
Submitted:
26 July 2023
Posted:
27 July 2023
You are already at the latest version
Abstract
The Myc gene, widely studied in numerous cancer types, serves as a pivotal regulator of crucial cellular processes implicated in tumorigenesis, such as cell growth, proliferation, invasion, metastasis, and therapy resistance. LncRNAs have been identified as relevant partners of Myc. LncRNAs modulate chromatin structure and function, transcription, splicing, stability, and translation of mRNAs, thereby emerging as pivotal regulators in cell differentiation, metabolism, signaling. An intricate crosstalk exists between Myc and lncRNAs in a wide range of cancers, wherein they can physically and functionally interact through direct and indirect mechanisms. Some lncRNAs can regulate Myc activity at post-transcriptional level, affecting its stability, translation, or interactions with other proteins, thus acting as oncogenes, while others act as tumor suppressors. On the other hand, Myc can target numerous lncRNA genes and modulate their expression, affecting the expression of downstream targets involved in tumorigenesis. Given the significant role of the Myc-lncRNA network in various cancer types, exploring therapeutic strategies targeting these crucial cellular regulators is of utmost importance. This review article focuses on breast cancer, analyzing those lncRNAs influencing Myc as well as the Myc-regulated lncRNAs.
Keywords:
MYC
; LncRNAs
; miRNA
; Breast Cancer
1. Introduction
The c- Myelocytomatosis viral oncogene homolog (Myc) gene and protein family act as an essential player in cancer pathophysiology. C-Myc protein is 62 kDa and contains 439 aminoacid residues. It contains several functional domains that facilitate the binding with a variety of other proteins. In the N-terminal region, there are a transcription transactivation domain (TAD) and three highly conserved Myc boxes (MB0, MBI, and MBII), which play a key role in transcriptional regulation and protein degradation [1]. In the middle part of the protein, there is a PEST domain (containing proline and glutamate as well as serine and threonine), a nuclear localization sequence (NLS), and three conserved Myc boxes (MBIIIa, MBIIIb, and MBIV), which are involved in cell transformation, transcription, and apoptosis [2]. In the C-terminal region, Myc family expresses basic helix-loop-helix leucine zipper (bHLHZip) DNA binding domains that regulate the expression of many genes involved in cell cycle progression, DNA replication, apoptosis, autophagy, ribosome biogenesis, protein synthesis, glycolysis, mitochondrial function, angiogenesis, and cell adhesion [3].
Overall, Myc proteins regulate approximately 2000 to 4000 different genes, as well as control transcription mediated by RNA polymerase I, II and III [4]. The Myc protein family consists of three members: C-Myc (sometimes referred to as MYC), l-Myc (MYCL), and n-Myc (MYCN) [5]. Approximately 70% of human cancers are associated with dysregulation of Myc gene expression [4]. Among all the cancer types, Myc (c-Myc) is the most frequently amplified gene, exhibiting amplification in 21% of tumors, while n-Myc and l-Myc exhibit amplification in 7% [6]. Copy numer gains of Myc gene in solid tumors is the primary mechanism driving c-Myc dependent proliferation of somatic cells and oncogenesis. Three distinct types of Myc amplifications are observed: low copy gain, intermediate copy gain, and a high copy gain. According to The Cancer Genome Atlas (TCGA), Myc is the most commonly amplified in breast cancer (BC), accounting for 61.27% of all samples [7].
The effects of Myc on gene expression depend on its ability to bind to specific DNA sequences, known as E-boxes, in the promoter regions of target genes. This binding leads to gene transcription activation or repression, depending on the context and the specific genes. TRRAP (Transactivation/ Transformation-Domain), CBP/p300 (cAMP-Response-Element-Binding Protein), WDR5 (WD Repeat Domain 5), TBP (TATA-Binding Protein), P-TEFb (Positive Transcription Elongation Factor b), ASH2L (Set1/Ash2 Histone Methyltransferase Complex Subunit ASH2) are also transcription activator factors that interact with Myc and regulate gene expression [2].
Myc family proto-oncogenes are commonly altered in human cancers and represent an attractive therapeutic target [4]. Myc can regulate the expression of genes involved in cell cycle control (cyclins, CDKs), cellular metabolism (glucose metabolism, glutamine metabolism), apoptosis (Bcl-2, BAX), and other crucial pathways for cancer development [8]. In addition to maintaining the tumor microenvironment, Myc regulates tissue remodeling, angiogenesis, and inflammation, even when not overexpressed [9]. The ability of Myc to activate tumor-associated macrophages and regulate epithelial-to-mesenchymal transition (EMT) effectors may indeed contribute to the acquisition of malignant features [10].
As an oncogene, Myc can be activated by multiple mechanisms in cancer. Chromosomal rearrangement is believed to be the most common Myc genetic alteration. Several other mechanisms contribute to this process, including point mutations, transcriptional regulation, mRNA/protein stabilization, and protein overexpression. The molecular mechanism underlying increased Myc protein levels in certain cancer types without a corresponding increase in Myc mRNA levels is unclear. Three hypotheses are proposed for this process: Myc protein stability by post-translational modifications, Myc mRNA stability by RNA binding proteins, and increased efficiency of Myc translation by ncRNA and lncRNAs [11,12].
2. lncRNAs function
In the cell, non-coding RNAs (ncRNAs) are a heterogeneous group of RNA molecules that have multiple regulatory functions, despite not encoding proteins. There are many different types of ncRNAs, which can be categorized by their size, origin, structure, function, and interaction. In general, ncRNAs are categorized based on their size: small ncRNAs (<200 nt), and long ncRNAs (>200 nt). Several classes of small non-coding RNAs have been described based on their biogenesis, function and interaction partners, such as microRNAs (miRNAs), small interfering RNAs (siRNAs), piwi-interacting RNAs (piRNAs), small nucleolar RNAs (snorRNAs), small nuclear RNAs (snRNAs) and transfer-derived RNAs (tRFs) [13,14].
Long noncoding RNAs (lncRNAs) are a heterogeneous group of RNA molecules, not usually coding for proteins, which can be classified into different categories based on various criteria, such as their genomic location, biogenesis, structure and function [15]. One of the simplest and most widely used criteria for classifying lncRNAs is their genomic location respect to protein-coding genes (intergenic lncRNAs “lincRNAs”, intronic lncRNAs, antisense lncRNAs, bidirectional lncRNAs, enhancer lncRNAs “eRNAs”, promoter lncRNAs “pRNAs”)[16]. According to the type of RNA polymerase used for lncRNA biogenesis, they can be categorized into three categories. The most common enzyme involved in lncRNA biogenesis appears to be RNA polymerase II [16]. As the second most dominant enzyme in lncRNA biogenesis, RNA polymerase III can transcribe lncRNAs from Alu elements, tRNA genes, and 5S rRNA genes. Only a few lncRNAs are instead synthesized from rRNA genes by RNA polymerase I [17]. Accordingly, a structure-based classification of lncRNAs can divide them into three main categories: linear lncRNAs, circular lncRNAs "circRNAs", and complicated secondary structures lncRNAs. Also, lncRNAs are classified as guide, signaling, decoy, enhancer, or scaffolding based on their mechanism of function (Figure 1). ceRNA targets and recruits transcriptional complexes to specific genes in order to regulate gene expression [15]. A signaling lncRNA acts as a transcription factor and participates in the cell signaling pathway. A decoy lncRNA acts as a sponge RNA, inhibiting transcription factors from binding to their targets. elncRNAs bind to enhancer elements and recruit transcription factors to activate specific target genes. Scaffolding lncRNAs can bring together proteins involved in transcriptional regulation and maintain their interaction [18,19].
LncRNAs can regulate, directly or indirectly, a variety of cancer pathways. lncRNA are involved in a wide range of biological processes, including regulation of gene expression at various levels (transcription, splicing, translation, and epigenetics), chromatin remodeling, cell- and stem cell- fate determination (embryonic development, cell differentiation, and cell proliferation, and survival), maintainance of tissue homeostasis, immune response and regulation of various cellular processes (apoptosis, autophagy, inflammation, cell migration and metabolism) [20,21].
3. Myc and breast cancer
Breast cancer (BC) is the most prevalent cancer among women globally. In 2020, it affected approximately 2.3 million women and resulted in 685,000 deaths worldwide, as reported by the World Health Organization (WHO). Additionally, by the end of that year, there were 7.8 million women living with a BC diagnosis. According to the International Agency for Research on Cancer (IARC), there is a projected 40% increase in new BC cases, surpassing 3 million per year by 2040, along with a 50% rise in deaths, exceeding 1 million per year. The BC Research Foundation (BCRF) reveals that every 14 seconds, a woman somewhere in the world is diagnosed with BC.
In normal somatic cells, Myc activity is tightly controlled by mitogenic signals, dedifferentiation, and stromal, inflammatory, or immune signals [22]. BC cells often overexpress Myc and this can be attributed to genetic and epigenetic factors that result in misregulation of Myc signaling, resulting in the development of BC [23]. Also, Myc expression is deregulated in BC through gene amplification, increased transcription, translation, or increased protein stability [24]. After chemotherapy for BC, Myc amplification was observed in the breast tissues. This indicates that Myc may contribute to chemotherapy resistance in the BC [25]. BC cells expressed high levels of cC-Myc through cross-talk between oestrogen receptors (ER) and human epidermal growth factor receptors 2 (HER2) [26]. The expression of Myc is associated with the down-regulation of inflammatory pathways such as IFN and JAK-STAT signaling in human BC [7]. Also, the increase in Myc expression in a cell during development can also affect the proliferation and apoptosis of neighboring cells [26]. However, BC tumors with high Myc expression are typically interspersed among tumor cells expressing lower Myc levels [27]. Additionally, Myc is the most commonly amplified oncogene in BCs associated with BRCA1/2 mutations [28]. It has been demonstrated that knocking out c-Myc inhibits tumor angiogenesis in BC cells by regulating VEGF [29].
4. Myc and lncRNAs
Myc and lncRNAs play an important role in the development of various cancers, including breast, lung, colorectal, and gastric cancers. Myc is both an upstream regulator and a target of many lncRNAs. Although Myc and lncRNAs have been studied in cancer for many years, their interactions remain unclear. Considering that Myc plays a crucial role in human cancer, we herein explore and summarize the implications of these interactions to better understand interactive pathways in cancer.
Myc-lncRNA interactions are complex and dynamic because they occur at multiple levels. The relationship between Myc and lncRNAs is a critical focal point in understanding cancer development, progression and molecular mechanisms underscoring cancer onset and progression. LncRNAs have the potential to modulate Myc expression and activity through a variety of mechanisms. They can act as molecular scaffolds, forming complexes with Myc and its associated proteins to influence its transcriptional activity. LncRNAs can also directly interact with Myc mRNA, affecting its stability or translation efficiency. Additionally, lncRNAs are capable of regulating Myc expression at the epigenetic level, by modulating histone modifications and chromatin accessibility.
On the other hand, Myc can regulate lncRNA expression and function. Myc can affect lncRNAs by regulating the expression of transcription factors or chromatin modifiers that influence their expression. These reciprocal interactions contribute to the pathogenesis of various types of cancer as they form an intricate regulatory network between Myc and lncRNAs.
This review focuses on the functions of lncRNAs both upstream and downstream of Myc. We will explore how lncRNAs influence Myc expression and activity, as well as how Myc impacts lncRNA expression and function. By elucidating the intricate crosstalk between Myc and lncRNAs, we aim to provide valuable insights into the molecular mechanisms underlying cancer development, with a potential impact on cancer diagnosis, prognosis, and treatment.
5. The relationship between lncRNAs and MYC in BC
The oncogenic transcription factor Myc is known to be dysregulated in many cancers including BC. As discussed previously, abnormal activation of Myc signaling promotes cancer pathogenesis through stimulating cell proliferation, metabolic reprogramming, inhibiting apoptosis and facilitating metastasis. Therefore, elucidating the Myc regulatory factors in cancer is imperative to gain insights into disease mechanisms and identify novel therapeutic approaches.
