Submitted:
14 July 2023
Posted:
17 July 2023
You are already at the latest version
Abstract
Farmland edge plants can support high butterfly richness. Our objective was determine if, even in farms under intensive cultivation, the diversity of plants (mostly native) in crop edges is enough to sustain a high species richness of butterfly. We characterized the vegetation on the edges of Medicago sativa farms in central Mexico and identified the species of diurnals butterflies (Rhopalocera) associated. Butterflies and plants were counted along transects at the edges of a cul-tivated field during 24 months. We found 2710 individuals of plants, belonging to 48 different species from 24 families; 1749 individuals of diurnal butterflies, belonging to 57 species from six families. Most than half of the plant species found were native flora of central Mexico and with ethnobotanical use. A similarity analysis test showed significant differences in floristic composi-tion between transects. The Canonical Correspondence Analysis between butterfly species and plant families showed three groups. Six butterfly species were migratory and four mexican en-demic species, most of them associated with a group formed by Amaranthaceae, Euphorbiaceae, Annonaceae Lamiaceae Apiaceae and Fabaceae families. The high diversity of plants in our agro-ecosystem plays an important role to sustain a high diversity of butterflies and could be useful as biological corridors.
Keywords:
diurnals butterflies
; farmland
; edges flora
; migratory butterflies
1. Introduction
Some agro-ecosystems have a high diversity of insect species that take refuge in the edges of crop fields, which are formed by native vegetation or by an array of different crops [13,16,41]. Many studies [10,37,38,40,41,52] have reported that the structure and diversity of plants associated with farmland can support a high richness of butterfly species, which are, after Hymenoptera, the second most important pollinators [50]. Also, it have showed that the diversity of Lepidoptera was higher in transitional areas, located between agricultural and protected areas, because transitional areas have more flowering plants [6,23,32]. Furthermore, the heterogeneity of topographic conditions, and the richness and structural complexity of plant species provide many habitats, by protection for larvae and nectar for adult butterflies [41,52]. Other environmental conditions and cultivation techniques can have an influence on the abundance and diversity of Lepidoptera. Many works [1,47,52,56,59] have shown that an increase in soil moisture and the presence of crop field edges formed by native vegetation lead to a greater abundance and diversity of butterflies. Although some agricultural management systems rely on the use of chemical substances, intensive use of plough and closeness to urban areas can negatively affect the abundance and diversity of butterflies [4,36]. Another negative factor is the periodical cutting of crops [12,18,46], but its effect is diminished when the cutting is performed at the end of summer, when the life cycle of butterflies is complete [64].
The natural habitats of butterflies have been rapidly disappearing in recent years, while Alfalfa (Medicago sativa L.) crops occupy 32, 266, 605 ha in the world and 21.5% of the agricultural land in central Mexico [48]. Also, agricultural intensification through landscape homogenization is the main drivers of the butterflies diversity declines [19]. Therefore, the aim of this study is to determine that, even in farms under intensive cultivation, the diversity of plants (most of them native) in crop edges is enough to sustain high species richness and give refuge to endemic and migratory butterfly species.
2. Materials and Methods
Study site. The study was carried out in a Medicago sativa crop located in the state of Puebla (18°52’32’’ N and 98°25’51’’ W; 1686 m.a.s.l.) in Central Mexico. The climate is mainly temperate, with summer rainfall. The annual precipitation ranges from 700 to 1000 mm and the temperature ranges from 18° to 21° C. The soils are mostly Feozems and Fluvisols, which are good for agriculture. The original vegetation was dominated by woodlands in which Pinus species were associated with Abies religiosa (Kunth) Schltdl. & Cham or Quercus sp. [14]. Today, a high proportion of the land surface has been converted to farmland for growing corn, beans and lucerne. Small woodland areas remain in the northern and northwestern parts of the state, containing native vegetation.
Collection of butterflies and plants. Butterflies and plants were counted for five days every month, during 12 months, along four transects (300 m each one) in the edges of a cultivated field. Twenty-three Van Someren Rydon butterfly traps were set along each transect [61]. The traps were placed at 1.30 m from the ground, at 50 m from each other. We also collected butterflies manually using entomological nets. Temperature, relative humidity and wind speed were recorded using a pocket weather tracker (Kestrel 4000, Niels-Kellerman Co., Boothwyn). The butterflies were preserved in ethanol (70%). The plants surrounding each trap (an area of 1 m x 1 m) were counted, collected and dried at 35 °C. Butterflies and plants were identified using taxonomic keys.
Data analysis. A Simpson index [42] of diversity was calculated for both plants and butterflies, and the diversity and abundance between transects was compared using a Kruskal Wallis test. The similarity in the composition of plant species between transects was analyzed using a Jaccard index [33]. A similarity analysis test (ANOSIM [5]) was used to identify significant differences in floristic composition (software PAST v.1.15 [25]). A species accumulation curve was obtained by the nonparametric estimator Chao 1, to determine the sampling efficiency of butterflies [34], using EstimateS v. 9.1.0 [8]. Environmental variables (temperature, relative humidity and speed of wind) were compared between transects and during 12 months using a repeated-measures ANOVA [24]. A Canonical Correspondence Analysis (CCA [35]) was used to evaluate the effect of environmental variables on the butterfly community, and the relationship between different plant families and the butterfly species under study (software MVSP v. 3.12c [39]). The results were then subjected to a correlation analysis between plant and butterfly species (software NCSS 2001 [30]).
