Sarno, S.; Mazzorana, M.; Traynor, R.; Ruzzene, M.; Cozza, G.; Pagano, M.A.; Meggio, F.; Zagotto, G.; Battistutta, R.; Pinna, L.A. Structural Features Underlying the Selectivity of the Kinase Inhibitors NBC and dNBC: Role of a Nitro Group That Discriminates between CK2 and DYRK1A. Cell Mol Life Sci 2012, 69, 449–460. [Google Scholar] [CrossRef]
- Łukowska-Chojnacka, E.; Wińska, P.; Wielechowska, M.; Poprzeczko, M.; Bretner, M. Synthesis of Novel Polybrominated Benzimidazole Derivatives-Potential CK2 Inhibitors with Anticancer and Proapoptotic Activity. Bioorg Med Chem 2016, 24, 735–741. [Google Scholar] [CrossRef]
- Morooka, S.; Hoshina, M.; Kii, I.; Okabe, T.; Kojima, H.; Inoue, N.; Okuno, Y.; Denawa, M.; Yoshida, S.; Fukuhara, J.; et al. Identification of a Dual Inhibitor of SRPK1 and CK2 That Attenuates Pathological Angiogenesis of Macular Degeneration in Mice. Mol Pharmacol 2015, 88, 316–325. [Google Scholar] [CrossRef]
- Sato, K.; Padgaonkar, A.A.; Baker, S.J.; Cosenza, S.C.; Rechkoblit, O.; Subbaiah, D.R.C.V.; Domingo-Domenech, J.; Bartkowski, A.; Port, E.R.; Aggarwal, A.K.; et al. Simultaneous CK2/TNIK/DYRK1 Inhibition by 108600 Suppresses Triple Negative Breast Cancer Stem Cells and Chemotherapy-Resistant Disease. Nat Commun 2021, 12, 4671. [Google Scholar] [CrossRef]
- Zien, P.; Duncan, J.S.; Skierski, J.; Bretner, M.; Litchfield, D.W.; Shugar, D. Tetrabromobenzotriazole (TBBt) and Tetrabromobenzimidazole (TBBz) as Selective Inhibitors of Protein Kinase CK2: Evaluation of Their Effects on Cells and Different Molecular Forms of Human CK2. Biochim Biophys Acta 2005, 1754, 271–280. [Google Scholar] [CrossRef]
- Koo, J.H.; Yu, H.C.; Nam, S.; Kim, D.C.; Lee, J.H. Casein Kinase 2 Alpha Inhibition Protects against Sepsis-Induced Acute Kidney Injury. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Nitta, R.T.; Gholamin, S.; Feroze, A.H.; Agarwal, M.; Cheshier, S.H.; Mitra, S.S.; Li, G. Casein Kinase 2α Regulates Glioblastoma Brain Tumor-Initiating Cell Growth through the β-Catenin Pathway. Oncogene 2015, 34, 3688–3699. [Google Scholar] [CrossRef]
- Ye, H.; Fu, D.; Fang, X.; Xie, Y.; Zheng, X.; Fan, W.; Hu, F.; Li, Z. Casein Kinase II Exacerbates Rheumatoid Arthritis via Promoting Th1 and Th17 Cell Inflammatory Responses. Expert Opin Ther Targets 2021, 25, 1017–1024. [Google Scholar] [CrossRef]
- Luo, Y.; Lei, Y.; Guo, X.; Zhu, D.; Zhang, H.; Guo, Z.; Xu, Z.; Zhao, H.; Xi, Y.; Peng, X.; et al. CX-4945 Inhibits Fibroblast-like Synoviocytes Functions through the CK2-P53 Axis to Reduce Rheumatoid Arthritis Disease Severity. Int Immunopharmacol 2023, 119, 110163. [Google Scholar] [CrossRef] [PubMed]
- Okoumassoun, L.E.; Russo, C.; Denizeau, F.; Averill-Bates, D.; Henderson, J.E. Parathyroid Hormone-Related Protein (PTHrP) Inhibits Mitochondrial-Dependent Apoptosis through CK2. J Cell Physiol 2007, 212, 591–599. [Google Scholar] [CrossRef]
- Lee, S.W.; Rho, J.H.; Lee, S.Y.; Yoo, S.H.; Kim, H.Y.; Chung, W.T.; Yoo, Y.H. Alpha B-Crystallin Protects Rat Articular Chondrocytes against Casein Kinase II Inhibition-Induced Apoptosis. PLoS One 2016, 11, e0166450. [Google Scholar] [CrossRef]
- Kim, K.M.; Song, J.D.; Chung, H.T.; Park, Y.C. Protein Kinase CK2 Mediates Peroxynitrite-Induced Heme Oxygenase-1 Expression in Articular Chondrocytes. Int J Mol Med 2012, 29, 1039–1044. [Google Scholar] [CrossRef]
- Kim, K.M.; Sohn, D.H.; Kim, K.; Park, Y.C. Inhibition of Protein Kinase CK2 Facilitates Cellular Senescence by Inhibiting the Expression of HO-1 in Articular Chondrocytes. Int J Mol Med 2019, 43, 1033–1040. [Google Scholar] [CrossRef]
