
Submitted:
20 September 2023
Posted:
22 September 2023
You are already at the latest version
Abstract
The shoot apical meristem (SAM) gives rise to the aerial structure of plants by producing lateral organs and secondary meristems. The SAM is responsible for plant developmental patterns, thus determining plant morphology and, consequently, many agronomic traits such as the number and size of fruits and flowers and kernel yield. Our current understanding of SAM morphology and regulation is based on studies conducted mainly on some angiosperms, including economically important crops such as maize (Zea mays) and rice (Oryza sativa), and the model species Arabidopsis (Arabidopsis thaliana). However, studies in other plant species from the gymnosperm class are scant, making difficult comparative analyses that help us understand SAM regulation in diverse plant species. This limitation prevents deciphering the mechanisms by which evolution gave rise to the multiple plant structures within the plant kingdom, and determined the conserved mechanisms involved in SAM maintenance and operation. This review aims to integrate and analyze the current knowledge of SAM evolution by combining the morphological and molecular information recently reported from the plant kingdom.
Keywords:
shoot apical meristem (SAM)
; SAM conservation
; Sc-RNA-seq
; Streptophyta
1. Introduction
Plants can maintain indeterminate postembryonic growth by reserving pools of stem cells. These reservoirs are confined to specialized regions called meristems, which are present in various plant structures. The apical meristems (AMs), located on the tips of the germinating seedling, are responsible for the plant's primary growth or length. There are two types of AMs: the shoot apical meristem (SAM) and the root apical meristem (RAM), which are responsible for the growth of the aerial part and the root system of plants, respectively. Additionally, plants have lateral meristems, which are responsible for secondary growth that determines the thickness of the plant, and intercalary meristems, which are responsible for growth at the base of the nodes and leaf blades in monocots [1].
Beyond growth, meristems also play a crucial role in defining plant anatomical patterns. With noticeable differences in their structure, meristems serve as a valuable model for studying plant evolution and understanding the structural and functional aspects of plant development. In general, primitive plants have a simple meristem consisting of only one apical cell, whereas late divergent plants have multicellular meristems [2]. Despite these important structural variations, meristematic cells share two essential and highly conserved functions: maintaining the meristem integrity, including its stem cell population, and producing cells to support plant growth [3]. This review summarizes current knowledge on meristem structure, function, and evolution. Given the broad scope of this topic and the scarcity of studies dedicated exclusively to studying the SAM from an evolutionary perspective, we integrate and synthesize all relevant literature on meristem reserach. The goal is to provide a comprehensive review that facilitates the understanding of SAM evolution. We divide this review into four main sections:
- In the first section, we present essential concepts to understand plant meristems. We introduce the concept of stem cells and compare the stem cell niche of RAM and SAM. We emphasize the importance of plant life cycles, as this concept gives rise to SAM evolution theories.
- Then, the meristem shape is analyzed throughout phylogeny. This section offers a morphological description of SAM across plant evolution. Given the extensive literature on SAM morphology, it provides perspectives on the differences in SAM between clades.
- We provide a comprehensive review of the regulatory and maintenance mechanisms in the SAM. This section focuses on the regulatory loops described for angiosperms and the conserved elements across clades.
- Taking advantage of single-cell transcriptomics to understand SAM, this section delves into the research of single-cell transcriptomics and single-nucleus transcriptomics on SAM. We describe key studies and discuss their findings.
2. Evolutionary Origin of the Meristem
2.1. The Concept of Stem Cells
Plant stem cells are innately undifferentiated and are contained mainly in the AMs. They are commonly defined by their developmental capabilities, namely, self-renewal and the ability to differentiate into multiple cell types [4]. Therefore, one of the main interests in studying AMs is related to their stem cell pool. Plant cells can be de-differentiated and re-differentiated into various cell types, which is why they are considered totipotent. The term totipotency refers to the capacity of one cell to produce different tissues and eventually regenerate into an entire organism [5]. However, the specific conditions that activate their differentiation into different cell types are still largely unknown [6]. To study this process, the standard models in plants are embryos [7], protoplasts [8], calli [9], and meristems [10]. These models are valuable as they contain dedifferentiated cells, which can develop into differentiated cells, ultimately giving rise to different plant tissues. Hence, an interesting perspective to understand this phenomenon is to focus on the evolution of totipotency by comparing the meristems of early and modern diverged land plants, signaling out the common factors on totipotency through evolution.
2.2. SAM and RAM
Extensive research has been conducted to understand RAM evolution, but little is known about the SAM evolutionary process. Since both AMs are structures with similar characteristics, studying the RAM can be a starting point for understanding the SAM. Both are reservoirs of self-renewing stem cells that can be considered primary meristems because, in most cases, they are determined during embryogenesis [11]. Stem cells in the RAM and SAM are located in regions referred to as the quiescent center (QC) and organizing center (OC), respectively. The QC and the OC are the signaling centers responsible for stem cell maintenance in both meristems [12] and have been proposed to be functionally equivalent [13,14].
Studies conducted on the model plant Arabidopsis thaliana suggest that the organization of the signaling components required for stem cell initiation and maintenance in the RAM and SAM are relatively conserved. Consequently, similar classes of genes have been co-opted as central regulators in both types of meristems [14,15]. However, despite these shared mechanisms, the hormonal function has been shown to be inverse [16,17]. In the SAM, auxins trigger differentiation, whereas in the RAM, they maintain the stem cell niche and support cell proliferation [18]. On the other hand, in the SAM, cytokinin promotes tissue proliferation of undifferentiated cells, whereas it promotes cell differentiation in the RAM [19,20]. The OC of the SAM is the site of maximal cytokinin activity, whereas the auxin maximum is in the QC of the RAM [20] (Figure 1).
Fossil evidence suggests that the RAM evolved at least twice independently rather than having a single origin [21,22]. The first appearance occurred in the lycophytes clade, followed by a second evolutionary event, which likely occurred in the ancestor of euphyllophytes (vascular plants non-lycophytes) [23]. It has even been proposed that RAM independently evolved multiple times in lycophytes [24]. Moreover, the similarities between lycophyte and euphyllophyte roots, including indeterminate growth, apical meristem protected by a root cap, root hairs, and a stele covered by a specialized endodermal cell layer, are clear examples of convergent evolution [25,26]. Genomic analyses revealed limited gene expansion occurred at the divergence between the lycophyte and euphyllophyte clades [27]. Consequently, all the similarities produced by convergent evolution did not require extra gene families, suggesting that the rewiring of existing genetic programs was sufficient to generate multiple independent emergences of the RAM [28].
2.3. Apical Metistem of Gametophyte and Sporophyte
The plant life cycle comprises two alternating phases: the haploid or gametophyte phase and the diploid or sporophyte phase. The diploid phase concludes with meiosis, while the haploid phase ends with the generation and union of gametes [31]. It is remarkable that, as land plants diversified, the sporophyte generation became increasingly specialized, whereas the gametophyte generation became reduced [32,33]; This tendency is reflected in SAMs, where non-seed plants usually have single-celled SAMs to sustain the growth of the sporophytes and gametophytes. In contrast, seed plants' sporophytes develop complex multicellular meristems, while their gametophytes lack meristems [34,35] (Figure 2). However, as both phases have SAMs, it is reasonable to discuss the origin of meristems in terms of the plant life cycle.
Two hypotheses have been proposed that explain the appearance of the alternation cycle [36]. First, the homologous theory describes the ancestor of land plants with an originally haplontic life cycle that gives rise to a sporophyte through delay in meiosis, resulting in the intercalation of a new diploid organism [33,37]. This assumption outlines the gametophytic SAM as the ancestor of the sporophytic SAM. The second hypothesis is the antithetic theory, which proposes that sporophytic SAM is independent of gametophytic SAM. According to this theory, the sporophytic SAM evolved de novo by intercalating a system between embryonic and reproductive growth [33,38]. Current evidence based on embryophytes and fossil records is more compatible with the antithetic theory than the homologous theory [39,40]. Hypotheses derived from the antithetic theory include 1) the embryophyte SAM originated from the transient seta meristem (sporophyte) of a bryophyte, 2) the hornwort sporophyte meristem is the possible ancestor of the SAM, and 3) the SAM in vascular plants arose de novo from a bryophyte ancestor [41].
Overall, evidence suggests that gametophyte and sporophyte SAM have an independent origin. However, as plants share the same startup gene toolkit since their separation from algae, they probably also share homologous regulatory loops and mechanisms on their SAM regulation. In fact, mosses and ferns gametophyte SAMs share key transcription factors (TFs) with angiosperm sporophyte SAMs [42,43], which suggests the existence fundamental mechanisms involved in stem cell regulation across land plants.
3. Meristem Shape Throughout Phylogeny
A traditional approach to studying meristems considers their morphology and focuses on their correlation to plant growth patterns. Cell division patterns and apical cell symmetry can outline plant morphology [44]. For example, in bryophytes, taxa with tetrahedrical apical cells tend to have leafy morphologies, while taxa with hemidiscoid apical cells tend to be thalloid [45]. The study of apical cells applies mainly to SAMs with a single apical cell found in macroalgae, bryophytes, lycophytes, and ferns. The simplest apical cell is the dome-shaped cell present in macroalgae (Charales) and moss gametophytes, and it allows one-dimensional (1D) growth. However, land plant clades such as lycophytes and ferns present a tetrahedral apical cell allowing three-dimensional (3D) growth [46,47] (Figure 3). It can be hypothesized that the emergence of apical cells capable of dividing following multiple planes resulted in drastic morphological innovations, ultimately allowing the colonization of land by plants.
3.1. Algae
The evolution from algae to plants is closely tied to the evolution of the body plan of plants. The Charophycean green algae, the closest relative of the extant land plants [50,51], provides an evolutionary example of the transition from unicellular to multicellularity. Some Chlorophycean algae are still at the midpoint of this transition [52]. However, the presence of a SAM is limited to multicellular organisms with organized tissues.
Among Charophyceans, apical growth has been demonstrated only in the order Charales, which have a single apical cell per shoot [53]. The dome-shaped apical cells of Charales have one cutting face opposite the apex of the surface, and this cutting plane is parallel to the orientation of the last cell plate. As a result, they present 1D growth, forming cell chains resembling filaments or mats [46,54]. Another charophyte, the Coleochaetalean algae, typically exhibits a marginal meristem where cells can divide into two possible planes, leading to the formation of radially symmetrical filamentous or monochromatic thalli that resemble hornworts gametophytes [55].
3.2. Bryophytes
Bryophytes comprise a taxonomic group consisting of three clades of non-vascular land plants: mosses, liverworts, and hornworts [56]. It is traditionally considered that land plant traits primarily evolved in bryophytes. Liverworts were the earliest-divergent clade of land plants, whereas mosses and hornworts are sister groups. However, there is only partial acceptance of the internal topology of this clade, despite evidence of its monophyly [57,58]. It has been proposed that the sporophyte SAM was derived from a sterilized bryophyte sporangium, but this theory remains untestable due to a lack of homologies or fossil records [59,60].
