Submitted:
30 August 2023
Posted:
31 August 2023
Read the latest preprint version here
Abstract
Cardiac hypertrophy typically follows myocardial damage, a process known to involve members of the Krüppel-Like Factors (KLFs) which have pro-hypertrophic and anti-hypertrophic roles. Our study delved into the molecular mechanisms underlying KLFs-cardiac hypertrophy interplay post myocardial infraction. We induced myocardial dam-age in rats using isoproterenol. Total RNA was extracted from the left ventricle and Quantitative Real Time RT-PCR was conducted to assess the expression of KLFs, cardiac commitment genes, inflammatory markers, and certain conduction-related genes. We devel-oped a computational approach to construct a proteomic network centered on KLFs. Initial results revealed early expression (2-3 days post-induction) of KLF3, KLF4 and KLF6, followed by the subsequent expression of KLF11 and KLF15 (5-8 days after induction). In the maturation stage, KLF12 and KLF13 regulators were found upregulated. IL-6 was gradually found to be upregulated through. Interaction analysis revealed KLF3, KLF8 and KLF12 interacted with cardiac electrical functions. RT-PCR confirmed up-regulation in cardiac genes linked to electrical function and scar maturation. Our findings underscore the central role of KLFs during the modulation of the cardiac hypertrophic response. Dysregulation of KLF expression resulted in damaged myocardium, participating in the progression of abnormal hypertrophy, highlighting their potential as therapeutic targets for heart diseases.
Keywords:
Krüppel-like factors
; myocardial damage
; cardiovascular diseases
; hypertrophy.
1. Introduction
Cardiovascular diseases continue to hold the unenviable position as the leading cause of death worldwide[1]. Within this group of diseases, prominent exhibitors include ischemic heart disease, stroke, and heart failure, all of which contribute significantly to the global burden of morbidity and mortality [2]. Among them, ischemic heart disease stands out as one of the most severe due to its rapid onset, particularly when presenting as acute myocardial infarction, claiming over a million lives each year in the United States alone [3]. The sudden interruption of blood flow to the cardiac muscle triggers hypoxia, and if blood supply is not promptly restored, it culminates in tissue necrosis with irreversible loss of cardiomyocytes. From this moment on, a series of pathophysiological responses are unleashed, from which the myocardial tissue undergoes structural remodeling (pathological hypertrophy) to maintain cardiac contraction and prevent progression towards heart failure. This process involves the release of proinflammatory mediators, an increase in the secretion of extracellular matrix, and compensatory hypertrophy of the remaining cardiomyocytes [4].
The Krüppel-like factors (KLFs) are a family of transcription factors known to be associated with many biological processes, including the regulation of cardiac hypertrophy. These belong to a group of DNA-binding proteins that are part of the zinc finger transcription factor family. Each one of them is composed of three zinc fingers of the Cys2His2 type, which are capable of binding to three pairs of bases, respectively, in GC-rich regions, such as the consensus sequences CACCC-, GC-, and GT-box [5,6]. Since their discovery in 1993 up to the current date, a total of 18 KLFs have been reported, each with unique expression patterns and the ability to regulate gene transcription, either activating or repressing, numerous targets involved in physiological and pathological processes at the subcellular level [7]. These processes encompass growth, differentiation, and cellular apoptosis, as well as the maintenance of specialized tissues [8].
Moreover, KLFs are key players in the regulation of cardiac hypertrophy, and their activity can have both pro-hypertrophic and anti-hypertrophic effects, depending on the context and interactions with other regulatory factors. The development of cardiac hypertrophy can be influenced by several aspects, including changes in gene expression, modifications to cellular signaling pathways, and alterations to cellular processes within cardiomyocytes. For instance, KLF2, which is highly expressed in endothelial cells, has been identified as a negative regulator of cardiac hypertrophy [9,10]. Previous research has shown KLF2´s ability to inhibit TGFβ signaling in endothelial cells by promoting the expression of Smad7 and blocking the activator protein 1 (AP-1) activity [9]. Moreover, KLF2 induction by Simvastatin, achieved a significant reduction in the expression of hypertrophic genes, such as atrial natriuretic peptide (ANP), brain natriuretic peptide (BNP), and b-myosin heavy chain (b-MHC), in a Transverse Aortic Constriction (TAC) model [10]. In contrast, KLF5 has been found to be increased in models of cardiac damage, namely end-stage heart failure or diabetic cardiomyopathy [11,12]. Additionally, its role as a mediator of hypertrophy has been demonstrated in heterozygous KLF5 mice, in which a reduced response to Angiotensin II infusion was observed, resulting in less cardiomyocyte growth and fibrosis development, emphasizing the importance of its presence for the onset of hypertrophy [13]. The significance of KLF5 in hypertrophy development lies in its capacity to transactivate targets such as Platelet-Derived Growth Factor (PDGF)-A/B, Egr-1, Plasminogen Activator Inhibitor-1 (PAI-1), inducible Nitric Oxide Synthase (iNOS), and Vascular Endothelial Growth Factor (VEGF) receptors, amongst others, all of which play a pivotal role during cardiovascular remodeling [14].
In this study, we aimed to investigate the gene expression of specific KLFs following myocardial infarction induction and the development of hypertrophy. Our research group's primary goal is to unravel the intricate molecular mechanisms underlying cardiac hypertrophy and its potential links to inflammatory genes and cardiac conduction. By examining the patterns of KLFs expression and their interactions with key mediators, we aim to shed light on the regulatory processes governing cardiac hypertrophy.
2. Materials and Methods
2.1. Animal Study Approval
All experiments were carried out using 150g ± 20g Wistar rats. These were housed in polymethylmethacrylate boxes and kept at 20 – 24 °C in a room with 12-hour light/dark cycle. Food and water were available ad libitum in the cage. Research with animals carried out for this study was performed according to approved protocols (BI21-00006) and animal welfare regulations of Nuevo León Autonomous University´s Institutional Bioethics Committee.
2.2. Rat model and cardiotomy
For myocardial damage induction, 65 mg/kg of the beta-adrenergic drug, isoproterenol hydrochloride (SKU: I5627 Sigma-Aldrich, St. Louis, MO, USA) was administered intraperitoneally, dissolved in 0.5 mL of physiological solution (0.9% Sodium Chloride) at room temperature, in a single dose. The control group was administered with 0.5 mL of physiological solution intraperitoneally in a single dose. The experimental animals were euthanized under anesthesia by cervical dislocation. To collect the hearts, the thorax was accessed through an inverted T incision and longitudinal sternotomy. The heart was freed by clamping and cutting the great vessels of the corona cordis, allowing the removal of the organ from the rat's body. Myocardial damage was confirmed by H&E staining. This procedure was performed at different time lapses after chemical induction of cardiac insult with isoproterenol.
2.3. Cardiac morphometry and heart volume calculation
Heart´s length, width and height were measured with a digital vernier. Using the ellipsoid volume formula (V=3/4𝜋×𝐷1×𝐷2×𝐿), heart volumes were calculated and normalized to body weight of each experimental animal.
