
Submitted:
12 September 2023
Posted:
13 September 2023
You are already at the latest version
Abstract
Auckland Island pigs represent an inbred population of feral pigs isolated on the subantarctic island for over 100 years. The animals have been maintained under pathogen-free conditions in New Zealand, they are well characterized virologically and have been used as donor sources in first clinical trials of porcine neonatal islet cell transplantation for the treatment of human dia-betes patients. The animals do not carry any of the xenotransplantation-relevant viruses and in the first clinical trials no porcine viruses including porcine endogenous retroviruses (PERVs) have been transmitted to the human recipients. PERVs pose a special risk in xenotransplantation since they are part of the pig genome. When the copy number of PERVs in these animals was analyzed using droplet digital PCR, a copy number typical for Western pigs was found, confirming previous phylogenetic analyses of microsatellites and mitochondrial analyses showing a closer relationship to European pigs than to Chinese pigs. When kidney cells from very young piglets were analyzed, only around 20 PERVpol copies were detected. Using these cells as nuclear donors in a somatic cell nuclear transfer (SCNT), animals were born showing PERVpol copy numbers between 35 and 56. This data indicates that Auckland Island pigs have a lower copy number in comparison with other Western pig breeds and that the copy number is higher in adult animals compared with cells from young piglets. Most importantly, PERV-C free animals were selected and the absence of additional eight porcine viruses was demonstrated.
Keywords:
porcine endogenous retroviruses
; Auckland Island pigs
; islet cell xenotransplantation
1. Introduction
The Auckland Islands are a group of subantarctic islands which lie some 560 kilometers south of New Zealand. Pigs were originally introduced onto these islands in 1807 by Captain Abraham Bristow and in 1840 James Ross brought more pigs onto the island [1, 2]. From the time of the last release the pig population on Auckland Island remained isolated for the next hundred years. Pigs were removed from Auckland Island in 1999 to help restore the environment to its former state and were brought to the mainland of New Zealand for conservation. 17 animals were housed, initially in quarantine, in a special, purpose-built facility in Invercargill, where they have been successfully bred [3]. The pigs are black or white to brown with black markings, have a long snout, high hair and bristle coverage for the cold conditions [1]. When the mitochondrial D-loop DNA sequences of the Auckland Island pigs were compared with sequences from domestic and wild boar, the sequence from the Auckland Island pigs clustered with domestic European breeds and they represent a single breeding population [1, 2]. Phylogenetic analyses of microsatellites also showed that they are more closely related to European pigs than Chinese pigs, which is consistent with the mitochondrial analyses [3].
These pigs have been monitored for 8 years and were found negative for all tested porcine infectious agents [4, 5]. Among the tested viruses potentially posing a risk for xenotransplantation were the porcine endogenous retro-virus (PERV), porcine cytomegalovirus (PCMV), a porcine roseolovirus (PCMV/PRV), porcine lymphotropic herpesvirus (PLHV), and porcine circovirus (PCV), for testing diagnostic PCR and serology methods were applied. In comparison with other pig breeds, the Auckland Island pigs are the “cleanest” pigs concerning virological safety. This was the reason to use pig islets from Auckland Island pigs for the treatment of humans with diabetes. In a prospective preclinical trial transplanting islet cells from Auckland Island pigs into non-human primates, no transmission of pig viruses was observed [6]. No transmission of pig viruses including PERVs was also observed in two clinical trials performed in New Zealand [7] and Argentina [8].
Here we analyze the number of PERV proviruses in the genome of adult Auckland Island pigs, in cells from young piglets as well as in animals obtained by somatic cell nuclear transfer (SCNT) using these piglet cells. Furthermore, we screened these animals for an additional 8 xenotransplantation-relevant viruses, among them PCMV/PRV. PCMV/PRV is of special interest, it is known to significantly reduce the survival time of non-human primates after transplantation of PCMV/PRV-positive kidneys or hearts [9-12]. PCMV/PRV was also transmitted in the first clinical trial transplanting a pig heart into a patient in Baltimore and contributed to the death of the patient [13, 14]. We confirmed the absence of PCMV/PRV and other potentially zoonotic viruses in the Auckland Island pigs.
It is known that PERV-C can recombine with PERV-A in living pigs and the resulting PERV-A/C recombinant viruses are characterized by higher virus titers compared with the paternal PERV-A [15-17]. Release of human tropic PERV-A/C from mitogen-triggered peripheral mononuclear cells (PBMCs) was observed in the case of different minipigs as well as ill pigs [18-22]. Since PERV-C is in contrast to PERV-A and PERV-B not present in all pigs, we screened the Auckland Island pigs for PERV-C and selected PERV-C animals for further breeding. The absence of PERV-C will prevent a recombination with PERV-A.
2. Materials and Methods
2.1. Animals and Cells
Auckland Island pigs were bred as described by the NZ Xeno Ltd, Auckland, New Zealand [4, 5]. DNA and blood from two groups of animals were sent to Berlin and the PERV copy numbers were analyzed. Later, kidney cells from four animals (two male, two female) at an age of about 4 weeks were sent to Germany and the PERV copy numbers were analyzed. Nuclei from these cells were used for an in vitro SCNT into matured oocytes which were collected from abattoir derived ovaries from German Landrace/Piétrain hybrid gilts. The SCNT was performed at the Chair for Molecular Animal Breeding and Biotechnology, and Center for Innovative Medical Models (CiMM) of the Ludwig Maximilian University Munich. DNA was obtained from animals of the first and second generation and analyzed for the PERV copy number. As an internal standard the porcine embryonic kidney cell line PK15 (ACC 640, from the Leibniz Institute DSMZ German Collection of Microorganisms and Cell lines, Braunschweig, Germany) was used.
2.2. DNA Isolation
DNA was extracted at XenoNZ and CiMM, DNA was isolated from blood using two DNA extraction methods: DNeasy Blood and Tissue kit (Qiagen GmbH, Hilden, Germany) or phenol chloroform extraction using TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer′s instructions. DNA was quantified using a Qubit 3.0 Fluorometer (Thermo Fischer), and the 260 nm/280 nm ratio was determined using a NanoDrop ND-1000 (Thermo Fisher Scientific Inc., Worcester, MA, USA).
2.3. Droplet Digital PCR
Droplet digital PCR (ddPCR) was performed according to the manufacturer's instructions (Bio-Rad, Hercules, CA, USA, [http://www.bio-rad.com/de-de/applications-technologies/droplet-digital-pcr-ddpcr-technology?ID=MDV31M4VY]) using a QX200 droplet generator and a QX100 droplet reader (Bio-Rad). Purified genomic DNA (100 ng genomic DNA) was digested with MseI (New England Biolabs, Ipswich, MA, USA) (20U) at 37°C for 1 hour, and the restriction enzyme was heat-inactivated. The DNA digest was diluted to 5-10 ng/μL for the ddPCR. The ddPCR mix consisted of 10 μL 2X ddPCR Master mix, 1.8 μL of each 10 μmol/L target primers (Table 1), 0.5 μL of each 10 μmol/L probes (FAM/HEX) (Table 1), 2.5-10 ng digested DNA, and water to a total volume of 20 μL. The following cycling conditions were used: 10-minute initial enzyme activation at 95°C, 30-second denaturation at 94°C, 1-min annealing.
2.3. Spleen and Liver Tissues
Frozen samples of spleen and liver tissues from dead born (day 0), 9-30 days old and older than 12 weeks piglets from the CiMM wer used. DNA was isolated as described above and the PERV copy number was measured in both organs. and extension at 59°C (45 cycles), and final 10-min enzyme deactivation at 98°C by using a Master cycler ProS (Eppendorf, Hamburg, Germany). The temperature ramp rate was 2°C per second. To run a gradient, temperatures between 57 and 62°C were used.
2.4. Tests for PERV-C
DNA from the Auckland Island pigs were used to analyze the presence of PERV-C. Primers and conditions used for were described [25, 26] (Table 2).
2.5. Tests for Additional Porcine Viruses
DNA, RNA and blood from the Auckland Island pigs were used to analyze the presence of different DNA viruses and the RNA virus HEV. DNA was isolated as described above, and RNA was isolated using the RNeasy Mini kit (Qiagen, Hilden, Germany). Primers and condition used for the PCR or real-time PCR assays have been described for PCMV/PRV [27, 28], PCV2 [29], PCV3 [30], PLHV-1, PLHV-2, and PLHV-3 [31] as well as for HEV [31]. The Western blot and ELISA to detetct antibodies against HEV using recombinant proteins were described as well [31].