An emerging class of Myc regulators are long non-coding RNAs (lncRNAs). LncRNAs have been shown to interact with Myc through various mechanisms, forming an intricate network affecting Myc expression, stability and transcriptional functions. For instance, some lncRNAs do act as scaffolds that recruit chromatin modifiers to alter Myc promoter accessibility. Other lncRNAs can directly bind to Myc mRNA and enhance its stability. There is also evidence that certain lncRNAs modulate miRNAs targeting Myc, thereby increasing Myc levels.
Conversely, Myc as a transcription factor can regulate the expression of many lncRNAs involved in BC. By inducing lncRNAs that in turn promote processes like proliferation, angiogenesis and metastasis, Myc amplifies its oncogenic signals. Of note, the crosstalk between Myc and lncRNAs establishes regulatory feedback loops that may profoundly influence BC pathogenesis. A deeper understanding of this relationship may open new avenues to combat BC progression and improve patient outcomes. All data about the function and mechanism between Myc and lncRNA have in BC been summarized in Table 1. Also, the interaction and crosstalk between lncRNA and Myc in BC have been illustrated in Figure 2.
5.1. lncRNAs regulated by MYC
SNHG7
Small Nucleolar RNA Host Gene 7(SNHG7) is an lncRNA that is overexpressed and involved in many types of cancer. The highest level of SNHG7 was found in a BC cell line [30]. There is a positive correlation between SNHG7 expression and tumor stage, lymph node metastasis, poor prognosis, and distant metastasis in BC [31]. SNHG7 expression increases drug resistance and inhibits apoptosis in chemoresistant BC cells [32]. In BC cells, SNHG7 acts as miRNA sponge, increasing Myc levels through the miR-34a/Notch-1 and miR-186 axis [30,33]. Downregulation of SNHG7 and consequent upregulation of sponged miRNAs (miR-34a or miR-186) in BC cells lead to sensitivity to Adriamycin and Trastuzumab [34]. Also, SNHG7 directly sponges miRNA-381, miR-15a to promote BC cell proliferation and invasion [35,36]. Following SNHG7 knockdown, several proteins were found upregulated, such as Bax and p21, in contrast, β-catenin, and its target molecules c-Myc and cyclin D1, were downregulated [37]. There is evidence that Myc controls the SNHG7 loci that encode genes that are essential for regulating snoRNAs [38]. Also, c-Myc binds to the promoter region of SNHG7 and influences glycolysis through the SNHG7/miR34a-5p/LDHA (lactate dehydrogenase A) axis in BC cells [39].
SNHG12
Long non-coding RNA small nucleolar RNA host gene 12 (SNHG12) was found upregulated in BC tissues and cells. Activation of SNHG7 is associated with proliferation, tumor growth, migration, and invasion of BC cells by sponging miR-451a through inhibiting AKT/mTOR [40]. There is increasing evidence that altered SNHG7 expression may play a role in various malignant tumors [41]. SNHG7 upregulation in the cytoplasm also significantly increased Matrix Metallopeptidase 13 (MMP13) expression, whereas SNHG7 knockdown significantly inhibited MMP13 expression [42]. C-Myc interacts directly with the promoter region of SNHG7 thus transcriptionally activating it in TNBC cells, where their expression correlates with lymph node and tumor size [43].
BCYRN1
A long non-coding RNA called BC200 (brain cytoplasmic RNA 1, BCYRN1) is highly expressed in invasive carcinomas of BC cells [44]. BCYRN1 may be used for prevention, cell survival screening, cell migration, invasion diagnosis, and prognosis of BC. The overexpression of BCYRN1 causes luminal and TNBC proliferation, migration, and invasion [45]. Estrogen can regulate BC200 expression in BC cell lines [46]. The expression of estrogen in BC cells induces BCYRN1, which leads to sponge Bcl-x mRNA [47]. Furthermore, myc binding to the E-box of BCYRN promoter increases its expression, which is important for cell migration and invasion [48]. BCYRN1 expression was significantly reduced by Myc knockdown, indicating that BCYRN1 is also controlled by Myc [46].
5.2. LncRNAs affecting MYC Expression
MALAT1
Several types of tumors, including BC, have overexpression of Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1). It has been demonstrated that MALAT1 expression in BC is positively associated with tumor size, lymph metastasis, angiogenesis, stage tumor, progression and metastasis [15]. A number of transcription factors control MALAT1 transcription, either positively, like SP1, SP3, β-catenin, HIF1α and HIF2α, c-Myc, YAP1, NRF1, or negatively, like p53 and SOX17 [49]. By binding and inactivating TEAD, MALAT1 prevents TEAD from interacting with its co-activator Yes-associated protein 1 (YAP1) and target gene promoters such as ITGB4 and VEGFA [50]. In a hypoxic environment, HIF-1/2 (Hypoxia-Induced Factor 1/2) could transcriptionally activate MALAT1 to promote tumor growth and migration in BC cell [51]. Also, MALAT1 promotes the Wnt-β-catenin pathway, which leads to c-Myc expression [52].
LacRNA
LacRNA (LINC00478-associated cytoplasmic RNA), is a novel cytoplasmic lncRNA derived from LINC00478 located in the nucleus [53]. LncRNA LINC00478 was substantially downregulated in the metastatic tumor samples [54]. LacRNA acts as a regulatory molecule that inhibits Myc activity. Through repression of Myc targets, LacRNA disrupts downstream signaling pathways involved in tumor cell migration, invasion, and colonization. LacRNA stabilizes Prohibitin 2 (PHB2), leading to BC metastasis suppression [53]. PHB2 is a protein found in the inner mitochondrial membrane, where it plays a crucial role in regulating mitochondrial function and maintaining stability [55]. Furthermore, PHB2 is involved in various cellular processes, including cell cycle control, apoptosis, transcription, and signal transduction. LacRNA interacts specifically with PHB2, recruiting c-Myc and promoting c-Myc ubiquitination and degradation. This negative regulation of the Myc signaling pathway ultimately inhibits BC metastasis [56].
Linc00839
The expression of LINC00839 is increased in various cancer types, especially chemoresistant BCs associated with poor prognosis [57]. BC patients with elevated Linc00839 have higher pathological grade, tumor size, and clinical stage [58]. Activation of Linc00839 by Myc leads to proliferation and chemoresistance in BC through its association with Lin28B [59]. A Myc/Linc00839/Lin28B loop activatse the PI3K/AKT signaling pathway and promotes proliferation and chemoresistance, and inhibits cell apoptosis in BC [59]. Notably, LINC00839 is up-regulated in tamoxifen- and ADR-resistant BC cells and tissues, and predicts a poor prognosis of BC patients [59].
Lnc408
The expression of long non-coding RNA 408 (Lnc408) was increased in BC stem cells (BCSCs) [60]. BCSCs require LNC408 for self-renewal maintenance via modulation of the β-catenin protein, and suppresses CBY1 transcription. Lnc408/ β-catenin functions as part of the Wnt signaling pathway that regulates stemness in a variety of malignancies [61]. The unregulated levels of nuclear β-catenin/Wnt resulting from the suppression of CBY1 by Lnc408 preserve the stemness characteristics of BCSCs expressing high levels of CD44, SOX2, Nanog, Klf4, and c-Myc [62].
LINC01287
In BC, LINC01287, a newly discovered lncRNA, acts as an oncogene. There was a significant reduction in overall survival of patients with higher LINC01287 expressions as compared to patients with lower LINC01287 [63]. In response to LINC01287 downregulation, E-cadherin expression was increased, whereas N-cadherin and vimentin expression decreased [64]. As a result of upregulation of LINC01287, BC cells proliferated and metastasized, while apoptosis was inhibited. LINC01287 activates Wnt/β-catenin pathway and upregulated Myc expression, thereby enhancing the proliferation and metastatic capability of BC cells [63].
MCM3AP-AS1
Minichromosome maintenance complex component 3-associated protein antisense RNA 1 (MCM3AP-AS1) is an lncRNA molecule that is transcribed from the opposite strand of the MCM3AP gene. The up-regulation of MCM3AP-AS1 significantly promotes BC cell proliferation, apoptosis, migration, and invasion [65]. Also, LncRNA MCM3AP-AS1 promotes BC progression via modulating the miR-28-5p/Centromere protein F (CENPF) axis [66]. Chemotherapeutic agents, such as cisplatin and doxorubicin, are affected by MCM3AP-AS1 [67]. A significant correlation was observed between MCM3AP-AS1 expression and the ER or PR expression profiles in BC patients, whereas there was no significant correlation between MCM3AP-AS1 expression and the HER2 expression profiles [68]. Overexpression of MCM3AP-AS1 in BC led to increased tumor size and cell proliferation through MEG3 downregulation [69]. In BC, MCM3AP-AS1 binds with the RNA-binding protein ZFP36 ring finger protein (ZFP36) to regulate the key genes involved in cancer progression, including cyclin D1 (CCND1), c-Myc, and MMP1 [65].
LINC00511
Long intergenic non-protein coding RNA 511, also known as LINC00511, encodes a protein called LINC00511-133aa. The peptide encoded by this lncRNA plays a role in blocking apoptosis and promoting stemness and invasiveness in BC cells. There was increased expression of LINC00511 in BC samples and cell lines associated with poor prognosis [70]. LINC00511 expression has been closely associated with lymph node metastasis, larger tumor size, and molecular subtypes of BC, regulating MMP13 expression through miR-150 sponging [71]. It is possible that LINC00511 leads to an increased level of aggressiveness in cancerous cells by sponging miRNAs and modifying their targets' levels such as E2F1/Nanog [72]. Knockdown of LINC00511 suppressed Wnt/β-catenin signaling activity and its downstream target genes, including c-Myc and Cyclin D1 [73].
sONE
sONE is a lncRNA located on 1q21.3 with a size of 2.4 kb and two exons. sONE is a tumor suppressor lncRNA that represses the expression of endothelial nitric oxide synthase3 (NOS3) and influences nitric oxide production (NO) [74]. The expression of sONE is down-regulated in TNBC patients compared to normal tissues, and it is negative correlated with disease aggressiveness, tumor size and lymph node metastasis [75]. sONE represses c-Myc expression, whereas TP53 expression is promoted. Additionally, sONE induces tumor suppressor miRNAs downstream of c-Myc and Tp53, including miR-34a, miR-15a, miR-16, and let-7a [75].
PICART1
PICART1 is a novel 2.5 kb in length lncRNA located at 17q21.33 and contains three exons, and it has been identified as a tumor suppressor [76]. TP53 expression increases in response to DNA damage caused by doxorubicin, which leads to an increase in PICART1 expression through a p53 response element in its promoter [76]. PICART1 plays a regulatory role in cell proliferation in BC by influencing the AKT/GSK3β/β-catenin signaling pathway [77]. In BC cells, down-regulation of PICART1 led to an increase in pAKT, pGSK3B, β-catenin, cyclin D1 and c-Myc expression, while p21Waf/cip1 expression was decreased [76].
5.3. MYC-lncRNAs regulatory loops
CCAT2
A significant increase in CCAT2 mRNA expression was observed in BC metastasis. Up-regulation of CCAT2 promotes BC cell proliferation, invasion and migration. By downregulating CCAT2, the cell cycle of BC cells is conversely arrested in the G0/G1 phase and apoptosis is promoted. Additionally, down-regulation of CCAT2 significantly reduced the expression of TGF-β, Smad2 and α-SMA proteins in BC cells [78]. Knockdown of CCAT2 improves Tamoxifen sensitivity in BC [79]. The CCAT2 level in BC regulated proliferation, apoptosis, and chemosensitivity through the mTOR, Notch and TGF-β signaling pathway [80,81]. As a result of CCAT2 interaction with EZH2, it inhibits the expression of p15 via histone methylation, contributing to the proliferation of BC cells [82]. Vitamin D can inhibit BC development by targeting CCAT2 [83]. Significant suppression of proliferation and stemness of BC cells was observed with cytoplasmic CCAT2 in luminal subtypes by miR-221-p27 signaling. In contrast, nuclear overexpression of CCAT2 led to upregulation of OCT4-PG1 and the induction of stemness features [84]. The long non-coding RNA CCAT2 has been shown to exhibit high expression in BC tissues and cell lines. Silencing CCAT2 has shown to suppress tumor formation and inhibit cell growth via a negative regulation of Wnt signaling pathway, consequently leading to c-Myc down-regulation [85].
