
Submitted:
26 September 2023
Posted:
28 September 2023
You are already at the latest version
Abstract
The copolyester of 3-hydroxybutyrate (3HB) and 3-hydoxyhexanoate (3HHx), PHBHHx, is a bi-odegradable plastic that has a high flexibility, softness, wide process window and marine bio-degradability. PHBHHx is usually produced from structurally related carbon sources such as vegetable oils or fatty acids but not from inexpensive carbon sources like sugars. In the previous studies, we demonstrated that engineered strains of hydrogen-oxidizing bacterium, Cupriavidus necator synthesized PHBHHx with high cellular content from not only sugars but also from CO2 as the sole carbon source in the flask culture. In this study, highly efficient production of PHBHHx from CO2 was investigated by pH-stat jar cultivation of the recombinant C. necator strains with feeding the substrate gas mixture (H2/O2/CO2=8:1:1) to complete mineral medium in recycled-gas closed-circuit culture system. As the result with the strain MF01/pBPP-ccrMeJAc-emd, the dry cell mass and PHBHHx concentration reached up to 59.62 ± 3.18 g・L-1 and 49.31 ± 3.14 g・L-1, respectively after 212 h of jar cultivation with limited addition of ammonia and phosphate solutions. The 3HHx composition was close to 10 mol% which is suitable for practical applica-tions. It is expected that autotrophic cultivation of the recombinant C. necator can be a feasible process for mass production of PHBHHx from CO2.
Keywords:
polyhydroxyalkanoate
; PHBHHx
; CO2
; Cupriavidus necator
; hydrogen-oxidizing bacterium
; high cell density cultivation
; jar-cultivation
1. Introduction
Bacterial polyesters, polyhydroxyalkanoates (PHAs), are eco-friendly biodegradable polymers which are potential alternatives to petroleum-based polymeric materials. A homopolyester of (R)-3-hydroxybutyric acid, P(3HB), is well known to exhibit thermoplastic and biodegradable properties, therefore P(3HB) was expected to be put into practical uses. Unfortunately, P(3HB) is stiff and poor in impact strength [1] and has a very narrow “processing window” as the melting temperature (Tm, 170°C –180 °C) is close to the thermal degradation temperature (180°C–190°C) [2]. Copolyesters of 3HB and other hydroxyalkanoic acids (HAs) are the candidates for more practical biodegradable plastics than P(3HB). Particularly, a copolyester of 3HB and (R)-3-hydroxyhexanoic acid (3HHx), P(3HB-co-3HHx) [PHBHHx] is very attractive owing to its superior thermal, mechanical, and physical properties [3]. In addition, the bacterial PHAs are biodegradable not only in soil but also marine environments [4] in contrast to many of other biodegradable plastics which shows almost no biodegradability in marine environments [5].
Figure 1.
Chemical structure of P(3HB) and P(3HB-co-3HHx), PHBHHx.
The increase in 3HHx unit composition in PHBHHx decreases the crystallinity and the melting temperature, leading to wider processing window with better thermal stability than P(3HB) and PHBV (the copolyester of 3HB and 3-hydroxyvarelic acid). P(3HB-co-10mol% 3HHx) shows one of the best property for practical application [6]. PHBHHx was originally found in Aeromonas caviae FA440 in 1993, although the cell growth and polymer accumulation of A. caviae was not so high [7]. Much research has been made to improve biosynthesis of PHBHHx, for instance 3HHx composition, molecular weight, and production efficiency by adopting several strategies and other bacteria, particularly Cupriavidus necator (the former name of this bacterium was Alcaligenes eutropha, Hydrogenomonas eutrophus, Ralstonia eutropha and Wautersia eutropha). The technology for PHBHHx production has progressed to commercial scale. Kaneka Corporation (Tokyo, Japan) constructed a plant for PHBHHx production by recombinant C. necator from vegetable oil with the capacity around 5000 ton/year in 2019 [8]. Kaneka announced to expand PHBH production to 20,000 ton/year [9]. Most of PHA copolymers including PHBHHx are produced from vegetable oils or fatty acids, otherwise by adding precursor compounds structurally related to the second unit other than 3HB during the cultivation. Those organic compounds are usually expensive as raw material in fermentation industry, furthermore such precursor compounds are often toxic to bacterial cells. The efficient production of useful PHAs from inexpensive feedstocks is an urgent task needed for wider use in society [10]. Genetic modification is available to construct the pathways for biosynthesis of the PHA copolymers from structurally unrelated carbon sources. There have been many reports for the biosynthesis of 3HB-based copolymers from plant oils or fatty acids. In those cases, the (R)-3HA-CoAs are provided via β-oxidation of the acyl-moieties [1]. Fukui and co-workers designed an artificial pathway for the generation of (R)-3HHx-CoA from sugar-derived acetyl-CoA molecules and installed into C. necator. Their engineered strain, MF01ΔB1/pBPP-ccrMeJ4a-emd accumulated P(3HB-co-22mol% 3HHx) in the cells at a high cellular content on fructose [11]. They also created the engineered strain to accumulate P(3HB-co-11.7mol% 3HHx) at the cellar content of 72 w/w% on glucose [12].
C. necator is a hydrogen-oxidizing bacterium that can grow and accumulate P(3HB) in the chemoautotrophic culture condition provided with CO2 as the sole carbon source, and H2 and O2 as the energy source and terminal electron acceptor, respectively. Among known autotrophic organisms, C. necator shows high levels of growth rate, cell yield, and P(3HB) accumulation on CO2 [13]. Furthermore, C. necator has been the best studied bacterium for the biosynthesis of PHAs from organic compounds because the bacterium has a high ability to produce PHAs in heterotrophic conditions. In the previous study, we investigated the biosynthesis of PHBHHx by flask culture of the four engineered strains of C. necator in the autotrophic condition provided with a substrate gas mixture (H2/O2/CO2 = 8:1:1) [14]. These recombinant strains accumulated PHBHHx with high 3HHx composition and high cellular content from CO2 as the sole carbon source. Especially, the strain MF01∆B1/pBBP-ccrMeJ4a-emd synthesized the copolymer with remarkably high 3HHx ratio of 47.7 ± 6.2 mol%. The strain MF01/pBPP-ccrMeJAc-emd harboring short-chain-length-specific (R)-enoyl-CoA hydratase (PhaJ) synthesized PHBHHx with high cellular content, where the 3HHx composition was about 11 mol% suitable for practical applications. The highest cell concentration (12.2 g・L-1) in the flask culture was obtained using MF01/pBPP-ccrMeJ4a. It was suspected that the cell growth and PHBHHx accumulation in the flask culture was stopped due to the decrease in pH.
In this study, we investigated the production of PHBHHx from CO2 in pH-stat batch culture of the four recombinant strains of C. necator using a jar fermenter and a recycled-gas closed-circuit culture (RGCC) system in a chemoautotrophic condition. The cell growth, PHBHHx accumulation, and 3HHx composition in the resulting copolyester were compared among these strains. As a result, the concentration of cells and PHBHHx in pH-stat batch culture increased up to about 5 times as that in the flask culture. Production of PHBHHx from CO2 by autotrophic culture of the recombinant C. necator can be comparable to heterotrophic production from organic carbon source such as plant oil when the control of gas composition in RGCC system and the addition of inorganic nutrients during cultivation is further improved.
