
Submitted:
23 August 2023
Posted:
23 August 2023
You are already at the latest version
Abstract
Lung cancer accounts for the majority of cancer cases. In recent years, checkpoint inhibitor immunotherapy (ICI) has emerged as a new treatment. A better understanding of the tumor microenvironment, or TMJ, or the immune system surrounding the tumor is needed. Cytokines are small proteins that carry messages between cells and are known to play an important role in the body's response to inflammation and infection. Cytokines are important for immunity in lung cancer. It promotes tumor growth (oncogenic cytokines) or inhibits tumor growth (anti-tumor cytokines) by controlling signaling pathways for growth, proliferation, metastasis, and apoptosis. The immune system relies heavily on cytokines. They can also be produced in the laboratory for therapeutic use. Cytokine therapy helps the immune system stop the growth or kill cancer cells. Interleukins and interferons are two types of cytokines used to treat cancer. This article begins by addressing the role of the tumor microenvironment (TMJ) and its components in lung cancer. This review also highlights the functions of various cytokines such as IL, TGF and TNF.
Keywords:
Lung cancer
; Cytokines
; Therapy
; Immune system
; Anti-tumor
1. Introduction
Lung carcinoma, also known as lung cancer, is a type of lung cancer that usually results from uncontrolled cell proliferation of the lungs. Tissue containing epithelial cells that have transformed into malignant cells, or in some cases all epithelial cells, is one type of lung cancer. Abnormal masses or nodules may be seen on chest X-rays [1]. CT scans can also see small lung lesions that X-rays may miss.
Lung cancer is currently the second leading cause of cancer worldwide. Although lung cancer can occur in people who have never smoked, smokers are the group most likely to develop lung cancer. Both the amount and frequency of smoking can affect a person's risk of developing lung cancer. People who quit smoking after smoking for a long time will have a decreased risk of lung cancer. In addition, the risk of lung cancer increases with urban pollution.
To reduce the number of deaths from lung cancer, the best option is to increase the effectiveness of cancer prevention and to use screening strategies for risk assessment and detection in the early stages of lung cancer treatment [2]. According to the study of R. Kuner et al., the estimated number of new cases and deaths from lung and lung cancer in the United States in 2013 was 228,190 and 159,480, respectively. About 55% to 60% of patients have distant metastases and are diagnosed at an advanced, incurable stage. Therefore, the five-year overall survival rate of each stage is between 13% and 16%.
Nonsmall cell lung cancer, also known as NSCLC, is the most common type of bronchial malignancy. Adenocarcinoma and squamous cell carcinoma are the two main histological groups from which they are often separated. Adenocarcinoma and squamous carcinoma cells differ in DNA copy number, DNA methylation, gene mutations, transcriptome, proteome, and potential biomarkers.
In lung cancer, cytokines, or proteins, are important to support the immune system. It can promote tumor growth (oncogenic cytokines) or inhibit tumor growth (anti-tumor cytokines) by modulating related factors such as growth, proliferation, metastasis and apoptosis.
1.1. Type and treatment of lung cancer
This is divided into small cell lung cancer (SCLC), which accounts for 15% of patients, and non-small cell lung cancer (NSCLC), which accounts for 85% of patients [70]. Histologically, NSCLC is divided into three subtypes: adenocarcinoma (ADC), large cell lung cancer (LCC), and squamous cell carcinoma (SCC). Both smokers and nonsmokers have been found to have NSCLC, the most common variety that is gender-neutral and affects 40% of ADC victims. ADC consists of type II alveolar cells in the outer lung, which are mucus sealing cells [9].
Other subtypes of SCC occur in flattened squamous cells in the airways, centrally in the lungs, and account for approximately 25% to 30% of the most common causes of smoking [10].
Some asymptomatic cancer patients cannot be accurately diagnosed during conventional treatment. However, approximately 70% of NSCLC patients progress to an advanced stage after diagnosis, either locally or in organ metastases[11]. In other words, early warning signs may be the key to improving cancer patient survival. An important part of the immune system's specific response to dangerous stimuli is inflammation.Many studies have demonstrated the role of inflammation in cancer [12].
Due to the disease's complexity, it develops genetic and epigenetic variants. Tumor differentiation, growth, invasion, and metastasis are generally controlled by these modifications. Approximately 70% to 80% of patients do not believe that surgery, which is considered the best treatment for early-stage NSCLC, is the best option, especially because of locoregional tumor swelling, extrathoracic spread, or poor physical and functional state at the time of diagnosis. Therefore, patients often choose radiotherapy or chemotherapy alone to achieve a better prognosis [13].
Alternatively, surgical resection followed the neoadjuvant approach. Radiotherapy (RT) is important in the treatment of lung cancer. According to various studies, approximately 60% of patients received radiation therapy in the early stages of the disease – approximately 44% at diagnosis and approximately 16% in advanced or relapsed state[14]. Chemotherapy is considered a better option for lung cancer treatment because twothirds of patients have cancer or lung cancer at diagnosis [15]. Most FDA approved drugs focus on angiogenesis and immunosuppression to demonstrate the biological and physiological dynamics of the tumor microenvironment and cancer-related symptoms.
1.2. Tumor microenvironment and content: their contribution to NSCLC metastasis
Tumors creates the tumor microenvironment, which is dominated by interactions brought on by the tumor. The anti-tumor potential of different immune effector cells are down-regulated at the tumor site, although they are usually selected in response to signals from the tumor. The chronic infiltration of inflammatory cells seen in human malignancies is enriched with regulatory T cells (Treg) and myeloid suppressor cells (MSC). The lymphocytes, macrophages, dendritic cells and natural killer cells that make up the immune system communicate through cells, surfaces and other mediators (cytokines and chemokines). Innate immunity typically works ahead of schedule during the course of an insusceptible reaction and includes a specific configuration of receptors, with the help of macrophages, neutrophils, and NK cells.
T lymphocytes (CD4+ and CD8+ T cells) and B cells, on the other hand, regulate adaptive immunity. These cells are prepared by antigen-presenting dendritic cells; B cells are activated by antigen entering their local environment; and CD8+ and CD4+ lymphocytes are sensitive to antigen presented as peptides that complex with MHC class I and class II particles, respectively. Germinal gene sequences that produce a large population of T-cell and B-cell receptors can sensitively detect multiple targets, resulting in an immune system that takes days to survive and causes strong memory.
The immune response is determined by the cytokine profile transmitted by the immune system: cell mediated immunity is favored by TH1 cytokines such as IL-2, IFN- and tumor necrosis factor (TNF); TH2 cytokines (eg., IL-4, 5, 10 and 13) are important for immunity and well-being; In addition, TH17 cytokines such as IL-17, 22 and granulocyte sedimentation enhancing component (G-CSF) act to elicit responses. It indicates that effector and focused memory T helper type 1 (Th1) and T killer type 1 (Tc1) cells, rather than a humoral immune response, are responsible for defensive long-range resistance against cancer and the ability to combat chronic inflammation.
T cells or NK cells are members of the TNF receptor family (eg. For example, FAS and TRAIL) or performance and degranulation by granzymes, the ultimate result of cancer prevention may or may not be the end of foragers. Similarly, the latter results in the delivery of Th1 cytokines that transform tumor-associated M2 macrophages that secrete arginase and inducible nitric oxide synthase into M1 killer cells that secrete IL-12 and TNFα. In addition, antibodies are effective in inhibiting tumor growth when they bind to oncogenic growth receptors such as HER2/NEU and EGFR and prefer NK or Fc receptors that activate macrophage or complement protein cascades[16]. When cancer cells metastasize, they leave their original site and live elsewhere where they can continue to grow. The tumor and surrounding ME initiates the process of infiltrating, forming, and dividing into different tissues, leading to metastasis. The transformation of a normal cell into a tumor is dependent on the environment, so it may occur in one TME but not in another. TME contains bothcellular and non-cellular components (Figure 1).
Lung cancer has a complex process of growth, initiation and metastasis. Genetic abnormalities acquired by the tumor lead to the development of the disease and interaction with the immune system leading to localized ME. Immobility and flexibility in TME use different mechanisms. First, it is the main defense mechanism against foreign invaders and consists of dendritic cells (DC) (CD1c (+), natural killer (NK) cells (CD49a, CD69 and CD103), macrophages (CD68+), NK-T cells CD56+ and CD3+) and neutrophil phagocyte (NK) cells. Cancer cells promote tumor growth, angiogenesis and metastasis.
If the system is reprogrammed, they also cause tumors to grow. Tumor growth slows from the second phase, which includes B cells (CD20+) and two subsets of T cells, T helper cells (CD4+) and cytotoxic T cells (CD8+) [18]. Tumor infiltrating leukocytes (TILs), which promote anti-tumor responses, account for 67% of TME in lung tumors. This is followed by tumor-associated macrophages (TAMs), followed by small numbers of DC and NK cells [19].