- Nie, J.; Liu, L.; He, F.; Fu, X.; Han, W.; Zhang, L. CKIP-1: A Scaffold Protein and Potential Therapeutic Target Integrating Multiple Signaling Pathways and Physiological Functions. Ageing Research Reviews 2013, 12, 276–281. [Google Scholar] [CrossRef]
- Fu, L.; Zhang, L. Physiological Functions of CKIP-1: From Molecular Mechanisms to Therapy Implications. Ageing Research Reviews 2019, 53, 100908. [Google Scholar] [CrossRef]
- Yuan, Y.; Sun, J.; Zhou, H.; Wang, S.; He, C.; Chen, T.; Fang, M.; Li, S.; Kang, S.; Huang, X.; et al. The Effect of QiangGuYin on Osteoporosis through the AKT/mTOR/Autophagy Signaling Pathway Mediated by CKIP-1. Aging (Albany NY) 2022, 14, 892–906. [Google Scholar] [CrossRef]
- Peng, X.; Wu, X.; Zhang, J.; Zhang, G.; Li, G.; Pan, X. The Role of CKIP-1 in Osteoporosis Development and Treatment. Bone & Joint Research 2018, 7, 173–178. [Google Scholar] [CrossRef]
- Xiao, P.-L.; Cui, A.-Y.; Hsu, C.-J.; Peng, R.; Jiang, N.; Xu, X.-H.; Ma, Y.-G.; Liu, D.; Lu, H.-D. Global, Regional Prevalence, and Risk Factors of Osteoporosis According to the World Health Organization Diagnostic Criteria: A Systematic Review and Meta-Analysis. Osteoporos Int 2022, 33, 2137–2153. [Google Scholar] [CrossRef]
- Liu, J.; Lu, C.; Wu, X.; Zhang, Z.; Li, J.; Guo, B.; Li, D.; Liang, C.; Dang, L.; Pan, X.; et al. Targeting Osteoblastic Casein Kinase-2 Interacting Protein-1 to Enhance Smad-Dependent BMP Signaling and Reverse Bone Formation Reduction in Glucocorticoid-Induced Osteoporosis. Sci Rep 2017, 7, 41295. [Google Scholar] [CrossRef]
- Izeradjene, K.; Douglas, L.; Delaney, A.; Houghton, J.A. Influence of Casein Kinase II in Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand-Induced Apoptosis in Human Rhabdomyosarcoma Cells. Clinical Cancer Research 2004, 10, 6650–6660. [Google Scholar] [CrossRef]
- Takahashi, K.; Setoguchi, T.; Tsuru, A.; Saitoh, Y.; Nagano, S.; Ishidou, Y.; Maeda, S.; Furukawa, T.; Komiya, S. Inhibition of Casein Kinase 2 Prevents Growth of Human Osteosarcoma. Oncol Rep 2017, 37, 1141–1147. [Google Scholar] [CrossRef]
- Lisberg, A.; Ellis, R.; Nicholson, K.; Moku, P.; Swarup, A.; Dhurjati, P.; Nohe, A. Mathematical Modeling of the Effects of CK2.3 on Mineralization in Osteoporotic Bone: Mathematical Modeling of the Effects of CK2.3. CPT Pharmacometrics Syst. Pharmacol. 2017, 6, 208–215. [Google Scholar] [CrossRef]
- Akkiraju, H.; Bonor, J.; Nohe, A. CK2.1, a Novel Peptide, Induces Articular Cartilage Formation in Vivo. J Orthop Res 4AD, 35, 876–885. [CrossRef]
- Akkiraju, H.; Srinivasan, P.P.; Xu, X.; Jia, X.; Safran, C.B.K.; Nohe, A. CK2.1, a Bone Morphogenetic Protein Receptor Type Ia Mimetic Peptide, Repairs Cartilage in Mice with Destabilized Medial Meniscus. Stem Cell Res Ther 04 18, 8, 82. [CrossRef]
- Halloran, D.; Pandit, V.; MacMurray, C.; Stone, V.; DeGeorge, K.; Eskander, M.; Root, D.; McTague, S.; Pelkey, H.; Nohe, A. Age-Related Low Bone Mineral Density in C57BL/6 Mice Is Reflective of Aberrant Bone Morphogenetic Protein-2 Signaling Observed in Human Patients Diagnosed with Osteoporosis. IJMS 2022, 23, 11205. [Google Scholar] [CrossRef]
- Weidner, H.; Yuan Gao, V.; Dibert, D.; McTague, S.; Eskander, M.; Duncan, R.; Wang, L.; Nohe, A. CK2.3, a Mimetic Peptide of the BMP Type I Receptor, Increases Activity in Osteoblasts over BMP2. IJMS 2019, 20, 5877. [Google Scholar] [CrossRef]
- Vrathasha, V.; Weidner, H.; Nohe, A. Mechanism of CK2.3, a Novel Mimetic Peptide of Bone Morphogenetic Protein Receptor Type IA, Mediated Osteogenesis. IJMS 2019, 20, 2500. [Google Scholar] [CrossRef]
Figure 1.
