
Submitted:
20 July 2023
Posted:
21 July 2023
You are already at the latest version
Abstract
Sodium butyrate (NaB) is one of the short-chain fatty acids and is notably produced in large amounts from dietary fiber in the gut. Recent evidence suggests that NaB induces cell proliferation and apoptosis. The skeletal muscle is rich in plenty of mitochondrial. However, it is unclear how NaB acts on host muscle cells and whether it is involved in mitochondria-related functions in myocytes. The present study aimed to investigate the role of NaB treatment on the proliferation, apoptosis, and mitophagy of bovine skeletal muscle satellite cells (BSCs). The results showed that NaB inhibited the proliferation and promoted apoptosis of BSCs, and promoted mitophagy in a time and dose-dependent manner in BSCs. In addition, 1mM NaB increased the mitochondrial ROS level, decreased the mitochondrial membrane potential (MMP), increased the number of autophagic vesicles in mitochondria, and increased the mitochondrial DNA(mtDNA) and ATP level. The effects of the mTOR pathway on BSCs were investigated, and the results showed that 1mM NaB inhibited the mRNA and protein expression of mTOR and genes AKT1, FOXO1, and EIF4EBP1 in the mTOR signaling pathway, while the addition of PP242, an inhibitor of the mTOR signaling pathway, also inhibited mRNA and protein expression levels of mTOR, AKT1, FOXO1, and EIF4EBP1, and promoted mitophagy and apoptosis, which were consistent with the effect of NaB treatment. In conclusion, NaB might promote mitophagy and apoptosis in BSCs by inhibiting the mTOR signaling pathway. Our results would expand the knowledge of sodium butyrate on bovine skeletal muscle cell state and mitochondrial function.
Keywords:
bovine skeletal muscle satellite cells
; sodium butyrate
; mitophagy
; apoptosis
; mTOR signaling pathway
1. Introduction
Sodium butyrate(NaB) is the sodium salt of short-chain fatty acids, a byproduct of dietary fiber produced during microbial fermentation in the gastrointestinal tract; NaB can cause cell cycle arrest[1], inhibit the growth of cancer cells[2], regulate energy metabolism and mitochondrial function[3], inhibit angiogenesis, immune response and oxidative stress, activate apoptosis, and even regulate the expression of non-coding RNAs in malignant cells[4]; Study showed that, compared with the control group, NaB could inhibit the viability and significantly enhance the apoptosis of Hela cells in a time- and dose-dependent manner[5]; NaB also inhibits proliferation, stimulates apoptosis, and promotes differentiation of human colon cancer cells along the absorptive phenotype[6]. For cattle, NaB induces profound changes in gene expression in bovine kidney epithelial cells, which were associated with cell cycle control [7,8], regulating chromatin states changes in bovine rumen epithelial primary cells[9,10], promotes milk fat synthesis in bovine mammary epithelial cell[11], and bovine immune and inflammatory responses[12], showing the pleiotropic effects of NaB on biological processes in host cells.
Mitochondria are key organelles for energy production and cellular sources of active substances and play an important role in maintaining energy homeostasis, cell growth and development, and intracellular calcium homeostasis in the organism. Skeletal muscle accounts for approximately 40% of the total body weight of mammal animals[13] and is the main organ for maintaining the energy metabolism and locomotion of the organism, as well as the main site of glucose uptake. Thus, skeletal muscle plays an important role in the glucose metabolism of the organism, which means that intracellular mitochondria in muscle cells are essential for maintaining muscle function[14]. Reactive oxygen species(ROS) are oxidants that cause peroxidation of unsaturated fatty acids in lipids, and the organism producing plenty of ROS will damage intracellular lipids, proteins, and mitochondrial DNA(mtDNA), and changes in mtDNA levels will lead to mitochondrial dysfunction and reduced biosynthesis, which in turn will affect cell growth and metabolism[15]. The mammalian target of rapamycin(mTOR) is a member of the proteins family related to a phosphatidylinositol kinase-related kinase(PIKK), which can integrate various extracellular signals such as nutrition, energy, and cytokines. mTOR is involved in transcription, protein translation, ribosomal synthesis, and other processes, and therefore plays an important role in cell growth, apoptosis, and the metabolic process of the organism[16,17]; when the mTOR signaling pathway was inhibited, the cell metabolism reacts abnormally, resulted in slower cell growth and faster catabolism, which in turn promotes apoptosis and mitophagy[18]. Studies have shown that the addition of NaB can induce mitochondrial function by promoting energy expenditure in mice[19]; when mitochondria were damaged, overproduction of NADH and deficiency of NAD+ lead to an imbalance of redox status, enhanced oxidative stress, decreased mitochondrial membrane potential(MMP), increased ROS production and abnormal ATP synthesis[20]. Butyrate treatment could alleviate oxidative stress by regulating NRF2 nuclear accumulation and histone acetylation via GPR109A in bovine mammary epithelial cells[21]. It has been reported that NaB regulates breast cancer cell growth and induces apoptosis by activating the activity of caspase3 and caspase8, increasing intracellular ROS levels, decreasing MMP, and inducing cell cycle arrested[22]. The mechanism of NaB-induced apoptosis of human colon cancer cells was that NaB acts on mitochondria, causes changes in mitochondrial MMP and caspase cascade activation, dissolves proteins in cells or stimulates caspase to activate DNA enzyme to cause degradation of nuclear DNA, and finally leads to apoptosis[23].
Skeletal muscle is an important organ of energy metabolism and contains large numbers of mitochondria, NaB has multiple roles in regulating cell state and gene expression of tissue from the host cell. However, it is unclear how NaB is involved in muscle cell-related activities and whether it affects mitochondria in myocytes. Therefore, this experiment was conducted to investigate the effects of NaB on apoptosis, mitophagy, and regulatory function of host skeletal muscle cells. This study will expand the knowledge of NaB and increase the understanding of the biological role of NaB on mitochondrial function and energy utilization of skeletal muscle cells in livestock.
2. Results
2.1. NaB inhibits BSCs proliferation and promotes apoptosis
The effects of 1 mM NaB on the proliferation of BSCs were detected by RT-qPCR and CCK-8. The results showed that, compared to the control group, the mRNA expression of CCNB2, CDK1, and PCNA were significantly down-regulated after 3h, 6h, and 12h of 1mM NaB treatment (P<0.01), and the inhibitory effect on BSCs proliferation was negatively correlated with time from 0h to 12h (Figure 1a); cell viability was detected by CCK-8 assay kit, the result showed that the viability significantly decreased after treatment with 1mM NaB for 12h, 24h, and 48h(P<0.01) (Figure 1b). For effect of NaB on BSCs apoptosis, the mRNA expression levels of pro-apoptosis genes Bax and p53 were significantly up-regulated at 12h (P<0.01), and the mRNA expression level of anti-apoptotic gene Bcl-2 was down-regulated significantly at 3h-12h after 1mM NaB treatment(P<0.01), indicating 1mM NaB promote apoptosis of BSCs and positively correlating with treating time from 0h to 12h (12h was the most significant ) (Figure 1c). Therefore, it was preliminarily determined that the best condition to treat BSCs with 1mM NaB for 12h. Compared with the control group, after 12h of 1mM NaB treatment that the rate of total apoptotic cells was significantly increased with flow cytometry assay (P<0.01) (Figure 1d); these results indicated that NaB inhibited the proliferation and promoted apoptosis of BSCs in a time-dependent manner.
2.2. NaB promotes mitophagy in BSCs
Studies have demonstrated that NaB induces mitophagy [24]. To clarify the effect of NaB on mitophagy in BSCs, RT-qPCR and WB assays were used to examine the influence of NaB on mitophagy at different concentrations and time. The results showed that, compared to the control group, the mRNA expression levels of pro-mitophagy genes LC3B and PINK1 were significantly up-regulated (P<0.01), and anti-mitophagy gene p62 expression level was significantly down-regulated (P<0.01) at 0.5mM, 1mM, 2mM, and 5mM NaB in BSCs after 12h treatment, and the most significant change was at 1mM NaB concentration (Figure 2a), the protein expression trend was consistent with the corresponding mRNA expression after 0.5mM, 1mM, 2mM, and 5mM NaB treatment at 12h (Figure 2b), this indicated that NaB promoted mitophagy in BSCs, and treatment with 1mM concentration has the most significant effect. To investigate the effect of NaB treatment on mitophagy at different time, BSCs were treated with 1mM NaB for 3h, 6h and 12h, and it was found that the expression levels of LC3B and PINK1 were increased significantly with time(P<0.01), and p62 expression levels was decreased significantly (P<0.01), but the degree of this down-regulation gradually diminishes with treating time and mitophagy was positively correlated with time from 0h to 12h, and the most significant at 12h (Figure 2c). This indicated that NaB promote mitophagy in a time and dose-dependent manner. Based on the above experiments, cells were treated with 1mM NaB for 12h in subsequent experiments.
