Submitted:
04 August 2023
Posted:
09 August 2023
Read the latest preprint version here
Abstract
Interferons were the original prototype cytokine system discovered during research of the 20th century. As the name suggests these were originally considered to be synthesised and secreted between cells. However, technological advancements since dictate processes involved in secreting these proteins can be extensively explained through both genetic and biochemical pathways comparatively clearer. Interferon (IFN) discovery occurred when genetic research was in its infancy. Simultaneous discovery by Franklin and Wilkins of deoxyribonucleic acid (DNA) structure and function occurred with Crick and Watson concurrently; however, two scientists Isaac and Lindemann described the first IFN in 1957. Technological advancement allows comparison, since many pathogens and genetic mutations can be factors in IFN regulation. Cancer cell regulation in research has long been central to host IFN synthesis and/or affected with differential IFN protein subunits defined further acting through 6 protein domains. Type II IFN remains central to immune cell function as it is released by a myriad of immune cells, mainly Natural Killer and T cells and is commonly used as a quantitative measurement of adaptive cellular immunity. Single–stranded and/or double–stranded RNA and DNA viruses as well as bacterial infections (e.g., Escherichia) and fungal infections (e.g., Mycobacteria) can affect IFN systems. These utilise intra/extracellular proteins like Toll–like Receptors (TLRs) affected by mutations within the overall IFN transduction pathways. Questions remain over immunological mechanisms contributing to innate and adaptive host immune regulation since type III IFN discovery in 2003 with immune cell phenotypes characterised further. Changes to synthesis of host type I/II/III IFNs may alter homeostatic cellular pathways differentially and beneficially in pathological disorders. We therefore aim to present the rationale of this regulatory protein mechanism of action in context with research developments recently with the functional effects
Keywords:
Interferon
; Innate
; Adaptive
; Genetic
; Molecular
Introduction
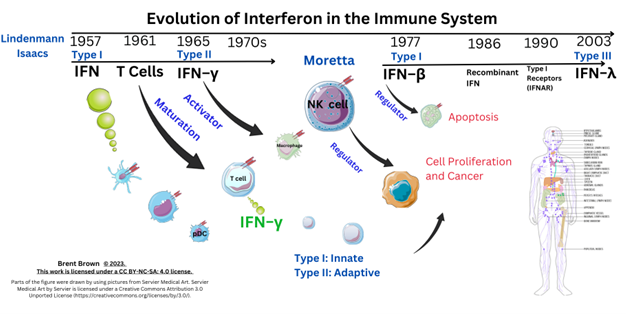
Interferons (IFN) are glycoproteins secreted that have historical unique anti–viral activity as well as cellular oncological regulation properties induced through immune cell regulation and secretion for example by dendritic cells (DCs) early in infection. The different types of IFN can stimulate immune system innate/adaptive compartments having pleotropic cellular properties regulated by many immune cells released proteins including cytokines (IL), chemokines (CC, CXC), acting as cellular autocrine/paracrine signals in a hormonal manner 1. Interferon nomenclature is derived historically as alpha (α, from leukocytes), beta (β, from fibroblasts), and gamma (γ), from mitogen activated lymphocytes stimulated to proliferate. Following the 1957 initial discovery of one IFN, three principal types of IFN are now known divided into type I (α/ β), type II (γ), and recent (2003) discovery of type III (λ), with each having distinct anti–proliferative and anti–viral activities and further sub–types. At least three IFN types have unique variable cell function and are expressed when differentially expressed genes (DEGs), are transcribed and translated during health and/or disease regulated by IFN–stimulated genes (ISGs), as well as IFN–inducible proteins (IFI), with IFI transmembrane (IFIT/M) proteins being additional cellular regulatory factors. Immunisation and therapeutic treatment historically did target IFN therapeutic benefit through pre–clinical development, from phase 1/ 2 to phase 3 and beyond, according to overall safety profile and success rates through regulatory and monitoring authorities like the United States Food and Drug Administration Agency (FDA), European Centre for Disease Control (ECDC), and other organisations. However, literature regarding type I/II/III IFN is required to detail the overall positive and negative controls
Figure 1.
Interferon Types.
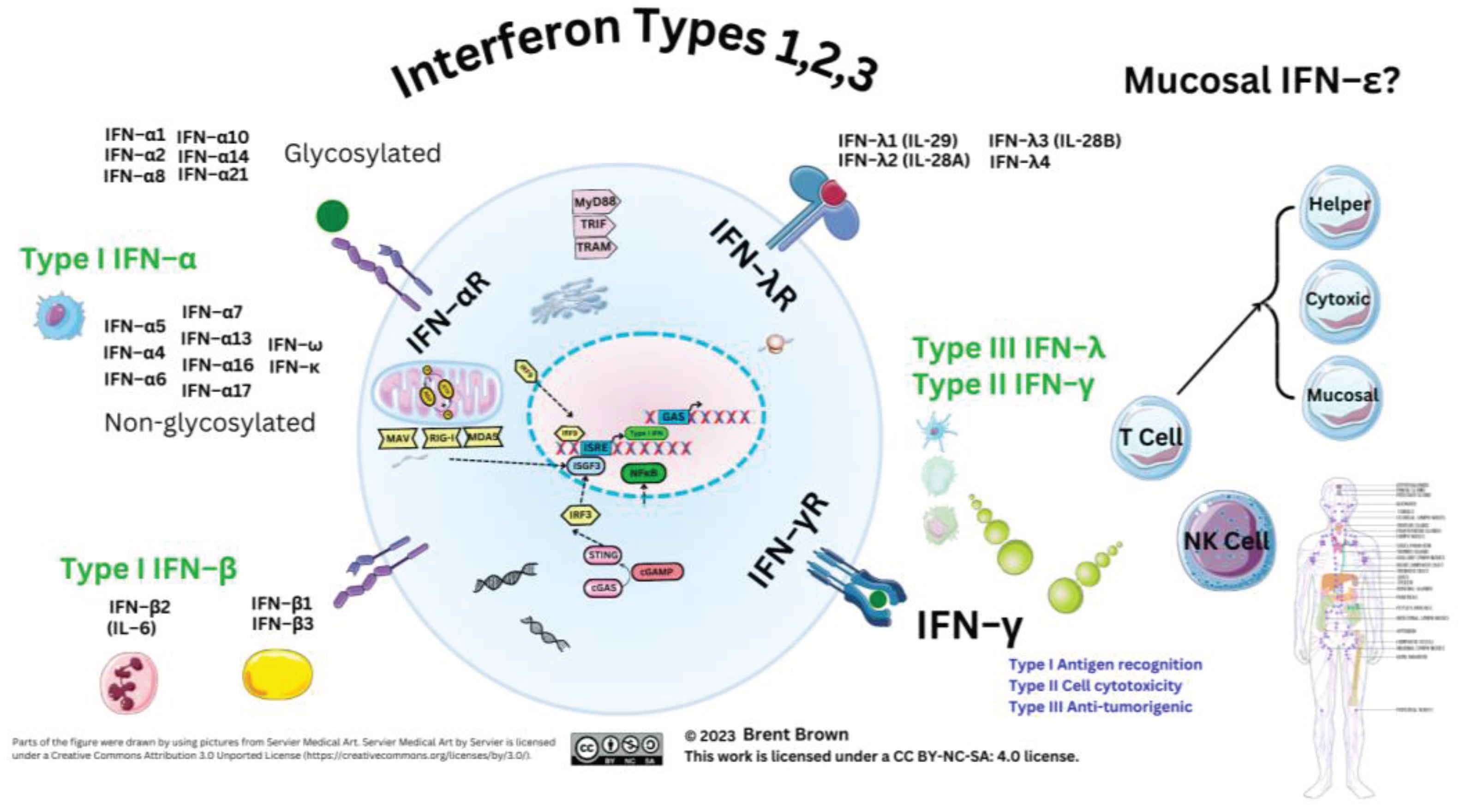
Methodology
Currently, indications were more than 100,000 pubmed results showed prior interferon research. Clinical trials (NCT) investigating utility of IFN as a potential therapeutic total number: Furthermore, these are currently divided into other clinical trials that include type I IFN–α (380), type I IFN–β (116), type I IFN–omega (6) and type I (epsilon (1) (5 type II IFN–γ (173), and type III IFN–λ (17) currently (see Supplementary Data S1).
Background
Through either natural IFN or recombinant IFN synthetic compounds, either human type I IFN and/or type II/III IFN concentrations within host populations can train and stimulate both innate/adaptive two immune system branches thereby honing an effective response during disease. One includes T cell synthesis and NK cell synthesis of type II IFN also produced by other antigen presenting cells (APCs), like macrophage phenotypes (M1ϕ/M2ϕ). The immune system senses pathogenic antigens additionally through pattern recognition receptors (PRRs), as well as cellular endosomal expressed Toll–like (TLR) receptors. Cancer pathologies also respond to type II IFN, whilst viral evolution affects type I/II IFN homeostatic immune cell function. This aspect during viral epidemics/pandemics is known over prior decades, evidenced with Dengue Fever virus (DENV), Ebola virus (EBOV), and recently Monkeypox virus (MPXV) 2,3. It is plausible that regulation of three types of IFN is modulated and can effect early therapeutic and/or clinical disease onset–delaying effects during viral evoked diseases like Influenza A virus (IAV), but also Measles (MeV), as well as Human Immunodeficiency virus (HIV); however other bacterial diseases are similar, such as lower respiratory tract bacterial infection caused by Haemophilus influenzae, Streptococcus pneumoniae and Staphylococcus aureus, as well as oncological diseases, like hepatic melanoma 4. Other reviews ascertain regulatory IFN proteins can be affected by viral proteins (VP) synthesised by Coronaviridae (SARS–CoV–2) as well as Flaviviridae (DENV, Yellow Fever) 5. Individual VP mutations also affect other cytosolic PRRs (e.g., retinoic acid–inducible gene I, RIG–I/ mitochondrial antiviral–signaling protein MAVS) pathways in at least two other virus families (Filoviridae/Nairovidiae) 2,3,5. There are unknowns regarding retinoic acid that is a metabolic component of immune cells. Viral mutations occur in both DNA/RNA viruses like the positive–sense single–stranded RNA virus (+ssRNA) Influenza A (Alphainfluenzavirus). This has 198 quantified potential antigen subtype combinations of the cellular expressed haemagglutinin/neuraminidase (HA/NA) protein antigens, which also affect immune cell phenotypes. Different serotypes of Gram negative coccobacilli, Haemophilus influenzae, are known to shed intracellular or extracellular membranes during infection (e.g., Hia–Hif or ncHI); whilst others like Avian Influenza virus (IAV, H5N1) have been observed during zoonotic spillover. Therefore, viral antigens co–exist circulating in nature, with increases in antigen circulation necessitate clarification of interferon as a regulatory factor within all host immune responses.