2.3. RNA Isolation and Quantitative Real-Time RT-PCR
Total RNA from the left ventricle area was extracted with TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA), according to the manufacturer's protocol. Complementary DNA (cDNA) was synthesized by reverse transcription reaction with 250 μg of total RNA using SuperScript™ VILO™ cDNA Synthesis Kit (Invitrogen; Thermo Fisher Scientific, Inc.). Real time PCR reactions were amplified and analyzed in triplicate using an Applied Biosystems 7500 Fast Real time PCR thermal cycler and following the SYBR-Green® FAST protocol (Thermo Fisher Scientific, Inc.) under the next reaction conditions: Step 1: 95°C for 20 seconds, Step 2: 40 cycles of 95°C for 3 seconds, Step 3: 40 cycles of 60°C for 30 seconds. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) levels were used to normalize the expression of target genes. The relative expression levels of the genes were calculated using the 2–∆∆Ct method. The list of primers sequences for target genes can be found in supplementary material.
2.4. Statistical analysis
The collected data was analyzed using SPSS software version 17.0 (SPSS, Inc. Chicago, IL, USA), data are expressed as mean ± SEM. Statistical significance was determined using one-way ANOVA with Dunnett correction. A P value <0.05 was considered statistically significant. Graphs and visual aids were made using GraphPad Prism Version 5 and the Biorender app, respectively.
2.5. STRING
STRING integrates information from multiple sources, including high-quality experiments, curated databases, and computational predictions, to construct large-scale protein interaction networks. We utilized STRING (version 11.5) with the multiple protein search option to explore their interactions. The resulting interactome was downloaded in TSV format and visualized using Excel. Our investigation primarily focused on the direct interactions involving the KLF proteins; priority was given to those with the highest correlation or weight, and we compared them with relevant literature. To facilitate a more intuitive comparison, we constructed a Venn diagram that depicted the distinct interactions of our genes of interest within the KLF protein families.
3. Results
Wistar rats of 150g ± 20g were initially injected with a single interperitoneally dose of isoproterenol (65mg/kg) to induce myocardial damage [15,16,17]. At 24-hour intervals, animals were sedated, euthanized, and subjected to cardiac removal. Figure 1A shows the representative surgical removal process. Cardiac hypertrophy was expected to occur in a period of approximately 21 days [18]. Figure 1B shows the comparative of the control size of the heart, and at 21 days post infarction induction.
Before surgery, animals were weighted, and after surgical removal of the organ, the heart was measured for length, width, as well as weighted (weights and measurements Supplemental Table 1). The measured variables cardiac volume was calculated (Figure 1C) and a graph of the cardiac volume vs cardiac weight was developed (Figure 1D), showing a continuous increase over time. As a final control of isoproterenol-induced cardiac damage, both ANP and aMHC, known markers of stress and cardiac hypertrophy, respectively, were shown to have an increase in expression over time, as seen on Figure 1E.
Since our main goal centered on better understanding the regulation of the Krüppel-like family of transcription factors, and their post-infarction role, we sought to measure their expression levels in combination to the cardiac commitment triad of GATA4, MEF2C, and TBX5 (GMT), as they represent the level of potential differentiation or loss of cardiomyocytes. As an insult to the heart promotes a state of inflammation, we further measured levels of known inflammatory cytokines IL-6 and IL1b.
Figure 2A shows a complete overview (RT-PCR heat map) of the entire KLF network of transcription factors, in addition to GMT and the inflammatory cytokines. Post infarction cardiac hypertrophy is segmented into three primary phases: an early inflammatory phase consisting of the first 4 days, followed by a proliferative stage (day 5 – 14) and a final onset of scar tissue maturation up to three weeks from the initial state [19]. Regarding this categorization, we conducted daily assessments of RNA expression from the initial 24 hours following the infarction event and extending up to day seven, which corresponds to the mid-inflammatory phase. Subsequently, samples were evaluated every alternate day for the duration of three weeks, encompassing the entire observation period.
Interestingly, regulatory effects were deemed close to the projected hypertrophic stages (Figure 2B). Several known KLFs related to cardiac function and growth exhibited significant alterations. First, our results showed an early up-regulation of KLF4, particularly at days 2 and 3, similarly KLF3 and 6 also have their highest peaks around day 3. These members of the KLF are noticeable as they are involved in pro-fibrotic and antiapoptotic signaling. Which could help explain the initial growth effects and fibrosis of the heart. Moreover, through the hypertrophic evolution related to the damage, in the proliferative stage, there is a remarkable upregulation of both KLF11 at days 5 and 6, and KLF15 at day 8. It is important to state that KLF11 is related to TGFb signaling which can lead to SMAD related cell growth [20,21]. Meanwhile, KLF15 is known directly as a hypertrophy modulator, known to inhibit both GATA4 and MEF2C [22]. Consistent with this observation we see a drastic drop in all 3 components of GMT: GATA4 on day 8 and both MEF2C and TBX5 on day 9. In the final mature stage, there is an up regulatory effect in KLF12 and KLF13, both pro-apoptotic regulators, which could indicate final patterning. Additionally, a quick look at IL6 showed a continuous increase in inflammation through the entire remodeling process (Figure 3 and Table 1).
Cardiac function is critically related to its electrical functions. As a first bioinformatic approach, we sought to identify genes interacting with KLFs. Using the STRING database, we constructed rudimentary networks, aimed to initiate the identification of direct correlations between electrical genes. Our initial screening yielded the presence of components such as ATP2A, CACNA1G, CX43, RYR2 (related to cardiac electrical function), GSK3b and SMAD3 (Wnt signaling elements), and TNF-a (inflammatory cytokine), Figure 4. Next, we proceeded to cluster these results based on the interacting KLFs subtypes. Our initial grouping effectively distinguished KLF3, 8, and 12 as displaying direct interactions, with the initially identified genes.
Finally, to confirm the regulatory effects of the electrical components, we proceeded with an opening expression screen. RNA samples were collected at intervals of 2 to 3 days for this preliminary screening. Figure 5 presents qRT-PCR heat map of the identified electrical elements, accompanied by plotted graphs for better visualization. These results show an up-regulatory tendency for most of the screened genes at the end of the proliferative stage, as well as during the scar maturation process.
4. Discussion
The heart is a critical organ in charge of distributing oxygen and nutrients to the entire body. Regrettably, this essential organ lacks the potential to self-regenerate, hence the heart is susceptible to the harsh effects of wear and tear, as well as environmental and genetic degeneration, which in turn lead to exacerbation of an inflammatory state, typically followed by loss of cardiomyocytes, fibrosis, and pathological remodeling [23,24,25,26]. Given its lack of natural capacity for renewal, it is critical to enhance our comprehension of the progression of these altered deteriorating stages. According to the American Heart Association myocardial infarction, better known as a heart attack, kills just over 800,000 people in the U.S. alone every year and is considered the leading killer of CVD´s [27]. The Krüppel-like factors family has recently received extensive attention, as novel research has centered in their involvement in many processes including embryogenesis and development of diseases related to several organs [24,28,29,30,31,32,33,34,35].
Isoproterenol is a b-adrenergic stimulant known to induce infarction-like lesions in the myocardium, leading to loss of cardiomyocytes, fibrosis, and remodeling, potentially leading to heart failure [15]. Several studies have confirmed that b-adrenergic stimulation increases cardiac contractility developing maladaptive hypertrophy, led by several mechanisms including enhanced protein synthesis, proto-oncogene expression, elevated oxidative stress, and stimulation of mitogen activated protein kinases [36]. As earlier stated, post infarction pathological remodeling typically takes 3 weeks (approximately 21 days). We tested a single intraperitoneal dose of isoproterenol (65 mg/kg), and over the course of 3 weeks, the relation of cardiac volume to cardiac weight more than doubled (Figure 1D). The cardiac hypertrophy was further confirmed by the increase in fibrosis as observed by H&E staining (supplemental Figure 1). The observed pathological maladaptive hypertrophy, as seen in our animal models is consistent with the earlier studies, wherein high dosages of isoproterenol induced significant irregularities in cardiac architecture, fibrotic infiltration, widening of the interstitium, and loss of cardiomyocytes [15]. Finally, Atrial Natriuretic Peptide (ANP) is considered a marker for cardiac stress. In clinical settings, ANP elevation has long been considered a staple of heart failure [37], as it has been shown to elevate during high demanding workflows, stimulating vasodilation and alleviates hypertension [38]. During a stress-related event such as the one promoted by isoproterenol, myocyte damage induces remodeling and an expected elevation in ANP [37], as seen in our results, Figure 1E shows a 5-fold surge in ANP levels and an initial 4-fold increase in aMHC expression, both indicative of cardiac damage.