3. Results
3.1. PERV Copy Number in the Genome of Adult Auckland Island Pigs
To determine the number of integrated PERVs, a ddPCR was developed which uses primers binding to a highly conserved region of the polymerase gene (pol) of PERV [24, 32]. DNA from 30 Auckland Island pigs, bred in New Zealand, were analyzed using the ddPCR and a PERVpol copy number of around 60 was measured (Figure 1). This copy number was similar compared with the copy number of Göttingen minipigs and lower compared with Aachen minipigs, investigated in parallel. DNA from kidneys of Göttingen minipigs revealed between 45 and 78 PERVpol copies using GAPDH as a reference and in a second study analyzing 16 Göttingen minipigs a median number of 64 (45-93) was found [32]. When DNA samples from Aachen minipigs were analyzed, a median number of 69 (34-97) was determined [32].
3.2. Selection of PERV-C Negative Animals
When the 14 animals tested for the PERV copy number above (Figure 2) and another pig (494) were tested using a PCR, eight out of 15 animals (53%) were PERV-C negative (Table 2). These investigations had the goal to select four PERV-C-free animals from which kidney cells were obtained, which could later be used for the SCNT.
Figure 2.
Determination of the PERVpol copy number of 14 further Auckland Island pigs bred in New Zealand by ddPCR at a later time point.
Figure 2.
Determination of the PERVpol copy number of 14 further Auckland Island pigs bred in New Zealand by ddPCR at a later time point.
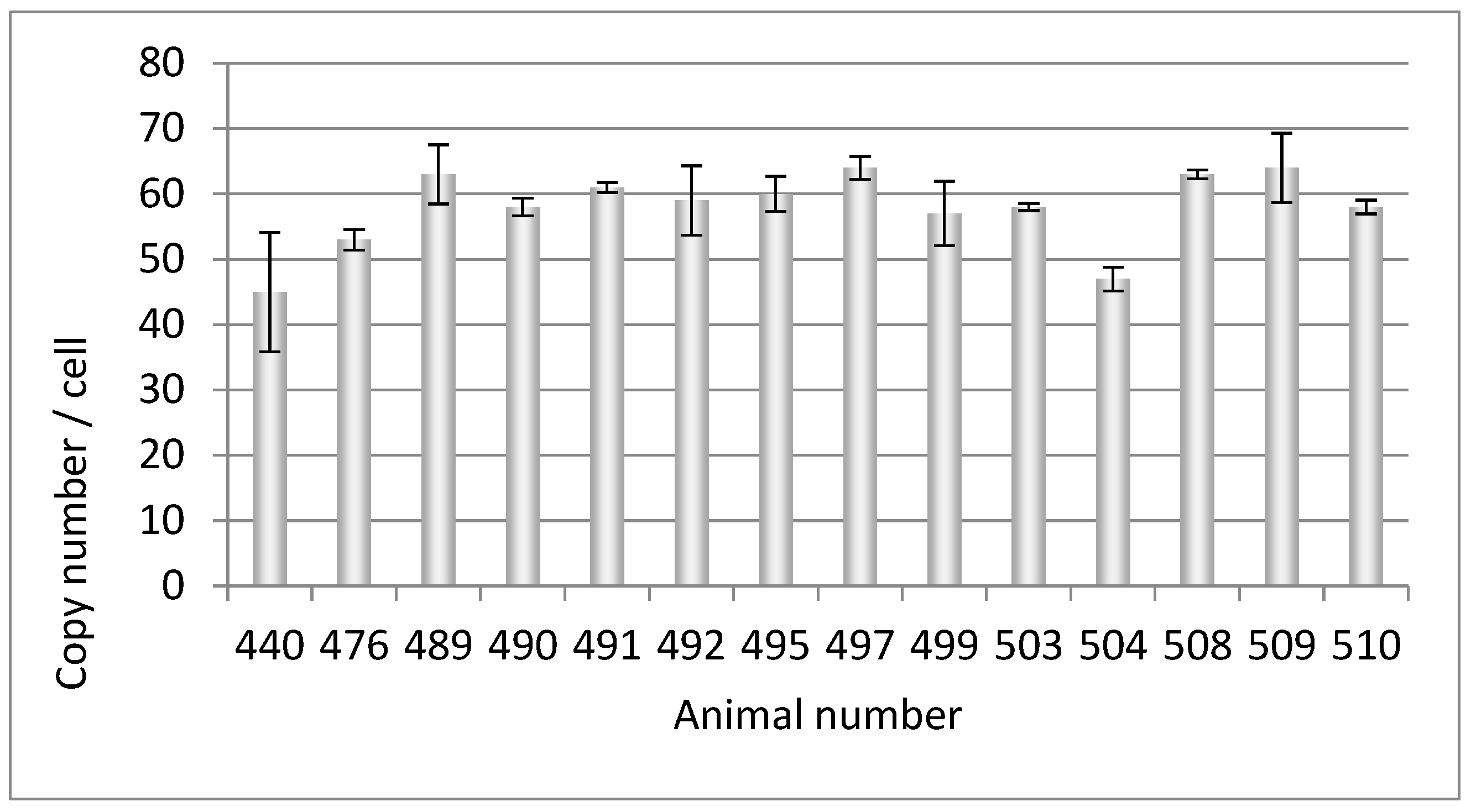

Table 2.
Testing of Auckland Island pigs (animals 440-510) and embryonic cells (w, female, m, male) for different viruses using PCR methods and a Western blot assay and ELISA (HEV). PERV, median copy number (Figure 1); n..t., not tested, n.a., not applicable.
Table 2.
Testing of Auckland Island pigs (animals 440-510) and embryonic cells (w, female, m, male) for different viruses using PCR methods and a Western blot assay and ELISA (HEV). PERV, median copy number (Figure 1); n..t., not tested, n.a., not applicable.
| Pig | PERV copy number |
PERV-C | PCMV | HEV | HEV | HEV | PCV1/2 | PCV3 | PLHV-1/2 | PLHV-3 |
|---|---|---|---|---|---|---|---|---|---|---|
| PCR | PCR | PCR | WB | ELISA | PCR | PCR | PCR | PCR | ||
| 440 | 69 | - | - | - | - | - | - | - | - | - |
| 476 | 64 | - | - | - | - | n.t. | - | - | - | + |
| 489 | 68 | + | - | - | - | - | - | - | - | - |
| 490 | 75 | + | - | - | - | n.t. | - | - | - | - |
| 491 | 68 | + | - | - | - | - | - | - | - | - |
| 492 | 62 | + | - | - | - | n.t. | - | - | - | - |
| 494 | 73 | + | - | - | - | - | - | - | - | - |
| 495 | 66 | - | - | - | - | - | - | - | - | - |
| 497 | 69 | - | - | - | - | - | - | - | - | - |
| 499 | 62 | - | - | - | - | - | - | - | - | - |
| 503 | 62 | - | - | - | - | - | - | - | - | - |
| 504 | 57 | - | - | - | - | n.t. | - | - | - | - |
| 508 | 64 | + | - | - | - | n.t. | - | - | - | - |
| 509 | 67 | + | - | - | - | n.t. | - | - | - | - |
| 510 | 60 | + | - | - | - | n.t. | - | - | - | |
| f1 | 21 | - | - | n.t. | n.a. | n.a. | n.t. | - | n.t. | n.t. |
| f2 | 20 | - | - | n.t. | n.a. | n.a. | n.t. | - | n.t. | n.t. |
| m1 | 20 | - | - | n.t. | n.a. | n.a. | n.t. | - | n.t. | n.t. |
| m2 | 22 | - | - | n.t. | n.a. | n.a. | n.t. | - | n.t. | n.t. |
3.3. PERV Copy Number in the Genome of Cell Lines from Very Young Piglets
DNA from four kidney cell lines from very young pigletswere analyzed which had been selected as PERV-C negative. A very low copy number of 20-22 copies was found (Figure 3).