Three or four or more types of IFN may have differential inhibitory or activation mechanisms on the immune system causal in lysing infectious viruses effectively through stimulating immune cellular effector cell activity through a myriad of proteins described here. This is effected though IFN receptors (IFNR) at the cell plasma membrane (PM) surface. This occurs through at least 18 types of IFN binding to 6 IFNR proteins expressed by dendritic cells (DCs) which also have variable phenotypes. Interferon receptors are also expressed by B–lymphocytes, monocytes, and macrophages (M1ϕ/M2ϕ), as well as T–lymphocytes. Receptor expression is also within the cellular PM including glial cells, neurons, and other cells. Interferon receptors (IFNR) therefore initiate downstream/upstream cellular effects as well as T cell secretion of type II IFN–γ upon host cellular viral infection. Plant products can also generate IFN stimulating proteins. Timing of cellular IFN synthesis and cellular secretion affects viral infection, propagation, replication. But also IFN acts through a myriad of cells and protein pathways to effect cellular lysis in organs, tissues and cell systems by regulating other cell cycle proteins, like p38. Immunodeficiency disorders or individual protein mutations may cause errors in IFN/IFNR signaling throughout development.
Therefore, the regulation of type I/II/III IFN response can have resultant detrimental and/or beneficial factors. The subtypes of these proteins directly affect and influence the two branches of the innate and adaptive immune response requiring clarity. Each IFN type fulfils individually unique immunological roles during 5 types of pathology including viral, fungal, bacterial, mycobacterial as well as oncogenic diseases. Immune system modulation and/or evasion may represent evolutionary development within animal hosts varying. Therefore here is the analysis of genetic, molecular, and cellular analysis of type I/II/III IFN mechanisms of action to date that will require further research.
Interferon Types
Overview to Interferon Cellular Types
Type I IFNs are synthesised/secreted after translation from cellular nuclear transcription factors (TFs) resulting in differential anti–viral activity against host pathogens that may vary. Each IFN protein is known as a small molecular weight (MW) molecule in humans; for example type I IFN–α1/13, IFN–α2, IFN– α8 and IFN–α21 are composed of 187–189 amino–acids, while type III IFN–λ are within the MW range 179–200 amino acids. Chemokines in comparison are smaller MW proteins (e.g., CCL2, 99 amino–acids), with pleiotropic effects directing immune cell migration throughout tissues. These small MW proteins are induced through gene synthesis transcription and can be translated earlier in response to pathogenic antigens outside the cell nucleus. Interferon subtypes can be synthesised by myeloid cells like plasmacytoid dendritic cells (pDCs) producing higher concentrations of type I IFN (IFN–α/IFN–β), effecting anti–viral responses in hosts; but also within skin epithelial cell tissues through tumour necrosis factor (TNF) related apoptosis–inducing ligand (TRAIL) and at least ten intra/extracellular PM and vesicular TLRs 6. On the other hand type II IFN–γ is secreted by at least two effector cells (NK and T cells) together with another two antigen presenting cells (DCs and Mϕ)with different phenotypes. Whilst type III IFN subtypes may also influence host immune responses within epithelial layers. It could generally be considered that through regulating cellular cycle function that each IFN performs different roles with type I IFN–β potentially regulating Mϕ cell cycle (M1ϕ/M2ϕ) and metabolism whilst type I IFN–α could be considered similar in regulation of homeostatic function and observed commonly in inflammation and autoimmune disorders .
Type I IFNs include IFN–β, IFN–δ, IFN–ε, IFN–κ, IFN–τ, IFN–ω, and IFN–ζ amongst others; whilst type III IFNs are composed of IFN–λ (IFN–λ1, IFN–λ2, IFN–λ3, IFN–λ4), known originally as IL29, IL28A, and IL28B with IFN–λ4 discovered in 2014 7. Two types of type III IFN (λ2,λ3) are considered to have 96% amino–acid homology 8. Other classifications of IFN subtype exist and all vary between host animal species encoded by IFN genes. To clarify, human IFN consists of at least 18 subtypes, some others of which are type I IFN–α4, IFN–α7, and IFN– α14; whilst in pigs and bats diversity of IFN–ω is worthy of consideration with less type I IFN–α described as discussed further 9–11. Amongst type I IFN–α subtypes, IFN–α2 is O–glycosylated with a recombinant IFN–α2b therapeutic version utilised 12. However, research studies since 2015 indicate that IFN–α2 is N–glycosylated and can be cleaved during secretion losing 23 amino acids and one aspartic acid (L) at position 44 13. Recently it was shown that type I IFN may contain pro–inflammatory glycans known to enhance binding of the predominant antibody (IgG) to immune cell FcγR PM receptors (CD16/CD32/CD64) that could train the immune system response 14,15. Furthermore, this could influence the T cell response , including both cytotoxic (TC) and NK cell function, through modulation of sialic acid residues present in other receptors like DC–SIGN or fucose residues.
Therefore in 2019, pharmacokinetic properties of recombinant type I IFN–α2 engineering indicated the production vector could affect pharmacokinetic half–life when glycoengineering indicated Pichia pastoris as a potential option together with purification of recombinant interferon; whilst all subtypes of IFN–β are also N–glycosylated 16–18. In comparison, other studies show the addition of a glycosyl group on IFN–λ4 may increase anti–inflammatory actions and anti–viral efficacy 19. It is notable that glycosylated IFNs vary in stability and display antimicrobial effects with research comparatively unknown 20. Glycosylated IFNs may interact with many carbohydrates and receptors on cell PMs and have higher/lower binding affinities to receptors necessitating clarification.
Respective receptors include type I IFN receptors (IFNAR1/2), type II IFN receptors (IFN–γR1/ IFN–γR2), as well as type III IFN receptors (IFN–λR1 IFN–λR2(IL10R2), each composed of two subunit domains 21,22. Just prior to 2011, comparisons of type I IFN–α assays allowed type I IFN–α receptor binding studies to show IFN binding to IFNAR1 occurred with higher (µm) affinity whilst binding to IFNAR2 was lower affinity in a smaller (nm) range 23. However, IFN–β and IFN–λ are produced by various cells, with IFN–α generally synthesised by immune cells, but specifically pDCs during infection with receptors throughout the bodily system 24. Other reviews establish type I IFN downregulation, while research into type III IFNs is in the early stages; however, we agree with other authors suggesting type III IFNs may have biological mechanisms requiring clarity 25,26. Much remains unknown with regards to type III IFN–λ signaling proteins. Specific data about therapy and immunisation comes through national clinical trials (NCTs) conducted throughout history before/after the first cloning of IFN receptors in 1990 and the usage of recombinant IFN–α2 (see Supplementary Data S1) 27. During the recent pandemic, type I IFNs were evidenced to have an effect in reducing SARS–CoV–2 viral genome load requiring further contextual detail 28–32.
The Three Type of Interferon Roles in the Immune System
Cellular effects of IFN are dependant on affinity of 3 types of IFN and subtypes with 6 subunit receptor domains and receptor expression in organs, systems, tissues and cells. In brief, type I IFN–α research to date does indicate unusual variance during host infections with evidential beneficial/detrimental effects, but yet regulates myeloid cell lineages as well as B/T cells and NK cells effects. This training of immune responses occurs through inhibition as well as stimulation of maturation/differentiation of DCs by regulating costimulatory molecules like CD80/CD86 increasing major histocompatibility complex (MHC) antigen presentation as well as stimulation of T cell phenotypes 33. However, DC maturation is known to occur through pDCs into three types conventional DCs (cDC1, cDC2, cDC3), with the most recent only known since 2017 that can reversibly differentiate into myeloid/monocytic lineages. It is suggested that type I IFN induces the suppressive cytokine IL–10 and mediates pro–inflammatory IFN gene product inhibition through suppressor of cytokine signaling–1 (SOCS–1) and be anti–viral/anti–proliferative as well having anti–tumour activity. This was evidenced from 2002 in more than 40 countries where recombinant type I IFN–α2 was used as a therapeutic to treat various types of leukaemia (B/T cell lymphomas) 34,35. In comparison, type II IFN–γ is largely produced by only cells of the immune system and primarily T cells that can upregulate MHC class II proteins. It is also produced and regulated by NK cells. Two primary T cell phenotypes produce type II IFN–γ with the majority expressing CD4 and/or CD8 molecule proteins 36. Historically, type II IFN has been utilised as a measure of T cell activity in research into adaptive immunity. Activity of type III IFN expression can now be measured by expression of the subunit receptors in tissues and cells through cellular mRNA expression. It is currently considered that the mRNA for one receptor subunit domain, IFNLR2 (IL10R2), has been observed in lungs, intestines, liver tissues as well as B cells, neutrophils, macrophages and pDCs but not in NK cells 37. Additionally, type III IFN is considered to have higher affinity for IFNLR1 with less affinity for IL10R2 possibly explaining some of the differential activity of IL–10 which shares this receptor. Previously, type III IFN was considered to be predominant on non–hematopoietic cells such as epithelial cells. Type III IFN has lower affinity binding to its respective receptors than type I IFN. Other reviews examine the relevance of single nucleotide point (SNP) mutations of type III IFN pathways during disease 37. The relevance of type III IFN is only now becoming clearer. Research in vivo is indicative that during type III IFN–λ2 (IL28A) deficiency, there is an effect on three crucial immune system branches. Namely, germinal B cell centre formation where antibody forming cells were observed to increase concurrently with increased activity of both T cell branches denoted by T helper (CD4+) cells as well as cytotoxic T (CD8+) cells. Moreover, type III IFN–λ3 has also similarly only just been highlighted as relevant to B cell proliferation and antibody production 37,38.
Immune system modulation and/or evasion may represent evolutionary development within animal host immune systems and vary. Recently, three types of cellular signaling are considered alongside IFN that are cytokines (interleukins , IL) and chemokines (CC/CXC). Individual cellular expression is stimulated by many pathogenic organisms, like Smallpox (VARV), Human Immunodeficiency virus (HIV), but also bacterial pathogens (Streptococcus), and others like Respiratory Syncytial virus (RSV) can also cause viral induced pathology., Viral mutations occur in DNA/RNA viruses like the positive–sense single–stranded RNA virus (+ssRNA) Influenza A (Alphainfluenzavirus), having 198 potential subtype combinations of haemagglutinin/neuraminidase (HA/NA) protein antigens that can differentially affect immune cell phenotypes. Different serotypes of Gram–negative coccobacilli, Haemophilus influenzae, are known to shed intracellular or extracellular membranes during infection (e.g. Hia–Hif or ncHI). Other viruses like Influenza A avian virus (H5N1) can affect variable animal hosts. Severe forms of Influenza A may cause morbidity following secondary bacterial infection.