2. Basic properties of cytokines
The balance of the immune system is controlled by cytokines, which are membrane-bound or released proteins that mediate intercellular signals. They are produced by innate and adaptive immune cells in response to tumor antigens and pathogens. Each cytokine has a unique effect on the immune system that is dependent on many factors, including local cytokine concentrations, cytokine receptor expression patterns, and the integration of various pathways into the immune response. The importance of cytokines in tumor immunity is demonstrated by increased tumor frequency in mice lacking type I or type II interferon (IFN) receptors or downstream IFN receptor signaling components [20].
An important aspect of cytokine signaling is pleiotropy, or the ability of a single cytokine to cause different cell types to produce different effects, some of which may lead to resistance (Table 1). This has been viewed as one of the main challenges of IL-2 therapy due to the dual role of IL-2 as a potent activator of the T regulatory and T effector regions. Another important aspect of cytokine signaling is redundancy, or the number of cytokines with the same function. This reactivity makes it difficult to change cytokines for therapy because changing one cytokine may induce the other to compensate.

Table 1.
Basic Characteristics of Cytokines.
| Cytokine | Primary Cell Source | Primary Target Cell | Biological Activity |
|---|---|---|---|
| IL-1 | Monocytes Macrophages Fibroblasts Epithelial cells Endothelial cells Astrocytes |
T cells B cells Endothelial cells Hypothalamus Liver |
Co-stimulation Cell activation Inflammation Fever Acute phase reactant |
| IL-2 | T cells NK cells |
T cells NK cells B cells Monocytes |
Cell growth Cell activation |
| IL-3 | T cells | Bone marrow progenitor cells | Cell growth and cell differentiation |
| IL-4 | T cells | T cells B cells |
Th2 differentiation Cell growth Cell activation IgE isotype switching |
| IL-5 | T cells | B cells Eosinophils |
Cell growth Cell activation |
| IL-6 | T cells Macrophages Fibroblasts |
T cells B cells Liver |
Co-stimulation Cell growth Cell activation Acute phase reactant |
| IL-7 | Fibroblasts Bone marrow stromal cells |
Immature lymphoid progenitors | T cell survival, proliferation, homeostasis B cell development |
| IL-8 | Macrophages Epithelial cells Platelets |
Neutrophils | Activation Chemotaxis |
| IL-10 | Th2 T cells | Macrophages T cells |
Inhibits antigen-presenting cells Inhibits cytokine production |
| IL-12 | Macrophages NK cells |
T cells | Th1 differentiation |
| IL-15 | Monocytes | T cells NK cells |
Cell growth Cell activation NK cell development Blocks apoptosis |
| IL-18 | Macrophages | T cells NK cells B cells |
Cell growth Cell activation Inflammation |
| IL-21 | CD4+ T cells NKT cells |
NK cells T cells B cells |
Cell growth/ activation Control of allergic responses and viral infections |
| IL-23 | Antigen-presenting cells | T cells NK cells DC |
Chronic inflammation Promotes Th17 cells |
| GM-CSF | Fibroblasts Mast cells T cells Macrophages Endothelial cells |
DC Macrophages NKT cells Bone marrow progenitor cells |
T cell homeostasis Promotes antigen presentation Hematopoietic cell growth factor |
| IFN-α | Plasmacytoid DC NK cells T cells B cells Macrophages Fibroblasts Endothelial cells Osteoblasts |
Macrophages NK cells |
Anti-viral Enhances MHC expression |
| IFN-γ | T cells NK cells NKT cells |
Monocytes Macrophages Endothelial Cells Tissue cells |
Cell growth/ activation Enhances MHC expression |
| TGF-β | T cells Macrophages |
T cells | Inhibits cell growth/activation |
| TNF- α | Macrophages T cells |
T cells B cells Endothelial cells Hypothalamus Liver |
Co-stimulation Cell activation Inflammation Fever Acute phase reactant |
Cytokines play complex and often antagonistic roles in immune maturation, host defense, and tumor immunobiology. Therefore, the development of cytokine-based immunotherapies for cancer treatment depends on knowing the biological activities and mechanisms of action of these agents.
Table 2.
The functions and duties of the cytokine-secreting cells in the tumor microenvironment.
| Cell type | Function in TME |
|---|---|
| Tumour-associated macrophages (TAMs) | TAMs exhibit the M2 macrophage phenotype, which includes protumorigenic characteristics, anti-inflammatory properties, and Th2 cytokine secretion. Help cancer cells invade secondary areas and promote angiogenesis. |
| Cancer-associated fibroblasts (CAFs) | Stromal cell populations that are active support the desmoplastic tumor microenvironment. By releasing cytokines, one can encourage angiogenesis and control tumor-promoting inflammation. |
| CD4+ Th cells | Th1 and Th2 lineages have been divided. Th1 secretes cytokines that are both proinflammatory and antitumorigenic, whereas Th2 secretes cytokines that are both proinflammatory and tumorigenic. |
| CD8+ Tc cells | Adaptive immune system effector cells that recognize and kill tumor cells by perforin-granzyme-mediated apoptosis. |
| Mast cells (MCs) | innate and adaptive immune responses to be produced and maintained. Release substances that encourage endothelial cell development to aid tumor cell angiogenesis. |
| B-cells | Modulators of humoral immunity and secrete cytokines. Alter Th1:Th2 ratio. |
| Natural killer (NK) cells | Without antigen presentation, cytotoxic lymphocytes obliterate stressed cells. Through "missing self" activation and "stress-induced" activation, detect and destroy tumor cells. |
| Dendritic cells (DCs) | Antigen-presenting cells (APCs) that control the immune system's adaptive response. They increase vascularization in the TME to encourage angiogenesis. |
| Neutrophils | N1-type cells are pro-inflammatory, anti-tumorigenic, and release Th1 cytokines. |
Depending on how they are affected, macrophages can be divided into two types: active M1 type and alternatively activate M2 type. The M1 type secretes Th1 cytokines with pro-inflammatory and anti-tumorigenic properties, while the M2 type secretes Th2 cytokines with anti-inflammatory and pro-tumorigenic properties. Tumor grade and stage correlate with the Th1:Th2 ratio
2.1. Classification of cytokines and their receptors
In order to better define cytokines, many partners and receptors have been identified that define the cytokine problem. To date, seven families of cytokine receptors have been identified (Table 3) : type I and type II cytokine receptors, immunoglobulin superfamily receptors, tumor necrosis factor (TNF) receptors, G protein-coupled receptors, transforming growth factor beta (TGF-β), and recently discovered IL -17 receptor. As these hold immediate clinical promise, this section will focus on cytokines that signal through the type I and type II cytokine receptor families.
Table 3.
Classification of cytokine receptors.
| Receptor Family | Ligands | Structure and Function |
|---|---|---|
| Type I Cytokine Receptors | IL-2 IL-3 Il-4 IL-5 IL-6 IL-7 IL-9 IL-11 IL-12 IL-13 IL-15 IL-21 IL-23 IL-27 Erythropoietin GM-CSF G-CSF Growth hormone Prolactin Oncostatin M Leukemia inhibitory factor |
Composed of multimeric chains. Signals through JAK-STAT pathway using common signaling chain. Contains cytokine binding chains. |
| Type II Cytokine Receptors | IFN-α/β IFN-γ IL-10 IL-20 IL-22 IL-28 |
Immunoglobulin-like domains. Uses heterodimer and multimeric chains. Signals through JAK-STAT. |
| Immunoglobulin Superfamily Receptors | IL-1 CSF1 c-kit IL-18 |
Shares homology with immunoglobulin structures. |
| IL-17 Receptor | IL-17 IL-17B IL-17C IL-17D IL-17E IL-17F |
|
| G Protein-Coupled Receptors (GPCR) | IL-8 CC chemokines CXC chemokine |
Functions to mediate cell activation and migration. |
| TGF-β receptors 1/2 | TGF-β | |
| Tumor Necrosis Factor Receptors (TNFR) | CD27 CD30 CD40 CD120 Lymphotoxin-β |
Functions as co-stimulatory and co-inhibitory receptors. |
2.1.1. Type I cytokine receptors
Type I cytokine receptors, including IL-2, IL-4, IL-7, IL-9, IL-15, and IL-21 receptors, common chain (c), this chain contains specific Janus kinases ( The cytokine component of JAK) 1 and 3 that initiates intracellular signaling via coordination of signal transduction and activating T (STAT) molecules (Figure 3). Other type I cytokine receptor subsets include the IL-6 and GM-CSF receptor families, which cooperate through the gp130 receptor to influence multiple signaling pathways at their targets[21].
Various receptor complexes such as IL-6, IL-11, leukemia inhibitory factor (LIF), oncostatin M, cardiotrophin-1, and ciliary neurotrophic factor use gp130 signaling components to influence immunological, hematopoietic excess activity, and pleiotropic effects[22]. Similarly, receptors of the distinct GM-CSF receptor subfamily, which has a common β chain that binds with the cytokine-specific chain, also recognize IL-3, IL-5, and GM-CSF [23].