3.4. PERV Copy Number in the Genome of Auckland Island Pigs Obtained by SCNT
The nuclei of these PERV-C-negative embryonic kidney cells were used for an in vitro SCNT into matured oocytes collected from abattoir derived ovaries from German Landrace/Piétrain hybrid gilts. The animals of the first and second generation were analyzed for the PERVpol copy number. The copy numbers of the individual pigs ranged from 32 to 57, but were lower compared with the numbers of the Auckland Island pigs analyzed in the past (Figure 1, Figure 2), and lower than other pig breeds such as the Göttingen minipigs, and the Aachen minipigs [32]. The copy number found in PK15 cells correlated well with the copy number detected previously. Since different copy numbers of integrated PERV were found in different PK15 cell lines [32], here the PK15 cells ordered from the Leibniz Institute DSMZ German Collection of Microorganisms and Cell lines, Braunschweig, Germany (ACC 640), were used and the measured copy number was 32.12 +/- 1.7, which was in agreement with our previous findings [24, 32].
3.5. Increase of the PERV Copy Number with Age
In order to obtain additional evidence for the increase of the PERV copy number with age, the copy number was determined in two different organs of dead born (day 0), 9-30 days old and older than 12 weeks piglets from the CiMM. The copy number was determined in the spleen and liver and the difference between the copy number was depicted dependend on the age of the animals (Figure 4). It was obvious that there were differences in the copy number between spleen and liver, indicating that in addition to the copy number in the germ line de novo integrations took place in one or the other organ. The de novo integrations was not tissue-specific, in some animals the higher copy number was in the spleen, in others in the liver. The difference was larger in older piglets, indicating that the copy number increased in dependence on the age of the animals (Figure 4).
3.6. Further Characterization of the Auckland Island Pigs
In order to characterize the Auckland Island pigs further, PCR-based methods for the detection of PERV-C, PCMV/PRV, PCV1, PCV2, PCV3, PLHV-1, PLHV-2, and PLHV-3 as well as well as a reverse transcriptase (RT) RT-PCR for HEV were performed (Table 2). The tests for all these viruses were negative. Negative was also a Western blot assay and an ELISA screening for antibodies against HEV, indicating that these animals are free of HEV. In addition to the Auckland Island pigs, the four kidney cell lines from very young piglets were tested for PERV-C, PCMV/PRV, PCV3 and were found negative (Table 2).
4. Discussion
For the first time, a determination of the number of integrated PERVs in Auckland Island pigs was performed using the ddPCR method. The Auckland Island pigs had been used in the first xenotransplantation trials in New Zealand and Argentina [7, 8], they are virologically well characterized [4-6] and are the “cleanest” pigs compared with other pig breeds.
In the past it had been shown that the PERV copy number differs in dependence on the pig strain, on the individual animal tested, and on the method used (for review see [33]). Auckland Island pigs had been reported previously to contain three to 37 copies using a real-time PCR assay and a light cycler [5] and four to 40 copies were found using a PCR-based limited dilution assay (PLDA) [7]. In another study, relative gene dosages of PERV in Auckland Island pigs ranged from 2.8 ± 0.1 to 68 ± 1.6 at an average of 17.6 ± 10.4 using a real-time PCR [34]. This indicates a generally lower copy number compared to the numbers found here, but some animals had even a higher copy number (68). In our study we found that the PERV copy number is much lower in cells from very young piglets used for the SCNT (20-22) compared with the copy number in adult animals (ranging from 32 to 59 in the animals obtained from these cells by SCNT) (Figure 3).
This data confirms that the copy number depends on the method used and probably on the age of the animals. It is well known that PERVs are active and in addition to the endogenous proviruses there are do novo integrated proviruses in somatic cells. In the result, the number of integrated PERV proviruses differs from organ to organ (for review of numerous publications see [33]) and even in different parts of a single organ [24] in living pigs. The finding that the copy number is lower in cells from very young animals correlates with findings in another study: When analyzing 14-21-day-old neonatal and 1-2-year-old adult Belgian landrace pigs a significant increase in the copy number over time was also observed [35]. In these animals, an difference between least five copies and 80 copies was reported, indicating that the viruses are active and replicating in the neonatal pig. In Figure 5 a schematic presentation of these events is given. Indicated are the copy numbers found in the somatic cells of the animals and in the cells from young animals used for SCNT measured by ddPCR. Considering that copy numbers in adult animals are the result of de novo integrations in somatic cells, it logically follows that the number of proviruses in the germline can be at most 20 (Figure 5).
The fact that the number of integrated proviruses increased by age was also supported by a comparison of the copy number in two different tissues, spleen and liver, of other than Auckland Island pigs (Figure 4). Differences in the number of integrated copies means that in one or the other organs additional copies appeared in addition to the copy number in the germ line. The fact that the difference increases with age indicates that more new copies were integrated with age.
The differences in the copy number of PERV in Auckland Island pigs when measuring by different methods [5, 7, 34 and this paper] do not only depend on the method. Based on our findings that the copy number increases due to de novo integrations, it also depends on the age of the animal which was not known to us and therefore cannot be taken into account. It would be highly interesting to test the PERV copy number in organs of a single animal over time. Furthermore, it may also depend on the tissue and the amount of PBMCs present in this tissue. PBMCs are prone for release of different PERVs [36] and de novo integration evidenced by the presence of integrated and release of infectious recombinant PERV-A/C from PBMCs [18, 20, 21, 36]. PERV-A/C have never been found in the germ line of pigs [37].
Auckland Island pigs are descendants of Western pigs brought to the island [1, 2]. This was confirmed by phylogenetic analyses of microsatellites and mitochondrial analyses showing that they are more closely related to European pigs than Chinese pigs [3]. The PERV copy number in Western pigs is higher compared with Chinese pigs. By analyzing 63 high-quality porcine whole-genome resequencing data, it was found that the PERV copy numbers in Chinese pigs were lower (32.0±4.0) than in Western pigs (49.1±6.5) [38].
From all methods ever used to count PERV copies in the DNA of pigs, such as Southern blot [39, 40], PCR titration [41], real-time PCR [42-47], fluorescence in situ hybridization (FISH) [30], PCR-based limited dilution assay [5], and genome wide sequencing [31], the ddPCR seems to be the most accurate method [24, 32, 33]. For example, using Southern blot analysis about 50 PERV copies were found in certain pig breeds [39, 40], but PCR titration [41] or real-time PCR [42-47] detected from one to 98 copies. By using FISH, 19 copies of PERV-A and 13 copies of PERV-B were found in Westran pigs [48]. Using genome-wide sequencing, 20 gammaretrovirus copies and 4 betaretrovirus copies were detected in Duroc pigs [49], indicating that sequencing does not detect all integrated viruses.
DdPCR was found to have numerous favorable features in comparison with all other methods including real-time PCR (for review see [23, 24, 32, 33]). One major advantage is that reference standard curves are not required, and bias effects arising from amplification efficiency and PCR inhibitors were reduced. The ddPCR technique provides absolute quantification based on the principles of sample partitioning and Poisson statistics, thus overcoming the normalization and calibrator issues; it has shown increased precision and sensitivity; it is relatively insensitive to PCR inhibitors and directly provides the result of the analysis expressed as number of copies of target per microliter of reaction [50].
Using ddPCR, fifty-nine copies were found when analyzing kidney fibroblasts from multi-transgenic pigs expressing the human transgenes CD46, CD55, CD59, HO1, and A20 [32]. When wild boars in and around Berlin were analyzed using droplet digital PCR, three to 69 PERV copies were found. A lower copy number, but a higher variability of this number was found in wild boars compared to domestic pigs, including minipigs [24].
In contrast to human endogenous retroviruses (HERVs), which are mostly defective and not replication-competent, PERVs can be released from normal pig cells and at least PERV-A and -B are infectious for certain human cells. Human embryonic kidney 293 cells are the most susceptible cells, mainly because these cells lost numerous intracellular restriction factors [51]. Other human cell lines such as THP-1 and C8166 cells could be infected, but only with a very low virus production compared with the 293 cells [52]. Important is the fact that primary human cells such as endothelial cells, vascular fibroblast, and mesangial cells could be infected with PERV [52, 53], but also with a low virus production and primary human PBMCs could only be infected with a human-cell-adapted PERV, characterized by a higher number of enhancer repeats and a high titer [54].
