Submitted:
24 July 2023
Posted:
26 July 2023
You are already at the latest version
Abstract
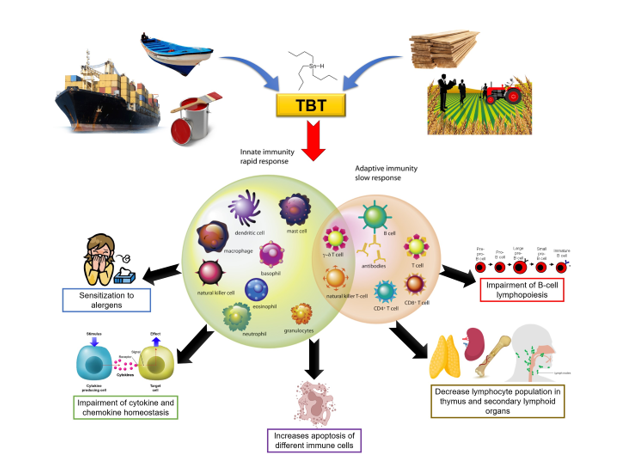
Tributyltin (TBT) is an environmental contaminant present on all continents, including Antarctica, with a potent biocidal action. Its use began to be intensified during the 1970s, being effectively banned in 2003, but remaining in the environment to this day due to several factors that increase its half-life and misuse despite the bans. In addition to the endocrine disrupting effect of TBT, that may lead to imposex induction in some invertebrate species, there are several studies that demonstrated that TBT also has an immunotoxic effect. The immunotoxic effects that have been observed experimentally in vertebrates using in vitro and in vivo models involve different mechanisms, but mainly alterations in the expression and/or secretion of cytokines. In this review, we summarize and update the literature on the impacts of TBT on the immune system, as well as discuss what still needs to be done to fill the knowledge gaps regarding the impact of this endocrine disrupting-chemical on immune system homeostasis.
Keywords:
endocrine-disrupting chemical
; tributyltin
; immune system
; organotin
; cytokines
; immunotoxicity
1. Introduction
1.1. Tributyltin
Organotin compounds are tin-based chemicals made up of hydrocarbons (Caseri, 2014). The use of these compounds ranges from industrial to agricultural biocidal agents such as antifungals, acaricides and molluscicides, being applied as wood preservatives or in antifouling paints. (Maguire, 1987; Davies, 2004). Among these organotin compounds, is tributyltin (TBT), which began to be widely used in antifouling paints in the mid-1960s, given its low cost and its effectiveness in protecting ship hulls and underwater marine equipment against biofouling (Kotrikla, 2009).
After years of extensive use, adverse effects of TBT were described as it spread into the marine environment, being harmful to aquatic organisms. For example, in the invertebrate Nucella lapillus, a low concentration of 1ng/L was able to induce imposex, the development of male sexual characteristics in females, as seen in populations of this invertebrate along the coast of the United Kingdom (Gibbs et al., 1987). In the commercial oyster Crassostrea gigas, the concentration of 20 ng/L of TBT affected larval growth and concentrations smaller than 2 ng/L inhibited its calcification, making it impossible to fish for oysters in Arcachon Bay, France (Alzieu, 2000). In vertebrates, imposex effect has also been described. The first report of imposex in vertebrates showed that doses of 0.1 µg/g of TBT present in the diet of Paralichthys olivaceus were sufficient for the masculinization of females (Shimasaki et al., 2003). Nevertheless, until 2011, the imposex effect of TBT had been described in more than 260 species of marine gastropods. Moreover, environmental damage and economic losses had already spread over several continents (Titley-O’Neal et al., 2011).
Several restrictions against TBT use on antifouling paints were firstly imposed in France, the United Kingdom, and other developed countries during the 1980s (Yebra et al., 2004). Years later, the International Maritime Organization (IMO) passed a global ban on the application of antifouling paints containing TBT (from January 1, 2003) and a ban on their presence on the surface of ships' hulls (from January 2008) (IMO, 2005). Even so, in 2004, it was estimated that 70-80% of the world naval fleet used TBT copolymer in its composition, given its economic benefits (Yebra et al., 2004). Also, in 2014 a United States company was fined for clandestinely producing and selling antifouling paints with TBT to various regions of the Caribbean. Nonetheless, the problem persists because as recently as 2021 these paints could still be found for sale on the internet and be sent to different regions such as the Caribbean, Central America and Oceania (Uc-Peraza et al., 2022).
Several factors can increase the half-life of TBT in the environment, such as its ability to deposit in marine sediments and continue to be released into the environment for up to 100 years, according to mathematical models (Hamer & Karius, 2005). These factors contributed for TBT values exceeding 7000 ng Sn/g to be found in an environmental reserve in the Virgin Islands (Hartwell et al., 2016). TBT also bioaccumulates in several marine species that are at the base of the food chain (Takahashi et al., 1999). TBT can be found in these organisms even after 20 years of the initial contamination, maintained by their lipid solubility. Furthermore, the biomagnification of contamination occurs in the food chain, making it possible to find TBT residues in fish, seabirds and marine mammals (Tanabe et al., 1998).
Given the risk to human health due to the ingestion of products contaminated with TBT (e.g., seafood, water), the World Health Organization (WHO) has defined an acceptable daily intake value of 250 ng/Kg/day of TBT (WHO- IPCS, 1999a). This value was extrapolated by a factor of 100 due to toxicity, kinetics and inter-individual differences tests performed in rats, in which the addition of TBT to the diet led to a reduction in the weight and function of the thymus of these animals (Penninks, 1993). Nevertheless, it is important to highlight that distinct diets may result in different levels of TBT contamination in human blood and tissues, but this is not the only determining factor for exposure to TBT (Antizar-Ladislao, 2008).
Some studies have shown that chronic exposure to TBT, even in values lower than acceptable daily intake, may lead to different complications. It has been shown that male rats treated with 0.5 and 50 µg/kg TBT become obese due to increased adiposity and present hepatic steatosis and insulin and leptin resistance (Zuo et al., 2011). Female rats treated with 100 ng/Kg/day of TBT showed signs of dysfunction of the hypothalamic-pituitary-adrenal axis, these being inflammation, oxidative stress, and fibrosis (Merlo et al., 2016). Finally, female rats treated via gavage with 100 or 500ng/Kg/day of TBT showed abnormalities in renal function, with decreased glomerular filtration, increased levels of proteinuria, inflammation, oxidative stress, and kidney fibrosis (Coutinho et al., 2016).
Another important adverse effect of TBT is its immunotoxicity. In the present review we aimed to summarize and update the current literature on the impact of TBT on the immune system, opening the door for new questions about the impact of this endocrine disruptor.
1.2. The Immune System
In response to a stimulus, like a challenge by a pathogen, the immune system generates innate and adaptive immune responses. Briefly, the innate response is immediate, less specific, and primarily involves the action of monocytes, neutrophils, and NK cells. In contrast, the adaptive response is built gradually, being more specific, long-lasting, and mainly orchestrated by B lymphocytes, through the production of antibodies, and by T lymphocytes that coordinate cell-mediated immune response (Nunes-Silva et al., 2018; Moss et al., 2004).