The moss sporophyte has three transient cells recognized as meristems: the apical cell, active in the early embryo; the central seta cell that maintains the seta's growth; and a transient single apical cell of the sporophytic tissue [61]. This anatomy is similar to that of liverworts, except that, in mosses, the seta arises from a transient meristem developing in the middle of the spindle-shaped embryo. In contrast, hornworts lack a seta and, consequently, a seta meristem on their sporophyte. The hornwort sporophyte is composed of a foot and a sporangial axis, growing from a basal meristem that remains active throughout the sporophyte's life [33,62]; they possess a unifacial and multicellular basal meristem, called the basal meristem, which provides upwards indeterminate growth. This type of "indeterminate" growth is unprecedented to any fossil or living plant and is similar to the indeterminate growth of sporophytes in tracheophytes. However, the resemblance to the angiosperm SAM is considered unrelated or superficial [63,64]. The hornwort's indeterminate growth contrasts with the limited proliferative growth of sporophytes in mosses and liverworts.
Bryophytes have an extended gametophyte phase during their life cycle. The gametophyte of bryophytes presents a long-lasting single-celled SAM composed of an apical cell (AC). Among bryophytes, mosses exhibit a singular development compared to hornworts and liverworts [34]. After spore germination, mosses develop filamentous tissues called protonema, consisting of two substages. In the early stage of germination, the protonema is composed of chloronemata filaments. The chloronemata exhibits 1D growth during filament extension until an apical stem cell transitions into a caulonemal apical stem cell. The caulonema consists of an extended 2D filamentous stage. Both substages comprise a protonema apical cell. Lastly, moss initiates 3D growth and leafy shoot buds called gametophores with a single tetrahedral apical cell [54]. In contrast, hornworts and liverworts do not possess a protonemal stage; after spore germination, they grow directly into thalli or "leafy" forms [48,65]. Liverworts grow from apical cells located in an invaginated notch. There are four different types of apical cells found in this group. These include a tetrahedral cell with three cutting faces, a Wedge-shaped with two lateral, one dorsal, and one ventral cutting faces, a lenticular or lens-shaped cell with two lateral cutting faces, and a hemidiscoid cell with two lateral and one posterior cutting face [48]. On the other hand, hornwort gametophytes usually present wedge-shaped apical cells with four cutting faces located in notches around the thallus [66].
3.4. Tracheophytes
Numerous efforts have been made to categorize the SAMs of tracheophytes, primarily through their histological and cytological features. It is generally accepted that there are three types of SAMs. This classification is based on the division plane within the SAM cells and the location and number of initial cells [67,68]. Initial cells divide to maintain the meristem as a continuing source of new cells. The three types of tracheophyte SAMs are 1) monoplex type, usually capped by one apical initial cell, also known as apical cell, that contributes to stem growth [69]. This SAM type is found on some lycophytes and ferns; 2) simplex type, whose configuration contains multiple initial cells within a single zone. These are common in gymnosperms [69]; and 3) duplex meristem type that incorporates initial cells distributed across at least two layers [69,70]. These are prevalent in angiosperms.
The indeterminate meristem of tracheophytes is a crucial innovation in plant evolution, which pushed the sporophyte life cycle as the dominant phase in the plant's life cycle [71]. Lycophytes comprise a taxonomic group representing a small portion of vegetal diversity. This clade consists of three living orders: Lycopodiales, Isoëtales, and Selaginellales. However, there are many other extinct orders [72]. Lycophytes display considerable variation in the organization of their meristems, ranging from the monoplex meristem with one or two apical cells found in Selaginellales [73,74] to the simplex meristems found in Lycopodiales and Isoëtales [70]. Monilophytes (ferns) usually have a meristem composed of an apical initial cell and its surrounding cells, this multicellular structure has also been proposed for Selaginella [42,75]. Apical cells of Lycophytes and Monilophytes (ferns) are considered convergent structures [42,76].
SAMs in seed plants tend to be larger and more complex compared to other land plants. They consist of multiple initial cells and distinct functional layers. The outer layer (L1), also known as tunica, gives rise to the epidermis; and L2 in maize, or L2 and L3 (corpus) in Arabidopsis are the internal layers [77,78,79]. It has been suggested that each layer comprises its own group of initials [77]. Gymnosperms SAMs lack tunica but its cells in the upper portion of the gymnosperm SAM form a lens-shaped zone with cytological features similar to those of angiosperms. Remarkably, there are gymnosperm genera with an apparent or discrete outer L1 layer, such as Gnetum and Ephedra [70].
Irrespective of cell layers, in both gymnosperms and angiosperms SAMs exist zonation patterns (known as well as cellular domains), that mirror differences in cell division or cellular activity. Different zones of the meristem possess different functions. Typically, there is a central zone (CZ) surrounded by a peripheral zone (PZ) on the flanks and a rib zone (RZ) behind the CZ. Cells within the CZ self-renew and replenish the surrounding (PZ) and subtending (RZ). Cells within the PZ and RZ proliferate more rapidly than in the CZ and ultimately differentiate to produce lateral organs and ground tissue, respectively [80,81].
4. Shoot Apical Meristem Regulation and Maintenance
The phenomena of branch development along the primary axis of plants, known as apical dominance, have been studied since the 1930s [82,83,84]. Over the course of nine decades of research, a general concept emerged: the phytohormone auxin produced by the shoot tip is transported in a basipetal fashion by the polar transport stream, which inhibits axillary bud outgrowth [82,85]. Although this concept has been observed in the vegetative developmental stages of many short-lived plants and the juvenile stages of some trees [86], it has not been validated in perennial plants [87,88] and remains obscure in plants with dichotomous branching.
Auxin was the first plant regulator explored in SAM activity, and much has been learned about its signaling pathways in different organs. However, particular attention has been given to its interaction with cytokinin, another plant hormone. Auxins and cytokinins act synergistically or antagonistically to control SAM organization, formation, and maintenance [89]. For example, in Arabidopsis, cytokinins promote cell expansion, increasing SAM size, while auxin indirectly promotes differentiation through multiple mechanisms [90]. Despite being the most extensively studied regulators of plant development across plants, our understanding of the crosstalk between these hormones is still limited to a few plant models.
4.1. The Regulatory Model: Angiosperms
With advances in molecular biology, SAM research has expanded from physiological aspects to the regulatory molecular mechanism. Currently, we understand that phytohormones and TFs cooperate to balance meristem maintenance and organ production. The canonical model of SAM regulation is based on the plant model Arabidopsis thaliana, and stands around the key genes CLAVATA (CLV, encoding ligand peptides) and WUSCHEL (WUS, encoding a homeobox TF) [91]. This regulatory loop also involves SHOOT MERISTEMLESS (STM), a KNOTTED1-LIKE HOMEOBOX (KNOX) TF [10,92]. The CLV gene family comprises the CLV peptides CLV3, CLV2, and CLV1. In this regulatory loop, WUS promotes stem cell identity, and is regulated by CVL and STM. In addition, CLV favors organ initiation, and STM prevents the incorporation of central meristem cells into organ primordia [91,93] (Figure 4).
Other genes, such as the plant-specific GRAS TFs, and HAIRY MERISTEM (HAM1–HAM4), interact with WUS/WOX5 (another member of the WUS family) [94]. A HAM concentration gradient modulates the WUS-CLVs interaction, promoting zonation of the SAM. In addition, members of the NAC group of leucine-rich TFs, including CUP-SHAPED COTYLEDON1 (CUC1), CUC2, and CUC3 [95], repeat receptor-like kinase genes (LRR-RLK), REVOLUTA (REV, a homeodomain TF), and the Apetala 2 (AP2) TF family, have been shown to have critical functions in the SAM [96,97].
4.2. SAM TFs Conserved Throughout Evolution
KNOX TFs
The KNOX genes belong to a large family of TFs called homeobox. They are involved in morphogenesis in all major eukaryotic lineages. In plants, members such as KN1, STM, and KNAT are directly associated with SAM maintenance [53]. Conservation of the KNOX gene family can be traced back to the plant's ancestors, including Chlorophytes and Charophycean algae [53]. It has been suggested that KNOX transcription factors are part of the genetic toolkit that enabled the development of multicellularity, cell differentiation, and increased SAM complexity in seed plants [98]. Their functional roles are diverse across the phylogeny, including activation of the diploid phase, sporophyte and spore formation, meristem maintenance, and organogenesis [99,100]. In organisms such as Chlamydomonas reinhardtii, KNOX, and BELL TFs are inherited by gametes of the opposite mating types and heterodimerize in zygotes to activate diploid development [100,101]. Similarly, in Marchantia polymorpha, the expression in the gametophyte of KNOX and BELL is required to initiate zygotic development, while in Physcomitrella patens, KNOX expression is associated with sporophyte development and meristem regulation rather than the gametophyte [62]. In Lycophytes such as Sellaginella and spermatophytes, KNOX genes have been associated with cell proliferation and meristem maintenance [102,103] Unfortunately, until now, there is not a clear pattern of KNOX function across the phylogeny. However, duplication events followed by a sub-functionalization within each lineage appear to explain the presence of paralogs specialized for several developmental functions [53].
MADS TFs
The MADS-box gene family encodes TFs with a DNA-binding MADS domain, which was named after the proteins MINICHROMOSOME MAINTENANCE 1, AGAMOUS, DEFICIENS, and SERUM RESPONSE FACTOR (SRF). They are classified into two major classes based on their structure and phylogeny, Type I and Type II [104]. Type II classic MADS-box genes have been particularly well studied, as many of them have roles in determining floral organs. Type II MADS-box genes further diverged into two groups: MIKCC and MIKC* a [105]. Phylogenetic analyses suggest that algae MIKCC MADS-box genes could be considered as the ancestral MIKCC before their divergence into the MIKC* and MIKCc clades. In Charophycean algae, MIKCC genes play a role in gamete differentiation [106]
In non-seed plants, MIKCC have roles in gametophytic and sporophytic generations and contribute to the development of vegetative and reproductive structures [107,108,109]. However, in seed plants, their function is primarily linked to sporophyte development and the determination of floral organs [110,111]. On the other hand, MIKC* genes of non-seed and seed plants have a conserved role during gametophyte development [109,112]. It has been hypothesized that the function of MADS-box genes became restricted to specific plant organs after duplication events coinciding with the diversification of seed plants. Before that, MADS-box genes had multiple roles throughout plant development [113].
AP2/ERF TFs
The plant-specific APETALA 2/ethylene-responsive factor (AP2/ERF) family is characterized by the AP2 DNA-binding domain [114]. AP2/ERF genes are divided into classes based on the number of AP2 domains present. ERF-like genes contain one AP2 domain, while AP2-like genes contain two AP2 domains [115]. AP2-like genes can be further divided into the clades euANT, basalANT, and euAP2 according to the amino acid sequence of the double AP2 domain and the nuclear localization sequence [116]. In the model plant Arabidopsis thaliana and other angiosperms, all AP2-like clades play key roles in developmental processes and SAM regulation [98,117,118].