All PCR-based methods used to estimate the PERV copy number have a common limitation, in that they used primers mostly for the highly conserved pol gene. This allows to detect all PERVs, i.e. PERV-A, PERV-B and PERV-C. However, the detection of this sequence does not mean that there is an intact provirus able to produce infectious virus particles. Therefore, the estimated number of integrated proviruses measured with PCR-based methods tells us nothing concerning the safety of xenotransplantation, e.g., the risk posed by PERVs [55]. Only sequencing the pig genome could give more reliable information. However, sequencing of repetitive, nearly identical sequences is always associated with problems as has been seen with hidden sequences of the human endogenous retrovirus HERV-K [56, 57]. In these investigations, in addition to 91 full-length HERV-K proviruses previously found in the human genome draft sequence hundreds more proviruses were detected. Sequencing the genome of a domestic Duroc pig, 212 endogenous retrovirus sequences were detected, among them 9 PERV-A, 11 PERV-B and 4 betaretrovirus copies [48], a lower number of PERVs compared with other methods [33].
One major result of the work was the elimination of PERV-C. Among the Auckland Island pigs analyzed in the first experiment approximately 53% of the animals were PERV-C-negative (Table 2). PERV-C is an ecotropic virus, infecting only pig cells and therefore does not pose a direct risk for xenotransplantation. However, recombinants between the human-tropic PERV-A and PERV-C have been observed, which are able to infect human cells and which are characterized by higher replication rates [58-60]. At present PERV-A/C were found mainly in minipigs, either released from their PBMCs [37, 60-62] or present in some organs of minipigs with melanomas [63] or even apparently healthy pigs [64]. One exception was the finding of PERV-A/C in farm animals with infectious diseases [22]. However, PERV-A/C was only found in somatic cells, never in the germ line [21, 65-67].
Compared with PERV-A and -B, PERV-C appears to be even younger in the pig lineage as not all domestic pigs have PERV-C sequence [39]. Miniature swine may be a unique pig breed as they possess more copies of PERV-C sequence (typically five to nine copies) than many other pig herds (one to three copies [40]), suggesting that PERV-C is active in miniature swine [19]. Although only limited studies have been performed on PERV transmission to human cells using primary cultures from other pig herds, it is intriguing that A/C recombinant PERV have been reported mainly from miniature swine but not from other herds [19]. However, there is no risk of PERV-A/C release from Auckland Island pigs generated by SCNT using nuclei from PERV-C-free Auckland Island pigs.
5. Conclusions
Auckland Island pigs had been used repeatedly as donors for pig islet cell transplantation into non-human primates and diabetic patients. In all preclinical and clinical trials no porcine viruses including PERV had been transmitted.
Auckland Island pigs are well characterized, they are free of many potentially pathogenic viruses and have a low PERV copy number in comparison with other pig breeds. The copy number is much lower in cells from very young piglets and in animals obtained by SCNT using nuclei from Auckland Island pig cells. To enhance the virus safety, PERV-C negative animals were selected.
Author Contributions
Conceptualization, J.D..; methodology, U.F. and L.K., validation, U.F., L. K. and J.D.; writing—original draft preparation, J.D.; writing—review and editing, U.F. and J.D.; supervision, J.D.; funding acquisition, J.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Deutsche Forschungsgemeinschaft, TR127.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data supporting reported results are included in this manuscript.
Acknowledgments
We thank Olga Garkavenko and Paul Tan, NZ Xeno Ltd, Auckland, for supplying the DNA and the blood from the New Zealand Auckland Island pigs and for many helpful discussions leading not only to the completion of this manuscript, but also of reference 55. We thank Eckhard Wolf and Barbara Keßler, Chair for Molecular Animal Breeding and Biotechnology and CiMM, Munich, for the DNA of the cells from the very young piglets and the DNA from the Munich Auckland Island pigs as well as for valuable discussions. We thank Bruno Reichart for constant support and advice.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gongora J, Garkavenko, O, Moran C. Origins of Kune Kune and Auckland Island pigs in New Zealand. 7th World Congress on Genetic Applied to Livestock Production, August 19-23, 2002, Montpellier, France. 19 August 2002; -23.
- Robins, J.H.; Matisoo-Smith, E.; Ross, H.A. The origins of the feral pigs on the Auckland Islands. J. Royal Society of New Zealand 2003, 33, 561–569. [Google Scholar] [CrossRef]
- Fan, B.; Gongora, J.; Chen, Y.; Garkavenko, O.; Li, K.; Moran, C. Population genetic variability and origin of Auckland Island feral pigs. J. R. Soc. N. Z. 2005, 35, 279–285. [Google Scholar] [CrossRef]
- Garkavenko, O.; Muzina, M.; Muzina, Z.; Powels, K.; Elliott, R.B.; Croxson, M.C. Monitoring for potentially xenozoonotic viruses in New Zealand pigs. J. Med Virol. 2003, 72, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Garkavenko, O.; Wynyard, S.; Nathu, D.; Simond, D.; Muzina, M.; Muzina, Z.; Scobie, L.; Hector, R.D.; Croxson, M.C.; Tan, P.; et al. Porcine Endogenous Retrovirus (PERV) and its Transmission Characteristics: A Study of the New Zealand Designated Pathogen-Free Herd. Cell Transplant. 2008, 17, 1381–1388. [Google Scholar] [CrossRef]
- Garkavenko, O.; Dieckhoff, B.; Wynyard, S.; Denner, J.; Elliott, R.B.; Tan, P.L.; Croxson, M.C. Absence of transmission of potentially xenotic viruses in a prospective pig to primate islet xenotransplantation study. J. Med Virol. 2008, 80, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Wynyard, S.; Nathu, D.; Garkavenko, O.; Denner, J.; Elliott, R. Microbiological safety of the first clinical pig islet xenotransplantation trial in New Zealand. Xenotransplantation 2014, 21, 309–323. [Google Scholar] [CrossRef]
- Morozov, V.A.; Wynyard, S.; Matsumoto, S.; Abalovich, A.; Denner, J.; Elliott, R. No PERV transmission during a clinical trial of pig islet cell transplantation. Virus Res. 2017, 227, 34–40. [Google Scholar] [CrossRef]
- Denner, J.; Längin, M.; Reichart, B.; Krüger, L.; Fiebig, U.; Mokelke, M.; Radan, J.; Mayr, T.; Milusev, A.; Luther, F.; et al. Impact of porcine cytomegalovirus on long-term orthotopic cardiac xenotransplant survival. Sci. Rep. 2020, 10, 17531. [Google Scholar] [CrossRef]
- Yamada, K.; Tasaki, M.; Sekijima, M.; Wilkinson, R.A.; Villani, V.; Moran, S.G.; Cormack, T.A.; Hanekamp, I.M.; Arn, J.S.; Fishman, J.A.; et al. Porcine Cytomegalovirus Infection Is Associated With Early Rejection of Kidney Grafts in a Pig to Baboon Xenotransplantation Model. Transplantation 2014, 98, 411–418. [Google Scholar] [CrossRef]