2.3. NaB induces ROS/MMP response in mitochondria
Mitochondrial ROS and mitophagy were key factors in regulating cellular homeostasis, and decrease in MMP levels was considered one of the hallmarks of mitophagy and apoptosis[25]. Therefore, the study explored the changes in the levels of ROS and MMP after NaB-induced mitophagy. The results showed after 1mM NaB treatment, the proportion of cells with decreased MMP increased significantly (P<0.01)(Figure 3a); ROS levels were detected by ROS assay kit, the result showed that ROS level in the positive control group was higher than that in the negative control group significantly, which was approximately 3.5 times higher, while the ROS level was significantly higher than the blank control after 1mM NaB treatment (P<0.01)(Figure 3b); and laser confocal assay showed that the ROS levels in the blank control and negative control groups were the same, and NaB treatment group was higher than that in the negative control group significantly (Supplemental Figure S1); To elucidate the morphological and structural changes of mitochondria after NaB promotes mitophagy, the ultrastructure of mitochondria was observed using transmission electron microscopy after 1mM NaB treatment cells at 12h, the results showed that compared to the control, NaB treatment increased the number of mitochondrial vacuolization and autophagosomes (Figure 3c); The above experiments showed that NaB induced the decrease of MMP, increased the ROS level and the number of autophagosomes, which further verified the previous molecular results, and suggest that NaB induced mitophagy.
2.4. NaB promotes mtDNA copy number
Because mtDNA replication and transcription levels contribute to mitochondrial biogenesis, energy metabolism, and antioxidant capacity field[26]; Therefore, this study investigated the effect of NaB on mtDNA copy number. The results showed that NaB treatment significantly up-regulated the mRNA expression levels of ND1 and COX1, which were marker genes of mtDNA copy number, indicating an increase in intracellular mitochondria(Figure 4a). The mitochondria were the primary source of ATP production, and a large amount of ATP production accompanied the increase in the number of mitochondria, so the test further verified the changes in intracellular ATP. The result found that the content of intracellular ATP increased significantly after NaB treatment (P<0.01) (Figure 4b), which was consistent with the expected results. These results indicated that 1mM NaB treatment increased the number and activity of intracellular mitochondria, and then might produce a large amount of ATP to maintain the energy metabolism of the organism.
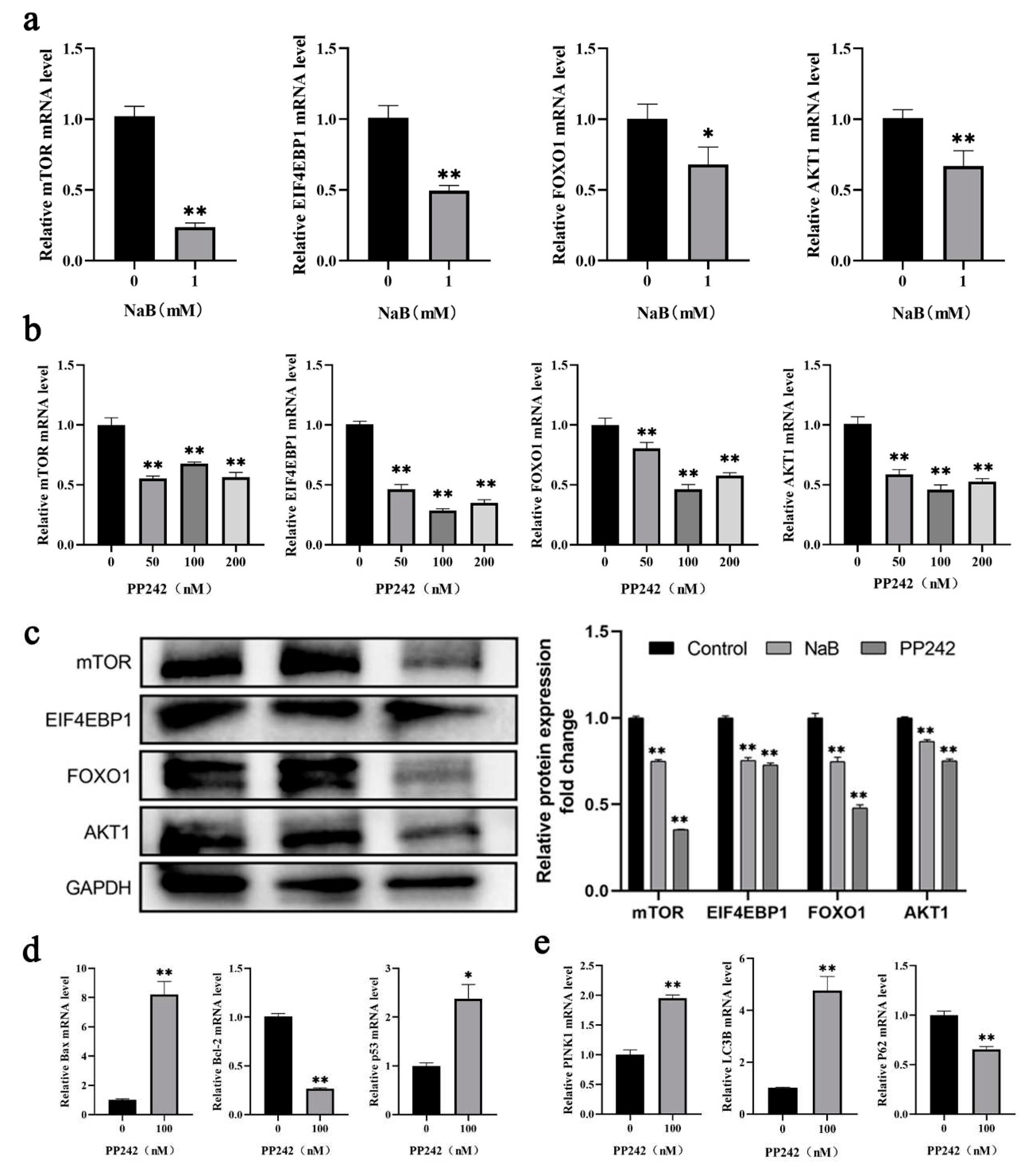
2.5. NaB mediated mTOR signaling pathway promotes mitophagy and apoptosis
Increased mitochondrial ROS levels and MMP deficiency were common phenomena during mitophagy-mediated cell apoptosis[27]. Numerous studies have shown that the mTOR signaling pathway mediated mitophagy and apoptosis in colon cancer[28], ovarian cancer[29], and other cancer cells[30,31]. To investigate the potential mechanisms of NaB on mitophagy and apoptosis in BSCs, this study detected the changes in the mTOR signaling pathway after 1 mM NaB treatment. The results showed that, compared to the control group, the mRNA expression levels of critical genes mTOR, EIF4EBP1, and AKT1 in the mTOR signaling pathway were highly significantly down-regulated (P<0.01), and FOXO1 was significantly down-regulated (P<0.05) after 1mM NaB treatment of BSCs (Figure 5a). PP242 was an effective inhibitor of the mTOR signaling pathway. PP242 inhibitor with different concentrations (50nM, 100nM, 200nM) was used to treat BSCs. It was found that compared to the control group, the mRNA expression levels of mTOR, EIF4EBP1, FOXO1, and AKT1 in the mTOR signaling pathway were significantly down-regulated after treatment with PP242 inhibitors at 50nM, 100nM, 200nM (P<0.01), and most significantly at 100nM (Figure 5b). After treatment of BSCs with 1mM NaB and 100nM PP242 inhibitor respectively, the protein expression levels of mTOR, EIF4EBP1, FOXO1, and AKT1 in the mTOR pathway were consistent with the mRNA levels and were down-regulated significantly (P<0.01)(Figure 5c); while in the 100nM PP242 treatment BSCs, the expression levels of the pro-apoptotic genes Bax and p53 were significantly up-regulated (P<0.01), and the anti-apoptotic genes Bcl-2 were significantly down-regulated (P<0.01) (Figure 5d); the expression levels of pro-mitophagy genes LC3B and PINK1 were significantly up-regulated (P<0.01), and anti-mitophagy p62 was significantly down-regulated (P<0.01) (Figure 5e). This indicated that PP242 could promote mitophagy and apoptosis after BSCs were treated by PP242 inhibitor, which could be speculated that PP242 inhibitor and NaB have the same effect on BSCs regulation. The above results showed that NaB might promote mitophagy and apoptosis in BSCs by inhibiting the mTOR signaling pathway.