Comparison of effect of inhibition of kinase activity and individual subunit knockout on mesenchymal stem cell line. Dysregulation of Wnt signaling pathway causes oncogenic activation in myogenic C2C12 cells. CK2 is a regulator of Wnt pathway. Individual subunits have other unidentified roles in myogenesis or regulation of Wnt pathway. Knockout of α ot β subunit causes complete suppression of myogenesis. Wheeras, inhibition of kinase activity causes partial suppression of myogenesis.
Figure 1.
Comparison of effect of inhibition of kinase activity and individual subunit knockout on mesenchymal stem cell line. Dysregulation of Wnt signaling pathway causes oncogenic activation in myogenic C2C12 cells. CK2 is a regulator of Wnt pathway. Individual subunits have other unidentified roles in myogenesis or regulation of Wnt pathway. Knockout of α ot β subunit causes complete suppression of myogenesis. Wheeras, inhibition of kinase activity causes partial suppression of myogenesis.
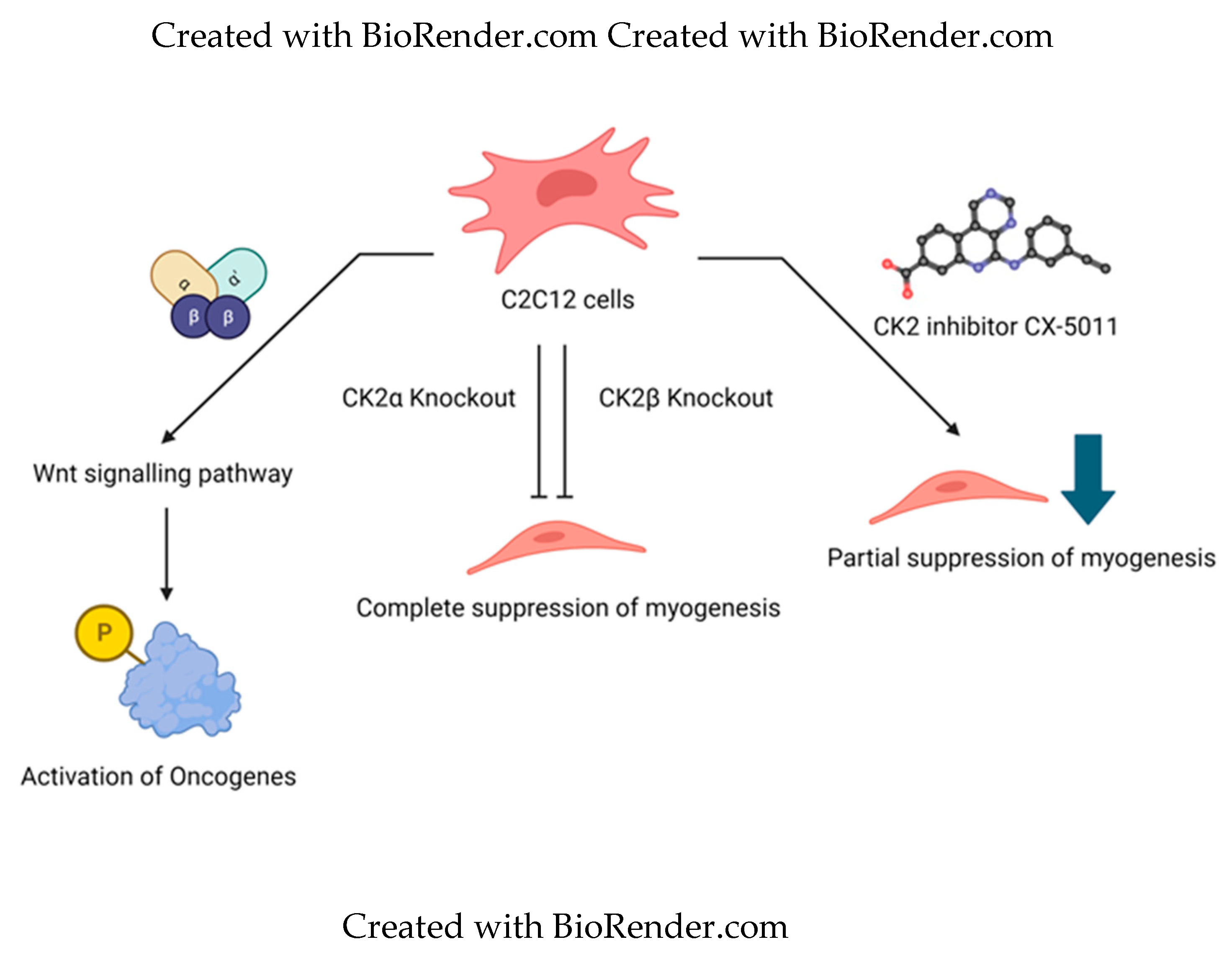
Figure 2.