- Sekijima, M.; Waki, S.; Sahara, H.; Tasaki, M.; Wilkinson, R.A.; Villani, V.; Shimatsu, Y.; Nakano, K.; Matsunari, H.; Nagashima, H.; et al. Results of Life-Supporting Galactosyltransferase Knockout Kidneys in Cynomolgus Monkeys Using Two Different Sources of Galactosyltransferase Knockout Swine. Transplantation 2014, 98, 419–426. [Google Scholar] [CrossRef]
- Denner, J. Reduction of the survival time of pig xenotransplants by porcine cytomegalovirus. Virol. J. 2018, 15, 171. [Google Scholar] [CrossRef] [PubMed]
- Griffith, B.P.; Goerlich, C.E.; Singh, A.K.; Rothblatt, M.; Lau, C.L.; Shah, A.; Lorber, M.; Grazioli, A.; Saharia, K.K.; Hong, S.N.; et al. Genetically Modified Porcine-to-Human Cardiac Xenotransplantation. New Engl. J. Med. 2022, 387, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, M.M.; Singh, A.K.; Scobie, L.; E Goerlich, C.; Grazioli, A.; Saharia, K.; Crossan, C.; Burke, A.; Drachenberg, C.; Oguz, C.; et al. Graft dysfunction in compassionate use of genetically engineered pig-to-human cardiac xenotransplantation: a case report. Lancet 2023, 402, 397–410. [Google Scholar] [CrossRef]
- Denner, J.; Specke, V.; Thiesen, U.; Karlas, A.; Kurth, R. Genetic alterations of the long terminal repeat of an ecotropic porcine endogenous retrovirus during passage in human cells. Virology 2003, 314, 125–133. [Google Scholar] [CrossRef]
- Karlas, A.; Irgang, M.; Votteler, J.; Specke, V.; Ozel, M.; Kurth, R.; Denner, J. Characterisation of a human cell-adapted porcine endogenous retrovirus PERV-A/C. Ann Transplant. 2010, 15, 45–54. [Google Scholar] [PubMed]
- Harrison, I.; Takeuchi, Y.; Bartosch, B.; Stoye, J.P. Determinants of High Titer in Recombinant Porcine Endogenous Retroviruses. J. Virol. 2004, 78, 13871–13879. [Google Scholar] [CrossRef]
- Wilson, C.A.; Wong, S.; Muller, J.; Davidson, C.E.; Rose, T.M.; Burd, P. Type C Retrovirus Released from Porcine Primary Peripheral Blood Mononuclear Cells Infects Human Cells. J. Virol. 1998, 72, 3082–3087. [Google Scholar] [CrossRef]
- Denner, J.; Schuurmann, K.J. High prevalence of recombinant porcine endogenous retroviruses (PERV-A/Cs) in minipigs: a review on origin and presence. Viruses 2021, 13, 1869. [Google Scholar] [CrossRef]
- Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs. Viruses 2019, 12, 38. [Google Scholar] [CrossRef]
- Halecker, S.; Krabben, L.; Kristiansen, Y.; Krüger, L.; Möller, L.; Becher, D.; Laue, M.; Kaufer, B.; Reimer, C.; Denner, J. Rare isolation of human-tropic recombinant porcine endogenous retroviruses PERV-A/C from Göttingen minipigs. Virol J. 2022, 19, 30. [Google Scholar] [CrossRef]
- Pal, N.; Baker, R.; Schalk, S.; Scobie, L.; Tucker, A.W.; Opriessnig, T. Detection of Porcine Endogenous Retrovirus (PERV) Viremia in Diseased Versus Healthy US Pigs by Qualitative and Quantitative Real-Time RT-PCR. Transbound. Emerg. Dis. 2011, 58, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Güell, M.; Niu, D.; George, H.; Lesha, E.; Grishin, D.; Aach, J.; Shrock, E.; Xu, W.; Poci, J.; et al. Genome-wide inactivation of porcine endogenous retroviruses (PERVs). Science 2015, 350, 1101–1104. [Google Scholar] [CrossRef]
- Krüger, L.; Stillfried, M.; Prinz, C.; Schröder, V.; Neubert, L.K.; Denner, J. Copy Number and Prevalence of Porcine Endogenous Retroviruses (PERVs) in German Wild Boars. Viruses 2020, 12, 419. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Patience, C.; Magre, S.; Weiss, R.A.; Banerjee, P.T.; Le Tissier, P.; Stoye, J.P. Host range and interference studies of three classes of pig endogenous retrovirus. J Virol 1998, 72, 9986–9991. [Google Scholar] [CrossRef] [PubMed]
- Kaulitz, D.; Mihica, D.; Adlhoch, C.; Semaan, M.; Denner, J. Improved pig donor screening including newly identified variants of porcine endogenous retrovirus-C (PERV-C). Arch. Virol. 2012, 158, 341–348. [Google Scholar] [CrossRef]
- Morozov, V.A.; Plotzki, E.; Rotem, A.; Barkai, U.; Denner, J. Extended microbiological characterization of Göttingen minipigs: porcine cytomegalovirus and other viruses. Xenotransplantation 2016, 23, 490–496. [Google Scholar] [CrossRef]
- Morozov, V.A.; Morozov, A.V.; Denner, J. New PCR diagnostic systems for the detection and quantification of porcine cytomegalovirus (PCMV). Arch. Virol. 2016, 161, 1159–1168. [Google Scholar] [CrossRef]
- Heinze, J.; Plotzki, E.; Denner, J. Virus Safety of Xenotransplantation: Prevalence of Porcine Cicrovirus 2 (PCV2) in Pigs. Ann. Virol. Res. 2016, 2, 1023. [Google Scholar]
- Prinz, C.; Stillfried, M.; Neubert, L.K.; Denner, J. Detection of PCV3 in German wild boars. Virol. J. 2019, 16, 25. [Google Scholar] [CrossRef]
- Morozov, V.A.; Morozov, A.V.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Extended Microbiological Characterization of Göttingen Minipigs in the Context of Xenotransplantation: Detection and Vertical Transmission of Hepatitis E Virus. PLOS ONE 2015, 10, e0139893. [Google Scholar] [CrossRef]
- Fiebig, U.; Fischer, K.; Bähr, A.; Runge, C.; Schnieke, A.; Wolf, E.; Denner, J. Porcine endogenous retroviruses: Quantification of the copy number in cell lines, pig breeds, and organs. Xenotransplantation 2018, 25, e12445. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. How Active Are Porcine Endogenous Retroviruses (PERVs)? Viruses 2016, 8, 215. [Google Scholar] [CrossRef] [PubMed]
- Wynyard, S.; Garkavenko, O.; Elliot, R. Multiplex high resolution melting assay for estimation of Porcine Endogenous Retrovirus (PERV) relative gene dosage in pigs and detection of PERV infection in xenograft recipients. J. Virol. Methods 2011, 175, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Mourad, N.I.; Crossan, C.; Cruikshank, V.; Scobie, L.; Gianello, P. Characterization of porcine endogenous retrovirus expression in neonatal and adult pig pancreatic islets. Xenotransplantation 2017, 24. [Google Scholar] [CrossRef] [PubMed]
- Tacke, S.J.; Specke, V.; Denner, J. Differences in Release and Determination of Subtype of Porcine Endogenous Retroviruses Produced by Stimulated Normal Pig Blood Cells. Intervirology 2003, 46, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Bartosch, B.; Stefanidis, D.; Myers, R.; Weiss, R.; Patience, C.; Takeuchi, Y. Evidence and Consequence of Porcine Endogenous Retrovirus Recombination. J. Virol. 2004, 78, 13880–13890. [Google Scholar] [CrossRef]
- Chen, J.-Q.; Zhang, M.-P.; Tong, X.-K.; Li, J.-Q.; Zhang, Z.; Huang, F.; Du, H.-P.; Zhou, M.; Ai, H.-S.; Huang, L.-S. Scan of the endogenous retrovirus sequences across the swine genome and survey of their copy number variation and sequence diversity among various Chinese and Western pig breeds. Zool. Res. 2022, 43, 423–441. [Google Scholar] [CrossRef]
- Le Tissier, P.; Stoye, J.P.; Takeuchi, Y.; Patience, C.; Weiss, R.A. Two sets of human-tropic pig retrovirus. Nature 1997, 389, 681–682. [Google Scholar] [CrossRef]
- Patience, C.; Takeuchi, Y.; Weiss, R.A. Infection of human cells by an endogenous retrovirus of pigs. Nat. Med. 1997, 3, 282–286. [Google Scholar] [CrossRef]