Regarding T lymphocytes, the population of Tαβ lymphocytes is mainly divided into two main subpopulations, CD4+ and CD8+ T lymphocytes, named according to their expression of some surface protein, called clusters of differentiation (CD). These cells have different functions; CD8+ T lymphocytes can induce the death of tumor cells or cells infected by viral pathogens, via class I major histocompatibility complex (MHC) recognition, and producing inflammatory mediators interleukin (IL)-2, interferon gamma (IFN-γ) and tumor necrosis factor alpha (TNF-α) (Koyasu & Moro, 2012; Wang et al., 2018). CD4+ T lymphocytes are characterized by the recognition of MHC class II, expressed on the surface of antigen-presenting cells (APCs), and can differentiate into different subpopulations depending on the cytokines present in the microenvironment (Gutcher & Becker, 2007; Koyasu & Moro, 2012). In fact, the balance of synthesis and secretion (autocrine and paracrine signaling) of different cytokines by immune cells and other cell types such as endothelial cells, fibroblasts, and bone marrow cells, maintains the multicellular network of communication in the microenvironment (Borish & Steinke, 2003). The main subpopulations of CD4+ T lymphocytes are: T helper (Th)1 cells that are capable of promoting a cell-mediated effector response, against viruses, intracellular bacteria and protozoa, and characterized by the production of cytokines such as IFN-γ and IL-2 (Belizario et al., 2016); Th2 cells, that are involved in the coordination of the humoral response, mainly against parasites (for example, helminths), and in the pathophysiology of several allergies (including asthma and atopic dermatitis) and are characterized by the production of IL-4, IL-5 and IL-13 (Koyasu & Moro, 2012; Belizário et al., 2016); Th17 cells that are involved in the defense against pathogens in the mucosa and in autoimmune diseases, where there is hyperactivation of Th17 (as in rheumatoid arthritis), and are characterized by the production of IL-17 and IL-22 (Koyasu & Moro, 2012; Zambrano-Zaragosa et al., 2014; Zayas & Mamede, 2022). Another important subtype of T lymphocytes is regulatory T cells, which have a primary function of suppressing the activation of the immune system, more specifically inhibiting the activation and expansion of CD4+ and CD8+ T lymphocytes and B cell activation, preventing the exacerbation of inflammation. Regulatory T cells can be characterized by the expression of CD4, CD25 and FOXP3 molecules and the secretion of granzyme B, TGF-β and IL-10 (Rajendeeran & Tenbrock, 2021).
The balance of synthesis and secretion (autocrine and paracrine signaling) of different cytokines by immune cells and other cell types such as endothelial cells, fibroblasts, and bone marrow cells, maintains the multicellular network of communication in the microenvironment (Borish & Steinke, 2003). In fact, cytokines are the basis of communication for initiation, maintenance, and termination of immune responses to infections and any change can lead to extremes, such as immunosuppression or the establishment of chronic inflammation (Lawrence et al., 2016). TNF-α, for example, modulates the function of effector immune cells, such as neutrophil, promoting an increase in their activity, adherence, and chemotaxis. On the other hand, IFN-γ promotes an increase in antitumor activity of natural killer cells, phagocytic activity of macrophages and production of nitric oxide, in addition to the increased expression of MHC class I and II molecules. Interleukin-1β (IL-1β) affects the maturation, proliferation, and synthesis of immunoglobulins by B lymphocytes, in addition to stimulating the synthesis of acute phase proteins at the beginning of the inflammatory process (Borish & Steinke, 2003; Gabay et al., 2010; Feng et al., 2015). In contrast, other cytokines, such as IL-10, have an immunoregulatory function, inhibiting the synthesis of cytokines such as IFN-γ, TNF-α, IL-12, IL-2 and IL-1β, thus inhibiting the inflammatory process and promoting a change in the profile of macrophages from the activated state to the tissue-resident phenotype (Borish & Steike, 2003; Galdiero et al., 2013; Nagata & Nishiyama, 2021).
It is worth noting that the immune system does not act alone, but in close connection with other systems, such as the endocrine system (Ponce-Regalado et al., 2022). The mechanism of communication between the immune and endocrine systems occurs in “two-way”, as some immune cells are capable of producing hormones, and are also sensitive to hormonal action (Sakazaki et al., 2002; Lardone et al., 2011; Csaba, 2014; Csaba, 2018; Fan et al., 2019; Tramontana et al., 2021). A good example is that of estrogen, which can modulate the production of cytokines, differentiation, proliferation and even apoptosis of cells of the immune system such as T and B lymphocytes, mast cells, basophils, and eosinophils via estrogen receptors (ERs) that are expressed by these cells (Fan et al., 2019). Cell populations of the immune system may differ in the expression of estrogen receptors, such as CD4+ T lymphocytes that express greater amounts of ERα than B lymphocytes, which, in contrast, express greater amounts of ERβ than CD4+ T cells. In parallel, CD8+ T lymphocytes express both ERα and ERβ receptors in low amounts, but at equal rates (Phiel et al., 2005; Cunningham & Gilkeson, 2011). Interestingly, TBT has been described as an antagonist of human estrogen receptors by inhibiting the transcriptional activation of the ER-dependent reporter gene and the interaction between ERβ LBD and the coactivator SRC1 (Cho et al., 2012; Beg et al., 2022). It is also known that different cytokines and chemokines have targets in the hypothalamic-pituitary-thyroid axis and in situations of deregulation in the balance of these signals an impact on this axis, mainly on thyroid functioning, can occur (Croce et al., 2021). Therefore, exposure to endocrine disruptors, like TBT, can impact innate and/or adaptive response, more specifically interfering with the cellular and humoral immune responses, as well as the lifespan of immune cells (Nowak et al., 2019).
1.3. TBT and the Immune System
The relationship between TBT contamination and immunotoxicity was first observed in animals that live exclusively or mainly in aquatic environments. In in vivo experiments with the fish Oncorhynchus mykiss, TBT led to thymic atrophy and reduced circulating lymphocyte population (Schwaiger et al., 1994). Moreover, Bottlenose dolphins (Tursiops truncatus) and sea otters (Enhydra lutris), which lived in contaminated sites, were found dead along the US coast and had high tissue concentrations of TBT. The mortality of both species has been linked to a lower ability of the immune system to fight infectious diseases (Kannan et al., 1997; Murata et al., 2008). In vitro experiments that used seal cells (Phoca vitulina) showed that doses between 50-200 nM of TBT and its metabolite dibutyltin (DBT) reduced the antitumor capacity of natural killer (NK) cells, decreased the proliferation of T cells, and that 100-200 nM of DBT were able to decrease macrophage phagocytic activity (Frouin et al., 2008).
More recently studies have shown the impacts of TBT on several other species, including mammals (Schwaiger et al., 1994; Kannan et al., 1997; Murata et al., 2008; Frouin et al., 2008). Moreover, over the last decades tests have been carried out using animal models (mainly mice) and human cells, which will be addressed throughout this article, as a way of inferring whether TBT immunotoxic effects would also be seen in humans.
C57BL/6 mice that had TBT inserted in their diet for a period of two weeks showed a reduction in the number of lymphocytes in the spleen and lymph nodes, thymic atrophy, and an increase in serum immunoglobulin M, but a decrease in immunoglobulin G (Ohtaki et al., 2007). In addition, BALB/c mice injected with 100nM of TBT showed an increase in serum levels of the cytokines IFN-γ, TNF-α and IL-13 and the chemokines MIP-1β and RANTES. A decrease in the cytokine IL-2, which is essential for the maturation of B and T lymphocytes, was also observed (Lawrence et al., 2016). In splenic cells of CBA/J mice stimulated in vitro with anti-mouse CD3 antibodies, exposure to 0,1µM of TBT was sufficient to inhibit the secretion of IFN-γ and IL-4 by T CD4 cells (Yano et al., 2003). ICR mice treated in vivo with doses of 4 and 20 mg/Kg of TBT showed a positive linear increase in thymocyte apoptosis and Fas expression, indicating that TBT may be leading to cell apoptosis (Chen et al., 2011). Furthermore, low doses of TBT (0.1, 1, 3 and 10 nM) were also able of inducing apoptosis of Balb/c thymocytes in vitro via an increase in reactive oxygen species (ROS), reduction of growth hormone (GSH), depolarization of mitochondrial membrane and activation of caspase-3 (Sharma & Kumar, 2014). In Balb/c and C57BL/6 T lymphocytes, it was seen that TBT (100 nM) in vitro induces the differentiation of T cells to the Th2 phenotype, which is characterized by the production of IL-10 and suppression of the production of IL-12, one of the main cytokines responsible for the differentiation of CD4+ T lymphocytes into the Th1 phenotype (Kato et al., 2004; Kato et al., 2006). It was also observed that in C57BL/6 mice treated in vivo with TBT (6 μmol/Kg) there was greater ovalbumin-induced airway inflammation than in untreated mice. Furthermore, in the bronchoalveolar lavage (BALF) of these TBT-treated animals there was increased number of eosinophils and IL-5 levels, as well as IgE levels in serum, which was expected, given that Th2 cells are involved in the response to allergens (Kato et al., 2006). Part of this effect was due to an increase in oxidative stress in secondary lymphoid organs of C57BL/6 animals due to TBT treatment and, interestingly, Th17 lymphocytes had no role in the increased inflammation observed in the respiratory tract (Kato et al., 2013).