As we followed myocardial damage over time, GMT alterations were expected. Post infarction-like stimulus is known to induce a strong MAPK kinase signaling cascade. As a result, considerable Gata4 phosphorylation occurs, favoring pro-hypertrophic gene transcription [39]. GATA4 is a transcription factor that directly regulates the expression of several heart-specific genes, including the aMHC, troponin C (Tnnc1), troponin I (Tnni3), NPPA and NPPB, respectively), and ion transport genes, such as the cardiac sodium-calcium exchanger (Slc8a1). Previously, murine research has shown that specific Gata4 overexpression can cause cardiac hypertrophy, as demonstrated by [40] who used transgenic animals capable of expressing Gata4 2.5 times more than wild type. In Figure 2 we observed this similar increase by day 8. Moreover, Mef2C and Tbx5, both essential players in Gata4 regulation show similar trends of upregulation by day 9. Our interactome data (Figure 4B) further confirmed this interaction and showed a strong GMT relation to KLF 4, 5, 13 and 15. Moreover, Gata4 showed involvement with KLF 6 which related to SMAD signaling. TGFb, a driver of SMAD signaling, functions as an effector of tissue fibrosis, which can eventually lead to scaring and elevation of collagen related genes [41]. Both KLF4 and 6 exhibited early post myocardial damage elevations, potentially leading the way for hypertrophy. Under this perspective hypertrophy could lead to high levels of fibrosis in the heart, reducing its function and increasing its size, yet around 1 week the upregulation of KLF 11 and 15 peak potentially blocking hypertrophy progression [32,42,43], marking the initiation of a final maturation stage, wherein we can observe an elevation of KLF 12 and 13. While information regarding KLF 12 is limited [44] its involvement in the upregulation of Survivin, -an antiapoptotic factor- has been demonstrated. In addition, KLF 13 seems to be in balance as it is potentially inhibiting BCL-XL and consequently triggering cell death. Left Anterior Descending Coronary Artery Ligation (LAD) murine models have shown an increase in KLF13 expression related to early phase cardiomyocyte death, a contrasting effect to that of KLF4 [45,46].
GMT genes contribute to cardiac maintenance, contraction, and growth of the heart [47,48,49,50]. Unsurprisingly, electrical function is also related to these genes. Tbx5 has been related to SRF and CX43 (early activation post myocardial damage), which are directly related to myocardial stress, which, when exacerbated, leads to pathological hypertrophy [40,47]. Cardiac muscle contraction (RYR, ATP2A, aMHC) and electrical activity are intrinsically related. Interestingly our findings reveal a sharp decrease in several of these genes, particularly by day 10 (Figure 5), strongly suggesting a decline in function, along with the preceding drop of CX43 expression [51]. Notably, our initial bioinformatic screening did directly link KLFs to these genes, implying indirect interactions. These interactions could further explain both up- and down- regulatory effects, making further exploration imperative to better elucidate these mechanisms.
5. Conclusion
The heart is a vital organ that can be damaged by various factors, including stress, genetic defects, and environmental factors. At high dosage Isoproterenol can induce myocardial damage, leading to inflammation, loss of heart muscle cells, scarring, and remodeling or hypertrophy.
KLFs are involved in a variety of cellular processes, including gene transcription, cell growth, and differentiation. Our study showed that KLFs are dysregulated by myocardial damage promoting the progression of abnormal hypertrophy. We showed that expression profiles of different KLFs change during different stages of cardiac damage, suggesting that KLFs may play a role in modulating the hypertrophic response and influencing downstream signaling pathways. The findings of this study provide new insights into the molecular mechanisms underlying cardiac damage and abnormal hypertrophy. A better understanding of their targets and interacting partners could help us lead to the development of new therapeutic interventions to treat heart diseases.
Supplementary Materials
Supplemental Figure 1. Infarct Model. H&E staining A, B) 21d post infarct high fibroblast infiltration, C) Loss of muscle fibers, D) Control no infarct. Supplemental Table 1. Primer list. Supplemental Table 2. Weights and lengths.
Author Contributions
Conceptualization: JFI, GRP-R, JLD-G; methodology MGS-S, JAG-L, JDM-M, JLD-G; software JAG-L, PZ-M; validation GPR-R; formal analysis GRP-R, JAG-L, PZ-M; investigation JDM-M, AE-R; resources: LG-O, AS-D, AE-R, AC-M; writing and editing: JFI, DFB-C, ENG-T; supervision JFI, project administration JFI, GRP-R, ENG-T, funding acquisition JFI, AR-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CONAHCYT, grant Ciencia de Fronteras No. CF-MG-20191004101335125-51208
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of Facultad de Medicina, Universidad Autonoma de Nuevo Leon, protocol number BI21-00006.
Conflicts of Interest
The authors declare no conflict of interest.
References
- WHO The Top 10 Causes of Death Available online:. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 2 August 2023).
- Gaziano, T.; Reddy, S.; Paccaud, F.; Horton, S. Disease Control Priorities in Developing Countries. In ; Jamison, D., Berman, J., Measham, A., Eds.; Oxford University Press: Washington.
- Mechanic, O.J.; Gavin, M.; Grossman, S.A.; Ziegler, K. Acute Myocardial Infarction (Nursing). 2022.