3. Discussion
NaB can induce growth, differentiation, and apoptosis in different cancer cell lines, such as NaB inhibits proliferation and induces apoptosis in human breast cancer cells in a dose-dependent manner, thus being an effective inhibitor of breast cancer [32], inhibiting nasopharyngeal carcinoma cells and promotes apoptosis by regulating the progression of the G0/G1 phase of the cell cycle and activating apoptotic factors field[33]. NaB alleviates the adaptive response to inflammation and modulates fatty acid metabolism in lipopolysaccharide-stimulated bovine hepatocytes[34], promoting milk fat synthesis in bovine mammary epithelial cells via GPR41 gene and its downstream signaling pathways[11], reducing the excessive formation of neutrophil extracellular traps by inhibiting autophagy, thus reducing the damaging effect of neutrophil extracellular traps on bovine mammary epithelial cells[35], and affecting rumen epithelial morphology and plasma concentrations of hormones by increasing plasma concentration of glucagon-like peptide-2, lower gene expression of TNF-α, IL-1β, and TLR-2 in the cow’s rumen epithelium cells[36]. NaB induced lipid accumulation and increased expression of peroxisome proliferator-activated receptor gamma (PPARγ) by enriching H3 acetylated K27 (H3K27), thereby affecting growth performance, lipid metabolism, and fatty acid composition in skeletal muscle of offspring piglets[37]; inhibiting platelet-derived growth factor-induced proliferation and migration in pulmonary artery smooth muscle cells through Akt inhibition[38]; NaB could modulated the muscle and adipose tissue metabolism at the transcriptional level by decreasing amino acid metabolism pathways of growing pigs[39]. However, the effect of NaB on the proliferation and apoptosis of BSCs and the mechanism of action are not apparent, so this experiment investigated the effect of 1mM NaB on the proliferation and apoptosis of BSCs. The results showed that adding NaB to the cells significantly inhibited the mRNA expression levels of proliferating marker genes CCNB2, CDK1, CDK2, and PCNA. Bcl-2 is a critical anti-apoptotic gene, and Bax and p53 genes play an activating role in apoptosis. In this study, the expression levels of the Bcl-2 gene were significantly down-regulated, while Bax and p53 were significantly up-regulated after 1mM NaB treatment; the apoptosis rate was detected by flow cytometry, and it was found that the total number of apoptotic cells was significantly increased after 1mM NaB treatment. These results showed that NaB inhibited the proliferation and promoted apoptosis of BSCs. The study investigated NaB inhibited U937 cell growth and apoptosis in a dose-dependent manner, mainly by up-regulation of the expression of the pro-apoptotic gene Bax and down-regulation of the anti-apoptotic gene Bcl-2[40]. For detecting the effect of NaB on the proliferation and apoptosis of osteosarcoma cells and its regulatory mechanism, the results found that the protein expression level of Bcl-2 decreased while Bax increased after NaB treatment. In addition, p53 inhibited proliferation and promoted apoptosis, while MDM2 promoted proliferation and inhibited apoptosis. NaB enhanced p53 expression and decreased MDM2 expression, indicating that NaB promoted apoptosis by regulating the MDM2-p53 signaling pathway[41]. The above studies were consistent with our results, indicating that NaB promoted cell apoptosis.
Cell apoptosis was essential for tissue development, cellular homeostasis, and disease control, and it was usually activated by intrinsic mitochondria-dependent pathways and extrinsic cell apoptotic pathways[42]. This study found that NaB promoted apoptosis in BSCs, but the molecular mechanism causing apoptosis still needs clarification. Therefore, the test further explored the relationship between NaB-induced mitophagy and cell apoptosis. The results showed that the mRNA and protein expression levels of pro-mitophagy genes LC3B and PINK1 were promoted. In contrast, the mRNA and protein expression levels of anti-mitophagy gene p62 were significantly down-regulated after 0mM, 0.5mM, 1mM, and 2mM NaB treatment cells for 3h, 6h, and 12h, which indicated that NaB promoted mitophagy. Mitophagy acts as a protective scavenger to maintain intracellular environmental homeostasis by digesting different products of the organism field[43]. Many studies have shown that mitophagy affects the process of cell apoptotic[22,44], and excessive mitophagy exerts cytotoxic effects and activates the cell apoptotic process[45]. Thus, mitophagy can act as an activator of cell apoptosis. Mitophagy has usually accompanied the production of ROS, damage to MMP, and increased mtDNA, which damage to MMP and increased mtDNA were markers of cell apoptosis. NaB promoted the expression of mitophagy marker proteins LC3 and PINK1. It decreased MMP in hamster ovary cells in a time-and dose-dependent manner while the expression levels of the apoptosis marker proteins PARP, Bcl-2, and caspase-3 gradually increased. The mechanism of NaB regulation of apoptosis in hamster ovary cells was due to the activation of caspase 9 by cytochrome C (Cyt C) in the mitochondrial electron transport chain, and the activation of caspase 9 led to the activity of downstream effectors caspase 3 and Bcl-2, which eventually led to cell apoptosis[46]. In this study, NaB-induced mitophagy was followed by a decrease in MMP, and significant up-regulation of ROS levels, along with a large amount of mtDNA and ATP production by the cells, which tentatively suggested that NaB-induced cell apoptosis was mediated by mitophagy.
In this study, NaB promoted mitophagy and apoptosis in BSCs, but the molecular mechanisms and whether downstream signaling pathways regulated them remain to be explored. mTOR proteins were highly conserved serine/threonine (Ser/Thr) protein kinases belonging to the PIKK protein family. mTOR exists as two protein complexes: mTORC1 and mTORC2, which regulate cellular metabolism, growth, mitophagy, and apoptosis by participating in multiple signaling pathways in vivo[47]. mTOR inhibitors block mTOR signaling pathways and produce anti-inflammatory, anti-proliferative, mitophagy, and apoptosis-inducing effects[48]. Based on the above functions of mTOR, we studied the mTOR signaling pathway using NaB treatment of BSCs. We found that NaB treatment caused significant down-regulation of mRNA and protein expression levels of mTOR, EIF4EBP1, FOXO1, and AKT1 in the mTOR signaling pathway, which indicated that NaB inhibited the mTOR signaling pathway. The mTOR signaling pathway inhibitor PP242 also inhibited the expression levels of mTOR, EIF4EBP1, FOXO and AKT1. In addition, the expression levels of apoptotic genes Bax and p53 were significantly up-regulated, and the expression levels of anti-apoptotic genes Bcl-2 were significantly down-regulated. In contrast, the expression levels of mitophagy genes LC3B and PINK1 were significantly up-regulated. Anti-mitophagy gene p62 expression levels were significantly down-regulated, suggesting that NaB has a synergistic effect with PP242 inhibitor, which promoted mitophagy in BSCs by inhibiting the mTOR signal pathway, thereby inducing cell apoptosis. NaB induces mitophagy-related apoptosis by inhibiting the AKT/mTOR axis in nasopharyngeal carcinoma cells and then exerts anti-tumor effects in nasopharyngeal carcinoma[49]; Research showed that NaB inhibited bladder cancer cell migration and induced AMPK/mTOR pathway-activated mitophagy and ROS overproduction through the miR-139-5p/Bmi-1 axis, and ROS overproduction contributed to NaB-induced cysteine-dependent apoptosis[50]; α-synuclein aggregation is central to the pathogenesis of Parkinson disease, and it was found that in murine neuroendocrine STC-1 cells, NaB leads to α-synaptic nuclear protein degradation and apoptosis through Atg5-dependent and PI3K/Akt/mTOR-related mitophagy pathways[51].
4. Materials and Methods
4.1. Reagents and antibodies
Fetal bovine serum(FBS) and 0.25% Trypsin-EDTA were obtained from Gibco (Gibco, USA); whole protein extraction kit and BCA protein content assay kit were purchased from (KeyGEN Biotechnology, China); CCK-8 assay kit (APExBIO, USA), mTOR pathway inhibitor PP242(APExBIO, USA), ROS assay kit Elabscience (Elabscience Biotechnology, China), sodium butyrate(Sigma-Aldrich, St. Louis, MO, USA), and MMP assay kit was purchased from Beyotime(Shanghai, China); SDS-PAGE gel kit (Solarbio, China), GAPDH antibody(Cambridge, UK), and P62 antibody were obtained from Abcam(Cambridge, UK); mTOR antibody, AKT1 antibody, and FOXO antibody was purchased from Proteintech (Proteintech, China).