After oral treatment of ICR mice with 1, 10 and 20 mg/Kg of TBT, a decrease in the percentage of CD4+ and CD8+ T cell populations was observed in the thymus, but with an increase in the percentage of the CD4+CD8+ and CD4-CD8- T cell populations and decreased expression of IL-7. (Im et al., 2015). According to the same study, the spleens of ICR animals were also analyzed during TBT treatment and a decrease in the percentage of naive CD44-CD62L+ and effector/memory CD44+ T cells was observed. Collectively, these data suggest that TBT impairs the development of T cells in thymus and in spleen (Im et al., 2015). Regarding B lymphocytes, in an C57BL/6 in vitro model, exposure to TBT promoted apoptosis of pro-B cells. In an C57BL/6 ex vivo model, TBT directly affects the differentiation of B cells in addition to altering the bone marrow microenvironment (Baker et al., 2017). In macrophages, exposure of the murine J774.1 cell line to TBT (1 - 1.5 µM) induced an increase in TNF-α expression and an increase in caspase-3 activity leading to an increase in the apoptosis of these cells (Nakano et al., 2004). Additionally, treatment of J774A.1 cell line with 0.4, 0.6, 1 and 1.2 µM of TBT promoted cell death possibly via RP1 and RP3 receptors, which are related to necroptosis. Furthermore, bone marrow-derived macrophages from TBT-treated C57BL/6 mice also displayed increased apoptosis (Ling et al., 2019). Exposure to TBT also induced changes in RAW 264.7 murine macrophages leading to activation of the inflammasome complex (Childers et al., 2021). Finally, macrophages from C57BL/6 mice treated with doses of 250 µg/kg and 500 µg/kg of TBT showed activation of the PPARγ pathway and increased expression of genes related to lipogenesis and lipid metabolism, which could be related to a role of the innate immune system in the TBT promoted obesogenic effect (Jie et al., 2021).
In humans, TBT (1 μM) induced neutrophil apoptosis in vitro via a caspase-dependent mechanism (Lavastre & Girard, 2002). Moreover, TBT (2.5 - 200 nM) in vitro decreased the viability of peripheral blood mononuclear cells (PBMCs) and was able to modulate the production of IL-1β and IFN-γ, in a dose-dependent manner with TBT effect varying according to the concentration and duration of the exposure (Brown & Whalen, 2015; Lawrence et al., 2015). Specifically in monocyte-free human PBMCs, in addition to an increase in IL-1β synthesis and secretion, an increase in IL-6 synthesis and secretion was observed using the same concentrations of TBT (Brown et al., 2018a; Brown et al., 2018b). This data is corroborated given that the inhibition of Toll-like receptors (TLRs 4, 1/2 and 8), in PBMCs without monocytes treated with TBT, leads to a significant decrease in the production of IL-1β and IL-6. This effect probably occurs because exposure to TBT causes the activation of these TLRs, promoting the activation of the MAPK pathway (Alcala et al., 2022). In human natural killer cells (NKs), TBT also modulates IL-1β and TNF-α, promoting an increase in the secretion of these cytokines at low doses (5 - 50 nM for IL-1β and 5-100 nM for TNF-α), but a decrease at high doses (200 nM for both) (Brown & Whalen, 2015; Hurt et al., 2013). Other in vitro studies also showed that NKs exposed to TBT (200 - 300 nM) had lower expression of perforins and granzyme B, as well as lower cytotoxic activity against tumors (TBT 25-500 nM) (Thomas et al., 2003; Dudimah et al., 2007). Specifically in human B lymphocytes, in vitro treatment with TBT (100nM) reduced proliferation, survival, and differentiation of mature B cells (De Santiago & Aguilar-Santelises, 1999). Finally, in human marrow cells, TBT (1 nM) led to a decrease in the percentage of CD19+CD22+ B cells in a mechanism independent of the PPARy pathway (Carfi et al., 2008; Carfi et al., 2010).
In the zebrafish model (Danio rerio), chronic exposure for 8 weeks with different doses of TBT (1, 10 and 100 ng/L) led to a decrease in the activity of antioxidant enzymes (superoxide dismutase, catalase and glutathione peroxidase), intestinal lysozyme and immunoglobulin M (IgM) and to an increase expression of TNF-α, IL-1β, IL-6, nuclear factor-kappa B p65 (NF-κ B p65) and heat shock proteins HSP70 and HSP90 in intestines, indicating that TBT induces oxidative stress and immunotoxicity in zebrafish (Zhang et al., 2017). In another protocol of chronic exposure to TBT with zebrafish for 6 weeks at doses of 10 ng/L, 100 ng/L and 300 ng/L, a decrease in the amount of lysozyme and IgM was also observed along with dysregulation in the production of thyroid hormones (Li & Li., 2021). In Takifugu obscurus fish, different water concentrations of TBT (1.962, 3.924 and 9.81μg/L) for 96 h led to increased production of ROS and concomitantly upregulation of CD28 (a known costimulatory receptor present in T cells surface) in the gills and liver of these animals, suggesting that CD28 plays a role in the response to TBT toxicity (Fang et al., 2019). Also, in the Takifugu obscurus model, the same concentrations of TBT in water (1.962, 3.924 and 9.81µg/L) for 96h and chronic exposure with 900 ng/L for 30 days induced lower mRNA expression of Toll-like receptors (TLRs) 2 and 3 in the gills and higher mRNA expression of TLR18 and TLR22 in the liver and gills when compared to animals not exposed to TBT, confirming these tissues as vital sites in the initial response to exposure to TBT (Zhao et al., 2020).
1.4. Tributyltin and Others Endocrine Disrupting-Chemicals
Sensitization to allergens is one of the side effects caused by endocrine disruptors in the immune system, as they induce the breakdown of homeostasis in the production of cytokines and chemokines, as already summarized in this work in relation to exposure to TBT (Table 1). In the context of bisphenol S (BPS), water intake of 0.4 μg/Kg/day of BPS for 6 weeks concomitantly to the exposure to ovalbumin (OVA) every 2 weeks increased lung inflammation in C3H/HeJ mice, as well as anti-OVA IgE and IgG1 levels in serum and IL-5, IL-13, IL-33 and eotaxin levels in BALF (Yanagisawa et al., 2022). In the mediastinal lymph node of these same animals, there was an increase in the number of total cells and antigen-presenting cells such as dendritic cells. In addition, a restimulation of lymph node cells with OVA in vitro led to an increase in cell proliferation and cytokine production of the Th2 lymphocyte profile, such as IL-4, IL-5 and IL-13, indicating that BPS may be leading to an increase in the number of Th2 cells with greater sensitization to allergens. Similar effects have already been observed for TBT in C57BL/6 mice model (Kato et al., 2004; Kato et al., 2006; Yanagisawa et al., 2022). Furthermore, increased levels of IL-4 and decreased levels of IL-12 were observed in the umbilical cord blood of newborns whose mothers had higher amounts of monoethyl phthalate (MEP), a metabolite of di-ethyl phthalate (DEP), in blood and urine (during weeks 24 to 28 of gestation), indicating a possible polarization of newborn T cells towards the Th2 phenotype (Tsai et al., 2021). Bisphenol A (BPA) at doses of 10 µM, 30 µM and 50 µM for 12h led to a greater translocation of the transcription factor NF-κ B p65 and to an increase in the production of cytokines IL-1β, IL-6 and TNF-α together with nitric oxide and prostaglandin E2 (PGE2) in murine macrophages of the RAW264.7 lineage, suggesting that BPA can induce a pro-inflammatory response in these cells (Huang et al., 2019).