2. Materials and Methods
2.1. Bacterial Strains
The recombinant strains of C. necator used in this study were MF01/pBPP-ccrMeJ4a-emd, MF01ΔB1/pBPP-ccrMeJ4a-emd, MF01/pBPP-ccrMeJAc-emd, MF01ΔB1/pBPP-ccrMeJAc-emd. The bacterial strains and plasmids used to create those recombinants are listed in Table 1. The details for the construction of recombinant strains were described in the previous report [11,12].
PhaA, β-ketothiolase gene; bktB, broad-substrate range β-ketothiolase gene; phaB1, NADPH-dependent acetoacetyl-CoA reductase gene; phaC, PHA synthase gene in pha operon on chromosome 1; phaCNSDG, a gene of N149S/D171G double mutant of broad-substrate range PHA synthase from A. cavaie; PphaP1, promoter region of phaP1 from C. necator; TrrnB, transcription terminator region from E. coli; ccrMe, crotonyl-CoA carboxylase/reductase gene from Methylorubrum extorquens; emd, a codon-optimized gene encoding ethylmalonyl-CoA decarboxylase from Mus musculu; phaJ4a, medium chain-specific (R)-2-enoyl-CoA hydratase gene; phaJAc, short-chain-specific (R)-2-enoyl-CoA hydratase gene from A. caviae.
2.2. Culture Medium
Mineral salts medium (MS medium) was used for autotrophic culture of the recombinant strains of C. necator. Basic composition of the MS medium was NH4Cl 2.42g, KH2PO4 4.0g, NaHPO4 0.8g, NaHCO3 1.0g, MgSO4・7H2O 0.2g and 1 L of distilled water. Magnesium and phosphate were individually dissolved in water and autoclaved at 121°C for 20 min after the pH was adjusted to 7.0. After cooling, they were aseptically mixed with the salt solution. Zero-point one mL of trace elements solution and 10 mL of kanamycin solution (20 mg/mL), which were filter-sterilized, were added to 1 L of the medium. The composition of the trace elements solution was FeCl3 9.7g, CaCl2 7.8g, CoCl2・6H2O 0.218g, NiCl3・6H2O 0.118g, CrCl3・6H2O 0.105g, and CuSO4・6H2O 0.156g per 1 L of 0.1 M HCl.
2.3. Subculture and Preculture
All the strains were subcultured every 3 weeks on the MS plate medium containing 1.5 w/w% agar. The stock strain was inoculated onto the MS agar plates and placed in a desiccator equipped with a vacuum gauge. The air was evacuated by a vacuum pump and the inner space was filled with H2, O2 and CO2 with a ratio of 8:1:1 from the high-pressure gas cylinders, respectively. The gas composition was adjusted by reading the vacuum gauge. The desiccator was placed at 30°C for 72 h in an incubator for cell growth. These plate cultures were stored at 5°C until the next subculture.
Preculture was autotrophically performed using 15 mL of MS medium in a 300-mL Erlenmeyer flask for 72 h at a reciprocal shaking speed of 170 rpm to prepare the seed culture for jar cultivation. One loop of the cells was inoculated from the stock culture described above to the MS medium in the flask. The detail of flask culture in the autotrophic condition was described previously [14,15].
2.4. Recycled-Gas Closed-Circuit Culture System (RGCC Culture System)
To investigate the polymer production from CO2 employing the wild strain and recombinant strains of C. necator, pH-stat batch culture was carried using a recycled-gas closed-circuit (RGCC) culture system. The RGCC system was used in our previous studies [13,15,16,17,18,19,20,21] for chemoautotrophic cultivation of hydrogen-oxidizing bacteria and those reported by a few studies [22,23,24]. In the culture system, each substrate gas was introduced to a reservoir from the high-pressure gas cylinder, respectively. The schematic diagram of the culture system is shown in Figure 2. The details were explained in our previous reports [15,21].
A glass jar fermenter with a total volume of 1 L was used in this study instead of a 200 mL-scale micro jar fermenter used in our previous studies. The agitator unit was a two-blade type magnetic stirrer bar with a rotating shaft. The volume coefficient of mass transfer for oxygen (KLa) measured by sulfite oxidation method was 250 h-1 at 900 rpm and 310 h-1 at 1,300 rpm. A handmade water-sealed gas holder with the head space capacity of 30 L were used as a new gas reservoir. The water-sealed gas holder was composed of two plastic buckets with different sizes. Four holes were provided at the bottom of the smaller bucket (total volume 45 L) and plastic pipes were tightly inserted into all holes. The pipes were fixed with adhesive material to prevent the gasses from leaking out. They were connected to a vacuum pump, gas cylinders, gas sampling apparatus and the culture system with silicone rubber tubes. This smaller size bucket was floated upside down in saturated saline water in the larger size bucket (total volume 60 L). The air within the smaller bucket was evacuated then substrate gasses were introduced from the cylinders to store them on the saline. The volume and composition of the gas mixture within the smaller basket was adjusted by reading the depth of the smaller bucket sinking in the saline with a ruler written on the surface. The gas mixture in the gas reservoir was fed to the MS medium in the fermenter using a diaphragm pump. The exhaust gas from the fermenter, which was not absorbed in the medium was returned to the gas reservoir and it was repeatedly fed to the fermenter to reduce the risk of detonation and the waste of substrate gasses.
2.5. Condition for Main Culture
In pH-stat batch culture, the working volume of the fermenter was 600 mL and the feeding rate of the gas mixture to the fermenter was 600 mL・min-1 (equivalent to 1.0 vvm). The agitation speed was set to 900 rpm at the start of cultivation then it was increased to 1,300 rpm after 32 h. The pH was maintained at 7.0 by automatically adding 7 w/w% ammonia solution with a pH controller (PHC-2201. Biott Co., Ltd., Japan). The gas composition within the reservoir was set to H2/O2/CO2=8:1:1 and it was reset to the initial ratio every 12 h by refilling new gasses from the high-pressure cylinders. To prevent the culture broth flowing out from the fermenter due to the foaming, a diluted antifoaming reagent, A-nol (Biott) was added during cultivation.
2.6. Analyses
Cell concentration was monitored every 12 h by measuring the optical density at a wavelength of 600 nm (OD600) of the culture broth sampled from the fermenter. For determination of dry cell mass (DCM) per 1 L, 10 mL of the culture broth was heated in a boiling water bath for 1 min then it was centrifuged and washed with distilled water. The weight of the harvested cells was measured after drying for 24 h 105°C. Dissolved oxygen concentration (DO) was measured with a membrane type DO sensor and DO meter (DJ-1033, Biott). The gas mixture was periodically extracted from the gas reservoir (the sample volume; 5 mL) and the composition was analyzed by a gas chromatograph (Shimadzu GC-8A) with a column 4 mm × 6 m into which a molecular sieve 5A and a Porapack Q were packed. The content and monomer composition of PHBHHx accumulated within the cells were determined by gas chromatography. The cells were harvested by centrifugation from the heated culture broth then lyophilized. The dried cells were subjected to direct methanolysis in methanol containing 15% sulfuric acid at 100 °C for 140 min. The resulting methyl esters of 3HB and 3HHx were quantified by gas chromatography [25].