Therefore, many viral antigens co–exist alongside the novel SARS–CoV–2 antigens in nature. This effective increase in antigen circulation may inhibit or stimulate or sentisite the immune system differentially affecting effective lysis of infectious viruses through host IFN synthesis or unknown metabolic factors. The three shared methods of immune system differential kinetics comprise of pathogenic DNA/RNA 5’ capping, by addition of a methyl (CH3–) group to the 5’ genome with pattern recognition receptors (PRR) including Toll–like receptors (TLRs) affected. Secondly, cellular mitochondrial metabolic changes may change the synthesis rate of reactive oxygen species (ROS), whilst viruses utilise inter–cellular channeling nanotubes 39.The final evolutionary objective considered could be modulation of types of type I/III IFN or indeed the rate of IFN synthesis by either immune cells or infected cells with much remaining unknown.
Supplementary Materials
Welcome to Interferome (monash.edu.au).
Author Contributions
Original manuscript draft written by B.B.; conceptualisation by: B.B., formal analysis by: B.B.; data curation by: B.B.; writing—reviewing and editing by: B.B..: software: B.B.: visualization: B.B.; supervision by B.B.; validation: .B.; Figures designed by Brent/Brown/Biochem123 Ltd. All authors have signed, read and agreed to the published version of the manuscript according to IJCME/COPE guidelines.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not–for–profit sectors.
Ethical Approval Statement and/or Informed Consent Statement
See originating citations and Supplementary Materials. Author(s) have given signed consent. Individual citations are subject to the Helsinki Declaration and individual regulatory authorities.
Data Availability Statement
Contained within manuscript.
Trial registration details
CTG Labs - NCBI (clinicaltrials.gov).
Conflict or Competing Interest
The author(s) declares no conflict of interest in the production of this manuscript.
Use of Artificial Intelligence (AI) Disclaimer
During the preparation of this work the author(s) did not use AI tools or service. Citations are referenced using Mendeleey. After using the above tool/service, each author(s) reviewed and edited the content.
References
- Brown, B. Innate and Adaptive Immunity during SARS-CoV-2 Infection: Biomolecular Cellular Markers and Mechanisms. Vaccines (Basel) 2023, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Brown, B. Immunopathogenesis of Orthopoxviridae: Insights into Immunology from Smallpox to Monkeypox (Mpox). ( 2023. [CrossRef]
- Brown, B. Filoviridae: Insights into Immune Responses to Ebolavirus. ( 2023. [CrossRef]
- Sheng, L.; Chen, X.; Wang, Q.; Lyu, S.; Li, P. Interferon-α2b enhances survival and modulates transcriptional profiles and the immune response in melanoma patients treated with dendritic cell vaccines. BioMedicine 2020, 125, 109966. [Google Scholar] [CrossRef]
- Mesev, E.V.; LeDesma, R.A.; Ploss, A. Decoding type I and III interferon signalling during viral infection. Nat. Microbiol. 2019, 4, 914–924. [Google Scholar] [CrossRef]
- Högner, K.; Wolff, T.; Pleschka, S.; Plog, S.; Gruber, A.D.; Kalinke, U.; Walmrath, H.-D.; Bodner, J.; Gattenlöhner, S.; Lewe-Schlosser, P.; et al. Macrophage-expressed IFN-β Contributes to Apoptotic Alveolar Epithelial Cell Injury in Severe Influenza Virus Pneumonia. PLOS Pathog. 2013, 9, e1003188. [Google Scholar] [CrossRef]
- Reizis, B. Plasmacytoid Dendritic Cells: Development, Regulation, and Function. Immunity 2019, 50, 37–50. [Google Scholar] [CrossRef]
- O'Brien, T.R.; Prokunina-Olsson, L.; Donnelly, R.P. IFN-λ4: The Paradoxical New Member of the Interferon Lambda Family. J. Interferon. Cytokine Res. 2014, 34, 829–838. [Google Scholar] [CrossRef]
- Shields, L.E.; Jennings, J.; Liu, Q.; Lee, J.; Ma, W.; Blecha, F.; Miller, L.C.; Sang, Y. Cross-Species Genome-Wide Analysis Reveals Molecular and Functional Diversity of the Unconventional Interferon-ω Subtype. Front. Immunol. 2019, 10, 1431. [Google Scholar] [CrossRef]
- Wittling, M.C.; Cahalan, S.R.; Levenson, E.A.; Rabin, R.L. Shared and Unique Features of Human Interferon-Beta and Interferon-Alpha Subtypes. Front. Immunol. 2021, 11. [Google Scholar] [CrossRef]
- Zhou, P. et al. Contraction of the type I IFN locus and unusual constitutive expression of IFN-α in bats. Proceedings of the National Academy of Sciences 113, 2696–2701 (2016).
- George, J.; Mattapallil, J.J. Interferon-α Subtypes As an Adjunct Therapeutic Approach for Human Immunodeficiency Virus Functional Cure. Front. Immunol. 2018, 9, 299. [Google Scholar] [CrossRef] [PubMed]
- Paul, F.; Pellegrini, S.; Uzé, G. IFNA2: The prototypic human alpha interferon. Gene 2015, 567, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, X. IgG N-glycans. 2021, 105, 1–47. [CrossRef]
- Giron, L.B.; Colomb, F.; Papasavvas, E.; Azzoni, L.; Yin, X.; Fair, M.; Anzurez, A.; Damra, M.; Mounzer, K.; Kostman, J.R.; et al. Interferon-α alters host glycosylation machinery during treated HIV infection. EBioMedicine 2020, 59, 102945. [Google Scholar] [CrossRef]
- Katla, S.; Yoganand, K.; Hingane, S.; Kumar, C.R.; Anand, B.; Sivaprakasam, S. Novel glycosylated human interferon alpha 2b expressed in glycoengineered Pichia pastoris and its biological activity: N-linked glycoengineering approach. Enzym. Microb. Technol. 2019, 128, 49–58. [Google Scholar] [CrossRef] [PubMed]
- How, J.; Hobbs, G. Use of Interferon Alfa in the Treatment of Myeloproliferative Neoplasms: Perspectives and Review of the Literature. Cancers 2020, 12, 1954. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef]
- Chung, J.-H.; Hong, S.-H.; Seo, N.; Kim, T.-S.; An, H.J.; Lee, P.; Shin, E.-C.; Kim, H.M. Structure-based glycoengineering of interferon lambda 4 enhances its productivity and anti-viral potency. Cytokine 2020, 125, 154833–154833. [Google Scholar] [CrossRef]
- Li, S.-F.; Gong, M.-J.; Zhao, F.-R.; Shao, J.-J.; Xie, Y.-L.; Zhang, Y.-G.; Chang, H.-Y. Type I Interferons: Distinct Biological Activities and Current Applications for Viral Infection. Cell. Physiol. Biochem. 2018, 51, 2377–2396. [Google Scholar] [CrossRef]
- Zanin, N.; de Lesegno, C.V.; Lamaze, C.; Blouin, C.M. Interferon Receptor Trafficking and Signaling: Journey to the Cross Roads. Front. Immunol. 2021, 11. [Google Scholar] [CrossRef]
- Shemesh, M.; Lochte, S.; Piehler, J.; Schreiber, G. IFNAR1 and IFNAR2 play distinct roles in initiating type I interferon–induced JAK-STAT signaling and activating STATs. Sci. Signal. 2021, 14, eabe4627. [Google Scholar] [CrossRef]
- Lavoie, T.B.; Kalie, E.; Crisafulli-Cabatu, S.; Abramovich, R.; DiGioia, G.; Moolchan, K.; Pestka, S.; Schreiber, G. Binding and activity of all human alpha interferon subtypes. Cytokine 2011, 56, 282–289. [Google Scholar] [CrossRef]
- Kim, Y.-M. & Shin, E.-C. Type I and III interferon responses in SARS-CoV-2 infection. Exp Mol Med 53, 750–760 (2021).
- Ali, S.; Mann-Nüttel, R.; Schulze, A.; Richter, L.; Alferink, J.; Scheu, S. Sources of Type I Interferons in Infectious Immunity: Plasmacytoid Dendritic Cells Not Always in the Driver's Seat. Front. Immunol. 2019, 10, 778. [Google Scholar] [CrossRef] [PubMed]
- LaSalle, T.J.; Gonye, A.L.; Freeman, S.S.; Kaplonek, P.; Gushterova, I.; Kays, K.R.; Manakongtreecheep, K.; Tantivit, J.; Rojas-Lopez, M.; Russo, B.C.; et al. Longitudinal characterization of circulating neutrophils uncovers phenotypes associated with severity in hospitalized COVID-19 patients. Cell Rep. Med. 2022, 3, 100779. [Google Scholar] [CrossRef] [PubMed]
- de Weerd, N.A.; Samarajiwa, S.A.; Hertzog, P.J. Type I Interferon Receptors: Biochemistry and Biological Functions. Perspect. Surg. 2007, 282, 20053–20057. [Google Scholar] [CrossRef]
- Lam, J.-Y. et al. Loss of orf3b in the circulating SARS-CoV-2 strains. Emerg Microbes Infect 9, 2685–2696 (2020).
- Overholt, K.J.; Krog, J.R.; Zanoni, I.; Bryson, B.D. Dissecting the common and compartment-specific features of COVID-19 severity in the lung and periphery with single-cell resolution. iScience 2021, 24, 102738. [Google Scholar] [CrossRef] [PubMed]
- Sposito, B.; Broggi, A.; Pandolfi, L.; Crotta, S.; Clementi, N.; Ferrarese, R.; Sisti, S.; Criscuolo, E.; Spreafico, R.; Long, J.M.; et al. The interferon landscape along the respiratory tract impacts the severity of COVID-19. Cell 2021, 184, 4953–4968. [Google Scholar] [CrossRef]
- Mantlo, E. , Bukreyeva, N., Maruyama, J., Paessler, S. & Huang, C. Antiviral activities of type I interferons to SARS-CoV-2 infection. Antiviral Res 179, 104811 (2020).
- Brzoska, J. , von Eick, H. & Hündgen, M. Interferons in COVID-19: missed opportunities to prove efficacy in clinical phase III trials? Front Med (Lausanne) 10, (2023).