In the context of cancer, a single dose of BPA (250ug/Kg) in newborn Balb/c mice is capable of inducing lung metastasis with increased intratumoral production of IL-1β, IL-6, IFN-γ, TNF-α and VEGF in a model of induced mammary tumorigenesis in which animals were injected in situ with 4T1 tumor cells when they reach sexual maturity (Palacios-Arreola et al., 2022). Interestingly, 10−8 M of BPA was also able to increase the migration of breast ductal carcinoma in situ (DCIS) cells and RAW264.7 macrophages in an in vitro co-culture system. In in vivo experiments with Balb/c mice, exposure to 2.5 µg/L of BPA for 70 days promoted an augment of DCIS primary tumor growth rate, lymph node metastasis, and concomitant increase of protumorigenic M2 macrophages (Kim et al., 2019). It has already been observed that treatment of C57BL/6 mice with 4 mg/Kg DEHP for 21 days before the injection of B16F10 melanoma cells, and for 7 days after the injection, reduced the polarization of macrophages into M1 profile, but increased the polarization into M2 profile, leading to tumor formation and growth (Lee et al., 2018). This same polarization for the M2 profile is seen during in vitro exposure to benzophenone-3 (BP-3) in primary human macrophages (Gregory et al., 2020). Currently, there are no published data on how the exposure of macrophages to TBT would alter the immune response of these cells in the context of cancer, as previously described for other endocrine disruptors.
Similarly, to TBT, individual and co-exposure to the disruptors mancozeb (8000 mg/Kg/day) and fipronil (95 mg/Kg/day) for 29 days by oral gavage led to immunotoxicity in the spleen and thymus of Swiss albino mice (being more prominent in the treatment with both disruptors), as indicated by lower organ weight and cellularity, lower proliferation of splenocytes and thymocytes and higher rates of apoptosis and necrosis of these cells (Bano & Mohanty, 2020). However, injection of Swiss mice with 50 μg/Kg of BPA for 6 weeks led to an increase in the number of lymphocytes and monocytes in the blood and an invasion of lymphocytes and eosinophils into the red pulp of the spleen (Shaibi et al., 2022). In addition, doses of 100 µM of BPA, BPS, BPF and dimethyl terephthalate (DMTP) led to lower proliferation and viability of B lymphocytes, removed from the spleen of mice, and stimulated with LPS, with BPA being the most toxic for B cells among them (Jang et al., 2020). Interestingly, even though TBT is a PPARy agonist, it can induce a reduction in the mature population of B lymphocytes regardless of activation of the PPARy pathway by inducing changes in the bone marrow microenvironment that leads to adipogenesis in favor of lymphopoiesis (Carfi et al., 2008; Carfi et al., 2010; Baker et al., 2017; Beg et al., 2022). It is important to mention that other endocrine disruptors who are agonists of the PPARy pathway (such as phthalate metabolites) are well known to affect the differentiation of B lymphocytes in the bone marrow by inducing the apoptosis of B lymphocytes precursors (Schlezinger et al., 2004; Bissonnette et al., 2008; Giuliani et al., 2020).
1.5. Current Gaps in Literature
Many gaps need to be filled in the current knowledge about the effects of TBT on the immune system. For example, its effect on regulatory T cells, cells that are crucial for the maintenance of immune homeostasis through the synthesis and secretion of cytokines such as IL-10, TGF-β and IL-35 (Dominguez-Villar & Hafler., 2018; Shaw et al., 2018). Until this date there is just one study available only in Japanese that demonstrates that, in the presence of TBT, regulatory T lymphocytes are more likely to enter apoptosis than Th2 lymphocytes. (Tada-Oikawa et al., 2010). Moreover, it is still unknown how TBT would impact the functionality of γδ T cells, that are widely present in peripheral tissues, promoting tissue repair and immune surveillance in barrier tissues, through the synthesis and secretion of various molecules such as IFN-γ, IL-17, IL-22, keratinocyte growth factor (KGF), insulin growth factor 1 (IGF1) and fibroblast growth factor 9 (FGF9) (Ribot et al., 2021).
Unfortunately, there are also no studies on the effect of TBT on dendritic cells, even though they are very important for the effectiveness of the immune system, given that they bridge the gap between innate and adaptive immunity by capturing, processing, and presenting antigens to T lymphocytes, mediating their polarization into effector cells (Shortman & Liu, 2002). In the context of other disruptors, it has already been seen that in vitro exposure to a mixture of BPA and BPF (10 - 50μM) led to a decrease in the differentiation and maturation of human monocyte-derived dendritic cells, in addition to the loss of endocytic capacity and suppression of activation of NF-κB and ERK 1/2 pathways (Švajger et al., 2016). Inhibition of NF-κB and ERK pathways was also observed, along with lower expression of interferon-α (IFN-α) and interferon-β (IFN-β), in human plasmacytoid dendritic cells exposed to DEHP. This led to changes in the cytokine secretion profile of CD4+ T cells activated by these dendritic cells, suppressing the production of IFN-γ, but increasing the production of IL-13 (Kuo et al., 2013).
Another relevant point for this discussion is the lack of data about the effects of TBT on the polarization of macrophages into M1 and M2 profiles, as already described for other endocrine disruptors. This knowledge is important because M1 macrophages are mainly involved in the inflammatory response with secretion of cytokines such as IL-1, IL-6, IFN-γ and TNF-α, while M2 macrophages are mostly involved in anti-inflammatory responses producing cytokines such as IL-10 and TGF-β, so an imbalance between the two populations fatally lead to loss of immune homeostasis (Yunna et al., 2020).
On mast cells, there are only two studies involving TBT. The first, from 30 years ago, shows that in vitro exposure of rat serosal mast cells (2,4 × 105 in 0.8mL of medium) to 1mM of TBT for 5 minutes leads to a strong inhibition of histamine secretion from isolated rat mast cells (Iwai et al., 1992). The second only reports an increase in mast cells in cardiac vessels of Wistar rats exposed to 100 ng/Kg of TBT per day by oral gavage, in a study of the impact of TBT on the coronary function (dos Santos et al., 2012). As mast cells are cells present in all (but not only) mucosa of the body and synthesize and secrete various products such as IL-4, IL-6, TGF-β, biogenic amines, growth factors and proteases (Wernersson & Pejler, 2014) it is worrisome not to understand how TBT would affect these cells, as, for example, they may even be contributing to the inflammation in the respiratory system. In fact, there are already enough data to demonstrate that exposure to TBT causes inflammation in the respiratory system (Kato et al., 2004; Kato et al 2006; Kato et al., 2013).
Moreover, to the date of this publication, there is also no data comparing immunological alterations in males and females exposed to TBT. Evidence for divergence in responses between both sexes was already shown for other endocrine disruptors, such as bisphenol (Malaisé et al., 2018) and even for TBT, but in the context of the nervous system (Hernandez et al., 2021).
Conclusions
In this article we summarize what is already known about the effects of TBT on immune system homeostasis. Studies using different animal and human models have shown that exposure to TBT is able to alter the function and viability of immune cells, which may impact immune responses. Changes in the homeostasis of cytokine and chemokine production (summarized in Table 1) were also described in human PBMCs and NK cells (Hurt et al., 2013; Brown & Whalen., 2015; Lawrence et al., 2015; Brown et al., 2018). Furthermore, murine models showed similar alterations in cytokine profile in the serum of TBT-exposed animals, as well as thymic atrophy and changes in cell populations of primary and secondary lymphoid organs. (Yano et al., 2003; Ohtaki et al., 2007; Kato et al., 2004; Kato et al., 2006; Kato et al., 2013; Im et al., 2015; Lawrence et al., 2016). These changes, along with others directly or indirectly induced by TBT, lead to an augmented sensitization of mice to allergens, as seen by the increase in the differentiation of CD4+ T lymphocytes to Th2 profile (Kato et al., 2006), decrease in the lymphocyte population present in secondary lymphoid organs (Ohtaki et al., 2007; Im et al., 2015) and apoptosis of murine thymocytes (Chen et al., 2011; Sharma & Kumar, 2014). Finally, in both human and murine models, TBT directly impacted B cell lymphopoiesis, decreasing the mature B cells (De Santiago & Aguilar-Santelises, 1999; Carfi et al., 2008; Carfi et al., 2010; Baker et al., 2017).
Here, we revised decades of observation and experimentation studies on the effects of TBT on the immune system. We hope that this work brings light not only on what is already known, but also on how scare our knowledge about this topic really is and the urgent need for additional studies. Moreover, we hope to call attention to TBT impacts on the environment and human health, even in doses considered safe by government agencies worldwide.