Implication of CK2 in RA and effect of inhibition of CK2 kinase activity. Expression of CK2 is upregulated in RA. Increase in CK2 activity increases inflammation by various mechanisms. a) Inhibition of CK2 in Fibloblast like synovyocytes reduced their proliferation and hence reduced the synovial inflammation. b) In RA, CD4+ cells treated with CK2 inhibitors, had reduction in activation of Th1 and Th17 T cell response and it activated Th2 type cell response. Inhibition of CK2 in CD4+ cells reduced their proliferation and hence Th1 and Th17 T cell response was activated. These effects were seen in in-vivo conditions as well.
Figure 2.
Implication of CK2 in RA and effect of inhibition of CK2 kinase activity. Expression of CK2 is upregulated in RA. Increase in CK2 activity increases inflammation by various mechanisms. a) Inhibition of CK2 in Fibloblast like synovyocytes reduced their proliferation and hence reduced the synovial inflammation. b) In RA, CD4+ cells treated with CK2 inhibitors, had reduction in activation of Th1 and Th17 T cell response and it activated Th2 type cell response. Inhibition of CK2 in CD4+ cells reduced their proliferation and hence Th1 and Th17 T cell response was activated. These effects were seen in in-vivo conditions as well.
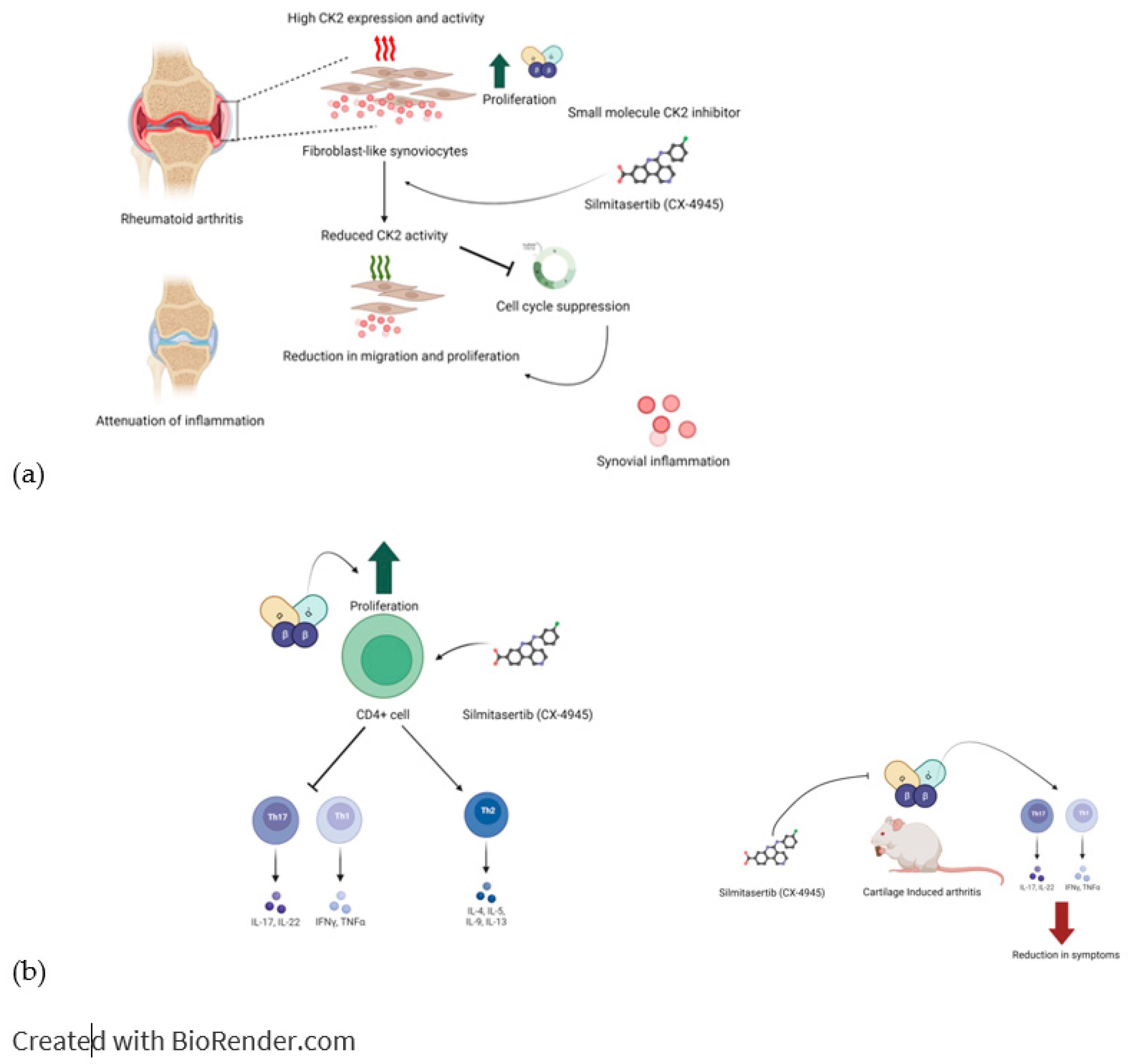
Figure 3.
