Submitted:
28 October 2023
Posted:
30 October 2023
You are already at the latest version
Abstract
The farnesoid X receptor (FXR)/βKlotho/fibroblast growth factors (FGFs) pathway is crucial for maintaining the intestinal barrier and preventing colorectal cancer (CRC). We used an FXR ago-nist, GW4064, and FXR knockout (KO) mice to investigate the role of FXR/Klothos/FGFs pathways in lipopolysaccharide (LPS)-induced intestinal barrier dysfunction and colon carcinogenesis. The results showed that upregulation of FXR in enterocytes effectively ameliorated intestinal tight junction markers (claudin1, Zonula occludens-1), inflammation, and bile acid levels, thereby protecting mice from intestinal barrier dysfunction and colon carcinogenesis. GW4064 treatment increased FXR, αKlotho, βKlotho, FGF19, FGF21, and FGF23 in wild-type mice exposed to LPS, while FXR KO mice had decreased levels. FXR KO mice exhibited elevated colon cancer markers (β-catenin, LGR5, CD44, CD34, Cyclin D1) under LPS, underscoring the pivotal role of FXR in inhibiting the development of colon tumorigenesis. The varying gut microbiota responses in FXR KO mice versus wild-type mice post-LPS exposure emphasize the pivotal role of FXR in pre-serving intestinal microbial health, involving Bacteroides thetaiotaomicron, Bacteroides acidifaciens, and Helicobacter hepaticus. Our study validates the effectiveness of GW4064 in alleviating LPS-induced disruptions to the intestinal barrier and colon carcinogenesis, emphasizing the importance of the FXR/αKlotho/βKlotho/FGFs pathway and the interplay between bile acids and gut microbiota.
Keywords:
Farnesoid X receptor
; gut microbiota
; intestinal epithelial barrier
; colon cancer
; Klotho
; fibroblast growth factor
1. Introduction
Farnesoid X receptor (FXR) is a regulator of bile acid (BA) and lipid, that meticulously regulates bile acid production and circulation. The role of FXR activation has been shown to be important in colitis [1] and nonalcoholic steatohepatitis (NASH) [2] in mouse models. FXR knockout (KO) mice exhibit increased BA pool concentration, which leads to spontaneous tumor development in liver, intestinal epithelial cell proliferation, and increased colon cancer susceptibility [3,4,5]. Bile acids and FXR play an important role in the modulation of a range of inflammatory responses, barrier function and the prevention of bacterial translocation in the intestinal tract. Although it has been determined that FXR plays a variety of functions, further research is still needed to determine how it assists in intestinal epithelial barrier function and colon tumorigenesis [3,4,6].
The three subfamilies of Klotho are αKlotho, βKlotho, and γKlotho [7]. βKlotho is a co-receptor for fibroblast growth factor 19 (FGF19) and FGF21, two hormones that play important roles in regulating energy and bile acid metabolism. In response to bile acid binding, FXR induces the expression of FGF19 in the ileum, which then binds to beta-klotho and activates downstream signaling pathways, leading to inhibition of bile acid synthesis in the liver. βklotho protein levels were substantially decreased in FXR KO mice; on the other hand, overexpression of βklotho in FXR-lacking hepatocytes partially restored FGF19 signaling and inhibition by FGF19 of Cyp7a1, which encodes the rate-limiting BA biosynthetic enzyme [8]. Overall, the interaction between FXR, βklotho, and FGF19/21 signaling plays an important role in regulating bile acid synthesis and metabolism. In the gut, the decreased βklotho expression caused by gene variation is associated with increased intestinal permeability in patients with irritable bowel syndrome with diarrhea [9] Furthermore, βklotho activation enhanced tight junction (TJ) proteins, thereby protecting against alcohol-induced TJ proteins endocytosis and degradation as well as intestinal barrier impairment [10].
The gene αKlotho has been associated with anti-aging effects and regulates several pathways involved in aging, such as phosphate homeostasis, insulin signaling, and Wnt/β-catenin signaling [11,12]. It also affects intracellular signaling pathways, including p53/p21, cyclic adenosine monophosphate (cAMP), protein kinase C (PKC) and transforming growth factor β (TGFβ) [13,14]. Studies have shown that αKlotho is expressed in the intestinal epithelium, which is the layer of cells that forms the intestinal barrier. αKlotho has been shown to promote the formation of tight junctions, which are the specialized structures that hold intestinal epithelial cells together. αKlotho levels are epigenetically downregulated in cancer, and overexpression or treatment with soluble klotho or the αKlotho domain slows growth of cancer cells in vitro and in vivo. αKlotho is also downregulated in colorectal cancer and αklotho overexpression inhibits growth of colorectal cancer cells [15,16]. αKlotho has been identified as a tumor suppressor and inhibits the insulin-like growth factor 1 (IGF1), FGF, and Wnt/β-catenin pathways; yet, the mode of action of αKlotho in cancer is a matter of confirm. To determine how much this interaction between FXR and Klotho affects the regulation of bile acid metabolism and other biological processes, more research into the molecular mechanisms underpinning it is necessary.
There is a strong interplay between intestinal flora and bile acid (BA) metabolism, which plays a crucial role in digestion and shaping the gut microbial community. Recent research has shown that gut microbiota-mediated deconjugation of bile acids influences de novo bile acid synthesis in the liver in an FXR-FGF15/19 axis-dependent manner in mice [17]. Gut microbiome performs essential bile acids and also influences host metabolism via the modulation of metabolites [18]. Gut microbiota also influences host metabolism via the modulation of metabolites, including the endotoxin LPS, bile acids, and short chain fatty acids. Therefore, they partially mediate the interaction between the gastrointestinal system and other organs [19]. Prevalent genera identified in the guts include both beneficial and harmful microorganisms, such as Bacteroides, Eubacterium, Bifidobacterium, Ruminococcus, Clostridium, Lactobacillus, Escherichia, Streptococcus [20], and naturally acquired enterohepatic Helicobacter spp. infection [21]. While most infected mice develop minimal pathologic changes, susceptible strains exhibit typhlocolitis and hepatitis, which can further progress to colon cancer and hepatocellular carcinoma [22]. Intestinal bacterial dysbiosis and Helicobacter hepaticus infection act synergistically during inflammation and neoplastic progression [23].
In this study, we demonstrated that downregulating FXR levels by applying LPS-treated and FXR-KO resulted in a decline in levels of α/βKlotho, FGF19, FGF21, and FGF23. GW4064 was found to improve the αKloth/βKlotho/FGFs pathway and decrease intestinal inflammation and β-catenin formation induced by LPS-induced intestinal barrier failure and colon carcinogenesis. The study also revealed that GW4064 prevented LPS-induced intestinal tight-junction damage, regulated bile acids, and improved microbiota dysbiosis in an FXR-dependent mechanism. Furthermore, LPS treatment induced severe intestinal barrier damage, higher bile acid levels, microbiota dysbiosis, and inflammatory response in FXR-deficient mice. The findings suggest that activating FXR and the αKloth/βKlotho/FGFs pathway might be a new strategy for preventing intestinal epithelial barrier dysfunction and colon tumorigenesis.
2. Results
2.1. GW4064 improves tight junction disruption in LPS-treated mice
Compared with the WT mice, LPS-treated mice presented with colon injuries that disrupted the epithelial barrier and caused infiltration of inflammatory cells into the mucosa and submucosa (Figure 1A). 13-month-old FXR KO colon showed villi-form papillary folds, had moderately reduced colon crypt height, and inflammatory cell infiltration (Figure 1A). However, LPS treatment in FXR KO mice resulted in serious injuries, such as loss of histological structures and increased inflammatory cell infiltration compared to LPS-treated WT mice. GW4064 restored the intestinal mucosal morphology and tight junction disruption in LPS-treated WT mice, but not in FXR KO mice (Figure 1A). GW4064 treatment increased FXR protein levels in LPS-treated WT mice (Figure 1A–C). Treatment with LPS or GW4064 did not alter the FXR protein levels in KO mice (Figure 1A-C). The zonula occludens-1 (ZO-1) and claudin-1 protein levels appeared to be increased by GW4064 administration in WT mice after LPS treatment, whereas no such effect was observed in FXR KO mice (Figure 1A, D and E). These data suggest that GW4064 rectifies LPS-induced intestinal barrier disruption and that this process requires functional FXR signaling.
GW4064 alters bile acid composition in LPS-treated mice. To determine whether GW4064 treatment alters BA profiles in the plasma, liver, and colon, BA species were assayed (Figure 2B-H). Compared with WT mice, LPS increased BA levels in the plasma and liver (~3-fold and ~2.25-fold, respectively). In contrast, the BA levels were decreased in the colon of WT mice (Figure 1B, C). FXR KO mice displayed an increase in total BA pool, which was decreased in the liver and further enhanced the level of BAs in the colon of LPS-treated FXR KO mice (Figure 1B-D). GW4064 reversed BA levels in the plasma, liver, and colon of WT and FXR KO mice treated with LPS. Moreover, all FXR KO mice showed considerably higher BA levels compared with WT mice, suggesting that FXR-deficiency causes BA homeostasis disorder. Of particular interest, the levels of tauro-cholic acid (TCA), tauro-chenodeoxycholate (TCDCA), tauro-ursodesoxy cholic acid (TUDCA), and tauro-deoxycholic acid (TDCA) were increased in the plasma and liver of WT mice after LPS injection (Figure 2 E-H), and GW4064 treatment was able to reverse most BAs and tauro-conjunction BA levels in the plasma and liver. In FXR KO mice, the levels of TCA increased over 100-fold in the plasma and 25-fold in the liver compared with WT mice. Interestingly, TCA, TUDCA, and TDCA levels were decreased in plasma and liver, as well as TCDCA in liver of LPS-treated FXR KO mice. Although GW4064 administration reversed almost bile acid levels in plasma and liver of WT and FXR KO mice-treated LPS, GW4064 markedly enhanced tauro-conjunction BA levels (Figure 2G and H), suggesting that these conditions lead to inflammatory disease susceptibility.
2.2. GW4064 improves tight junction disruption in LPS-treated mice
To test whether or not the alteration in the bile acid profile due to LPS treatment affects intestinal FXR signaling or not, the FXR signaling-related factors protein and mRNA expression was assessed. LPS enhanced MRP2, MRP3, OATP1, and decreased OSTβ, ASBT protein levels in WT mice, and GW4064 almost completely abolished this condition (Figure 3A and B). FXR KO mice had higher MRP2, MRP3, ASBT, OATP1, and lower OSTβ compared with WT mice. LPS further enhanced MRP2, MRP3, ASBT, and reduced OSTβ protein levels in FXR KO mice, and GW4064 administrated did not reverse this condition (Figure 3C and D). LPS reduced Fxr and pregnane X receptor (Pxr) and enhanced constitutive androstane receptor (Car), cytochrome P4503A11 (Cyp3a11), and sulfotransferase family 2A member 1 (Sult2a1) mRNA expression in WT mice, and GW4064 reversed this condition (Figure 3E). FXR KO mice had lower Fxr and Pxr mRNA expression than WT mice and their expression did not change after LPS and GW4064 administration in colon tissues (Figure 3E). Additionally, compared to WT, FXR KO mice had higher Car, Cyp3a11, and lower Sult2a1 mRNA expression, which increased furthermore following LPS administration (Figure 3E). Interestingly, GW4064 further enhanced Car and Sult2a1 mRNA expression in LPS-treated FXR KO mice (Figure 3E).
LPS also induced organic anion-transporting polypeptide 2B1 (Oatp2b1), Mrp2, Mrp3, multidrug resistance protein 1b (Mdr1b), Mdr2, and breast cancer resistance protein (Bcrp), and reduced apical sodium-dependent bile acid transporter (Asbt), ileal bile acid-binding protein (Ibabp), organic solute transporter α (Ostα), Ostβ mRNA expression in WT mice, and GW4064 almost completely abolished this condition. FXR KO mice had higher Asbt, Ibabp, Mrp2, Mrp3, Mdr1b, Mdr2, Bcrp, and lower Oatp2b1 and Ostβ, while unperturbed Ostα mRNA expression (Figure 3F and G) compared with WT mice. GW4064 reduced Oatp2b1 and Mrp2 expression and increased Ostβ, Mdr2, and Bcrp mRNA expression furthermore in LPS-treated FXR KO mice (Figure 3F and G). Hence, our results demonstrate that the imbalance in bile acid transporters due to LPS treatment could be significantly improved by GW4064 administration in WT mice but only reversed partially mRNA expression in FXR KO mice (Figure 3).
2.3. GW4064 improves FXR dysfunction and regulates the BA receptors and transporters
To test whether or not the alteration in the bile acid profile due to LPS treatment affects intestinal FXR signaling or not, the FXR signaling-related factors protein and mRNA expression was assessed. LPS enhanced MRP2, MRP3, OATP1, and decreased OSTβ, ASBT protein levels in WT mice, and GW4064 almost completely abolished this condition (Figure 3A and B). FXR KO mice had higher MRP2, MRP3, ASBT, OATP1, and lower OSTβ compared with WT mice. LPS further enhanced MRP2, MRP3, ASBT, and reduced OSTβ protein levels in FXR KO mice, and GW4064 administrated did not reverse this condition (Figure 3C and D). LPS reduced Fxr and pregnane X receptor (Pxr) and enhanced constitutive androstane receptor (Car), cytochrome P4503A11 (Cyp3a11), and sulfotransferase family 2A member 1 (Sult2a1) mRNA expression in WT mice, and GW4064 reversed this condition (Figure 3E). FXR KO mice had lower Fxr and Pxr mRNA expression than WT mice and their expression did not change after LPS and GW4064 administration in colon tissues (Figure 3E). Additionally, compared to WT, FXR KO mice had higher Car, Cyp3a11, and lower Sult2a1 mRNA expression, which increased furthermore following LPS administration (Figure 3E). Interestingly, GW4064 further enhanced Car and Sult2a1 mRNA expression in LPS-treated FXR KO mice (Figure 3E).