Author Contributions
Ricardo Correia da Silva: Original Draft, Investigation and Writing. Mariana Pires Cavalcanti Teixeira: Investigation, and Supervision and Writing. Luciana Souza de Paiva: Writing -Review & Editing. Leandro Miranda-Alves: Conceptualization, Investigation, Writing - Review & Editing, Supervision and Funding Acquisition.
Funding
This study was supported by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (CNE-FAPERJ E-26/200.798/2021; Apoio a Grupo Emergentes de Pesquisa do Estado do Rio de Janeiro, E-26/010.002198/2019 & Programa de Apoio a Projetos Temáticos FAPERJ, E-26/211.265/2021), and CNP (PQ2 310654/2021-1). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Acknowledgments
The authors would like to thank Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for financial support.
Conflicts of Interest
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of this manuscript.
References
- Alcala, A.; Osborne, B.; Allen, B.; Seaton-Terry, A.; Kirkland, T.; Whalen, M. Toll-like receptors in the mechanism of tributyltin-induced production of pro-inflammatory cytokines, IL-1β and IL-6. Toxicology 2022, 472, 153177–153177. [Google Scholar] [CrossRef] [PubMed]
- Alzieu, C. Environmental impact of TBT: the French experience. Sci. Total. Environ. 2000, 258, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Antizar-Ladislao, B. Environmental levels, toxicity and human exposure to tributyltin (TBT)-contaminated marine environment. A review. Environ. Int. 2008, 34, 292–308. [Google Scholar] [CrossRef]
- Baker, A.H.; Wu, T.H.; Bolt, A.M.; Gerstenfeld, L.C.; Mann, K.K.; Schlezinger, J.J. From the Cover: Tributyltin Alters the Bone Marrow Microenvironment and Suppresses B Cell Development. Toxicol. Sci. 2017, 158, 63–75. [Google Scholar] [CrossRef]
- Bano, F.; Mohanty, B. Thyroxine modulation of immune toxicity induced by mixture pesticides mancozeb and fipronil in mice. Life Sci. 2019, 240, 117078. [Google Scholar] [CrossRef] [PubMed]
- Beg, M.A.; Beg, A.; Zargar, U.R.; Sheikh, I.A.; Bajouh, O.S.; Abuzenadah, A.M.; Rehan, M. Organotin Antifouling Compounds and Sex-Steroid Nuclear Receptor Perturbation: Some Structural Insights. Toxics 2022, 11, 25. [Google Scholar] [CrossRef]
- Belizário, J.E.; Brandão, W.; Rossato, C.; Peron, J.P. Thymic and Postthymic Regulation of Naïve CD4+T-Cell Lineage Fates in Humans and Mice Models. Mediat. Inflamm. 2016, 2016, 1–16. [Google Scholar] [CrossRef]
- Bissonnette, S.L.; Teague, J.E.; Sherr, D.H.; Schlezinger, J.J. An Endogenous Prostaglandin Enhances Environmental Phthalate-Induced Apoptosis in Bone Marrow B Cells: Activation of Distinct but Overlapping Pathways. Perspect. Surg. 2008, 181, 1728–1736. [Google Scholar] [CrossRef]
- Borish, L.C.; Steinke, J.W. 2. Cytokines and chemokines. J. Allergy Clin. Immunol. 2003, 111, S460–S475. [Google Scholar] [CrossRef]
- Brown, S.; Boules, M.; Hamza, N.; Wang, X.; Whalen, M. Synthesis of interleukin 1 beta and interleukin 6 in human lymphocytes is stimulated by tributyltin. Arch. Toxicol. 2018, 92, 2573–2586. [Google Scholar] [CrossRef]
- Brown, S.; Wilburn, W.; Martin, T.; Whalen, M. Butyltin compounds alter secretion of interleukin 6 from human immune cells. J. Appl. Toxicol. 2017, 38, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Whalen, M. Tributyltin alters secretion of interleukin 1 beta from human immune cells. J. Appl. Toxicol. 2014, 35, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.-M. Organotin Compounds Act as Inhibitor of Transcriptional Activation with Human Estrogen Receptor. J. Microbiol. Biotechnol. 2012, 22, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, J.V.; Freitas-Lima, L.C.; Freitas, F.F.; Freitas, F.P.; Podratz, P.L.; Magnago, R.P.; Porto, M.L.; Meyrelles, S.S.; Vasquez, E.C.; Brandao, P.A.; et al. Tributyltin chloride induces renal dysfunction by inflammation and oxidative stress in female rats. Toxicol. Lett. 2016, 260, 52–69. [Google Scholar] [CrossRef]
- Croce, L.; Gangemi, D.; Ancona, G.; Liboà, F.; Bendotti, G.; Minelli, L.; Chiovato, L. The cytokine storm and thyroid hormone changes in COVID-19. J. Endocrinol. Investig. 2021, 44, 891–904. [Google Scholar] [CrossRef]
- Carfi’, M.; Croera, C.; Ferrario, D.; Campi, V.; Bowe, G.; Pieters, R.; Gribaldo, L. TBTC induces adipocyte differentiation in human bone marrow long term culture. Toxicology 2008, 249, 11–18. [Google Scholar] [CrossRef]
- Carfi’, M.; Bowe, G.; Pieters, R.; Gribaldo, L. Selective inhibition of B lymphocytes in TBTC-treated human bone marrow long-term culture. Toxicology 2010, 276, 33–40. [Google Scholar] [CrossRef]
- Caseri W. 2014. Initial Organotin Chemistry. Journal of Organometallic Chemistry 751:20-24. [CrossRef]
- Chen, Q.; Zhang, Z.; Zhang, R.; Niu, Y.; Bian, X.; Zhang, Q. Tributyltin chloride-induced immunotoxicity and thymocyte apoptosis are related to abnormal Fas expression. Int. J. Hyg. Environ. Heal. 2011, 214, 145–150. [Google Scholar] [CrossRef]
- Childers, G.M.; Perry, C.A.; Blachut, B.; Martin, N.; Bortner, C.D.; Sieber, S.; Li, J.-L.; Fessler, M.B.; Harry, G.J. Assessing the Association of Mitochondrial Function and Inflammasome Activation in Murine Macrophages Exposed to Select Mitotoxic Tri-Organotin Compounds. Environ. Heal. Perspect. 2021, 129, 47015. [Google Scholar] [CrossRef]
- Csaba, G. Effect of endocrine disruptor phytoestrogens on the immune system: Present and future. Acta Microbiol. et Immunol. Hung. 2018, 65, 1–14. [Google Scholar] [CrossRef]
- Csaba, G. Immunoendocrinology: Faulty hormonal imprinting in the immune system. Acta Microbiol. et Immunol. Hung. 2014, 61, 89–106. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.; Gilkeson, G. Estrogen Receptors in Immunity and Autoimmunity. Clin. Rev. Allergy Immunol. 2010, 40, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Davies AG. 2004. Organotin Chemistry, 2nd Edition Weinheim: Wiley-VCH. ISBN 978-3-527-31023-4.