- Ferrini, A.; Stevens, M.M.; Sattler, S.; Rosenthal, N. Toward Regeneration of the Heart: Bioengineering Strategies for Immunomodulation. Front Cardiovasc Med 2019, 6, 26. [Google Scholar] [PubMed]
- Oishi, Y.; Manabe, I. Krüppel-like Factors in Metabolic Homeostasis and Cardiometabolic Disease. Front Cardiovasc Med 2018, 5, 69. [Google Scholar] [CrossRef] [PubMed]
- Prosdocimo, D.A.; Sabeh, M.K.; Jain, M.K. Kruppel-like Factors in Muscle Health and Disease. Trends Cardiovasc Med 2015, 25, 278–287. [Google Scholar] [CrossRef]
- McConnell, B.B.; Yang, V.W. Mammalian Krüppel-like Factors in Health and Diseases. Physiol Rev 2010, 90, 1337–1381. [Google Scholar] [PubMed]
- Tetreault, M.-P.; Yang, Y.; Katz, J.P. Krüppel-like Factors in Cancer. Nat Rev Cancer 2013, 13, 701–713. [Google Scholar]
- Boon, R.A.; Fledderus, J.O.; Volger, O.L.; Van Wanrooij, E.J.A.; Pardali, E.; Weesie, F.; Kuiper, J.; Pannekoek, H.; Ten Dijke, P.; Horrevoets, A.J.G. KLF2 Suppresses TGF-β Signaling in Endothelium through Induction of Smad7 and Inhibition of AP-1. Arterioscler Thromb Vasc Biol 2007, 27, 532–539. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Liu, J.; Chen, X.; Duan, Y.; Wang, X.; Shen, Y.; Kuang, Y.; Zhuang, T.; Tomlinson, B. Endothelial Klf2-Foxp1-TGFβ Signal Mediates the Inhibitory Effects of Simvastatin on Maladaptive Cardiac Remodeling. Theranostics 2021, 11, 1609. [Google Scholar] [CrossRef]
- Kyriazis, I.D.; Hoffman, M.; Gaignebet, L.; Lucchese, A.M.; Markopoulou, E.; Palioura, D.; Wang, C.; Bannister, T.D.; Christofidou-Solomidou, M.; Oka, S. KLF5 Is Induced by FOXO1 and Causes Oxidative Stress and Diabetic Cardiomyopathy. Circ Res 2021, 128, 335–357. [Google Scholar] [CrossRef]
- Hoffman, M.; Palioura, D.; Kyriazis, I.D.; Cimini, M.; Badolia, R.; Rajan, S.; Gao, E.; Nikolaidis, N.; Schulze, P.C.; Goldberg, I.J. Cardiomyocyte Krüppel-like Factor 5 Promotes de Novo Ceramide Biosynthesis and Contributes to Eccentric Remodeling in Ischemic Cardiomyopathy. Circulation 2021, 143, 1139–1156. [Google Scholar] [CrossRef]
- Shindo, T.; Manabe, I.; Fukushima, Y.; Tobe, K.; Aizawa, K.; Miyamoto, S.; Kawai-Kowase, K.; Moriyama, N.; Imai, Y.; Kawakami, H. Krüppel-like Zinc-Finger Transcription Factor KLF5/BTEB2 Is a Target for Angiotensin II Signaling and an Essential Regulator of Cardiovascular Remodeling. Nat Med 2002, 8, 856–863. [Google Scholar] [CrossRef]
- Nagai, R.; Suzuki, T.; Aizawa, K.; Shindo, T.; Manabe, I. Significance of the Transcription Factor KLF5 in Cardiovascular Remodeling. Journal of Thrombosis and Haemostasis 2005, 3, 1569–1576. [Google Scholar] [PubMed]
- Grimm, D. Development of Heart Failure Following Isoproterenol Administration in the Rat: Role of the Renin–Angiotensin System. Cardiovasc Res 1998, 37, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.; Rajabian, A.; Sobhanifar, M.-A.; Alavi, M.S.; Taghipour, Z.; Hasanpour, M.; Iranshahi, M.; Boroumand-Noughabi, S.; Banach, M.; Sahebkar, A. Attenuation of Isoprenaline-Induced Myocardial Infarction by Rheum Turkestanicum. Biomedicine & Pharmacotherapy 2022, 148, 112775. [Google Scholar] [CrossRef]
- Liu, Y.; Jia, H.; Chang, X.; Ding, G.; Zhang, H.; Zou, Z.-M. The Metabolic Disturbances of Isoproterenol Induced Myocardial Infarction in Rats Based on a Tissue Targeted Metabonomics. Mol Biosyst 2013, 9, 2823. [Google Scholar] [CrossRef]
- Ferrini, A.; Stevens, M.M.; Sattler, S.; Rosenthal, N. Toward Regeneration of the Heart: Bioengineering Strategies for Immunomodulation. Front Cardiovasc Med 2019, 6. [Google Scholar] [CrossRef]
- Venugopal, H.; Hanna, A.; Humeres, C.; Frangogiannis, N.G. Properties and Functions of Fibroblasts and Myofibroblasts in Myocardial Infarction. Cells 2022, 11, 1386. [Google Scholar] [CrossRef] [PubMed]
- Pardali, E.; Sanchez-Duffhues, G.; Gomez-Puerto, M.C.; Dijke, P. TGF- β -Induced Endothelial-Mesenchymal Transition in Fibrotic Diseases. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Liu, Y.; Wang, N.; Qi, Y.; Du, J. Krüppel-Like Factor 4 Transcriptionally Regulates TGF-Β1 and Contributes to Cardiac Myofibroblast Differentiation. PLoS One 2013, 8, 0–9. [Google Scholar] [CrossRef]
- Fisch, S.; Gray, S.; Heymans, S.; Haldar, S.M.; Wang, B.; Pfister, O.; Cui, L.; Kumar, A.; Lin, Z.; Sen-Banerjee, S.; et al. Kruppel-like Factor 15 Is a Regulator of Cardiomyocyte Hypertrophy. Proc Natl Acad Sci U S A 2007, 104, 7074–7079. [Google Scholar] [CrossRef]
- Islas, J.; Moreno-Cuevas, J. A MicroRNA Perspective on Cardiovascular Development and Diseases: An Update. Int J Mol Sci 2018, 19, 2075. [Google Scholar] [CrossRef]
- Santoyo-Suarez, M.G.; Mares-Montemayor, J.D.; Padilla-Rivas, G.R.; Delgado-Gallegos, J.L.; Quiroz-Reyes, A.G.; Roacho-Perez, J.A.; Benitez-Chao, D.F.; Garza-Ocañas, L.; Arevalo-Martinez, G.; Garza-Treviño, E.N.; et al. The Involvement of Krüppel-like Factors in Cardiovascular Diseases. Life 2023, 13, 420. [Google Scholar] [CrossRef] [PubMed]
- Sarre-Álvarez, D.; Cabrera-Jardines, R Rodríguez-Weber, F. Enfermedad Cardiovascular Aterosclerótica. Revisión de Las Escalas de Riesgo y Edad Cardiovascular. Med Int Mex 2018, 6, 910–923. [Google Scholar]
- Goradel, N.H.; Hour, F.G.; Negahdari, B.; Malekshahi, Z.V.; Hashemzehi, M.; Masoudifar, A.; Mirzaei, H. Stem Cell Therapy: A New Therapeutic Option for Cardiovascular Diseases. J Cell Biochem 2018, 119, 95–104. [Google Scholar] [CrossRef]
- Tsao, C.W.; Aday, A.W.; Almarzooq, Z.I.; Anderson, C.A.M.; Arora, P.; Avery, C.L.; Baker-Smith, C.M.; Beaton, A.Z.; Boehme, A.K.; Buxton, A.E.; et al. Heart Disease and Stroke Statistics—2023 Update: A Report From the American Heart Association. Circulation 2023, 147. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Sukhorukov, V.N.; Kalmykov, V.A.; Grechko, A. V.; Shakhpazyan, N.K.; Orekhov, A.N. The Role of KLF2 in the Regulation of Atherosclerosis Development and Potential Use of KLF2-Targeted Therapy. Biomedicines 2022, 10, 254. [Google Scholar] [CrossRef]