The AP2/ERF family is involved in diverse roles through plant evolution. Although there is no experimental evidence of AP2-like function and no ANT sequences in algae. Phylogenetic analysis showed that an AP2 domain (AP2-R1 AA insertion) of the microalgae C. reinhardtii and Chlorokybus atmophyticus form a sister clade to the major clade of the AP2-like sequences of plants. However, they are distinct from euANTgene sequences, forming their own clade [119]. This suggests that C. reinhardtii and C. atmophyticus AP2 sequences could represent the putative ancestral sequence of the ANT group. In contrast, the AP2-like genes of multicellular algae, such as Mesotaenium caldariorum and Klebsormidium nitens, form a clade with land plants. It has been suggested that the ancestor of embryophytes may have a preANT-like gene which gave rise to the land plant exclusive basalANT and euANT lineages [118,120]. However, the diverse functions of this gene family increased following the plant's evolutionary novelties; for example, in ferns, the expression of ANT has been reported in young sporangia, gametes, and spores, in gymnosperms in the ovule and during seed development. In angiosperms, they are involved in the meristem, flower organ, and fruit development [118,121,122].
5. Taking Advantage of Single-Cell Transcriptomics to Understand SAM
Studies on SAM transcriptional pathways have evolved from early techniques, such as in situ hybridization and microdissection to more high-throughput methods, such as single-cell RNA-sequencing (sc-RNA-seq) and single-nucleus RNA-sequencing (sn-RNA-seq). In situ hybridization is used to map specific genes, making it difficult to obtain a comprehensive view of the transcriptome. Microdissection involves dissecting tissue pieces, from which RNA is extracted for analysis. Unfortunately, microdissection studies are susceptible to RNA degradation and low yields, potentially leading to low-quality data [122,123]. scRNA-seq and snRNA-seq (sc/snRNA-seq) is a technique that avoids these problems. sc/snRNA-seq relies on microfluidics (10X Genomics) to capture single cells or nuclei and obtain an RNA sequencing library for each cell or nuclei. After sequencing, the transcriptomes of individual cells are used to group cells into clusters, and clusters are assigned to cell types using previously described markers from fluorescent protein reporters or in situ hybridization [124].
sc/snRNA-seq simultaneously allows the study of cellular dynamics, including the cell cycle, cell differentiation, and gene regulatory network construction [125]. This is particularly useful in studying meristem development, as it allows the identification of stem cells and their exploration during transitional states up to their differentiation [126]. SAM transcriptomic landscapes are a turning point in plant development research. Sc/snRNA-seq offers the possibility to study division and differentiation at cell resolution, quantifying the contribution of individual cells during the formation of new aerial structures, a task that was impossible before this technology [11]. Currently, three SAM scRNA-seq studies in Arabidopsis [127], one in maize [128], and one in pea [129], and two SAM snRNA-seq studies, one in tomato [130] and one in Populus [131], are available.
The transcriptomic landscape of the Arabidopsis SAM is an invaluable reference due to its status as the most extensively studied plant model species and has led to numerous resources for single-cell analysis. Three main conclusions can be drawn from Arabidopsis SAM scRNA-seq research. First, despite the wealth of available markers, there remain undefined groups of cells, hinting at the potential existence of new cell types. Second, this research also explores the dynamics of transcriptomic changes during temporal processes, enabling the assignment of cells into distinct cell cycle phases, which revealed a cell cycle continuum in meristematic tissues, suggesting that subtle variations in cell division duration and patterns play a role in the formation of different cell types. And third, the overlap of single-cell RNA sequencing data from Arabidopsis RAM and SAM uncovered previously unrecognized similarities between shoot and root apex epidermis and vascular tissues, adding to our understanding of plant development [127].
Conde and colleagues [131] developed a SAM cross-species analysis between Populus and Arabidopsis at single-cell resolution. These results provide novel insights into the conservation of transcriptional programs during primary vasculature formation and yield valuable information about xylem and wood formation. The tomato SAM transcriptomic landscape described by Tian and colleagues [130] showed the enrichment of Arabidopsis homologous genes associated with the cell-specific domains, such as meristem, epidermis, mesophyll, and vasculature. Furthermore, Tian and colleagues [130] used a single-cell approach to infer gene regulatory networks, facilitating the identification of key regulators.
For the first time, Chen and colleagues [129] used sc-RNA-seq technology on pea SAMs to understand plant growth under nutrient stress, specifically boron (B) deficiency. Their study links the progression of SAM development with the up regulation of genes encoding histones and chromatin assembly and remodeling proteins under B deficiency conditions. The expression of these genes was suppressed, suggesting a reorganization of chromatin during SAM development and a possibly impaired SAM activity under B deficiency. These results emphasize the importance of considering cell type-specific stress responses, which may be key to unraveling complex biological processes [129].
Satterly and colleagues [128] obtained a transcriptional landscape of the maize SAM. Maize belongs to the monocot clade, while Arabidopsis, Populus, tomato, and pea are eudicots, and one would expect to observe more differences between the transcriptional profiles of monocots vs eudicots. Based on their scRNA-seq data, Satterly and colleagues [128] did not find evidence of a canonical WUS-CLV3 stem-cell OC as in other SAM transcriptional landscapes. The Arabidopsis SAM exhibits a repression mechanism of the stem-cell-promoting transcription factor WUS, which relies on the peptide CLV3 and its receptor CLV1, which surrounds the OC. WUS is crucial in maintaining the stem-cell fate in SAM and floral meristems (FM). However, in maize SAM, there is no clear regulatory loop similar to that in Arabidopsis.
Maize SAM has several Leucine-Rich-Repeat (LRR) receptors and their peptide ligands that could be involved in stem cell control [132,133], but the identity of the TF(s) that promote the stem-cell fate remains unclear. WUS1 has been considered an essential meristem size regulator in maize [96]. Its expression has been detected in the inflorescence meristem (IM) and FM [134]. In addition, an in-situ hybridization localizes WUS in SAM, but against Arabidopsis, it is not restricted to an OC; it is detected in peripheral cell layers at the primordium height [135]. Remarkable in maize, SAM sc-RNA-seq did not detect WUS or any other functionally homologous to WUS [128]. Laureyns and colleagues [136] develop an in-situ sequencing (ISS) on maize SAM meristem. They probed the expression of 90 genes and could not obtain a clear expression pattern for 15 of them, including ZmWUS1 and ZmWUS2.
Several challenges deriving from the protoplasting technique could hinder Maize SAM heterogeneity due to several issues arising from this process. Protoplasting has been linked with issues such as ectopic gene expression, bias on proportions of cell types [8,127], and noise from generation-related gene expression [96,130]. Therefore, it is possible that WUS-containing cells were affected by protoplasting or its low expression level could not capture by the in-situ sequencing (ISS). Considerations on this phenomenon are: first, the expression of WUS in maize meristem varies depending on its developmental state vegetative (SAM), IM, and FM. Second, no evidence exists that maize SAM expresses a canonical OC in normal conditions. Third, WUS functions as a mobile TF [134], so it is possible that we can only see the location of its expression or accumulation. Remarkably, this example in angiosperms between monocots and dicots exposes the possibility of different mechanisms or regulatory mechanisms on their SAM regulation. Therefore, it is exciting to think about the diversity in SAM regulation on different clades.
Using sc/snRNA-seq to study the SAM in non-model plant species has additional challenges. Sc/snRNA-seq relies on previous knowledge of cell type-specific markers to assign cell clusters to cell types. Such markers are rare to non-existent in non-model plant species. Spatial transcriptomics has emerged as an alternative to overcome this limitation, although it has drawbacks. The spatial transcriptomics technique enables creating in situ libraries, thus obtaining transcriptomic landscapes from tissue sections, avoiding the need to cluster and identify each cell population [137]. Giacomello and colleagues [138] established the basis of the spatial transcriptomics method for SAMs of different plant species [11,139]. The possibility of matching the anatomical and molecular information and accelerating gene and tissue discovery is a valuable resource for future functional genomics studies. Additionally, technologies such as spatial transcriptomics and snRNA-seq will facilitate the study of non-model species as it becomes unnecessary to develop protocols for protoplasting or extensive background on marker genes for particular species. These technologies alleviate the technical difficulties of working on SAMs, as seen in the examples above. However, the advantage of sc/snRNA-seq for genes and TFs discovery and dynamics of regulation cannot be assessed but only by in vitro or in vivo approaches.
6. Perspectives
The study of the SAM is a topic that has captivated humanity historically. Initially, based on anatomical approaches, the prevailing evolutive notion was that the simple meristems composed of one or a few cells were the most primitive, with increased complexity during plant evolution. However, our current understanding suggests that the SAM has multiple origins and that meristems with an apical cell may reappear in vascular clades from ancestors with multicellular meristems [2]. However, further evidence is needed to confirm and specify the multiple origins of SAM. Among all groups of plants, angiosperms are the most studied. However, research into the conservation of SAM regulatory pathways is still in its early stages.
Angiosperm meristems, mainly dicot meristems, have been extensively studied, and there is an increasing interest in deciphering if SAM regulatory pathways are conserved among diverse groups of plants. Evidence suggests that monocot and dicot meristems could exhibit distinct regulatory loops. However, if such diversity exists within angiosperms, one might wonder about its prevalence in other plant groups. For instance, does research on meristem regulation in bryophytes and lycophytes need to commence without assuming similar regulatory mechanisms?
Significant progress has been made in understanding the maize SAM through single-cell and spatial transcriptomics. However, developing this technique in non-model species remains challenging mainly for three reasons. First, protocols for sc/snRNA-seq in plants are species-dependent. Second, developing these techniques on meristems with only one apical cell is challenging as the cell recovery on the overall efficiency of current scRNA-seq protocols can vary between <1% to >60% across cells, depending on the method used [140]. Therefore, ensuring the representation of a sufficient percentage of apical cells would necessitate adept technical skills. Third, as sc/snRNA-seq analyses rely on gene markers to identify clusters, the poor genomic information available for non-model species will limit their significance. This will also be impacted by the lack of information regarding cellular domains of the SAM and gene expression and regulatory pathways involved in regulation. Therefore, all this information must be generated in advance.
Comparative studies between multiple species have the potential to shed light on the regulatory pathways of the SAM, help clarify its origin, and address related questions. For instance, are there other regulatory cycles besides the CLV-WUS? Do they act in a concerted way or independently? Or are they species-specific? That would help explore the presence of conserved or non-conserved regulatory loops in monocots and other plant groups. Furthermore, it will be essential to investigate whether the expansion of SAM regulatory genes relates to plant morphological traits across plant phylogeny. Another exciting perspective is to search for conserved mechanisms that promote and maintain cell dedifferentiation and proliferation, which would have a great impact on different fields of research.
Author Contributions
Conceptualization, writing—original draft preparation, T.K-G; figure preparation T.K-G and DLLA; writing—Review and editing, DLLA and LHE. All authors reviewed the results and approved the final version of the manuscript.