- De Santiago, A.; Aguilar-Santelises, M. Organotin compounds decrease in vitro survival, proliferation and differentiation of normal human B lymphocytes. Hum. Exp. Toxicol. 1999, 18, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Villar, M.; Hafler, D.A. Regulatory T cells in autoimmune disease. Nat. Immunol. 2018, 19, 665–673. [Google Scholar] [CrossRef]
- dos Santos, R.L.; Podratz, P.L.; Sena, G.C.; Filho, V.S.D.; Lopes, P.F.I.; Gonçalves, W.L.S.; Alves, L.M.; Samoto, V.Y.; Takiya, C.M.; Miguel, E.d.C.; et al. Tributyltin Impairs the Coronary Vasodilation Induced by 17β-Estradiol in Isolated Rat Heart. J. Toxicol. Environ. Heal. Part A 2012, 75, 948–959. [Google Scholar] [CrossRef]
- Dudimah, F.D.; Odman-Ghazi, S.O.; Hatcher, F.; Whalen, M.M. Effect of tributyltin (TBT) on ATP levels in human natural killer (NK) cells: Relationship to TBT-induced decreases in NK function. J. Appl. Toxicol. 2006, 27, 86–94. [Google Scholar] [CrossRef]
- Fan, Z.; Che, H.; Yang, S.; Chen, C. Estrogen and estrogen receptor signaling promotes allergic immune responses: Effects on immune cells, cytokines, and inflammatory factors involved in allergy. Allergol. et Immunopathol. 2019, 47, 506–512. [Google Scholar] [CrossRef]
- Fang, D.-A.; Zhao, C.-S.; Jiang, S.-L.; Zhou, Y.-F.; Xu, D.-P. Toxic function of CD28 involving in the TLR/MyD88 signal pathway in the river pufferfish (Takifugu obscurus) after exposed to tributyltin chloride (TBT-Cl). Gene 2019, 688, 84–92. [Google Scholar] [CrossRef]
- Feng, P.; Jyotaki, M.; Kim, A.; Chai, J.; Simon, N.; Zhou, M.; Bachmanov, A.A.; Huang, L.; Wang, H. Regulation of bitter taste responses by tumor necrosis factor. Brain, Behav. Immun. 2015, 49, 32–42. [Google Scholar] [CrossRef]
- Frouin, H.; Lebeuf, M.; Saint-Louis, R.; Hammill, M.; Pelletier. ; Fournier, M. Toxic effects of tributyltin and its metabolites on harbour seal (Phoca vitulina) immune cells in vitro. Aquat. Toxicol. 2008, 90, 243–251. [Google Scholar] [CrossRef]
- Gabay, C.; Lamacchia, C.; Palmer, G. IL-1 pathways in inflammation and human diseases. Nat. Rev. Rheumatol. 2010, 6, 232–241. [Google Scholar] [CrossRef]
- Galdiero, M.R.; Garlanda, C.; Jaillon, S.; Marone, G.; Mantovani, A. Tumor associated macrophages and neutrophils in tumor progression. J. Cell. Physiol. 2013, 228, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, P.E.; Bryan, G.W.; Pascoe, P.L.; Burt, G.R. The use of the dog-whelk, Nucella lapillus, as an indicator of tributyltin (TBT) contamination. J. Mar. Biol. Assoc. United Kingd. 1987, 67, 507–523. [Google Scholar] [CrossRef]
- Giuliani, A.; Zuccarini, M.; Cichelli, A.; Khan, H.; Reale, M. Critical Review on the Presence of Phthalates in Food and Evidence of Their Biological Impact. Int. J. Environ. Res. Public Heal. 2020, 17, 5655. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.J.; Morin, S.M.; Kubosiak, A.; Ser-Dolansky, J.; Schalet, B.J.; Jerry, D.J.; Schneider, S.S. The use of patient-derived breast tissue explants to study macrophage polarization and the effects of environmental chemical exposure. Immunol. Cell Biol. 2020, 98, 883–896. [Google Scholar] [CrossRef]
- Gutcher, I.; Becher, B. APC-derived cytokines and T cell polarization in autoimmune inflammation. J. Clin. Investig. 2007, 117, 1119–1127. [Google Scholar] [CrossRef]
- Hamer, K.; Karius, V. Tributyltin release from harbour sediments—Modelling the influence of sedimentation, bio-irrigation and diffusion using data from Bremerhaven. Mar. Pollut. Bull. 2005, 50, 980–992. [Google Scholar] [CrossRef]
- Hartwell, S.I.; Apeti, D.A.; Mason, A.L.; Pait, A.S. An assessment of butyltins and metals in sediment cores from the St. Thomas East End Reserves, USVI. Environ. Monit. Assess. 2016, 188, 1–11. [Google Scholar] [CrossRef]
- Scudder, M.E.H.; Young, R.L.; Thompson, L.M.; Kore, P.; Crews, D.; A Hofmann, H.; Gore, A.C. EDCs Reorganize Brain-Behavior Phenotypic Relationships in Rats. J. Endocr. Soc. 2021, 5, bvab021. [Google Scholar] [CrossRef]
- Huang, F.-M.; Chang, Y.-C.; Lee, S.-S.; Yang, M.-L.; Kuan, Y.-H. Expression of pro-inflammatory cytokines and mediators induced by Bisphenol A via ERK-NFκB and JAK1/2-STAT3 pathways in macrophages. Environ. Toxicol. 2019, 34, 486–494. [Google Scholar] [CrossRef]
- Hurt, K.; Hurd-Brown, T.; Whalen, M. Tributyltin and dibutyltin alter secretion of tumor necrosis factor alpha from human natural killer cells and a mixture of T cells and natural killer cells. J. Appl. Toxicol. 2012, 33, 503–10. [Google Scholar] [CrossRef]
- Im, E.; Kim, H.; Kim, J.; Lee, H.; Yang, H. Tributyltin acetate-induced immunotoxicity is related to inhibition of T cell development in the mouse thymus. Mol. Cell. Toxicol. 2015, 11, 231–239. [Google Scholar] [CrossRef]
- IMO. 2005. International Marine Organization. Antifouling Systems. International Convention on the Control of Harmful Anti-fouling Systems on Ships. International Maritime Organization. London.
- Iwai, H.; Kurosawa, M.; Matsui, H.; Wada, O. Inhibitory Effects of Organotin Compounds on Histamine Release from Rat Serosal Mast Cells. Ind. Heal. 1992, 30, 77–84. [Google Scholar] [CrossRef]
- Kannan, K.; Senthilkumar, K.; Loganathan, B.G.; Takahashi, S.; Odell, D.K.; Tanabe, S. Elevated Accumulation of Tributyltin and Its Breakdown Products in Bottlenose Dolphins (Tursiops truncatus) Found Stranded along the U.S. Atlantic and Gulf Coasts. Environ. Sci. Technol. 1996, 31, 296–301. [Google Scholar] [CrossRef]
- Kato, T.; Uchikawa, R.; Yamada, M.; Arizono, N.; Oikawa, S.; Kawanishi, S.; Nishio, A.; Nakase, H.; Kuribayashi, K. Environmental pollutant tributyltin promotes Th2 polarization and exacerbates airway inflammation. Eur. J. Immunol. 2004, 34, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Tada-Oikawa, S.; Takahashi, K.; Saito, K.; Wang, L.; Nishio, A.; Hakamada-Taguchi, R.; Kawanishi, S.; Kuribayashi, K. Endocrine disruptors that deplete glutathione levels in APC promote Th2 polarization in mice leading to the exacerbation of airway inflammation. Eur. J. Immunol. 2006, 36, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Tada-Oikawa, S.; Wang, L.; Murata, M.; Kuribayashi, K. Endocrine disruptors found in food contaminants enhance allergic sensitization through an oxidative stress that promotes the development of allergic airway inflammation. Toxicol. Appl. Pharmacol. 2013, 273, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, H.S.; Piao, Y.J.; Moon, W.K. Bisphenol A Promotes the Invasive and Metastatic Potential of Ductal Carcinoma In Situ and Protumorigenic Polarization of Macrophages. Toxicol. Sci. 2019, 170, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Kotrikla, A. Environmental management aspects for TBT antifouling wastes from the shipyards. J. Environ. Manag. 2009, 90, S77–S85. [Google Scholar] [CrossRef]