- Vinjamur, D.S.; Wade, K.J.; Mohamad, S.F.; Haar, J.L.; Sawyer, S.T.; Lloyd, J.A. Krüppel-like Transcription Factors KLF1 and KLF2 Have Unique and Coordinate Roles in Regulating Embryonic Erythroid Precursor Maturation. Haematologica 2014, 99, 1565–1573. [Google Scholar] [CrossRef]
- Shankman, L.S.; Gomez, D.; Cherepanova, O.A.; Salmon, M.; Alencar, G.F.; Haskins, R.M.; Swiatlowska, P.; Newman, A.A.C.; Greene, E.S.; Straub, A.C.; et al. KLF4-Dependent Phenotypic Modulation of Smooth Muscle Cells Has a Key Role in Atherosclerotic Plaque Pathogenesis. Nat Med 2015, 21, 628–637. [Google Scholar] [CrossRef]
- Pabona, J.M.P.; Zeng, Z.; Simmen, F.A.; Simmen, R.C.M. Functional Differentiation of Uterine Stromal Cells Involves Cross-Regulation between Bone Morphogenetic Protein 2 and Krüppel-like Factor (KLF) Family Members KLF9 and KLF13. Endocrinology 2010, 151, 3396–3406. [Google Scholar] [CrossRef]
- Leenders, J.J.; Wijnen, W.J.; van der Made, I.; Hiller, M.; Swinnen, M.; Vandendriessche, T.; Chuah, M.; Pinto, Y.M.; Creemers, E.E. Repression of Cardiac Hypertrophy by KLF15: Underlying Mechanisms and Therapeutic Implications. PLoS One 2012, 7, 1–10. [Google Scholar] [CrossRef]
- Zheng, H.; Pritchard, D.M.; Yang, X.; Bennett, E.; Liu, G.; Liu, C.; Ai, W. KLF4 Gene Expression Is Inhibited by the Notch Signaling Pathway That Controls Goblet Cell Differentiation in Mouse Gastrointestinal Tract. AJP: Gastrointestinal and Liver Physiology 2009, 296, G490–G498. [Google Scholar] [CrossRef]
- Yang, H.; Xi, X.; Zhao, B.; Su, Z.; Wang, Z. KLF4 Protects Brain Microvascular Endothelial Cells from Ischemic Stroke Induced Apoptosis by Transcriptionally Activating MALAT1. Biochem Biophys Res Commun 2018, 495, 2376–2382. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.P.; Wang, R.; Yang, X.; Wang, X.; Wang, L.; Ting, K.K.; Foley, M.; Cogger, V.; Yang, Z.; Liu, F.; et al. Ponatinib (AP24534) Inhibits MEKK3-KLF Signaling and Prevents Formation and Progression of Cerebral Cavernous Malformations. Sci Adv 2018, 4. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, D.; Tangutur, A.D.; Khatua, T.N.; Saxena, P.; Banerjee, S.K.; Bhadra, M.P. A Proteomic View of Isoproterenol Induced Cardiac Hypertrophy: Prohibitin Identified as a Potential Biomarker in Rats. J Transl Med 2013, 11, 130. [Google Scholar] [CrossRef] [PubMed]
- Ilatovskaya, D. V.; Levchenko, V.; Winsor, K.; Blass, G.R.; Spires, D.R.; Sarsenova, E.; Polina, I.; Zietara, A.; Paterson, M.; Kriegel, A.J.; et al. Effects of Elevation of ANP and Its Deficiency on Cardiorenal Function. JCI Insight 2022, 7. [Google Scholar] [CrossRef] [PubMed]
- NISHIKIMI, T.; MAEDA, N.; MATSUOKA, H. The Role of Natriuretic Peptides in Cardioprotection. Cardiovasc Res 2006, 69, 318–328. [Google Scholar] [CrossRef]
- Tenhunen, O.; Sármán, B.; Kerkelä, R.; Szokodi, I.; Papp, L.; Tóth, M.; Ruskoaho, H. Mitogen-Activated Protein Kinases P38 and ERK 1/2 Mediate the Wall Stress-Induced Activation of GATA-4 Binding in Adult Heart. Journal of Biological Chemistry 2004, 279, 24852–24860. [Google Scholar] [CrossRef]
- Liang, Q.; De Windt, L.J.; Witt, S.A.; Kimball, T.R.; Markham, B.E.; Molkentin, J.D. The Transcription Factors GATA4 and GATA6 Regulate Cardiomyocyte Hypertrophy in Vitro and in Vivo. Journal of Biological Chemistry 2001, 276, 30245–30253. [Google Scholar] [CrossRef]
- Hu, H.-H.; Chen, D.-Q.; Wang, Y.-N.; Feng, Y.-L.; Cao, G.; Vaziri, N.D.; Zhao, Y.-Y. New Insights into TGF-β/Smad Signaling in Tissue Fibrosis. Chem Biol Interact 2018, 292, 76–83. [Google Scholar] [CrossRef]
- Leenders, J.J.; Wijnen, W.J.; Hiller, M.; Van Der Made, I.; Lentink, V.; Van Leeuwen, R.E.W.; Herias, V.; Pokharel, S.; Heymans, S.; De Windt, L.J.; et al. Regulation of Cardiac Gene Expression by KLF15, a Repressor of Myocardin Activity. Journal of Biological Chemistry 2010, 285, 27449–27456. [Google Scholar] [CrossRef]
- Wenying, L.; Lu, H.; Sun, J.; Zhao, G.; Wang, H.; Guo, Y.; Eitzman, D.; Chen, E.; Fan, Y.; Zhang, J. KLF11 Protects against Venous Thrombosis via Suppressing Tissue Factor Expression. Thromb Haemost 2021, 122, 777–788. [Google Scholar] [CrossRef]
- Mak, C.S.L.; Yung, M.M.H.; Hui, L.M.N.; Leung, L.L.; Liang, R.; Chen, K.; Liu, S.S.; Qin, Y.; Leung, T.H.Y.; Lee, K.-F.; et al. MicroRNA-141 Enhances Anoikis Resistance in Metastatic Progression of Ovarian Cancer through Targeting KLF12/Sp1/Survivin Axis. Mol Cancer 2017, 16, 11. [Google Scholar] [CrossRef]
- Bayoumi, A.S.; Park, K.; Wang, Y.; Teoh, J.; Aonuma, T.; Tang, Y.; Su, H.; Weintraub, N.L.; Kim, I. A Carvedilol-Responsive MicroRNA, MiR-125b-5p Protects the Heart from Acute Myocardial Infarction by Repressing pro-Apoptotic Bak1 and Klf13 in Cardiomyocytes. J Mol Cell Cardiol 2018, 114, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Herring, B.P. Mechanisms Responsible for the Promoter-Specific Effects of Myocardin. J Biol Chem 2005, 280, 10861–10869. [Google Scholar] [CrossRef] [PubMed]
- Christoforou, N.; Chellappan, M.; Adler, A.F.; Kirkton, R.D.; Wu, T.; Addis, R.C.; Bursac, N.; Leong, K.W. Transcription Factors MYOCD, SRF, Mesp1 and SMARCD3 Enhance the Cardio-Inducing Effect of GATA4, TBX5, and MEF2C during Direct Cellular Reprogramming. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Liu, Y. Earlier and Broader Roles of Mesp1 in Cardiovascular Development. Cellular and Molecular Life Sciences 2017, 74, 1969–1983. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, L.; Diaz, A.D.; Benham, A.; Xu, X.; Wijaya, C.S.; Fa’Ak, F.; Luo, W.; Soibam, B.; Azares, A.; et al. Mesp1 Marked Cardiac Progenitor Cells Repair Infarcted Mouse Hearts. Sci Rep 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Dirkx, E.; Da Costa Martins, P.A.; De Windt, L.J. Regulation of Fetal Gene Expression in Heart Failure ☆. 2013. [CrossRef]
- Zhang, Y.; Zheng, Y.; Wang, S.; Fan, Y.; Ye, Y.; Jing, Y.; Liu, Z.; Yang, S.; Xiong, M.; Yang, K.; et al. Single-Nucleus Transcriptomics Reveals a Gatekeeper Role for FOXP1 in Primate Cardiac Aging. Protein Cell 2022. [Google Scholar] [CrossRef]
Figure 1.