LPS also induced organic anion-transporting polypeptide 2B1 (Oatp2b1), Mrp2, Mrp3, multidrug resistance protein 1b (Mdr1b), Mdr2, and breast cancer resistance protein (Bcrp), and reduced apical sodium-dependent bile acid transporter (Asbt), ileal bile acid-binding protein (Ibabp), organic solute transporter α (Ostα), Ostβ mRNA expression in WT mice, and GW4064 almost completely abolished this condition. FXR KO mice had higher Asbt, Ibabp, Mrp2, Mrp3, Mdr1b, Mdr2, Bcrp, and lower Oatp2b1 and Ostβ, while unperturbed Ostα mRNA expression (Figure 3F and G) compared with WT mice. GW4064 reduced Oatp2b1 and Mrp2 expression and increased Ostβ, Mdr2, and Bcrp mRNA expression furthermore in LPS-treated FXR KO mice (Figure 3F and G). Hence, our results demonstrate that the imbalance in bile acid transporters due to LPS treatment could be significantly improved by GW4064 administration in WT mice but only reversed partially mRNA expression in FXR KO mice (Figure 3).
2.4. GW4064 regulates αKlotho/FGF23 and βKlotho/FGF19/FGF21 pathway
To assess the underlying mechanistic basis whereby GW4064 could alter αKlotho/FGF23 and βKlotho/FGF19/FGF21 pathways, that pathways were subsequently analyzed via immunohistochemical and western blot analyses. As shown in Figure 4A-C, GW4064 induced αKlotho, βKlotho, FGF19, FGF21, and FGF23 protein levels in LPS treated mice. Compared with WT mice, were significantly decreased αKlotho, βKlotho, FGF19, FGF21, and FGF23 protein levels in FXR KO mice (Figure 4A-D). Therefore, αKlotho, βKlotho, FGF19, FGF21, and FGF23 protein levels were reduced in LPS-treated FXR KO mice; moreover, GW4064 only partial enhanced FGF19 and FGF23 in FXR KO mice treated LPS (Figure 4B and D).
2.5. GW4064 suppresses LPS-induced inflammatory response, ER stress, and angiogenesis in mice
In LPS-treated WT mice, GW4064 significantly decreased colonic mRNA expression of the proinflammatory genes, Tnfα, interferon gamma (Ifnγ), and Il-1β (Figure 5A). GW4064 alleviated LPS-induced pro-caspase-1, adaptor protein apoptosis-associated speck-like protein containing a caspase recruitment domain (Asc), Nlrp3, and Pannexin-1 mRNA expression in WT mice. Compared to WT mice, the FXR KO mice had higher Tnfα, Il-1β, pro-caspase-1, Asc, Nlrp3, and Pannexin-1 mRNA expression. GW4064 did not affect LPS-induced inflammatory mediator and Nlrp3 inflammasome in FXR KO mice (Figure 5A). Furthermore, we investigated the effect of GW4064 on LPS-induced endoplasmic reticulum (ER) stress in the intestinal inflammation model. Compared to WT mice, the mRNA levels of activating transcription factor 4 (Atf4), Atf6, glucose-regulated protein 78 (Grp78), CCAAT-enhancer-binding protein homologous protein (Chop), and X-box binding protein 1 Spliced (Xbp1s) increased remarkably upon LPS administration, which was markedly suppressed by GW4064 treatment (Figure 5B). FXR KO mice had higher Atf4, Atf6, and Xbp1s mRNA levels than WT mice under un-induced conditions. Our results demonstrate that GW4064 had no effect on LPS-induced increase in ER stress in FXR KO mice (Figure 5B). The LPS effects on colonic inflammatory response and tight junction disruption and microbiota dysbiosis might be due to the upregulation of TLR4 signaling. Our results showed that GW4064 repressed LPS-induced TLR4, myeloid differentiation primary response 88 (MyD88), nuclear factor kappa-B (NF-κB), and Caspase 3 protein levels in WT mice, but not in FXR KO mice (Figure 5C-G).
To investigate the mechanism contributing to the formation of angiogenesis and intestinal epithelial cell proliferation in mice treated with LPS, GW4064 significantly diminished β-catenin and c-myc protein levels in WT mice treated with LPS, but not in FXR KO mice (Figure 5H-J). Additionally, GW4064 significantly decreased LPS-induced transforming growth factor beta receptor II (TGFβRII), intercellular adhesion molecule (ICAM), vascular cell adhesion molecule (VCAM), vascular endothelial growth factor (VEGF), vascular endothelial growth factor receptor 1 (VEGFR1), and matrix metallopeptidase 9 (MMP9) protein levels in WT mice (Figure 5K). FXR KO mice displayed higher levels of angiogenesis protein than WT mice. LPS stimulation in FXR KO mice resulted in a dramatic increase in TGRβRII, VCAM, VEGF1, and MMP9 protein levels compared to untreated. GW4064 treatment had no effect on TGRβRII, VCAM, VEGF1, and MMP9 protein levels in FXR KO mice with LPS treatment (Figure 5L). These data indicate that FXR activation decreases the expression of several inflammatory, Nlrp3 inflammasome, and ER stress genes by inhibiting TLR4 and β-catenin signaling, contributing to the amelioration of LPS-induced intestinal injury in WT mice.
2.6. GW4064 regulates LPS-induced intestinal epithelial cell proliferation
We hypothesized that FXR activation may alter the growth of colon stem cells (CSCs), which is generally expressed in the intestine. Mechanistically, we found that the expression levels of intestinal stem cell markers and their proliferation, including CD44, LGR5, PCNA, CD34, BrdU, CD133, GCSF, and cyclin D1 were downregulated 30-40% upon GW4064 administration in WT mice treated with LPS (Figure 6A-E). In addition, GW4064 decreased the Lgr5, Olfm4, and cyclin D1 mRNA expression (Figure 6F) in WT mice treated with LPS, but not in FXR KO mice. Our results corroborated the attenuation of stem cell proliferation by FXR, as expression of cancer stem cells expansion and proliferation marker protein increased significantly in FXR KO mice and further enhanced with LPS treatment. These data suggest that FXR may play an important role in early stage of colon cancer development.
2.7. GW0464 administration affects intestinal microbial diversity and composition
To investigate the effects of GW4064 on gut microbiota in LPS-treated WT and FXR KO mice, we examined the stool samples from WT and FXR KO mice post-treatment. Alpha diversity indices, including observed Chao1 and Shannon’s diversity indices, were lower in the LPS-treated WT mice than in the untreated mice; however, the diversity was restored to a normal level by GW4064 treatment (Figure 7A and B). The alpha diversity was reduced in FXR KO mice compared with WT mice, and it further decreased with LPS treatment. Notably, GW4064 treatment did not alter the alpha diversity in these mice (Figure 7A and B). Principal component analyses (PCA) revealed that the gut microbiota in the LPS treated group deviated from the untreated group, and GW4064 partially restored the level of gut microbiome in LPS-treated mice (Figure 7C). In addition, the gut microbiota profile of FXR KO group differed from the WT group.
Here, an analysis of 16S rRNA gene sequencing results revealed distinct clustering of colon microbiome communities isolated from WT and FXR KO mice. Changes in the composition of the gut microbiota induced by GW4064 were noted and it was observed that the colon community structure in LPS-injected mice was altered following GW4064 treatment in WT and FXR KO mice (Figure 7D-Z). The phyla Proteobacteria, Verrucomicrobia, Bacteroidetes, class Betaproteobacteria, Gammaproteobacteria, Verrucomicrobiae, order Enterobacteriales, Verrucomicrobiales, genus Bacteroides, Escherichia, and species Bacteroides thetaiotaomicron, Escherichia coli, Bacteroides acidifaciens, Akkermansia muciniphila, and Clostridium saccharogumia were significantly increased, while phyla Firmicutes, the Firmicutes/Bacteroidetes (F/B) ratio, class Clostridia, and genus Clostridium were reduced in WT mice treated with LPS (Figure 7D-Z). However, compared with WT mice, specifically, the phyla Bacteroidetes, genus Bacteroides, and species Bacteroides thetaiotaomicron were increased, and the phyla Verrucomicrobia, the Firmicutes/Bacteroidetes (F/B) ratio, class Betaproteobacteria, Gammaproteobacteria, Verrucomicrobiae, order Verrucomicrobiales, Burkholderiales, genus Clostridium, Escherichia, species Escherichia coli, Akkermansia muciniphila, and Clostridium saccharogumia were significantly reduced in FXR KO mice (Figure 7H-Z). The phyla Proteobacteria, Firmicutes, order Enterobacteriales, species Bacteroides acidifaciens, and Helicobacter hepaticus did not significantly change in FXR KO mice (Figure 7E-Z). Interestingly, FXR KO mice treated with LPS showed increased levels of Bacteroidetes, Betaproteobacteria, Clostridia, Bacteroides, Clostridium, and Helicobacter hepaticus. The abundance of specific bacterial species from Proteobacteria, Verrucomicrobia, Gammaproteobacteria, Clostridia, Verrucomicrobiae, Enterobacteriales, Verrucomicrobiales, Clostridium, Escherichia, Escherichia coli, and Akkermansia muciniphila exhibited oppositing changes in lipopolysaccharides (LPS) treatment between wild-type (WT) and FXR knockout (FXR KO) mice, as compared to their respective control groups (Figure 7E-X). After GW4064 treatment, the abundance of Proteobacteria, Verrucomicrobia, Firmicutes, Betaproteobacteria, Gammaproteobacteria, Clostridia, Verrucomicrobiales, Enterobacteriales, Verrucomicrobiales, Clostridium, Escherichia, Bacteroides thetaiotaomicron, Escherichia coli, Bacteroides acidifaciens, Akkermansia muciniphila, and Clostridium saccharogumia were significantly reversed in LPS-treated WT mice, while only a few of these microbes were restored in LPS-treated FXR KO mice (Figure 7E-Z). These results indicate that GW4064 has a potential role in the regulation of microbiota dysbiosis.
2.8. Impact of GW0464 Administration on Total SCFA and BCFA Concentrations
To understand the impact of GW4064 on intestinal injury, bile acid signaling, and changes in the gut microbiota, we further analyze the variations in Short Chain Fatty Acids (SCFA) and Branched Chain Fatty Acids (BCFA). LPS have been observed to significantly decrease the concentrations of total SCFA and BCFA in the feces of normal mice (Figure 7A, B). The administration of GW4064 further reduces total SCFA, but interestingly, it significantly elevates the concentration of total BCFA. In the feces of FXR KO mice, the concentrations of total SCFA and BCFA are significantly lower than those in normal mice. However, post LPS treatment, there is a significant increase in the concentrations of total SCFA and BCFA. When treated concurrently with GW4064, the concentration of total BCFA experiences a significant decrease (Figure 7A, B). Further analysis reveals that LPS significantly reduces the concentrations of acetic acid, butyric acid, valeric acid, isobutyric acid, isovaleric acid, 2-methylbutyric acid, and 4-methylvaleric acid in normal mouse feces. GW4064 administration significantly reverses these changes induced by LPS. However, GW4064 also reduces acetic acid concentration, which constitutes a large proportion of total SCFA. As a result, there is no significant improvement in the concentration of total SCFA. On the other hand, GW4064 significantly increases the changes in propionic acid, butyric acid, valeric acid, hexanoic acid, isobutyric acid, isovaleric acid, 2-methylbutyric acid, and 4-methylvaleric acid concentrations (except for formic acid and acetic acid) (Figure 7C-H). In FXR-deficient mice, apart from formic acid, the concentrations of other acids such as acetic acid, butyric acid, valeric acid, hexanoic acid, isobutyric acid, isovaleric acid, 2-methylbutyric acid, and 4-methylvaleric acid are significantly lower than those in normal mice. Post LPS treatment leads to a significant increase in the concentration of these acids. However, following the administration of GW4064 there is a significant decrease observed. The absence of FXR in FXR KO mice primarily results in no significant changes in isovaleric acid concentration regardless of whether it is treated with LPS or GW4064 (Figure 7J). Therefore, FXR and GW4064 administration affects Short Chain Fatty Acids (SCFA) and Branched Chain Fatty Acids (BCFA) concentrations in mice.
Figure 8.
GW4064 alters SCFA and BCFA in feces. (A) Total SCFA and (B) total BCFA levels in feces. (C-H) Formate acid, acetate acid, propinate acid, butyrate acid, valerate acid, and hexanoate acid levels in feces. (I-L) Isobutyrate acid, isovalerate acid, 2-methylbutyrate acid, and 4-methylvalerate acid levels in feces. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 8.