4.2. Cell culture
BSCs were isolated using 2% collagenase I. The cells were cultured in DMEM-F12 complete medium supplemented with 20% FBS and antibiotics(1% penicillin-streptomycin), under an incubator of 5% CO2 at 37°C, and the medium was replaced every 48h. When the cells density reached 70%-90%, cells were washed twice with phosphate-buffered saline (PBS)(HyClone, USA) and digested with Pancreatin for 4-5min. The digestion was terminated by adding medium immediately when the cells were observed to become rounded and suspended. The fresh cells were collected after centrifugation for use or frozen and stored in liquid nitrogen for the next experiment.
4.3. Cell viability assay
BSCs were seeded in 96-well plates at a density of 1×104 cells per well. The cells were cultured in the incubator for 0h, 12h, 24h, and 48h, respectively, with a final concentration of 1mM NaB after the cells were attached to the wall and the density reached 20%. Then 10μL CCK-8 reagent was added to each well and incubated for 2h at 37°C, and absorbance at 450nm was measured using a microplate reader (BioTek Instruments, Winooski, VT, USA).
4.4. Cell apoptosis assay
BSCs were inoculated with 1×106 cells per well in 6-well plates. When the density reached 70%, the culture medium was discarded and washed twice with PBS buffer, after which 1mM NaB was added and incubated in a CO2 incubator for 0h, 3h, 6h, and 12h, respectively. The effect of NaB on cell apoptosis was detected by RT-qPCR and flow cytometry (FACSAria II, Becton Dickinson, Franklin Lakes, NJ, USA); The RT-qPCR reaction system consisted of 2×ChamQ Universal SYBR qPCR Master Mix 10 μL, primer F 0.8 μL, primer R 0.8 μL, cDNA template 2 μL, and RNase free ddH2O 6.4 μL. The reaction procedure was as follows: predenaturation at 95°C for 30 s, denaturation at 95°C for 10 s, and annealing extension for 30 s, 40 cycles in total.
4.5. Assay of mitophagy and ROS
Cells were treated with NaB at concentrations of 0.5mM, 1mM, 2mM, and 5mM for 0h, 3h, 6h, and 12h respectively, and total cellular mRNA and total protein were extracted. The effects of NaB on mitophagy genes and proteins at different times and concentrations were detected by RT-qPCR and western blotting (WB). ROS levels were detected by ROS assay kit and laser confocal method, according to the assay kit operation manual.
4.6. MMP assay
MMP was assayed by JC-1 staining. Cells were treated with NaB at 1 mM for 12h, cells were collected and incubated at 37°C for 20min with 5mg/mL JC-1 dye, 500μL PBS was added to wash the cells three times, and placed in a fresh serum-free medium. The level of MMP was detected by flow cytometry.
4.7. Mitophagy vesicles assay
Cells treated with 1mM NaB were incubated for 12h. The medium was discarded, and 2.5% glutaraldehyde fixative was added and fixed at 4℃ for 6h, followed by 1% osmium tetroxide for 2h at room temperature. Samples were dehydrated twice by gradients of 30%, 50%, 70%, 80%, 85%, 90%, 95%, and 100% ethanol for 20min each time. The mitophagy vesicles in mitochondria were observed using transmission electron microscopy by permeabilization, embedding, ultrathin sectioning, and double staining.
4.8. mtDNA copy number
Genomic DNA was extracted from the cells using a commercially available kit following the manufacturer’s instructions, calculated standard curve, and the expression of mtDNA copy number genes ND1 and COX1 in BSCs by 1mM NaB treatment were detected for RT-qPCR, GAPDH was used as an internal reference gene to calculate the relative expression of each gene. The genes sequences were obtained from NCBI, and the primers were designed using primer5.0 software (Premier Bosoft, Palo Alto, CA, USA). The sequences of all primers were listed in the supplementary Table S1.
4.9. ATP content assays
When the cell density reached 70%, the culture medium was discarded, and the cells were washed twice with PBS buffer. 200μL of cell lysate was added to each well, and the cells were repeatedly blown using a pipette to lyse the cells fully. Centrifuge at 4℃, 12, 000g for 5min, the supernatant was taken, and the content of ATP in the whole cell lysate was measured using the ATP assay kit (Beyotime Biotech, Shanghai, China), according to the assay kit operation manual.
4.10. Effect of mTOR signaling pathway on mitophagy and apoptosis
Cells were treated with 1mM NaB for 12h, total cellular mRNA and total protein were extracted, and the mRNA and protein expression levels of mTOR, EIF4EBP1, FOXO1, and AKT1 in the mTOR signaling pathway were detected by RT-qPCR and WB. To further investigate the effect of the mTOR signaling pathway on mitophagy and apoptosis, the mTOR signaling pathway inhibitor PP242 was cultured with final concentrations of 0nM, 50nM, 100nM, and 200nM for 12h, respectively. Total cellular mRNA and total protein were extracted to detect the effect of PP242 inhibitor on the expression of key genes in the mTOR signaling pathway, such as mTOR, EIF4EBP1, FOXO1, and AKT1, as well as the effects on mitophagy and apoptosis.
4.11. Statistical analysis
The experimental data were statistically normalized using the 2-ΔΔCt method and analyzed using the SPSS 25.0 software. Image J software was used to analyze the protein content. One-way ANOVA was used for test analysis, and GraphPad Prism 8.0 software was used for graphing. The results were expressed by mean ± standard error. P<0.05 indicated a significant difference.
5. Conclusions
Present study results demonstrated that NaB subdued the proliferation and increased apoptosis in BSCs. Further, NaB treatment significantly promoted mitophagy and ROS response and decreased the MMP. It was also shown that NaB might regulate apoptosis and mitophagy in BSCs by downregulating the mTOR signaling pathway. The present results indicated that NaB may be a critical metabolic molecule in muscle development, energy metabolism, and regulation of mitochondrial function.
Author Contributions
X.K. and Y.S. conceived and designed the research, Y.D., P.W., C.Y., X.Z., and Z.S. performed experiment; Y.Z., C.L., and X.W. analyzed data; W.M and L.Z prepared figures; CJ.L. and X.K. revised the manuscript; Y.D. organized the data and drafted the manuscript, all authors read and approved the final manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32160776), grant from the China Agriculture Research System (CARS-36), the Natural Science Foundation of Ningxia Province, China (Grant No. 2021AAC03027) and West Light Foundation of the Chinese Academy of Sciences (Grant No. XAB2022YW11).
Institutional Review Board Statement
All experimental animal studies were reviewed and approved by the Animal Welfare Committee of Ningxia University (permit number NXUC20180306).