Isoproterenol induced cardiac damage. A) Cardiac surgical removal procedure, B) Left, control vs right, 21-day Isoproterenol induced damage (hypertrophy). C) Cardiac volume schematic and volume formula, D) Cardiac volume / cardiac weight plot, E) ANP and aMHC expression control vs isoproterenol induced damage.
Figure 1.
Isoproterenol induced cardiac damage. A) Cardiac surgical removal procedure, B) Left, control vs right, 21-day Isoproterenol induced damage (hypertrophy). C) Cardiac volume schematic and volume formula, D) Cardiac volume / cardiac weight plot, E) ANP and aMHC expression control vs isoproterenol induced damage.
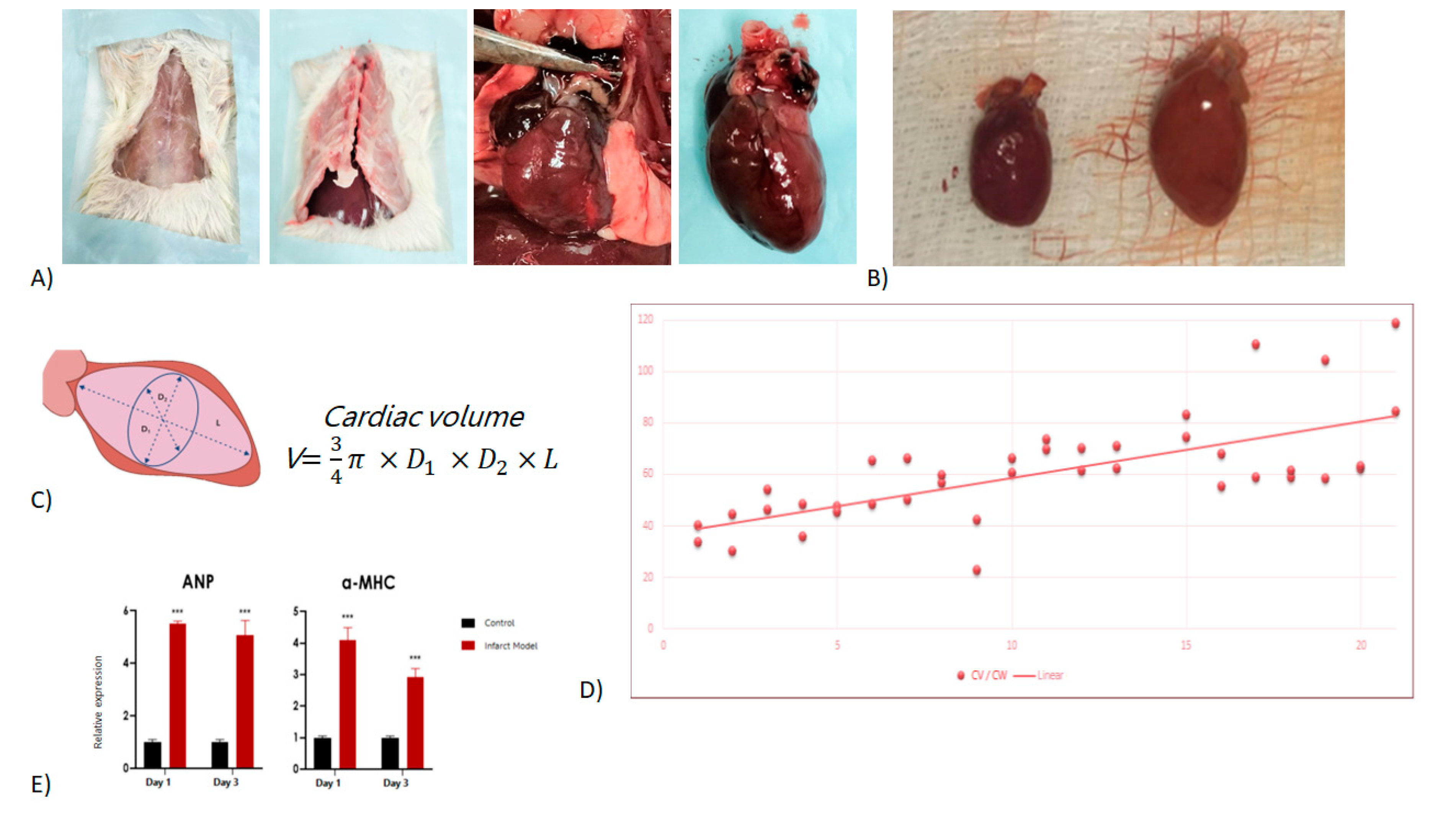
Figure 2.
Gene expression profiles. A) Heatmap of representative gene expression for the entire Krüppel-like family and related genes, B) qRT-PCR showed gene expression for KLFs, inflammation and GMT representative of distinctive early-, mid- and late-stage remodeling phases.
Figure 2.
Gene expression profiles. A) Heatmap of representative gene expression for the entire Krüppel-like family and related genes, B) qRT-PCR showed gene expression for KLFs, inflammation and GMT representative of distinctive early-, mid- and late-stage remodeling phases.
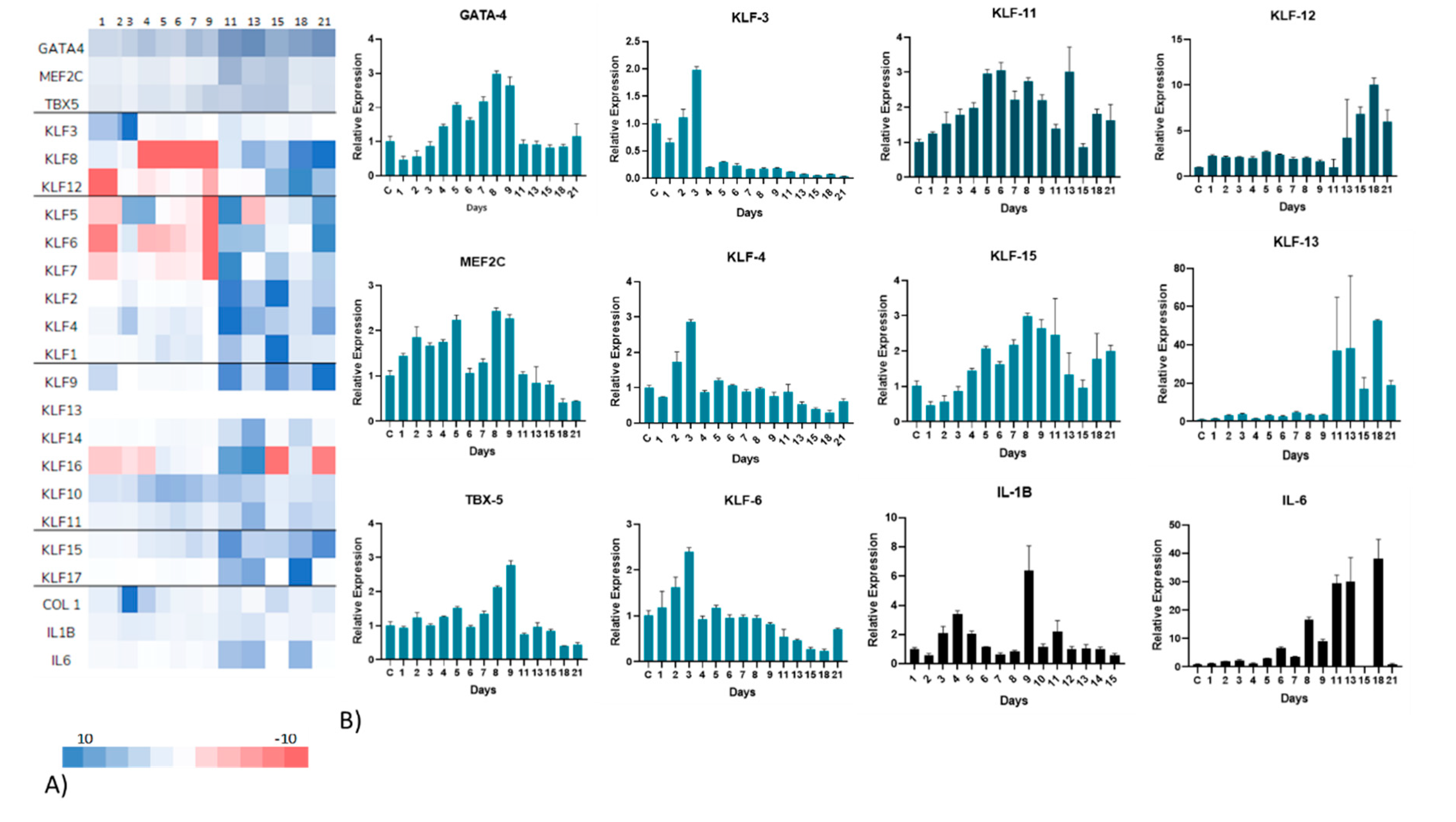
Figure 3.