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O'Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef]
- Medrano, R.F.V.; Hunger, A.; Mendonça, S.A.; Barbuto, J.A.M.; Strauss, B.E. Immunomodulatory and antitumor effects of type I interferons and their application in cancer therapy. Oncotarget 2017, 8, 71249–71284. [Google Scholar] [CrossRef]
- Belardelli, F.; Ferrantini, M.; Proietti, E.; Kirkwood, J.M. Interferon-alpha in tumor immunity and immunotherapy. Cytokine Growth Factor Rev. 2002, 13, 119–134. [Google Scholar] [CrossRef]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in tumor progression and regression: a review. Biomark. Res. 2020, 8, 1–16. [Google Scholar] [CrossRef]
- Syedbasha, M.; Egli, A. Interferon Lambda: Modulating Immunity in Infectious Diseases. Front. Immunol. 2017, 8, 119. [Google Scholar] [CrossRef] [PubMed]
- Egli, A.; Santer, D.M.; O'Shea, D.; Barakat, K.; Syedbasha, M.; Vollmer, M.; Baluch, A.; Bhat, R.; Groenendyk, J.; Joyce, M.A.; et al. IL-28B is a Key Regulator of B- and T-Cell Vaccine Responses against Influenza. PLOS Pathog. 2014, 10, e1004556. [Google Scholar] [CrossRef] [PubMed]
- Sherer, N.M.; Mothes, W. Cytonemes and tunneling nanotubules in cell–cell communication and viral pathogenesis. Trends Cell Biol. 2008, 18, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Dubois, A.; François, C.; Descamps, V.; Fournier, C.; Wychowski, C.; Dubuisson, J.; Castelain, S.; Duverlie, G. Enhanced anti-HCV activity of interferon alpha 17 subtype. Virol. J. 2009, 6, 70–70. [Google Scholar] [CrossRef]
- Scagnolari, C.; Trombetti, S.; Selvaggi, C.; Carbone, T.; Monteleone, K.; Spano, L.; Di Marco, P.; Pierangeli, A.; Maggi, F.; Riva, E.; et al. In Vitro Sensitivity of Human Metapneumovirus to Type I Interferons. Viral Immunol. 2011, 24, 159–164. [Google Scholar] [CrossRef]
- Markušić, M.; Šantak, M.; Košutić-Gulija, T.; Jergović, M.; Jug, R.; Forčić, D. Induction of IFN-αSubtypes and Their Antiviral Activity in Mumps Virus Infection. Viral Immunol. 2014, 27, 497–505. [Google Scholar] [CrossRef]
- Yang, L.; Wang, S.; Wang, Y.; Zhao, P.; Cui, C.; Tu, L.; Li, X.; Yu, Y.; Li, H.; Wang, L. Diversity of locally produced IFN-α subtypes in human nasopharyngeal epithelial cells and mouse lung tissues during influenza virus infection. Appl. Microbiol. Biotechnol. 2020, 104, 6351–6361. [Google Scholar] [CrossRef]
- El Safadi, D.; Lebeau, G.; Lagrave, A.; Mélade, J.; Grondin, L.; Rosanaly, S.; Begue, F.; Hoareau, M.; Veeren, B.; Roche, M.; et al. Extracellular Vesicles Are Conveyors of the NS1 Toxin during Dengue Virus and Zika Virus Infection. Viruses 2023, 15, 364. [Google Scholar] [CrossRef]
- Brown, B.; Gravier, T.; Fricke, I.; Al-Sheboul, S.A.; Carp, T.-N.; Leow, C.Y.; Imarogbe, C.; Arabpour, J. Immunopathogenesis of Nipah Virus Infection and Associated Immune Responses. Immuno 2023, 3, 160–181. [Google Scholar] [CrossRef]
- Ilinykh, P.A.; Lubaki, N.M.; Widen, S.G.; Renn, L.A.; Theisen, T.C.; Rabin, R.L.; Wood, T.G.; Bukreyev, A. Different Temporal Effects of Ebola Virus VP35 and VP24 Proteins on Global Gene Expression in Human Dendritic Cells. J. Virol. 2015, 89, 7567–7583. [Google Scholar] [CrossRef]
- Mateo, M.; Reid, S.P.; Leung, L.W.; Basler, C.F.; Volchkov, V.E. Ebolavirus VP24 Binding to Karyopherins Is Required for Inhibition of Interferon Signaling. J. Virol. 2010, 84, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Leung, L.W.; Park, M.; Martinez, O.; Valmas, C.; López, C.B.; Basler, C.F. Ebolavirus VP35 suppresses IFN production from conventional but not plasmacytoid dendritic cells. Immunol. Cell Biol. 2011, 89, 792–802. [Google Scholar] [CrossRef]
- Kuroda, M.; Halfmann, P.J.; Hill-Batorski, L.; Ozawa, M.; Lopes, T.J.S.; Neumann, G.; Schoggins, J.W.; Rice, C.M.; Kawaoka, Y. Identification of interferon-stimulated genes that attenuate Ebola virus infection. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Woolsey, C.; Menicucci, A.R.; Cross, R.W.; Luthra, P.; Agans, K.N.; Borisevich, V.; Geisbert, J.B.; Mire, C.E.; Fenton, K.A.; Jankeel, A.; et al. A VP35 Mutant Ebola Virus Lacks Virulence but Can Elicit Protective Immunity to Wild-Type Virus Challenge. Cell Rep. 2019, 28, 3032–3046. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT signaling pathway: from bench to clinic. Signal Transduct. Target. Ther. 2021, 6, 1–33. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, G.; Zhu, L.; Zhao, Z.; Liu, Y.; Han, W.; Zhang, X.; Zhang, Y.; Xiong, T.; Zeng, H.; et al. HIV-1 Vif suppresses antiviral immunity by targeting STING. Cell. Mol. Immunol. 2021, 19, 108–121. [Google Scholar] [CrossRef]
- Khatamzas, E.; Hipp, M.M.; Gaughan, D.; Pichulik, T.; Leslie, A.; A Fernandes, R.; Muraro, D.; Booth, S.; Zausmer, K.; Sun, M.; et al. Snapin promotes HIV -1 transmission from dendritic cells by dampening TLR 8 signaling. EMBO J. 2017, 36, 2998–3011. [Google Scholar] [CrossRef]
- Passarelli, C.; Civino, A.; Rossi, M.N.; Cifaldi, L.; Lanari, V.; Moneta, G.M.; Caiello, I.; Bracaglia, C.; Montinaro, R.; Novelli, A.; et al. IFNAR2 Deficiency Causing Dysregulation of NK Cell Functions and Presenting With Hemophagocytic Lymphohistiocytosis. Front. Genet. 2020, 11, 937. [Google Scholar] [CrossRef]
- Bucciol, G.; Moens, L.; Ogishi, M.; Rinchai, D.; Matuozzo, D.; Momenilandi, M.; Kerrouche, N.; Cale, C.M.; Treffeisen, E.R.; Al Salamah, M.; et al. Human inherited complete STAT2 deficiency underlies inflammatory viral diseases. J. Clin. Investig. 2023, 133. [Google Scholar] [CrossRef]
- Lee, D. et al. Inborn errors of OAS–RNase L in SARS-CoV-2–related multisystem inflammatory syndrome in children. Science (1979) 379, (2023).
- Awasthi, N.; Liongue, C.; Ward, A.C. STAT proteins: a kaleidoscope of canonical and non-canonical functions in immunity and cancer. J. Hematol. Oncol. 2021, 14, 1–17. [Google Scholar] [CrossRef]
- Ahn, D. & Prince, A. Participation of the IL-10RB Related Cytokines, IL-22 and IFN-λ in Defense of the Airway Mucosal Barrier. Front Cell Infect Microbiol 10, (2020).
- Gal-Ben-Ari, S.; Barrera, I.; Ehrlich, M.; Rosenblum, K. PKR: A Kinase to Remember. Front. Mol. Neurosci. 2019, 11, 480. [Google Scholar] [CrossRef] [PubMed]
- Samuel, C. E. Adenosine deaminase acting on RNA (ADAR1), a suppressor of double-stranded RNA–triggered innate immune responses. Journal of Biological Chemistry 294, 1710–1720 (2019).
- Zhang, Y.; Zhang, J.; Xue, Y. ADAR1: a mast regulator of aging and immunity. Signal Transduct. Target. Ther. 2023, 8, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Ehrhardt, C.; Wolff, T.; Pleschka, S.; Planz, O.; Beermann, W.; Bode, J.G.; Schmolke, M.; Ludwig, S. Influenza A Virus NS1 Protein Activates the PI3K/Akt Pathway To Mediate Antiapoptotic Signaling Responses. J. Virol. 2007, 81, 3058–3067. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-S. et al. SARS-CoV-2 inhibits induction of the MHC class I pathway by targeting the STAT1-IRF1-NLRC5 axis. Nat Commun 12, 6602 (2021).
- Zhang, M.; Meng, Y.; Ying, Y.; Zhou, P.; Zhang, S.; Fang, Y.; Yao, Y.; Li, D. Selective activation of STAT3 and STAT5 dictates the fate of myeloid progenitor cells. Cell Death Discov. 2023, 9, 1–11. [Google Scholar] [CrossRef]
- Deimel, L.P.; Li, Z.; Roy, S.; Ranasinghe, C. STAT3 determines IL-4 signalling outcomes in naïve T cells. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Meyer zu Horste, G. et al. Fas Promotes T Helper 17 Cell Differentiation and Inhibits T Helper 1 Cell Development by Binding and Sequestering Transcription Factor STAT1. Immunity 48, 556-569.e7 (2018).
- Kerner, G.; Rosain, J.; Guérin, A.; Al-Khabaz, A.; Oleaga-Quintas, C.; Rapaport, F.; Massaad, M.J.; Ding, J.-Y.; Khan, T.; Al Ali, F.; et al. Inherited human IFN-γ deficiency underlies mycobacterial disease. J. Clin. Investig. 2020, 130, 3158–3171. [Google Scholar] [CrossRef]
- Boisson-Dupuis, S.; Kong, X.-F.; Okada, S.; Cypowyj, S.; Puel, A.; Abel, L.; Casanova, J.-L. Inborn errors of human STAT1: allelic heterogeneity governs the diversity of immunological and infectious phenotypes. Curr. Opin. Immunol. 2012, 24, 364–378. [Google Scholar] [CrossRef]
- Bogunovic, D.; Byun, M.; Durfee, L.A.; Abhyankar, A.; Sanal, O.; Mansouri, D.; Salem, S.; Radovanovic, I.; Grant, A.V.; Adimi, P.; et al. Mycobacterial Disease and Impaired IFN-γ Immunity in Humans with Inherited ISG15 Deficiency. Science 2012, 337, 1684–1688. [Google Scholar] [CrossRef]
- Crow, Y.J.; Stetson, D.B. The type I interferonopathies: 10 years on. Nat. Rev. Immunol. 2021, 22, 471–483. [Google Scholar] [CrossRef]
- Shih, H.-P.; Ding, J.-Y.; Yeh, C.-F.; Chi, C.-Y.; Ku, C.-L. Anti-interferon-γ autoantibody-associated immunodeficiency. Curr. Opin. Immunol. 2021, 72, 206–214. [Google Scholar] [CrossRef]
- Dupuis, S.; Döffinger, R.; Picard, C.; Fieschi, C.; Altare, F.; Jouanguy, E.; Abel, L.; Casanova, J.-L. Human interferon-g-mediated immunity is a genetically controlled continuous trait that determines the outcome of mycobacterial invasion. Immunol. Rev. 2000, 178, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Kreins, A.Y.; Ciancanelli, M.J.; Okada, S.; Kong, X.-F.; Ramírez-Alejo, N.; Kilic, S.S.; El Baghdadi, J.; Nonoyama, S.; Mahdaviani, S.A.; Ailal, F.; et al. Human TYK2 deficiency: Mycobacterial and viral infections without hyper-IgE syndrome. J. Exp. Med. 2015, 212, 1641–1662. [Google Scholar] [CrossRef] [PubMed]
- van de Vosse, E. et al. IL-12Rβ1 Deficiency: Mutation Update and Description of the IL12RB1 Variation Database. Hum Mutat 34, 1329–1339 (2013).