Funding
This work was partially supported by Cotton Incorporated Cary, NC grant 20-675, USDA-AFRI grant 2021-67013-34738, and the State of Texas Governor's University Research Initiative (GURI)/Texas Tech University grant 05-2018.
Data Availability Statement
No new data were created in this study.
Acknowledgments
We thank Ricardo A. Chávez Montes for critical reading of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Murray, J.A.H.; Jones, A.; Godin, C.; Traas, J. Systems Analysis of Shoot Apical Meristem Growth and Development: Integrating Hormonal and Mechanical Signaling. Plant Cell 2012, 24, 3907–3919. [Google Scholar] [CrossRef]
- Jill Harrison, C. Development and Genetics in the Evolution of Land Plant Body Plans. Philosophical Transactions of the Royal Society B: Biological Sciences 2017, 372. [Google Scholar] [CrossRef]
- Banks, J.A. The Evolution of the Shoot Apical Meristem from a Gene Expression Perspective. New Phytologist 2015, 207, 486–487. [Google Scholar] [CrossRef]
- Grafi, G.; Florentin, A.; Ransbotyn, V.; Morgenstern, Y. The Stem Cell State in Plant Development and in Response to Stress. Front Plant Sci 2011, 2. [Google Scholar] [CrossRef]
- Takebe, I.; Labib, G.; Melchers, G. Regeneration of Whole Plants from Isolated Mesophyll Protoplasts of Tobacco. Naturwissenschaften 1971, 58, 318–320. [Google Scholar] [CrossRef]
- Fehér, A. Callus, Dedifferentiation, Totipotency, Somatic Embryogenesis: What These Terms Mean in the Era of Molecular Plant Biology? Front Plant Sci 2019, 10, 442509. [Google Scholar] [CrossRef]
- Boutilier, K.; Offringa, R.; Sharma, V.K.; Kieft, H.; Ouellet, T.; Zhang, L.; Hattori, J.; Liu, C.M.; Van Lammeren, A.A.M.; Miki, B.L.A.; et al. Ectopic Expression of BABY BOOM Triggers a Conversion from Vegetative to Embryonic Growth. Plant Cell 2002, 14, 1737–1749. [Google Scholar] [CrossRef]
- Xu, M.; Du, Q.; Tian, C.; Wang, Y.; Jiao, Y. Stochastic Gene Expression Drives Mesophyll Protoplast Regeneration. Sci Adv 2021, 7, 8466–8477. [Google Scholar] [CrossRef]
- Shim, S.; Kim, H.K.; Bae, S.H.; Lee, H.; Lee, H.J.; Jung, Y.J.; Seo, P.J. Transcriptome Comparison between Pluripotent and Non-Pluripotent Calli Derived from Mature Rice Seeds. Scientific Reports 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A Member of the KNOTTED Class of Homeodomain Proteins Encoded by the STM Gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef]
- Stahl, Y.; Simon, R. Plant Primary Meristems: Shared Functions and Regulatory Mechanisms. Curr Opin Plant Biol 2010, 13, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Seago, J.L.; Fernando, D.D. Anatomical Aspects of Angiosperm Root Evolution. Ann Bot 2013, 112, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Haecker, A.; Groß-Hardt, R.; Geiges, B.; Sarkar, A.; Breuninger, H.; Herrmann, M.; Laux, T. Expression Dynamics of WOX Genes Mark Cell Fate Decisions during Early Embryonic Patterning in Arabidopsis Thaliana. Development 2004, 131, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Bennett, T.; Scheres, B. Root Development-Two Meristems for the Price of One? Curr Top Dev Biol 2010, 91, 67–102. [Google Scholar] [CrossRef]
- Sarkar, A.K.; Luijten, M.; Miyashima, S.; Lenhard, M.; Hashimoto, T.; Nakajima, K.; Scheres, B.; Heidstra, R.; Laux, T. Conserved Factors Regulate Signalling in Arabidopsis Thaliana Shoot and Root Stem Cell Organizers. Nature 2007, 446, 811–814. [Google Scholar] [CrossRef]
- Skylar, A.; Wu, X. Regulation of Meristem Size by Cytokinin SignalingF. J Integr Plant Biol 2011, 53, 446–454. [Google Scholar] [CrossRef]
- Greb, T.; Lohmann, J.U. Plant Stem Cells. Curr Biol 2016, 26, R816–R821. [Google Scholar] [CrossRef]
- Mähönen, A.P.; Ten Tusscher, K.; Siligato, R.; Smetana, O.; Díaz-Triviño, S.; Salojärvi, J.; Wachsman, G.; Prasad, K.; Heidstra, R.; Scheres, B. PLETHORA Gradient Formation Mechanism Separates Auxin Responses. Nature 2014, 515, 125–129. [Google Scholar] [CrossRef]
- Dello Ioio, R.; Galinha, C.; Fletcher, A.G.; Grigg, S.P.; Molnar, A.; Willemsen, V.; Scheres, B.; Sabatini, S.; Baulcombe, D.; Maini, P.K.; et al. A PHABULOSA/Cytokinin Feedback Loop Controls Root Growth in Arabidopsis. Curr Biol 2012, 22, 1699–1704. [Google Scholar] [CrossRef]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The Yin-Yang of Hormones: Cytokinin and Auxin Interactions in Plant Development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef]
- Friedman, W.E.; Moore, R.C.; Purugganan, M.D. The Evolution of Plant Development. Am J Bot 2004, 91, 1726–1741. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.J.; Dolan, L. Stepwise and Independent Origins of Roots among Land Plants. Nature 2018, 561, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xu, L. Recruitment of IC-WOX Genes in Root Evolution. Trends Plant Sci 2018, 23, 490–496. [Google Scholar] [CrossRef]
- Fujinami, R.; Yamada, T.; Imaichi, R. Root Apical Meristem Diversity and the Origin of Roots: Insights from Extant Lycophytes. J Plant Res 2020, 133, 291–296. [Google Scholar] [CrossRef]
- Huang, L.; Schiefelbein, J. Conserved Gene Expression Programs in Developing Roots from Diverse Plants. Plant Cell 2015, 27, 2119–2132. [Google Scholar] [CrossRef] [PubMed]
- Augstein, F.; Carlsbecker, A. Getting to the Roots: A Developmental Genetic View of Root Anatomy and Function from Arabidopsis to Lycophytes. Front Plant Sci 2018, 9, 411407. [Google Scholar] [CrossRef] [PubMed]
- Leebens-Mack, J.H.; Barker, M.S.; Carpenter, E.J.; Deyholos, M.K.; Gitzendanner, M.A.; Graham, S.W.; Grosse, I.; Li, Z.; Melkonian, M.; Mirarab, S.; et al. One Thousand Plant Transcriptomes and the Phylogenomics of Green Plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef]
- Ferrari, C.; Shivhare, D.; Hansen, B.O.; Pasha, A.; Esteban, E.; Provart, N.J.; Kragler, F.; Fernie, A.; Tohge, T.; Mutwil, M. Expression Atlas of Selaginella Moellendorffii Provides Insights into the Evolution of Vasculature, Secondary Metabolism, and Roots. Plant Cell 2020, 32, 853–870. [Google Scholar] [CrossRef]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin-Cytokinin Interaction Regulates Meristem Development. Mol Plant 2011, 4, 616–625. [Google Scholar] [CrossRef]
- Luo, L.; Zeng, J.; Wu, H.; Tian, Z.; Zhao, Z. A Molecular Framework for Auxin-Controlled Homeostasis of Shoot Stem Cells in Arabidopsis. Mol Plant 2018, 11, 899–913. [Google Scholar] [CrossRef]
- Allen, C.E. Am Erican Naturalist The American Society Of Naturalists Haploid And Diploid Generations’; 1937. [Google Scholar]
- Albert, V.A. Shoot Apical Meristems and Floral Patterning: An Evolutionary Perspective. Trends Plant Sci 1999, 4, 84–86. [Google Scholar] [CrossRef]
- Ligrone, R.; Duckett, J.G.; Renzaglia, K.S. The Origin of the Sporophyte Shoot in Land Plants: A Bryological Perspective. Ann Bot 2012, 110, 935. [Google Scholar] [CrossRef] [PubMed]
- Hata, Y.; Kyozuka, J. Fundamental Mechanisms of the Stem Cell Regulation in Land Plants: Lesson from Shoot Apical Cells in Bryophytes. Plant Mol Biol 2021, 107, 213–225. [Google Scholar] [CrossRef]
- Wu, X.; Yan, A.; McAdam, S.A.M.; Banks, J.A.; Zhang, S.; Zhou, Y. Timing of Meristem Initiation and Maintenance Determines the Morphology of Fern Gametophytes. J Exp Bot 2021, 72, 6990–7001. [Google Scholar] [CrossRef] [PubMed]
- Haig, D. Homologous Versus Antithetic Alternation of Generations and the Origin of Sporophytes. The Botanical Review 2008, 74, 395–418. [Google Scholar] [CrossRef]
- Niklas, K.J.; Kutschera, U. The Evolution of the Land Plant Life Cycle. New Phytologist 2010, 185, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Bennici, A. Origin and Early Evolution of Land Plants Problems and Considerations; Volume 1.