3. Results
3.1. pH-Stat Batch Culture of Recombinant Strains
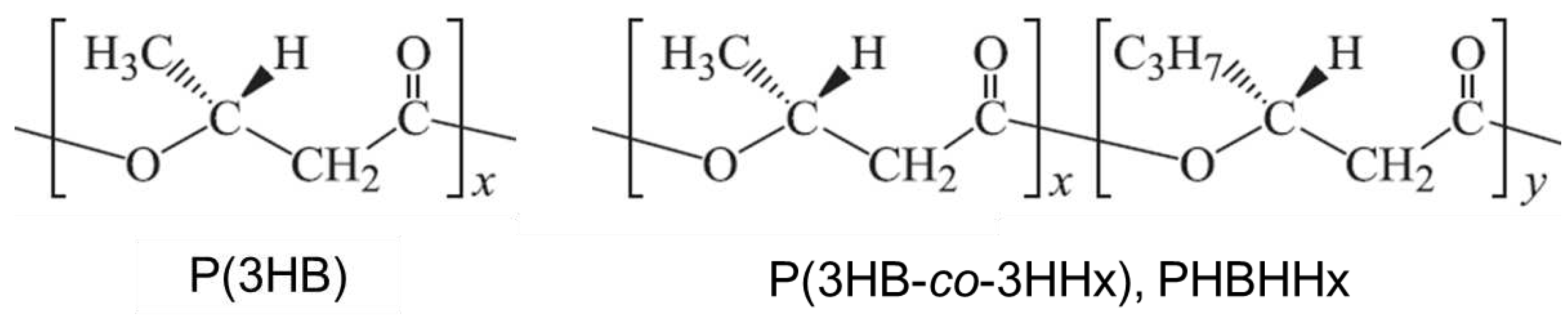
The wild strain (C. necator H16) and the four recombinant strains were cultivated in the MS medium containing 2.42 g・L-1 NH4Cl as a nitrogen source using the RGCC culture system by maintaining pH at 7.0 with 7 w/w% ammonia solution. Time courses of the cell concentration (OD600) and the DO in the culture medium from the start until 156 h of cultivation are shown in Figure 3 and Figure 4, respectively.
The growth of the recombinant strains was slower than that of the wild strain H16. The specific growth rates of each strain from 12 h to 24 h were 0.159 h-1 for H16, 0.137 h-1 for MF01/pBPP-ccrMeJ4a-emd, 0.094 h-1 for MF01ΔB1/pBPP-ccrMeJ4a-emd, 0.107 h-1 for MF01/pBPP-ccrMeJAc-emd, and 0.116 h-1 for MF01ΔB1/pBPP-ccrMeJAc-emd. The exponential cell growth of the strain H16 ceased at around 36 h. After that, the increase in cell concentration followed the linear kinetics. On the hand, the exponential cell growth of the recombinant strains ceased between 36 h and 48 h then the increase in the cell concentration changed to linear kinetics.
The DO of the culture medium decreased to almost 0 ppm after 24 h then it was increased when the gas composition within the reservoir was reset to the initial ratio of H2/O2/CO2=8:1:1 (Figure 4). This up and down in the DO was repeated every 12 h. The DO for all strains was always lower than 0.5 ppm after 72 h, which would promote the polymer accumulation. The decrease in the DO was caused by increase in the DO consumption by the cells and the decline in oxygen concentration of the gas mixture.
The volume of gas mixture within the reservoir (the water-sealed gas holder) decreased as the fermentation proceeded. The concentration of oxygen and CO2 of the gas mixture within the reservoir seriously decreased while that of H2 increased a little (Figure 5). The decrease in oxygen concentration in the gas mixture accelerates the decrease in the DO, which reduces the production rate of the cells and polyester. Therefore, in this study, the substrate gasses were supplied to the reservoir from the high-pressure gas cylinders every 12 h, and the gas composition was reset to the initial ratio. In the case of MF01/pBPP-ccrMeJAc-emd, the amount of substrate gasses consumed per 1 g of produced cells were estimated to be about H2, 9.1 L; O2, 2.4 L; CO2, 0.8 L from the change in the volume and composition of gas mixture in the reservoir between 48 h and 72 h of the cultivation. However, the amount of gas leak from the culture system occurred with unignorable level in this study then these estimations are not reliable. We are going to measure the consumption of substrate gasses by the cells precisely in future study.
Among the recombinant strains, the highest cell concentration at the end of cultivation was obtained for MF01/pBPP-ccrMeJ4a-emd (OD600, 160.0), which was a little higher than that in the wild strain H16. MF01/pBPP-ccrMeJAc-emd also reached high cell concentration. After the cultivation, the cells were harvested from the culture broth and freeze-dried. The lyophilized cells were applied for methanolysis then the samples were analyzed by gas chromatography to determine the polymer content in the cells and the monomer composition (Table 2). The highest content of PHBHHx in the cells (about 83.9 w/w%) was achieved by MF01/pBPP-ccrMeJAc-emd. The highest composition of 3HHx (21.2 mol%) was obtained in MF01ΔB1/pBPP-ccrMeJ4a-emd while the 3HHx composition in MF01/pBPP-ccrMeJAc-emd was 10.9 mol% that was very close to the best value for practical use of PHBHHx. In the case of MF01/pBPP-ccrMeJ4a-emd, the 3HHx composition obtained in the pH-stat batch culture was much lower than that of the flask culture [14]. In the flask culture using 2.42 g・L-1 NH4Cl, the pH decreased to around 4.0 at the end of cultivation (the data are not shown). The decrease in pH might affect the biosynthesis of 3HB-CoA unit and 3HHx-CoA unit, and/or incorporation of the monomer units into the copolymer. The pathway for the biosynthesis of PHBHHx from CO2 in the recombinant C. necator was deduced in our previous study [14]. Unfortunately, it is unknown which step on the pathway was affected by the decrease in the pH of culture medium.
3.1. pH-Stat Batch Culture of MF01/pBPP-ccrMeJAc-emd with Supplementing Phosphate
MF01/pBPP-ccrMeJAc-emd showed good results in the cell concentration, PHBH accumulation and 3HHx composition. Hence, to further increase the production of cells and PHBHHx, the pH-stat batch culture was carried out by adding 0.2 g・L-1 of KH2PO4 to the culture medium at 96 h and 168 h. The culture experiment was carried out three times at the same conditions. Figure 6 shows the increase in dry cell mass (DCM) and PHBHHx concentration at every 24 h during the cultivation. After 216 h, the average values of DCM and PHBHHx concentration increased to 59.62 ± 3.18 g・L-1 and 49.31 ± 3.14 g・L-1, respectively. The cellular content of PHBHHx and the 3HHx composition at the end were 82.7 ± 1.37 w/w% and 9.3 ± 3.7 mol%, respectively.