Implication of CK2 in Osteosarcoma and effect of inhibition of CK2 kinase activity. Inhibition of CK2 makes the cells more sensitive to Apoptotic signals.
Figure 3.
Implication of CK2 in Osteosarcoma and effect of inhibition of CK2 kinase activity. Inhibition of CK2 makes the cells more sensitive to Apoptotic signals.
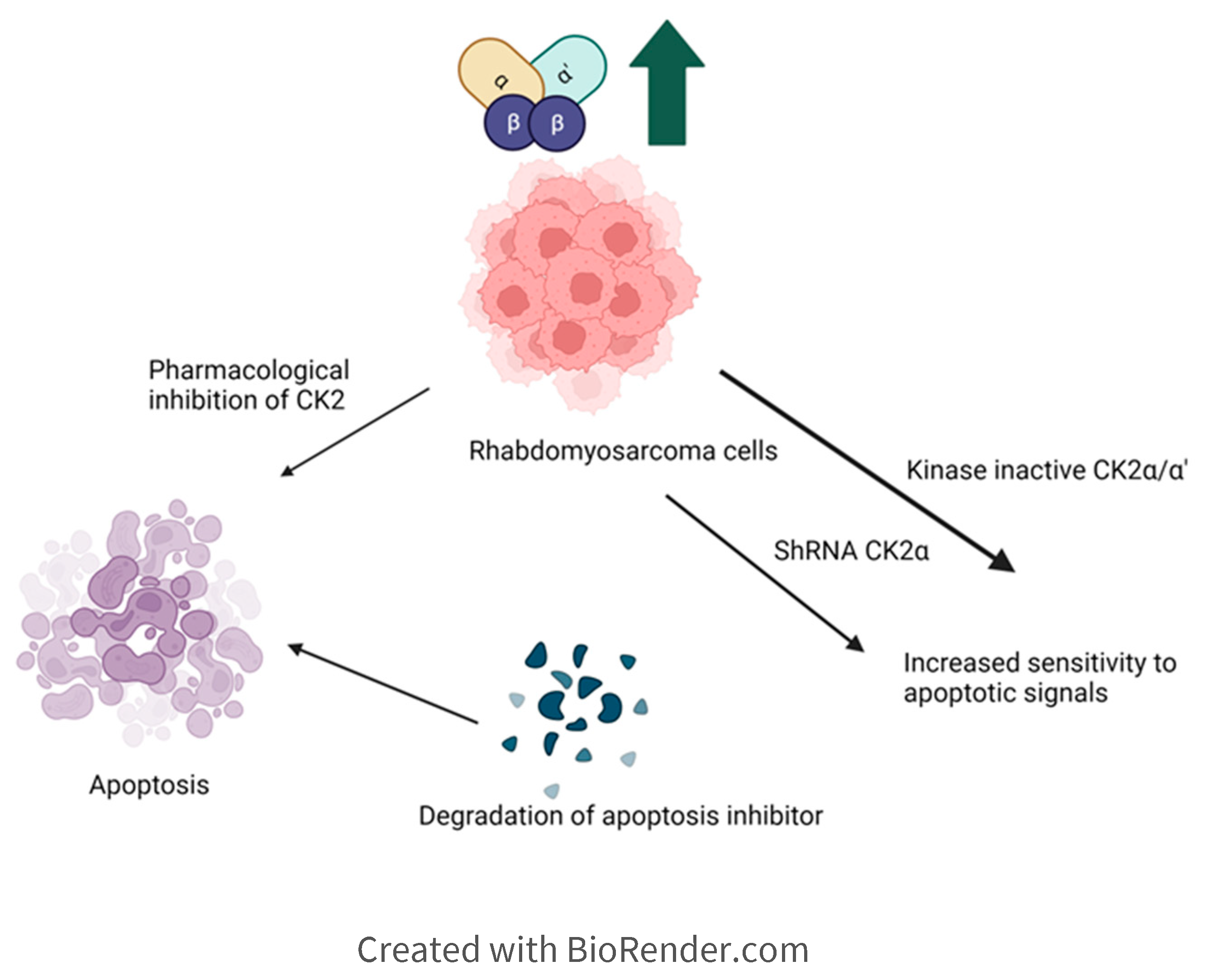
Figure 4.