- Hemsley, A.R. The origin of the land plant sporophyte: An interpolational scenario. Biological Reviews 1994, 69, 263–273. [Google Scholar] [CrossRef]
- Kenrick, P. Alternation of generations in land plants: New phylogenetic and palaeobotanical evidence. Biological Reviews 1994, 69, 293–330. [Google Scholar] [CrossRef]
- Harrison, C.J. Auxin Transport in the Evolution of Branching Forms. New Phytologist 2017, 215, 545–551. [Google Scholar] [CrossRef]
- Frank, M.H.; Scanlon, M.J. Transcriptomic Evidence for the Evolution of Shoot Meristem Function in Sporophyte-Dominant Land Plants through Concerted Selection of Ancestral Gametophytic and Sporophytic Genetic Programs. Mol Biol Evol 2015, 32, 355–367. [Google Scholar] [CrossRef]
- Youngstrom, C.E.; Geadelmann, L.F.; Irish, E.E.; Cheng, C.L. A Fern WUSCHEL-RELATED HOMEOBOX Gene Functions in Both Gametophyte and Sporophyte Generations. BMC Plant Biol 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Popham, R.A. principal types of vegetative shoot apex organization in vascular plants 1. Ohio J Sci 1951, 51. [Google Scholar]
- Renzaglia, K.S. A Comparative Developmental Investigation of the Gametophyte Generation in the Metzgeriales (Hepatophyta). 1982, 253. [Google Scholar]
- Korn, R.W. Apical Cells As Meristems; KluwerAcademic Publishers, 1993; Volume 41. [Google Scholar]
- Nishihama, R.; Naramoto, S. Apical Stem Cells Sustaining Prosperous Evolution of Land Plants. J Plant Res 2020, 133, 279–282. [Google Scholar] [CrossRef]
- Shaw, J.; Renzaglia, K. Phylogeny and Diversification of Bryophytes. Am J Bot 2004, 91, 1557–1581. [Google Scholar] [CrossRef]
- Niklas, K.J.; Wayne, R.; Benítez, M.; Newman, S.A. Polarity, Planes of Cell Division, and the Evolution of Plant Multicellularity. Protoplasma 2019, 256, 585–599. [Google Scholar] [CrossRef]
- Stewart, K.D.; Mattox, K.R. Some Aspects of Mitosis in Primitive Green Algae: Phylogeny and Function. Biosystems 1975, 7, 310–315. [Google Scholar] [CrossRef]
- Graham, L.E.; Cook, M.E.; Busse, J.S. The Origin of Plants: Body Plan Changes Contributing to a Major Evolutionary Radiation. Proc Natl Acad Sci U S A 2000, 97, 4535–4540. [Google Scholar] [CrossRef]
- Umen, J.G. Green Algae and the Origins of Multicellularity in the Plant Kingdom. Cold Spring Harb Perspect Biol 2014, 6. [Google Scholar] [CrossRef]
- Graham, L.E.; Cook, M.E.; Busse, J.S. The Origin of Plants: Body Plan Changes Contributing to a Major Evolutionary Radiation; 2000; Volume 97. [Google Scholar]
- Moody, L.A. Unravelling 3D Growth in the Moss Physcomitrium Patens. Essays Biochem 2022, 66, 769–779. [Google Scholar]
- Graham, L.E. Coleochaete And The Origin Of Land Plants. Am J Bot 1984, 71, 603–608. [Google Scholar] [CrossRef]
- Puttick, M.N.; Morris, J.L.; Williams, T.A.; Cox, C.J.; Edwards, D.; Kenrick, P.; Pressel, S.; Wellman, C.H.; Schneider, H.; Pisani, D.; et al. The Interrelationships of Land Plants and the Nature of the Ancestral Embryophyte. Current Biology 2018, 28, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Wolf, P.G.; Kugita, M.; Sinclair, R.B.; Sugita, M.; Sugiura, C.; Wakasugi, T.; Yamada, K.; Yoshinaga, K.; Yamaguchi, K.; et al. Chloroplast Phylogeny Indicates That Bryophytes Are Monophyletic. Mol Biol Evol 2004, 21, 1813–1819. [Google Scholar] [CrossRef]
- Renzaglia, K.S.; Garbary, D.J. Motile Gametes of Land Plants: Diversity, Development, and Evolution. Critical reviews in plant sciences 2010, 20, 107–213. [Google Scholar] [CrossRef]
- Campbell, D.H. The Structure & Development of the Mosses & Ferns (Archegoniatae); Macmillan, 1895. [Google Scholar]
- Tomescu, A.M.F.; Wyatt, S.E.; Hasebe, M.; Rothwell, G.W. Early Evolution of the Vascular Plant Body Plan — the Missing Mechanisms. Curr Opin Plant Biol 2014, 17, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Ligrone, R.; Duckett, J.G.; Renzaglia, K.S. Conducting Tissues and Phyletic Relationships of Bryophytes. Philos Trans R Soc Lond B Biol Sci 2000, 355, 795–813. [Google Scholar] [CrossRef]
- Sakakibara, K.; Nishiyama, T.; Deguchi, H.; Hasebe, M. Class 1 KNOX Genes Are Not Involved in Shoot Development in the Moss Physcomitrella Patens but Do Function in Sporophyte Development. Evol Dev 2008, 10, 555–566. [Google Scholar] [CrossRef]
- Kato, M.; Akiyama, H. Interpolation Hypothesis for Origin of the Vegetative Sporophyte of Land Plants; 2005; Volume 54. [Google Scholar]
- Renzaglia, K.S.; Schuette, S.; Duff, R.J.; Ligrone, R.; Shaw, A.J.; Mishler, B.D.; Duckett, J.G. Bryophyte Phylogeny: Advancing the Molecular and Morphological Frontiers; 2007; Volume 110. [Google Scholar]
- Solly, J.E.; Cunniffe, N.J.; Harrison, C.J. Regional Growth Rate Differences Specified by Apical Notch Activities Regulate Liverwort Thallus Shape. Curr Biol 2017, 27, 16–26. [Google Scholar] [CrossRef]
- Renzaglia, A.; Wehman, P.; Schutz, R.; Karan, O. Use of Cue Redundancy and Positive Reinforcement to Accelerate Production in Two Profoundly Retarded Workers. British Journal of Social and Clinical Psychology 1978, 17, 183–187. [Google Scholar] [CrossRef]
- Esau, K. Plant Anatomy. Plant Anatomy 1965. [Google Scholar] [CrossRef]
- Gifford, E.M.; Corson, G.E. The Shoot Apex in Seed Plants. The Botanical Review 1971, 37, 143–229. [Google Scholar] [CrossRef]
- Newman, I.V. Pattern in the Meristems of Vascular Plants: III. Pursuing the Patterns in the Apical Meristem Where No Cell Is a Permanent Cell*. Journal of the Linnean Society of London, Botany 1965, 59, 185–214. [Google Scholar] [CrossRef]
- Imaichi, R.; Hiratsuka, R. Evolution of Shoot Apical Meristem Structures in Vascular Plants with Respect to Plasmodesmatal Network. Am J Bot 2007, 94, 1911–1921. [Google Scholar] [CrossRef] [PubMed]
- Fouracre, J.P.; Harrison, C.J. How Was Apical Growth Regulated in the Ancestral Land Plant? Insights from the Development of Non-Seed Plants. Plant Physiol 2022, 190, 100–112. [Google Scholar] [CrossRef]
- Spencer, V.; Nemec Venza, Z.; Harrison, C.J. What Can Lycophytes Teach Us about Plant Evolution and Development? Modern Perspectives on an Ancient Lineage. Evol Dev 2021, 23, 174–196. [Google Scholar] [CrossRef]
- Harrison, C.J.; Rezvani, M.; Langdale, J.A. Growth from Two Transient Apical Initials in the Meristem of Selaginella Kraussiana. Development 2007, 134, 881–889. [Google Scholar] [CrossRef]
- Jones, C.S.; Drinnan, A.N. The Developmental Pattern of Shoot Apices in Selaginella Kraussiana (Kunze) a. Braun. Int J Plant Sci 2009, 170, 1009–1018. [Google Scholar] [CrossRef]
- Ambrose, B.A.; Vasco, A. Bringing the Multicellular Fern Meristem into Focus. New Phytologist 2016, 210, 790–793. [Google Scholar] [CrossRef]
- White, R.A.; Turner, M.D. Anatomy and Development of the Fern Sporophyte. The Botanical Review 1995, 61, 281–305. [Google Scholar] [CrossRef]
- Gifford, E.M. The Structure and Development of the Shoot Apex in Certain Woody Ranales; 1950; Volume 37. [Google Scholar]
- Kessler, S.; Townsley, B.; Sinha, N. L1 Division and Differentiation Patterns Influence Shoot Apical Meristem Maintenance. Plant Physiol 2006, 141, 1349. [Google Scholar] [CrossRef]
- Sinha, N.; Hake, S. Mutant Characters of Knotted Maize Leaves Are Determined in the Innermost Tissue Layers. Dev Biol 1990, 141, 203–210. [Google Scholar] [CrossRef]
- Gifford, E.M.; Wetmore, R.H. APICAL MERISTEMS OF VEGETATIVE SHOOTS AND STROBILI IN CERTAIN GYMNOSPERMS. Proc Natl Acad Sci U S A 1957, 43, 571–576. [Google Scholar] [CrossRef]
- Barton, M.K. Twenty Years on: The Inner Workings of the Shoot Apical Meristem, a Developmental Dynamo. Dev Biol 2010, 341, 95–113. [Google Scholar] [CrossRef]
- Kebrom, T.H. A Growing Stem Inhibits Bud Outgrowth – The Overlooked Theory of Apical Dominance. Front Plant Sci 2017, 8, 309506. [Google Scholar] [CrossRef] [PubMed]
- Thimann, K.V. Auxins And The Inhibition Of Plant Growth.