Acknowledgments
We thank all the cattle breeding staff who helped with sample collecting in this experiment and all members of this research group for all valuable discussions and feedback on this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Falkenberg, K.J.; Johnstone, R.W. Histone deacetylases and their inhibitors in cancer, neurological diseases and immune disorders. Nat Rev Drug Discov 2014, 13, 673–691. [Google Scholar] [CrossRef] [PubMed]
- Fung, K.Y.; Cosgrove, L.; Lockett, T.; Head, R.; Topping, D.L. A review of the potential mechanisms for the lowering of colorectal oncogenesis by butyrate. Br J Nutr 2012, 108, 820–831. [Google Scholar] [CrossRef]
- Tang, X.; Ma, S.; Li, Y.; Sun, Y.; Zhang, K.; Zhou, Q.; Yu, R. Evaluating the Activity of Sodium Butyrate to Prevent Osteoporosis in Rats by Promoting Osteal GSK-3beta/Nrf2 Signaling and Mitochondrial Function. J Agric Food Chem 2020, 68, 6588–6603. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Jiang, W.; Xiao, C.; Yang, W.; Qin, Q.; Mao, A.; Tan, Q.; Lian, B.; Wei, C. Sodium Butyrate Combined with Docetaxel for the Treatment of Lung Adenocarcinoma A549 Cells by Targeting Gli1. Onco Targets Ther 2020, 13, 8861–8875. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Sun, X.; Wang, J.; Lei, K.; Shan, Z.; Zhao, C.; Ning, Y.; Gong, R.; Ren, H.; Cui, Z. Synergistic effects of sodium butyrate and cisplatin against cervical carcinoma in vitro and in vivo. Front Oncol 2022, 12, 999667. [Google Scholar] [CrossRef]
- Augenlicht, L.; Shi, L.; Mariadason, J.; Laboisse, C.; Velcich, A. Repression of MUC2 gene expression by butyrate, a physiological regulator of intestinal cell maturation. Oncogene 2003, 22, 4983–4992. [Google Scholar] [CrossRef]
- Boschiero, C.; Gao, Y.; Baldwin RLt Ma, L.; Li, C.J.; Liu, G.E. Butyrate Induces Modifications of the CTCF-Binding Landscape in Cattle Cells. Biomolecules 2022, 12. [Google Scholar] [CrossRef]
- Li, R.W.; Li, C. Butyrate induces profound changes in gene expression related to multiple signal pathways in bovine kidney epithelial cells. BMC Genomics 2006, 7, 234. [Google Scholar] [CrossRef]
- Fang, L.; Liu, S.; Liu, M.; Kang, X.; Lin, S.; Li, B.; Connor, E.E.; Baldwin RLt Tenesa, A.; Ma, L.; et al. Functional annotation of the cattle genome through systematic discovery and characterization of chromatin states and butyrate-induced variations. BMC Biol 2019, 17, 68. [Google Scholar] [CrossRef]
- Kang, X.; Liu, S.; Fang, L.; Lin, S.; Liu, M.; Baldwin, R.L.; Liu, G.E.; Li, C.J. Data of epigenomic profiling of histone marks and CTCF binding sites in bovine rumen epithelial primary cells before and after butyrate treatment. Data Brief 2020, 28, 104983. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, Y.; Ge, Y.; Li, W.; Cao, Y.; Qu, Y.; Liu, S.; Guo, Y.; Fu, S.; Liu, J. Sodium butyrate promotes milk fat synthesis in bovine mammary epithelial cells via GPR41 and its downstream signalling pathways. Life Sci 2020, 259, 118375. [Google Scholar] [CrossRef]
- Hill, T.M.; Vandehaar, M.J.; Sordillo, L.M.; Catherman, D.R.; Bateman, H.G., 2nd; Schlotterbeck, R.L. Fatty acid intake alters growth and immunity in milk-fed calves. J Dairy Sci 2011, 94, 3936–3948. [Google Scholar] [CrossRef]
- Guller, I.; Russell, A.P. MicroRNAs in skeletal muscle. their role and regulation in development, disease and function. J Physiol 2010, 588, 4075–4087. [Google Scholar] [CrossRef] [PubMed]
- Abate, M.; Festa, A.; Falco, M.; Lombardi, A.; Luce, A.; Grimaldi, A.; Zappavigna, S.; Sperlongano, P.; Irace, C.; Caraglia, M.; et al. Mitochondria as playmakers of apoptosis, autophagy and senescence. Semin Cell Dev Biol 2020, 98, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Kudryavtseva, A.V.; Krasnov, G.S.; Dmitriev, A.A.; Alekseev, B.Y.; Kardymon, O.L.; Sadritdinova, A.F.; Fedorova, M.S.; Pokrovsky, A.V.; Melnikova, N.V.; Kaprin, A.D.; et al. Mitochondrial dysfunction and oxidative stress in aging and cancer. Oncotarget 2016, 7, 44879–44905. [Google Scholar] [CrossRef] [PubMed]
- Holczer, M.; Besze, B.; Zambo, V.; Csala, M.; Banhegyi, G.; Kapuy, O. Epigallocatechin-3-Gallate (EGCG) Promotes Autophagy-Dependent Survival via Influencing the Balance of mTOR-AMPK Pathways upon Endoplasmic Reticulum Stress. Oxid Med Cell Longev 2018, 2018, 6721530. [Google Scholar] [CrossRef] [PubMed]
- Nowosad, A.; Jeannot, P.; Callot, C.; Creff, J.; Perchey, R.T.; Joffre, C.; Codogno, P.; Manenti, S.; Besson, A. Publisher Correction. p27 controls Ragulator and mTOR activity in amino acid-deprived cells to regulate the autophagy-lysosomal pathway and coordinate cell cycle and cell growth. Nat Cell Biol 2021, 23, 1048. [Google Scholar] [CrossRef]
- Yang, H.; Bai, X.; Zhang, H.; Zhang, J.; Wu, Y.; Tang, C.; Liu, Y.; Yang, Y.; Liu, Z.; Jia, W.; et al. Antrodin C, an NADPH Dependent Metabolism, Encourages Crosstalk between Autophagy and Apoptosis in Lung Carcinoma Cells by Use of an AMPK Inhibition-Independent Blockade of the Akt/mTOR Pathway. Molecules 2019, 24. [Google Scholar] [CrossRef]
- Henagan, T.M.; Stefanska, B.; Fang, Z.; Navard, A.M.; Ye, J.; Lenard, N.R.; Devarshi, P.P. Sodium butyrate epigenetically modulates high-fat diet-induced skeletal muscle mitochondrial adaptation, obesity and insulin resistance through nucleosome positioning. Br J Pharmacol 2015, 172, 2782–2798. [Google Scholar] [CrossRef]
- Claflin, D.R.; Jackson, M.J.; Brooks, S.V. Age affects the contraction-induced mitochondrial redox response in skeletal muscle. Front Physiol 2015, 6, 21. [Google Scholar] [CrossRef]
- Guo, W.; Liu, J.; Sun, J.; Gong, Q.; Ma, H.; Kan, X.; Cao, Y.; Wang, J.; Fu, S. Butyrate alleviates oxidative stress by regulating NRF2 nuclear accumulation and H3K9/14 acetylation via GPR109A in bovine mammary epithelial cells and mammary glands. Free Radic Biol Med 2020, 152, 728–742. [Google Scholar] [CrossRef]
- Salimi, V.; Shahsavari, Z.; Safizadeh, B.; Hosseini, A.; Khademian, N.; Tavakoli-Yaraki, M. Sodium butyrate promotes apoptosis in breast cancer cells through reactive oxygen species (ROS) formation and mitochondrial impairment. Lipids Health Dis 2017, 16, 208. [Google Scholar] [CrossRef]
- Heerdt, B.G.; Houston, M.A.; Anthony, G.M.; Augenlicht, L.H. Mitochondrial membrane potential (delta psi(mt)) in the coordination of p53-independent proliferation and apoptosis pathways in human colonic carcinoma cells. Cancer Res 1998, 58, 2869–2875. [Google Scholar]
- Bian, Z.; Zhang, Q.; Qin, Y.; Sun, X.; Liu, L.; Liu, H.; Mao, L.; Yan, Y.; Liao, W.; Zha, L.; et al. Sodium Butyrate Inhibits Oxidative Stress and NF-kappaB/NLRP3 Activation in Dextran Sulfate Sodium Salt-Induced Colitis in Mice with Involvement of the Nrf2 Signaling Pathway and Mitophagy. Dig Dis Sci 2023.