GW4064 alters SCFA and BCFA in feces. (A) Total SCFA and (B) total BCFA levels in feces. (C-H) Formate acid, acetate acid, propinate acid, butyrate acid, valerate acid, and hexanoate acid levels in feces. (I-L) Isobutyrate acid, isovalerate acid, 2-methylbutyrate acid, and 4-methylvalerate acid levels in feces. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
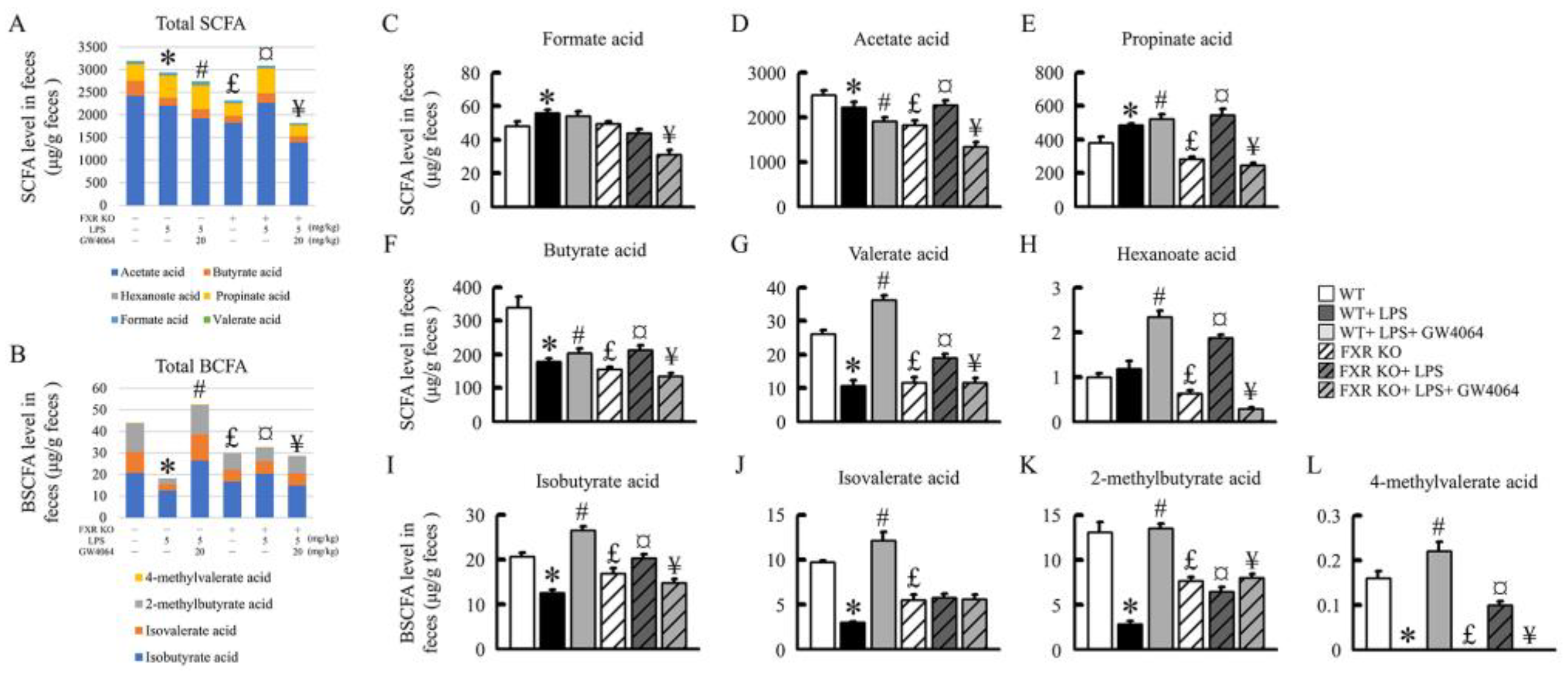
Figure 9.
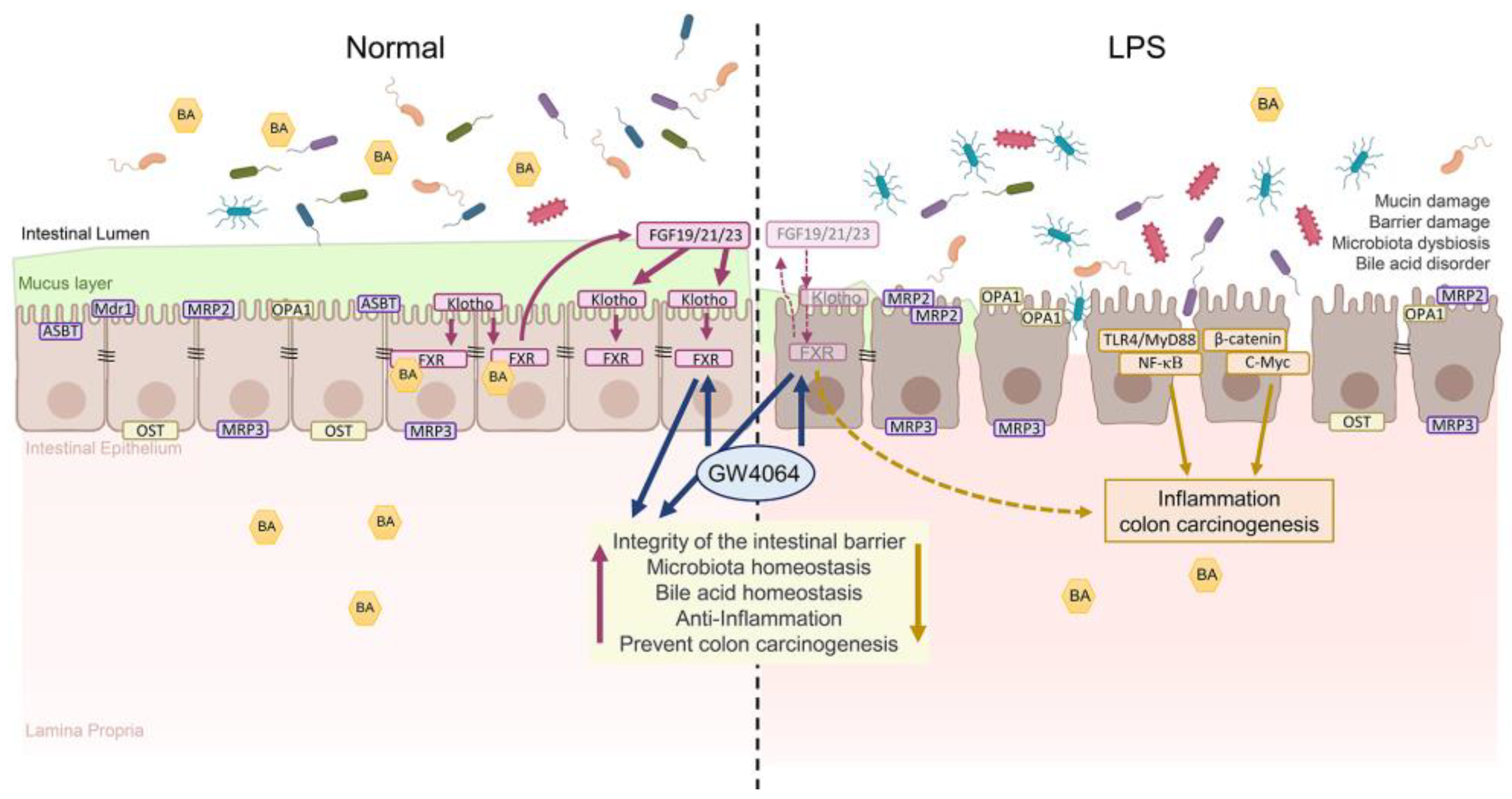
Representation of the inflammatory process induced in the colonic mucosa of LPS models. LPS causes disruption of the intestinal epithelial barrier and, thereby, the entry of luminal bacteria or bacterial antigens into the mucosa. The FXR/αKlotho/βKlotho/FGFs pathway is crucial for maintaining the integrity of the intestinal barrier and preventing colon cancer. Upregulation of FXR helps protect against intestinal barrier dysfunction and colon carcinogenesis by improving tight junction markers, reducing inflammation, and regulating β-catenin levels. GW4064, an FXR agonist, enhances FXR, αKlotho, βKlotho, FGF19, FGF21, and FGF23 protein levels in the colon, while FXR deficiency leads to a decrease in FXR/Klothos/FGFs protein levels and increased colon tumorigenesis markers. FXR plays a role in modulating gut microbiota and bile acid metabolism, highlighting the interplay between these factors. Activation of the FXR/αKlotho/βKlotho/FGFs pathway may be a potential strategy for preventing intestinal barrier dysfunction and colon tumorigenesis.
Figure 9.
Representation of the inflammatory process induced in the colonic mucosa of LPS models. LPS causes disruption of the intestinal epithelial barrier and, thereby, the entry of luminal bacteria or bacterial antigens into the mucosa. The FXR/αKlotho/βKlotho/FGFs pathway is crucial for maintaining the integrity of the intestinal barrier and preventing colon cancer. Upregulation of FXR helps protect against intestinal barrier dysfunction and colon carcinogenesis by improving tight junction markers, reducing inflammation, and regulating β-catenin levels. GW4064, an FXR agonist, enhances FXR, αKlotho, βKlotho, FGF19, FGF21, and FGF23 protein levels in the colon, while FXR deficiency leads to a decrease in FXR/Klothos/FGFs protein levels and increased colon tumorigenesis markers. FXR plays a role in modulating gut microbiota and bile acid metabolism, highlighting the interplay between these factors. Activation of the FXR/αKlotho/βKlotho/FGFs pathway may be a potential strategy for preventing intestinal barrier dysfunction and colon tumorigenesis.
3. Discussion
FXR activation can decrease inflammation in the gut and improve the integrity of the intestinal epithelial barrier. Similar to our research, Watanabe M et al. investigated at which FXR activation affected affects the function of the intestinal barrier in a mouse model of intestinal injury [24]. Our preliminary research indicates that GW4064 treatment prevented the expression of ZO-1 and claudin-1 from declining only in WT mice, indicating that FXR activation may be important for maintaining TJPs expression in the gut [25].
FXR is a transcription factor that regulates bile acid (BA) homeostasis in the liver and gastrointestinal system. In FXR KO mice, BA production is impaired, resulting in increased BA concentrations [26]. Abnormalities in BA metabolism (such as TCA) are linked to hepatocellular carcinoma [3] and colon cancer [27]. Moreover, bile acid is absorbed via the ASBT and it is exported via the basolateral heteromeric OSTα-OSTβ in intestinal epithelial barrier [28]. In the present study, OSTβ protein levels were significantly lower in both LPS-treated mice. Specifically, the OSTβ protein levels were significantly lower in FXR KO mice than in WT mice. OST levels were severely diminished in intestinal injury tissue and markedly reduced in adjacent non-tumorous tissue [29]. In LPS-induced intestinal injury, FXR KO mice had higher serum levels of taurine-conjugated bile acids, which are linked to sepsis. The FXR-specific agonist GW4064 improved Tauro-BA profile in plasma and liver and enhanced ASBT and OSTα/β levels in WT mice, but not in FXR KO mice, suggesting that the effect of GW4064 was dependent on FXR. In LPS-induced intestinal injury, FXR KO mice showed increased expression of Asbt, Ibabp, and Mdr2 and decreased protein levels of OSTβ, leading to intracellular accumulation of BAs, increased cell apoptosis, and inflammatory response in enterocytes of the colon.
The βKlotho/FGF15/19/21 pathway activation via FXR results in the reduction of bile acid production through negative feedback control. The βKlotho/FGF19 signaling was hampered in FXR-KO mice hepatocytes, which is similar to our finding [8]. βKlotho KO mice presented hepatic alterations, combining a proinflammatory status and initiation of fibrosis. These defects are associated with a massive shift in BA composition in the enterohepatic system and the circulation, which is characterized by a large excess of microbiota-derived deoxycholic acid (DCA) known for its genotoxicity in the gastrointestinal tract [30]. Interestingly, despite increased synthesis and excretion of bile acids by the liver, ileal expression of ASBT was not reduced in β-Klotho-deficient mice [15,16]. FXR and βKlotho/FGF19/FGF15 plays an important role in modulation of the liver receptor homolog -1 (LRH-1) or pregnane X receptor (RAR)/retinoic acid receptor (RAR) pathway to regulate ASBT causing in alter bile acid translation. Our findings demonstrated that loss of FXR or treatment with GW4064, which enhances FXR activity, interferes with the response of β-Klotho to ASBT regulation. This reveals the mechanisms by which FXR influences β-Klotho expression and activity. Furthermore, βKlotho appears to regulate the endocytosis and degradation of Occludin and ZO-1. The endocytosis of TJ proteins from the plasma membrane is a key that mechanism regulates TJ plasticity and function in epithelial barrier tissues [31]. In the present study, we have shown that FXR activation by GW4064 can regulate BA homeostasis and intestinal barrier function involved in the βKlotho/FGF19/FGF21 pathway.
Mechanistically, we showed that BAs that antagonize intestinal Farnesoid X receptor (FXR) function, including tauro-β-muricholic acid (T-βMCA) and deoxycholic acid (DCA), induce proliferation and DNA damage in Lgr5+ cells. Conversely, selective activation of intestinal FXR can restrict abnormal Lgr5+ cell growth and curtail CRC progression. FXR silencing in chronic colitis mouse models of intestinal tumorigenesis results in early mortality and increased tumor progression [32]. In colon cancer, low FXR expression was correlated with worse clinical outcome [32]. The present study revealed that knockdown of FXR activated Wnt/β-catenin signaling [5]. The mechanism by which FXR suppresses tumor growth remains unclear, but it may involve protecting the colonic epithelium from inflammation and ameliorating BA toxicity by upregulating intracellular BA binding proteins and efflux transporters and downregulating influx transporters and de novo BA synthesis [33,34]. Thus, the re-establishment of FXR signaling not only restricts aberrant Lgr5+ stem cell proliferation but also promotes gut health including restoring the intestinal barrier [35,36] and BA homeostasis [37,38]. Beyond its well-established role in regulating cytotoxicity of hydrophobic BAs, early study highlights the role of FXR in restricting the tumorigenesis of Lgr5+ cells, which mediate the key adenoma-to-adenocarcinoma transformation. FXR deficiency increases colon cancer susceptibility by increasing epithelial permeability to bacteria, promoting Wnt/β-catenin signaling and increasing intestinal inflammation [29,36]. In this study, we shown that GW4064 significantly reduced colon tumorigenesis markers, LGR5, CD44, CD34, Cyclin D1 in LPS-treat WT mice, but not in FXR KO mice.
The αklotho is a classical aging suppressor, which is a subfamily of βKlotho. Klotho has been shown to regulate intestinal barrier function and protect against gut inflammation, which may help to prevent the development of colon cancer. This decrease in αKlotho levels may be observed in patients with several aging-related diseases such as cancer [39]. In colorectal cancer models, Klotho overexpression has been shown to reduce overall β-catenin expression, inhibiting transcriptional pathway activity by binding to the Wnt3a ligand, and thereby decreasing nuclear translocation of β-catenin [40]. The anti-neoplastic effects of Klotho and describe the modulation of downstream oncogenic signaling pathways, including Wnt/β-catenin, FGF, IGF1, PIK3K/AKT, TGFβ, and the Unfolded Protein Response [39]. In this study, we shown that GW4064 significantly reduced colon tumorigenesis may through FXR dependent in αKlotho/FGF23 pathway.