EPIC1
The epigenetically induced Myc interacting lncRNA 1 (EPIC1), a Myc-binding lncRNA, is an intergenic lncRNA of 2.4 kb in size, which is transcribed from the minus strand of DNA on chromosome 22q13.31. EPIC1 is epigenetically activated during BC and is associated with poor survival rates. According to Xu et al., EPIC1 is implicated in mechanisms involved in chemoresistance in BC, including the metabolism of daunorubicin, rapamycin and doxorubicin through the AKT-mTORC1 signaling pathway [86,87]. EPIC1 directly interacts with Myc in BC cells and promotes their cell cycle progression [88]. In response to EPIC1 activation, BC cells express lower levels of CD8+ T cell markers, including perforin 1 and granzyme A [89]. A high expression level of EPIC1 in luminal B BC patients has been associated with poor prognosis and increased tumor size. Also, EPIC1 interacts with Myc to promote cancer cell progression and colony formation, cell cycle, and tumor growth. [88,90]. In different types of cancer, EPIC1 gene expression is increased due to hypermethylation of its promoter [91]. EPIC1 knockdown reduces Myc recruitment to its target genes’ promoters such as CDKN1A, CCNA2, CDC20, and CDC45 [91].
HOTAIR
HOX Transcript Antisense Intergenic RNA (HOTAIR) is transcribed from the minus strand of the HOXC gene cluster. HOTAIR plays a role in promoting BC cell proliferation by regulating cell cycle and apoptosis. HOTAIR contributes to the recruitment of enhancer of zeste homolog 2 (EZH2) to the Myc promoter [92]. In BC, HOTAIR acts as a scaffold to attract transcription factors Lysine-specific demethylase 1 (LSD1) and Mammalian hepatitis BX-interacting protein (HBXIP) [93]. The HBXIP/Hotair/LSD1 complex interacts with c-Myc to activate the transcription of Myc target genes (Cyclin A, eIF4E and LDHA) [94]. HOTAIR is a direct target of c-Myc and the HOTAIR promoter exhibited significant enrichment of both NF-kB and c-Myc binding sites [95]. The increased HOTAIR expression in cancer has been also attributed to transcriptional activation by c-Myc through the E-box located upstream of the transcription start site [96].
RHPN1-AS1
The Rhophilin Rho GTPase Binding Protein 1 AntisenseRNA 1 (RHPN1-AS1) lncRNA is a direct transcriptional target of c-Myc, which regulates P53 expression in BC cells via the MDM2 gene. Also, RHPN1-AS1 acts as a molecular sponge for miR-4261 and prevents it from targeting c-Myc [97]. In BC cells, RHPN1-AS1 is highly expressed enhances the expression of epithelial-to-mesenchymal transition (EMT) markers [98], and RHPN1-AS1 knockdown inhibits MCF-7 cell proliferation [97].
5.4. LncRNAs affecting MYC stability/translation
AFAP1-AS1
There is an association between AFAP1-AS1 elevated expression and poor prognosis of BC patients [99]. AFAP1-AS1 knockdown downregulates septin-2 (SEPT2, a class of cytoskeletal proteins) via miR-497-5 sponging [100], repressing cell proliferation and migration and inducing apoptosis Further, AFAP1-AS1 plays a role in promoting EMT and tumorigenesis through the Wnt/β-catenin signaling pathway in TNBC [101]. AFAP1-AS1 can interact with Smad nuclear interacting protein 1 (SNIP1), a protein that suppresses the ubiquitination and destruction of c-Myc [102]. Due to this function, AFAP1-AS1 promotes the overexpression of c-Myc, as well as the increase in ZEB1, ZEB2, and SNAIL levels, thus triggering EMT [102].
LINC01638
There has been evidence of low levels of LINC01638 expression in several normal human tissues [103]. LINC01638 was found to be overexpressed and promote BC growth. [104]. In HER2-positive BC cells, inhibition of LINC01638 reduced the expression of DNMT1, DNMT3a, and DNMT3b while promoting the expression of BRCA1 and PTEN [105]. In HER2-positive BC cells, Linc01638 is upregulated, which acts inhibiting cell apoptosis while promoting proliferation and invasion. LINC01638 interacts with c-Myc to prevent its ubiquitination and degradation by SPOP (E3 ubiquitin ligase adapter speckle type POZ) via proteasomal degradation [106]; in turn, c-Myc transcriptionally enhances MTDH (metadherin) expression and subsequently activates Twist1 expression to induce EMT and cancer stem cell (CSC)-like features [103].
SNHG15
The small nucleolar RNA host gene 15 (SNHG15) is a long intergenic lncRNA with a size of 2.8 kb located on the 7q22.1 chromosome and transcribed from the plus strand of DNA. In BC, high SNHG15 expression positively correlated with TNM stage, lymph node metastasis, and survival. SNHG15 significantly increased proliferation and inhibited apoptosis in BC cells [15]. SNHG15 promotes human BC proliferation, migration and invasion by sponging miR-211-3p [107]. In BC cells, upregulation of SNHG15 promotes cell proliferation, migration, and invasion and inhibits apoptosis through regulation of miR-411-5p/VASP and miR-345-5p [108,109]. Additionally, SNHG15 enhanced cisplatin resistance of BC cells by sponging miR-381 [110]. SNHG15 expression is positively correlated with Myc expression and transcription of SNHG15 is directly regulated by Myc [111]. By increasing c-Myc expression in response to miR-451 sponging, SNHG15 improved BC progression [112]. SNHG15 is a direct c-Myc transcriptional target. In BC cells, SNHG15 acts as a molecular sponge also for miR-873-5p, a miRNA capable of destabilizing c-Myc mRNA [113], ultimately leading to an increase in c-Myc activity, driving growth and survival of of BC cells [113].
Linc-RoR
Linc-RoR is an oncogene that was originally identified as involved in induced pluripotent stem cells (iPSC) reprogramming [114]. The overexpression of linc-RoR appears to promote proliferation, metastasis, drug resistance, and invasion in BC cells [115]. Overexpression of Linc-RoR increased stemness features and induced the epithelial-to-mesenchymal transition (EMT) by activating the Wnt/β-catenin pathway [116]. Linc-RoR promotes the TGF-β signaling pathway to induce downstream signals such as Smad2 and α-SMA that contribute to BC progression [117]. Linc-RoR increases c-Myc mRNA stability in BC through interactions with heterogeneous nuclear ribonucleoprotein 1 (hnRNP1) and AUF1 (AU-rich element RNA-binding protein 1) [115]. Mechanistically, Linc-RoR acts facilitating hnRNP I interaction with c-Myc mRNA and inhibiting AUF1 binding to c-Myc mRNA for degradation [118]. Increased expression of LncRoR, enhancing Myc stability in BC cells, is usually associated with poor patient survival.
KB-1980E6.3
Hypoxia modulates a specific group of hypoxia-responsive lncRNAs (HRLs), like NORAD, LncHIFCAR, RAB11B-AS1, AC020978 and KB-1980E6.3, which may underlie cancer cells' survival and promote disease progression [119]. Breast tumors had significantly higher levels of KB-1980E6.3 than normal tissues, which was closely correlated with poor survival facilitate c-Myc mRNA stability [120]. BC cells are stimulated to proliferate, invade and migrate by KB-1980E6.3 through the activation of PI3K/AKT signaling [121]. KB-1980E6.3 expression positively correlates with migration and invasion markers such as MMP-2, MMP-9 and vimentin in BC [121]. By recruiting IGF2BP1 to regulate c-Myc mRNA stability under hypoxia conditions, KB-1980E6.3 promotes the self-renewal and stemness maintenance of BC stem cells [122]. During mild chronic hypoxia, HIF enhances the stability and activity of Myc, while under normoxia, IGF2BP1 protects Myc mRNA from degradation [122,123]. This mechanism promotes BC stem cell stemness, metabolic rewiring and tumorigenesis.
H19
The H19 transcript was one of the first lncRNAs to be discovered. It acts as oncogene, whose expression levels are elevated in breast tissues and associated with an increased BC risk [124]. At the H19/IGF2 (insulin-like growth factor 2) locus, genomic imprinting occurs, leading to differential expression of H19 from the maternal allele and IGF2 from the paternal allele [125]. A protein known as the coding region determinant-binding protein (CRD-BP) binds to c-Myc, insulin-like growth factor II, beta-actin, and H19 RNA [126]. Histone acetylation and transcriptional initiation of the H19 promoter are promoted by c-Myc protein binding to E-boxes, increasing H19 expression [127]. As a result of the binding to c-Myc and H19, CRD-BP affects the stability, localization and translation of RNA [128]. BC tumorigenesis is facilitated by the complex of CRD-BP, c-Myc, and H19. Furthermore, the reciprocally imprinted gene at H19 locus, IGF2, is also down-regulated by c-Myc [129].
DCST1-AS1
There has been evidence that the Domain Containing 1 Antisense RNA 1 (DCST1-AS1) lncRNA is dysregulated in numerous cancers, including BC, cervical cancer, glioblastoma, and hepatocellular carcinoma [130]. Overexpression of DCST1-AS1 in cancer is commonly observed, and it is associated with tumor progression, metastasis, poor prognosis, cell cycle, invasion, and inhibition of apoptosis [131]. By sponging miR-1254, lncRNA DCST1-AS1 significantly enhances proliferation of hepatocellular carcinoma (HCC) by upregulating Fas apoptosis inhibitor 2 (FAIM2) expression [132]. As a result of DCST1-AS1 overexpression in cervical cancer, miR-874-3p expression is downregulated, promoting proliferation, migration, and invasion [133]. Also, DCST1-AS1 directly binds to Annexin A1 (ANXA1) and enhances TGF-β/Smad and subsequently promotes EMT-related proteins such as E-cadherin, SNAI1, vimentin, MMP2, and MMP9 in BC cells [134]. DCST1-AS1 is a direct transcriptional target of c-Myc, and acts as a molecular sponge for miR-873-5p in triple-negative breast cancer (TNBC). MIR-873-5p binds directly to IGF2BP1 (stabilizes c-Myc mRNA) and regulates downstream proteins of IGF2BP1 such as Myc, LEF1 (activates c-Myc expression) and CD44, inhibiting Myc expression. [134]. in BC, a positive feedback regulation was found between Myc and DCST1-AS1, by which Myc promotes DCST1-AS1 transcription and DCST1-AS1 increases Myc translation [134].
PVT1
There is a significant increase in Plasmacytoma Variant Translocation 1 (PVT1) levels in BC patients. Cell proliferation and metastasis are promoted by PVT1 both in vitro and in vivo [15]. Inhibition of PVT1 expression inhibited cell proliferation and induced apoptosis in cell lines [135]. CRISPR interference of PVT1 enhances BC cell growth. PVT1 contains two MYC-binding sites that are bound and transactivated by c-Myc [136]. PVT1 and Myc promoters compete for intragenic enhancers in the PVT1 locus (8q24 region contains PVT1 and Myc), thus PVT1 promoter can regulate release of Myc transcription [137]. Also, it has been suggested that PVT1 can enhance Myc protein stability by decreasing its degradation. The stable Myc protein (Myc STAB) binds to the canonical E-boxes at the PVT1 promoter region, and upregulates PVT1 RNA [138]. High levels of Myc and PVT1 can transcriptionally activate RSPO1, a β-catenin signaling regulator which is important for female development and is associated with tumor formation [139].
FGF13-AS1
Fibroblast growth factor 13 Antisense RNA 1(FGF13-AS1) is transcribed from the opposite strand of the FGF13 gene. FGF13-AS1 expression is decreased in BC tissues compared with corresponding normal tissues, and reduced FGF13-AS1 is associated with poor prognosis [140]. FGF13-AS1 is downregulated in BC and acts as a tumor suppressor by inhibiting glycolysis and stemness properties [140]. FGF13-AS1 reduces stability of Myc mRNA by disrupting the interaction between IGF2BPs and Myc mRNA. It also forms a feedback loop with Myc, since Myc transcriptionally inhibits FGF13-AS1 expression [140].