4. Discussion
There has been growing interest in PHA production from CO2 by autotrophic microorganisms and the number of the research reports for hydrogen-oxidizing bacteria is also increasing. For instance, those research focus on the fermentation process to keep O2 concentration in the substrate gas below the lower limit for detonation [26,27,28,29], the precise measurement of the gas consumption and determination of the stoichiometry for PHA production [26,28,30,31], the improvement of productivity [32], and the production of copolymer PHAs. However, most of the studies for production of copolymer PHAs from CO2 were studied by mixotrophic culture where organic compounds structurally related to the secondary hydroxyacyl monomer units were added to culture medium during PHA accumulation period [24,33,34]. Although some researchers reported biosynthesis of copolymer PHAs from CO2 as a sole carbon source [14,27,35], the concentration of the resulting copolyesters was still very low. The PHBHHx concentration that we obtained by obligate autotrophic culture of MF01/pBPP-ccrMeJAc-emd in this study was much higher than those observed in previous related reports.
Inorganic nutrients contained in the MS medium are utilized for biosynthesis of cell components such as proteins and nucleic acids, and for activation of enzymes and other physiological functions in cells. While it is known that PHA accumulation is promoted by limitation of inorganic nutrients especially nitrogen (N) compounds and phosphorus (P) compounds, that forms an imbalanced composition to carbon source in the culture medium. Limitation of dissolved oxygen also promotes PHA accumulation. In heterotrophic production of PHAs with feeding organic carbon sources, very high PHA concentration was often obtained by limiting the feed amount of nitrogen [36] or phosphorus compound [37] during PHA accumulation period. The PHA accumulation in the autotrophic cultivation of hydrogen-oxidizing bacteria is also promoted by limitation of N, P and/or DO. However, it is difficult to feed CO2 and other nutrients with a constant C/N or C/P ratio. Hence, in this study, ammonia solution (N source) was gradually added as the neutralizer to maintain the pH of culture medium, and small amount of KH2PO4 solution (P source) was added twice during the PHA accumulation. This N- and P-feeding strategy appeared to be successful to increase PHBHHx concentration to a certain extent. However, the concentrations of NH4-N and PO43- were changeable during the cultivation and they often significantly increased when the gas reservoir was refilled with new gasses at every 12 h. The concentration of oxygen and CO2 in the gas reservoir was quickly increased to the initial level after the refill of the reservoir, as shown in Figure 5. The increase in CO2 concentration accelerated the addition of ammonia solution to maintain the pH of culture medium. It will be possible to produce the cells and PHBHHx beyond their concentrations obtained in this study by improving the method for the feed of ammonia solution and K2HPO4 to maintain constantly low concentrations of NH4-N and PO43-.
The productivity of cells and PHA by autotrophic culture depends on the mass transfer of the substrate gasses in the culture medium. The oxygen transfer rate is particularly important because DO limitation is inevitable at high cell concentration. The oxygen concentration of gas phase within the culture system of hydrogen-oxidizing bacteria is usually kept lower than that of the air for some reasons. Most of the bacteria is sensitive to oxygen and the optimal oxygen concentration is much lower than that of the air. Furthermore, it is effective to keep oxygen concentration below the lower limit for explosion (around 4.0 v/v%) to prevent detonation. Oxygen transfer rate in culture medium decreases according to the decline in oxygen concentration of the gas mixture, which diminishes the productivity of cells and PHA [18]. Therefore, the productivity of PHBHHx in the autotrophic culture of the recombinant strains of C. necator can be increased by improving the supply of the substrate gasses into the reservoir to maintain the gas composition at a constant ratio and by enhancing the agitation effect on the dissolution of the gasses in the culture medium. We already succeeded to increase the cell growth and homopolyester P(3HB) production to 91.3 g・L-1 and 62 g・L-1, respectively after 40 h of autotrophic cultivation of the wild strain H16 using specially designed basket type agitation unit [18].
Now we are investigating further high cell density cultivation of the engineered C. necator with the shorter fermentation time by improving the method to control the concentration of NH4-N and PO43- in the medium and the gas composition within the reservoir, and by using a high-performance agitation unit.
Author Contributions
K.T.: conceptualization; investigations on autotrophic culture experiment and analyze; methodology; writing—original draft preparation. I.O.: methodology and investigations for recombination of C. necator H16; T.F.: resources of C. necator recombinant; funding acquisition; methodology and investigations for recombination of C. necator H16; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This paper is based on results obtained from a project, JPNPX18016, commissioned by the New Energy and Industrial Technology Development Organization (NEDO).
References
- Lu, J.; Tappel, R.C.; Nomura, C.T. Mini-review: Biosynthesis of poly(hydroxyalkanoates). Polym. Rev. 2009, 49, 226–248. [Google Scholar] [CrossRef]
- Turco, R.; Santagata, G.; Corrado, I.; Pezzella, C.; Serio, M.D. In vivo and post-synthesis strategies to enhance the properties of PHB-based materials: A Review. Front. Bioeng. Biotechnol. 2021, 8, 619266. [Google Scholar] [CrossRef] [PubMed]
- Eraslan, k.; Aversa, C.; Nofar, M.; Barletta, M.; Gisario, A.; Salehiyan, R.; Goksu, Y.A. Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHBH): Synthesis, properties, and applications – review. Eur. Polym. J. 2022, 167, 15. [Google Scholar] [CrossRef]
- Sashiwa, H.; Fukuda, R.; Okura, T.; Sato, S.; Nakayama, A. Microbial degradation behavior in seawater of polyester blends containing poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) (PHBHHx). Mar. Drugs. 2018, 16, 34. [Google Scholar] [CrossRef] [PubMed]
- Barron, A.; Taylor, D.S. Commercial marine-degradable polymers for flexible packaging. iScience. 2020, 23, 8–101353. [Google Scholar] [CrossRef]
- Shimamura, E.; Kasuya, K.; Kobayashi, G.; Shiotani, T.; Shima, Y.; Doi, Y. Physical properties and biodegradability of microbial poly(3-hydroxybutyrate-co-3-hydroxyhexanoate). Macromolecules. 1994, 27, 878–880. [Google Scholar] [CrossRef]
- Doi, Y.; Kitamura, S.; Abe, H. Microbial synthesis and characterization of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate). Macromolecules. 1995, 28, 4822–4828. [Google Scholar] [CrossRef]
- Kaneka Corporation News Release. Completion of KANEKA Biodegradable Polymer PHBH™ plant with annual production of 5,000 tons. 19 December 2019. https://www.kaneka.co.jp/en/topics/news/nr20191219/ (accessed on 20 August 2021).