- Jin, M.H.; Yu, J.B.; Sun, H.N.; Jin, Y.H.; Shen, G.N.; Jin, C.H.; Cui, Y.D.; Lee, D.S.; Kim, S.U.; Kim, J.S.; et al. Peroxiredoxin II Maintains the Mitochondrial Membrane Potential against Alcohol-Induced Apoptosis in HT22 Cells. Antioxidants (Basel) 2019, 9. [Google Scholar] [CrossRef]
- Vriens, A.; Nawrot, T.S.; Baeyens, W.; Den Hond, E.; Bruckers, L.; Covaci, A.; Croes, K.; De Craemer, S.; Govarts, E.; Lambrechts, N.; et al. Neonatal exposure to environmental pollutants and placental mitochondrial DNA content. A multi-pollutant approach. Environ Int 2017, 106, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Li, A.X.; Sun, M.; Li, X. Withaferin-A induces apoptosis in osteosarcoma U2OS cell line via generation of ROS and disruption of mitochondrial membrane potential. Eur Rev Med Pharmacol Sci 2017, 21, 1368–1374. [Google Scholar] [PubMed]
- Wang, G.Y.; Zhang, L.; Geng, Y.D.; Wang, B.; Feng, X.J.; Chen, Z.L.; Wei, W.; Jiang, L. beta-Elemene induces apoptosis and autophagy in colorectal cancer cells through regulating the ROS/AMPK/mTOR pathway. Chin J Nat Med 2022, 20, 9–21. [Google Scholar] [PubMed]
- Shan, Y.; Guan, F.; Zhao, X.; Wang, M.; Chen, Y.; Wang, Q.; Feng, X. Macranthoside B Induces Apoptosis and Autophagy Via Reactive Oxygen Species Accumulation in Human Ovarian Cancer A2780 Cells. Nutr Cancer 2016, 68, 280–289. [Google Scholar] [CrossRef]
- Wang, F.; Wang, L.; Qu, C.; Chen, L.; Geng, Y.; Cheng, C.; Yu, S.; Wang, D.; Yang, L.; Meng, Z.; et al. Kaempferol induces ROS-dependent apoptosis in pancreatic cancer cells via TGM2-mediated Akt/mTOR signaling. BMC Cancer 2021, 21, 396. [Google Scholar] [CrossRef]
- Yuan, Y.; He, X.; Li, X.; Liu, Y.; Tang, Y.; Deng, H.; Shi, X. Narciclasine induces autophagy-mediated apoptosis in gastric cancer cells through the Akt/mTOR signaling pathway. BMC Pharmacol Toxicol 2021, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Semaan, J.; El-Hakim, S.; Ibrahim, J.N.; Safi, R.; Elnar, A.A.; El Boustany, C. Comparative effect of sodium butyrate and sodium propionate on proliferation, cell cycle and apoptosis in human breast cancer cells MCF-7. Breast Cancer 2020, 27, 696–705. [Google Scholar] [CrossRef]
- Li, W.F.; Zhang, Y.; Huang, X.B.; Du, X.J.; Tang, L.L.; Chen, L.; Peng, H.; Guo, R.; Sun, Y.; Ma, J. Prognostic value of plasma Epstein-Barr virus DNA level during posttreatment follow-up in the patients with nasopharyngeal carcinoma having undergone intensity-modulated radiotherapy. Chin J Cancer 2017, 36, 87. [Google Scholar] [CrossRef]
- Xu, T.; Ma, N.; Wang, Y.; Shi, X.; Chang, G.; Loor, J.J.; Shen, X. Sodium Butyrate Supplementation Alleviates the Adaptive Response to Inflammation and Modulates Fatty Acid Metabolism in Lipopolysaccharide-Stimulated Bovine Hepatocytes. J Agric Food Chem 2018, 66, 6281–6290. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Cui, Y.; Cao, Y.; Xu, P.; Kan, X.; Guo, W.; Fu, S. Sodium butyrate attenuates bovine mammary epithelial cell injury by inhibiting the formation of neutrophil extracellular traps. Int Immunopharmacol 2022, 110, 109009. [Google Scholar] [CrossRef] [PubMed]
- Fukumori, R.; Doi, K.; Mochizuki, T.; Oikawa, S.; Gondaira, S.; Iwasaki, T.; Izumi, K. Sodium butyrate administration modulates the ruminal villus height, inflammation-related gene expression, and plasma hormones concentration in dry cows fed a high-fiber diet. Anim Sci J 2022, 93, e13791. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, D.; Huang, Y.; Wu, D.; Ji, X.; Zhou, X.; Xia, D.; Yang, X. Maternal butyrate supplementation affects the lipid metabolism and fatty acid composition in the skeletal muscle of offspring piglets. Anim Nutr 2021, 7, 959–966. [Google Scholar] [CrossRef]
- Cantoni, S.; Galletti, M.; Zambelli, F.; Valente, S.; Ponti, F.; Tassinari, R.; Pasquinelli, G.; Galie, N.; Ventura, C. Sodium butyrate inhibits platelet-derived growth factor-induced proliferation and migration in pulmonary artery smooth muscle cells through Akt inhibition. FEBS J 2013, 280, 2042–2055. [Google Scholar] [CrossRef]
- Zhang, H.; Ren, E.; Xu, R.; Su, Y. Transcriptomic Responses Induced in Muscle and Adipose Tissues of Growing Pigs by Intravenous Infusion of Sodium Butyrate. Biology (Basel) 2021, 10. [Google Scholar] [CrossRef]
- Choi, Y.H. Apoptosis of U937 human leukemic cells by sodium butyrate is associated with inhibition of telomerase activity. Int J Oncol 2006, 29, 1207–1213. [Google Scholar] [CrossRef]
- Xie, C.; Wu, B.; Chen, B.; Shi, Q.; Guo, J.; Fan, Z.; Huang, Y. Histone deacetylase inhibitor sodium butyrate suppresses proliferation and promotes apoptosis in osteosarcoma cells by regulation of the MDM2-p53 signaling. Onco Targets Ther 2016, 9, 4005–4013. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Lopez-Soto, A.; Kumar, S.; Kroemer, G. Caspases Connect Cell-Death Signaling to Organismal Homeostasis. Immunity 2016, 44, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.M.M.; Towers, C.G.; Thorburn, A. Targeting autophagy in cancer. Nat Rev Cancer 2017, 17, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Lei, Y.H.; Yao, N.; Wang, C.R.; Hu, N.; Ye, W.C.; Zhang, D.M.; Chen, Z.S. Autophagy and multidrug resistance in cancer. Chin J Cancer 2017, 36, 52. [Google Scholar] [CrossRef]
- Song, S.; Tan, J.; Miao, Y.; Li, M.; Zhang, Q. Crosstalk of autophagy and apoptosis. Involvement of the dual role of autophagy under ER stress. J Cell Physiol 2017, 232, 2977–2984. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, G.M. Effect of sodium butyrate on autophagy and apoptosis in Chinese hamster ovary cells. Biotechnol Prog 2012, 28, 349–357. [Google Scholar]
- Chen, Y.; Zhou, X. Research progress of mTOR inhibitors. Eur J Med Chem 2020, 208, 112820. [Google Scholar] [CrossRef] [PubMed]
- Heras-Sandoval, D.; Perez-Rojas, J.M.; Hernandez-Damian, J.; Pedraza-Chaverri, J. The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration. Cell Signal 2014, 26, 2694–2701. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zeng, C.; Liu, J.; Yuan, L.; Liu, W.; Wang, L.; Zhu, H.; Xu, Y.; Luo, Y.; Xie, D.; et al. Sodium butyrate induces autophagic apoptosis of nasopharyngeal carcinoma cells by inhibiting AKT/mTOR signaling. Biochem Biophys Res Commun 2019, 514, 64–70. [Google Scholar] [CrossRef]
- Wang, F.; Wu, H.; Fan, M.; Yu, R.; Zhang, Y.; Liu, J.; Zhou, X.; Cai, Y.; Huang, S.; Hu, Z.; et al. Sodium butyrate inhibits migration and induces AMPK-mTOR pathway-dependent autophagy and ROS-mediated apoptosis via the miR-139-5p/Bmi-1 axis in human bladder cancer cells. FASEB J 2020, 34, 4266–4282. [Google Scholar] [CrossRef]
- Qiao, C.M.; Sun, M.F.; Jia, X.B.; Shi, Y.; Zhang, B.P.; Zhou, Z.L.; Zhao, L.P.; Cui, C.; Shen, Y.Q. Sodium butyrate causes alpha-synuclein degradation by an Atg5-dependent and PI3K/Akt/mTOR-related autophagy pathway. Exp Cell Res 2020, 387, 111772. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of 1mM NaB on the proliferation and apoptosis of BSCs.(a) The mRNA expression of proliferation genes with 1mM NaB treatment. (b) The effect of 1mM NaB treatment on cell viability. (c) The mRNA expression of apoptosis genes with 1mM NaB treatment. (d) The apoptosis rate of cells with 1mM NaB treatment by flow cytometry. Data were expressed as the mean ± standard deviation of three replicate experiments, *P < 0.05 , **P < 0 .01.
Figure 1.
Effect of 1mM NaB on the proliferation and apoptosis of BSCs.(a) The mRNA expression of proliferation genes with 1mM NaB treatment. (b) The effect of 1mM NaB treatment on cell viability. (c) The mRNA expression of apoptosis genes with 1mM NaB treatment. (d) The apoptosis rate of cells with 1mM NaB treatment by flow cytometry. Data were expressed as the mean ± standard deviation of three replicate experiments, *P < 0.05 , **P < 0 .01.