Schematic representation of activation of particular Krüppel-like factors at different stages of remodeling post isoproterenol induced cardiac damage.
Figure 3.
Schematic representation of activation of particular Krüppel-like factors at different stages of remodeling post isoproterenol induced cardiac damage.
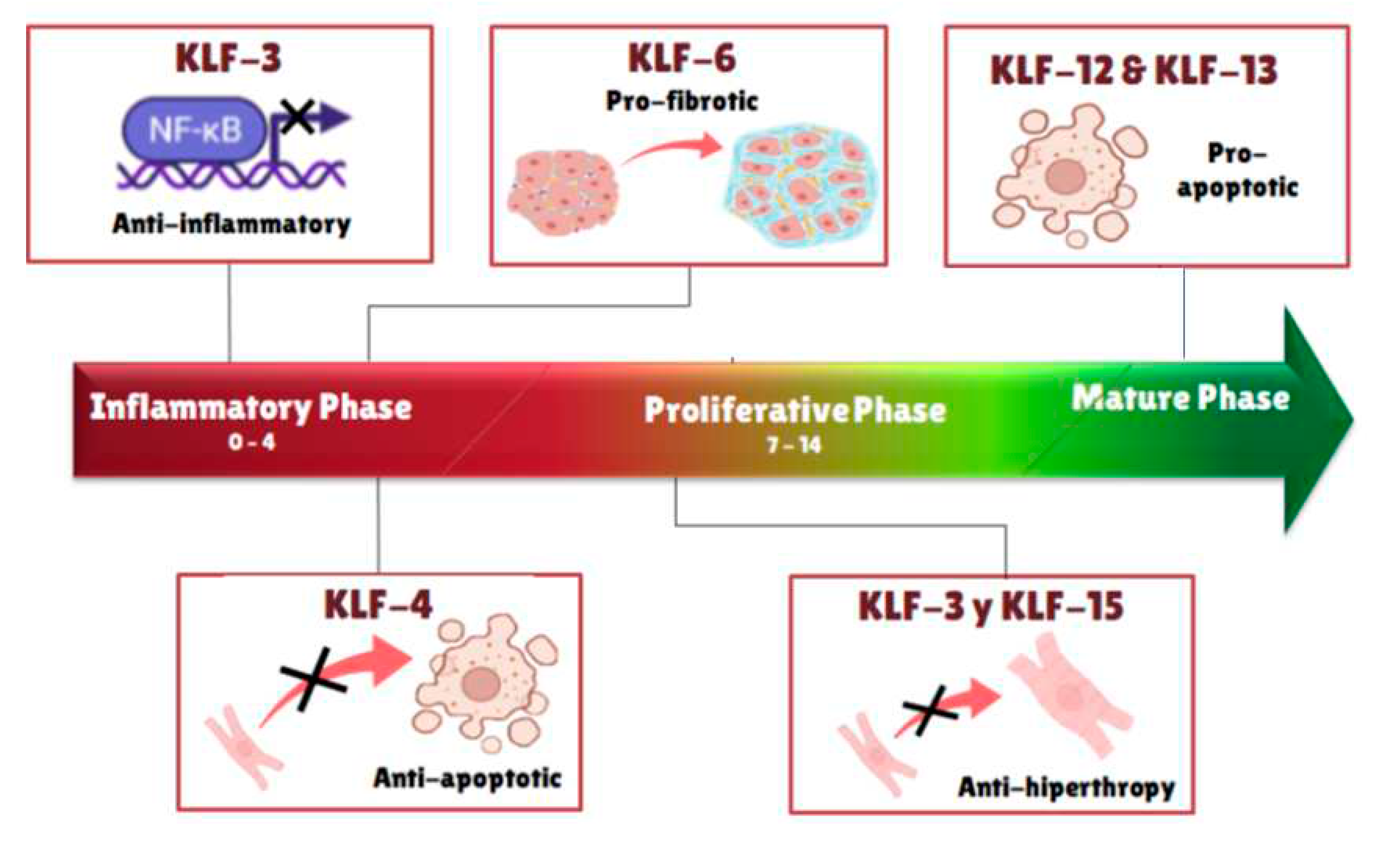
Figure 4.
Left, Venn diagram of interactions given by the predicting string interactome (right). Right, interactome shows direct and indirect relation of KLF´s, GMT, electrical, and inflammatory genes. No initial observable interactions for KLF3, KLF8 and KLF12.
Figure 4.
Left, Venn diagram of interactions given by the predicting string interactome (right). Right, interactome shows direct and indirect relation of KLF´s, GMT, electrical, and inflammatory genes. No initial observable interactions for KLF3, KLF8 and KLF12.