- Kaneka Corporation News Release. Kaneka to significantly increase its production capacity for KANEKA Biodegradable Polymer Green Planet™ in Japan. February 7, 2022. https://www.kaneka.co.jp/en/topics/news/2022/ennr2202071.html (accessed 1 October 2023).
- Boey, J.Y.; Mohamad, L.; Khok, Y.S.; Tay, G.S.; Baidurah, S. A Review of the applications and biodegradation of polyhydroxyalkanoates and poly(lactic acid) and its composites. Polymers. 2021, 13, 1544. [Google Scholar] [CrossRef]
- Insomphun, C.; Xie, H.; Mifune, J.; Kawashima, Y.; Orita, I.; Nakamura, S.; Fukui, T. Improved artificial pathway for biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) with high C6-monomer composition from fructose in Ralstonia eutropha. Metab. Eng. 2015, 27, 38–45. [Google Scholar] [CrossRef]
- Zhang, M.; Kurita, S.; Orita, I.; Nakamura, S.; Fukui, T. Modification of acetoacetyl-CoA reduction step in Ralstonia eutropha for biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from structurally unrelated compounds. Microb. Cell Fact. 2019, 18, 147. [Google Scholar] [CrossRef]
- Ishizaki, A.; Tanaka, K.; Taga, N. Microbial production of poly-D-3-hydroxybutyrate from CO2. Appl. Microbiol. Biotechnol. 2001, 57, 6–12. [Google Scholar] [PubMed]
- Tanaka, K.; Kazumasa Yoshida, K.; Orita, I.; Fukui, T. Biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a recombinant Cupriavidus necator. Bioengineering, 2021, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, A.; Tanaka, K. Batch culture of Alcaligenes eutrophus ATCC 17697T using recycled gas closed circuit culture system. J. Ferment. Bioeng 1990, 69, 170–174. [Google Scholar] [CrossRef]
- Ishizaki, A.; Tanaka, K. Production of poly-β-hydroxybutyric acid from carbon dioxide by Alcaligenes eutrophus ATCC 17697T. J. Ferment. Bioeng. 1991, 71, 254–257. [Google Scholar] [CrossRef]
- Takeshita, T.; Tanaka, K.; Ishizaki, A.; Stanbury, P.F. Development of a dissolved hydrogen sensor and its application to evaluation of hydrogen mass transfer. J. Ferment. Bioeng. 1993, 76, 148–150. [Google Scholar] [CrossRef]
- Tanaka, K.; Ishizaki, A.; Kanamaru, T.; Kawano, T. Production of poly(D-3-hydroxybutyrate) from CO2, H2, and CO2 by high cell density autotrophic cultivation of Alcaligenes eutrophus. Biotechnol. Bioeng. 1995, 45, 268–275. [Google Scholar] [CrossRef]
- Taga, N.; Tanaka, K.; Ishizaki, A. Effects of rheological change by addition of carboxymethylcellulose in culture media of an air-lift fermentor on poly-D-3-hydroxybutyric acid productivity in autotrophic culture of hydrogen-oxidizing bacterium, Alcaligenes eutrophus. Biotechnol. Bioeng. 1997, 53, 529–533. [Google Scholar] [CrossRef]
- Sugimoto, T.; Tsuge, T.; Tanaka, K.; Ishizaki, A. Control of acetic acid concentration by pH-stat continuous substrate feeding in heterotrophic culture phase of two-stage cultivation of Alcaligenes eutrophus for production of P(3HB) from CO2, H2 and O2 under non-explosive condition. Biotechnol. Bioeng. 1999, 62, 625–631. [Google Scholar] [CrossRef]
- Tanaka, K.; Miyawaki, K.; Yamaguchi, A.; Khosravi-Darani, K.; Matsusaki, H. Cell growth and P(3HB) accumulation from CO2 of a carbon monoxide-tolerant hydrogen-oxidizing bacterium, Ideonella sp. O-1. Appl Microbiol Biotechnol. 2011, 92, 1161–1169. [Google Scholar] [CrossRef]
- Schlegel, H.G.; Lafferty, R.M. Novel energy and carbon sources. The production of biomass from hydrogen and carbon dioxide. Adv. Biochem. Eng. 1971, 143, 143–168. [Google Scholar] [CrossRef]
- Kodama, T.; Igarashi, Y.; Minoda, Y. Isolation and culture conditions of a bacterium grown on hydrogen and carbon dioxide. Agr. Biol. Chem. 1975, 39, 77–82. [Google Scholar] [CrossRef]
- Volova, T.G.; Kiselev, E.G.; Shishatskaya, E.I.; Zhila, E.I.; Boyandin, A.N.; Syrvacheva, D.A.; Vinogradova, O.N.; Kalacheva, G.S.; Vasiliev, A.D.; Peterson, I.V. Cell growth and accumulation of polyhydroxyalkanoates from CO2 and H2 of a hydrogen-oxidizing bacterium, Cupriavidus eutrophus B-10646. Bioresour. Technol. 2013, 146, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Bao, H.J.; Kang, C.-K.; Fukui, T.; Doi, Y. Production of a novel copolyester of 3-hydroxybutyric acid and medium-chain-length 3-hydroxyalkanoic acids by Pseudomonas sp. 61-3 from sugars. Appl. Microbiol. Biotechnol. 1996, 45, 363–370. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, L.; Mozumder, M.S.; Dubreuil, M.; Volcke, E.; Wever, H. Sustainable autotrophic production of polyhydroxybutyrate (PHB) from CO2 using a two-stage cultivation system. Catal.Today. 2015, 257 Pt 2, 237–245. [Google Scholar] [CrossRef]
- Miyahara, Y.; Yamamoto, M.; Thorbecke, R.; Mizuno, S.; Tsuge, T. Autotrophic biosynthesis of polyhydroxyalkanoate by Ralstonia eutropha from non-combustible gas mixture with low hydrogen content. Biotechnol. Lett. 2020, 42, 1655–1662. [Google Scholar] [CrossRef]
- Lambauer, V.; Kratzer, R. Lab-scale cultivation of Cupriavidus necator on explosive gas mixtures: Carbon dioxide fixation into polyhydroxybutyrate. Bioengineering. 2022, 9, 204. [Google Scholar] [CrossRef] [PubMed]
- Lambauer, V.; Permann, A.; Petrášek, Z.; Subotić, V.; Hochenauer, C.; Kratzer, R.; Reichhartinger, M. Automatic control of chemolithotrophic cultivation of Cupriavidus necator: Optimization of oxygen supply for enhanced bioplastic production. Fermentation. 2023, 9, 619. [Google Scholar] [CrossRef]
- Mozumder, M.S.; Garcia-Gonzalez, L.; Wever, H.; Volcke, E. Poly(3-hydroxybutyrate) (PHB) production from CO2: Model development and process optimization. Biochem. Eng. J. 2015, 98, 107–116. [Google Scholar] [CrossRef]
- Yu, J.; Lu, Y. Carbon dioxide fixation by a hydrogen-oxidizing bacterium: Biomass yield, reversal respiratory quotient, stoichiometric equations and bioenergetics. Biochem. Eng. J. 2019, 152, 15–107369. [Google Scholar] [CrossRef]
- Yu, J.; Munasinghe, P. Gas fermentation enhancement for chemolithotrophic growth of Cupriavidus necator on carbon dioxide. Fermentation. 2018, 4, 63. [Google Scholar] [CrossRef]
- Inseon Park, I.; Jho, E.h.; Nam, K. Optimization of carbon dioxide and valeric acid utilization for polyhydroxyalkanoates synthesis by Cupriavidus necator. J. Polym. Environ. 2014, 22, 244–251. [Google Scholar] [CrossRef]
- Ghysels, S.; Mozumder, M.S.; Wever, H.; Volcke, E. .; Garcia-Gonzalez, L. Targeted poly(3-hydroxybutyrate-co-3-hydroxyvalerate) bioplastic production from carbon dioxide. Bioresour. Technol. 2018, 249, 858–868. [Google Scholar] [CrossRef]
- Nangle, S.N.; Ziesack, M. : Buckley, S.; Trivedi, D.; Loh, D.M.; Nocera, D.G.; Silvera, P.A. Valorization of CO2 through lithoautotrophic production of sustainable chemicals in Cupriavidus necator. Metab. Eng. 2020, 62, 207–220. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamane, T.; Shimizu, S. ; Mass production of poly-β-hydroxybutyric acid by fed-batch culture with controlled carbon/nitrogen feeding. Appl. Microbiolo. Biotechnol. 1986, 24, 370–374. [Google Scholar] [CrossRef]
- Ryu, H.W.; Hahn, S.K.; Chang, Y.K.; Chang, H.N. Production of poly (3-hydroxybutyrate) by high cell density fed-batch culture of Alcaligenes eutrophus with phosphate limitation, Biotechnol. Bioeng. 1997, 55, 28–32. [Google Scholar] [CrossRef]
Figure 2.