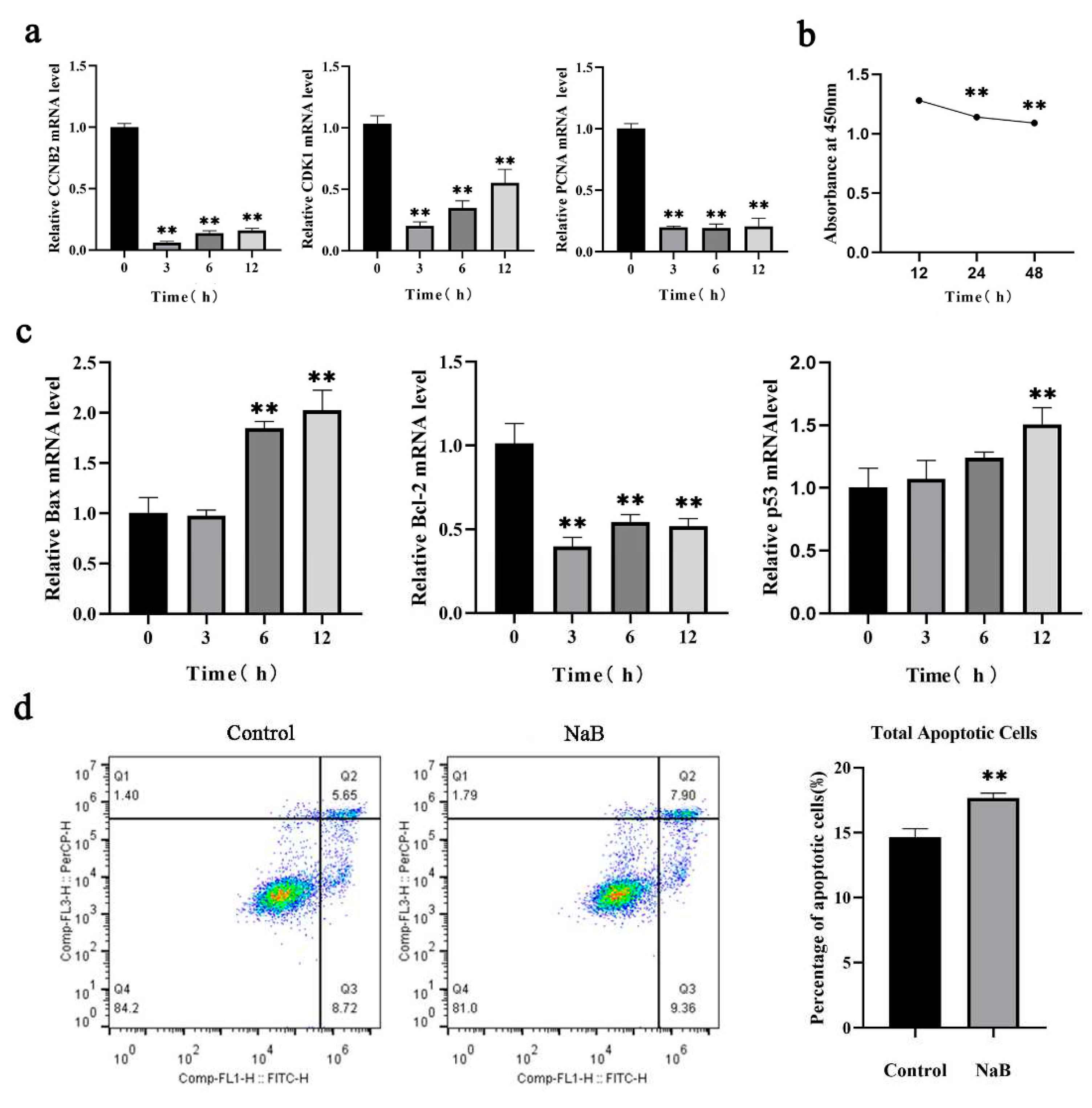
Figure 2.
Effect of NaB on mitophagy in BSCs.(a) Expression of mitophagy genes with different concentrations of NaB treatment. (b) Expression of mitophagy proteins with different concentrations of NaB treatment. (c) Expression of mitophagy genes with different time of NaB treatment.
Figure 2.
Effect of NaB on mitophagy in BSCs.(a) Expression of mitophagy genes with different concentrations of NaB treatment. (b) Expression of mitophagy proteins with different concentrations of NaB treatment. (c) Expression of mitophagy genes with different time of NaB treatment.
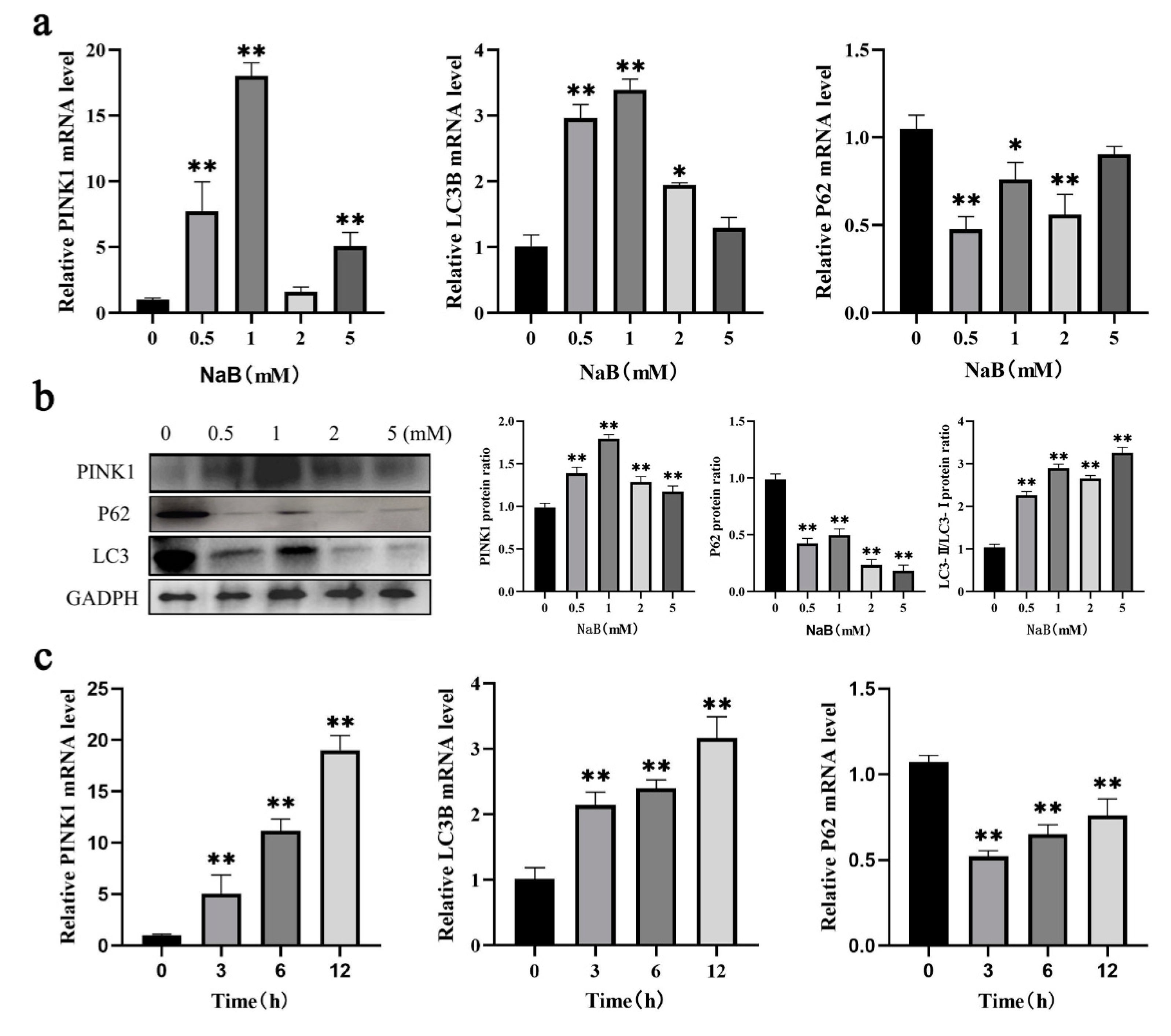
Figure 3.
Effect of NaB on ROS and MMP in BSCs. (a) The percentage of cells with decreased MMP. (b) The effect of 1mM NaB on cellular ROS level. (c) Ultrastructure of mitochondria under a transmission electron microscope (TEM).(A: control group; B: sodium butyrate treatment group; The red circle in B represents autophagosomes).
Figure 3.
Effect of NaB on ROS and MMP in BSCs. (a) The percentage of cells with decreased MMP. (b) The effect of 1mM NaB on cellular ROS level. (c) Ultrastructure of mitochondria under a transmission electron microscope (TEM).(A: control group; B: sodium butyrate treatment group; The red circle in B represents autophagosomes).