6. Conclusion
Overall, this comprehensive review provides a detailed analysis of the intricate cross-talk between long noncoding RNAs (lncRNAs) and the Myc gene, highlighting their pivotal roles in various aspects of BC biology. The Myc gene, a highly studied oncogene, serves as a central regulator of critical cellular processes such as cell growth, proliferation, and metabolism. The aberrant expression and dysregulation of Myc have been extensively implicated in tumorigenesis, metastasis, invasion, and therapy resistance across different cancer types. LncRNAs are a diverse group of transcripts longer than 200 nucleotides that do not encode proteins, but play significant regulatory role in gene expression. The interactions between Myc and lncRNAs occur through a multitude of mechanisms, including direct and indirect modulation of expression at transcriptional, post-transcriptional, and post-translational levels. LncRNAs can impact Myc activity by influencing its stability, translation, or interactions with other proteins. Conversely, Myc can regulate the expression of numerous lncRNA genes, thereby influencing down-stream genes involved in tumorigenesis.
Understanding the functional implications of the Myc-lncRNA cross-talk is of great significance for cancer research and clinical applications. Dysregulated Myc-lncRNA interactions can disrupt normal cellular processes, leading to uncontrolled proliferation, genomic instability, and evasion of cell death, all hallmarks of cancer. Conversely, targeted modulation of Myc-lncRNA interactions holds therapeutic potential. Strategies aimed at restoring balance in Myc regulation through the manipulation of oncogenic or tumor-suppressive lncRNAs may provide novel avenues for cancer treatment, blunting Myc signaling which still remains undruggable
However, despite the substantial progress made in unraveling the Myc-lncRNA network, several challenges and gaps in knowledge still remain. Further research is needed to uncover the precise mechanisms underlying Myc-lncRNA interactions, including the identification of specific binding sites and associated co-factors. Moreover, comprehensive functional studies are necessary to determine the specific roles of individual lncRNAs in Myc-mediated tumorigenesis and their interactions with other molecular players.
In conclusion the Myc-lncRNA network represents a complex regulatory system that plays a significant role in BC initiation, progression, and therapeutic responses. Expanding our knowledge on the dysregulated lncRNAs, their mechanisms of action, and their clinical implications will pave the way for the development of targeted therapies aimed at antagonizing Myc oncogenic functions, improving patient outcomes in BC and possibly other Myc-addicted cancer types. Further investigation is required to decipher the specific roles of individual lncRNAs in modulating Myc and viceversa. Additionally, it is necessary to investigate the downstream effects of these interactions on cellular processes relevant to cancer biology.
Author Contributions
R.H. and A.M.B. conceived and designed the study; S.P.M. , M.J.A , A.H.M. and R.A.B. performed the search; S.Z.S., and M.C. obtained the funding to perform this project; I.M.E. and A.J. wrote the first draft of the manuscript and R.H, S.Z.S, M.C. N.A. and R.H. performed the review and supervision of the final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Madden, S.K.; de Araujo, A.D.; Gerhardt, M.; Fairlie, D.P.; Mason, J.M. Taking the Myc out of cancer: toward therapeutic strategies to directly inhibit c-Myc. Molecular Cancer 2021, 20, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gao, X.; Yuan, M.; Yang, B.; He, Q.; Cao, J. Targeting Myc Interacting Proteins as a Winding Path in Cancer Therapy. Frontiers in Pharmacology 2021, 12. [Google Scholar] [CrossRef]
- Dhanasekaran, R.; Deutzmann, A.; Mahauad-Fernandez, W.D.; Hansen, A.S.; Gouw, A.M.; Felsher, D.W. The MYC oncogene — the grand orchestrator of cancer growth and immune evasion. Nature Reviews Clinical Oncology 2022, 19, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.J.; O'Grady, S.; Tang, M.; Crown, J. MYC as a target for cancer treatment. Cancer Treatment Reviews 2021, 94, 102154. [Google Scholar] [CrossRef]
- Llombart, V.; Mansour, M.R. Therapeutic targeting of "undruggable" MYC. eBioMedicine 2022, 75. [Google Scholar] [CrossRef]
- Schaub, F.X.; Dhankani, V.; Berger, A.C.; Trivedi, M.; Richardson, A.B.; Shaw, R.; Zhao, W.; Zhang, X.; Ventura, A.; Liu, Y.; et al. Pan-cancer Alterations of the MYC Oncogene and Its Proximal Network across the Cancer Genome Atlas. Cell Systems 2018, 6, 282–300e282. [Google Scholar] [CrossRef]
- Zimmerli, D.; Brambillasca, C.S.; Talens, F.; Bhin, J.; Linstra, R.; Romanens, L.; Bhattacharya, A.; Joosten, S.E.P.; Da Silva, A.M.; Padrao, N.; et al. MYC promotes immune-suppression in triple-negative BC via inhibition of interferon signaling. Nature Communications 2022, 13, 6579. [Google Scholar] [CrossRef]
- Dang, C.V. c-Myc target genes involved in cell growth, apoptosis, and metabolism. Mol Cell Biol 1999, 19, 1–11. [Google Scholar] [CrossRef]
- Sodir, N.M.; Swigart, L.B.; Karnezis, A.N.; Hanahan, D.; Evan, G.I.; Soucek, L. Endogenous Myc maintains the tumor microenvironment. Genes Dev 2011, 25, 907–916. [Google Scholar] [CrossRef]
- Massó-Vallés, D.; Beaulieu, M.-E.; Jauset, T.; Giuntini, F.; Zacarías-Fluck, M.F.; Foradada, L.; Martínez-Martín, S.; Serrano, E.; Martín-Fernández, G.; Casacuberta-Serra, S.; et al. MYC Inhibition Halts Metastatic BC Progression by Blocking Growth, Invasion, and Seeding. Cancer Research Communications 2022, 2, 110–130. [Google Scholar] [CrossRef]
- Gao, Y.; Jiang, M.; Guo, F.; Liu, X.; Zhang, Q.; Yang, S.; Yeung, Y.T.; Yang, R.; Wang, K.; Wu, Q.; et al. A novel lncRNA MTAR1 promotes cancer development through IGF2BPs mediated post-transcriptional regulation of c-MYC. Oncogene 2022, 41, 4736–4753. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, C.; Resetca, D.; Redel, C.; Lin, P.; MacDonald, A.S.; Ciaccio, R.; Kenney, T.M.G.; Wei, Y.; Andrews, D.W.; Sunnerhagen, M.; et al. MYC protein interactors in gene transcription and cancer. Nature Reviews Cancer 2021, 21, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Karami, M.; Mousavi, S.H.; Rafiee, M.; Heidari, R.; Shahrokhi, S.Z. Biochemical and molecular biomarkers: unraveling their role in gestational diabetes mellitus. Diabetol Metab Syndr 2023, 15, 5. [Google Scholar] [CrossRef] [PubMed]
- Jacovetti, C.; Bayazit, M.B.; Regazzi, R. Emerging Classes of Small Non-Coding RNAs With Potential Implications in Diabetes and Associated Metabolic Disorders. Frontiers in Endocrinology 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Heidari, R.; Akbariqomi, M.; Asgari, Y.; Ebrahimi, D.; Alinejad-Rokny, H. A systematic review of long non-coding RNAs with a potential role in BC. Mutation Research/Reviews in Mutation Research 2021, 787, 108375. [Google Scholar] [CrossRef]
- Heidari, R.; Akbariqomi, M.; Asgari, Y.; Ebrahimi, D.; Alinejad-Rokny, H. A systematic review of long non-coding RNAs with a potential role in BC. Mutat Res Rev Mutat Res 2021, 787, 108375. [Google Scholar] [CrossRef]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol Immunol 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Wang, X.; Song, X.; Glass, C.K.; Rosenfeld, M.G. The long arm of long noncoding RNAs: roles as sensors regulating gene transcriptional programs. Cold Spring Harbor perspectives in biology 2011, 3, a003756. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nature Reviews Genetics 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Li, G.; Deng, L.; Huang, N.; Sun, F. The Biological Roles of lncRNAs and Future Prospects in Clinical Application. Diseases 2021, 9. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nature Reviews Molecular Cell Biology 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed]
- Kortlever, R.M.; Sodir, N.M.; Wilson, C.H.; Burkhart, D.L.; Pellegrinet, L.; Brown Swigart, L.; Littlewood, T.D.; Evan, G.I. Myc Cooperates with Ras by Programming Inflammation and Immune Suppression. Cell 2017, 171, 1301–1315e1314. [Google Scholar] [CrossRef] [PubMed]
- Fallah, Y.; Brundage, J.; Allegakoen, P.; Shajahan-Haq, A.N. MYC-Driven Pathways in BC Subtypes. Biomolecules 2017, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Risom, T.; Wang, X.; Liang, J.; Zhang, X.; Pelz, C.; Campbell, L.G.; Eng, J.; Chin, K.; Farrington, C.; Narla, G.; et al. Deregulating MYC in a model of HER2+ BC mimics human intertumoral heterogeneity. J Clin Invest 2020, 130, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Giltnane, J.M.; Balko, J.M.; Schwarz, L.J.; Guerrero-Zotano, A.L.; Hutchinson, K.E.; Nixon, M.J.; Estrada, M.V.; Sánchez, V.; Sanders, M.E.; et al. MYC and MCL1 Cooperatively Promote Chemotherapy-Resistant BC Stem Cells via Regulation of Mitochondrial Oxidative Phosphorylation. Cell Metab 2017, 26, 633–647e637. [Google Scholar] [CrossRef]
- Johnston, L.A. Socializing with MYC: cell competition in development and as a model for premalignant cancer. Cold Spring Harb Perspect Med 2014, 4, a014274. [Google Scholar] [CrossRef]
- Kreuzaler, P.; Clarke, M.A.; Brown, E.J.; Wilson, C.H.; Kortlever, R.M.; Piterman, N.; Littlewood, T.; Evan, G.I.; Fisher, J. Heterogeneity of Myc expression in BC exposes pharmacological vulnerabilities revealed through executable mechanistic modeling. Proceedings of the National Academy of Sciences 2019, 116, 22399–22408. [Google Scholar] [CrossRef]