- Zheng, J.; van de Veerdonk, F.L.; Crossland, K.L.; Smeekens, S.P.; Chan, C.M.; Al Shehri, T.; Abinun, M.; Gennery, A.R.; Mann, J.; Lendrem, D.W.; et al. Gain-of-function STAT1 mutations impair STAT3 activity in patients with chronic mucocutaneous candidiasis (CMC). Eur. J. Immunol. 2015, 45, 2834–2846. [Google Scholar] [CrossRef] [PubMed]
- Passarelli, C.; Civino, A.; Rossi, M.N.; Cifaldi, L.; Lanari, V.; Moneta, G.M.; Caiello, I.; Bracaglia, C.; Montinaro, R.; Novelli, A.; et al. IFNAR2 Deficiency Causing Dysregulation of NK Cell Functions and Presenting With Hemophagocytic Lymphohistiocytosis. Front. Genet. 2020, 11, 937. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.K.; Huang, S.X.; Chen, J.; Kerner, G.; Gilliaux, O.; Bastard, P.; Dobbs, K.; Hernandez, N.; Goudin, N.; Hasek, M.L.; et al. Severe influenza pneumonitis in children with inherited TLR3 deficiency. J. Exp. Med. 2019, 216, 2038–2056. [Google Scholar] [CrossRef]
- Sakata, S.; Tsumura, M.; Matsubayashi, T.; Karakawa, S.; Kimura, S.; Tamaura, M.; Okano, T.; Naruto, T.; Mizoguchi, Y.; Kagawa, R.; et al. Autosomal recessive complete STAT1 deficiency caused by compound heterozygous intronic mutations. Int. Immunol. 2020, 32, 663–671. [Google Scholar] [CrossRef]
- Alphonse, N.; Wanford, J.J.; Voak, A.A.; Gay, J.; Venkhaya, S.; Burroughs, O.; Mathew, S.; Lee, T.; Evans, S.L.; Zhao, W.; et al. A family of conserved bacterial virulence factors dampens interferon responses by blocking calcium signaling. Cell 2022, 185, 2354–2369. [Google Scholar] [CrossRef]
- Chen, J.; Jing, H.; Martin-Nalda, A.; Bastard, P.; Rivière, J.G.; Liu, Z.; Colobran, R.; Lee, D.; Tung, W.; Manry, J.; et al. Inborn errors of TLR3- or MDA5-dependent type I IFN immunity in children with enterovirus rhombencephalitis. J. Exp. Med. 2021, 218. [Google Scholar] [CrossRef]
- Sakata, S.; Tsumura, M.; Matsubayashi, T.; Karakawa, S.; Kimura, S.; Tamaura, M.; Okano, T.; Naruto, T.; Mizoguchi, Y.; Kagawa, R.; et al. Autosomal recessive complete STAT1 deficiency caused by compound heterozygous intronic mutations. Int. Immunol. 2020, 32, 663–671. [Google Scholar] [CrossRef]
- Duncan, C.J.A.; Hambleton, S. Human Disease Phenotypes Associated with Loss and Gain of Function Mutations in STAT2: Viral Susceptibility and Type I Interferonopathy. J. Clin. Immunol. 2021, 41, 1446–1456. [Google Scholar] [CrossRef]
- Holland, S.; DeLeo, F.R.; Elloumi, H.Z.; Hsu, A.P.; Uzel, G.; Brodsky, N.; Freeman, A.F.; Demidowich, A.; Davis, J.; Turner, M.L.C.; et al. STAT3 Mutations in the Hyper-IgE Syndrome. N. Engl. J. Med. 2007, 357, 1608–1619. [Google Scholar] [CrossRef] [PubMed]
- Lupov, I.P.; Voiles, L.; Han, L.; Schwartz, A.; De La Rosa, M.; Oza, K.; Pelloso, D.; Sahu, R.P.; Travers, J.B.; Robertson, M.J.; et al. Acquired STAT4 deficiency as a consequence of cancer chemotherapy. Blood 2011, 118, 6097–6106. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Mai, H.; Peng, J.; Zhou, B.; Hou, J.; Jiang, D. STAT4: an immunoregulator contributing to diverse human diseases. Int. J. Biol. Sci. 2020, 16, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.-Z.; Liu, W.; Xia, Y.; Yin, H.-M.; Zhang, C.-Y.; Su, D.; Yan, L.-F.; Gu, A.-H.; Zhou, Y. The pro-inflammatory signalling regulator Stat4 promotes vasculogenesis of great vessels derived from endothelial precursors. Nat. Commun. 2017, 8, 14640. [Google Scholar] [CrossRef] [PubMed]
- Torpey, N.; Maher, S.E.; Bothwell, A.L.M.; Pober, J.S. Interferon α but Not Interleukin 12 Activates STAT4 Signaling in Human Vascular Endothelial Cells. J. Biol. Chem. 2004, 279, 26789–26796. [Google Scholar] [CrossRef]
- Wiedemann, G.M.; Grassmann, S.; Lau, C.M.; Rapp, M.; Villarino, A.V.; Friedrich, C.; Gasteiger, G.; O’shea, J.J.; Sun, J.C. Divergent Role for STAT5 in the Adaptive Responses of Natural Killer Cells. Cell Rep. 2020, 33, 108498–108498. [Google Scholar] [CrossRef]
- Hatami, M.; Salmani, T.; Arsang-Jang, S.; Omrani, M.D.; Mazdeh, M.; Ghafouri-Fard, S.; Sayad, A.; Taheri, M. STAT5a and STAT6 gene expression levels in multiple sclerosis patients. Cytokine 2018, 106, 108–113. [Google Scholar] [CrossRef]
- Sumida, T.S.; Dulberg, S.; Schupp, J.C.; Lincoln, M.R.; Stillwell, H.A.; Axisa, P.-P.; Comi, M.; Unterman, A.; Kaminski, N.; Madi, A.; et al. Type I interferon transcriptional network regulates expression of coinhibitory receptors in human T cells. Nat. Immunol. 2022, 23, 632–642. [Google Scholar] [CrossRef]
- Kanai, T.; Jenks, J.; Nadeau, K.C. The STAT5b Pathway Defect and Autoimmunity. Front. Immunol. 2012, 3, 234. [Google Scholar] [CrossRef]
- Karpathiou, G.; Papoudou-Bai, A.; Ferrand, E.; Dumollard, J.M.; Peoc’h, M. STAT6: a review of a signaling pathway implicated in various diseases with a special emphasis in its usefulness in pathology. Pathol. - Res. Pr. 2021, 223, 153477. [Google Scholar] [CrossRef]
- Shi, H. et al. The inhibition of IL-2/IL-2R gives rise to CD8+ T cell and lymphocyte decrease through JAK1-STAT5 in critical patients with COVID-19 pneumonia. Cell Death Dis 11, 429 (2020).
- Parackova, Z.; Zentsova, I.; Vrabcova, P.; Sediva, A.; Bloomfield, M. Aberrant tolerogenic functions and proinflammatory skew of dendritic cells in STAT1 gain-of-function patients may contribute to autoimmunity and fungal susceptibility. Clin. Immunol. 2023, 246, 109174. [Google Scholar] [CrossRef] [PubMed]
- Bogunovic, D.; Byun, M.; Durfee, L.A.; Abhyankar, A.; Sanal, O.; Mansouri, D.; Salem, S.; Radovanovic, I.; Grant, A.V.; Adimi, P.; et al. Mycobacterial Disease and Impaired IFN-γ Immunity in Humans with Inherited ISG15 Deficiency. Science 2012, 337, 1684–1688. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, L.; Fracella, M.; Frasca, F.; D’auria, A.; Santinelli, L.; Maddaloni, L.; Bugani, G.; Bitossi, C.; Gentile, M.; Ceccarelli, G.; et al. Alterations in the Expression of IFN Lambda, IFN Gamma and Toll-like Receptors in Severe COVID-19 Patients. Microorganisms 2023, 11, 689. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Birkenbach, M.; Chen, S. Patterns of Inflammatory Cell Infiltration and Expression of STAT6 in the Lungs of Patients With COVID-19: An Autopsy Study. Appl. Immunohistochem. Mol. Morphol. 2022, 30, 350–357. [Google Scholar] [CrossRef]
- Meloni, A.; Furcas, M.; Cetani, F.; Marcocci, C.; Falorni, A.; Perniola, R.; Pura, M.; Wolff, A.S.B.; Husebye, E.S.; Lilic, D.; et al. Autoantibodies against Type I Interferons as an Additional Diagnostic Criterion for Autoimmune Polyendocrine Syndrome Type I. J. Clin. Endocrinol. Metab. 2008, 93, 4389–4397. [Google Scholar] [CrossRef]
- Ghale, R.; Spottiswoode, N.; Anderson, M.S.; Mitchell, A.; Wang, G.; Calfee, C.S.; DeRisi, J.L.; Langelier, C.R. Prevalence of type-1 interferon autoantibodies in adults with non-COVID-19 acute respiratory failure. Respir. Res. 2022, 23, 1–4. [Google Scholar] [CrossRef]
- Hong, G.H.; Ortega-Villa, A.M.; Hunsberger, S.; Chetchotisakd, P.; Anunnatsiri, S.; Mootsikapun, P.; Rosen, L.B.; Zerbe, C.S.; Holland, S.M. Natural History and Evolution of Anti-Interferon-γ Autoantibody-Associated Immunodeficiency Syndrome in Thailand and the United States. Clin. Infect. Dis. 2019, 71, 53–62. [Google Scholar] [CrossRef]
- Martin-Sancho, L. et al. Functional landscape of SARS-CoV-2 cellular restriction. Mol Cell 81, 2656-2668.e8 (2021).