- Went, F.W. Auxin, the Plant Growth-Hormone. Review 1935, 1, 162–182. [Google Scholar] [CrossRef]
- Sussex, I.M.; Kerk, N.M. The Evolution of Plant Architecture. Curr Opin Plant Biol 2001, 4, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Cline, M.G. Execution of the Auxin Replacement Apical Dominance Experiment in Temperate Woody Species. Am J Bot 2000, 87, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Norstog, K.; Nicholls, T.J. The Biology of the Cycads. The Biology of the Cycads 2020. [Google Scholar] [CrossRef]
- Napoli, C.A.; Beveridge, C.A.; Snowden, K.C. 5 Reevaluating Concepts of Apical Dominance and the Control of Axillary Bud Outgrowth. Curr Top Dev Biol 1998, 44, 127–169. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin Biosynthesis and Its Role in Plant Development. Annual review of plant biology 2010, 61, 49–64. [Google Scholar] [CrossRef]
- Azizi, P.; Rafii, M.Y.; Maziah, M.; Abdullah, S.N.A.; Hanafi, M.M.; Latif, M.A.; Rashid, A.A.; Sahebi, M. Understanding the Shoot Apical Meristem Regulation: A Study of the Phytohormones, Auxin and Cytokinin, in Rice. Mech Dev 2015, 135, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Schoof, H.; Lenhard, M.; Haecker, A.; Mayer, K.F.X.; Jürgens, G.; Laux, T. The Stem Cell Population of Arabidopsis Shoot Meristems Is Maintained by a Regulatory Loop between the CLAVATA and WUSCHEL Genes. Cell 2000, 100, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Laux, T.; Mayer, K.F.X.; Berger, J.; Jürgens, G. The WUSCHEL Gene Is Required for Shoot and Floral Meristem Integrity in Arabidopsis. Development 1996, 122, 87–96. [Google Scholar] [CrossRef]
- Endrizzi, K.; Moussian, B.; Haecker, A.; Levin, J.Z.; Laux, T. The SHOOT MERISTEMLESS Gene Is Required for Maintenance of Undifferentiated Cells in Arabidopsis Shoot and Floral Meristems and Acts at a Different Regulatory Level than the Meristem Genes WUSCHEL and ZWILLE. The Plant Journal 1996, 10, 967–979. [Google Scholar] [CrossRef]
- Zhou, X.; Guo, Y.; Zhao, P.; Sun, M.X. Comparative Analysis of WUSCHEL-Related Homeobox Genes Revealed Their Parent-of-Origin and Cell Type-Specific Expression Pattern during Early Embryogenesis in Tobacco. Front Plant Sci 2018, 9, 337885. [Google Scholar] [CrossRef] [PubMed]
- Aida, M.; Tasaka, M. Genetic Control of Shoot Organ Boundaries. Curr Opin Plant Biol 2006, 9, 72–77. [Google Scholar] [CrossRef]
- Zhang, W. Putting Genes on the Map: Spatial Transcriptomics of the Maize Shoot Apical Meristem. Plant Physiol 2022, 188, 1931–1932. [Google Scholar] [CrossRef]
- Shimotohno, A.; Scheres, B. Topology of Regulatory Networks That Guide Plant Meristem Activity: Similarities and Differences. Curr Opin Plant Biol 2019, 51, 74–80. [Google Scholar] [CrossRef]
- Floyd, S.K.; Bowman, J.L. The Ancestral Developmental Tool Kit of Land Plants. Int J Plant Sci 2007, 168, 1–35. [Google Scholar] [CrossRef]
- Dierschke, T.; Flores-Sandoval, E.; Rast-Somssich, M.I.; Althoff, F.; Zachgo, S.; Bowman, J.L. Gamete Expression of Tale Class Hd Genes Activates the Diploid Sporophyte Program in Marchantia Polymorpha. Elife 2021, 10. [Google Scholar] [CrossRef]
- Hisanaga, T.; Fujimoto, S.; Cui, Y.; Sato, K.; Sano, R.; Yamaoka, S.; Kohchi, T.; Berger, F.; Nakajima, K. Deep Evolutionary Origin of Gamete-Directed Zygote Activation by KNOX/ BELL Transcription Factors in Green Plants. Elife 2021, 10. [Google Scholar] [CrossRef]
- Lee, J.H.; Lin, H.; Joo, S.; Goodenough, U. Early Sexual Origins of Homeoprotein Heterodimerization and Evolution of the Plant KNOX/BELL Family. Cell 2008, 133, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Harrison, C.J.; Coriey, S.B.; Moylan, E.C.; Alexander, D.L.; Scotland, R.W.; Langdale, J.A. Independent Recruitment of a Conserved Developmental Mechanism during Leaf Evolution. Nature 2005, 434, 509–514. [Google Scholar] [CrossRef]
- Kawai, J.; Tanabe, Y.; Soma, S.; Ito, M. Class 1 KNOX Gene Expression Supports the Selaginella Rhizophore Concept. Journal of Plant Biology 2010, 53, 268–274. [Google Scholar] [CrossRef]
- Alvarez-Buylla, E.R.; Liljegren, S.J.; Pelaz, S.; Gold, S.E.; Burgeff, C.; Ditta, G.S.; Vergara-Silva, F.; Yanofsky, M.F. MADS-Box Gene Evolution beyond Flowers: Expression in Pollen, Endosperm, Guard Cells, Roots and Trichomes. The Plant Journal 2000, 24, 457–466. [Google Scholar] [CrossRef]
- Henschel, K.; Kofuji, R.; Hasebe, M.; Saedler, H.; Münster, T.; Theißen, G. Two Ancient Classes of MIKC-Type MADS-Box Genes Are Present in the Moss Physcomitrella Patens. Mol Biol Evol 2002, 19, 801–814. [Google Scholar] [CrossRef]
- Tanabe, Y.; Hasebe, M.; Sekimoto, H.; Nishiyama, T.; Kitani, M.; Henschel, K.; Mü nster, T.; nter Theissen, G.; Nozaki, H.; Ito, M. Characterization of MADS-Box Genes in Charophycean Green Algae and Its Implication for the Evolution of MADS-Box Genes; 2005.
- Singer, S.D.; Krogan, N.T.; Ashton, N.W. Clues about the Ancestral Roles of Plant MADS-Box Genes from a Functional Analysis of Moss Homologues. Plant Cell Rep 2007, 26, 1155–1169. [Google Scholar] [CrossRef]
- Koshimizu, S.; Kofuji, R.; Sasaki-Sekimoto, Y.; Kikkawa, M.; Shimojima, M.; Ohta, H.; Shigenobu, S.; Kabeya, Y.; Hiwatashi, Y.; Tamada, Y.; et al. Physcomitrella MADS-Box Genes Regulate Water Supply and Sperm Movement for Fertilization. Nature Plants 2018, 4, 36–45. [Google Scholar] [CrossRef]
- Thangavel, G.; Nayar, S. A Survey of MIKC Type MADS-Box Genes in Non-Seed Plants: Algae, Bryophytes, Lycophytes and Ferns. Front Plant Sci 2018, 9, 342764. [Google Scholar] [CrossRef]
- Becker, A.; Theißen, G. The Major Clades of MADS-Box Genes and Their Role in the Development and Evolution of Flowering Plants. Mol Phylogenet Evol 2003, 29, 464–489. [Google Scholar] [CrossRef]
- Ferrario, S.; Shchennikova, A.V.; Franken, J.; Immink, R.G.H.; Angenent, G.C. Control of Floral Meristem Determinacy in Petunia by MADS-Box Transcription Factors. Plant Physiol 2006, 140, 890–898. [Google Scholar] [CrossRef]
- Zobell, O.; Faigl, W.; Saedler, H.; Münster, T. MIKC* MADS-Box Proteins: Conserved Regulators of the Gametophytic Generation of Land Plants. Mol Biol Evol 2010, 27, 1201–1211. [Google Scholar] [CrossRef]
- Ambrose, B.A.; Smalls, T.L.; Zumajo-Cardona, C. All Type II Classic MADS-Box Genes in the Lycophyte Selaginella Moellendorffii Are Broadly yet Discretely Expressed in Vegetative and Reproductive Tissues. Evol Dev 2021, 23, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP Family of Plant Transcription Factors. Biol Chem 1998, 379, 633–654. [Google Scholar] [CrossRef] [PubMed]
- Gutterson, N.; Reuber, T.L. Regulation of Disease Resistance Pathways by AP2/ERF Transcription Factors. Curr Opin Plant Biol 2004, 7, 465–471. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, R.; Xu, D.; Bi, H.; Xia, Z.; Peng, H. Genome-Wide Identification and Analysis of the AP2 Transcription Factor Gene Family in Wheat (Triticum Aestivum L.). Front Plant Sci 2019, 10, 486684. [Google Scholar] [CrossRef]
- Kim, S.; Soltis, P.S.; Wall, K.; Soltis, D.E. Phylogeny and Domain Evolution in the APETALA2-like Gene Family. Mol Biol Evol 2006, 23, 107–120. [Google Scholar] [CrossRef]
- Zumajo-Cardona, C.; Vasco, A.; Ambrose, B.A. The Evolution of the KANADI Gene Family and Leaf Development in Lycophytes and Ferns. Plants 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Dipp-álvarez, M.; Cruz-Ramírez, A. A Phylogenetic Study of the ANT Family Points to a PreANT Gene as the Ancestor of Basal and EuANT Transcription Factors in Land Plants. Front Plant Sci 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Gutterson, N.; Reuber, T.L. Regulation of Disease Resistance Pathways by AP2/ERF Transcription Factors. Curr Opin Plant Biol 2004, 7, 465–471. [Google Scholar] [CrossRef]
- Bui, L.T.; Pandzic, D.; Youngstrom, C.E.; Wallace, S.; Irish, E.E.; Szövényi, P.; Cheng, C.L. A Fern AINTEGUMENTA Gene Mirrors BABY BOOM in Promoting Apogamy in Ceratopteris Richardii. The Plant Journal 2017, 90, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Krizek, B.A.; Bantle, A.T.; Heflin, J.M.; Han, H.; Freese, N.H.; Loraine, A.E. AINTEGUMENTA and AINTEGUMENTA-LIKE6 Directly Regulate Floral Homeotic, Growth, and Vascular Development Genes in Young Arabidopsis Flowers. J Exp Bot 2021, 72, 5478–5493. [Google Scholar] [CrossRef] [PubMed]
- Farris, S.; Wang, Y.; Ward, J.M.; Dudek, S.M. Optimized Method for Robust Transcriptome Profiling of Minute Tissues Using Laser Capture Microdissection and Low-Input RNA-Seq. Front Mol Neurosci 2017, 10, 272672. [Google Scholar] [CrossRef] [PubMed]
- Rakszewska, A.; Tel, J.; Chokkalingam, V.; Huck, W.T.S. One Drop at a Time: Toward Droplet Microfluidics as a Versatile Tool for Single-Cell Analysis. NPG Asia Materials 2014, 6, e133. [Google Scholar] [CrossRef]
- Scialdone, A.; Natarajan, K.N.; Saraiva, L.R.; Proserpio, V.; Teichmann, S.A.; Stegle, O.; Marioni, J.C.; Buettner, F. Computational Assignment of Cell-Cycle Stage from Single-Cell Transcriptome Data. Methods 2015, 85, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Bendall, S.C.; Davis, K.L.; Amir, E.A.D.; Tadmor, M.D.; Simonds, E.F.; Chen, T.J.; Shenfeld, D.K.; Nolan, G.P.; Pe’Er, D. Single-Cell Trajectory Detection Uncovers Progression and Regulatory Coordination in Human B Cell Development. Cell 2014, 157, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Q.; Chen, Y.; Wang, J.W. A Single-Cell Analysis of the Arabidopsis Vegetative Shoot Apex. Dev Cell 2021, 56, 1056–1074. [Google Scholar] [CrossRef]
- Satterlee, J.W.; Strable, J.; Scanlon, M.J. Plant Stem-Cell Organization and Differentiation at Single-Cell Resolution. Proceedings of the National Academy of Sciences 2020, 117. [Google Scholar] [CrossRef]
- Chen, X.; Ru, Y.; Takahashi, H.; Nakazono, M.; Shabala, S.; Smith, S.M.; Yu, M. Single-Cell Transcriptomic Analysis of Pea Shoot Development and Cell-Type-Specific Responses to Boron Deficiency. bioRxiv 2023. [Google Scholar] [CrossRef]
- Tian, C.; Du, Q.; Xu, M.; Du, F.; Jiao, Y. Single-Nucleus RNA-Seq Resolves Spatiotemporal Developmental Trajectories in the Tomato Shoot Apex. bioRxiv 2020. [Google Scholar] [CrossRef]
- Conde, D.; Triozzi, P.M.; Pereira, W.J.; Schmidt, H.W.; Balmant, K.M.; Knaack, S.A.; Redondo-López, A.; Roy, S.; Dervinis, C.; Kirst, M. Single-Nuclei Transcriptome Analysis of the Shoot Apex Vascular System Differentiation in Populus. Development (Cambridge) 2022, 149. [Google Scholar] [CrossRef] [PubMed]
- Bommert, P.; Lunde, C.; Nardmann, J.; Vollbrecht, E.; Running, M.; Jackson, D.; Hake, S.; Werr, W. Thick Tassel Dwarf1 Encodes a Putative Maize Ortholog of the Arabidopsis CLAVATA1 Leucine-Rich Repeat Receptor-like Kinase. Development 2005, 132, 1235–1245. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, F.; Jackson, D. All Together Now, a Magical Mystery Tour of the Maize Shoot Meristem. Curr Opin Plant Biol 2018, 45, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Somssich, M.; Je, B.I.; Simon, R.; Jackson, D. CLAVATA-WUSCHEL Signaling in the Shoot Meristem. Development 2016, 143, 3238–3248. [Google Scholar] [CrossRef] [PubMed]
- Nardmann, J.; Werr, W. The Invention of WUS-like Stem Cell-Promoting Functions in Plants Predates Leptosporangiate Ferns. Plant Mol Biol 2012, 78, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Laureyns, R.; Joossens, J.; Herwegh, D.; Pevernagie, J.; Pavie, B.; Demuynck, K.; Debray, K.; Coussens, G.; Pauwels, L.; van Hautegem, T.; et al. An in Situ Sequencing Approach Maps PLASTOCHRON1 at the Boundary between Indeterminate and Determinate Cells. Plant Physiol 2022, 188, 782–794. [Google Scholar] [CrossRef]
- Ståhl, P.L.; Salmén, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and Analysis of Gene Expression in Tissue Sections by Spatial Transcriptomics. Science (1979) 2016, 353, 78–82. [Google Scholar] [CrossRef]
- Giacomello, S.; Lundeberg, J. Preparation of Plant Tissue to Enable Spatial Transcriptomics Profiling Using Barcoded Microarrays. Nat Protoc 2018, 13, 2425–2446. [Google Scholar] [CrossRef]
- Giacomello, S.; Salmén, F.; Terebieniec, B.K.; Vickovic, S.; Navarro, J.F.; Alexeyenko, A.; Reimegård, J.; McKee, L.S.; Mannapperuma, C.; Bulone, V.; et al. Spatially Resolved Transcriptome Profiling in Model Plant Species. Nat Plants 2017, 3. [Google Scholar] [CrossRef]
- Svensson, V.; Natarajan, K.N.; Ly, L.H.; Miragaia, R.J.; Labalette, C.; Macaulay, I.C.; Cvejic, A.; Teichmann, S.A. Power Analysis of Single-Cell RNA-Sequencing Experiments. Nature Methods 2017, 14, 381–387. [Google Scholar] [CrossRef]
Figure 1.