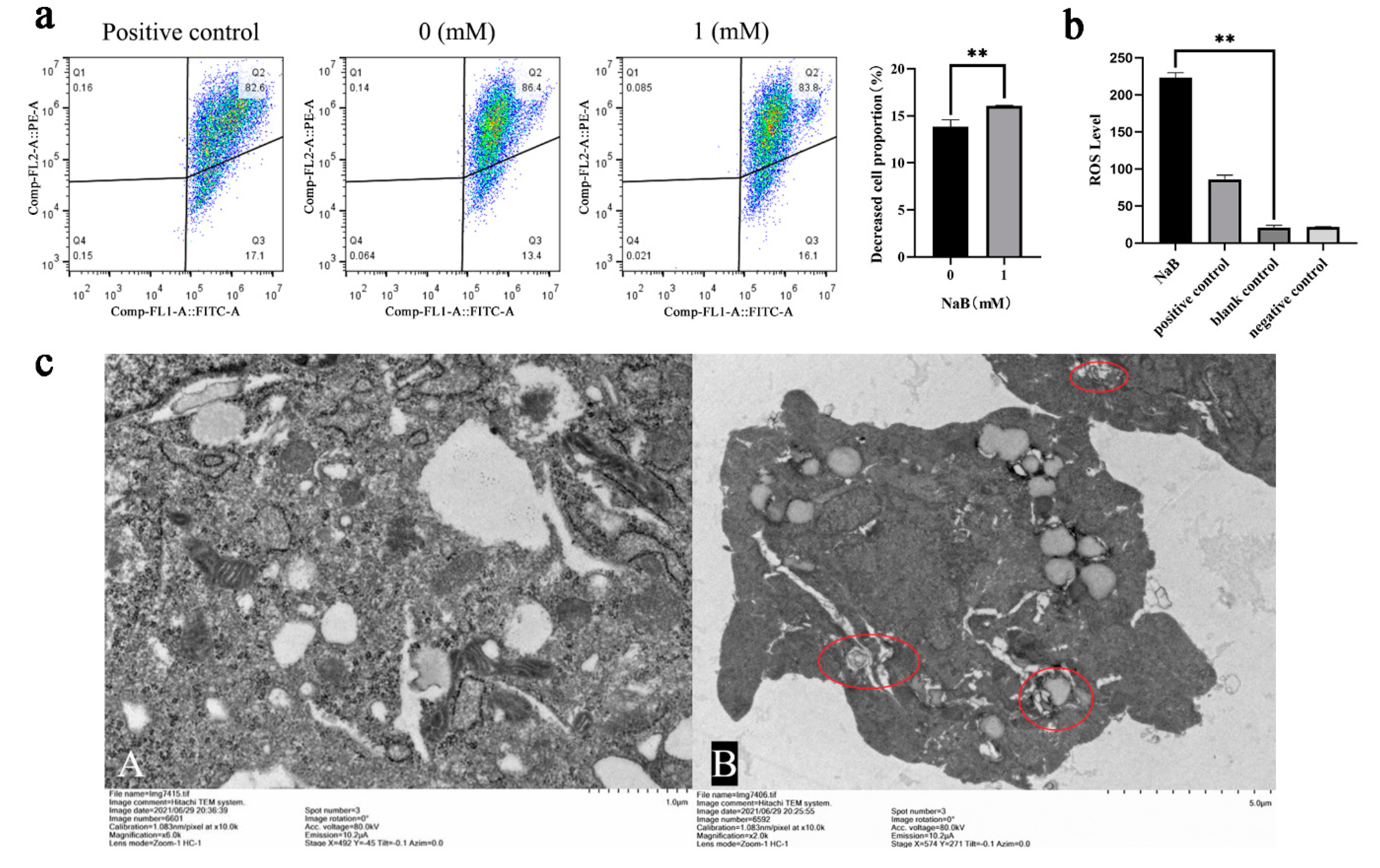
Figure 4.
Effect of NaB on mtDNA and ATP content. (a) The expression of mtDNA copy number genes with 1mM NaB treatment. (b) Changes of cellular ATP content with 1mM NaB treatment.
Figure 4.
Effect of NaB on mtDNA and ATP content. (a) The expression of mtDNA copy number genes with 1mM NaB treatment. (b) Changes of cellular ATP content with 1mM NaB treatment.
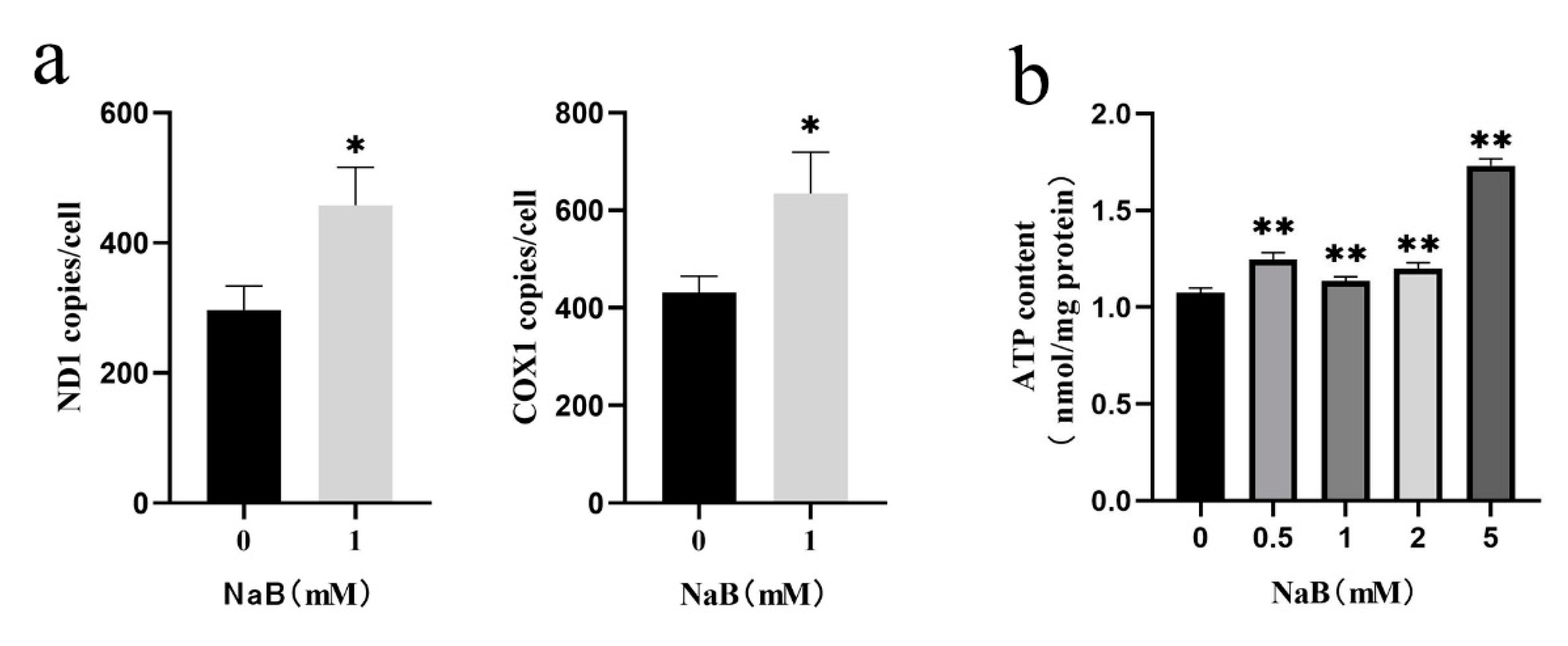
Figure 5.
Effect of NaB on mitophagy and apoptosis through the mTOR signaling pathway.(a) The mRNA expression of key genes in the mTOR signaling pathway with NaB treatment. (b) The mRNA expression of key genes in the mTOR signaling pathway with the PP242 treatment. (c) Key proteins expression in the mTOR signaling pathway with NaB and PP242 treatment, respectively. (d) The mRNA expression of apoptosis genes with the PP242 treatment. (e) The mRNA expression of mitophagy genes with the PP242 treatment.
Figure 5.
Effect of NaB on mitophagy and apoptosis through the mTOR signaling pathway.(a) The mRNA expression of key genes in the mTOR signaling pathway with NaB treatment. (b) The mRNA expression of key genes in the mTOR signaling pathway with the PP242 treatment. (c) Key proteins expression in the mTOR signaling pathway with NaB and PP242 treatment, respectively. (d) The mRNA expression of apoptosis genes with the PP242 treatment. (e) The mRNA expression of mitophagy genes with the PP242 treatment.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