Schematic diagram of RGCC culture system for pH-stat batch culture of hydrogen-oxidizing bacterium, C. necator in chemoautotrophic conditions.
Figure 2.
Schematic diagram of RGCC culture system for pH-stat batch culture of hydrogen-oxidizing bacterium, C. necator in chemoautotrophic conditions.

Figure 3.
Time course for increase in cell concentration (OD600) in pH-stat batch culture of C. necator H16 and the recombinant strains in autotrophic condition.
Figure 3.
Time course for increase in cell concentration (OD600) in pH-stat batch culture of C. necator H16 and the recombinant strains in autotrophic condition.
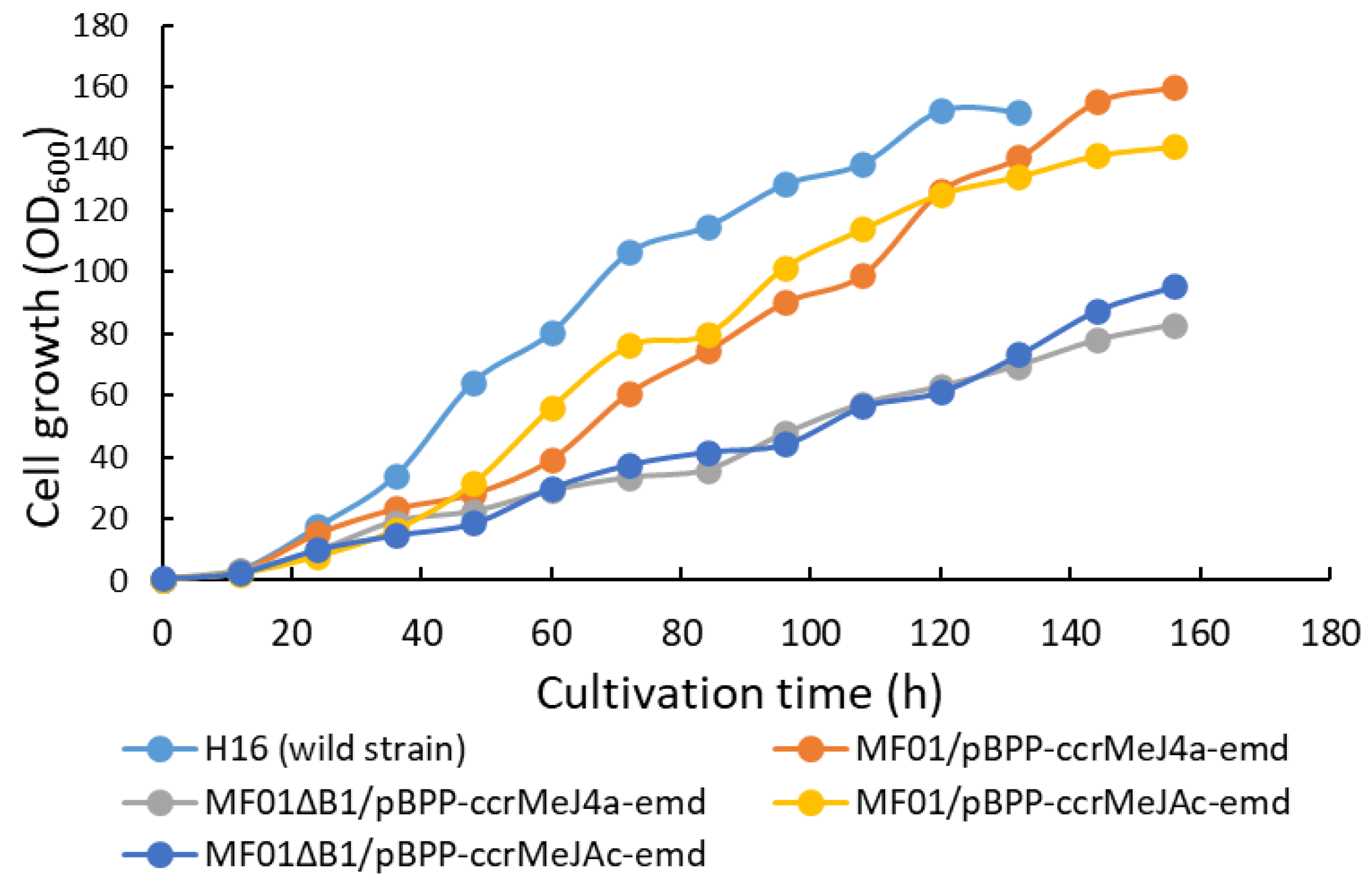
Figure 4.
Time course for change in dissolved oxygen concentration in pH-stat batch culture of C. necator H16 and the recombinant strains in autotrophic condition.
Figure 4.
Time course for change in dissolved oxygen concentration in pH-stat batch culture of C. necator H16 and the recombinant strains in autotrophic condition.