Diagram of SAM and RAM in Arabidopsis thaliana. In the SAM, a stem cell pool is located above the organizing center (OC, purple), which expresses the WUSCHEL transcription factor (TF) (yellow). Cells that pass the boundary defined by CLAVATA (CLV) function start differentiation establishing the organ founder cell population. The RAM cell poll surrounds the quiescent center (QC, green). This regulatory model highlights the complex interplay of phytohormones and TFs in the WOX domain (green). Arrows indicate activation and barred lines indicate inhibition. The dotted arrows indicate the direction of auxin (AUX) and cytokinin (CK) flow. AUX produced in the SAM and young leaves is basipetal transported through the stem by the polar auxin transport (PAT) stream toward the RAM. CK biosynthesis genes are expressed in the RAM differentiation zone and acropetally transported. CZ, central zone; PZ, peripheral zone; LP lateral primordium [29,30].
Figure 1.
Diagram of SAM and RAM in Arabidopsis thaliana. In the SAM, a stem cell pool is located above the organizing center (OC, purple), which expresses the WUSCHEL transcription factor (TF) (yellow). Cells that pass the boundary defined by CLAVATA (CLV) function start differentiation establishing the organ founder cell population. The RAM cell poll surrounds the quiescent center (QC, green). This regulatory model highlights the complex interplay of phytohormones and TFs in the WOX domain (green). Arrows indicate activation and barred lines indicate inhibition. The dotted arrows indicate the direction of auxin (AUX) and cytokinin (CK) flow. AUX produced in the SAM and young leaves is basipetal transported through the stem by the polar auxin transport (PAT) stream toward the RAM. CK biosynthesis genes are expressed in the RAM differentiation zone and acropetally transported. CZ, central zone; PZ, peripheral zone; LP lateral primordium [29,30].
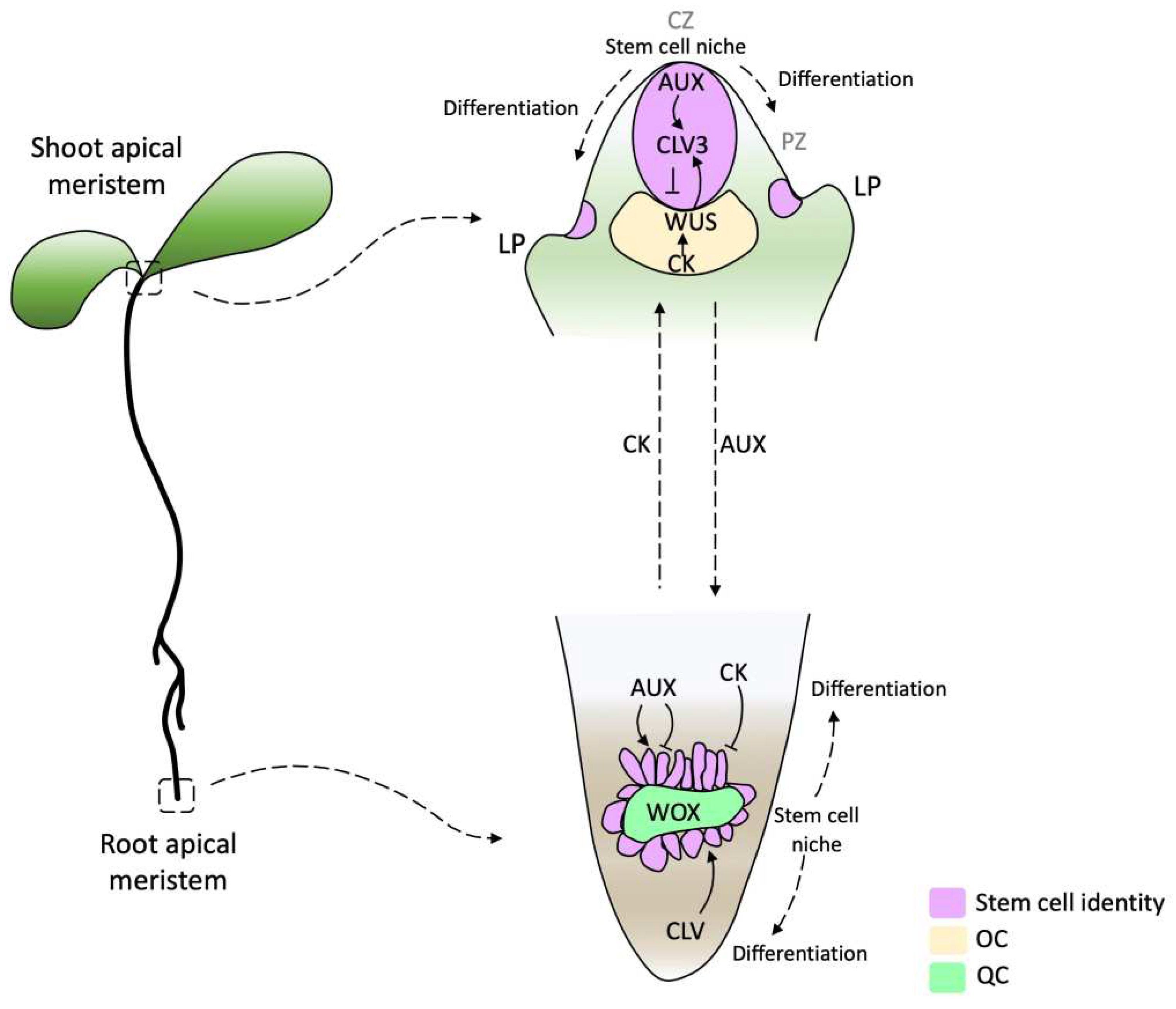
Figure 2.
Phylogeny of streptophyte and schematic location of shoot apical meristem (SAM) through gametophytic and sporophytic life states. The pink arrows point to the position of the SAM, while blue arrows indicate a close-up of specific structures. Within the non-vascular plant (bryophytes) and charophytes, the gametophyte is the dominant life state with vegetative growth. While in vascular plants, vegetative growth occurs in a sporophyte state. The gametophytes of lycophytes and spermatophytes lack meristems, and the sporophytic meristem in charophytes is unknown.
Figure 2.
Phylogeny of streptophyte and schematic location of shoot apical meristem (SAM) through gametophytic and sporophytic life states. The pink arrows point to the position of the SAM, while blue arrows indicate a close-up of specific structures. Within the non-vascular plant (bryophytes) and charophytes, the gametophyte is the dominant life state with vegetative growth. While in vascular plants, vegetative growth occurs in a sporophyte state. The gametophytes of lycophytes and spermatophytes lack meristems, and the sporophytic meristem in charophytes is unknown.
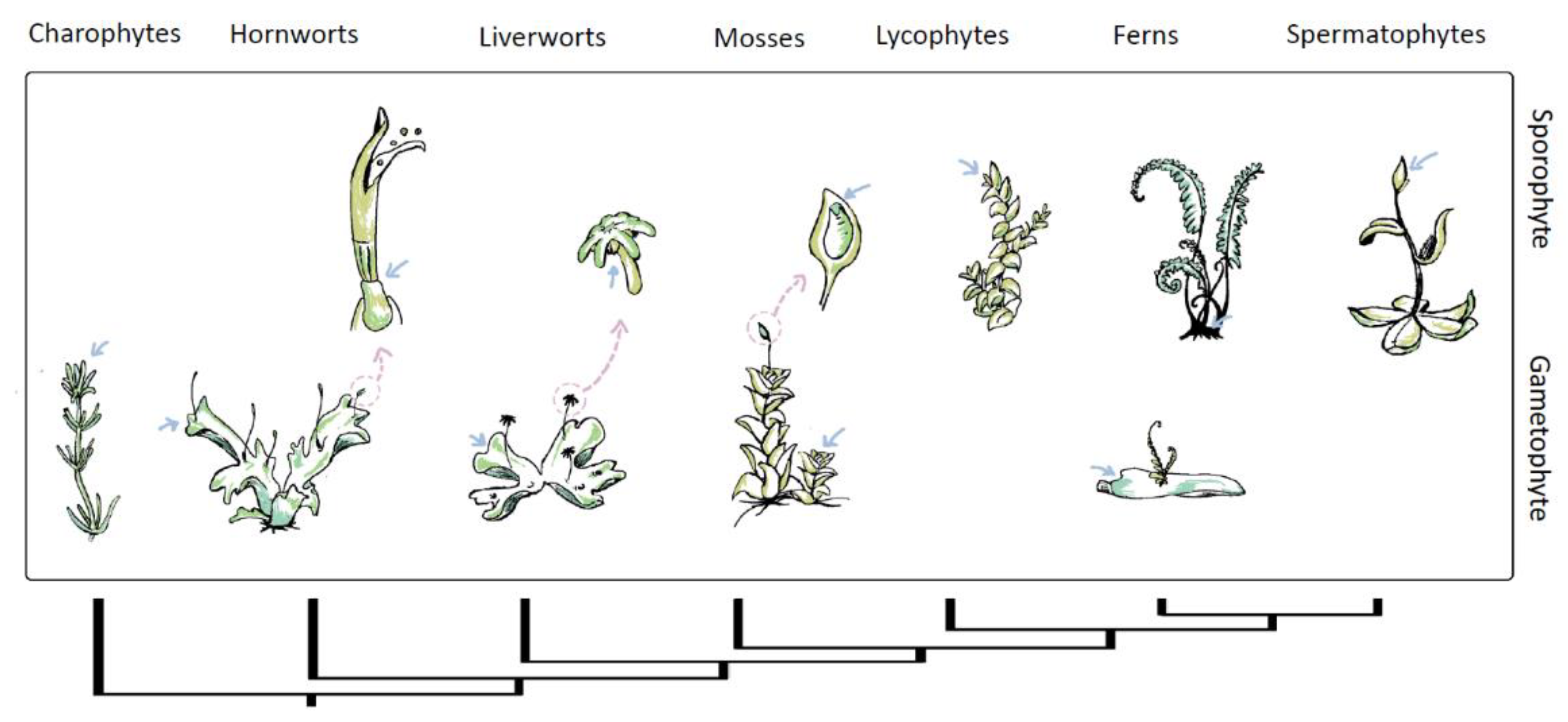
Figure 3.