In the present study, Bacteroidetes, and Bacteroides genera were the predominant gut anaerobes, which increased as a result of FXR deactivation in both WT mice and FXR KO mice treated with LPS; these data are consistent with those of Jena et al. [41] Bacteroides species are bile acid-resistant pathogens, which are found in most anaerobic infections with a mortality of >19% [42]. In this study, we focused on the TCA, whose components, taurine and cholic acid, support the growth of microbial groups of low abundance. These less less-abundant microbes are mechanistically implicated in DNA-damage and tumor-promotion mediated by their metabolic by-products [27]. Our data revealed that Bacteroides thetaiotaomicron, and Bacteroides acidifaciens were eliminated by GW4064 in LPS-treated WT mice, but not in FXR KO mice. However, the persistent Bacteroides thetaiotaomicron in FXR KO mice might lead to maintaining chronic intestinal inflammation after GW4064 treatment. Furthermore, the commensal gut bacterium Bacteroides thetaiotaomicron has a robust ability to degrade dietary polysaccharides and host mucin glycans. This, in turn, leads to a notable reduction in the thickness of the colonic mucus layer and exacerbates enteric infections by altering the metabolic environment [43]. In the present study, GW4064 reversed LPS-induced relative abundance of Bacteroides acidifaciens in WT mice. Compared to WT mice, Bacteroides thetaiotaomicron was increased in FXR KO mice, and neither LPS nor GW4064 treatment showed any significant change in it. While FXR deficiency strongly affects the expression of genes related to immunity and bile acid metabolism, as well as the composition of the microbiome; its deficiency was not able to produce significant histopathological changes in the absence of Helicobacter hepaticus infection. This data confirms those reported by Swennes et al. [23] Bacteroides acidifaciens, a colitis-associated species and an abundant member of the Bacteroidaceae family [44], was also elevated during inflammation, but decreased rapidly after colitis. Specifically, LPS decreased FXR levels and induced microbiota dysbiosis in the colon, which elevated TCA, TCDCA, T-ursodeoxycholic acid (TUDCA), and TDCA in the plasma and liver, leading to the impairment of intestinal tight junctions and inflammation. Conversely, direct activation of FXR restores bile acid levels, microbiota profiles, intestinal tight junctions, and reduces inflammation in LPS-treated WT.
The contribution of SCFAs to maintenance of gut homeostasis has been investigated extensively, there is an increasing body of evidence that commensal bacteria and bacterial metabolites have opposing roles in inflammatory responses and carcinogenesis depending on the cell type and the environment. Based on our findings from the gut microbiota and SCFAs results, we can highlight three important aspects: (1) The genus Clostridium has been associated with the production of several SCFAs, including butyrate acid, valerate acid, isobutyrate acid, and 4-methylvalerate acid. The trends in Clostridium populations appear to correspond with the levels of these SCFAs [45]. (2) The genus Bacteroides has been linked with the production of propionate, another type of SCFA that has important roles in gut health and metabolism. The trends in Bacteroides populations appear to correspond with the levels of propionate. Bacteroides and Clostridium may be responsible for the increase in SCFA levels and bile acid metabolism in FXR KO treatments. (3) Isovalerate (or isovaleric acid) is a branched-chain fatty acid produced by the gut microbiota. The trends in isovalerate levels appear to correspond with the populations of Clostridium saccharogumia. However, research on the relationship between Clostridium saccharogumia and SCFAs is currently lacking. In a steady-state situation, butyrate is present in the mM range in the gut lumen and serves as the primary energy source for colonocytes [46]. However, in the context of cancerous colonocytes, butyrate was shown to act paradoxically. In addition to that, butyrate is capable of promoting carcinogenesis in a genetic mouse model based on mutations in the Apc and the mismatch repair gene Msh2 (ApcMin/+; Msh2–/–) [47]. And superior inhibitory efficacy of butyrate over propionate and acetate against human colon cancer cell proliferation via cell cycle arrest and apoptosis [48]. Our study shows that GW4064 positively affects SCFA and BCFA levels in normal mice. However, in FXR KO mice, the regulatory effect of GW4064 on SCFA is different from that in normal mice, possibly due to an increase in total bile acids and pathogenic intestinal tract bacteria, including Clostridium and Bacteroidetes spp.
Ultimately, the study search elucidates that the activation of the FXR/αKlotho/βKlotho/FGFs pathway may hold promise as a potential strategy for averting intestinal barrier dysfunction and colon tumorigenesis. Lastly, our research highlights those elevated levels of bile acids and the high abundance of certain microbiota, such as Clostridium, Bacteroidetes, and Helicobacter hepaticus, Bacteroides thetaiotaomicron, in FXR KO mice are crucial factors leading to the reduction of colonic tight junction proteins and consequent damage to the intestinal barrier. Additionally, LPS treatment exacerbates structural damage to the intestine, rendering the administration of GW4064 less effective in mitigating these effects. As such, this study emphasizes two key aspects: the paramount importance of FXR in the context of intestinal injury and the secondary significance of the gut microbiota-bile acid-FXR axis as a therapeutic target for intestinal diseases.
4. Materials and Methods
4.1. Animals and treatments
13-month-old male C57BL/6J wild-type (WT) mice (Taiwan National Laboratory Animal Center) and FXR knockout (KO) mice (strain B6.129X1(FVB)-Nr1h4tm1Gonz/J; stock number 007214, Jackson Laboratory) were maintained in the Animal Experimental Center of Chang Gung University under 12-h light/dark cycle. Food and water were provided ad libitum. Lipopolysaccharide (LPS) is a vital component of the outer membrane in Gram-negative bacteria. LPS serves as the principal toxin derived from the gut microbiota, triggering inflammation in gut epithelial cells. Notably, the serum concentration of LPS is significantly elevated in patients with inflammatory bowel disease (IBD) [49], and LPS has been employed in experimental animal models to investigate intestinal injury [50]. In this study, WT and FXR KO mice were intraperitoneally injected with a single dose of LPS (5 mg/kg) and after 12 hours injected with GW4064 (20 mg/kg i.p. every 12 hours) twice. After 6 hours of the last GW4064 treatment, all animals were terminated by gradual fill isoflurane asphyxiation.
4.2. Histology
The colon tissues were fixed in 4% paraformaldehyde for 48 h and cut into 5 μm-thick serial sections, while hematoxylin and eosin (HE) staining for histology. Slides were incubated with 7.5% non-immune goat serum for 30 min, and then incubated with primary antibody for 2 hours. Immunofluorescence (IF) and immunohistochemical (IHC) staining were performed as previously described [51]. The antibodies used for IF and IHC are listed in Table 1.
4.3. Western blot analysis
The colon tissue protein extraction for use NE-PER nuclear and cytoplasmic extraction reagents. The protein concentration was determined by the use of Bio-Red Protein Assay Kit; in which protein extracts were added to the prepared SDS-PAGE for protein electrophoresis. Next, protein extracts were transferred onto PVDF membranes and blocked with 5 % skim milk powder solution, while incubated with diluted primary antibodies overnight at 4°C and incubated with HRP-conjugated secondary antibody for 1 h. The signals were detected using an enhanced chemiluminescence kit, and quantified using ImageQuant 5.2 software.
4.4. Quantitative reverse transcription polymerase chain reaction (qRT-PCR)
RNA was isolated from colon of mice using TRIzol reagent and cDNA was generated from 1 μg total RNA using High-Capacity cDNA Reverse transcription kit. qRT-PCR analysis was carried out using SYBR green PCR mastermix and analyzed on a LightCycler® 1.5 Real-Time PCR System. Values were normalized to glyceraldehyde-3-phosphate dehydrogenase (Gapdh). The sequences of primers used for qRT-PCR are listed in Table 2.
4.5. 16S rRNA gene amplification and phylogenetic analysis
Fecal samples were collected and immediately transferred to a -80℃ freezer. Total bacterial DNA was extracted and purified using the QIAGEN mini stool kit (QIAGEN). The total DNA was extracted with the reported methods [52]. The primers 16s_illumina_V3F (5’-CCTACGGGNGGCWGCAG-3’) and 16s_illumina_V4R (5’-GACTACHVGGGTATCTAATCC-3’) were used for bacterial 16S rRNA variable region V3-V4. The 16S rRNA sequences were compared for similarity with the reference species of bacteria using the NCBI nucleotide database and PhiX Control Library. To begin, sequences were clustered into operational taxonomic units (OTUs) using UPARSE [53]. To generate taxonomic assignments, Bowtie2 was used to align sequencing reads against the collection of a 16S rRNA sequences database [53]. Principal component analysis (PCA) plot was performed using ClustVis software [54], which is a web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap from Oxford University.
4.6. Plasma, liver, and colonic bile acids analysis
Plasma (50 uL) was added to 150 μL of methanol with internal standard (DCA-d6) for protein precipitation and stay in -80°C for 10 min. Samples were centrifuged for 30 min at 12000 g at 4°C. The frozen liver and colon tissue were weighed and placed in homogenization tubes containing ceramic beads. The tissues were homogenized in a Precelly24 homogenizer for two times over 15 s at 6000 rpm with 30 s pause intervals with extraction solvent (150 uL) containing internal standard. After homogenization, samples were centrifuged for 30 min at 12000 g at 4°C. The supernatant was analyzed with UPLC/MS/MS [55].
4.7. Fecal SCFAs extraction and Derivatization
Thawed frozen fecal samples at room temperature, then combined 50 mg of fecal material with 1 mL of 10% isobutanol (J.T. baker 9044-01). The mixture was homogenized for 20 seconds at 6000 rpm, with two cycles and a 30-second pause in between, using a Precelly24 homogenizer (Bertin Technologies, Montigmy le Bretonnexux, France). Subsequently, the homogenized mixture was centrifuged for 30 minutes at 12,000 rpm at 4°C. Following centrifugation, 270 µL of the supernatant was transferred into a fresh glass vial. Sequentially, 50 µL of 20 mM NaOH and 160 µL of chloroform were added to the same glass vial for extraction. After vortexing for 5 minutes, the sample was centrifuged for 15 minutes at 1200 xg. The upper aqueous layer (200 µL) was then carefully transferred to a new glass vial for further derivatization. For both the calibration standards and the aqueous layer (200 µL), the following steps were taken: 70 µL of the internal standard (100 µM butyrate-d8), 80 µL of isobutanol, 100 µL of pyridine, and 200 µL of ultrapure water were added to the sample. Subsequently, 50 µL of isobutyl chloroformate was carefully introduced into the sample or standard solution. To release the gases generated by the reaction, the cap was left open for 20 seconds, then closed and mixed for 1 minute. The mixture was then sonicated for 5 minutes. Finally, 150 µL of hexane was added and thoroughly mixed. The vials were centrifuged at 1200 xg for 10 minutes. The upper hexane phase was transferred into an autosampler sample vial and securely capped for GC/MS analysis.
4.8. GC/MS analysis
The GC/MS analysis was conducted using a Bruker Scion 436 gas chromatograph/mass spectrometry system (Bruker Daltonics, Billerica, MA) equipped with a VF-5 ms capillary column (30 meters in length, 0.25 mm in diameter, and a 0.5 µm film thickness) from Agilent Technologies. Analyte quantification was performed in the selected ion monitoring (SIM) mode. The temperature settings for various components were as follows: the injector was set at 260°C, the ion source at 250°C, and the transfer line at 280°C. The helium carrier gas flowed at a rate of 1 mL/min. A 2 µl volume of derivatized sample was injected with a split ratio of 50:1. The temperature program for the column was as follows: it started at 40 °C and was maintained for 5 minutes, then ramped to 200 °C at a rate of 10 °C/min and held for 1 minute. Finally, the temperature was increased to 310 °C at a rate of 50 °C/min and maintained at this level for 3 minutes.
4.9. Statistical analysis
Results are presented as mean ± standard error of the mean (SEM). Statistical determined by the Student t test and analysis of variance with Student Newman-Keuls multiple-range test as appropriate. All p-values values < 0.05 were considered statistically significant.
Author Contributions
Conceptualization, H.-M. L. and T.-Y. L.; software, H.-M. L.; validation, H.-M. L., C.-W. Y., Z.-Y. C., H.-H. C., and T.-Y. L.; investigation, H.-M. L. and T.-Y. L.; resources, Z.-Y. C.; data curation, H.-M. L. and C.-W. Y.; writing—original draft preparation, H.-M. L.; writing—review and editing, H.-M. L., H.-H. C., and T.-Y. L.; supervision, Z.-Y. C. and T.-Y. L.; project administration, Z.-Y. C., H.-H. C., and T.-Y. L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science and Technology Council, Taipei, Taiwan (grant number NSTC 102-2320-B182-015-MY3, 103-2320-B182-002-MY3, and 106-2320-B-182-005-MY3, 109-2320-B-182-023-MY3), and the Chang Gung Memorial Hospital, Linkou, Taiwan (grant number CMRPD1B0261, CMRPD1B0262, CMRPD1D0351, CMRPD1D0352).
Institutional Review Board Statement
This study was approved by the Animal Care and Use Committee of Chang Gung University Institutional Animal Care and Use Committee (protocol code CGU111-202).