Inhibition of the interaction between CK2 and BMPRIa is effective strategy for the treatment of Osteoarthtis and Osteoporosis. a) Stimulation with BMP2 ligand causes heterodimerization of BMPRIa receptor with BMPRII. Release of CK2 from BMPRIa and its phosphorylation by BMPRII activates BMP signaling pathway. b) BMPRIa receptor has three phosphosites on the cytoplasmic/intracellular domain. These phosphosites were validated using CK2 mutants. c) BMPRIa mimetic peptides containing phosphosites sequence corresponding to ‘SYED’ activates articular chondrocytes to repair Osteoarthtic lesions in knee articular cartilage. The BMPRIa mimetic peptide containing phosphosite sequence corresponding to ‘SLKD’ corrects the BMP pathway in senile mice. Thereby it helps regain balance between Osteoblastogenesis and Osteoclastic activity, which is affected in Osteoporosis. .
Figure 4.
Inhibition of the interaction between CK2 and BMPRIa is effective strategy for the treatment of Osteoarthtis and Osteoporosis. a) Stimulation with BMP2 ligand causes heterodimerization of BMPRIa receptor with BMPRII. Release of CK2 from BMPRIa and its phosphorylation by BMPRII activates BMP signaling pathway. b) BMPRIa receptor has three phosphosites on the cytoplasmic/intracellular domain. These phosphosites were validated using CK2 mutants. c) BMPRIa mimetic peptides containing phosphosites sequence corresponding to ‘SYED’ activates articular chondrocytes to repair Osteoarthtic lesions in knee articular cartilage. The BMPRIa mimetic peptide containing phosphosite sequence corresponding to ‘SLKD’ corrects the BMP pathway in senile mice. Thereby it helps regain balance between Osteoblastogenesis and Osteoclastic activity, which is affected in Osteoporosis. .
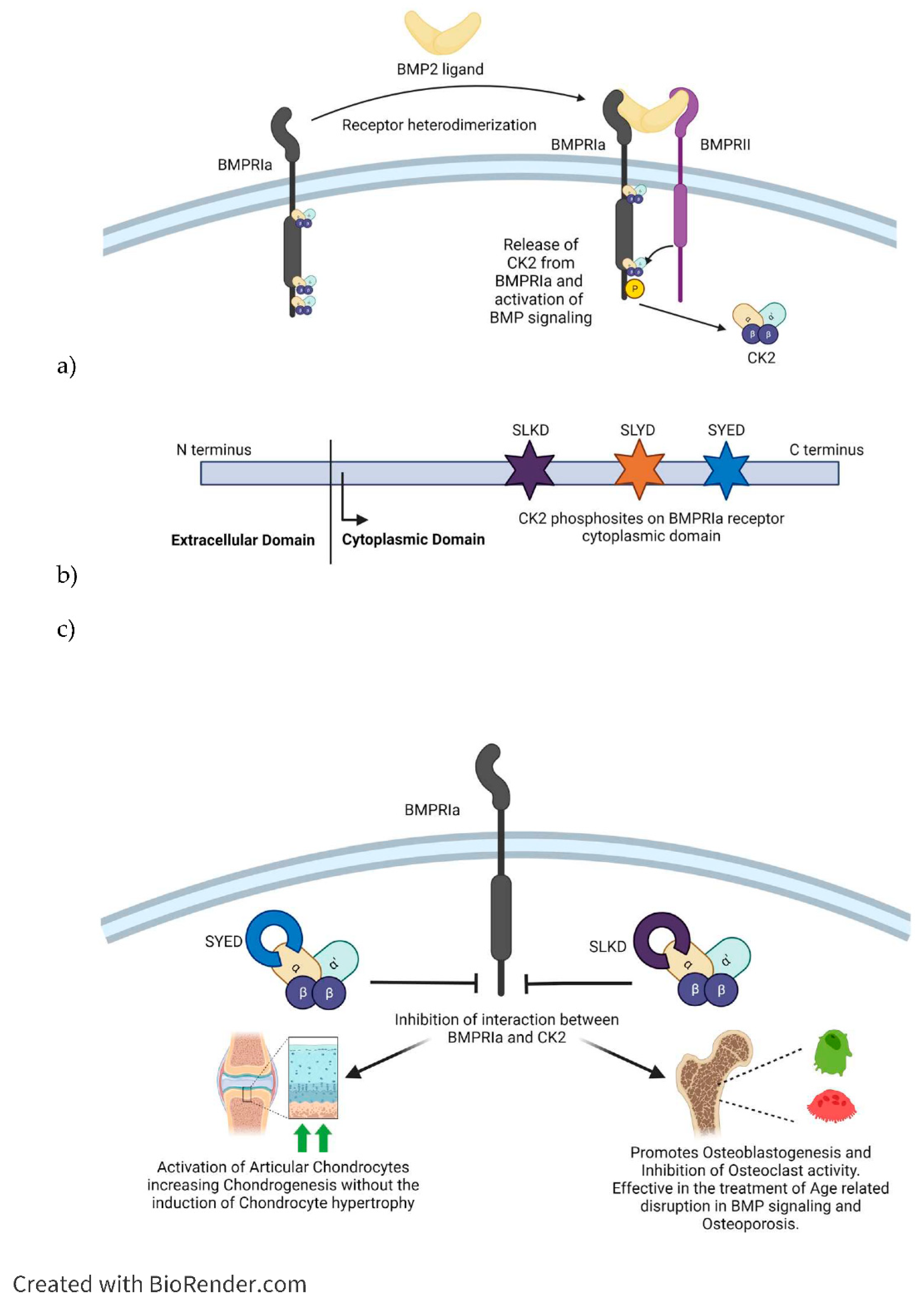
Figure 5.