- Annunziato, S.; de Ruiter, J.R.; Henneman, L.; Brambillasca, C.S.; Lutz, C.; Vaillant, F.; Ferrante, F.; Drenth, A.P.; van der Burg, E.; Siteur, B.; et al. Comparative oncogenomics identifies combinations of driver genes and drug targets in BRCA1-mutated BC. Nat Commun 2019, 10, 397. [Google Scholar] [CrossRef]
- Debnath, S.; Mukherjee, A.; Saha, D.; Dash, J.; Chatterjee, T.K. Poly-l-Lysine inhibits VEGF and c-Myc mediated tumor-angiogenesis and induces apoptosis in 2D and 3D tumor microenvironment of both MDA-MB-231 and B16F10 induced mice model. International Journal of Biological Macromolecules 2021, 183, 528–548. [Google Scholar] [CrossRef]
- Luo, X.; Song, Y.; Tang, L.; Sun, D.H.; Ji, D.G. LncRNA SNHG7 promotes development of BC by regulating microRNA-186. Eur Rev Med Pharmacol Sci 2018, 22, 7788–7797. [Google Scholar] [CrossRef]
- Bian, Z.; Ji, W.; Xu, B.; Huang, W.; Jiao, J.; Shao, J.; Zhang, X. The role of long noncoding RNA SNHG7 in human cancers (Review). Mol Clin Oncol 2020, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Yu, N.S.; Deng, Q.; Zhang, Y.; Hu, Y.Y.; Liu, G.; Huang, K. LncRNA SNHG7 Mediates the Chemoresistance and Stemness of BC by Sponging miR-34a. Front Oncol 2020, 10, 592757. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Long, Z.; Wu, S.; Xiao, M.; Hu, W. LncRNA-SNHG7 regulates proliferation, apoptosis and invasion of bladder cancer cells assurance guidelines. J buon 2018, 23, 776–781. [Google Scholar] [PubMed]
- Najafi, S.; Ghafouri-Fard, S.; Hussen, B.M.; Jamal, H.H.; Taheri, M.; Hallajnejad, M. Oncogenic Roles of Small Nucleolar RNA Host Gene 7 (SNHG7) Long Noncoding RNA in Human Cancers and Potentials. Frontiers in Cell and Developmental Biology 2022, 9. [Google Scholar] [CrossRef]
- Gao, Y.T.; Zhou, Y.C. Long non-coding RNA (lncRNA) small nucleolar RNA host gene 7 (SNHG7) promotes BC progression by sponging miRNA-381. Eur Rev Med Pharmacol Sci 2019, 23, 6588–6595. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Guo, X.; Wei, Y. LncRNA SNHG7 inhibits proliferation and invasion of BC cells by regulating miR-15a expression. J BUON 2020, 25, 1792–1798. [Google Scholar]
- Zhou, Y.; Tian, B.; Tang, J.; Wu, J.; Wang, H.; Wu, Z.; Li, X.; Yang, D.; Zhang, B.; Xiao, Y.; et al. SNHG7: A novel vital oncogenic lncRNA in human cancers. Biomedicine & Pharmacotherapy 2020, 124, 109921. [Google Scholar] [CrossRef]
- Destefanis, F.; Manara, V.; Bellosta, P. Myc as a Regulator of Ribosome Biogenesis and Cell Competition: A Link to Cancer. International Journal of Molecular Sciences 2020, 21, 4037. [Google Scholar] [CrossRef]
- Zhang, L.; Fu, Y.; Guo, H. c-Myc-Induced Long Non-Coding RNA Small Nucleolar RNA Host Gene 7 Regulates Glycolysis in BC. J BC 2019, 22, 533–547. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, G. Knockdown of lncRNA SNHG12 suppresses cell proliferation, migration and invasion in BC by sponging miR-451a. Int J Clin Exp Pathol 2020, 13, 393–402. [Google Scholar]
- Wang, X.; Jiang, Q.; Zhang, C.; Yang, Q.; Wang, L.; Zhang, J.; Wang, L.; Chen, X.; Hou, X.; Han, D.; et al. Long noncoding RNA SNHG12 is a potential diagnostic and prognostic biomarker in various tumors. Chinese Neurosurgical Journal 2021, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Tamang, S.; Acharya, V.; Roy, D.; Sharma, R.; Aryaa, A.; Sharma, U.; Khandelwal, A.; Prakash, H.; Vasquez, K.M.; Jain, A. SNHG12: An LncRNA as a Potential Therapeutic Target and Biomarker for Human Cancer. Frontiers in Oncology 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, O.; Yang, F.; Liu, Y.; Lv, L.; Ma, R.; Chen, C.; Wang, J.; Tan, Q.; Cheng, Y.; Xia, E.; et al. C-MYC-induced upregulation of lncRNA SNHG12 regulates cell proliferation, apoptosis and migration in triple-negative BC. Am J Transl Res 2017, 9, 533–545. [Google Scholar] [PubMed]
- Iacoangeli, A.; Lin, Y.; Morley, E.J.; Muslimov, I.A.; Bianchi, R.; Reilly, J.; Weedon, J.; Diallo, R.; Böcker, W.; Tiedge, H. BC200 RNA in invasive and preinvasive BC. Carcinogenesis 2004, 25, 2125–2133. [Google Scholar] [CrossRef] [PubMed]
- Barton, M.; Santucci-Pereira, J.; Vaccaro, O.G.; Nguyen, T.; Su, Y.; Russo, J. BC200 overexpression contributes to luminal and triple negative BC pathogenesis. BMC Cancer 2019, 19, 994. [Google Scholar] [CrossRef]
- Booy, E.P.; McRae, E.K.S.; Koul, A.; Lin, F.; McKenna, S.A. The long non-coding RNA BC200 (BCYRN1) is critical for cancer cell survival and proliferation. Molecular Cancer 2017, 16, 109. [Google Scholar] [CrossRef]
- Singh, R.; Gupta, S.C.; Peng, W.X.; Zhou, N.; Pochampally, R.; Atfi, A.; Watabe, K.; Lu, Z.; Mo, Y.Y. Regulation of alternative splicing of Bcl-x by BC200 contributes to BC pathogenesis. Cell Death & Disease 2016, 7, e2262–e2262. [Google Scholar] [CrossRef]
- Hu, T.; Lu, Y.R. BCYRN1, a c-MYC-activated long non-coding RNA, regulates cell metastasis of non-small-cell lung cancer. Cancer Cell Int 2015, 15, 36. [Google Scholar] [CrossRef]
- Amodio, N.; Raimondi, L.; Juli, G.; Stamato, M.A.; Caracciolo, D.; Tagliaferri, P.; Tassone, P. MALAT1: a druggable long non-coding RNA for targeted anti-cancer approaches. Journal of Hematology & Oncology 2018, 11, 63. [Google Scholar] [CrossRef]
- Kim, J.; Piao, H.L.; Kim, B.J.; Yao, F.; Han, Z.; Wang, Y.; Xiao, Z.; Siverly, A.N.; Lawhon, S.E.; Ton, B.N.; et al. Long noncoding RNA MALAT1 suppresses BC metastasis. Nat Genet 2018, 50, 1705–1715. [Google Scholar] [CrossRef]
- Shih, C.-H.; Chuang, L.-L.; Tsai, M.-H.; Chen, L.-H.; Chuang, E.Y.; Lu, T.-P.; Lai, L.-C. Hypoxia-Induced MALAT1 Promotes the Proliferation and Migration of BC Cells by Sponging MiR-3064-5p. Frontiers in Oncology 2021, 11. [Google Scholar] [CrossRef]
- Wang, Z.; Katsaros, D.; Biglia, N.; Shen, Y.; Fu, Y.; Loo, L.W.M.; Jia, W.; Obata, Y.; Yu, H. High expression of long non-coding RNA MALAT1 in BC is associated with poor relapse-free survival. BC Res Treat 2018, 171, 261–271. [Google Scholar] [CrossRef]
- Guo, R.; Su, Y.; Zhang, Q.; Xiu, B.; Huang, S.; Chi, W.; Zhang, L.; Li, L.; Hou, J.; Wang, J.; et al. LINC00478-derived novel cytoplasmic lncRNA LacRNA stabilizes PHB2 and suppresses BC metastasis via repressing MYC targets. Journal of Translational Medicine 2023, 21, 120. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Su, Y.-H.; Xue, J.-y.; Si, J.; Chi, Y.-y.; Wu, J. Abstract P6-05-01: A novel cleaved cytoplasmic lncRNA LacRNA interacts with PHB2 and suppresses BC metastasis via repressing MYC targets. Cancer Research 2019, 79, P6–05. [Google Scholar] [CrossRef]
- Hu, L.L.; Zou, K.; Chen, Y.; Wu, L.J.; Cao, J.; Xiong, X.Y.; Wang, L.; Cheng, X.S.; Xiao, Q.Z.; Yang, R.Q. Functional role and molecular mechanisms underlying prohibitin 2 in platelet mitophagy and activation. Mol Med Rep 2021, 23. [Google Scholar] [CrossRef]
- Guo, R.; Su, Y.; Zhang, Q.; Xiu, B.; Huang, S.; Chi, W.; Zhang, L.; Li, L.; Hou, J.; Wang, J.; et al. LINC00478-derived novel cytoplasmic lncRNA LacRNA stabilizes PHB2 and suppresses BC metastasis via repressing MYC targets. J Transl Med 2023, 21, 120. [Google Scholar] [CrossRef]
- Liu, X.; Chen, J.; Zhang, S.; Liu, X.; Long, X.; Lan, J.; Zhou, M.; Zheng, L.; Zhou, J. LINC00839 promotes colorectal cancer progression by recruiting RUVBL1/Tip60 complexes to activate NRF1. EMBO Rep 2022, 23, e54128. [Google Scholar] [CrossRef]
- Su, J.; Deng, L.; Wang, Y.-D. Roles and Mechanisms of Long Non-Coding RNAs in BC. International Journal of Molecular Sciences 2023, 24, 89. [Google Scholar] [CrossRef]
- Chen, Q.; Shen, H.; Zhu, X.; Liu, Y.; Yang, H.; Chen, H.; Xiong, S.; Chi, H.; Xu, W. A nuclear lncRNA Linc00839 as a Myc target to promote BC chemoresistance via PI3K/AKT signaling pathway. Cancer Sci 2020, 111, 3279–3291. [Google Scholar] [CrossRef]
- Wen, S.; Qin, Y.; Wang, R.; Yang, L.; Zeng, H.; Zhu, P.; Li, Q.; Qiu, Y.; Chen, S.; Liu, Y.; et al. A novel Lnc408 maintains BC stem cell stemness by recruiting SP3 to suppress CBY1 transcription and increasing nuclear β-catenin levels. Cell Death Dis 2021, 12, 437. [Google Scholar] [CrossRef]
- Teng, Y.; Wang, X.; Wang, Y.; Ma, D. Wnt/beta-catenin signaling regulates cancer stem cells in lung cancer A549 cells. Biochem Biophys Res Commun 2010, 392, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Qin, Y.; Wang, R.; Yang, L.; Zeng, H.; Zhu, P.; Li, Q.; Qiu, Y.; Chen, S.; Liu, Y.; et al. A novel Lnc408 maintains BC stem cell stemness by recruiting SP3 to suppress CBY1 transcription and increasing nuclear β-catenin levels. Cell Death & Disease 2021, 12, 437. [Google Scholar] [CrossRef]
- Song, C.; Sun, P.; He, Q.; Liu, L.L.; Cui, J.; Sun, L.M. Long non-coding RNA LINC01287 promotes BC cells proliferation and metastasis by activating Wnt/ß-catenin signaling. Eur Rev Med Pharmacol Sci 2019, 23, 4234–4242. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; He, L.; Lai, Z.; Wan, Z.; Chen, Q.; Pan, S.; Li, L.; Li, D.; Huang, J.; Xue, F.; et al. LINC01287 regulates tumorigenesis and invasion via miR-298/MYB in hepatocellular carcinoma. J Cell Mol Med 2018, 22, 5477–5485. [Google Scholar] [CrossRef]
- Tang, T.P.; Qin, C.X.; Yu, H. MCM3AP-AS1 regulates proliferation, apoptosis, migration, and invasion of BC cells via binding with ZFP36. Transl Cancer Res 2021, 10, 4478–4488. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, H.; Zhu, J.; Feng, K.; Hu, C. LncRNA MCM3AP-AS1 promotes BC progression via modulating miR-28-5p/CENPF axis. Biomed Pharmacother 2020, 128, 110289. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Khoshbakht, T.; Hussen, B.M.; Taheri, M.; Samadian, M. A review on the role of MCM3AP-AS1 in the carcinogenesis and tumor progression. Cancer Cell International 2022, 22, 225. [Google Scholar] [CrossRef]