- Brass, A.L.; Huang, I.-C.; Benita, Y.; John, S.P.; Krishnan, M.N.; Feeley, E.M.; Ryan, B.J.; Weyer, J.L.; van der Weyden, L.; Fikrig, E.; et al. The IFITM Proteins Mediate Cellular Resistance to Influenza A H1N1 Virus, West Nile Virus, and Dengue Virus. Cell 2009, 139, 1243–1254. [Google Scholar] [CrossRef]
- Compton, A. A. et al. Natural mutations in IFITM3 modulate post-translational regulation and toggle antiviral specificity. EMBO Rep 17, 1657–1671 (2016).
- John, S.P.; Chin, C.R.; Perreira, J.M.; Feeley, E.M.; Aker, A.M.; Savidis, G.; Smith, S.E.; Elia, A.E.H.; Everitt, A.R.; Vora, M.; et al. The CD225 Domain of IFITM3 Is Required for both IFITM Protein Association and Inhibition of Influenza A Virus and Dengue Virus Replication. J. Virol. 2013, 87, 7837–7852. [Google Scholar] [CrossRef]
- Klein, S.; Golani, G.; Lolicato, F.; Lahr, C.; Beyer, D.; Herrmann, A.; Wachsmuth-Melm, M.; Reddmann, N.; Brecht, R.; Hosseinzadeh, M.; et al. IFITM3 blocks influenza virus entry by sorting lipids and stabilizing hemifusion. Cell Host Microbe 2023, 31, 616–633. [Google Scholar] [CrossRef]
- Yount, J.S.; Zani, A.; Kenney, A.D. Interferon-induced transmembrane protein 3 (IFITM3) limits lethality of SARS-CoV-2 in mice. 2022, 208, 51.17–51.17. [CrossRef]
- Hur, J.-Y. et al. The innate immunity protein IFITM3 modulates γ-secretase in Alzheimer’s disease. Nature 586, 735–740 (2020).
- Pidugu, V.K.; Pidugu, H.B.; Wu, M.-M.; Liu, C.-J.; Lee, T.-C. Emerging Functions of Human IFIT Proteins in Cancer. Front. Mol. Biosci. 2019, 6, 148. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.K.S.; Xu, D.; Licursi, M.; Joyce, M.A.; Saffran, H.A.; Liu, K.; Gohda, J.; Tyrrell, D.L.; Kawaguchi, Y.; Hirasawa, K. Interferon regulatory factor 3 mediates effective antiviral responses to human coronavirus 229E and OC43 infection. Front. Immunol. 2023, 14, 930086. [Google Scholar] [CrossRef] [PubMed]
- Cao, D. et al. The SARS-CoV-2 papain-like protease suppresses type I interferon responses by deubiquitinating STING. Sci Signal 16, (2023).
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.-H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Auto-antibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, D.; Mezidi, M.; Bastard, P.; Perret, M.; Saker, K.; Fabien, N.; Pescarmona, R.; Lombard, C.; Walzer, T.; Casanova, J.; et al. Antibodies against type I interferon: detection and association with severe clinical outcome in COVID-19 patients. Clin. Transl. Immunol. 2021, 10, e1327. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zheng, F.; Li, L.; Jin, Y.; Luo, Y.; Li, Z.; Zeng, J.; Tang, L.; Li, Z.; Xia, N.; et al. A Chinese host genetic study discovered IFNs and causality of laboratory traits on COVID-19 severity. iScience 2021, 24, 103186. [Google Scholar] [CrossRef]
- Yu, R.; Zhu, B.; Chen, D. Type I interferon-mediated tumor immunity and its role in immunotherapy. Cell. Mol. Life Sci. 2022, 79, 1–24. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, D.; Cui, X.; Dai, Y.; Wang, C.; Feng, W.; Lv, X.; Li, Y.; Wang, L.; Ru, Y.; et al. Loss of IRF7 accelerates acute myeloid leukemia progression and induces VCAM1-VLA-4 mediated intracerebral invasion. Oncogene 2022, 41, 2303–2314. [Google Scholar] [CrossRef]
- Lin, C.-F. et al. Escape from IFN-γ-dependent immunosurveillance in tumorigenesis. J Biomed Sci 24, 10 (2017).
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in tumor progression and regression: a review. Biomark. Res. 2020, 8, 1–16. [Google Scholar] [CrossRef]
- Yu, R.; Zhu, B.; Chen, D. Type I interferon-mediated tumor immunity and its role in immunotherapy. Cell. Mol. Life Sci. 2022, 79, 1–24. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, Y.; Wang, Y.; Zhang, M.; Sun, W.; Dai, T.; Wang, A.; Wu, X.; Zhang, S.; Wang, S.; et al. A Dual Role of Type I Interferons in Antitumor Immunity. Adv. Biosyst. 2020, 4, e1900237. [Google Scholar] [CrossRef]
- Lauber, C.; Vieyres, G.; Terczyńska-Dyla, E.; Anggakusuma, X.; Dijkman, R.; Gad, H.H.; Akhtar, H.; Geffers, R.; Vondran, F.W.R.; Thiel, V.; et al. Transcriptome analysis reveals a classical interferon signature induced by IFNλ4 in human primary cells. Genes Immun. 2015, 16, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Ruetsch, C.; Brglez, V.; Crémoni, M.; Zorzi, K.; Fernandez, C.; Boyer-Suavet, S.; Benzaken, S.; Demonchy, E.; Risso, K.; Courjon, J.; et al. Functional Exhaustion of Type I and II Interferons Production in Severe COVID-19 Patients. Front. Med. 2021, 7, 603961. [Google Scholar] [CrossRef] [PubMed]
- Scagnolari, C.; Pierangeli, A.; Frasca, F.; Bitossi, C.; Viscido, A.; Oliveto, G.; Scordio, M.; Mazzuti, L.; Di Carlo, D.; Gentile, M.; et al. Differential induction of type I and III interferon genes in the upper respiratory tract of patients with coronavirus disease 2019 (COVID-19). Virus Res. 2021, 295, 198283–198283. [Google Scholar] [CrossRef]
- Contoli, M.; Papi, A.; Tomassetti, L.; Rizzo, P.; Sega, F.V.D.; Fortini, F.; Torsani, F.; Morandi, L.; Ronzoni, L.; Zucchetti, O.; et al. Blood Interferon-α Levels and Severity, Outcomes, and Inflammatory Profiles in Hospitalized COVID-19 Patients. Front. Immunol. 2021, 12, 648004. [Google Scholar] [CrossRef] [PubMed]
- López-Rodríguez, J.C.; Hancock, S.J.; Li, K.; Crotta, S.; Barrington, C.; Suárez-Bonnet, A.; Priestnall, S.L.; Aubé, J.; Wack, A.; Klenerman, P.; et al. Type I interferons drive MAIT cell functions against bacterial pneumonia. J. Exp. Med. 2023, 220. [Google Scholar] [CrossRef] [PubMed]
- Syedbasha, M.; Bonfiglio, F.; Linnik, J.; Stuehler, C.; Wüthrich, D.; Egli, A. Interferon-λ Enhances the Differentiation of Naive B Cells into Plasmablasts via the mTORC1 Pathway. Cell Rep. 2020, 33, 108211. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xue, M.; Fu, F.; Yin, L.; Feng, L.; Liu, P. IFN-Lambda 3 Mediates Antiviral Protection Against Porcine Epidemic Diarrhea Virus by Inducing a Distinct Antiviral Transcript Profile in Porcine Intestinal Epithelia. Front. Immunol. 2019, 10, 2394. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.; Maleh, P.A.; Bagherzadeh, M.; Moulana, Z.; Sepidarkish, M.; Rezanejad, M.; Mirzakhani, M.; Ebrahimpour, S.; Ghorbani, H.; Ahmadnia, Z.; et al. Linkage of Lambda Interferons in Protection Against Severe COVID-19. J. Interf. Cytokine Res. 2021, 41, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Syedbasha, M.; Linnik, J.; Santer, D.; O'Shea, D.; Barakat, K.; Joyce, M.; Khanna, N.; Tyrrell, D.L.; Houghton, M.; Egli, A. An ELISA Based Binding and Competition Method to Rapidly Determine Ligand-receptor Interactions. J. Vis. Exp. 2016, e53575–e53575. [Google Scholar] [CrossRef]
- da Silva, J.; Hilzendeger, C.; Moermans, C.; Schleich, F.; Henket, M.; Kebadze, T.; Mallia, P.; Edwards, M.R.; Johnston, S.L.; Louis, R. Raised interferon-β, type 3 interferon and interferon-stimulated genes - evidence of innate immune activation in neutrophilic asthma. Clin. Exp. Allergy 2016, 47, 313–323. [Google Scholar] [CrossRef]
- Ye, L.; Schnepf, D.; Ohnemus, A.; Ong, L.C.; Gad, H.H.; Hartmann, R.; Lycke, N.; Staeheli, P. Interferon-λ Improves the Efficacy of Intranasally or Rectally Administered Influenza Subunit Vaccines by a Thymic Stromal Lymphopoietin-Dependent Mechanism. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.W.; Constant, D.A.; Nice, T.J. Interferon Lambda in the Pathogenesis of Inflammatory Bowel Diseases. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Villamayor, L.; López-García, D.; Rivero, V.; Martínez-Sobrido, L.; Nogales, A.; DeDiego, M.L. The IFN-stimulated gene IFI27 counteracts innate immune responses after viral infections by interfering with RIG-I signaling. Front. Microbiol. 2023, 14, 1176177. [Google Scholar] [CrossRef]
- Villamayor, L.; Rivero, V.; López-García, D.; Topham, D.J.; Martínez-Sobrido, L.; Nogales, A.; DeDiego, M.L. Interferon alpha inducible protein 6 is a negative regulator of innate immune responses by modulating RIG-I activation. Front. Immunol. 2023, 14, 1105309. [Google Scholar] [CrossRef] [PubMed]
- Chilunda, V.; Martinez-Aguado, P.; Xia, L.C.; Cheney, L.; Murphy, A.; Veksler, V.; Ruiz, V.; Calderon, T.M.; Berman, J.W. Transcriptional Changes in CD16+ Monocytes May Contribute to the Pathogenesis of COVID-19. Front. Immunol. 2021, 12, 665773. [Google Scholar] [CrossRef] [PubMed]
- Fernández, D.J.; Hess, S.; Knobeloch, K.-P. Strategies to Target ISG15 and USP18 Toward Therapeutic Applications. Front. Chem. 2020, 7, 923. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, P. et al. Expression profiles of human interferon-alpha and interferon-lambda subtypes are ligand- and cell-dependent. Immunol Cell Biol 90, 774–783 (2012).