Current CK2 inhibitors are evolved for increased specificity for CK2, reducing off-target effects and better cell permeability. In the case of anti-cancer drugs, higher cytotoxicity is achieved in CK2 inhibitors. Future CK2 inhibitors could potentially target pathology-specific substrate interactions with CK2 using the knowledge of validated phosphosites of CK2 and the role of subunits or molecular forms of CK2 in processes.
Figure 5.
Current CK2 inhibitors are evolved for increased specificity for CK2, reducing off-target effects and better cell permeability. In the case of anti-cancer drugs, higher cytotoxicity is achieved in CK2 inhibitors. Future CK2 inhibitors could potentially target pathology-specific substrate interactions with CK2 using the knowledge of validated phosphosites of CK2 and the role of subunits or molecular forms of CK2 in processes.
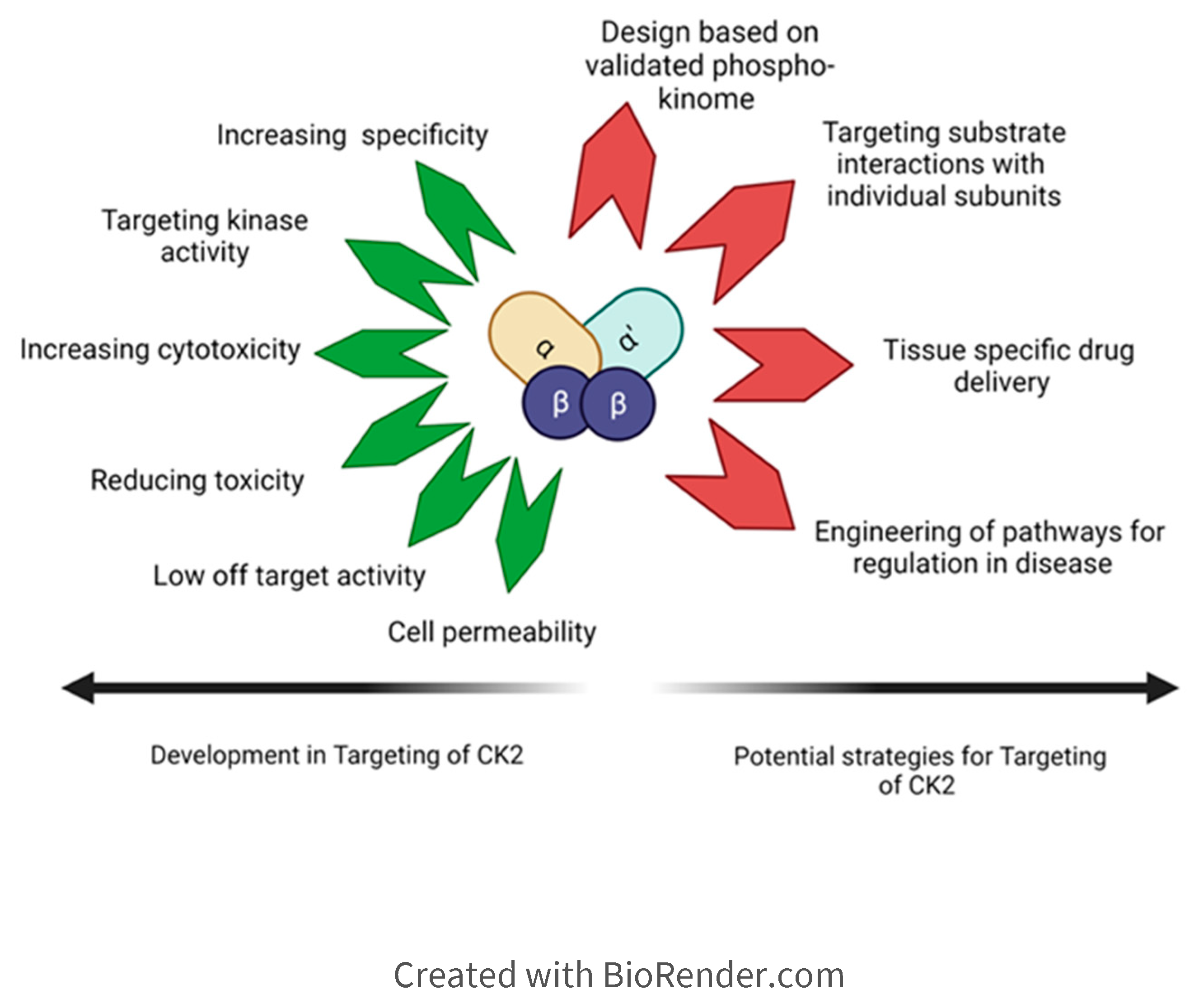
Table 1.