- Yu, X.; Zheng, Q.; Zhang, Q.; Zhang, S.; He, Y.; Guo, W. MCM3AP-AS1: An Indispensable Cancer-Related LncRNA. Frontiers in Cell and Developmental Biology 2021, 9. [Google Scholar] [CrossRef]
- Ren, G.; Han, G.; Song, Z.; Zang, A.; Liu, B.; Hu, L.; Jia, L.; Hong, D.; Yang, L.; Qie, S. LncRNA MCM3AP-AS1 Downregulates LncRNA MEG3 in Triple Negative BC to Inhibit the Proliferation of Cancer Cells. Crit Rev Eukaryot Gene Expr 2021, 31, 81–87. [Google Scholar] [CrossRef]
- Lu, G.; Li, Y.; Ma, Y.; Lu, J.; Chen, Y.; Jiang, Q.; Qin, Q.; Zhao, L.; Huang, Q.; Luo, Z.; et al. Long noncoding RNA LINC00511 contributes to BC tumourigenesis and stemness by inducing the miR-185-3p/E2F1/Nanog axis. J Exp Clin Cancer Res 2018, 37, 289. [Google Scholar] [CrossRef]
- Shi, G.; Cheng, Y.; Zhang, Y.; Guo, R.; Li, S.; Hong, X. Long non-coding RNA LINC00511/miR-150/MMP13 axis promotes BC proliferation, migration and invasion. Biochim Biophys Acta Mol Basis Dis 2021, 1867, 165957. [Google Scholar] [CrossRef]
- Lu, G.; Li, Y.; Ma, Y.; Lu, J.; Chen, Y.; Jiang, Q.; Qin, Q.; Zhao, L.; Huang, Q.; Luo, Z.; et al. Long noncoding RNA LINC00511 contributes to BC tumourigenesis and stemness by inducing the miR-185-3p/E2F1/Nanog axis. Journal of Experimental & Clinical Cancer Research 2018, 37, 289. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Meng, F.; Fu, L.; Kong, C. Knockdown of long non-coding RNA linc00511 suppresses proliferation and promotes apoptosis of bladder cancer cells via suppressing Wnt/β-catenin signaling pathway. Biosci Rep 2018, 38. [Google Scholar] [CrossRef]
- Youness, R.A.; Assal, R.A.; Abdel Motaal, A.; Gad, M.Z. A novel role of sONE/NOS3/NO signaling cascade in mediating hydrogen sulphide bilateral effects on triple negative BC progression. Nitric Oxide 2018, 80, 12–23. [Google Scholar] [CrossRef]
- Youness, R.A.; Hafez, H.M.; Khallaf, E.; Assal, R.A.; Abdel Motaal, A.; Gad, M.Z. The long noncoding RNA sONE represses triple-negative BC aggressiveness through inducing the expression of miR-34a, miR-15a, miR-16, and let-7a. J Cell Physiol 2019, 234, 20286–20297. [Google Scholar] [CrossRef]
- Cao, Y.; Lin, M.; Bu, Y.; Ling, H.; He, Y.; Huang, C.; Shen, Y.; Song, B.; Cao, D. p53-inducible long non-coding RNA PICART1 mediates cancer cell proliferation and migration. Int J Oncol 2017, 50, 1671–1682. [Google Scholar] [CrossRef]
- Tetsu, O.; McCormick, F. Beta-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999, 398, 422–426. [Google Scholar] [CrossRef]
- Wu, Z.J.; Li, Y.; Wu, Y.Z.; Wang, Y.; Nian, W.Q.; Wang, L.L.; Li, L.C.; Luo, H.L.; Wang, D.L. Long non-coding RNA CCAT2 promotes the BC growth and metastasis by regulating TGF-β signaling pathway. Eur Rev Med Pharmacol Sci 2017, 21, 706–714. [Google Scholar] [PubMed]
- Caia, Y.; He, J.; Zhang, D. Suppression of long non-coding RNA CCAT2 improves tamoxifen-resistant BC cells’ response to tamoxifen. Molecular Biology 2016, 50, 725–730. [Google Scholar] [CrossRef]
- Zhou, D.; Gu, J.; Wang, Y.; Luo, B.; Feng, M.; Wang, X. Long noncoding RNA CCAT2 reduces chemosensitivity to 5-fluorouracil in BC cells by activating the mTOR axis. Journal of Cellular and Molecular Medicine 2022, 26, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Liu, C.; Zhao, Q.; Lü, J.; Ding, X.; Luo, A.; He, J.; Wang, G.; Li, Y.; Cai, Z.; et al. Long non-coding RNA CCAT2 promotes oncogenesis in triple-negative BC by regulating stemness of cancer cells. Pharmacol Res 2020, 152, 104628. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhao, Y.; Wu, X.; Song, G. Upregulation of CCAT2 promotes cell proliferation by repressing the P15 in BC. Biomed Pharmacother 2017, 91, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, S.; Guo, B. Vitamin D suppresses ovarian cancer growth and invasion by targeting long non-coding RNA CCAT2. International Journal of Molecular Sciences 2020, 21, 2334. [Google Scholar] [CrossRef]
- Xie, H.; Guo, Y.; Xu, Z.; Wang, Q.; Wang, T.; Gu, Y.; Li, D.; Liu, Y.; Ma, W.; Liu, P.; et al. Dual Function of CCAT2 in Regulating Luminal Subtype of BC Depending on the Subcellular Distribution. Cancers 2023, 15, 538. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; He, J.; Zhang, D. Long noncoding RNA CCAT2 promotes breast tumor growth by regulating the Wnt signaling pathway. OncoTargets and therapy 2015, 2657–2664. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Yuan, C.; Sheng, X.; Sha, R.; Dai, H.; Zhang, S.; Wang, Y.; Lin, Y.; Zhou, L.; et al. Predictive and prognostic value of EPIC1 in patients with BC receiving neoadjuvant chemotherapy. Ther Adv Med Oncol 2020, 12, 1758835920940886. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, M.; Wang, Z.; Guo, W.; Yang, D. MYC-binding lncRNA EPIC1 promotes AKT-mTORC1 signaling and rapamycin resistance in breast and ovarian cancer. Mol Carcinog 2020, 59, 1188–1198. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, B.; Zhang, M.; Guo, W.; Wu, Z.; Wang, Y.; Jia, L.; Li, S.; Xie, W.; Yang, D. lncRNA Epigenetic Landscape Analysis Identifies EPIC1 as an Oncogenic lncRNA that Interacts with MYC and Promotes Cell-Cycle Progression in Cancer. Cancer Cell 2018, 33, 706–720e709. [Google Scholar] [CrossRef]
- Guo, W.; Wang, Y.; Yang, M.; Wang, Z.; Wang, Y.; Chaurasia, S.; Wu, Z.; Zhang, M.; Yadav, G.S.; Rathod, S.; et al. LincRNA-immunity landscape analysis identifies EPIC1 as a regulator of tumor immune evasion and immunotherapy resistance. Sci Adv 2021, 7. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, B.; Zhang, M.; Guo, W.; Wu, Z.; Wang, Y.; Jia, L.; Li, S.; Cancer Genome Atlas Research, N.; Xie, W.; et al. lncRNA Epigenetic Landscape Analysis Identifies EPIC1 as an Oncogenic lncRNA that Interacts with MYC and Promotes Cell-Cycle Progression in Cancer. Cancer cell 2018, 33, 706–720e709. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, B.; Zhang, M.; Guo, W.; Wu, Z.; Wang, Y.; Jia, L.; Li, S.; Caesar-Johnson, S.J.; Demchok, J.A.; et al. lncRNA Epigenetic Landscape Analysis Identifies EPIC1 as an Oncogenic lncRNA that Interacts with MYC and Promotes Cell-Cycle Progression in Cancer. Cancer Cell 2018, 33, 706–720e709. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Fei, Q.; Zhang, H.; Qiu, M.; Zhang, B.; Wang, Q.; Yu, Y.; Guo, C.; Ren, Y.; Mei, M.; et al. lncRNA HOTAIR Promotes DNA Repair and Radioresistance of BC via EZH2. DNA Cell Biol 2020. [CrossRef]
- Li, Y.; Wang, Z.; Shi, H.; Li, H.; Li, L.; Fang, R.; Cai, X.; Liu, B.; Zhang, X.; Ye, L. HBXIP and LSD1 Scaffolded by lncRNA Hotair Mediate Transcriptional Activation by c-Myc. Cancer Res 2016, 76, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Pawłowska, E.; Szczepanska, J.; Blasiak, J. The Long Noncoding RNA HOTAIR in BC: Does Autophagy Play a Role? International Journal of Molecular Sciences 2017, 18, 2317. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, J.; Tang, W. The molecular mechanism of HOTAIR in tumorigenesis, metastasis, and drug resistance. Acta Biochim Biophys Sin (Shanghai) 2014, 46, 1011–1015. [Google Scholar] [CrossRef]
- Loewen, G.; Jayawickramarajah, J.; Zhuo, Y.; Shan, B. Functions of lncRNA HOTAIR in lung cancer. J Hematol Oncol 2014, 7, 90. [Google Scholar] [CrossRef]
- Zhu, P.; Li, Y.; Li, P.; Zhang, Y.; Wang, X. c-Myc induced the regulation of long non-coding RNA RHPN1-AS1 on BC cell proliferation via inhibiting P53. Mol Genet Genomics 2019, 294, 1219–1229. [Google Scholar] [CrossRef]
- Zheng, S.; Lv, P.; Su, J.; Miao, K.; Xu, H.; Li, M. Silencing of the long non-coding RNA RHPN1-AS1 suppresses the epithelial-to-mesenchymal transition and inhibits BC progression. Am J Transl Res 2019, 11, 3505–3517. [Google Scholar]
- Ma, D.; Chen, C.; Wu, J.; Wang, H.; Wu, D. Up-regulated lncRNA AFAP1-AS1 indicates a poor prognosis and promotes carcinogenesis of BC. BC 2019, 26, 74–83. [Google Scholar] [CrossRef]
- LncRNA AFAP1-AS1 Knockdown Represses Cell Proliferation, Migration, and Induced Apoptosis in BC by Downregulating SEPT2 Via Sponging miR-497-5p. Cancer Biotherapy and Radiopharmaceuticals 2022, 37, 662–672. [CrossRef]
- Zhang, X.; Li, F.; Zhou, Y.; Mao, F.; Lin, Y.; Shen, S.; Li, Y.; Zhang, S.; Sun, Q. Long noncoding RNA AFAP1-AS1 promotes tumor progression and invasion by regulating the miR-2110/Sp1 axis in triple-negative BC. Cell Death & Disease 2021, 12, 627. [Google Scholar] [CrossRef]
- Zhong, Y.; Yang, L.; Xiong, F.; He, Y.; Tang, Y.; Shi, L.; Fan, S.; Li, Z.; Zhang, S.; Gong, Z.; et al. Long non-coding RNA AFAP1-AS1 accelerates lung cancer cells migration and invasion by interacting with SNIP1 to upregulate c-Myc. Signal Transduction and Targeted Therapy 2021, 6, 240. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Tang, H.; Ling, L.; Li, N.; Jia, X.; Zhang, Z.; Wang, X.; Shi, L.; Yin, J.; Qiu, N.; et al. LINC01638 lncRNA activates MTDH-Twist1 signaling by preventing SPOP-mediated c-Myc degradation in triple-negative BC. Oncogene 2018, 37, 6166–6179. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Luan, X.; Han, G.; Guo, K.; Wang, S.; Zhang, X. LINC01638 lncRNA mediates the postoperative distant recurrence of bladder cancer by upregulating ROCK2. Oncol Lett 2019, 18, 5392–5398. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Tang, H.; Wu, J.; Qiu, X.; Kong, Y.; Zhang, L.; Xie, X.; Xiao, X. Linc01638 Promotes Tumorigenesis in HER2+ BC. Curr Cancer Drug Targets 2019, 19, 74–80. [Google Scholar] [CrossRef]