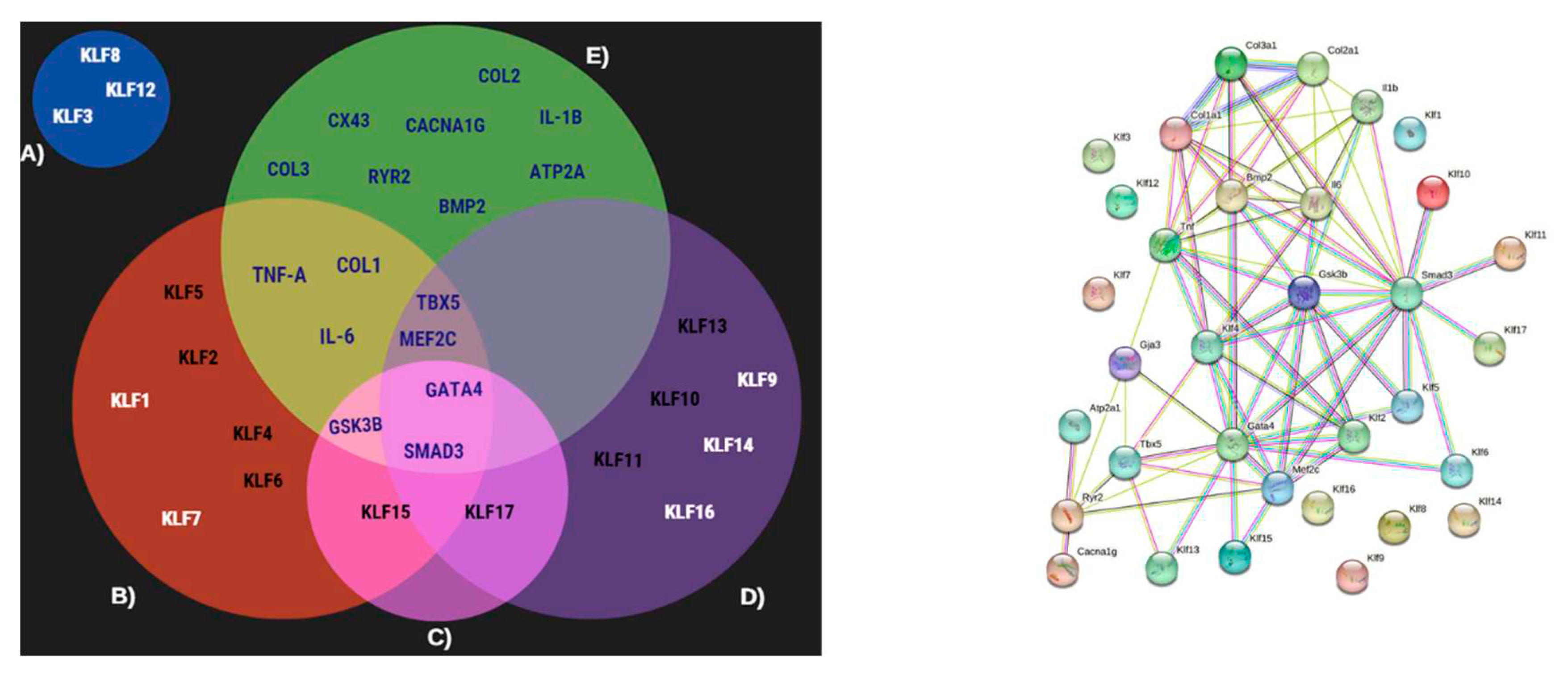
Figure 5.
Electrical gene profile, Left-top, shows the representative heatmap expression. Additionally, qRT-PCR graphs plots presenting gene expression profiles.
Figure 5.
Electrical gene profile, Left-top, shows the representative heatmap expression. Additionally, qRT-PCR graphs plots presenting gene expression profiles.
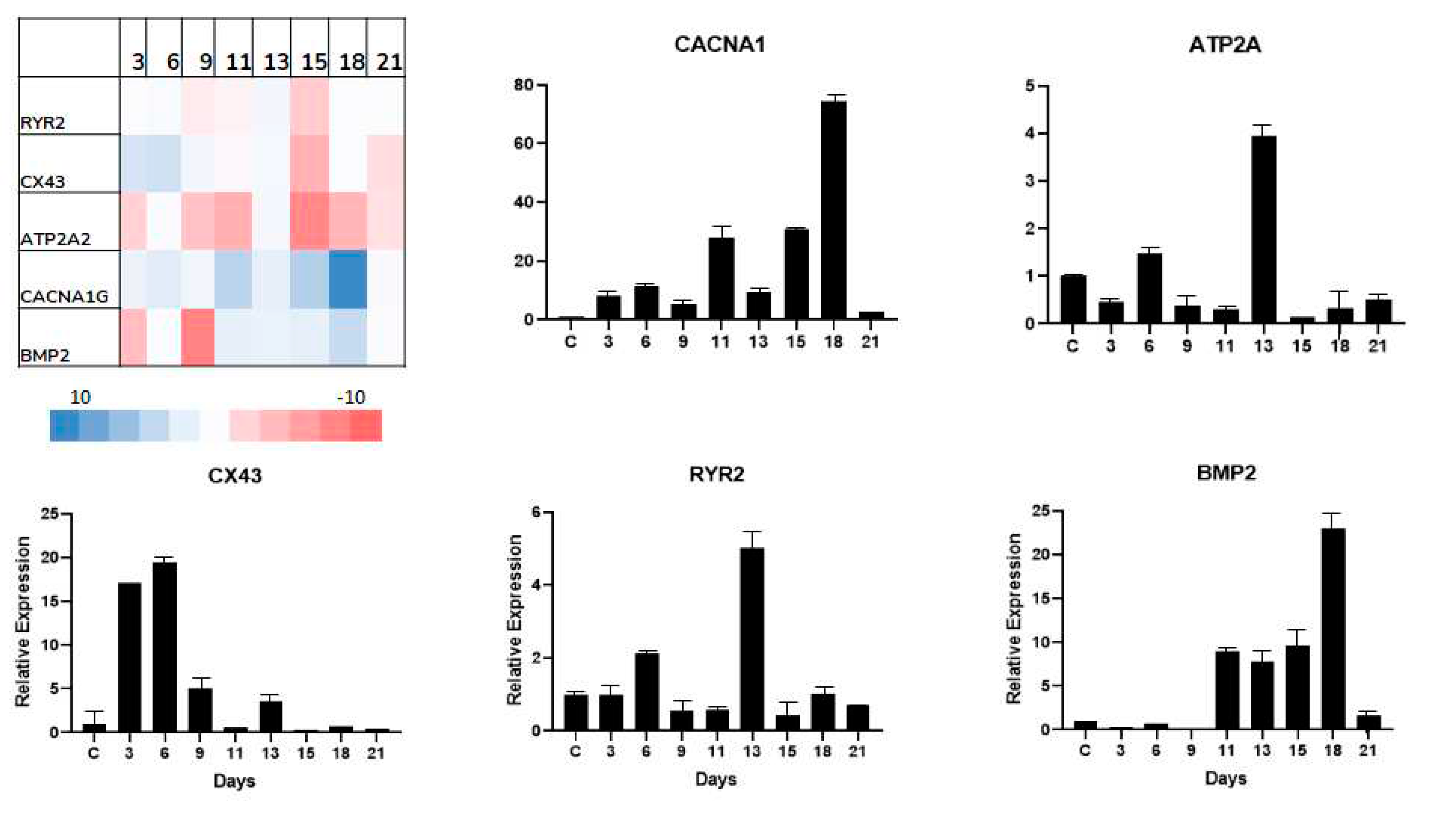

Table 1.
Krüppel-like factor activation and function related to characteristic members and their potential effects.
Table 1.
Krüppel-like factor activation and function related to characteristic members and their potential effects.
| Gene | Regulation over time | Max Expression Day | Relative Expression | Potential Effect |
|---|---|---|---|---|
| KLF-3 | Downregulated | 3 | 1.98x | Hypertrophy repressor |
| KLF-4 | Downregulated | 2 | 1.7x | Antiapoptotic |
| 3 | 2.8x | |||
| KLF-6 | Upregulated | 3 | 2.4 | Profibrotic |
| KLF-11 | Upregulated | 5 | 2.96x | Unknown |
| 6 | 3.05x | |||
| 13 | 3x | |||
| KLF-12 | Upregulated | 16 | 7.2x | Proapoptotic |
| 18 | 9.7x | |||
| KLF-13 | Upregulated | 18 | 53x | Proapoptotic / Anti-inflammatory |
| KLF-15 | Upregulated | 8 | 3x | Hypertrophy modulation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