2.1.2. Type II cytokine receptors
By use of Type II cytokine receptors, which are made up of a signaling chain and a ligand-binding chain, IFN-α, IFN-β, IFN-γ, and IL-10 actions are mediated. The intracellular domains of type II cytokine receptors frequently interact with Janus kinase (JAK) family tyrosine kinases whose sequences resemble tandem Ig-like domains [24].
2.1.3. Immunoglobulin superfamily receptors
IL-1, IL-18, stem cell factor and monocyte colony stimulating factor receptors are members of the immunoglobulin superfamily and have extracellular immunoglobulin domains.
3. The role of cytokines in immunotherapy and cancer
Since lung cancer outperforms all currently used treatment modalities, it's important to consider some options, especially since the immune system is an important organ of cancer prevention. TILs (immune cells) of the TME, particularly lymphocytes and macrophages, produce cytokines (<30 kDa) that regulate various cellular functions, and high molecular weight nonstructural proteins called chemokines. This process must interact with specific cellular receptors that have an effect on the body through autocrine, paracrine and endocrine types of action. These include metabolism, growth, cell and tissue repair, and chemotaxis [25]. However, cytokines and chemokines are recognized to play an important role in local inflammation and infection.
In addition to treatment, these factors also play an important role in TME for cancer and infection. Considering their role as biomarkers, cytokines (IL-6, 10, 17, 27, 35; TNF-; IFN-; and TGF-) and chemokines (CCL-2, 5, 18; CCR-4; CXCR -4 ; CX3CL) -1; and CXCL-1, 5, 8, 13) are frequently targeted for the treatment of lung cancer[26]. Among other stromal cell types in theTME, macrophages are the major secretors of the cytokine IL-6. When it binds to IL-6R (ligand-binding receptor), its hormonal effects (autocrine and paracrine) are triggered. Its role in TME requires reconciliation of many responses.
It promotes apoptosis, invasion, angiogenesis, EMT and metastasis through the immune system. According to recent research, it also transplants tumor cells into new areas, such as tumor cells containing T lymphocytes [27]. T cells can switch tumors from one state to another, for example from a suppressed state to a reactive state, and also stop the growth and spread of tumors. In addition to innate immunity, TAM-based IL-6 supports the development and repair of cancer stem cells (CSCs). Phosphatidylinositol 4,5-biphosphate 3-kinase/threonine kinase 1/AKT serine (PI3K/AKT) signaling is known to be activated by IL-6 to promote the growth of the A549 brain cancer cell line.
A recent study using in vivo mouse and in vitro human lung tumor models demonstrated that the mechanism by which IL-6 stimulates CCL-2/5 causes EMT and resistance in different stages of lung cancer [26]. Blocking IL-6 modifies TME, complicating the as yet undiagnosed model of carcinogenesis. A number of methods targeting IL-6 signaling have been explored, including blocking STAT3 stimulation with antibodies and peptides, phosphorylating Janus kinase (JAK), and using the IL-6, IL-6R, and IL-6-sIL-6R complex. Body [28]. CAFs are important in NSCLC as they enhance EMT signaling to regulate the immune response. IL-10 activity is known to inhibit IL-6 production.
TNF- has been termed a "pyrogenic cytokine" because of its tendency to proliferate in a harsh environment and in response to infection, causing tumor necrosis and cytotoxicity. It binds to its target cells using one of the two receptor families (TNFR1 or TNFR2). According to studies using cancer cells in vivo and in vitro, TNF-- like IL-6 stimulates the process. These include metastasis, angiogenesis, invasion, and anti-apoptosis. Doxorubicin treatment produces TNF-α, while cyclin-dependent kinase inhibitor 1A (CDKN1A) is down-regulated.
This then induces apoptosis in lung tumors lacking TP53. It affects the communication between TNF-TAM and other cells in the TME and controls the survival and growth pathway as well as causing death by TNFR1. Although TNF-- is antitumorigenic because it slows the growth of tumors, its side effects are not normal. Therefore, some studies warn against altering TNF-α, while other studies strongly increase acceptance [29].
The normal regulation of Th1/Th2 T helper cell development was disrupted in 2005 with the identification of T helper 17 (Th17) cells as a third subset of T helper cells.
Th17 cells differ from other T cells in that they produce IL-17, express specific mutations and fulfill specific biological roles. TGF- and IL-6 are required in some cytokine cocktails to differentiate mouse Th17 cells. Additionally, IL-6 promotes the production of IL-21, which helps TGF and IL-23 promote the development of Th17 cells in mice. IL-1 is essential for and promotes the early development of Th17 cells in mice. IL-1 is required for the differentiation of human Th17 cells and together IL-1, IL-6 and IL-23 form a good cytokine environment produced by human Th17 cells.
In addition to cytokine regulation, Th17 development is also influenced by molecular programs of transcriptional regulation. Various, including Th17 cell growth (HIF1-a), signal transducer and activator of transcription 3 (Stat3), retinoid-associated orphan receptor gt (ROR-gt), nuclear receptor ROR-a, IFN-regulated Factor 4 (IRF-4) depends on proteins. . ), B cell activating transcription factor (B-ATF) and hypoxia-inducible factor 1 [30].
There is increasing evidence of an association between chronic disease, chronic pain, and cancer. Many immune cells, such as Ab T cells, gd T cells, and natural killer (NK) T cells, can play an important role in tumor prevention and are attracted to local inflammation in the tumor microenvironment. Th17 cells are known to play an important role in the immune system, so they must be present in the tumor microenvironment.
Although Th17 cells are abundant in the tumor microenvironment, it is unclear how they contribute to tumor protection. Most studies investigating the relationship between Th17 cells and cancer have used animal models with varying degrees of success [30].
The IL-12 family of immunosuppressive cytokines includes IL-35. It is the same subunit commonly expressed by regulatory T cells (Treg). STAT1 and STAT4 mediate signaling through IL-35. It does this by binding to the IL-35R receptor. IL-35 inhibits T cell proliferation and effector activity.
Several studies have shown that Tregs induced by IL-35 in TME inhibit CD4 and CD8 function and NK cell antitumor activity. To achieve this, it increases IL-10 and TGF- production. In addition to these results, it has been reported that high plasma IL-35 levels are associated with infection in cancer patients. Potential therapeutic targets include the use of certain monoclonal antibodies to suppress the immune response to IL-35 [31]. B and T cells and activated macrophages release IL-10, a cytokine with protective properties.
It works by binding to the IL-10R receptor. Its main function is to prevent the activation of macrophages. It inhibits the production of proinflammatory cytokines such as IL-1, IL-6, IL-8, IL-12, TNF- and granulocyte-macrophage colony stimulating factor (GM-CSF) [32]. It also reduces the expression of major histocompatibility complex-II (MHC-II) by inhibiting antigen presentation in active macrophages. In addition, IFN- (interferon-) production of Th1 and NK cells is inhibited by IL-10.
Serum IL-10 uptake can be seen clearly in many diseases, especially in the immune system. In cancer patients, TAM-(M2-)-dependent IL-10 has consistently shown significant clinical significance. While both IL-10 and TAM-based IL-10 have been studied and show some similarities, the signal of the former involves a complex chemical process involving 76 processes and 37 molecules (less extremes) to promote tumor growth [33]. IL-10 from TAMs preferentially promotes lung cancer stalk. In vivo cancer mouse model studies use the NFKB/JAK1/STAT1/NOTCH1 signaling pathway. It also promotes the spread of lung cancer throughout the body. It targets the induction of the CCL-2/CCR-2 and CXC3CL-1/CX3CR-1 axes in the interaction between macrophages and tumor cells. Not only is this IL-10 signaling issue being studied, but many other methods are also being evaluated at the clinical level. Although IL-10 has been extensively studied, its role in cancer therapy remains unclear. Different strategies, such as the development of monoclonal antibodies and peptides that block IL-10, have been tested in various diseases.
In addition, efforts are being made to develop molecular inhibitors and receptor blocking (IL-10R) strategies that target JAK/STAT3 signaling [34]. Several results indicate that IL-10+ Bregs (regulatory B cells) suppress human lung cancer, particularly in ADCs. They have been shown to be involved in cancer. In addition, IL-10 is thought to be important for maintaining the homeostasis of inflammation-fighting regulatory T cells (Treg). In addition, it is important for the suppression of proinflammatory T cells (Th17) that express the IL-17 gene [64].
Control of tumor growth depends on different T cell populations that maintain a balance between pro-inflammatory and anti-inflammatory factors [35]. Despite its many functions, IL-10's ability to downregulate MHC I renders tumor cells more susceptible to NK. This indicates that IL-10 works in concert with excitatory cytokines such as IL-2, which can increase NK cell-mediated responses to tumor cells. However, conflicting data on the role of IL-10 in cancer are beginning to emerge. According to Konjevi et al.