- Koyasu, S.; Moro, K. Role of Innate Lymphocytes in Infection and Inflammation. Front. Immunol. 2012, 3, 101. [Google Scholar] [CrossRef]
- Kuo, C.-H.; Hsieh, C.-C.; Kuo, H.-F.; Huang, M.-Y.; Yang, S.-N.; Chen, L.-C.; Huang, S.-K.; Hung, C.-H. Phthalates suppress type I interferon in human plasmacytoid dendritic cells via epigenetic regulation. Allergy 2013, 68, 870–879. [Google Scholar] [CrossRef]
- Jang, J.-W.; Lee, J.-W.; Yoon, Y.D.; Kang, J.-S.; Moon, E.-Y. Bisphenol A and its substitutes regulate human B cell survival via Nrf2 expression. Environ. Pollut. 2020, 259, 113907. [Google Scholar] [CrossRef]
- Jie, J.; Ling, L.; Yi, Y.; Tao, L.; Liao, X.; Gao, P.; Xu, Q.; Zhang, W.; Chen, Y.; Zhang, J.; et al. Tributyltin triggers lipogenesis in macrophages via modifying PPARγ pathway. Environ. Pollut. 2020, 271, 116331. [Google Scholar] [CrossRef]
- Lardone, P.J.; Guerrero, J.M.; Fernández-Santos, J.M.; Rubio, A.; Martín-Lacave, I.; Carrillo-Vico, A. Melatonin synthesized by T lymphocytes as a ligand of the retinoic acid-related orphan receptor. J. Pineal Res. 2011, 51, 454–462. [Google Scholar] [CrossRef]
- Lavastre, V.; Girard, D. TRIBUTYLTIN INDUCES HUMAN NEUTROPHIL APOPTOSIS AND SELECTIVE DEGRADATION OF CYTOSKELETAL PROTEINS BY CASPASES. J. Toxicol. Environ. Heal. Part A 2002, 65, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, S.; Reid, J.; Whalen, M. Secretion of interferon gamma from human immune cells is altered by exposure to tributyltin and dibutyltin. Environ. Toxicol. 2013, 30, 559–571. [Google Scholar] [CrossRef]
- Lawrence, S.; Pellom, S.T.; Shanker, A.; Whalen, M.M. Tributyltin exposure alters cytokine levels in mouse serum. J. Immunotoxicol. 2016, 13, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-W.; Park, S.; Han, H.-K.; Gye, M.C.; Moon, E.-Y. Di-(2-ethylhexyl) phthalate enhances melanoma tumor growth via differential effect on M1-and M2-polarized macrophages in mouse model. Environ. Pollut. 2018, 233, 833–843. [Google Scholar] [CrossRef]
- Li, Z.-H.; Li, P. Effects of the tributyltin on the blood parameters, immune responses and thyroid hormone system in zebrafish. Environ. Pollut. 2020, 268, 115707. [Google Scholar] [CrossRef]
- Ling, L.; Wen, J.; Tao, L.; Zhao, M.; Ge, W.; Wang, L.; Zhang, J.; Weng, D. RIP1 and RIP3 contribute to Tributyltin-induced toxicity in vitro and in vivo. Chemosphere 2018, 218, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Maguire, R.J. Environmental aspects of tributyltin. Appl. Organomet. Chem. 1987, 1, 475–498. [Google Scholar] [CrossRef]
- Malaisé, Y.; Lencina, C.; Cartier, C.; Olier, M.; Ménard, S.; Guzylack-Piriou, L. Bisphenol A, S or F mother’s dermal impregnation impairs offspring immune responses in a dose and sex-specific manner in mice. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef]
- Merlo, E.; Podratz, P.L.; Sena, G.C.; de Araújo, J.F.P.; Lima, L.C.F.; Alves, I.S.S.; Gama-De-Souza, L.N.; Pelição, R.; Rodrigues, L.C.M.; Brandão, P.A.A.; et al. The Environmental Pollutant Tributyltin Chloride Disrupts the Hypothalamic-Pituitary-Adrenal Axis at Different Levels in Female Rats. Endocrinology 2016, 157, 2978–2995. [Google Scholar] [CrossRef]
- Moss, R.B.; Moll, T.; El-Kalay, M.; Kohne, C.; Hoo, W.S.; Encinas, J.; Carlo, D.J. Th1/Th2 cells in inflammatory disease states: therapeutic implications. Expert Opin. Biol. Ther. 2004, 4, 1887–1896. [Google Scholar] [CrossRef]
- Murata, S.; Takahashi, S.; Agusa, T.; Thomas, N.J.; Kannan, K.; Tanabe, S. Contamination status and accumulation profiles of organotins in sea otters (Enhydra lutris) found dead along the coasts of California, Washington, Alaska (USA), and Kamchatka (Russia). Mar. Pollut. Bull. 2008, 56, 641–649. [Google Scholar] [CrossRef]
- Nagata, K.; Nishiyama, C. IL-10 in Mast Cell-Mediated Immune Responses: Anti-Inflammatory and Proinflammatory Roles. Int. J. Mol. Sci. 2021, 22, 4972. [Google Scholar] [CrossRef]
- Nakano, K.; Tsunoda, M.; Konno, N. Tributyltin (TBT) increases TNFα mRNA expression and induces apoptosis in the murine macrophage cell line in vitro. Environ. Heal. Prev. Med. 2004, 9, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Nowak, K.; Jabłońska, E.; Ratajczak-Wrona, W. Immunomodulatory effects of synthetic endocrine disrupting chemicals on the development and functions of human immune cells. Environ. Int. 2019, 125, 350–364. [Google Scholar] [CrossRef]
- Nunes-Silva, A.; Dittz, D.; Santana, H.S.; Faria, R.A.; Freitas, K.M.; Coutinho, C.R.; Rodrigues, L.C.d.M.; Miranda-Alves, L.; Silva, I.V.; Graceli, J.B.; et al. The Pollutant Organotins Leads to Respiratory Disease by Inflammation: A Mini-Review. Front. Endocrinol. 2018, 8, 369. [Google Scholar] [CrossRef] [PubMed]
- Ohtaki, K.; Aihara, M.; Takahashi, H.; Fujita, H.; Takahashi, K.; Funabashi, T.; Hirasawa, T.; Ikezawa, Z. Effects of tributyltin on the emotional behavior of C57BL/6 mice and the development of atopic dermatitis-like lesions in DS-Nh mice. J. Dermatol. Sci. 2007, 47, 209–216. [Google Scholar] [CrossRef]
- Palacios-Arreola, M.I.; Moreno-Mendoza, N.A.; Nava-Castro, K.E.; Segovia-Mendoza, M.; Perez-Torres, A.; Garay-Canales, C.A.; Morales-Montor, J. The Endocrine Disruptor Compound Bisphenol-A (BPA) Regulates the Intra-Tumoral Immune Microenvironment and Increases Lung Metastasis in an Experimental Model of Breast Cancer. Int. J. Mol. Sci. 2022, 23, 2523. [Google Scholar] [CrossRef] [PubMed]
- Penninks, A.H. The evaluation of data-derived safety factors for bis(tri-n-butyltin)oxide. Food Addit. Contam. 1993, 10, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Phiel, K.L.; Henderson, R.A.; Adelman, S.J.; Elloso, M.M. Differential estrogen receptor gene expression in human peripheral blood mononuclear cell populations. Immunol. Lett. 2005, 97, 107–113. [Google Scholar] [CrossRef]
- Ponce-Regalado, M.D.; Pérez-Sánchez, G.; Rojas-Espinosa, O.; Arce-Paredes, P.; Girón-Peréz, M.I.; Pavón-Romero, L.; Becerril-Villanueva, E. NeuroImmunoEndocrinology: A brief historic narrative. J. Leukoc. Biol. 2022, 112, 97–114. [Google Scholar] [CrossRef]
- Rajendiran, A.; Tenbrock, K. Regulatory T cell function in autoimmune disease. J. Transl. Autoimmun. 2021, 4, 100130. [Google Scholar] [CrossRef] [PubMed]
- Ribot, J.C.; Lopes, N.; Silva-Santos, B. γδ T cells in tissue physiology and surveillance. Nat. Rev. Immunol. 2020, 21, 221–232. [Google Scholar] [CrossRef]
- Schwaiger J, Falk HF, Bucher F, Orthuber G, Hoffmann R, Negele RD. 1994. Prolonged exposure of rainbow trout (Oncorhynchus mykiss) to sublethal concentrations of bis(tri-n-butyltin) oxide: effects on leukocytes, lymphatic tissues and phagocytosis activity. In: Muller R, Lloyd R (eds) Sublethal and chronic effects of pollutants on freshwater fish. Blackwell Scientific, Cambridge, MA, pp 113.