- Geng, C.; Kaochar, S.; Li, M.; Rajapakshe, K.; Fiskus, W.; Dong, J.; Foley, C.; Dong, B.; Zhang, L.; Kwon, O.J.; et al. SPOP regulates prostate epithelial cell proliferation and promotes ubiquitination and turnover of c-MYC oncoprotein. Oncogene 2017, 36, 4767–4777. [Google Scholar] [CrossRef]
- Kong, Q.; Qiu, M. Long noncoding RNA SNHG15 promotes human BC proliferation, migration and invasion by sponging miR-211-3p. Biochem Biophys Res Commun 2018, 495, 1594–1600. [Google Scholar] [CrossRef]
- Liu, L.B.; Jiang, Z.J.; Jiang, X.L.; Wang, S. Up-regulation of SNHG15 facilitates cell proliferation, migration, invasion and suppresses cell apoptosis in BC by regulating miR-411-5p/VASP axis. Eur Rev Med Pharmacol Sci 2020, 24, 1899–1912. [Google Scholar] [CrossRef]
- Zhu, M.; Xiang, H.; Peng, Z.; Ma, Z.; Shen, J.; Wang, T.; Chen, L.; Cao, D.; Gu, S.; Wang, M.; et al. Silencing the expression of lncRNA SNHG15 may be a novel therapeutic approach in human BC through regulating miR-345-5p. Annals of Translational Medicine 2022, 10, 1173. [Google Scholar] [CrossRef]
- Mi, H.; Wang, X.; Wang, F.; Li, L.; Zhu, M.; Wang, N.; Xiong, Y.; Gu, Y. SNHG15 Contributes To Cisplatin Resistance In BC Through Sponging miR-381. Onco Targets Ther 2020, 13, 657–666. [Google Scholar] [CrossRef]
- Saeinasab, M.; Bahrami, A.R.; González, J.; Marchese, F.P.; Martinez, D.; Mowla, S.J.; Matin, M.M.; Huarte, M. SNHG15 is a bifunctional MYC-regulated noncoding locus encoding a lncRNA that promotes cell proliferation, invasion and drug resistance in colorectal cancer by interacting with AIF. Journal of Experimental & Clinical Cancer Research 2019, 38, 172. [Google Scholar] [CrossRef]
- Du, J.; Zhong, H.; Ma, B. Targeting a novel LncRNA SNHG15/miR-451/c-Myc signaling cascade is effective to hamper the pathogenesis of BC (BC) in vitro and in vivo. Cancer Cell International 2021, 21, 186. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Zhong, H.; Ma, B. Targeting a novel LncRNA SNHG15/miR-451/c-Myc signaling cascade is effective to hamper the pathogenesis of BC (BC) in vitro and in vivo. Cancer Cell Int 2021, 21, 186. [Google Scholar] [CrossRef]
- Loewer, S.; Cabili, M.N.; Guttman, M.; Loh, Y.H.; Thomas, K.; Park, I.H.; Garber, M.; Curran, M.; Onder, T.; Agarwal, S.; et al. Large intergenic non-coding RNA-RoR modulates reprogramming of human induced pluripotent stem cells. Nat Genet 2010, 42, 1113–1117. [Google Scholar] [CrossRef] [PubMed]
- Peña-Flores, J.A.; Enríquez-Espinoza, D.; Muela-Campos, D.; Álvarez-Ramírez, A.; Sáenz, A.; Barraza-Gómez, A.A.; Bravo, K.; Estrada-Macías, M.E.; González-Alvarado, K. Functional Relevance of the Long Intergenic Non-Coding RNA Regulator of Reprogramming (Linc-ROR) in Cancer Proliferation, Metastasis, and Drug Resistance. Non-Coding RNA 2023, 9, 12. [Google Scholar] [CrossRef]
- Jiang, B.; Zhu, H.; Tang, L.; Gao, T.; Zhou, Y.; Gong, F.; Tan, Y.; Xie, L.; Wu, X.; Li, Y. Apatinib Inhibits Stem Properties and Malignant Biological Behaviors of BC Stem Cells by Blocking Wnt/β-catenin Signal Pathway through Downregulating LncRNA ROR. Anti-Cancer Agents in Medicinal Chemistry 2022, 22, 1723–1734. [Google Scholar] [CrossRef]
- Hou, L.; Tu, J.; Cheng, F.; Yang, H.; Yu, F.; Wang, M.; Liu, J.; Fan, J.; Zhou, G. Long noncoding RNA ROR promotes BC by regulating the TGF-β pathway. Cancer Cell International 2018, 18, 142. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, A.; Ho, T.T.; Zhang, Z.; Zhou, N.; Ding, X.; Zhang, X.; Xu, M.; Mo, Y.Y. Linc-RoR promotes c-Myc expression through hnRNP I and AUF1. Nucleic Acids Res 2016, 44, 3059–3069. [Google Scholar] [CrossRef]
- Shih, J.-W.; Kung, H.-J. Long non-coding RNA and tumor hypoxia: new players ushered toward an old arena. Journal of Biomedical Science 2017, 24, 53. [Google Scholar] [CrossRef]
- Gao, F.-y.; Li, X.-t.; Xu, K.; Wang, R.-t.; Guan, X.-x. c-MYC mediates the crosstalk between BC cells and tumor microenvironment. Cell Communication and Signaling 2023, 21, 28. [Google Scholar] [CrossRef]
- He, L.; Tang, L.; Wang, R.; Liu, L.; Zhu, P.; Jiang, K.; Tu, G. Long noncoding RNA KB-1980E6.3 promotes BC progression through the PI3K/AKT signalling pathway. Pathology - Research and Practice 2022, 234, 153891. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; He, F.; Hou, Y.; Tu, G.; Li, Q.; Jin, T.; Zeng, H.; Qin, Y.; Wan, X.; Qiao, Y.; et al. A novel hypoxic long noncoding RNA KB-1980E6.3 maintains BC stem cell stemness via interacting with IGF2BP1 to facilitate c-Myc mRNA stability. Oncogene 2021, 40, 1609–1627. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Yu, G.; Li, H.; Wang, M.; Cui, Y.; Zhang, T.; Song, T.; Liu, C. Mild chronic hypoxia-induced HIF-2α interacts with c-MYC through competition with HIF-1α to induce hepatocellular carcinoma cell proliferation. Cellular Oncology 2021, 44, 1151–1166. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Fu, F.; Chen, Y.; Qiu, W.; Lin, S.; Yang, P.; Huang, M.; Wang, C. Genetic variants in long noncoding RNA H19 contribute to the risk of BC in a southeast China Han population. Onco Targets Ther 2017, 10, 4369–4378. [Google Scholar] [CrossRef]
- Reik, W.; Walter, J. Genomic imprinting: parental influence on the genome. Nature Reviews Genetics 2001, 2, 21–32. [Google Scholar] [CrossRef]
- Tessier, C.R.; Doyle, G.A.; Clark, B.A.; Pitot, H.C.; Ross, J. Mammary tumor induction in transgenic mice expressing an RNA-binding protein. Cancer Res 2004, 64, 209–214. [Google Scholar] [CrossRef]
- Barsyte-Lovejoy, D.; Lau, S.K.; Boutros, P.C.; Khosravi, F.; Jurisica, I.; Andrulis, I.L.; Tsao, M.S.; Penn, L.Z. The c-Myc oncogene directly induces the H19 noncoding RNA by allele-specific binding to potentiate tumorigenesis. Cancer Res 2006, 66, 5330–5337. [Google Scholar] [CrossRef]
- Wang, J.; Sun, J.; Yang, F. The role of long non-coding RNA H19 in BC. Oncol Lett 2020, 19, 7–16. [Google Scholar] [CrossRef]
- Barsyte-Lovejoy, D.; Lau, S.K.; Boutros, P.C.; Khosravi, F.; Jurisica, I.; Andrulis, I.L.; Tsao, M.S.; Penn, L.Z. The c-Myc Oncogene Directly Induces the H19 Noncoding RNA by Allele-Specific Binding to Potentiate Tumorigenesis. Cancer Research 2006, 66, 5330–5337. [Google Scholar] [CrossRef]
- Wang, J.; Shi, P.; Teng, H.; Lu, L.; Guo, H.; Wang, X. LncRNA DCST1-AS1 Promotes Endometrial Cancer Progression by Modulating the MiR-665/HOXB5 and MiR-873-5p/CADM1 Pathways. Frontiers in Oncology 2021, 11. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, J.; Hu, Y.; Zou, H.; Zhang, X.; Hu, X. Inhibition of lncRNA DCST1-AS1 suppresses proliferation, migration and invasion of cervical cancer cells by increasing miR-874-3p expression. J Gene Med 2021, 23, e3281. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, D.; Zhang, Y.; Yang, Y.; Duan, Y.; An, Y. LncRNA DCST1-AS1 functions as a competing endogenous RNA to regulate FAIM2 expression by sponging miR-1254 in hepatocellular carcinoma. Clin Sci (Lond) 2019, 133, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, J.; Hu, Y.; Zou, H.; Zhang, X.; Hu, X. Inhibition of lncRNA DCST1-AS1 suppresses proliferation, migration and invasion of cervical cancer cells by increasing miR-874-3p expression. The Journal of Gene Medicine 2021, 23, e3281. [Google Scholar] [CrossRef]
- Tang, L.; Chen, Y.; Tang, X.; Wei, D.; Xu, X.; Yan, F. Long Noncoding RNA DCST1-AS1 Promotes Cell Proliferation and Metastasis in Triple-negative BC by Forming a Positive Regulatory Loop with miR-873-5p and MYC. J Cancer 2020, 11, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Kuo, W.L.; Stilwell, J.L.; Takano, H.; Lapuk, A.V.; Fridlyand, J.; Mao, J.H.; Yu, M.; Miller, M.A.; Santos, J.L.; et al. Amplification of PVT1 contributes to the pathophysiology of ovarian and BC. Clin Cancer Res 2007, 13, 5745–5755. [Google Scholar] [CrossRef]
- Carramusa, L.; Contino, F.; Ferro, A.; Minafra, L.; Perconti, G.; Giallongo, A.; Feo, S. The PVT-1 oncogene is a Myc protein target that is overexpressed in transformed cells. J Cell Physiol 2007, 213, 511–518. [Google Scholar] [CrossRef]
- Cho, S.W.; Xu, J.; Sun, R.; Mumbach, M.R.; Carter, A.C.; Chen, Y.G.; Yost, K.E.; Kim, J.; He, J.; Nevins, S.A. Promoter of lncRNA gene PVT1 is a tumor-suppressor DNA boundary element. Cell 2018, 173, 1398–1412 e1322. [Google Scholar] [CrossRef]
- Tseng, Y.-Y.; Bagchi, A. The PVT1-MYC duet in cancer. Molecular & Cellular Oncology 2015, 2, e974467. [Google Scholar] [CrossRef]
- Sarver, A.L.; Murray, C.D.; Temiz, N.A.; Tseng, Y.Y.; Bagchi, A. MYC and PVT1 synergize to regulate RSPO1 levels in BC. Cell Cycle 2016, 15, 881–885. [Google Scholar] [CrossRef]
- Ma, F.; Liu, X.; Zhou, S.; Li, W.; Liu, C.; Chadwick, M.; Qian, C. Long non-coding RNA FGF13-AS1 inhibits glycolysis and stemness properties of BC cells through FGF13-AS1/IGF2BPs/Myc feedback loop. Cancer Lett 2019, 450, 63–75. [Google Scholar] [CrossRef]
Figure 1.
Mechanism of action of lncRNA in cancer cells. LncRNAs can be classified as guides, signaling molecules, decoy molecules, enhancer molecules, or scaffold molecules. LncRNAs can interact with DNA, RNA, and proteins in the cell. This interaction can regulate gene expression, cell proliferation, invasion, and metastasis. They can also modulate transcription factors, RNA-binding proteins, and microRNAs.
Figure 1.
Mechanism of action of lncRNA in cancer cells. LncRNAs can be classified as guides, signaling molecules, decoy molecules, enhancer molecules, or scaffold molecules. LncRNAs can interact with DNA, RNA, and proteins in the cell. This interaction can regulate gene expression, cell proliferation, invasion, and metastasis. They can also modulate transcription factors, RNA-binding proteins, and microRNAs.