A plausible explanation for why its function is not regulated by environmental stimuli is cytokine homeostasis [36].
Myeloid-derived suppressor cells (MDSCs) support peripheral immunity by acting as anti-inflammatory agents. Because of its complexity, the role of IL-10 in preventing cancer has been controversial since its discovery in the 1990s. While decreased IL-10 levels have been shown to increase the risk of gastric adenocarcinoma and prostate cancer, it is known to increase tumor growth in various types of cancer (HIV-positive cancer). According to research by Lee and colleagues, IL-10 slows tumor growth by blocking the effects of the IL-6/STAT3 axis on MDSCs.
Among T lymphocytes, IL-10 is thought to have both stimulating and inhibitory effects. According to Fujii et al.'s research, administration of IL-10 immediately after vaccination can enhance immunity and increase the effectiveness of vaccination. Splenocyte-based transfection studies showed that IL-10 can better regulate CTL activity by reducing CD+ T lymphocytes. This finding suggests that IL-10 has a negative effect on immune and immune-promoting CD4+ and CD8+ T lymphocytes, respectively.
The secondary function of IL-10 suggests that it could be used in the development of increasingly complex anti-inflammatory drugs [37]. Several studies have shown impaired IL-10 signaling in young cancer patients, and other studies have shown that IL-10 mutant animals exhibit an increased risk of cancer. These studies highlight the impact of IL-10 deficiency on the perception of tumor-promoting inflammation and the importance of IL-10 in the control of inflammation [38].
IL-27, another anti-inflammatory cytokine, is recognized for its ability to reduce tumor growth. This immunostimulatory two-chain cytokine, together with IL-12, promotes CD4+ T cell proliferation, Th1 cell differentiation and IFN- production. It has pleiotropic effects, including EB13 and IL-27p28 subunits. IL-27 mainly activates STAT1 and STAT3, gp130 chain and IL-27R.
It slows the growth of NSCLC while promoting apoptosis. In xenograft models, in addition to downregulatory and EMT-related genes, IL-27 forced intratumoral myeloid cells to show anti-tumor activity [39]. The COX2 inhibitor called apricotoxib combined with IL-27 prevents EMT in NSCLC cells in a STAT1-dependent manner. One study showed that IL-27 inhibits proliferation, migration and invasion of NSCLC cells. Another anti-cytokine is IL-37 with 5 variants (IL-37a to IL-37e).
IL-37 is a member of the IL-1 family. The second type, IL-37b, is the most studied isoform as it is widely expressed in many human tissues and brain tumors. It has been reported to reduce pain and fatigue. It is known that proinflammatory factors such as IL-1/, IL-6 and TNF- are inhibited by it. The mechanisms underlying IL-37 activity are thought to be modulation of the IL-6/STAT3 pathway, inhibition of β-catenin, inhibition of the pSmad3C/P21 tumor signaling pathway, and recruitment of CD57+ NK cells[40].
M2 macrophages release tumor growth factor (TGF-), another anti-inflammatory cytokine that promotes angiogenesis and metastasis. It uses type I and type II transmembrane serine/threonine kinase receptors to regulate its biological processes. These macrophages produce VEGF and COX-PGE2, as well as other mediators that promote tumor growth. IFN-, a cytokine that promotes inflammation, reactive nitrogen and oxygen intermediates, inducible nitric oxide synthase production, and NSCLC-associated MHC molecules secreted by another macrophage subtype called M1 [41]. The role of its signaling is pleiotropic.
In addition to immunity, these activities affect cell growth, differentiation, death, motility and invasion. TGF stimulates differentiation, induces immunosuppressive cells, and subsequently suppresses the activity of T cells and anergic NK cells in the TME. The main source of TGF- is also an anti-inflammatory agent. TGF-β can be used as an effective therapeutic agent to promote NK cell-mediated anti-tumor protection by modifying the TME, thereby promoting tumor growth and metastasis. TGF-β induces EMT in lung tumors using the JAK/STAT3 signaling pathway [42].
Another cytokine, IL-17, is dysregulated in many human diseases such as cancer and inflammation. IL-17A to IL-17F is a family of six cytokines with different homology and roles in the immune system. These cytokines bind to IL-17 receptors (IL-17R, IL-17RA to IL-17RE), which function as homodimeric or heterodimeric complexes. IL-17A, the oldest member of the IL-17 family, is produced by Th17 cells [63]. Its overproduction is associated with cancer as well as autoimmune and inflammatory diseases [43].
The IL-17B/IL-17RB pathway is a signal that promotes not only cancer survival but also growth and spread. Tumor-promoting activity of the IL-17B/IL-17RB pathway is unique and complex due to ongoing tumor processes and unexpected mechanisms that cause TME modification. Various in vitro and in vivo studies have demonstrated that IL-17B signaling is essential to support tumor health and growth. All studies have shown that the IL-17B/IL-17RB axis is blocked by inhibiting receptor expression in tumor cells, eliciting anti-IL-17RB antibodies, and restoring chemosensitivity in vitro and in vivo [77]. Surprisingly, IL-17RB signaling along the ERK/GSK-3 β/β-catenin pathway has been associated with EMT in lung cancer [95].
Both human cancer patients and mouse tumor models showed tumor-promoting therapeutic activity via IL-17 and Th17. The main role in the tumor-promoting activity of IL-17 or Th17 cells ultimately leading to the promotion of tumor growth is angiogenesis and cytokine activation in the tumor microenvironment. IL-17 has been shown to promote human cervical cancer in nude mice. This effect was associated with increased IL-6 and IL-8 levels and recruitment of macrophages to the tumor site. In addition, research using a mouse model of colon adenocarcinoma has shown that the anti-inflammatory effect of IL-17 is associated with the activation of various angiogenic factors from fibroblasts and tumor cells, including VEGF, PGE2, keratinocyte-derived factor. Talent. promotes tumor angiogenesis and nitric oxide.
According to the same research group, IL-17 promotes CXCR2-dependent angiogenesis in vivo by increasing net angiogenic activity and growth in human non-small cell lung cancer. In addition to its role in angiogenesis, IL-17 can also stimulate the synthesis of IL-6, which activates oncogenic Stat3 signaling and increases expression of genes that promote survival and angiogenesis [86].
4. Conclusions and future directions
Cytokines have been shown to be effective in the treatment of cancer, but it is not yet clear that some effective targets affect the different immune system. For cell populations that have been extensively studied, such as T cells, the labeling and role of surface receptors is well understood, and researchers use the same set of CD markers to identify the populations at hand using flow cytometry. However, the difficulty of detecting the receptor set increases with the level of the immune system being studied, making it difficult to compare results and identify patterns for a long time.
By analyzing changes in the activity and expression of surface markers of all different types of immune cells after immunotherapy, evaluation of treatment can be made easier and more consistent. This analysis can also use surface markers of the immune system in cancer patients to predict the effectiveness of the immune system.
It is important to examine the role and surface markers of the immune system in tumors and to pursue cytokine-based immunotherapy, but the main purpose of the article is to provide detailed information about the prospects of cancer cytokine therapy. One of the most important features of cytokines as modulators of the immune system is their pleiotropic effects. Each cytokine has a different effect on the immune system, enabling it to promote both pro-inflammatory and anti-tumor responses.
Therefore, for the future of cytokine-based cancer therapy, it will be important to develop a combination strategy that will enhance the anti-inflammatory effect while inhibiting the tumor-enhancing immune system. High doses of cytokines are required to induce a beneficial response in cancer patients, but doing so leads to many problems, including their short lifespan and toxicity (pro-inflammatory and autoimmune response). New technologies are coming in that improve the targeting of cytokines and alter their pharmacokinetics, such as cellular or other delivery systems based on transporters and drug transfer proteins Voluntarily helping to solve many of the shortcomings of cytokine therapy. According to the latest developments in cancer prevention, they will become the most important part of the treatment when used together with other drugs such as cytokines, anti-inflammatory drugs, oncolytics or as part of the immune response of DC and tumor cells.
Author’s contribution
All of the authors above have made major contributions to the development and writing of this review.
Data availability statement
Not Applicable.
Acknowledgments
Ministry of Science and Higher Education of the Russian Federation within the framework of state support for the creation and development of World-Class Research Centers "Digital biodesign and personalized healthcare" №075-15-2022-304.
Conflicts of Interest
There are no conflicts of interest that the authors can disclose.