Schematic representation of the organization of different types of shoot apical meristems (SAMs) from various taxonomic groups. (a) Represents the shapes of meristems and its division. From top to bottom: cross-sections of duplex and simplex meristems where divisions of the initial cells can occur anticlinal to provide length or periclinal to increase breadth. Meristems with apical cell, Tetrahedral apical cell with three cutting sides, three-sided cell with two cutting sides and dome-shaped apical cell with one cutting side. Dashed lines represent division planes, and arrows represent the directions in which the cell's progeny is displaced during growth. Blue cells represent stem cells. (b) Schematic representation of streptophyte phylogeny of the anatomy on SAM, through gametophyte and sporophyte life states. Within the charophycean algae, the apical meristem is a single cell with one cutting face [47,48]. Within bryophytes, unicellular apical meristems with one, two, three, and four cutting sides occur, as well as simple multicellular meristems consisting of two cells with eight cutting sides [45,48]. Pteridophytes and lycophytes, exhibit unicellular apical meristems with two or three sides persist in addition to simple and more complex multicellular meristems [65,136,137,138]. Large multicellular meristems occur within the seed plants. However, this figure shows only some of the common SAM shapes described, while multiple shapes can occur between different species [65,75,137]. Panel (b) adapted from Niklas and colleagues [49].
Figure 3.
Schematic representation of the organization of different types of shoot apical meristems (SAMs) from various taxonomic groups. (a) Represents the shapes of meristems and its division. From top to bottom: cross-sections of duplex and simplex meristems where divisions of the initial cells can occur anticlinal to provide length or periclinal to increase breadth. Meristems with apical cell, Tetrahedral apical cell with three cutting sides, three-sided cell with two cutting sides and dome-shaped apical cell with one cutting side. Dashed lines represent division planes, and arrows represent the directions in which the cell's progeny is displaced during growth. Blue cells represent stem cells. (b) Schematic representation of streptophyte phylogeny of the anatomy on SAM, through gametophyte and sporophyte life states. Within the charophycean algae, the apical meristem is a single cell with one cutting face [47,48]. Within bryophytes, unicellular apical meristems with one, two, three, and four cutting sides occur, as well as simple multicellular meristems consisting of two cells with eight cutting sides [45,48]. Pteridophytes and lycophytes, exhibit unicellular apical meristems with two or three sides persist in addition to simple and more complex multicellular meristems [65,136,137,138]. Large multicellular meristems occur within the seed plants. However, this figure shows only some of the common SAM shapes described, while multiple shapes can occur between different species [65,75,137]. Panel (b) adapted from Niklas and colleagues [49].
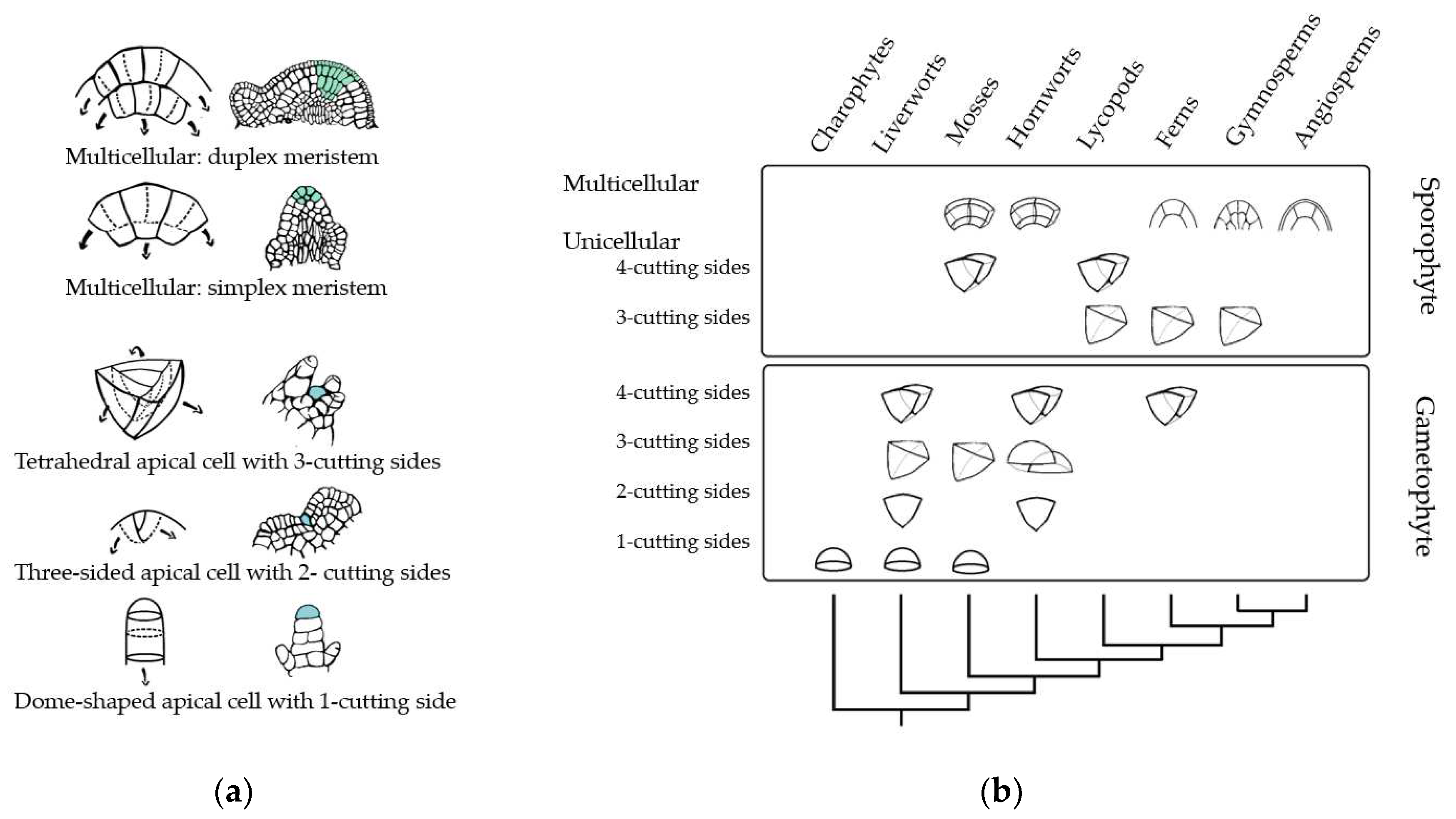
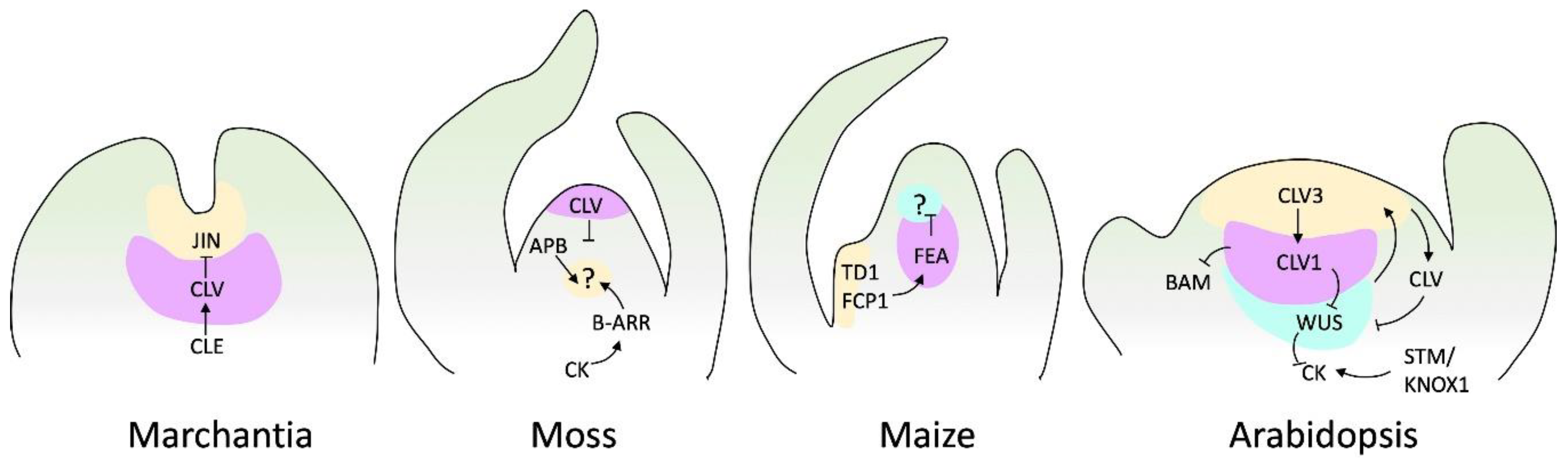
Figure 4.
SAM signaling pathways of various species. Genes and other factors as cytokinin (CK) with characterized genetic and biochemical interactions, are shown. Arrows depict positive regulation, and barred lines depict negative regulation. Question marks denote unidentified receptors. Marchantia does not have a well-described loop to maintain meristem regulation. However, it has been reported that JINGASA (JIN) acts downstream of CLV3/ESR-related (CLE) peptide signaling and controls stem cell behavior in the gametophyte [140]. Moss diagram adapted from Hata and Kyozuka [32], and maize and Arabidopsis diagrams adapted from Fletcher [141].
Figure 4.
SAM signaling pathways of various species. Genes and other factors as cytokinin (CK) with characterized genetic and biochemical interactions, are shown. Arrows depict positive regulation, and barred lines depict negative regulation. Question marks denote unidentified receptors. Marchantia does not have a well-described loop to maintain meristem regulation. However, it has been reported that JINGASA (JIN) acts downstream of CLV3/ESR-related (CLE) peptide signaling and controls stem cell behavior in the gametophyte [140]. Moss diagram adapted from Hata and Kyozuka [32], and maize and Arabidopsis diagrams adapted from Fletcher [141].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