- Henig, N.; Avidan, N.; Mandel, I.; Staun-Ram, E.; Ginzburg, E.; Paperna, T.; Pinter, R.Y.; Miller, A. Interferon-Beta Induces Distinct Gene Expression Response Patterns in Human Monocytes versus T cells. PLOS ONE 2013, 8, e62366. [Google Scholar] [CrossRef]
- Forero, A.; Ozarkar, S.; Li, H.; Lee, C.H.; Hemann, E.A.; Nadjsombati, M.S.; Hendricks, M.R.; So, L.; Green, R.; Roy, C.N.; et al. Differential Activation of the Transcription Factor IRF1 Underlies the Distinct Immune Responses Elicited by Type I and Type III Interferons. Immunity 2019, 51, 451–464. [Google Scholar] [CrossRef]
- Zhao, M.; Li, L.; Zhai, L.; Yue, Q.; Liu, H.; Ren, S.; Jiang, X.; Gao, F.; Bai, S.; Li, H.; et al. Comparative Transcriptomic and Proteomic Analyses Prove that IFN-λ1 is a More Potent Inducer of ISGs than IFN-α against Porcine Epidemic Diarrhea Virus in Porcine Intestinal Epithelial Cells. J. Proteome Res. 2020, 19, 3697–3707. [Google Scholar] [CrossRef]
- Soday, L.; Potts, M.; Hunter, L.M.; Ravenhill, B.J.; Houghton, J.W.; Williamson, J.C.; Antrobus, R.; Wills, M.R.; Matheson, N.J.; Weekes, M.P. Comparative Cell Surface Proteomic Analysis of the Primary Human T Cell and Monocyte Responses to Type I Interferon. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Awwad, M.H.S.; Mahmoud, A.; Bruns, H.; Echchannaoui, H.; Kriegsmann, K.; Lutz, R.; Raab, M.S.; Bertsch, U.; Munder, M.; Jauch, A.; et al. Selective elimination of immunosuppressive T cells in patients with multiple myeloma. Leukemia 2021, 35, 2602–2615. [Google Scholar] [CrossRef]
- Simmons, D.P.; Nguyen, H.N.; Gomez-Rivas, E.; Jeong, Y.; Jonsson, A.H.; Chen, A.F.; Lange, J.K.; Dyer, G.S.; Blazar, P.; Earp, B.E.; et al. SLAMF7 engagement superactivates macrophages in acute and chronic inflammation. Sci. Immunol. 2022, 7, eabf2846–eabf2846. [Google Scholar] [CrossRef] [PubMed]
- Attur, M.; Zhou, H.; Samuels, J.; Krasnokutsky, S.; Yau, M.; Scher, J.U.; Doherty, M.; Wilson, A.G.; Bencardino, J.; Hochberg, M.; et al. Interleukin 1 receptor antagonist (IL1RN) gene variants predict radiographic severity of knee osteoarthritis and risk of incident disease. Ann. Rheum. Dis. 2019, 79, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Barragan, L. et al. Plasmacytoid dendritic cells produce type I interferon and reduce viral replication in 1 airway epithelial cells after SARS-CoV-2 infection 2 3 4. [CrossRef]
- Georgiades, J.; Babiuch, L. Results of the prolonged use of natural human interferon alpha (nHuIFNα) lozenges in the treatment of human immune deficiency virus infection. Cytokine 1994, 6, 572. [Google Scholar] [CrossRef]
- Pereda, R.; González, D.; Rivero, H.B.; Rivero, J.C.; Pérez, A.; Lopez, L.D.R.; Mezquia, N.; Venegas, R.; Betancourt, J.R.; Domínguez, R.E.; et al. Therapeutic Effectiveness of Interferon Alpha 2b Treatment for COVID-19 Patient Recovery. J. Interf. Cytokine Res. 2020, 40, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Pierangeli, A. et al. Comparison by Age of the Local Interferon Response to SARS-CoV-2 Suggests a Role for IFN-ε and -ω. Front Immunol 13, 873232 (2022).
- Emilie, D.; Burgard, M.; Lascoux-Combe, C.; Laughlin, M.; Krzysiek, R.; Pignon, C.; Rudent, A.; Molina, J.-M.; Livrozet, J.-M.; Souala, F.; et al. Early control of HIV replication in primary HIV-1 infection treated with antiretroviral drugs and pegylated IFNα: results from the Primoferon A (ANRS 086) Study. AIDS 2001, 15, 1435–1437. [Google Scholar] [CrossRef]
- Zhou, Q.; MacArthur, M.R.; He, X.; Wei, X.; Zarin, P.; Hanna, B.S.; Wang, Z.-H.; Xiang, X.; Fish, E.N. Interferon-α2b Treatment for COVID-19 Is Associated with Improvements in Lung Abnormalities. Viruses 2020, 13, 44. [Google Scholar] [CrossRef]
- Zhou, Q.; MacArthur, M.R.; He, X.; Wei, X.; Zarin, P.; Hanna, B.S.; Wang, Z.-H.; Xiang, X.; Fish, E.N. Interferon-α2b Treatment for COVID-19 Is Associated with Improvements in Lung Abnormalities. Viruses 2020, 13, 44. [Google Scholar] [CrossRef]
- Davoudi-Monfared, E.; Rahmani, H.; Khalili, H.; Hajiabdolbaghi, M.; Salehi, M.; Abbasian, L.; Kazemzadeh, H.; Yekaninejad, M.S. A Randomized Clinical Trial of the Efficacy and Safety of Interferon β-1a in Treatment of Severe COVID-19. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Tsiakos, K.; Tsakiris, A.; Tsibris, G.; Voutsinas, P.-M.; Panagopoulos, P.; Kosmidou, M.; Petrakis, V.; Gravvani, A.; Gkavogianni, T.; Klouras, E.; et al. Early Start of Oral Clarithromycin Is Associated with Better Outcome in COVID-19 of Moderate Severity: The ACHIEVE Open-Label Single-Arm Trial. Infect. Dis. Ther. 2021, 10, 2333–2351. [Google Scholar] [CrossRef]
- Shalamova, L. et al. Omicron variant of SARS-CoV-2 exhibits an increased resilience to the antiviral type I interferon response. PNAS Nexus 1, (2022).
- Lee-Kirsch, M.A. The Type I Interferonopathies. Annu. Rev. Med. 2017, 68, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.C.; Jeong, Y.; Lee, S.; Jang, H.; Zheng, A.; Kwon, S.; Repine, J.E. Nasopharyngeal Type-I Interferon for Immediately Available Prophylaxis Against Emerging Respiratory Viral Infections. Front. Immunol. 2021, 12, 660298. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.J.; Ashkar, A.A. The Dual Nature of Type I and Type II Interferons. Front. Immunol. 2018, 9, 2061. [Google Scholar] [CrossRef]
- Lee, J.S.; Shin, E.-C. The type I interferon response in COVID-19: implications for treatment. Nat. Rev. Immunol. 2020, 20, 585–586. [Google Scholar] [CrossRef]
- Chong, Z. et al. Nasally delivered interferon-λ protects mice against infection by SARS-CoV-2 variants including Omicron. Cell Rep 39, 110799 (2022).
- Cooles, F.A.H.; Tarn, J.; Lendrem, D.W.; Naamane, N.; Lin, C.M.; Millar, B.; Maney, N.J.; E Anderson, A.; Thalayasingam, N.; Diboll, J.; et al. Interferon-α-mediated therapeutic resistance in early rheumatoid arthritis implicates epigenetic reprogramming. Rheumatol. 2022, 81, 1214–1223. [Google Scholar] [CrossRef]
- Borden, E. C. Interferons α and β in cancer: therapeutic opportunities from new insights. Nat Rev Drug Discov 18, 219–234 (2019).
- Borden, E.C.; Sen, G.C.; Uze, G.; Silverman, R.H.; Ransohoff, R.M.; Foster, G.R.; Stark, G.R. Interferons at age 50: past, current and future impact on biomedicine. Nat. Rev. Drug Discov. 2007, 6, 975–990. [Google Scholar] [CrossRef] [PubMed]
- Niles, M. A. et al. Macrophages and Dendritic Cells Are Not the Major Source of Pro-Inflammatory Cytokines Upon SARS-CoV-2 Infection. Front Immunol 12, (2021).
- Trouillet-Assant, S.; Viel, S.; Gaymard, A.; Pons, S.; Richard, J.-C.; Perret, M.; Villard, M.; Brengel-Pesce, K.; Lina, B.; Mezidi, M.; et al. Type I IFN immunoprofiling in COVID-19 patients. J. Allergy Clin. Immunol. 2020, 146, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Yuen, C.-K.; Lam, J.-Y.; Wong, W.-M.; Mak, L.-F.; Wang, X.; Chu, H.; Cai, J.-P.; Jin, D.-Y.; To, K.K.-W.; Chan, J.F.-W.; et al. SARS-CoV-2 nsp13, nsp14, nsp15 and orf6 function as potent interferon antagonists. Emerg. Microbes Infect. 2020, 9, 1418–1428. [Google Scholar] [CrossRef]
- Vazquez, C. et al. SARS-CoV-2 viral proteins NSP1 and NSP13 inhibit interferon activation through distinct mechanisms. PLoS One 16, e0253089 (2021).
- Kumar, A. et al. SARS-CoV-2 Nonstructural Protein 1 Inhibits the Interferon Response by Causing Depletion of Key Host Signaling Factors. J Virol 95, (2021).
- Rincon-Arevalo, H.; Aue, A.; Ritter, J.; Szelinski, F.; Khadzhynov, D.; Zickler, D.; Stefanski, L.; Lino, A.C.; Körper, S.; Eckardt, K.; et al. Altered increase in STAT1 expression and phosphorylation in severe COVID-19. Eur. J. Immunol. 2021, 52, 138–148. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, Y.; Chen, Y.; Wang, J.; Feng, H.; Wei, Q.; Zhao, S.; Yang, S.; Ma, H.; Liu, D.; et al. Antiviral activity of porcine interferon delta 8 against pesudorabies virus in vitro. Int. J. Biol. Macromol. 2021, 177, 10–18. [Google Scholar] [CrossRef]
- Yin, L.; Liu, X.; Hu, D.; Luo, Y.; Zhang, G.; Liu, P. Swine Enteric Coronaviruses (PEDV, TGEV, and PDCoV) Induce Divergent Interferon-Stimulated Gene Responses and Antigen Presentation in Porcine Intestinal Enteroids. Front. Immunol. 2022, 12, 826882. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ke, H.; Blikslager, A.; Fujita, T.; Yoo, D. Type III Interferon Restriction by Porcine Epidemic Diarrhea Virus and the Role of Viral Protein nsp1 in IRF1 Signaling. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Zhang, Q.; Yoo, D. Immune evasion of porcine enteric coronaviruses and viral modulation of antiviral innate signaling. Virus Res. 2016, 226, 128–141. [Google Scholar] [CrossRef]
- Iaconis, G. et al. Rotavirus NSP1 Inhibits Type I and Type III Interferon Induction. Viruses 13, 589 (2021).