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhou:, X.; Cao, L.; Jiang, C.; Xie, Y.; Cheng, X.; Krausz, K.W.; Qi, Y.; Sun, L.; Shah, Y.M.; Gonzalez, F.J.; et al. PPARα-UGT axis activation represses intestinal FXR-FGF15 feedback signalling and exacerbates experimental colitis. Nat. commun. 2014, 5, 4573. [Google Scholar] [CrossRef] [PubMed]
- Vandel, J.; Dubois-Chevalier, J.; Gheeraert, C.; Derudas, B.; Raverdy, V.; Thuillier, D.; Gaal, L.; Francque, S.; Pattou, F.; Staels, B.; et al. Hepatic Molecular Signatures Highlight the Sexual Dimorphism of Nonalcoholic Steatohepatitis (NASH). Hepatology 2021, 73, 920–936. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Tanaka, N.; Fukami, T.; Xie, C.; Yagai, T.; Kim, D.; Velenosi, T.J.; Yan, T.; Krausz, K.W.; Levi, M.; et al. Role of Farnesoid X Receptor and Bile Acids in Hepatic Tumor Development. Hepatol. commun. 2018, 2, 1567–1582. [Google Scholar] [CrossRef] [PubMed]
- Maran, R.R.; Thomas, A.; Roth, M.; Sheng, Z.; Esterly, N.; Pinson, D.; Gao, X.; Zhang, Y.; Ganapathy, V.; Gonzalez, F.J.; et al. Farnesoid X receptor deficiency in mice leads to increased intestinal epithelial cell proliferation and tumor development. J. Pharmacol. Exp. Tther. 2009, 328, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.; Thomas, A.; Edwards, G.; Jaseja, R.; Guo, G.L.; Apte, U. Increased activation of the Wnt/β-catenin pathway in spontaneous hepatocellular carcinoma observed in farnesoid X receptor knockout mice. J. Pharmacol. Exp. Ther. 2011, 338, 12–21. [Google Scholar] [CrossRef]
- Liu, N.; Meng, Z.; Lou, G.; Zhou, W.; Wang, X.; Zhang, Y.; Zhang, L.; Liu, X.; Yen, Y.; Lai, L.; et al. Hepatocarcinogenesis in FXR-/- mice mimics human HCC progression that operates through HNF1α regulation of FXR expression. Mol. Endocrinol. 2012, 26, 775–785. [Google Scholar] [CrossRef]
- Dolegowska, K.; Marchelek-Mysliwiec, M.; Nowosiad-Magda, M.; Slawinski, M.; Dolegowska, B. FGF19 subfamily members: FGF19 and FGF21. J. Physiol. Biochem. 2019, 75, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Kim, Y.C.; Byun, S.; Kim, D.H.; Seok, S.; Suino-Powell, K.; Xu, H.E.; Kemper, B.; Kemper, J.K. FXR Primes the Liver for Intestinal FGF15 Signaling by Transient Induction of β-Klotho. Mol. Endocrinol. 2016, 30, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Wong, B.S.; Camilleri, M.; Carlson, P.J.; Guicciardi, M.E.; Burton, D.; McKinzie, S.; Rao, A.S.; Zinsmeister, A.R.; Gores, G.J. A Klothoβ variant mediates protein stability and associates with colon transit in irritable bowel syndrome with diarrhea. Gastroenterology 2011, 140, 1934–1942. [Google Scholar] [CrossRef]
- Hou, Z.; Ding, Q.; Li, Y.; Zhao, Z.; Yan, F.; Li, Y.; Wang, X.; Xu, J.; Chen, W.; Wu, G.; et al. Intestinal epithelial β Klotho is a critical protective factor in alcohol-induced intestinal barrier dysfunction and liver injury. EBioMedicine 2022, 82, 104181. [Google Scholar] [CrossRef]
- Kuro, O.M. The Klotho proteins in health and disease. Nat. Rev. Nephrol. 2019, 15, 27–44. [Google Scholar] [CrossRef]
- Quarles, L.D. Fibroblast growth factor 23 and α-Klotho co-dependent and independent functions. Curr. Opin. Nephrol. Hypertens. 2019, 28, 16–25. [Google Scholar] [CrossRef]
- Richter, B.; Faul, C. FGF23 Actions on Target Tissues-With and Without Klotho. Front. Endocrinol. 2018, 9, 189. [Google Scholar] [CrossRef]
- Li, S.; Yu, L.; He, A.; Liu, Q. Klotho Inhibits Unilateral Ureteral Obstruction-Induced Endothelial-to-Mesenchymal Transition via TGF-β1/Smad2/Snail1 Signaling in Mice. Front. Pharmacol. 2019, 10, 348. [Google Scholar] [CrossRef]
- Ligumsky, H.; Merenbakh-Lamin, K.; Keren-Khadmy, N.; Wolf, I.; Rubinek, T. The role of α-klotho in human cancer: molecular and clinical aspects. Oncogene 2022, 41, 4487–4497. [Google Scholar] [CrossRef]
- Li, X.X.; Huang, L.Y.; Peng, J.J.; Liang, L.; Shi, D.B.; Zheng, H.T.; Cai, S.J. Klotho suppresses growth and invasion of colon cancer cells through inhibition of IGF1R-mediated PI3K/AKT pathway. Int. J. Oncol 2014, 45, 611–618. [Google Scholar] [CrossRef]
- Degirolamo, C.; Rainaldi, S.; Bovenga, F.; Murzilli, S.; Moschetta, A. Microbiota modification with probiotics induces hepatic bile acid synthesis via downregulation of the Fxr-Fgf15 axis in mice. Cell Rep. 2014, 7, 12–18. [Google Scholar] [CrossRef]
- Fox, A.C.; McConnell, K.W.; Yoseph, B.P.; Breed, E.; Liang, Z.; Clark, A.T.; O'Donnell, D.; Zee-Cheng, B.; Jung, E.; Dominguez, J.A.; et al. The endogenous bacteria alter gut epithelial apoptosis and decrease mortality following Pseudomonas aeruginosa pneumonia. Shock 2012, 38, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Kho, Z.Y.; Lal, S.K. The Human Gut Microbiome - A Potential Controller of Wellness and Disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [PubMed]
- Zaborin, A.; Smith, D.; Garfield, K.; Quensen, J.; Shakhsheer, B.; Kade, M.; Tirrell, M.; Tiedje, J.; Gilbert, J.A.; Zaborina, O.; et al. Membership and behavior of ultra-low-diversity pathogen communities present in the gut of humans during prolonged critical illness. mBio 2014, 5, e01361–01314. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.S.; Xu, S.; Nambiar, P.; Dewhirst, F.E.; Fox, J.G. Enterohepatic Helicobacter species are prevalent in mice from commercial and academic institutions in Asia, Europe, and North America. J. Clin. Microbiol. 2007, 45, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Whary, M.T.; Fox, J.G. Natural and experimental Helicobacter infections. Comp. Med. 2004, 54, 128–158. [Google Scholar] [PubMed]
- Swennes, A.G.; Sheh, A.; Parry, N.M.; Muthupalani, S.; Lertpiriyapong, K.; García, A.; Fox, J.G. Helicobacter hepaticus infection promotes hepatitis and preneoplastic foci in farnesoid X receptor (FXR) deficient mice. PloS one 2014, 9, e106764. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Houten, S.M.; Mataki, C.; Christoffolete, M.A.; Kim, B.W.; Sato, H.; Messaddeq, N.; Harney, J.W.; Ezaki, O.; Kodama, T.; et al. Bile acids induce energy expenditure by promoting intracellular thyroid hormone activation. Nature 2006, 439, 484–489. [Google Scholar] [CrossRef]
- Liu, H.M.; Liao, J.F.; Lee, T.Y. Farnesoid X receptor agonist GW4064 ameliorates lipopolysaccharide-induced ileocolitis through TLR4/MyD88 pathway related mitochondrial dysfunction in mice. Biochem. Biophys Res. Commun. 2017, 490, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Jena, P.K.; Hu, Y.; Liu, H.X.; Nagar, N.; Kalanetra, K.M.; French, S.W.; French, S.W.; Mills, D.A.; Wan, Y.Y. Hepatic inflammation caused by dysregulated bile acid synthesis is reversible by butyrate supplementation. J. Pathol. 2017, 243, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Wolf, P.G.; Gaskins, H.R. Taurocholic acid metabolism by gut microbes and colon cancer. Gut microbes 2016, 7, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Dawson, P.A.; Lan, T.; Rao, A. Bile acid transporters. J. Lipid Res. 2009, 50, 2340–2357. [Google Scholar] [CrossRef]
- Bailey, A.M.; Zhan, L.; Maru, D.; Shureiqi, I.; Pickering, C.R.; Kiriakova, G.; Izzo, J.; He, N.; Wei, C.; Baladandayuthapani, V.; et al. FXR silencing in human colon cancer by DNA methylation and KRAS signaling. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G48–G58. [Google Scholar] [CrossRef]
- Somm, E.; Henry, H.; Bruce, S.J.; Bonnet, N.; Montandon, S.A.; Niederländer, N.J.; Messina, A.; Aeby, S.; Rosikiewicz, M.; Fajas, L.; et al. β-Klotho deficiency shifts the gut-liver bile acid axis and induces hepatic alterations in mice. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E833–E847. [Google Scholar] [CrossRef]
- Stamatovic, S.M.; Johnson, A.M.; Sladojevic, N.; Keep, R.F.; Andjelkovic, A.V. Endocytosis of tight junction proteins and the regulation of degradation and recycling. Ann. N. Y. Acad. Sci. 2017, 1397, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Lax, S.; Schauer, G.; Prein, K.; Kapitan, M.; Silbert, D.; Berghold, A.; Berger, A.; Trauner, M. Expression of the nuclear bile acid receptor/farnesoid X receptor is reduced in human colon carcinoma compared to nonneoplastic mucosa independent from site and may be associated with adverse prognosis. Int. J. Cancer 2012, 130, 2232–2239. [Google Scholar] [CrossRef] [PubMed]
- Frankenberg, T.; Rao, A.; Chen, F.; Haywood, J.; Shneider, B.L.; Dawson, P.A. Regulation of the mouse organic solute transporter alpha-beta, Ostalpha-Ostbeta, by bile acids. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G912–G922. [Google Scholar] [CrossRef] [PubMed]
- De Gottardi, A.; Touri, F.; Maurer, C.A.; Perez, A.; Maurhofer, O.; Ventre, G.; Bentzen, C.L.; Niesor, E.J.; Dufour, J.F. The bile acid nuclear receptor FXR and the bile acid binding protein IBABP are differently expressed in colon cancer. Dig. Dis. and Sci. 2004, 49, 982–989. [Google Scholar] [CrossRef] [PubMed]
- De Magalhaes Filho, C.D.; Downes, M.; Evans, R. Bile Acid Analog Intercepts Liver Fibrosis. Cell 2016, 166, 789. [Google Scholar] [CrossRef] [PubMed]
- Modica, S.; Murzilli, S.; Salvatore, L.; Schmidt, D.R.; Moschetta, A. Nuclear bile acid receptor FXR protects against intestinal tumorigenesis. Cancer Res. 2008, 68, 9589–9594. [Google Scholar] [CrossRef]
- Fu, T.; Choi, S.E.; Kim, D.H.; Seok, S.; Suino-Powell, K.M.; Xu, H.E.; Kemper, J.K. Aberrantly elevated microRNA-34a in obesity attenuates hepatic responses to FGF19 by targeting a membrane coreceptor β-Klotho. Proc. Natl. Acad. Sci. U S A 2012, 109, 16137–16142. [Google Scholar] [CrossRef] [PubMed]
- Parséus, A.; Sommer, N.; Sommer, F.; Caesar, R.; Molinaro, A.; Ståhlman, M.; Greiner, T.U.; Perkins, R.; Bäckhed, F. Microbiota-induced obesity requires farnesoid X receptor. Gut 2017, 66, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, A.; Gouge, J.; Kontovounisios, C.; Nikolaou, S.; Ashworth, A.; Lim, K.; Chong, I. Klotho and the Treatment of Human Malignancies. Cancers (Basel) 2020, 12, 1665. [Google Scholar] [CrossRef]
- Arbel Rubinstein, T.; Shahmoon, S.; Zigmond, E.; Etan, T.; Merenbakh-Lamin, K.; Pasmanik-Chor, M.; Har-Zahav, G.; Barshack, I.; Vainer, G.W.; Skalka, N.; et al. Klotho suppresses colorectal cancer through modulation of the unfolded protein response. Oncogene 2019, 38, 794–807. [Google Scholar] [CrossRef]
- Jena, P.K.; Sheng, L.; Liu, H.X.; Kalanetra, K.M.; Mirsoian, A.; Murphy, W.J.; French, S.W.; Krishnan, V.V.; Mills, D.A.; Wan, Y.Y. Western Diet-Induced Dysbiosis in Farnesoid X Receptor Knockout Mice Causes Persistent Hepatic Inflammation after Antibiotic Treatment. Am. J. Pathol. 2017, 187, 1800–1813. [Google Scholar] [CrossRef] [PubMed]
- Wexler, H.M. Bacteroides: the good, the bad, and the nitty-gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Schwab, C.; Milinovich, G.; Reichert, J.; Ben Mahfoudh, K.; Decker, T.; Engel, M.; Hai, B.; Hainzl, E.; Heider, S.; et al. Phylotype-level 16S rRNA analysis reveals new bacterial indicators of health state in acute murine colitis. ISME J. 2012, 6, 2091–2106. [Google Scholar] [CrossRef] [PubMed]
- Che, L.; Hu, Q.; Wang, R.; Zhang, D.; Liu, C.; Zhang, Y.; Xin, G.; Fang, Z.; Lin, Y.; Xu, S.; et al. Inter-correlated gut microbiota and SCFAs changes upon antibiotics exposure links with rapid body-mass gain in weaned piglet model. J. Nutr. Biochem 2019, 74, 108246. [Google Scholar] [CrossRef] [PubMed]
- Che, L.; Hu, Q.; Wang, R.; Zhang, D.; Liu, C.; Zhang, Y.; Xin, G.; Fang, Z.; Lin, Y.; Xu, S.; et al. Inter-correlated gut microbiota and SCFAs changes upon antibiotics exposure links with rapid body-mass gain in weaned piglet model. J. Nutr. Biochem. 2019, 74, 108246. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat Rev Gastroenterol Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Belcheva, A.; Irrazabal, T.; Robertson, S.J.; Streutker, C.; Maughan, H.; Rubino, S.; Moriyama, E.H.; Copeland, J.K.; Surendra, A.; Kumar, S.; et al. Gut microbial metabolism drives transformation of MSH2-deficient colon epithelial cells. Cell 2014, 158, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Hamlin, S.K.; Safratowich, B.D.; Cheng, W.H.; Johnson, L.K. Superior inhibitory efficacy of butyrate over propionate and acetate against human colon cancer cell proliferation via cell cycle arrest and apoptosis: linking dietary fiber to cancer prevention. Nutr. Res. 2020, 83, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Pastorelli, L.; Dozio, E.; Pisani, L.F.; Boscolo-Anzoletti, M.; Vianello, E.; Munizio, N.; Spina, L.; Tontini, G.E.; Peyvandi, F.; Corsi Romanelli, M.M.; et al. Procoagulatory state in inflammatory bowel diseases is promoted by impaired intestinal barrier function. Gastroenterol. Res. Pract. 2015, 2015, 189341. [Google Scholar] [CrossRef]
- Hotta, T.; Yoshida, N.; Yoshikawa, T.; Sugino, S.; Kondo, M. Lipopolysaccharide-induced colitis in rabbits. Res. Exp. Med. 1986, 186, 61–69. [Google Scholar] [CrossRef]
- Yeh, C.C.; Liu, H.M.; Lee, M.C.; Leu, Y.L.; Chiang, W.H.; Chang, H.H.; Lee, T.Y. Phytochemical-rich herbal formula ATG-125 protects against sucrose-induced gastrocnemius muscle atrophy by rescuing Akt signaling and improving mitochondrial dysfunction in young adult mice. Mol. Med. Rep. 2022, 25, 57. [Google Scholar] [CrossRef] [PubMed]
- Mirsepasi, H.; Persson, S.; Struve, C.; Andersen, L.O.; Petersen, A.M.; Krogfelt, K.A. Microbial diversity in fecal samples depends on DNA extraction method: easyMag DNA extraction compared to QIAamp DNA stool mini kit extraction. BMC Res. Notes 2014, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: a web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kaneko, R.; Nomura, M.; Naito, H.; Kitamori, K.; Nakajima, T.; Ogawa, T.; Hattori, H.; Seno, H.; Ishii, A. Simple and rapid quantitation of 21 bile acids in rat serum and liver by UPLC-MS-MS: effect of high fat diet on glycine conjugates of rat bile acids. Nagoya J. Med. Sci.i 2013, 75, 57–71. [Google Scholar]
Figure 1.