References
- Pascal Bezel, Alan Valaperti, Urs Steiner, Dieter Scholtze, Stephan Wieser, Maya Vonow-Eisenring, Andrea Widmer, Benedikt Kowalski, Malcolm Kohler & Daniel P. Franzen. Evaluation of cytokines in the tumor microenvironment of lung cancer using bronchoalveolar lavage fluid analysis. Cancer Immunol. Immunother. 2021, 70, 1867–1876. [CrossRef] [PubMed]
- Tianxia Lan, Li Chen,and Xiawei Wei. Inflammatory Cytokines in Cancer: Comprehensive Understanding and Clinical Progress in Gene Therapy. Cells 2021, 10, 100. [CrossRef] [PubMed]
- Juliana B. Schilsky, Ai Ni, Linda Ahn, Sutirtha Datta, William D. Travis,c,d Mark G Kris, Jamie E Chaft, Natasha Rekhtman, and Matthew D. Hellmann. Prognostic impact of TTF-1 expression in patients with stage IV lung adenocarcinomas. Lung Cancer 2017, 108, 205–211. [CrossRef]
- N H C Au, A M Gown, M Cheang, D Huntsman, E Yorida, W M Elliott, J Flint, J English, C B Gilks, H L Grimes. P63 expression in lung carcinoma: A tissue microarray study of 408 cases. Appl Immunohistochem Mol Morphol. 2004, 12, 240–247. [CrossRef] [PubMed]
- F. Pasini, M.A. Bassetto, R. Sabbioni, G.L. Cetto, F. Scognammiglio, G. Pizzolo. 1187 Soluble CD30 (SCD30) serum levels in patients with embrional carcinoma (EC) or mixed germ cell tumors (GCT) with embrional component. Eur. J. Cancer 1995, 31 (Supplement 6), S248. [CrossRef]
- Purdue, M.P.; Lan, Q.; Langseth, H.; Grimsrud, T.K.; Hildesheim, A.; Rothman, N. Prediagnostic serum sCD27 and sCD30 in serial samples and risks of non-Hodgkin lymphoma subtypes. Int. J. Cancer 2019, 146, 3312–3319. [Google Scholar] [CrossRef]
- Lynn I. Levin, Elizabeth C. Breen, Brenda M. Birmann, Julie L. Batista, Larry I. Magpantay, Yuanzhang Li, Richard F. Ambinder, Nancy E. Mueller, Otoniel Martinez-Maza. Elevated Serum Levels of sCD30 and IL6 and Detectable IL10 Precede Classical Hodgkin Lymphoma Diagnosis. Cancer Epidemiology Biomarkers Prev. 2017, 26, 1114–1123. [CrossRef]
- John Wiley and Sons. Soluble B-cell activation marker of sCD27 and sCD30 and future risk of B-cell lymphomas: A nested case-control study and meta-analyses. 2016, 138, 2357–2367. [CrossRef]
- Lococo, F.; Paci, M.; Rapicetta, C.; Rossi, T.; Sancisi, V.; Braglia, L.; Cavuto, S.; Bisagni, A.; Bongarzone, I.; Noonan, D.M.; et al. Preliminary Evidence on the Diagnostic and Molecular Role of Circulating Soluble EGFR in Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2015, 16, 19612–19630. [Google Scholar] [CrossRef]
- Mariano Provencio, Manuel Cobo, Delvys Rodriguez-Abreu, Virginia Calvo, Enric Carcereny, Alexandra Cantero, Reyes Bernabé, Gretel Benitez, Rafael López Castro, Bartomeu Massutí, Edel del Barco, Rosario García Campelo, Maria Guirado, Carlos Camps, Ana Laura Ortega, Jose Luis González Larriba, Alfredo Sánchez, Joaquín Casal, M. Angeles Sala, Oscar Juan-Vidal, Joaquim Bosch-Barrera, Juana Oramas, Manuel Dómine, Jose Manuel Trigo, …Maria Torrente. Determination of essential biomarkers in lung cancer: A real-world data study in Spain with demographic, clinical, epidemiological and pathological characteristics. BMC Cancer 2022, 22, 732. [Google Scholar]
- T. Yamashita, H. Kamada, S. Kanasaki, Y. Maeda, K. Nagano, Y. Abe, M. Inoue1, Y. Yoshioka, Y. Tsutsumi, S. Katayama, M. Inoue, S. Tsunoda. Epidermal growth factor receptor localized to exosome membranes as a possible biomarker for lung cancer diagnosis. April 26, 2013. Shin-ichi Tsunoda, Ph.D, Laboratory of Biopharmaceutical Research, National Institute of Biomedical Innovation, 7-6-8 Saito-Asagi, Ibaraki, Osaka 567-0085, Japan. 26 April.
- Thomas Jostock, JuÈ rgen MuÈ llberg, Suat OÈ zbek, Raja Atreya, Guido Blinn, Nicole Voltz, Martina Fischer, Markus F. Neurath and Stefan Rose-John. Soluble gp130 is the natural inhibitor of soluble interleukin-6 receptor transsignaling responses. Eur. J. Biochem. 2001, 268, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, E.; Kuehn, J. Measurements of IL-6, soluble IL-6 receptor and soluble gp130 in sera of B-cell lymphoma patients. Does Viscum album treatment affect these parameters? BioMedicine 2002, 56, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Wang, H.; Shen, G.; Lin, D.; Lin, Y.; Ye, N.; Guo, Y.; Li, Q.; Deng, C.; Meng, C. Recombinant soluble gp130 protein reduces DEN-induced primary hepatocellular carcinoma in mice. Sci. Rep. 2016, 6, 24397. [Google Scholar] [CrossRef] [PubMed]
- Rachel Bolanos, Otoniel Martinez-Maza, Zuo-Feng Zhang, Shehnaz Hussain, Mary Sehl, Janet S. Sinsheimer, Gypsyarn D’Souza, Frank Jenkins, Steven Wolinsky, and Roger Detels. Decreased levels of the serum inflammatory biomarkers, sGP130, IL-6, sCRP and BAFF, are associated with increased likelihood of AIDS related Kaposi’s sarcoma in men who have sex with men. Cancer Res Front. 2018, 4, 45–59. [Google Scholar] [CrossRef]
- Yang, Z.-Z.; Grote, D.M.; Ziesmer, S.C.; Manske, M.K.; Witzig, T.E.; Novak, A.J.; Ansell, S.M. Soluble IL-2Rα facilitates IL-2–mediated immune responses and predicts reduced survival in follicular B-cell non-Hodgkin lymphoma. Blood 2011, 118, 2809–2820. [Google Scholar] [CrossRef]
- Gonda, K.; Horita, S.; Maejima, Y.; Takenoshita, S.; Shimomura, K. Soluble interleukin-2 receptor as a predictive and prognostic marker for patients with familial breast cancer. Sci. Prog. 2021, 104. [Google Scholar] [CrossRef]
- Bien, E.; Balcerska, A. Serum soluble interleukin 2 receptor α in human cancer of adults and children: A review. Biomarkers 2008, 13, 1–26. [Google Scholar] [CrossRef]
- Liang-Shun Wang; Kuan-Chih Chow; Wing-Yin Li; Chia-Chuan Liu; Yu-Chung Wu; Min-Hsiung Huang. Clinical Significance of Serum Soluble Interleukin 2 Receptor-α in Esophageal Squamous Cell Carcinoma. Clin Cancer Res 2000, 6, 1445–1451.
- Chalaris, A.; Garbers, C.; Rabe, B.; Rose-John, S.; Scheller, J. The soluble Interleukin 6 receptor: Generation and role in inflammation and cancer. Eur. J. Cell Biol. 2011, 90, 484–494. [Google Scholar] [CrossRef]
- Okugawa, Y.; Miki, C.; Toiyama, Y.; Yasuda, H.; Yokoe, T.; Saigusa, S.; Hiro, J.; Tanaka, K.; Inoue, Y.; Kusunoki, M. Loss of tumoral expression of soluble IL-6 receptor is associated with disease progression in colorectal cancer. Br. J. Cancer 2010, 103, 787–795. [Google Scholar] [CrossRef]
- Isobe, A.; Sawada, K.; Kinose, Y.; Ohyagi-Hara, C.; Nakatsuka, E.; Makino, H.; Ogura, T.; Mizuno, T.; Suzuki, N.; Morii, E.; et al. Interleukin 6 Receptor Is an Independent Prognostic Factor and a Potential Therapeutic Target of Ovarian Cancer. PLoS ONE 2015, 10, e0118080. [Google Scholar] [CrossRef]
- Petra Tesarová 1, Marta Kalousová, Marie Jáchymová, Oto Mestek, Lubos Petruzelka, Tomás Zima. Receptor for advanced glycation end products (RAGE)--soluble form (sRAGE) and gene polymorphisms in patients with breast cancer. Cancer Invest. 2007, 25, 720–725. [CrossRef] [PubMed]
- Galvani, V.; Hartman, K.P.; Rupreht, R.R.; Novaković, S.; Štabuc, B.; Ocvirk, J.; Menart, V.; Porekar, V.G.; Štalc, A.; Rožman, P.; et al. Soluble tumor necrosis factor receptor I (sTNFRI) as a prognostic factor in melanoma patients in Slovene population. 2000, 440 (5 Suppl), R061–R063. [Google Scholar] [CrossRef] [PubMed]
- Selinsky, C.L.; Howell, M.D. Soluble Tumor Necrosis Factor Receptor Type I Enhances Tumor Development and Persistence in Vivo. Cell. Immunol. 2000, 200, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Loa, C.C.; Yan, W.; Zhou, J.; Wheeler, S.; Zhang, H.; Zondlo, S.; Chen, L. Abstract 4588: GLP validiation for quantitative determination of soluble tumor necrosis factor receptor I and II (sTNFRI and sTNFRII) in human serum utilizing a highly sensitive ELISA. Cancer Res 2010, 70 (8_Supplement), 4588. [Google Scholar] [CrossRef]
- .