- Fisher, T. et al. Parsing the role of NSP1 in SARS-CoV-2 infection. Cell Rep 39, 110954 (2022).
- Ferreira-Gomes, M. et al. SARS-CoV-2 in severe COVID-19 induces a TGF-β-dominated chronic immune response that does not target itself. Nat Commun 12, (2021).
- Barros-Martins, J.; Förster, R.; Bošnjak, B. NK cell dysfunction in severe COVID-19: TGF-β-induced downregulation of integrin beta-2 restricts NK cell cytotoxicity. Signal Transduct. Target. Ther. 2022, 7, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, M.; Tizian, C.; Ferreira-Gomes, M.; Niemeyer, D.; Jones, T.C.; Heinrich, F.; Frischbutter, S.; Angermair, S.; Hohnstein, T.; Mattiola, I.; et al. Untimely TGFβ responses in COVID-19 limit antiviral functions of NK cells. Nature 2021, 600, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Barros-Martins, J.; Förster, R.; Bošnjak, B. NK cell dysfunction in severe COVID-19: TGF-β-induced downregulation of integrin beta-2 restricts NK cell cytotoxicity. Signal Transduct. Target. Ther. 2022, 7, 1–3. [Google Scholar] [CrossRef]
- Matic, S. et al. SARS-CoV-2 infection induces mixed M1/M2 phenotype in circulating monocytes and alterations in both dendritic cell and monocyte subsets. PLoS One 15, e0241097–e0241097 (2020).
- Setaro, A. C. & Gaglia, M. M. All hands on deck: SARS-CoV-2 proteins that block early anti-viral interferon responses. Curr Res Virol Sci 2, 100015 (2021).
- Miorin, L. et al. SARS-CoV-2 Orf6 hijacks Nup98 to block STAT nuclear import and antagonize interferon signaling. Proceedings of the National Academy of Sciences 117, 28344–28354 (2020).
- Wu, J. et al. SARS-CoV-2 ORF9b inhibits RIG-I-MAVS antiviral signaling by interrupting K63-linked ubiquitination of NEMO. Cell Rep 34, 108761 (2021).
- Guo, K. et al. Interferon resistance of emerging SARS-CoV-2 variants. Proceedings of the National Academy of Sciences 119, (2022).
- Brisse, M.; Ly, H. Comparative Structure and Function Analysis of the RIG-I-Like Receptors: RIG-I and MDA5. Front. Immunol. 2019, 10, 1586. [Google Scholar] [CrossRef]
- Prelli Bozzo, C. et al. IFITM proteins promote SARS-CoV-2 infection and are targets for virus inhibition in vitro. Nat Commun 12, 4584 (2021).
- McKellar, J. , Rebendenne, A., Wencker, M., Moncorgé, O. & Goujon, C. Mammalian and Avian Host Cell Influenza A Restriction Factors. Viruses 13, 522 (2021).
- Cervantes, J.L.; Weinerman, B.; Basole, C.; Salazar, J.C. TLR8: the forgotten relative revindicated. Cell. Mol. Immunol. 2012, 9, 434–438. [Google Scholar] [CrossRef]
- Lee, A.J.; Ashkar, A.A. The Dual Nature of Type I and Type II Interferons. Front. Immunol. 2018, 9, 2061. [Google Scholar] [CrossRef]
- Sidahmed, A.M.; León, A.J.; Bosinger, S.E.; Banner, D.; Danesh, A.; Cameron, M.J.; Kelvin, D.J. CXCL10 contributes to p38-mediated apoptosis in primary T lymphocytes in vitro. Cytokine 2012, 59, 433–441. [Google Scholar] [CrossRef]
- Khalil, B.A.; Elemam, N.M.; Maghazachi, A.A. Chemokines and chemokine receptors during COVID-19 infection. Comput. Struct. Biotechnol. J. 2021, 19, 976–988. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Casillas, A. , Redwan, E. M. & Uversky, V. N. SARS-CoV-2: A Master of Immune Evasion. Biomedicines 10, 1339 (2022).
- Yoo, J.-S. et al. SARS-CoV-2 inhibits induction of the MHC class I pathway by targeting the STAT1-IRF1-NLRC5 axis. Nat Commun 12, 6602 (2021).
- Rashid, F. et al. Roles and functions of SARS-CoV-2 proteins in host immune evasion. Front Immunol 13, 940756 (2022).
- Glennon-Alty, L.; Moots, R.J.; Edwards, S.W.; Wright, H.L. Type I interferon regulates cytokine-delayed neutrophil apoptosis, reactive oxygen species production and chemokine expression. Clin. Exp. Immunol. 2020, 203, 151–159. [Google Scholar] [CrossRef]
- Zanoni, I.; Granucci, F.; Broggi, A. Interferon (IFN)-λ Takes the Helm: Immunomodulatory Roles of Type III IFNs. Front. Immunol. 2017, 8, 1661. [Google Scholar] [CrossRef]
- Margaroli, C.; Fram, T.R.; Sharma, N.S.; Patel, S.B.; Tipper, J.; Robison, S.W.; Russell, D.W.; Fortmann, S.D.; Banday, M.M.; Soto-Vazquez, Y.M.; et al. Interferon-dependent signaling is critical for viral clearance in airway neutrophils. J. Clin. Investig. 2023, 8. [Google Scholar] [CrossRef]
- Glennon-Alty, L.; Moots, R.J.; Edwards, S.W.; Wright, H.L. Type I interferon regulates cytokine-delayed neutrophil apoptosis, reactive oxygen species production and chemokine expression. Clin. Exp. Immunol. 2020, 203, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Hijano, D.R.; Vu, L.D.; Kauvar, L.M.; Tripp, R.A.; Polack, F.P.; Cormier, S.A. Role of Type I Interferon (IFN) in the Respiratory Syncytial Virus (RSV) Immune Response and Disease Severity. Front. Immunol. 2019, 10, 566. [Google Scholar] [CrossRef] [PubMed]
- Munir, S.; Hillyer, P.; Le Nouën, C.; Buchholz, U.J.; Rabin, R.L.; Collins, P.L.; Bukreyev, A. Respiratory Syncytial Virus Interferon Antagonist NS1 Protein Suppresses and Skews the Human T Lymphocyte Response. PLOS Pathog. 2011, 7, e1001336. [Google Scholar] [CrossRef]
- Mo, S. et al. Respiratory syncytial virus activates Rab5a to suppress IRF1-dependent IFN-λ production, subverting the antiviral defense of airway epithelial cells. J Virol 95, e02333-20 (2021).
- Mortezaee, K. & Majidpoor, J. Cellular immune states in SARS-CoV-2-induced disease. Front Immunol 13, 1016304 (2022).
- De Maeyer, E. & De Maeyer-Guignard, J. Type I Interferons. Int Rev Immunol 17, 53–73 (1998).
- Vremec, D.; O'Keeffe, M.; Hochrein, H.; Fuchsberger, M.; Caminschi, I.; Lahoud, M.; Shortman, K. Production of interferons by dendritic cells, plasmacytoid cells, natural killer cells, and interferon-producing killer dendritic cells. Blood 2006, 109, 1165–1173. [Google Scholar] [CrossRef]
- De Cecco, M.; Ito, T.; Petrashen, A.P.; Elias, A.E.; Skvir, N.J.; Criscione, S.W.; Caligiana, A.; Brocculi, G.; Adney, E.M.; Boeke, J.D.; et al. L1 drives IFN in senescent cells and promotes age-associated inflammation. Nature 2019, 566, 73–78. [Google Scholar] [CrossRef]
- Li, Y. et al. SARS-CoV-2-encoded inhibitors of human LINE-1 retrotransposition. J Med Virol 95, (2023).
- Crow, M.K. Long interspersed nuclear elements (LINE-1): Potential triggers of systemic autoimmune disease. Autoimmunity 2009, 43, 7–16. [Google Scholar] [CrossRef]
- Zhou, H.; Møhlenberg, M.; Terczyńska-Dyla, E.; Winther, K.G.; Hansen, N.H.; Vad-Nielsen, J.; Laloli, L.; Dijkman, R.; Nielsen, A.L.; Gad, H.H.; et al. The IFNL4 Gene Is a Noncanonical Interferon Gene with a Unique but Evolutionarily Conserved Regulation. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Marongiu, L.; Protti, G.; Facchini, F.A.; Valache, M.; Mingozzi, F.; Ranzani, V.; Putignano, A.R.; Salviati, L.; Bevilacqua, V.; Curti, S.; et al. Maturation signatures of conventional dendritic cell subtypes in COVID-19 suggest direct viral sensing. Eur. J. Immunol. 2021, 52, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Brown, B. Innate and Adaptive Immunity during SARS-CoV-2 Infection: Biomolecular Cellular Markers and Mechanisms. Vaccines (Basel) 2023, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Sposito, B.; Broggi, A.; Pandolfi, L.; Crotta, S.; Clementi, N.; Ferrarese, R.; Sisti, S.; Criscuolo, E.; Spreafico, R.; Long, J.M.; et al. The interferon landscape along the respiratory tract impacts the severity of COVID-19. Cell 2021, 184, 4953–4968. [Google Scholar] [CrossRef] [PubMed]
- Bourgoin, P. CD169 and CD64 could help differentiate bacterial from CoVID -19 or other viral infections in the Emergency Department. Cytometry Part A 2021, 99, 435–445. [Google Scholar] [CrossRef]
- Filipi, M. & Jack, S. Interferons in the Treatment of Multiple Sclerosis. Int J MS Care 2020, 22, 165–172. [Google Scholar]
- Errante, P.R.; Frazao, J.B.; Condino-Neto, A. The Use of Interferon-Gamma Therapy in Chronic Granulomatous Disease. Recent Patents Anti-Infective Drug Discov. 2008, 3, 225–230. [Google Scholar] [CrossRef]
- Li, S.-F.; Zhao, F.-R.; Shao, J.-J.; Xie, Y.-L.; Chang, H.-Y.; Zhang, Y.-G. Interferon-omega: Current status in clinical applications. Int. Immunopharmacol. 2017, 52, 253–260. [Google Scholar] [CrossRef]
- He, X. Anti-Lyssaviral Activity of Interferons κ and ω from the Serotine Bat, Eptesicus serotinus. J Virol 2014, 88, 5444–5454. [Google Scholar] [CrossRef]
- Pavlovich, S.S.; Darling, T.; Hume, A.J.; Davey, R.A.; Feng, F.; Mühlberger, E.; Kepler, T.B. Egyptian Rousette IFN-ω Subtypes Elicit Distinct Antiviral Effects and Transcriptional Responses in Conspecific Cells. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