- Patience, C.; Switzer, W.M.; Takeuchi, Y.; Griffiths, D.J.; Goward, M.E.; Heneine, W.; Stoye, J.P.; Weiss, R.A. Multiple Groups of Novel Retroviral Genomes in Pigs and Related Species. J. Virol. 2001, 75, 2771–2775. [Google Scholar] [CrossRef]
- Liu, G.; Li, Z.; Pan, M.; Ge, M.; Wang, Y.; Gao, Y. Genetic Prevalence of Porcine Endogenous Retrovirus in Chinese Experimental Miniature Pigs. Transplant. Proc. 2011, 43, 2762–2769. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Lee, J.; Yoon, J.-K.; Kim, N.Y.; Kim, G.-W.; Park, C.; Oh, Y.-K.; Kim, Y.B. Rapid Determination of Perv Copy Number From Porcine Genomic DNA by Real-Time Polymerase Chain Reaction. Anim. Biotechnol. 2011, 22, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.; Choi, J.; Lee, H.; Cho, Y.; Gwon, Y.; Jang, Y.; Kim, S.; Choi, H.; Lee, J.; Kim, Y. Distribution of Porcine Endogenous Retrovirus in Different Organs of the Hybrid of a Landrace and a Jeju Domestic Pig in Korea. Transplant. Proc. 2015, 47, 2067–2071. [Google Scholar] [CrossRef]
- Zhang, P.; Yu, P.; Wang, W.; Zhang, L.; Li, S.; Bu, H. An effective method for the quantitative detection of porcine endogenous retrovirus in pig tissues. Vitr. Cell. Dev. Biol. - Anim. 2010, 46, 408–410. [Google Scholar] [CrossRef]
- Quereda, J.J.; Herrero-Medrano, J.M.; Abellaneda, J.M.; García-Nicolás, O.; Martínez-Alarcón, L.; Pallarés, F.J.; Ramírez, P.; Muñoz, A.; Ramis, G. Porcine Endogenous Retrovirus Copy Number in Different Pig Breeds is not Related to Genetic Diversity. Zoonoses Public Heal. 2012, 59, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Mang, R.; Maas, J.; Chen, X.; Goudsmit, J.; van der Kuyl, A.C. Identification of a novel type C porcine endogenous retrovirus: evidence that copy number of endogenous retroviruses increases during host inbreeding. J. Gen. Virol. 2001, 82, 1829–1834. [Google Scholar] [CrossRef]
- Lee, J.H.; Webb, G.C.; Allen, R.D.; Moran, C. Characterizing and mapping porcine endogenous retrovirusesin Westran pigs. J. Virol. 2002, 76, 5548–5556. [Google Scholar] [CrossRef]
- Groenen, M.A.M.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.-J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef]
- Pinheiro, L.B.; Coleman, V.A.; Hindson, C.M.; Herrmann, J.; Hindson, B.J.; Bhat, S.; Emslie, K.R. Evaluation ofa droplet digital polymerase chain reaction format for DNA copy number quantification. Anal. Chem. 2012, 84, 1003–1011. [Google Scholar] [CrossRef]
- Lee, J.; Choi, J.Y.; Lee, H.-J.; Kim, K.-C.; Choi, B.-S.; Oh, Y.-K.; Kim, Y.B. Repression of porcine endogenous retrovirus infection by human APOBEC3 proteins. Biochem. Biophys. Res. Commun. 2011, 407, 266–270. [Google Scholar] [CrossRef]
- Specke, V.; Rubant, S.; Denner, J. Productive Infection of Human Primary Cells and Cell Lines with Porcine Endogenous Retroviruses. Virology 2001, 285, 177–180. [Google Scholar] [CrossRef]
- Martin, U.; Winkler, M.E.; Id, M.; Radeke, H.; Arseniev, L.; Takeuchi, Y.; Simon, A.R.; Patience, C.; Haverich, A.; Steinhoff, G. Productive infection of primary human endothelial cells by pig endogenous retrovirus (PERV). Xenotransplantation 2000, 7, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. Porcine endogenous retrovirus infection of human peripheral blood mononuclear cells. Xenotransplantation 2014, 22, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. What does the PERV copy number tell us? Xenotransplantation. 2022, 2, e12732. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, R.P.; Wildschutte, J.H.; Russo, C.; Coffin, J.M. Identification, characterization, and comparative genomic distribution of the HERV-K (HML-2) group of human endogenous retroviruses. Retrovirology 2011, 8, 90–90. [Google Scholar] [CrossRef]
- Contreras-Galindo, R.; Kaplan, M.H.; He, S.; Contreras-Galindo, A.C.; Gonzalez-Hernandez, M.J.; Kappes, F.; Dube, D.; Chan, S.M.; Robinson, D.; Meng, F.; et al. HIV infection reveals widespread expansion of novel centromeric human endogenous retroviruses. Genome Res. 2013, 23, 1505–1513. [Google Scholar] [CrossRef]
- Denner, J. Recombinant porcine endogenous retroviruses (PERV-A/C): a new risk for xenotransplantation? Adv. Virol. 2008, 153, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Harrison, I.; Takeuchi, Y.; Bartosch, B.; Stoye, J.P. Determinants of High Titer in Recombinant Porcine Endogenous Retroviruses. J. Virol. 2004, 78, 13871–13879. [Google Scholar] [CrossRef]
- Denner, J.; Specke, V.; Thiesen, U.; Karlas, A.; Kurth, R. Genetic alterations of the long terminal repeat of an ecotropic porcine endogenous retrovirus during passage in human cells. Virology 2003, 314, 125–133. [Google Scholar] [CrossRef]
- Wilson, C.A.; Wong, S.; VanBrocklin, M.; Federspiel, M.J. Extended Analysis of the In Vitro Tropism of Porcine Endogenous Retrovirus. J. Virol. 2000, 74, 49–56. [Google Scholar] [CrossRef]
- Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A Comprehensive Strategy for Screening for Xenotransplantation-Relevant Viruses in a Second Isolated Population of Göttingen Minipigs. Viruses 2019, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Dieckhoff, B.; Puhlmann, J.; Büscher, K.; Hafner-Marx, A.; Herbach, N.; Bannert, N.; Büttner, M.; Wanke, R.; Kurth, R.; Denner, J. Expression of porcine endogenous retroviruses (PERVs) in melanomas of Munich miniature swine (MMS) Troll. Veter- Microbiol. 2007, 123, 53–68. [Google Scholar] [CrossRef]
- Bittmann, I.; Mihica, D.; Plesker, R.; Denner, J. Expression of porcine endogenous retroviruses (PERV) in different organs of a pig. Virology 2012, 433, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Scobie, L.; Taylor, S.; Wood, J.C.; Suling, K.M.; Quinn, G.; Meikle, S.; Patience, C.; Schuurman, H.-J.; Onions, D.E. Absence of Replication-Competent Human-Tropic Porcine Endogenous Retroviruses in the Germ Line DNA of Inbred Miniature Swine. J. Virol. 2004, 78, 2502–2509. [Google Scholar] [CrossRef]
- Wood, J.C.; Quinn, G.; Suling, K.M.; Oldmixon, B.A.; Van Tine, B.A.; Cina, R.; Arn, S.; Huang, C.A.; Scobie, L.; Onions, D.E.; et al. Identification of Exogenous Forms of Human-Tropic Porcine Endogenous Retrovirus in Miniature Swine. J. Virol. 2004, 78, 2494–2501. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.I.; Wilkinson, R.; Fishman, J.A. Genomic presence of recombinant porcine endogenous retrovirus in transmitting miniature swine. Virol. J. 2006, 3, 91. [Google Scholar] [CrossRef]
Figure 1.
Determination of the PERVpol copy number of 30 Auckland Island pigs bred in New Zealand by ddPCR. For comparison die PERVpol copy number of Göttingen minipigs and Aachen minipigs is shown [32]. Porcine GAPDH was chosen as reference gene.
Figure 1.
Determination of the PERVpol copy number of 30 Auckland Island pigs bred in New Zealand by ddPCR. For comparison die PERVpol copy number of Göttingen minipigs and Aachen minipigs is shown [32]. Porcine GAPDH was chosen as reference gene.