GW4064-dependent FXR activation ameliorates histological characteristics of tight junctions. (A) Representative hematoxylin and eosin staining and immunohistochemistry of FXR, Claudin-1, and ZO-1 staining for colon sections are shown. Scale bar: 50 μm. The (B, C) FXR, (D, E) Claudin-1, and ZO-1 protein were determined by Western blot. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 1.
GW4064-dependent FXR activation ameliorates histological characteristics of tight junctions. (A) Representative hematoxylin and eosin staining and immunohistochemistry of FXR, Claudin-1, and ZO-1 staining for colon sections are shown. Scale bar: 50 μm. The (B, C) FXR, (D, E) Claudin-1, and ZO-1 protein were determined by Western blot. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
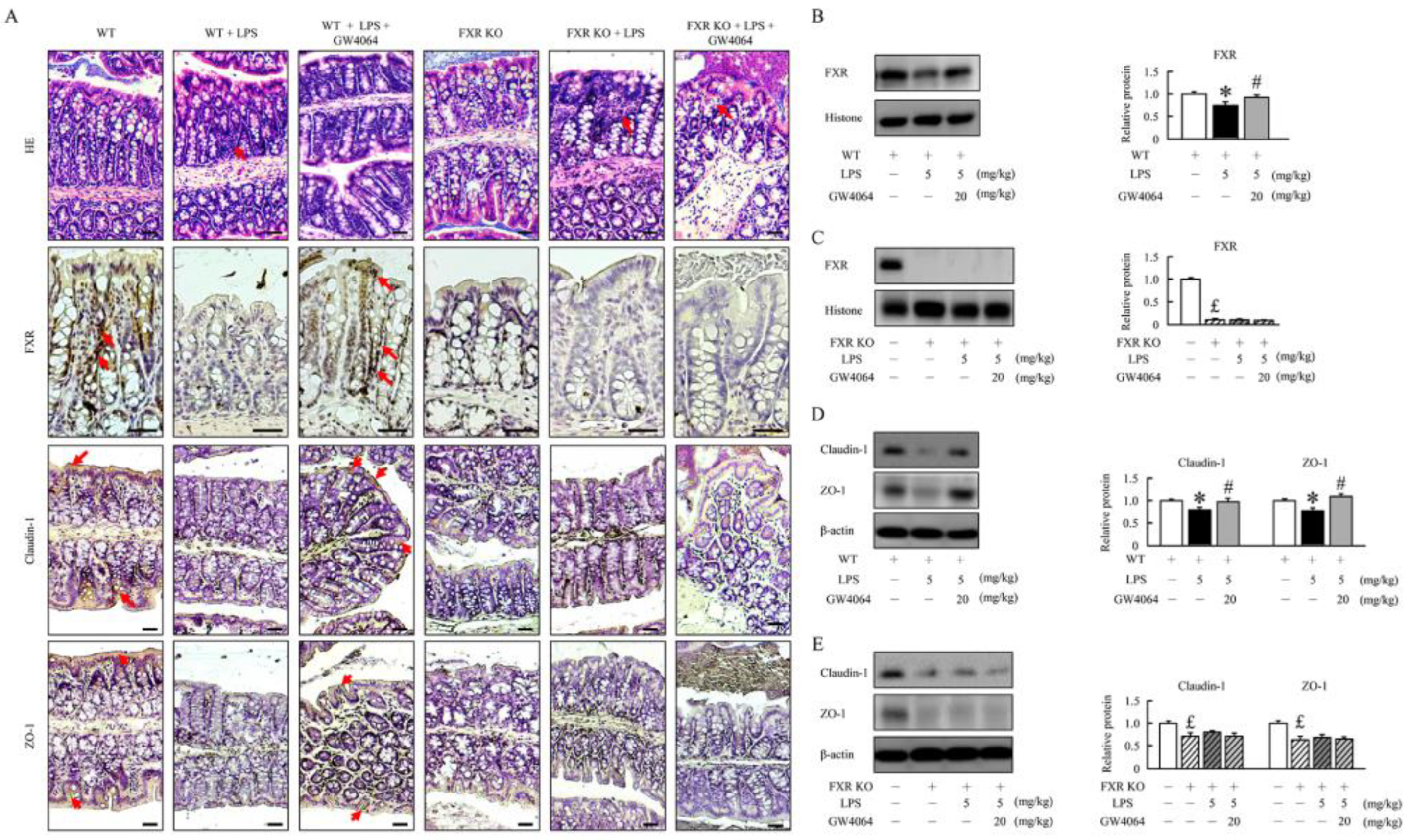
Figure 2.
GW4064 affects bile acids profile. (A) Schematic diagram of the experiment. The concentration of (B) plasma, (C) liver, (D) colon bile acids, and (E-H) individual bile acids in LPS-treated wild-type (WT) and FXR knockout (KO) mice with and without GW4064. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 2.
GW4064 affects bile acids profile. (A) Schematic diagram of the experiment. The concentration of (B) plasma, (C) liver, (D) colon bile acids, and (E-H) individual bile acids in LPS-treated wild-type (WT) and FXR knockout (KO) mice with and without GW4064. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
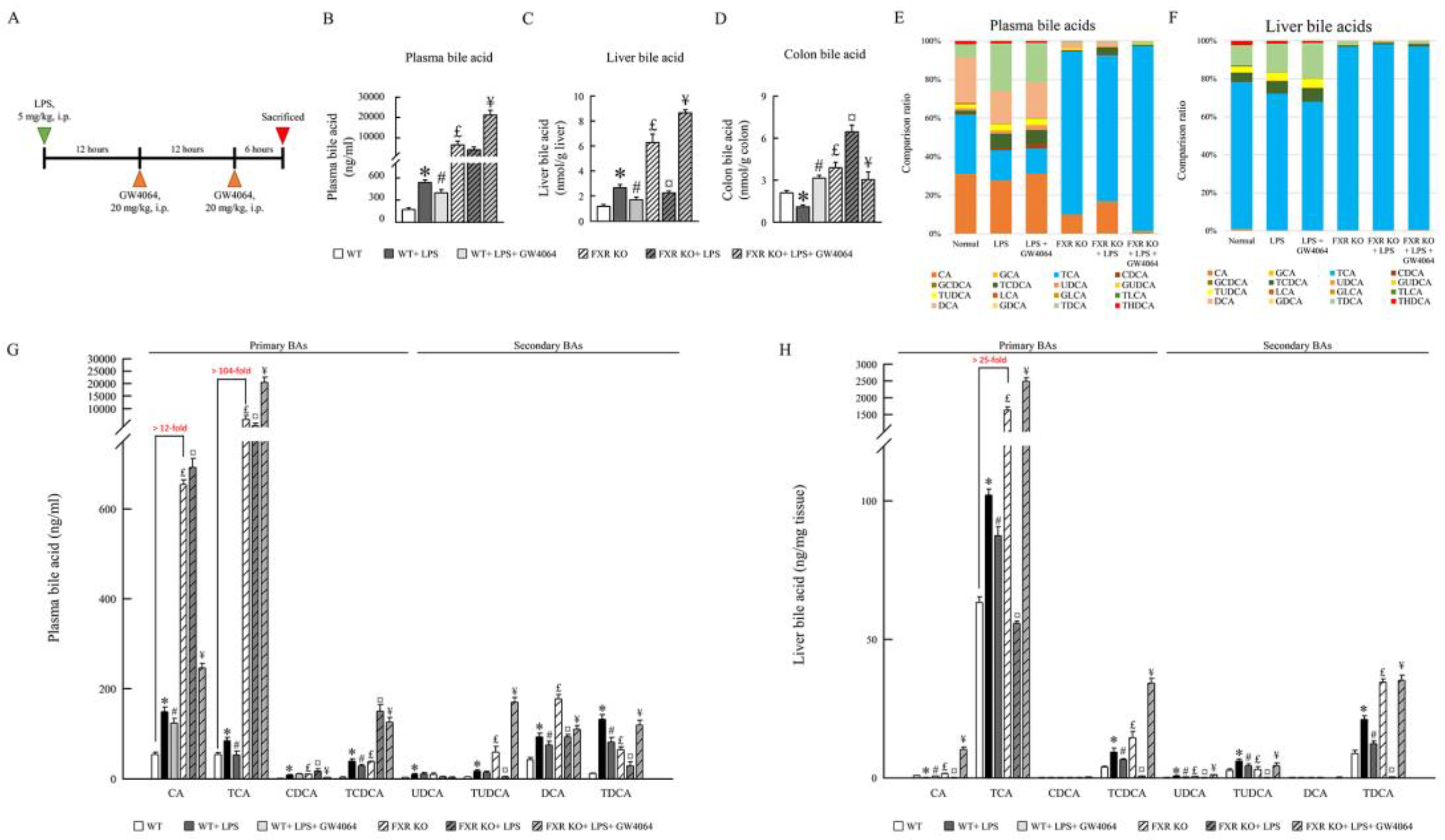
Figure 3.
The effects of GW4064 on bile acid receptors and transporters expression. Immunohistochemistry of colon sections for (A) MRP3, OATP1, (B) OSTβ, and ASBT staining. Scale bar: 100 μm. (C, D) The MRP2, MRP3, OSTβ, ASBT, and OATP1 proteins were determined by Western blot. qRT-PCR analyses of colonic mRNA expression of (E) Fxr, Pxr, Car, Cyp3a11, Sult2a1, and (F) Oatp2b1, Asbt, Ibabp, Ostα, Ostβ, and (G) Mrp2, Mrp3, Mdr1b, Mdr2, and Bcrp. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 3.