- Sakazaki, H.; Ueno, H.; Nakamuro, K. Estrogen receptor α in mouse splenic lymphocytes: possible involvement in immunity. Toxicol. Lett. 2002, 133, 221–229. [Google Scholar] [CrossRef]
- Schlezinger, J.J.; Howard, G.J.; Hurst, C.H.; Emberley, J.K.; Waxman, D.J.; Webster, T.; Sherr, D.H. Environmental and Endogenous Peroxisome Proliferator-Activated Receptor γ Agonists Induce Bone Marrow B Cell Growth Arrest and Apoptosis: Interactions between Mono(2-ethylhexyl)phthalate, 9-cis-Retinoic Acid, and 15-Deoxy-Δ12,14-prostaglandin J2. PEDIATRICS 2004, 173, 3165–3177. [Google Scholar] [CrossRef]
- Scudder, M.E.H.; Young, R.L.; Thompson, L.M.; Kore, P.; Crews, D.; A Hofmann, H.; Gore, A.C. EDCs Reorganize Brain-Behavior Phenotypic Relationships in Rats. J. Endocr. Soc. 2021, 5, bvab021. [Google Scholar] [CrossRef]
- Shaibi, T.; Balog, H.; Alghazeer, R.; Othman, M.; Benjama, A.; Elhensheri, M.; Lwaleed, B.; Griw, M. Exposure to low-dose bisphenol A induces spleen damage in a murine model: Potentially through oxidative stress? Open Veter- J. 2022, 12, 23–32. [Google Scholar] [CrossRef]
- Sharma, N.; Kumar, A. Mechanism of immunotoxicological effects of tributyltin chloride on murine thymocytes. Cell Biol. Toxicol. 2014, 30, 101–112. [Google Scholar] [CrossRef]
- Shaw, D.M.; Merien, F.; Braakhuis, A.; Dulson, D. T-cells and their cytokine production: The anti-inflammatory and immunosuppressive effects of strenuous exercise. Cytokine 2018, 104, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, Y.; Kitano, T.; Oshima, Y.; Inoue, S.; Imada, N.; Honjo, T. Tributyltin causes masculinization in fish. Environ. Toxicol. Chem. 2003, 22, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Shortman, K.; Liu, Y.-J. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2002, 2, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Švajger, U.; Dolenc, M.S.; Jeras, M. In vitro impact of bisphenols BPA, BPF, BPAF and 17β-estradiol (E2) on human monocyte-derived dendritic cell generation, maturation and function. Int. Immunopharmacol. 2016, 34, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Tada-Oikawa, S.; Murata, M.; Kato, T. Preferential Induction of Apoptosis in Regulatory T Cells by Tributyltin: Possible Involvement in the Exacerbation of Allergic Diseases. Nippon. Eiseigaku Zasshi (Japanese J. Hyg. 2010, 65, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Tanabe, S.; Takeuchi, I.; Miyazaki, N. Distribution and Specific Bioaccumulation of Butyltin Compounds in a Marine Ecosystem. Arch. Environ. Contam. Toxicol. 1999, 37, 50–61. [Google Scholar] [CrossRef]
- Tanabe S, Prudente M, Mizuno T, Hasegawa J, Iwata H, Miyazaki N. 1998. Butyltin contamination in marine mammals from North Pacific and Asian Coastal waters. Environ Sci Tech. 32:193–8. [CrossRef]
- Titley-O'Neal, C.P.; Munkittrick, K.R.; MacDonald, B.A. The effects of organotin on female gastropods. J. Environ. Monit. 2011, 13, 2360–2388. [Google Scholar] [CrossRef]
- Thomas, L.D.; Shah, H.; A Green, S.; Bankhurst, A.D.; Whalen, M.M. Tributyltin exposure causes decreased granzyme B and perforin levels in human natural killer cells. Toxicology 2004, 200, 221–233. [Google Scholar] [CrossRef]
- Tramontana, F.; Battisti, S.; Napoli, N.; Strollo, R. Immuno-Endocrinology of COVID-19: The Key Role of Sex Hormones. Front. Endocrinol. 2021, 12, 726696. [Google Scholar] [CrossRef]
- Tsai, C.-K.; Cheng, H.-H.; Hsu, T.-Y.; Wang, J.-Y.; Hung, C.-H.; Tsai, C.-C.; Lai, Y.-J.; Lin, Y.-J.; Huang, H.-C.; Chan, J.Y.H.; et al. Prenatal Exposure to Di-Ethyl Phthalate (DEP) Is Related to Increasing Neonatal IgE Levels and the Altering of the Immune Polarization of Helper-T Cells. Int. J. Environ. Res. Public Heal. 2021, 18, 6364. [Google Scholar] [CrossRef] [PubMed]
- Uc-Peraza, R.G.; Castro. B.; Fillmann, G. An absurd scenario in 2021: Banned TBT-based antifouling products still available on the market. Sci. Total. Environ. 2021, 805, 150377. [Google Scholar] [CrossRef] [PubMed]
- Yebra, D.M.; Kiil, S.; Dam-Johansen, K. Antifouling technology—past, present and future steps towards efficient and environmentally friendly antifouling coatings. Prog. Org. Coatings 2004, 50, 75–104. [Google Scholar] [CrossRef]
- Wang, Q.; Pan, W.; Liu, Y.; Luo, J.; Zhu, D.; Lu, Y.; Feng, X.; Yang, X.; Dittmer, U.; Lu, M.; et al. Hepatitis B Virus-Specific CD8+ T Cells Maintain Functional Exhaustion after Antigen Reexposure in an Acute Activation Immune Environment. Front. Immunol. 2018, 9, 219. [Google Scholar] [CrossRef]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- WHO-IPCS. 1999a. World Health Organization. International Programme on Chemical Safety. Concise international chemical assessment document 14. Tributyltin oxide. http://www.inchem.org/documents/cicads/cicad14.htm.
- Yanagisawa, R.; Koike, E.; Win-Shwe, T.-T.; Takano, H. Effects of Oral Exposure to Low-Dose Bisphenol S on Allergic Asthma in Mice. Int. J. Mol. Sci. 2022, 23, 10790. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Ohno, S.; Nakajima, Y.; Toyoshima, S.; Nakajin, S. Effects of Various Chemicals Including Endocrine Disruptors and Analogs on the Secretion of Th1 and Th2 Cytokines from Anti CD3-Stimulated Mouse Spleen Cells. J. Heal. Sci. 2003, 49, 195–204. [Google Scholar] [CrossRef]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Zambrano-Zaragoza, J.F.; Romo-Martínez, E.J.; Durán-Avelar, M.d.J.; García-Magallanes, N.; Vibanco-Pérez, N. Th17 Cells in Autoimmune and Infectious Diseases. Int. J. Inflamm. 2014, 2014, 1–12. [Google Scholar] [CrossRef]
- Zayas, J.P.; Mamede, J.I. HIV Infection and Spread between Th17 Cells. Viruses 2022, 14, 404. [Google Scholar] [CrossRef]
- Zuo, Z.; Chen, S.; Wu, T.; Zhang, J.; Su, Y.; Chen, Y.; Wang, C. Tributyltin causes obesity and hepatic steatosis in male mice. Environ. Toxicol. 2011, 26, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-N.; Zhang, J.-L.; Ren, H.-T.; Zhou, B.-H.; Wu, Q.-J.; Sun, P. Effect of tributyltin on antioxidant ability and immune responses of zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2017, 138, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-S.; Fang, D.-A.; Xu, D.-P. Toll-like receptors (TLRs) respond to tributyltin chloride (TBT-Cl) exposure in the river pufferfish (Takifugu obscurus): Evidences for its toxic injury function. Fish Shellfish. Immunol. 2020, 99, 526–534. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Summary of changes in cytokine/chemokine synthesis and/or secretion caused by TBT treatment.
Table 1.
Summary of changes in cytokine/chemokine synthesis and/or secretion caused by TBT treatment.
| Cytokine/Chemokine | Experimental Model | Effect of TBT |
|---|---|---|
| IL-1β | Human/in vitro - zebrafish |
|
| IL-2 | BALB/c |
|
| IL-4 | C57BL/6 & CBA/J mice spleen cells |
|
| IL-5 | C57BL/6 |
|
| IL-6 | Human/in vitro - zebrafish |
|
| IL-7 | ICR mice |
|
| IL-10 | C57BL/6 |
|
| IL-12 | C57BL/6 |
|
| IL-13 | BALB/c |
|
| IFN-γ | BALB/c, CBA/J mice spleen cells & Human/in vitro |
|
| TNF-α | BALB/c/J774.1 cell line & Human/in vitro - zebrafish |
|
| MIP-1β | BALB/c |
|
| RANTES | BALB/c |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