- Fuqian Yang Zhonghua Zhao Nana Zhao. Clinical implications of tumor necrosis factor receptor 2 in breast cancer. 2017, 2393–2398.
- Shiels, M.S.; Katki, H.A.; Hildesheim, A.; Pfeiffer, R.M.; Engels, E.A.; Williams, M.; Kemp, T.J.; Caporaso, N.E.; Pinto, L.A.; Chaturvedi, A.K. Circulating Inflammation Markers, Risk of Lung Cancer, and Utility for Risk Stratification. 2015, djv199. [Google Scholar] [CrossRef]
- Bastard, J.-P.; Fellahi, S.; Audureau, É.; Layese, R.; Roudot-Thoraval, F.; Cagnot, C.; Mahuas-Bourcier, V.; Sutton, A.; Ziol, M.; Capeau, J.; et al. Elevated adiponectin and sTNFRII serum levels can predict progression to hepatocellular carcinoma in patients with compensated HCV1 cirrhosis. Eur. Cytokine Netw. 2018, 29, 112–120. [Google Scholar] [CrossRef]
- Wu, F.T.H.; Stefanini, M.O.; Mac Gabhann, F.; Kontos, C.D.; Annex, B.H.; Popel, A.S. A systems biology perspective on sVEGFR1: Its biological function, pathogenic role and therapeutic use. J. Cell. Mol. Med. 2010, 14, 528–552. [Google Scholar] [CrossRef]
- Masakazu Toi, Hiroko Bando, Taeko Ogawa, Mariko Muta, Carsten Hornig, Herbert A Weich. Significance of vascular endothelial growth factor (VEGF)/soluble VEGF receptor-1 relationship in breast cancer. Int J Cancer. 2002, 98, 14–18. [Google Scholar] [CrossRef]
- Yilmaztepe, A.; Ulukaya, E.; Zik, B.; Yagci, A.; Sevimli, A.; Yilmaz, M.; Erdogan, B.B.; Koc, M.; Akgoz, S.; Karadag, M.; et al. Soluble Vascular Endothelial Growth Factor Receptor-1 (sVEGFR-1) is Decreased in Lung Cancer Patients Showing Progression: A Pilot Study. 2009, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Lamszus, K.; Ulbricht, U.; Matschke, J.; A Brockmann, M.; Fillbrandt, R.; Westphal, M. Levels of soluble vascular endothelial growth factor (VEGF) receptor 1 in astrocytic tumors and its relation to malignancy, vascularity, and VEGF-A. Clin. Cancer Res. 2003, 9, 1399–1405. [Google Scholar] [PubMed]
- Harris, H.; Wolk, A.; Larsson, A.; Vasson, M.-P.; Basu, S. Soluble vascular endothelial growth factor receptors 2 (sVEGFR-2) and 3 (sVEGFR-3) and breast cancer risk in the Swedish Mammography Cohort. Int. J. Mol. Epidemiol. Genet. 2016, 7, 81–86. [Google Scholar] [PubMed]
- Ebos, J.M.; Lee, C.R.; Bogdanovic, E.; Alami, J.; Van Slyke, P.; Francia, G.; Xu, P.; Mutsaers, A.J.; Dumont, D.J.; Kerbel, R.S. Vascular Endothelial Growth Factor–Mediated Decrease in Plasma Soluble Vascular Endothelial Growth Factor Receptor-2 Levels as a Surrogate Biomarker for Tumor Growth. Cancer Res 2008, 68, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Kanefendt, F.; Lindauer, A.; Mross, K.; Fuhr, U.; Jaehde, U. Determination of soluble vascular endothelial growth factor receptor 3 (sVEGFR-3) in plasma as pharmacodynamic biomarker. J. Pharm. Biomed. Anal. 2012, 70, 485–491. [Google Scholar] [CrossRef]
- Shibata, M.; Shibata, E.; Tanaka, Y.; Shiraoka, C.; Kondo, Y. Soluble Vegfr3 gene therapy suppresses multi-organ metastasis in a mouse mammary cancer model. Cancer Sci. 2020, 111, 2837–2849. [Google Scholar] [CrossRef]
- Singh, N.; Tiem, M.; Watkins, R.; Cho, Y.K.; Wang, Y.; Olsen, T.; Uehara, H.; Mamalis, C.; Luo, L.; Oakey, Z.; et al. Soluble vascular endothelial growth factor receptor 3 is essential for corneal alymphaticity. Blood 2013, 121, 4242–4249. [Google Scholar] [CrossRef]
- Mendelsohn, J.; Baselga, J. The EGF receptor family as targets for cancer therapy. Oncogene 2000, 19, 6550–6565. [Google Scholar] [CrossRef]
- Mohamed R Akl, Poonam Nagpal, Nehad M Ayoub, Betty Tai, Sathyen A Prabhu, Catherine M Capac, Matthew Gliksman, Andre Goy, K Stephen Suh.
- Mohamed R Akl, Poonam Nagpal, Nehad M Ayoub, Betty Tai, Sathyen A Prabhu, Catherine M Capac, Matthew Gliksman, Andre Goy, K Stephen Suh. Molecular and clinical significance of fibroblast growth factor 2 (FGF2 /bFGF) in malignancies of solid and hematological cancers for personalized therapies. Oncotarget. 2016, 7, 44735–44762. [Google Scholar] [CrossRef]
- Li, Y.; Guo, X.; Wang, J.S.; Wang, H.; Li, L. Function of fibroblast growth factor 2 in gastric cancer occurrence and prognosis. Mol. Med. Rep. 2019, 21, 575–582. [Google Scholar] [CrossRef]
- Dick Wågsäter, Sture Löfgren, Anders Hugander, Olaf Dienus & Jan Dimberg. Analysis of single nucleotide polymorphism in the promoter and protein expression of the chemokine Eotaxin-1 in colorectal cancer patients. 84 (2007).