Small molecular inhibitors of CK2 and their Clinical utility.
| Name | Status for Clinical Application | Reference |
|---|---|---|
| Silmitasertib (CX-4945) | The most selective CK2 inhibitor. Promotes apoptosis while inhibiting the PI3K/Akt signaling pathway and the cell cycle progression. It has little toxicity. It is in Clinical use. | [54,55] |
| dimethylamino-4,5,6,7-1H-tetrabromobenzimidazole (DMAT) | Permeates cell membranes and induces apoptosis in the Jerkat cell line. | [29,56] |
| Emodin | It’s a natural anthraquinone derivative extracted from rhubarb. Inhibits CK2. | [57,58,59] |
| Quinalizarin | One of the most selective CK2 inhibitors. Polar interactions are established with CK2 in at least three hydroxyl groups. Inhibits CK2 and promotes apoptosis in HEK-293 and Jurkat cells. | [60] |
| IC20 | Extremely high selectivity for CK2alpha. It doesn’t show cytotoxicity in cancer cells. Binds at multiple sites with CK2. | [61] [62,63] |
| SGC-CK2-1 | High selectivity and cell-membrane permeable CK2 inhibitor. Used as a cellular probe to investigate how CK2 functions in the cell. It has little inhibitory effect on the proliferation of most cancer cells, and it is only effective against a small subset of cancer cells. | [64,65] |
| Tetrabromocinnamic Acid (TBCA) | Promotes apoptosis in Jurkat cancer cells. Suppresses platelet aggregation/secretion and the cell cycle progression in prostate cancer cells. | [66] |
| 4-(6,8-Dibromo-3-hydroxy-4-oxo-4H-chromo-2-yl)-benzoic acid (FLC26) | Mild increase in selectivity over the predecessor compound FLC21. Permeable to cell membranes and caused a significant increase in apoptosis in PANC-1 cells. | [67,68] |
| 3,8-dibromo-7-hydroxy-4-methylchromen-2-one (DBC) | Cell permeable and induces apoptosis in Jurkat cells. | [69,70] |
| CAM4066 | Poor cell membrane permeability. However, a synthetic methyl ester derivative, pro-CAM4066, increases its cell permeability, making it effective against cancerous tumors. Similar to KN2, but less selective and therefore less optimal as an inhibitor of CK2. Acts on the αD region and ATP-binding sites of CK2. The moiety bound to the ATP binding site forms a hydrogen bond with Lys68 and two water molecules. The moiety in the αD site interacts with Pro159 and a conserved water molecule. The linker forms a network of hydrogen bonds. | [71,72,73] |
| CAM4712 | Has high cell permeability and anti-proliferation effects. | [74,73] |
| GO289 | High selectivity for CK2, with little inhibitory effect on other kinases. Extremely selective, ideal for clinical use. Inhibition of the phosphorylation sites of multiple clock proteins and suppressed the growth/proliferation of cells of a diverse array of cancers. CK2α and CK2α′ are the primary targets of. | [75] |
| HY1-Pt | Derived from CX-4945. Extremely high selectivity, ideal for clinical use. Reversed cisplatin-induced drug resistance. Suppresses DNA damage repair in cancer cells. It also inhibited the Wnt/beta-catenin signaling pathway while activating the mitochondrial apoptosis pathway. Displayed no toxicity to healthy hepatocytes and could be used as a therapeutic for NSCLC. | [52] |
Table 3.
CK2 inhibitors with pathway or subunit specific mechanism of action.
| Name | Status for Clinical Applica-tion | Reference |
|---|---|---|
| CIGB-300 |
Cell-permeable. Inhibits angiogenesis and metastasis. Used in early clinical trials in combination with chemoradiotherapy as a therapeutic against cervical cancer. Administered by injection into the tumor. Targets the phosphoacceptor domain. Releases histamine from the cells, possibly due to higher intracellular calcium levels in the cell. | [37] |
| 4,5,6,7-tetrabromobenzotriazole (TBBt) | Moderately effective as an anti-cancer drug. Induces apoptosis in tumor cells. Inhibits CK2α subunit. Used in Sepsis-Induced Acute Kidney Injury. | [86,87] |
| 4,5,6,7-tetrabromobenzimidazole (TBBz) | Able to target specific molecular forms of CK2. It is more effective in inducing apoptosis and necrosis in tumor cells compared to TBBt. Inhibits CK2α subunit activity. Tested in Glioblastoma Cell lines. | [86,88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