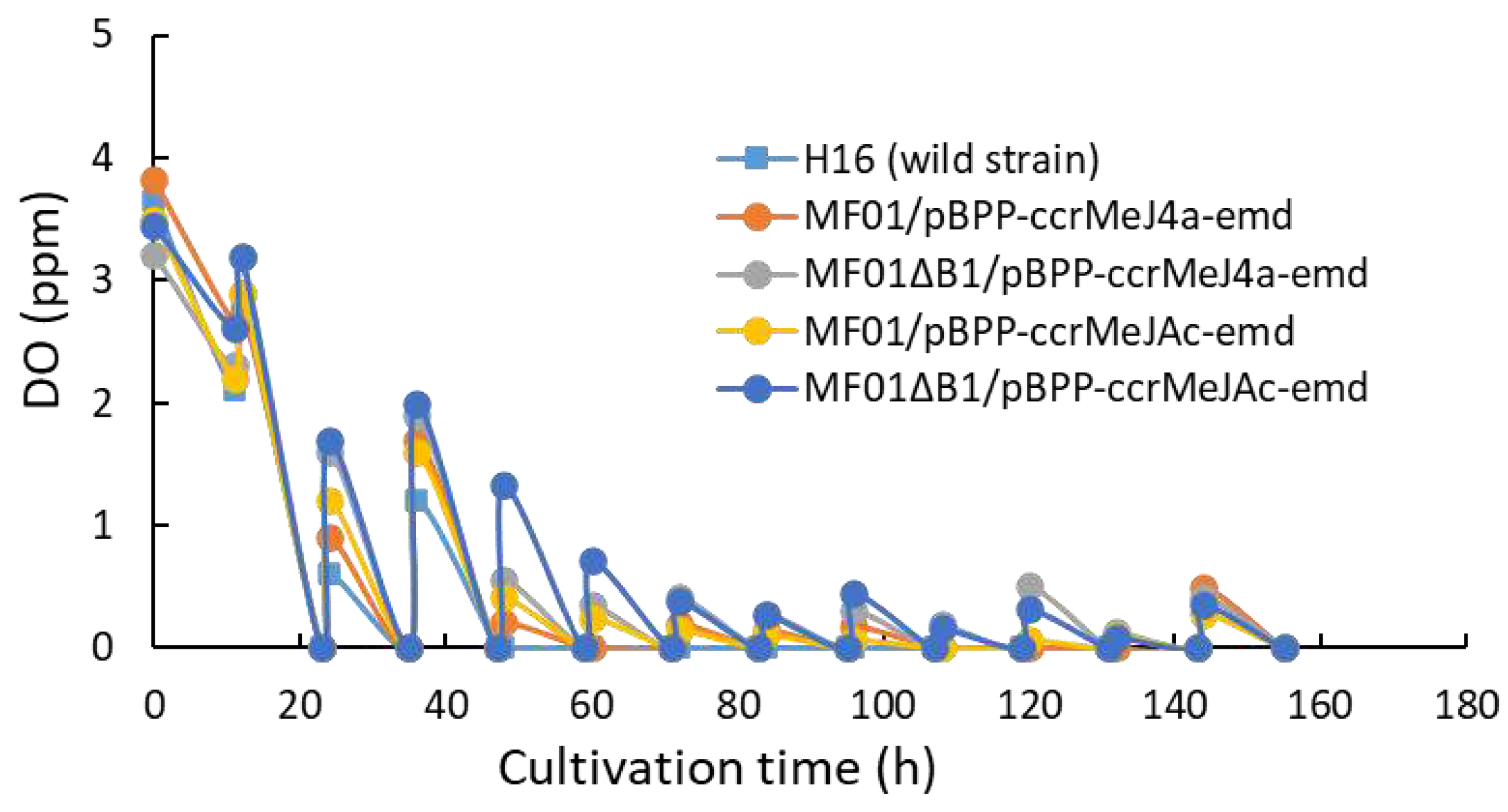
Figure 5.
Change of gas composition within gas reservoir in pH-stat batch culture of C. necator MF01/pBPP-ccrMeJAc-emd in autotrophic condition.
Figure 5.
Change of gas composition within gas reservoir in pH-stat batch culture of C. necator MF01/pBPP-ccrMeJAc-emd in autotrophic condition.
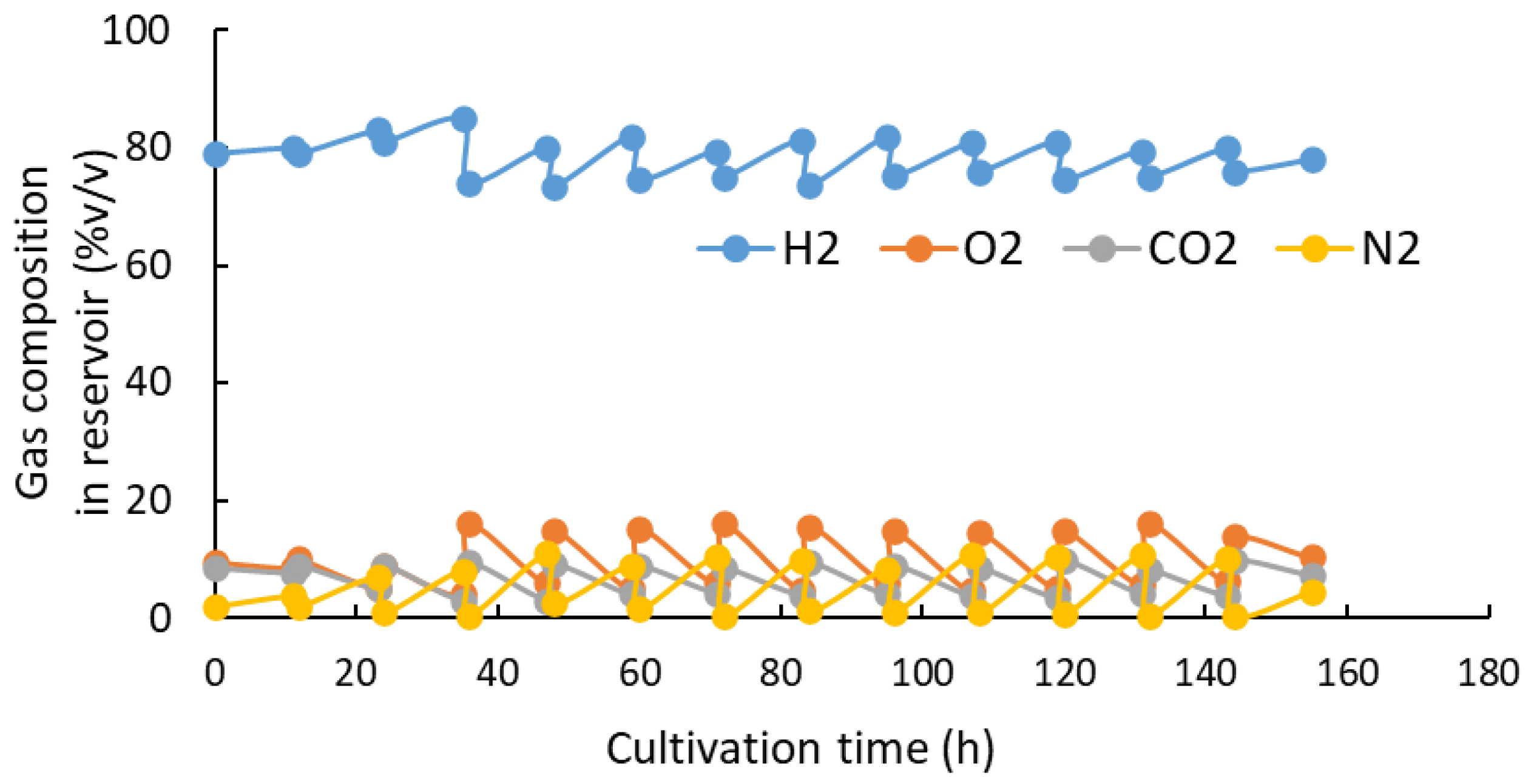
Figure 6.