- Rusch, V.; Klimstra, D.; Venkatraman, E.; Pisters, P.W.; Langenfeld, J.; Dmitrovsky, E. Overexpression of the epidermal growth factor receptor and its ligand transforming growth factor alpha is frequent in resectable non-small cell lung cancer but does not predict tumor progression. Clin. Cancer Res. 1997, 3, 515–522. [Google Scholar] [PubMed]
- F. Ciardiello, N. Kim, M. L. McGeady, D. S. Liscia, T. Saeki, C. Bianco & D. S. Salomon. Expression of transforming growth factor alpha (TGFa) in breast cancer. Anals of Oncology 1991, 2, 169–182. [Google Scholar]
- Liu, L.; Liu, Y.; Yan, X.; Zhou, C.; Xiong, X. The role of granulocyte colony-stimulating factor in breast cancer development: A review. Mol. Med. Rep. 2020, 21, 2019–2029. [Google Scholar] [CrossRef] [PubMed]
- Mouchemore, K.A.; Anderson, R.L. Immunomodulatory effects of G-CSF in cancer: Therapeutic implications. Semin. Immunol. 2021, 54, 101512. [Google Scholar] [CrossRef]
- Morris, K.T.; Khan, H.; Ahmad, A.; Weston, L.L.; Nofchissey, R.A.; Pinchuk, I.V.; Beswick, E.J. G-CSF and G-CSFR are highly expressed in human gastric and colon cancers and promote carcinoma cell proliferation and migration. Br. J. Cancer 2014, 110, 1211–1220. [Google Scholar] [CrossRef]
- Ohri, N.; Halmos, B.; Garg, M.; Levsky, J.M.; Cheng, H.; Gucalp, R.A.; Bodner, W.R.; Kabarriti, R.; Berkowitz, A.; Yellin, M.J.; et al. FLT3 ligand (CDX-301) and stereotactic radiotherapy for advanced non-small cell lung cancer. J. Clin. Oncol. 2020, 38 (15_suppl), 9618. [Google Scholar] [CrossRef]
- Drings, P.; Fischer, J.R. Biology and clinical use of GM-CSF in lung cancer. Lung 1990, 168, 1059–1068. [Google Scholar] [CrossRef]
- Hong, I.-S. Stimulatory versus suppressive effects of GM-CSF on tumor progression in multiple cancer types. Exp. Mol. Med. 2016, 48, e242. [Google Scholar] [CrossRef]
- Rong, Q.-X.; Wang, F.; Guo, Z.-X.; Hu, Y.; An, S.-N.; Luo, M.; Zhang, H.; Wu, S.-C.; Huang, H.-Q.; Fu, L.-W. GM-CSF mediates immune evasion via upregulation of PD-L1 expression in extranodal natural killer/T cell lymphoma. Mol. Cancer 2021, 20, 80. [Google Scholar] [CrossRef]
- Korbecki, J.; Simińska, D.; Kojder, K.; Grochans, S.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Fractalkine/CX3CL1 in Neoplastic Processes. Int. J. Mol. Sci. 2020, 21, 3723. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.; Erreni, M.; van Brakel, M.; Debets, R.; Allavena, P. Enhanced recruitment of genetically modified CX3CR1-positive human T cells into Fractalkine/CX3CL1 expressing tumors: Importance of the chemokine gradient. J. Immunother. Cancer 2016, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Liang, Y.; Chan, Q.; Jiang, L.; Dong, J. CX3CL1 promotes lung cancer cell migration and invasion via the Src/focal adhesion kinase signaling pathway. Oncol. Rep. 2019, 41, 1911–1917. [Google Scholar] [CrossRef]
- Jiang, G.; Wang, H.; Huang, D.; Wu, Y.; Ding, W.; Zhou, Q.; Ding, Q.; Zhang, N.; Na, R.; Xu, K. The Clinical Implications and Molecular Mechanism of CX3CL1 Expression in Urothelial Bladder Cancer. Front. Oncol. 2021, 11, 752860. [Google Scholar] [CrossRef] [PubMed]
- Erreni, M.; Siddiqui, I.; Marelli, G.; Grizzi, F.; Bianchi, P.; Morone, D.; Marchesi, F.; Celesti, G.; Pesce, S.; Doni, A.; et al. The Fractalkine-Receptor Axis Improves Human Colorectal Cancer Prognosis by Limiting Tumor Metastatic Dissemination. J. Immunol. 2016, 196, 902–914. [Google Scholar] [CrossRef]
- Hong, S.K.; Ko, D.W.; Park, J.; Kim, I.S.; Doo, S.H.; Yoon, C.Y.; Park, H.; Lee, W.K.; Kim, D.S.; Jeong, S.J.; et al. Alteration of Antithrombin III and D-dimer Levels in Clinically Localized Prostate Cancer. Korean J. Urol. 2010, 51, 25–29. [Google Scholar] [CrossRef]
- Polansky, M.; Varon, J.; Hoots, W. Use of antithrombin III in cancer patients with sepsis complicated with disseminated intravascular coagulopathy. Crit. Care 1998, 2, P024. [Google Scholar] [CrossRef]
- Paul, F.; Pellegrini, S.; Uzé, G. IFNA2: The prototypic human alpha interferon. Gene 2015, 567, 132–137. [Google Scholar] [CrossRef]
- Fimmel, S.; Devermann, L.; Herrmann, A.; Zouboulis, C. GRO-alpha: A Potential Marker for Cancer and Aging Silenced by RNA Interference. Ann. N. Y. Acad. Sci. 2007, 1119, 176–189. [Google Scholar] [CrossRef]
- Xiaxia Man, Xiaolin Yang, Zhentong Wei, Yuying Tan, Wanying Li, Hongjuan Jin & Baogang Wang. High expression level of CXCL1/GROα is linked to advanced stage and worse survival in uterine cervical cancer and facilitates tumor cell malignant processes. BMC Cancer 2022, 22, 712. [Google Scholar]
- Deok-Soo Son, Angelika K. Parl, Valerie Montgomery Rice & Dineo Khabele. Keratinocyte chemoattractant (KC)/human growthregulated oncogene (GRO) chemokines and proinflammatory chemokine networks in mouse and human ovarian epithelial cancer cells. Deok-Soo Son, Angelika K. Parl, Valerie Montgomery Rice & Dineo Khabele (2007) Keratinocyte chemoattractant (KC)/human growth-regulated oncogene (GRO) chemokines and pro-inflammatory chemokine networks in mouse and human ovarian epithelial cancer cells. Cancer Biol. Ther. 2007, 6, 1308–1318. [CrossRef] [PubMed]
- E Jablonska, J Jablonski, L Piotrowski, Z Grabowska. IL-1beta, IL-1Ra and sIL-1RII in the culture supernatants of PMN and PBMC and the serum levels in patients with inflammation and patients with cancer disease of the same location. Immunobiology 2001, 204, 508–516. [CrossRef] [PubMed]
- Park, S.Y.; Lee, J.G.; Kim, J.; Park, Y.; Lee, S.K.; Bae, M.K.; Lee, C.Y.; Kim, D.J.; Chung, K.Y. Preoperative serum CYFRA 21-1 level as a prognostic factor in surgically treated adenocarcinoma of lung. Lung Cancer 2013, 79, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xu, M.; Huang, H.; Jiang, X.; Gong, K.; Liu, Y.; Kuang, X.; Yang, X. Serum carcinoembryonic antigen elevation in benign lung diseases. Sci. Rep. 2021, 11, 19044. [Google Scholar] [CrossRef]
- Saad, H.M.; Tourky, G.F.; Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Khattab, A.M.; Elmasry, S.A.; Alsayegh, A.A.; Hakami, Z.H.; Alsulimani, A.; Sabatier, J.-M.; et al. The Potential Role of MUC16 (CA125) Biomarker in Lung Cancer: A Magic Biomarker but with Adversity. Diagnostics 2022, 12, 2985. [Google Scholar] [CrossRef]
- Ramachandran, S.; Verma, A.K.; Dev, K.; Goyal, Y.; Bhatt, D.; Alsahli, M.A.; Rahmani, A.H.; Almatroudi, A.; Almatroodi, S.A.; Alrumaihi, F.; et al. Role of Cytokines and Chemokines in NSCLC Immune Navigation and Proliferation. Oxidative Med. Cell. Longev. 2021, 2021, 5563746. [Google Scholar] [CrossRef]
- Couraud, S.; Zalcman, G.; Milleron, B.; Morin, F.; Souquet, P.-J. Lung cancer in never smokers – A review. Eur. J. Cancer 2012, 48, 1299–1311. [Google Scholar] [CrossRef]