The effects of GW4064 on bile acid receptors and transporters expression. Immunohistochemistry of colon sections for (A) MRP3, OATP1, (B) OSTβ, and ASBT staining. Scale bar: 100 μm. (C, D) The MRP2, MRP3, OSTβ, ASBT, and OATP1 proteins were determined by Western blot. qRT-PCR analyses of colonic mRNA expression of (E) Fxr, Pxr, Car, Cyp3a11, Sult2a1, and (F) Oatp2b1, Asbt, Ibabp, Ostα, Ostβ, and (G) Mrp2, Mrp3, Mdr1b, Mdr2, and Bcrp. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
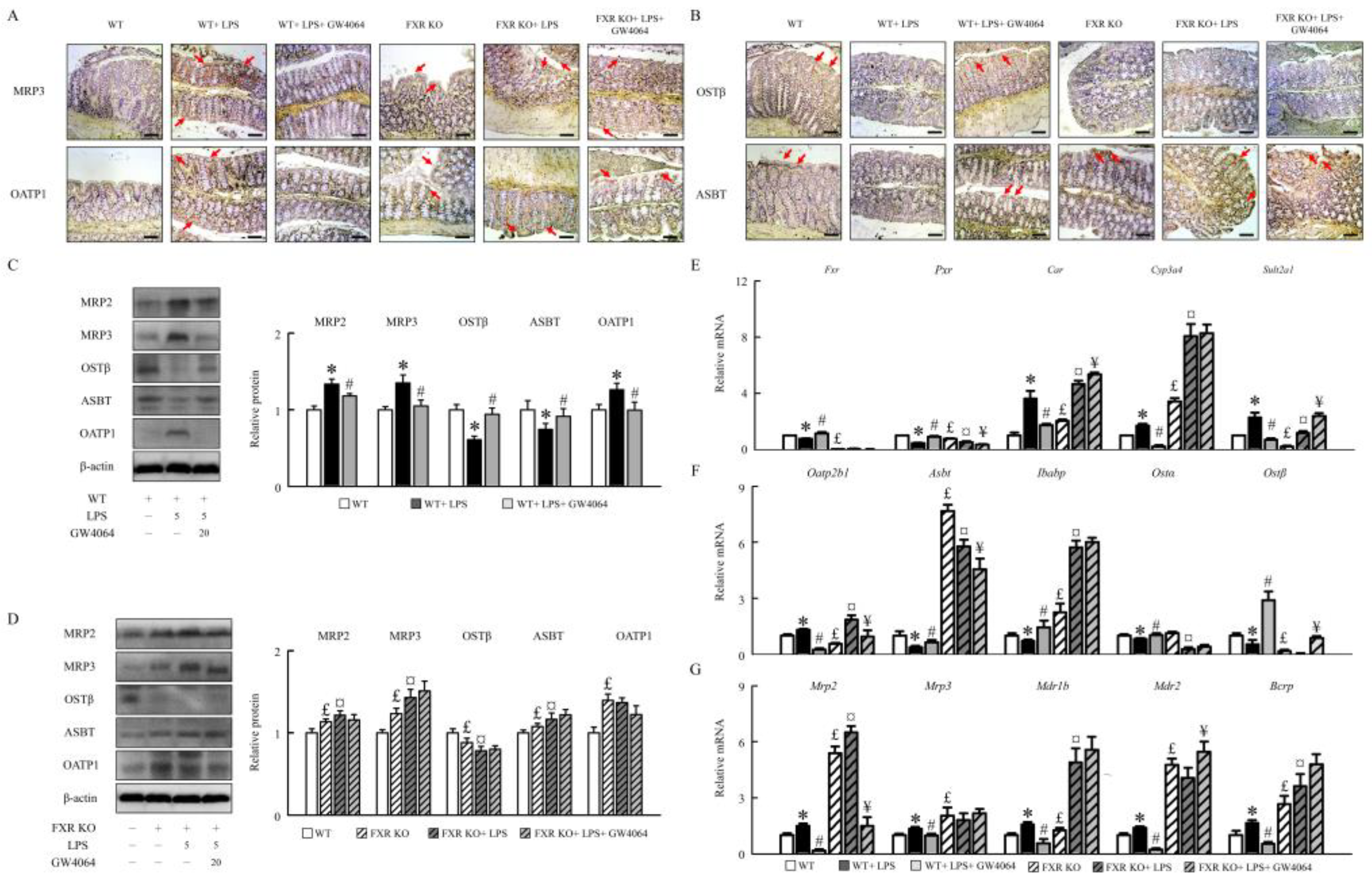
Figure 4.
GW4064 regulates αKlotho/βKlotho/FGF19/FGF21/FGF23 pathway. Immunohistochemistry of colon sections for (A) αKlotho, βKlotho, and (B) FGF19, FGF21, FGF23 staining. Scale bar: 50 μm. The (C) αKlotho, βKlotho, and (D) FGF19, FGF21, FGF23 protein levels were determined by Western blot. *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 4.
GW4064 regulates αKlotho/βKlotho/FGF19/FGF21/FGF23 pathway. Immunohistochemistry of colon sections for (A) αKlotho, βKlotho, and (B) FGF19, FGF21, FGF23 staining. Scale bar: 50 μm. The (C) αKlotho, βKlotho, and (D) FGF19, FGF21, FGF23 protein levels were determined by Western blot. *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
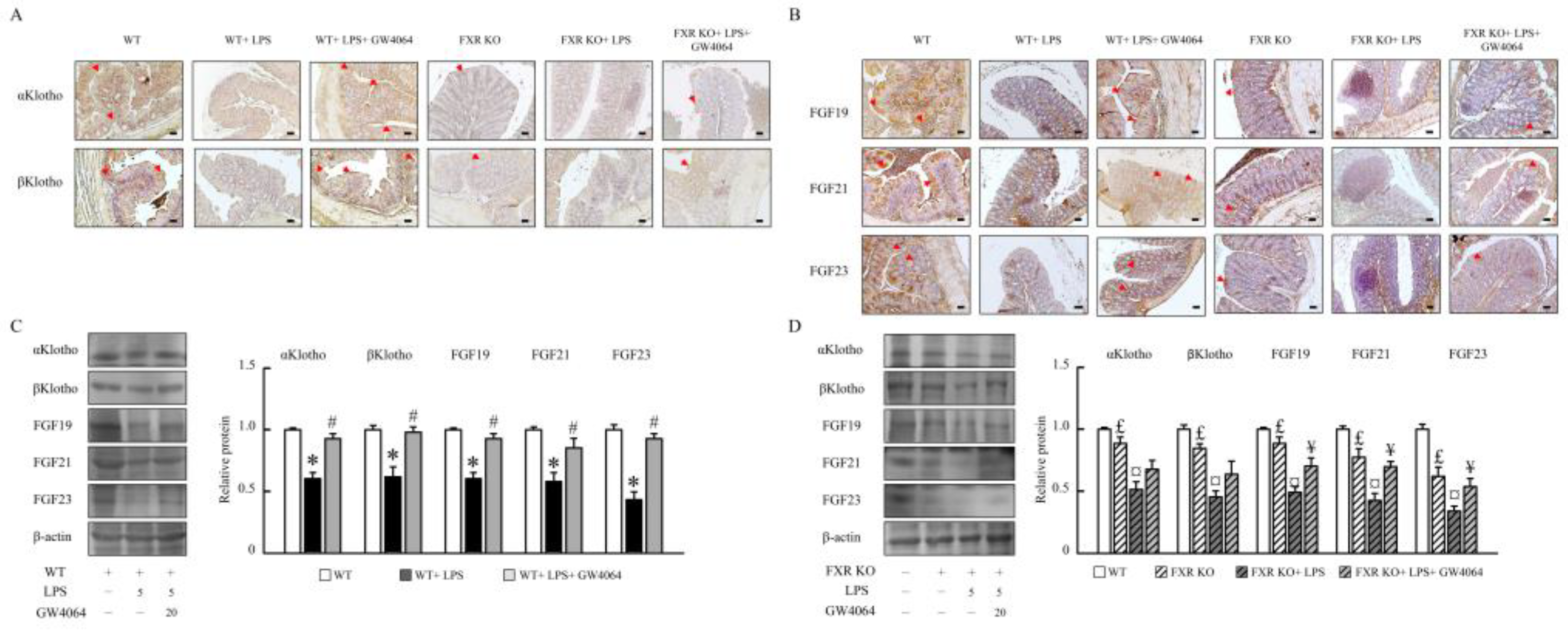
Figure 5.
FXR deletion exhibits signs of TLR4-mediated inflammatory response, apoptosis of IECs, and leads to ER stress. qRT-PCR analyses of colonic mRNA levels of (A) Tnfα, Ifnγ, Il-1β, pro-caspase-1, Asc, Nlrp3, Pannexin-1 and (B) Atf4, Atf6, Grp78, Chop and Xbp1s. (C) Immunohistochemistry of colon sections for TLR4 and Caspase 3 staining. Scale bar: 100 μm. (D, E) The TLR4, MyD88 and NF-κB protein were determined by Western blot. (F, G) The Caspase 3 protein levels were determined by Western blot. (H) Immunohistochemistry of colon sections for β-catenin and c-Myc staining. Scale bar: 100 μm. (I, J) The β-catenin, c-Myc, (K, L) TGFβRII, ICAM, VCAM, VEGF1, VEGFR1, and MMP9 protein levels were determined by Western blot. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 5.
FXR deletion exhibits signs of TLR4-mediated inflammatory response, apoptosis of IECs, and leads to ER stress. qRT-PCR analyses of colonic mRNA levels of (A) Tnfα, Ifnγ, Il-1β, pro-caspase-1, Asc, Nlrp3, Pannexin-1 and (B) Atf4, Atf6, Grp78, Chop and Xbp1s. (C) Immunohistochemistry of colon sections for TLR4 and Caspase 3 staining. Scale bar: 100 μm. (D, E) The TLR4, MyD88 and NF-κB protein were determined by Western blot. (F, G) The Caspase 3 protein levels were determined by Western blot. (H) Immunohistochemistry of colon sections for β-catenin and c-Myc staining. Scale bar: 100 μm. (I, J) The β-catenin, c-Myc, (K, L) TGFβRII, ICAM, VCAM, VEGF1, VEGFR1, and MMP9 protein levels were determined by Western blot. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
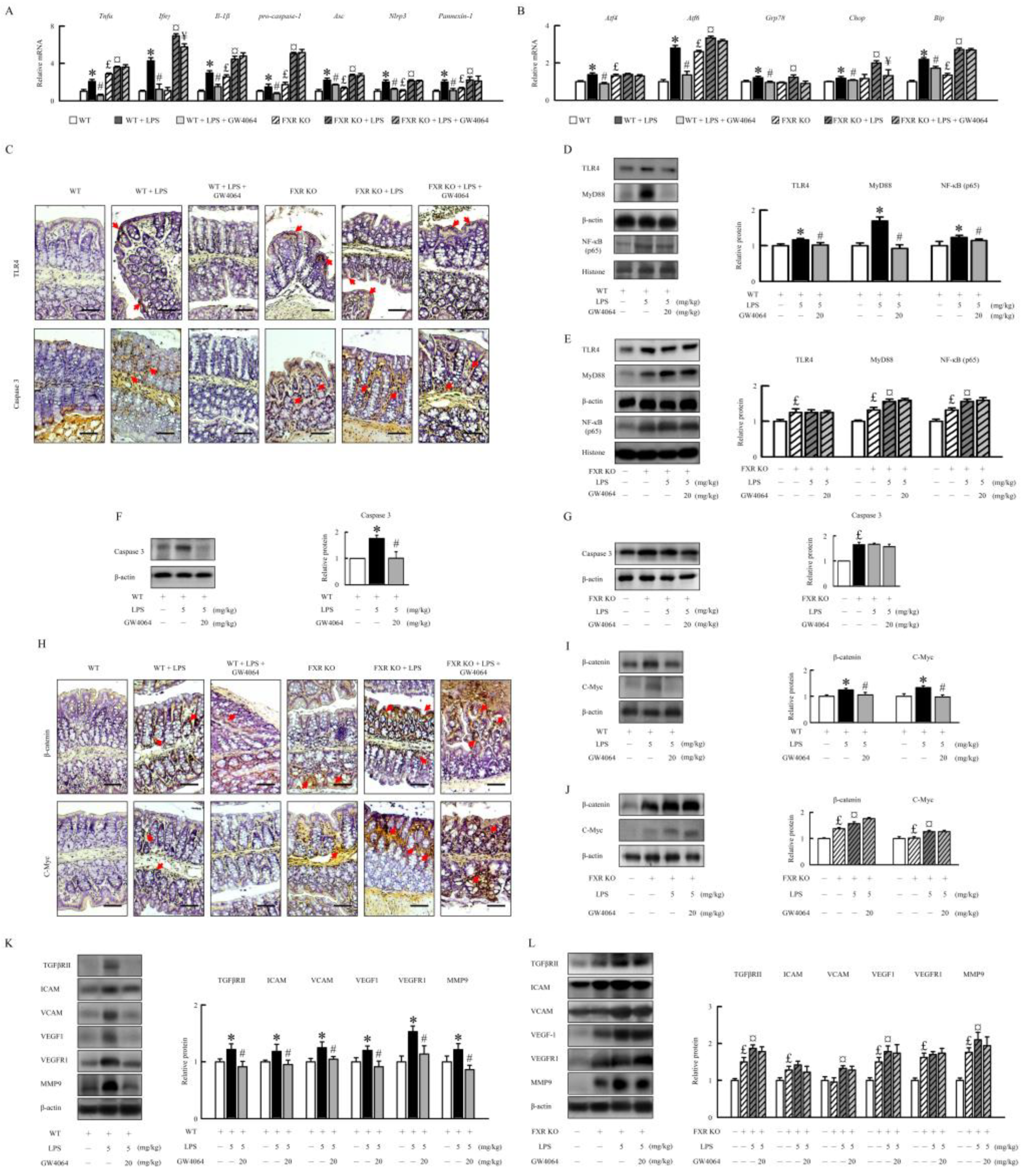
Figure 6.
GW4064 alters colonic stem cell proliferation. (A) Double immunofluorescence staining of LGR5 (red) and CD44 (green); (B) PCNA (red) and CD44 (green); BrdU (red) and (C) CD34 (green); (D) BrdU (red) and CD133 (green); (E) Cyclin D1 (red) and GCSF (green) with DAPI (blue) in colon. Scale bar = 50 μm. (F) qRT-PCR analyses of colonic mRNA level of Lgr5, Olfm4, and Cyclin D1. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 6.
GW4064 alters colonic stem cell proliferation. (A) Double immunofluorescence staining of LGR5 (red) and CD44 (green); (B) PCNA (red) and CD44 (green); BrdU (red) and (C) CD34 (green); (D) BrdU (red) and CD133 (green); (E) Cyclin D1 (red) and GCSF (green) with DAPI (blue) in colon. Scale bar = 50 μm. (F) qRT-PCR analyses of colonic mRNA level of Lgr5, Olfm4, and Cyclin D1. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 7.
GW4064 alters intestinal microbial composition in mice. Alpha diversity measurements of the microbiota across locations. (A) Chao1 (B) and Shannon’s diversity index. PCA plot based on the abundance of bacterial gene sequences in fecal content. (C) Axes correspond to principal component 1 (x axis) and 2 (y axis). (D) Microbial community bar plot by phylum relative abundance (%). (E-I) Phylum levels of Proteobacteria, Verrucomicrobia, Firmicutes, Bacteroidetes, and Firmicutes/Bacteroidetes (F/B) ratio. (J) Microbial community bar plot by class relative abundance (%). (K-N) Class levels of Betaproteobacteria, Gammaproteobacteria, Clostridia and Verrucomicrobiae. (O) Microbial community bar plot by order relative abundance (%). (P-R) Order levels of Enterobacteriales, Verrucomicrobiales and Burkholderiales. (S) Genus levels of Bacteroides, Clostridium, and Escherichia. (T) Microbial community bar plot by species relative abundance (%). (U-Z) Species levels of Bacteroides thetaiotaomicron, Escherichia coli, Bacteroides acidifaciens, Akkermansia muciniphila, Clostridium saccharogumia, and Helicobacter hepaticus. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
Figure 7.