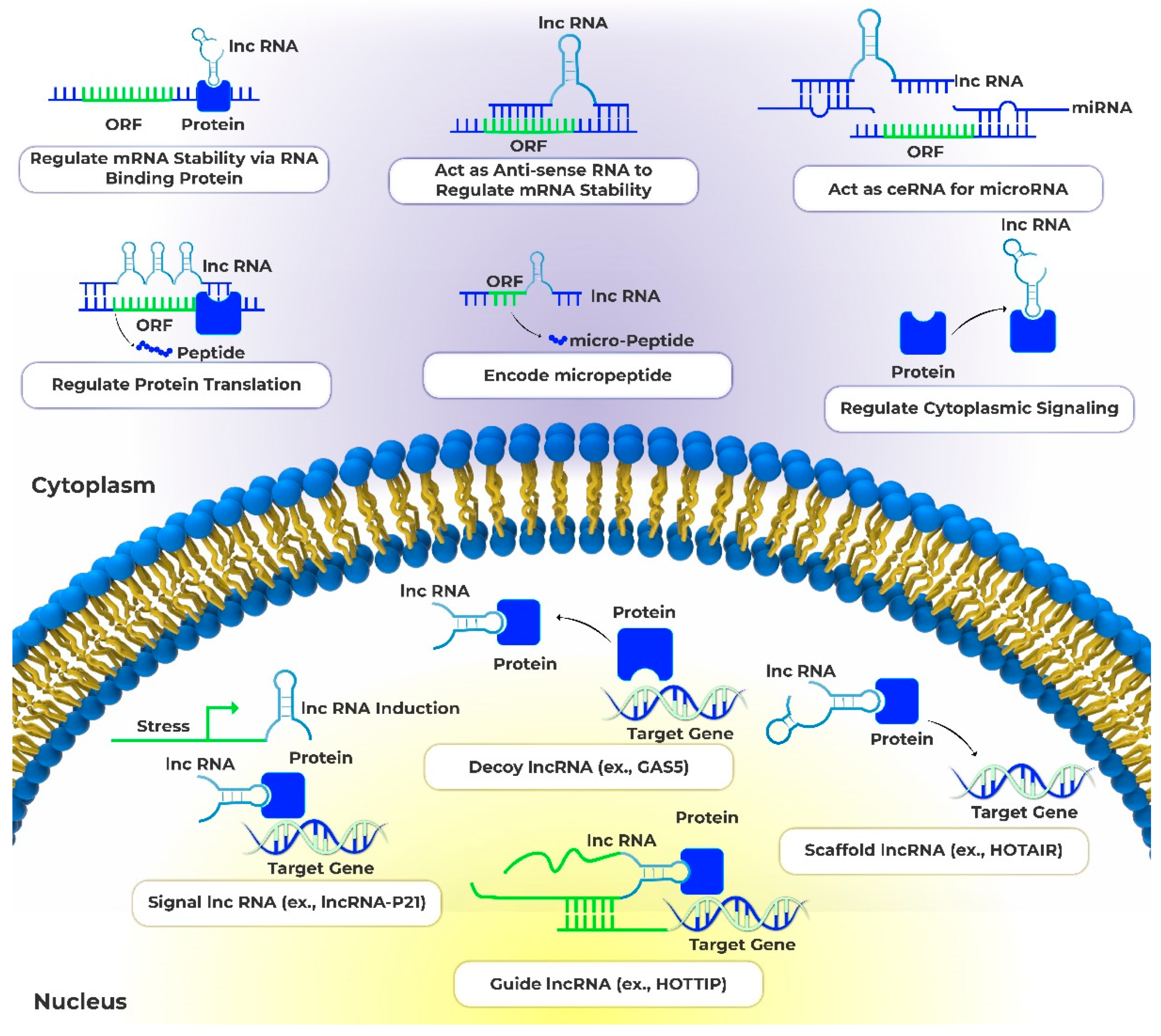
Figure 2.
Crosstalk between lncRNA and Myc in BC. The transcription factor Myc regulates the expression of many lncRNAs involved in cancer pathogenesis. Conversely, multiple lncRNAs can modulate Myc expression and activity through diverse mechanisms, forming complex bidirectional regulatory loops. Additionally, certain lncRNAs regulate miRNAs that target Myc, thereby increasing Myc levels. Through these mechanisms, the interactions between Myc and lncRNAs potentiate oncogenic signaling, promoting cell proliferation, tumor growth, metastasis and other cancer phenotypes. The picture was created using Adobe Illustrator CC version 20.
Figure 2.
Crosstalk between lncRNA and Myc in BC. The transcription factor Myc regulates the expression of many lncRNAs involved in cancer pathogenesis. Conversely, multiple lncRNAs can modulate Myc expression and activity through diverse mechanisms, forming complex bidirectional regulatory loops. Additionally, certain lncRNAs regulate miRNAs that target Myc, thereby increasing Myc levels. Through these mechanisms, the interactions between Myc and lncRNAs potentiate oncogenic signaling, promoting cell proliferation, tumor growth, metastasis and other cancer phenotypes. The picture was created using Adobe Illustrator CC version 20.
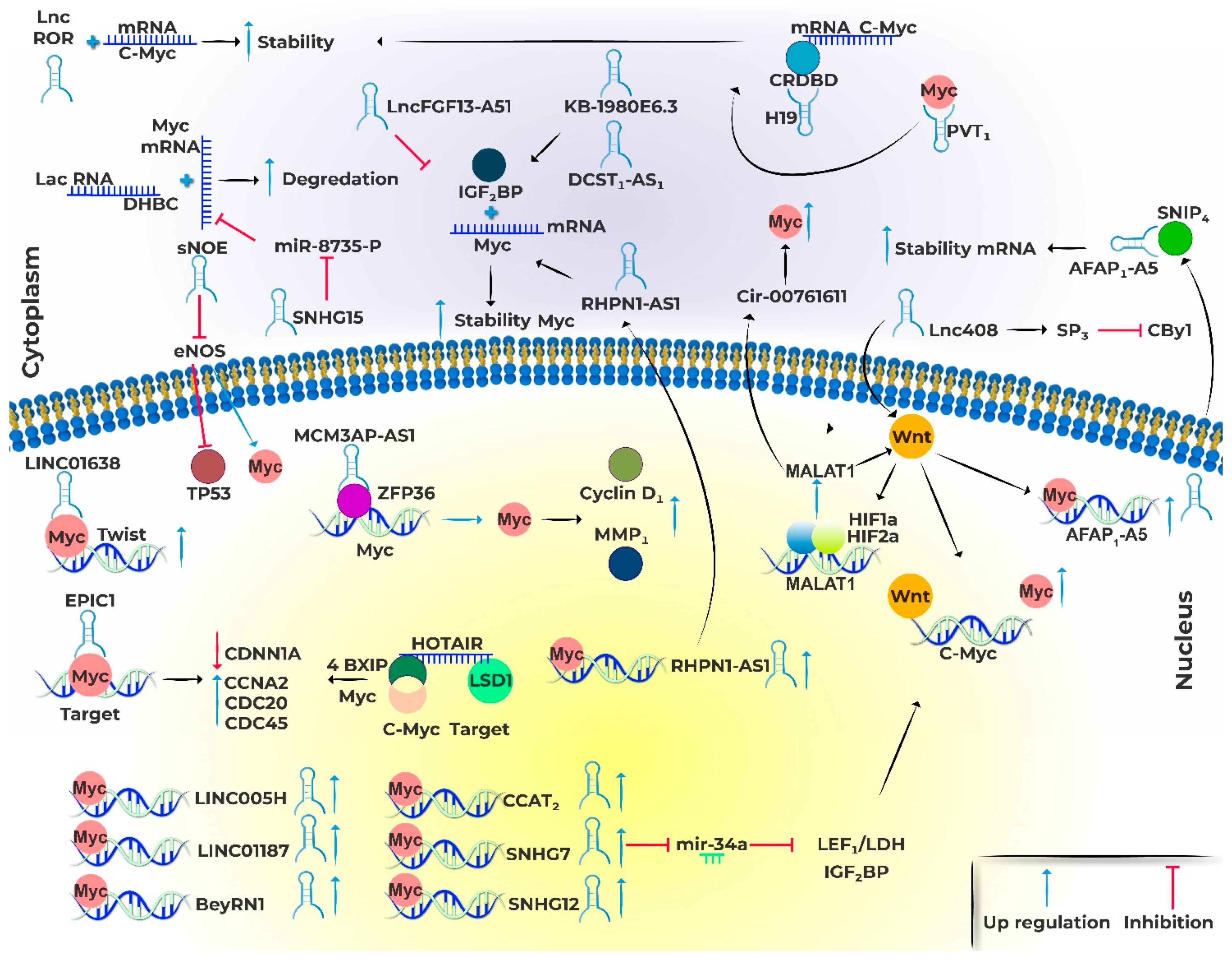

Table 1.
Function and cellular mechanism between Myc and lncRNAs involved in BC.
| LncRNA | Function | Cellular mechanism(s) | Regulation |
|---|---|---|---|
| lncRNAs regulated by MYC | |||
| SNHG7 | c-Myc binds to SNHG7 promoter | Cancer cell proliferation | Positive |
| SNHG12 | c-Myc direct regulated to SNHG7 expression | Cancer cell proliferation, tumor growth, migration, and invasion | Positive |
| BCYRN1 | BCYRN1 regulated by Myc | Cancer cell migration and invasion | Positive |
| lncRNAs affecting MYC Expression | |||
| LacRNA | LacRNA inhibits Myc activity LacRNA stabilizes PHB2 |
Cancer cell metastasis suppression | Negative |
| Linc00839 | Myc binds to Linc00839's promoter and activates its transcription Linc00839 increases the expression of Myc Linc00839 increases the Lin28B and activated the PI3K/AKT signaling |
cancer cell, proliferation, invasion, and migration | Positive |
| Lnc408 | Lnc408 recruits a SP3 Lnc408 suppresses CBY1 transcription |
Influences BC stem cells | Positive |
| LINC01287 | LINC01287 upregulates Myc expression activates the Wnt/β-catenin signaling |
Cancer cell proliferation and metastatic | Positive |
| MCM3AP-AS1 | MCM3AP-AS1 binds ZFP36 and regulates Myc | Cancer cell proliferation, apoptosis, migration, and invasion | Positive |
| LINC00511 | LINC00511 activates Wnt/β-catenin, increasing the expression of c-Myc |
Cancer cell promoting stemness and invasiveness, inhibit apoptosis | Positive |
| sONE | Knocking down of sONE decreases in TP53 and increases in c-Myc sONE can alter tumor suppressors miRNAs (miR-34a, miR-15, miR-16, and let-7a) |
cancer cell proliferation, colony-forming ability, migration, and invasion capacities | Negative |
| MALAT1 | MALAT1 activates Wnt/β-catenin, increasing the expression of c-Myc |
tumor size, lymph metastasis, angiogenesis, stage tumor, progression and metastasis | Positive |
| PICART1 | PICART1 influencing the AKT/GSK3β/β-catenin, activates the expression of cyclin D1 and c-Myc | Cancer cell proliferation | Positive |
| MYC-lncRNAs regulatory loops | |||
| CCAT2 | CCAT2 regulates Myc expression. c-Myc binds to CCAT2 promoter | cancer cell proliferation migration and metastasis | Positive |
| EPIC1 | EPIC1 promotes Myc recruitment to its target genes. | Cancer cell proliferation | Positive |
| HOTAIR | HOTAIR contributes to EZH2 recruitment to the Myc promoter HOTAIR promoter enriched for both NF-kB and c-Myc binding sequences |
Cancer cell proliferation by regulating cell cycle and apoptosis | Positive |
| RHPN1-AS1 | RHPN1-AS1 is a molecular sponge of miR-4261 RHPN1-AS1 is a transcriptional target of c-Myc RHPN1-AS1 exerts tumorigenesis by regulating P53 expression via MDM2 gene. |
cancer cells proliferation | Positive |
| lncRNAs affecting MYC stability/translation | |||
| SNHG15 | SNHG15 is directly regulated by Myc | Cancer cell proliferation, migration and invasion | Positive |
| Linc-RoR | linc-ROR increases c-Myc mRNA stability Linc-RoR activates Wnt/β-catenin |
Cancer cell proliferation, metastasis, drug resistance, and invasion | Positive |
| KB-1980E6.3 | It acts enhancing c-Myc mRNA stability via interaction with IGF2BP1 | Promote Cancer stem cell stemness, metabolic rewiring and tumorigenesis. | Positive |
| H19 | It acts enhancing c-Myc mRNA stability via interaction with CRD-BP | Cancer cell proliferation, migration and invasion | Positive |
| DCST1-AS1 | MYC regulates DCST1-AS1 expression DCST1-AS1 interaction with miR-873-5p upregulated IGF2BP1, Myc , CD44 and LEF1 |
cancer cell proliferation and metastasis | Positive |
| LINC01638 | LINC01638 interacts with c-Myc to prevent SPOP-mediated to c-Myc ubiquitination | cancer cell proliferation, metastasis and EMT | Positive |
| AFAP1-AS1 | AFAP1-AS1 activated Wnt/β-catenin, increasing the expression of c-Myc AFAP1-AS1 interacts with SNIP1 to prevent c-Myc ubiquitination |
Cancer cell proliferation and invasion inhibited cell apoptosis |
Positive |
| PVT1 | PVT1 can enhance Myc protein stability They amplify each other in a molecular loop |
cancer cell proliferation and metastasis | Positive |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