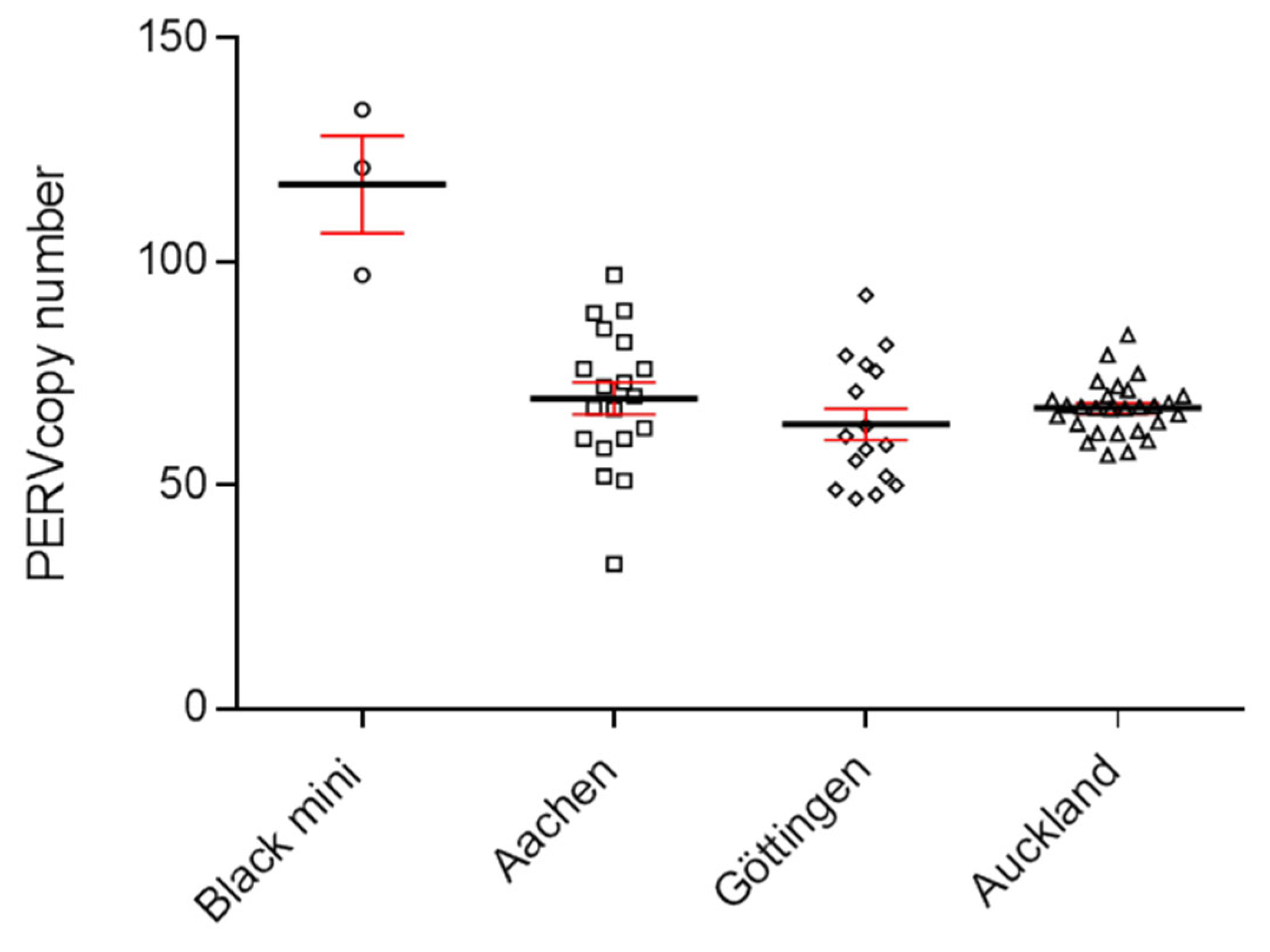
Figure 3.
Determination of the PERVpol copy number of four kidney cell lines from very young Auckland Island pigs, and from the first and second generation of pigs obtained by nuclear transfer from the embryonic kidney cells. The copy number of PK15 cells was determined as control. m, male, f, female.
Figure 3.
Determination of the PERVpol copy number of four kidney cell lines from very young Auckland Island pigs, and from the first and second generation of pigs obtained by nuclear transfer from the embryonic kidney cells. The copy number of PK15 cells was determined as control. m, male, f, female.
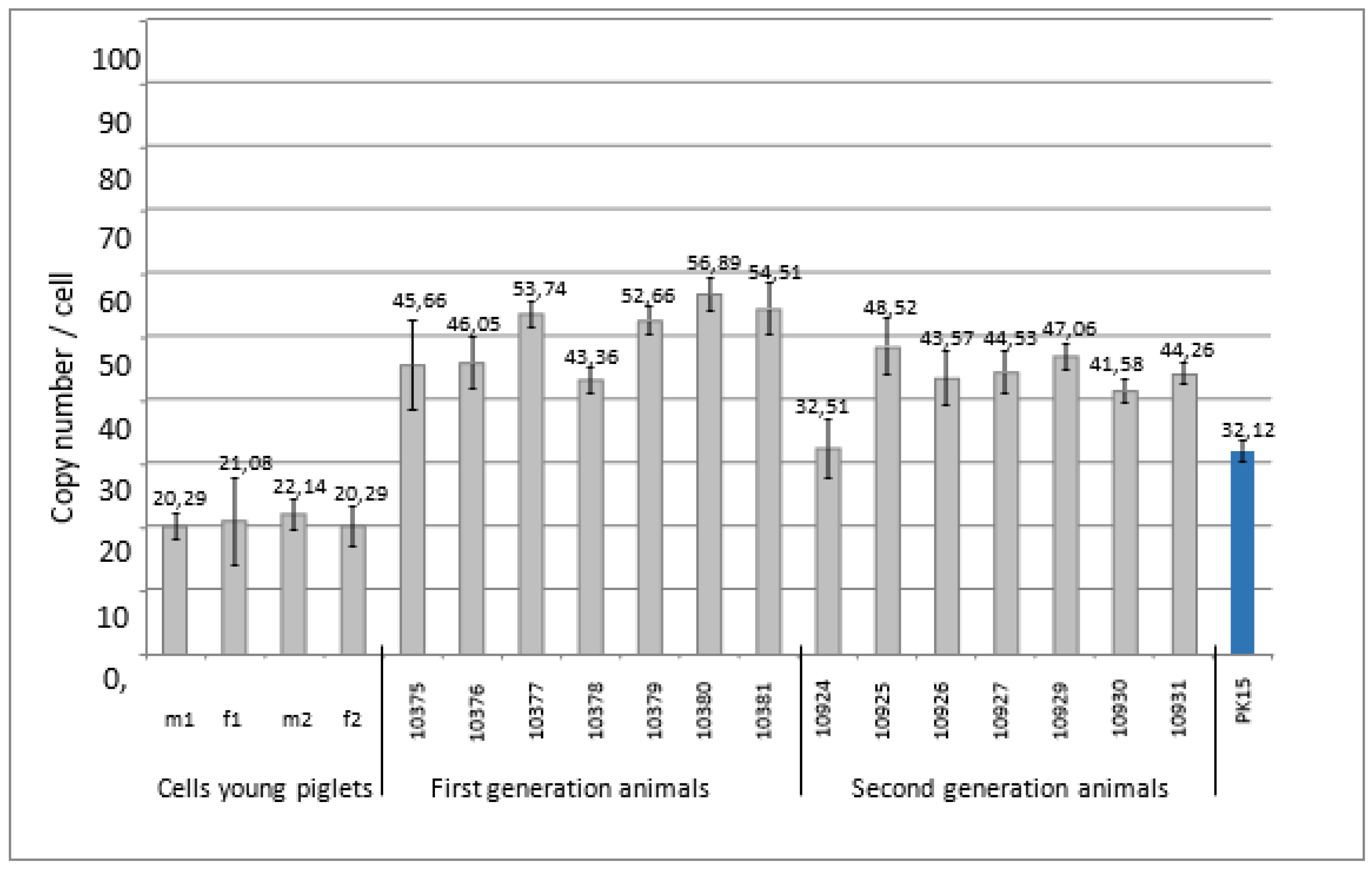
Figure 4.
Presentation of the differences of the PERV copy number in spleen and liver in dependence on the age of the animals. The PERV copy number was determined in both organs of dead born piglets (day 0), 9-30 days old and 12 weeks old animals using ddPCR and the difference was indicated in dependence on the age.
Figure 4.
Presentation of the differences of the PERV copy number in spleen and liver in dependence on the age of the animals. The PERV copy number was determined in both organs of dead born piglets (day 0), 9-30 days old and 12 weeks old animals using ddPCR and the difference was indicated in dependence on the age.