Time courses for increase in DCM and PHBHHx in pH-stat batch culture of MF01/pBPP-ccrMeJAc-emd under autotrophic condition with addition of 0.2 g・L-1 of KH2PO4 to culture medium at 96 h and 168 h. (n=3).
Figure 6.
Time courses for increase in DCM and PHBHHx in pH-stat batch culture of MF01/pBPP-ccrMeJAc-emd under autotrophic condition with addition of 0.2 g・L-1 of KH2PO4 to culture medium at 96 h and 168 h. (n=3).
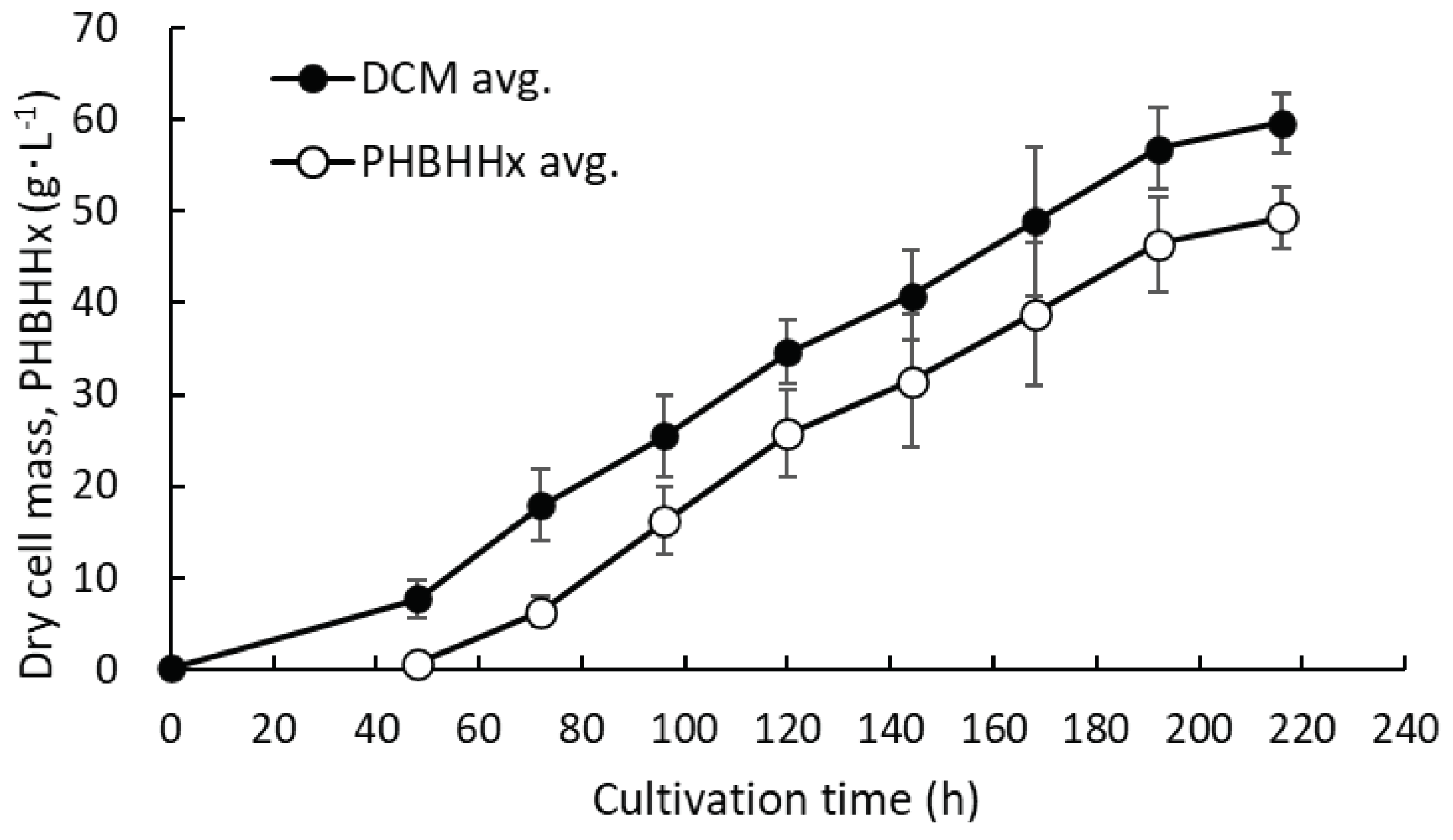

Table 1.
Bacterial strains and plasmids used in this study.
| Used strains | Relevant marker |
|---|---|
| C. necator H16 C. necator MF01 C. necator MF01ΔB1 |
Wild type H16 derivative; ΔphaC::phaCNSDG , ΔphaA::bktB MF01 derivative; ΔphaB1 |
| Used plasmids | |
| pBPP pBPP-ccrMeJ4a-emd pBPP-ccrMeJAc-emd |
pBBR1-MCS2 derivative; PphaP1, TrrnB, pBPP derivative; ccrMe, phaJ4a, emdMm pBPP-ccrMeJ4a-emd derivative; ΔphaJ4a::phaJAc |
Table 2.
Cell growth and PHA accumulation in autotrophic pH-stat batch culture of C. necator H16 and recombinant strains.
Table 2.
Cell growth and PHA accumulation in autotrophic pH-stat batch culture of C. necator H16 and recombinant strains.
| Strains/Plasmid | Dry cell mass (g・L-1) |
PHBHHx content in cells (w/w%) | Monomer composition (mol%) |
|
|---|---|---|---|---|
| 3HB | 3HHx | |||
| H16 (wild strain) | 44.05 | 78.5 | 100.0 | 0 |
| MF01/pBPP-ccrMeJ4a-emd | 45.42 | 57.7 | 92.9 | 7.1 |
| MF01ΔB1/pBPP-ccrMeJ4a-emd | 23.06 | 66.8 | 78.8 | 21.2 |
| MF01/pBPP-ccrMeJAc-emd | 40.12 | 83.9 | 89.1 | 10.9 |
| MF01ΔB1/pBPP-ccrMeJAc-emd | 26.70 | 75.5 | 90.9 | 9.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