- Thunnissen, E.; Noguchi, M.; Aisner, S.; Beasley, M.B.; Brambilla, E.; Chirieac, L.R.; Chung, J.-H.; Dacic, S.; Geisinger, K.R.; Hirsch, F.R.; et al. Reproducibility of Histopathological Diagnosis in Poorly Differentiated NSCLC: An International Multiobserver Study. J. Thorac. Oncol. 2014, 9, 1354–1362. [Google Scholar] [CrossRef]
- Kenfield, S.A.; Wei, E.K.; Rosner, B.A.; Glynn, R.J.; Stampfer, M.J.; Colditz, G.A. Burden of smoking on cause-specific mortality: Application to the Nurses' Health Study. Tob. Control. 2010, 19, 248–254. [Google Scholar] [CrossRef]
- Qian, F.; Yang, W.; Chen, Q.; Zhang, X.; Han, B. Screening for early stage lung cancer and its correlation with lung nodule detection. J. Thorac. Dis. 2018, 10, S846–S859. [Google Scholar] [CrossRef]
- Guo, J.; Wang, X.; Wang, Y.; Wang, L.; Hua, S. A promising role of interferon regulatory factor 5 as an early warning biomarker for the development of human non-small cell lung cancer. Lung Cancer 2019, 135, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Belderbos, J.; Sonke, J.J. State-of-the-art lung cancer radiation therapy. Expert Rev. Anticancer. Ther. 2009, 9, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Sunil, S.G.; Raviraj, R.; Nagarajan, D.; Zhao, W. Radiation-induced lung injury: Impact on macrophage dysregulation and lipid alteration–a review. Immunopharmacology and Immunotoxicology 2019, 41, 370–379. [Google Scholar]
- Tartour, E.; Zitvogel, L. Lung cancer: Potential targets for immunotherapy. Lancet Respir. Med. 2013, 1, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-S.; Liu, W.; Ly, D.; Xu, H.; Qu, L.; Zhang, L. Tumor-infiltrating B cells: Their role and application in anti-tumor immunity in lung cancer. Cell. Mol. Immunol. 2018, 16, 6–18. [Google Scholar] [CrossRef]
- Ramachandran, S.; Verma, A.K.; Dev, K.; Goyal, Y.; Bhatt, D.; Alsahli, M.A.; Rahmani, A.H.; Almatroudi, A.; Almatroodi, S.A.; Alrumaihi, F.; et al. Role of Cytokines and Chemokines in NSCLC Immune Navigation and Proliferation. Oxidative Med. Cell. Longev. 2021, 2021, 5563746. [Google Scholar] [CrossRef]
- Hermanns, H.M.; Radtke, S.; Haan, C.; de Leur, H.S.-V.; Tavernier, J.; Heinrich, P.C.; Behrmann, I. Contributions of Leukemia Inhibitory Factor Receptor and Oncostatin M Receptor to Signal Transduction in Heterodimeric Complexes with Glycoprotein 130. J. Immunol. 1999, 163, 6651–6658. [Google Scholar] [CrossRef]
- Sakamaki, K.; Miyajima, I.; Kitamura, T.; Miyajima, A. Critical cytoplasmic domains of the common beta subunit of the human GM-CSF, IL-3 and IL-5 receptors for growth signal transduction and tyrosine phosphorylation. EMBO J. 1992, 11, 3541–3549. [Google Scholar] [CrossRef]
- Kotenko, S.V.; Pestka, S. Jak-Stat signal transduction pathway through the eyes of cytokine class II receptor complexes. Oncogene 2000, 19, 2557–2565. [Google Scholar] [CrossRef]
- Poeta, V.M.; Massara, M.; Capucetti, A.; Bonecchi, R. Chemokines and Chemokine Receptors: New Targets for Cancer Immunotherapy. Front. Immunol. 2019, 10, 379. [Google Scholar] [CrossRef]
- Poeta, V.M.; Massara, M.; Capucetti, A.; Bonecchi, R. Chemokines and Chemokine Receptors: New Targets for Cancer Immunotherapy. Front. Immunol. 2019, 10, 379. [Google Scholar] [CrossRef] [PubMed]
- Karin, N. Chemokines and cancer: New immune checkpoints for cancer therapy. Curr. Opin. Immunol. 2018, 51, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Livergood, R.S.; Peng, G. The Role and Regulation of Human Th17 Cells in Tumor Immunity. Am. J. Pathol. 2012, 182, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Waugh, D.J.; Wilson, C. The Interleukin-8 Pathway in Cancer. Clin. Cancer Res. 2008, 14, 6735–6741. [Google Scholar] [CrossRef] [PubMed]
- Oft, M. IL-10: Master Switch from Tumor-Promoting Inflammation to Antitumor Immunity. Cancer Immunol. Res. 2014, 2, 194–199. [Google Scholar] [CrossRef]
- Soria, J.C.; Moon, C.; Kemp, B.L.; et al. Lack of interleukin-10 expression could predict poor outcome in patients with stage I non-small cell lung cancer. Clinical Cancer Research. 2003, 9, 1785–1791. [Google Scholar]
- Fujii, S.-I.; Shimizu, K.; Shimizu, T.; Lotze, M.T. Interleukin-10 promotes the maintenance of antitumor CD8+ T-cell effector function in situ. Blood 2001, 98, 2143–2151. [Google Scholar] [CrossRef]
- Ziblat, A.; Domaica, C.I.; Spallanzani, R.G.; Iraolagoitia, X.L.R.; Rossi, L.E.; Avila, D.E.; Torres, N.I.; Fuertes, M.B.; Zwirner, N.W. IL-27 stimulates human NK-cell effector functions and primes NK cells for IL-18 responsiveness. Eur. J. Immunol. 2015, 45, 192–202. [Google Scholar] [CrossRef]
- Banchereau, J.; Pascual, V.; O'Garra, A. From IL-2 to IL-37: The expanding spectrum of anti-inflammatory cytokines. Nat. Immunol. 2012, 13, 925–931. [Google Scholar] [CrossRef]
- Poeta, V.M.; Massara, M.; Capucetti, A.; Bonecchi, R. Chemokines and Chemokine Receptors: New Targets for Cancer Immunotherapy. Front. Immunol. 2019, 10, 379. [Google Scholar] [CrossRef]
- Liu, R.-Y.; Zeng, Y.; Lei, Z.; Wang, L.; Yang, H.; Liu, Z.; Zhao, J.; Zhang, H.-T. JAK/STAT3 signaling is required for TGF-β-induced epithelial-mesenchymal transition in lung cancer cells. Int. J. Oncol. 2014, 44, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.F.; Lee, Y.C.; Lo, S.; et al. A positive feedback loop of IL-17B-IL-17RB activates ERK/β-catenin to promote lung cancer metastasis. Cancer Letters. 2018, 422, 44–55. [Google Scholar] [CrossRef] [PubMed]
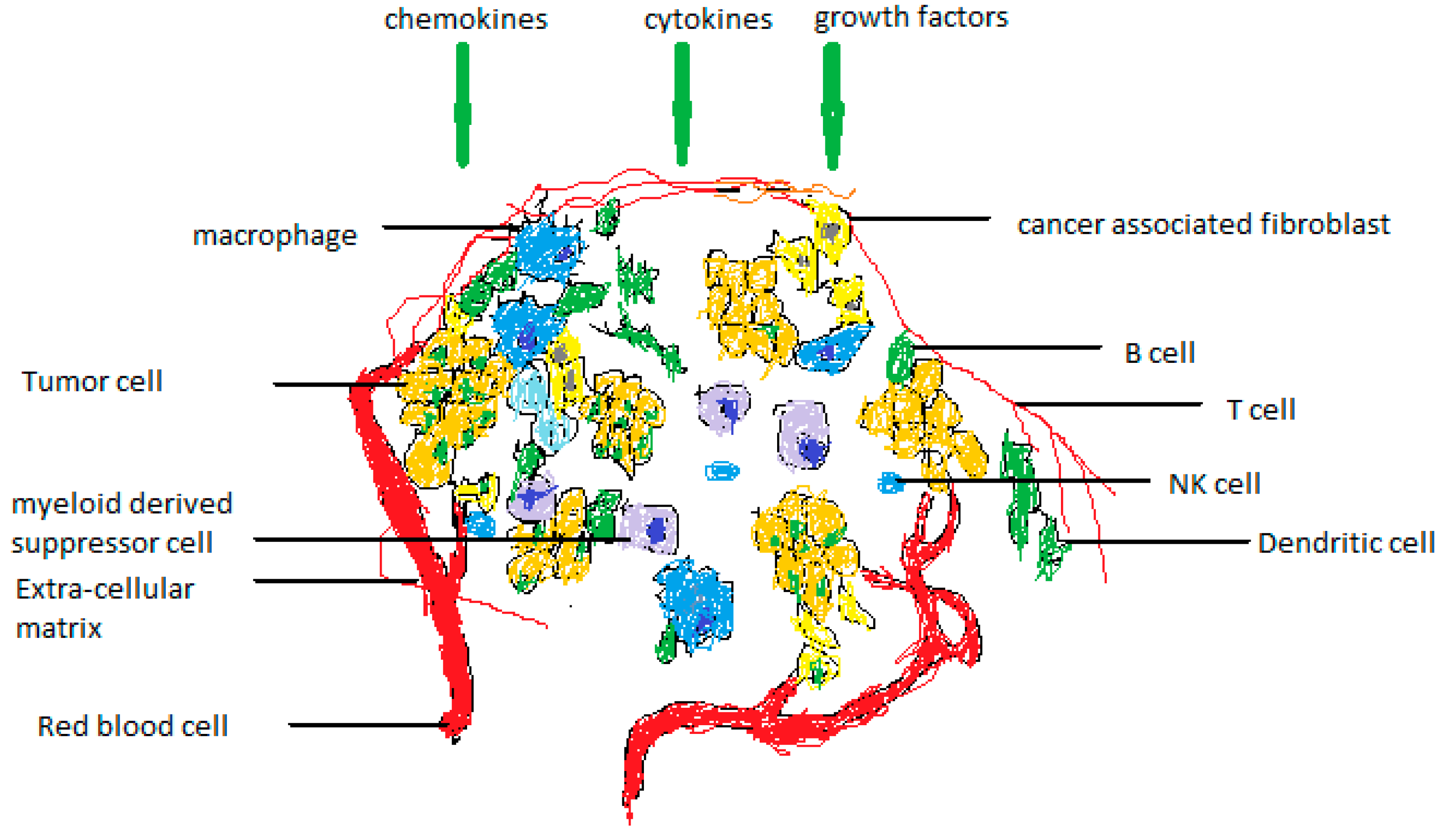
Figure 1.
The elements that make up the tumor's microenvironment. The TME is made up of both cellular and noncellular elements. Cancer-associated fibroblasts (CAFs), adipocytes (APs), niche, immune cells, carcinogenic and noncancerous cells, endothelial cells (ECs), mesenchymal stem cells (MSCs), and mesenchymal stem cells (MSCs) are all present in the former and all contribute to the tumor's progression. The mediators that make up the noncellular component include growth factors, cytokines, and chemokines. They could develop independently or promote a cancer cell's growth.
Figure 1.
The elements that make up the tumor's microenvironment. The TME is made up of both cellular and noncellular elements. Cancer-associated fibroblasts (CAFs), adipocytes (APs), niche, immune cells, carcinogenic and noncancerous cells, endothelial cells (ECs), mesenchymal stem cells (MSCs), and mesenchymal stem cells (MSCs) are all present in the former and all contribute to the tumor's progression. The mediators that make up the noncellular component include growth factors, cytokines, and chemokines. They could develop independently or promote a cancer cell's growth.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