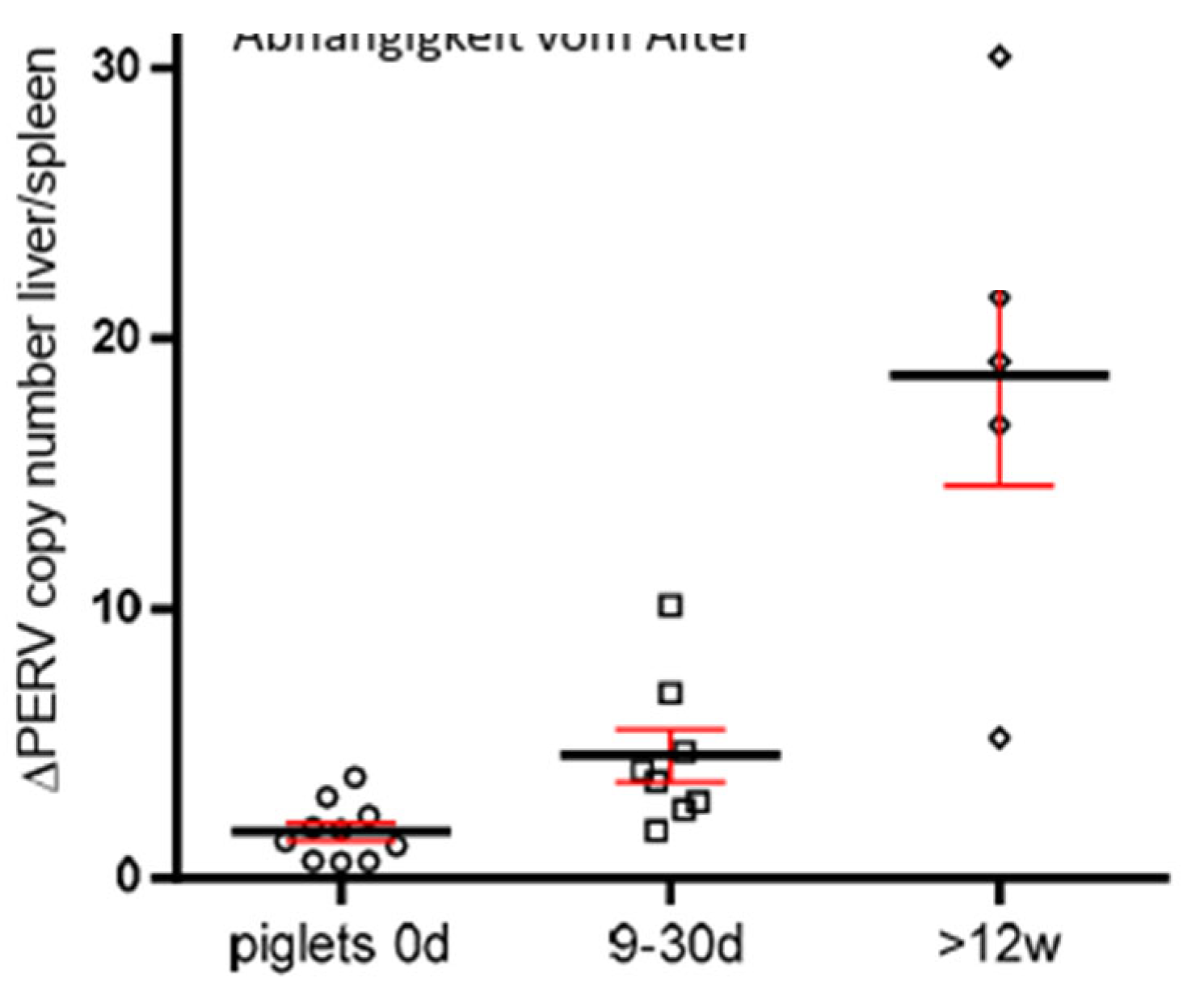
Figure 5.
Copy number of PERV in somatic cells and in cells used for SCNT indicating increase of the copy number in adult animals.
Figure 5.
Copy number of PERV in somatic cells and in cells used for SCNT indicating increase of the copy number in adult animals.
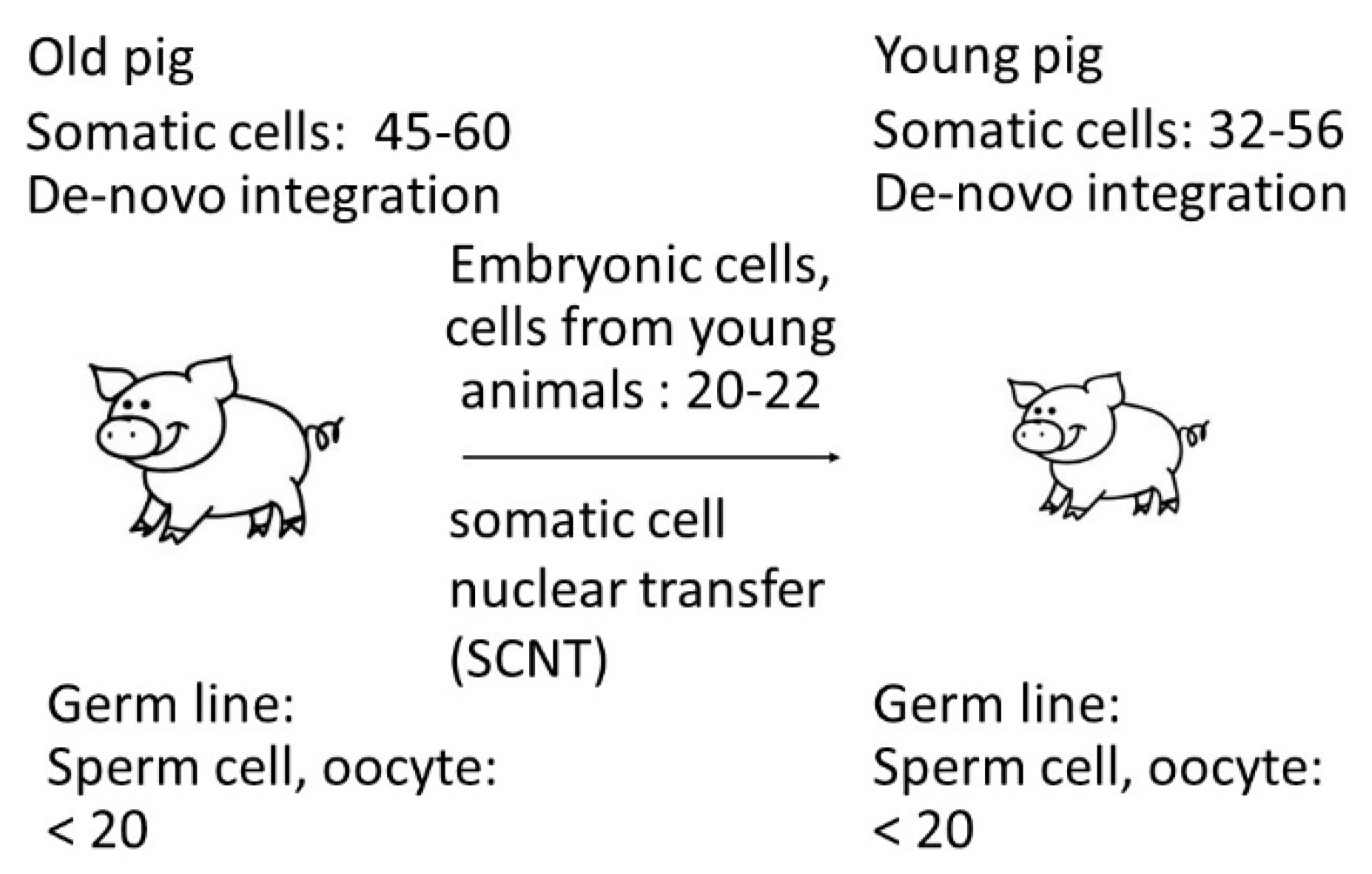
Table 1.
Primers and probes used for the estimation of the PERV copy number and the detection of PERV-C.
Table 1.
Primers and probes used for the estimation of the PERV copy number and the detection of PERV-C.
| Name | Sequence | Location (nucleotid number) | Accession number | Reference |
|---|---|---|---|---|
| PERV pol1-forward | CGACTGCCCCAAGGGTTCAA | 3568-3587 | HM159246 | Yang et al., 2015 [23] |
| PERV pol2-reverse | TCTCTCCTGCAAATCTGGGCC | 3803-3783 | ||
| PERV pol probe | /56FAM/CACGTACTGGAGGAGGGTCACCTG | 3678-3655 | ||
| Pig actin forward | TAACCGATCCTTTCAAGCATTT | Krüger et al., 2020 [24] | ||
| Pig actin reverse | TGGTTTCAAAGCTTGCATCATA | |||
| Pig actin probe | /5HEX/CGTGGGGATGCTTCCTGAGAAAG | |||
| Pig GAPDH forward Pig GAPDH reverse |
TTCACTCCGACCTTCACCAT CCGCGATCTAATGTTCTCTTTC |
3951-3970 4022-4001 |
NC_010447.5 (396823) | Krüger et al., 2020 [24] |
| Pig GAPDH probe | /5HEX/CAGCCGCGTCCCTGAGACAC | 3991-3972 | ||
| PERV-C forward PERV-C reverse |
CTGACCTGGATTAGAACTGG ATGTTAGAGGATGGTCCTGG |
6606-6625 6867-6886 |
AM229312 | Takeuchi et al. [25,26] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