GW4064 alters intestinal microbial composition in mice. Alpha diversity measurements of the microbiota across locations. (A) Chao1 (B) and Shannon’s diversity index. PCA plot based on the abundance of bacterial gene sequences in fecal content. (C) Axes correspond to principal component 1 (x axis) and 2 (y axis). (D) Microbial community bar plot by phylum relative abundance (%). (E-I) Phylum levels of Proteobacteria, Verrucomicrobia, Firmicutes, Bacteroidetes, and Firmicutes/Bacteroidetes (F/B) ratio. (J) Microbial community bar plot by class relative abundance (%). (K-N) Class levels of Betaproteobacteria, Gammaproteobacteria, Clostridia and Verrucomicrobiae. (O) Microbial community bar plot by order relative abundance (%). (P-R) Order levels of Enterobacteriales, Verrucomicrobiales and Burkholderiales. (S) Genus levels of Bacteroides, Clostridium, and Escherichia. (T) Microbial community bar plot by species relative abundance (%). (U-Z) Species levels of Bacteroides thetaiotaomicron, Escherichia coli, Bacteroides acidifaciens, Akkermansia muciniphila, Clostridium saccharogumia, and Helicobacter hepaticus. Values are expressed as the mean ± standard error of the mean (n = 5 mice per group). *p < 0.05, WT vs WT+ LPS; #p < 0.05, WT + LPS vs WT + LPS + GW4064; £p < 0.05, WT vs FXR KO; ¤p < 0.05, FXR KO vs FXR KO + LPS; ¥p < 0.05, FXR KO + LPS vs FXR KO + LPS + GW4064.
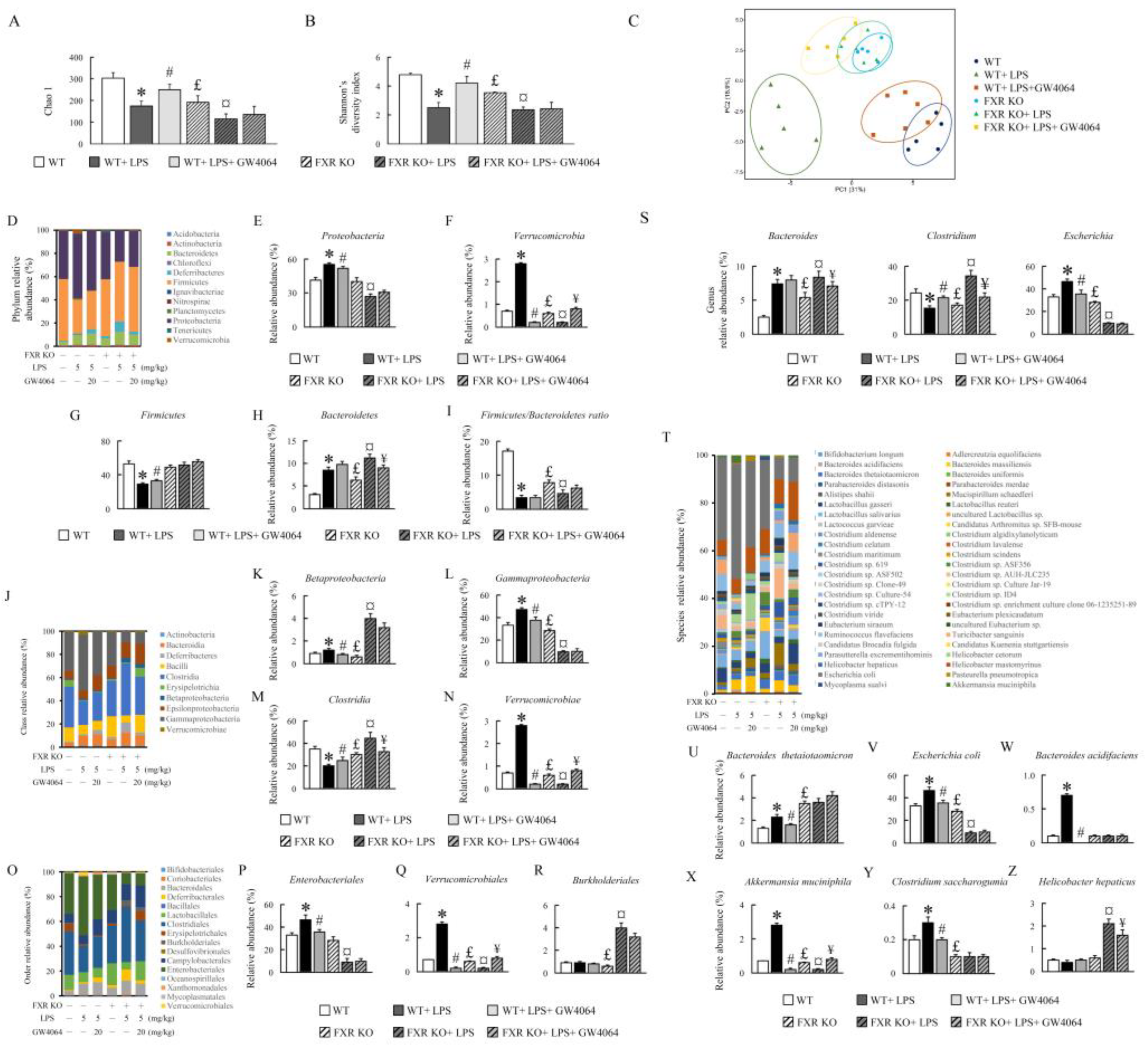

Table 1.
The antibodies used for IF, IHC, and Western blot.
| Antibodies | Company/State/ Country | Cat number |
| ASBT/SLC10A2 | Bioss Antibidies, Woburn, MA, USA | Cat# bs-4189R |
| αKlotho | Abcam, Trumpington, Cambridge, UK | Cat# ab181373 |
| β-actin | Proteintech, Rosemont, IL, USA | Cat# 60008-1-lg |
| β-catenin | Abcam, Trumpington, Cambridge, UK | Cat# ab6302 |
| βKlotho | GeneTex, Alton Pkwy Irvine, CA, USA | Cat# GTX122197 |
| Caspase 3 | Proteintech, Rosemont, IL, USA | Cat# 25546-1-AP |
| CD133 | Abcam, Trumpington, Cambridge, UK | Cat# ab19898 |
| CD34 | Abcam, Trumpington, Cambridge, UK | Cat# ab8158 |
| Claudin-1 | Abcam, Trumpington, Cambridge, UK | Cat# ab15098 |
| C-Myc | Abcam, Trumpington, Cambridge, UK | Cat# ab32 |
| FGF19 | Abcam, Trumpington, Cambridge, UK | Cat# ab225942 |
| FGF21 | Abcam, Trumpington, Cambridge, UK | Cat# ab171941 |
| FGF23 | Abcam, Trumpington, Cambridge, UK | Cat# ab307421 |
| FXR | Abcam, Trumpington, Cambridge, UK | Cat# ab129089 |
| Histone | Santa Cruz Biotechnology, Santa Cruz, California, USA | Cat# sc-56695 |
| ICAM | Thermo Fisher Scientific, Rockford, IL, USA | Cat# MA5407 |
| LGR5 | Abcam, Trumpington, Cambridge, UK | Cat# ab75850 |
| MMP9 | Merck Millipore, Burlington, Massachusetts, USA | Cat# NP_004985 |
| MRP2 | Thermo Fisher Scientific, Rockford, IL, USA | Cat# MA5-15700 |
| MRP3 | Biocompare, South San Francisco, CA, USA | Cat# L1010 |
| MyD88 | Abcam, Trumpington, Cambridge, UK | Cat# ab28763 |
| NF-κB | Santa Cruz Biotechnology, Santa Cruz, California, USA | Cat# sc-8008 |
| OATP/SLCO1A2 | Merck Millipore, Burlington, Massachusetts, USA | Cat# SAB4502814 |
| OSTβ | Biorbyt, Cambridge, UK | Cat# orb1964 |
| PCNA | Abcam, Trumpington, Cambridge, UK | Cat# ab18197 |
| TGFβRII | Thermo Fisher Scientific, Rockford, IL, USA | Cat# PA5-36115 |
| TLR4 | Thermo Fisher Scientific, Rockford, IL, USA | Cat# MA5-16216 |
| VCAM | Abcam, Trumpington, Cambridge, UK | Cat# ab78712 |
| VEGF | Boster Biological Technology, Pleasanton, CA, USA | Cat# PB9071 |
| VEGFR1 | Abcam, Trumpington, Cambridge, UK | Cat# ab32152 |
| ZO-1 | Abcam, Trumpington, Cambridge, UK, | Cat# ab59720 |
| Goat anti-mouse IgG | MilliporeSigma, Burlington, Massachusetts | Cat# Ap124P |
| Goat anti-rabbit IgG | Jackson ImmunoResearch Laboratories, West Grove, PA, USA | Cat# 111-065-003 |
| Goat anti-rat IgG | Jackson ImmunoResearch Laboratories, West Grove, PA, USA | Cat# 112-065-003 |
ASBT/SLC10A2, apical Sodium-dependent Bile acid Transporter; FGF19, fibroblast growth factor 19; FXR, farnesoid X receptor; ICAM, intercellular adhesion molecule; LGR5, leucine-rich repeat-containing G-protein coupled receptor 5; MMP9, matrix metalloproteinase 9; MRP2, multidrug resistance associated protein 2; MyD88, myeloid differentiation primary response protein 88; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; OATP/SLCO1A2, organic-anion-transporting polypeptides; OSTβ, organic solute transporter beta; PCNA, proliferating cell nuclear antigen; TGFβRII, rransforming growth factor beta receptor II; TLR4, toll like receptor 4; VCAM, vascular cell adhesion protein 1;VEGF, vascular endothelial growth factor; VEGFR1, vascular endothelial growth factor receptor 1; ZO-1, zonula occludens 1.
Table 2.
The sequences of primers used for qRT-PCR.
| Gene | Forward Primer sequences (5’ - 3’) | Reverse Primer sequences (5’ - 3’) |
| Nlrp3 | agccttccaggatcctcttc | cttgggcagcagtttctttc |
| Asc | gaagctgctgacagtgcaac | gccacagctccagactcttc |
| Panx-1 | ggccacggagtatgtgttct | tacagcagcccagcagtatg |
| Pro-casp 1 | agatggcacatttccaggac | gatcctccagcagcaacttc |
| Tnf-α | ttgacctcagcgctgagttg | cctgtagcccacgtcgtagc |
| Il-1β | gcaactgttcctgaactca | ctcggagcctgtagtgcag |
| Ifn-γ | tggcatagatgtggaagaaaagag | tgcaggattttcatgtcaccat |
| Mrp2 | gcttcccatggtgatctctt | atcatcgcttcccaggtact |
| Mrp3 | ctgggtcccctgcatctac | gccgtcttgagcctggataac |
| Mdr1b | gtgttaaaggggcgatgggcg | aggcttggccagacaacagctt |
| Mdr2 | atcctatgcactggccttctggt | gaaagcatcaatacagggggcag |
| Asbt | ggaactggctccaatatcctg | gttcccgagtcaacccacat |
| Ibabp | acaggacttcacctggtc | gcgctcataggtcacatc |
| Gapdh | tcaccaccatggagaaggc | gctaagcagttggtggtgca |
| Car | ggaggaccagatctcccttc | atttcattgccactcccaag |
| Cyp3a11 | gtgctcctagcaatcagctt | cagtgcctaaaaatggcagagg |
| Sult2a1 | ggaaggaccacgactcataac | gattcttcacaaggtttgtgttacc |
| Bcrp | tcgcagaaggagatgtgttgag | ccagaatagcattaaggccagg |
| Oatp2b1 | attgcaggcatcacacaaga | tagaagacctggcttttgcc |
| Ostα | gccaggcaggactcatatcaaa | ggcaactgagccagtggtaaga |
| Ostβ | caggaactgctggaagaaatgc | gcaggtcttctggtgtttctttgt |
| Grp78/Bip | acatggacctgttccgctcta | tggctccttgccattgaaga |
| Chop | tatctcatccccaggaaacg | gggcactgaccactctgttt |
| Xbp1s | ctgagtccgaatcaggtgcag | gtccatgggaagatgttctgg |
| Atf4 | atggccggctatggatgat | cgaagtcaaactctttcagatccatt |
| Lgr5 | caagccatgaccttggccctg | tttcccagggagtggattctatt |
| Atf6 | tgggcaggactatgaagtaatg | aggcttggccagacaacagctt |
| Cyclin D1 | ggggacaactcttaagtctcac | ccaataaaagaccaatctctc |
| Olfm4 | cagccactttccaatttcactg | gctggacatactccttcacctta |
Nlrp3, NACHT, LRR and PYD domains-containing protein 3; Asc, apoptosis-associated speck-like protein containing a caspase recruitment domain; Tnf-α, tumor necrosis factor-α; Il-1β, interleukin-1β; Ifn-γ, Interferon-γ; Fxr, farnesoid X receptor; Atf4, activating transcription factor; Grp78, glucose-regulated protein 78; Chop, CCAAT-enhancer-binding protein homologous protein (Bip, binding immunoglobulin protein); Xbp1s, X-box binding protein 1 Spliced; Gapdh, glyceraldehyde 3-phosphate dehydrogenase; Pxr, pregnane X receptor; Car, constitutive androstane receptor; Cyp3a11, Cytochrome P450 3A11; Sult2a1, sulfotransferase family 2A member 1; Mrp2, multidrug resistance-associated protein 2; Mdr1b, multidrug resistance protein; Bcrp, breast cancer resistance protein; Oatp2b1, organic anion-transporting polypeptide 2B1; Asbt, apical sodium dependent bile acid transporter; Ibabp, ileal bile acid-binding protein; Ost, organic solute transporter; Lgr5, Leucine-rich repeat-containing G-protein coupled receptor 5; Olfm4, olfactomedin 4.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