3. Results
3.1. Abundance and diversity
We found 2710 plant specimens belonging to 48 species and 24 families (Table 1). Most than half of the plant species found (58 %) are native flora of Central Mexico, and 91.66% of them have ethnobotanical use. The Simpson's diversity index showed no significant differences between transects, but the Jaccard index showed that the similarity in species composition was very low and the ANOSIM test showed significant differences in floristic composition (R= 0.042, p= 0.051, 9,999 permutations; α<0.10). Only 13 plant species were shared between transects.
We collected 1749 butterfly specimens belonging to 57 species and 6 families (Table 2). We found four Mexican endemic species Hamadryas atlantis Bates, Phyciodes pallescens Felder, Chlosyne ehrenbergii Geyer and Anthanassa sitalces Hall; six migratory species, Ascia monuste L., Smyrna blomfildia Fabricius, Eurema daira Godart, Eurema proterpia Fabricius, Vanessa atalanta Frühstorfer and Danaus plexippus L. and a species typical of woodland: Morpho polyphemus Westwood.
The environmental variables were not significantly different between transects throughout the year. But the abundance and diversity of butterflies showed significant differences between months; August and September always had the most abundance (H = 7.822, p = 0.0107), while February, March and April had the highest diversity (H = 5.92, p = 0.0034). The ACC between environmental variables and species families showed that environmental variables explained 38.6% of the variation in butterfly abundance in the first ordination axis and 10.2% in the second (Figure 1). Lycaenidae and Papilionidae were affected mostly by temperature, while Pieridae was affected by relative humidity and Nymphalidae was mostly affected by speed of wind. Sampling efficiency, as shown by a species accumulation curve was 87%. We identified three feeding guilds: nectar-feeding, mud-puddling and yeast-feeding (Table 2) that were present all year.
3.2. Influence of plants on the butterfly community
The Canonical Correspondence Analysis between butterfly species and plant families showed three groups (Figure 2). Plant families explained 58.4% of the abundance of butterfly species in the first ordination axis, and 42.6% in the second ordination axis. The plant families in the first group (Table 3) were associated with a higher number of butterfly species (17 species).
The correlation analysis between plant and butterfly species (Table 3) showed that the plant species Salvia longistyla Benth was correlated with more butterfly species (4). The butterflies that visited more species of plants were Leptophobia aripa (4) and Anthanassa sitalces (4). When applied this analysis only for migratory butterflies (Table 4), we found that A. sitalces and S. blomfildia were correlated with the highest number of plant species.
4. Discussion
4.1. Abundance and diversity
The results of the present study showed a similar number of plant families than other studies [2,17,26]. However, have been reported [11] a greater number of species (64) than our (48) in a lucerne crop, but with larger surface during a longer period of time. On the other hand, most than half of the plant species found are native flora of central Mexico, and 91.6% of them have ethnobotanical use. These results are similar to those reported by other authors [11,43,53]. This occurs when remnants of native flora colonize sites with adequate conditions such as the edges of crop fields; if these plants are not to removed from those sites, the diversity increase [10,37,40,41,52,55]. In our study, the ANOSIM test showed significant differences in floristic composition between transects. These results agree with literature [3,27,51], in areas with different crops, a consequence of the movement of seeds across neighboring habitats, especially when crop fields are distributed in a mosaic pattern. In our study site, the presence of farming plots with corn, beans and lucerne crops, as well as small remaining wooded areas containing native vegetation, could create a large-scale mosaic pattern with a high diversity of plants along the edges.
With respect to the butterfly community, the species accumulation curve showed that sampling effectiveness was adequate (87%), but the collection period could be extended. In spite of this, the butterfly community found had a higher species richness (57) than other studies that sampled for longer time over a larger area (27 species [10], 58 species [7], 31 species [9], 61 species [21], 30 species [13]). Also, only four of our species have been reported in other lucerne crops [9,22,57]. Our results showed that the butterfly community was sensible to environmental variables, as has been reported in other studies [15,58,65]. Nymphalidae was mostly affected by speed of wind, possibly because some species of this family are migratory (D. plexippus, S. blomfildia and V. atalanta). Furthermore some authors have reported that abundance of Nymphalidae increased in conserved areas or in biological corridors [15,49]. Particularly, Anaea aidea Guérin-Méneville (Nymphalidae) present in our study, has been reported by literature [49] as an indicator species in conserved areas; this called our attention, and because Nymphalidae was the second most abundant family in our study site, even though the agroecosystem under study was under an intense management (the crop is harvested every 28 days).
4.2. Influence of plants on the butterfly community
The correlation analysis between plant and butterfly species showed that endemic plants S. longistyla (Lamiaceae) and Erythrina coralloides DC. (Fabaceae) were positively correlated with more butterfly species (Table 3) and one migratory species: A. monuste. While Leptophobia aripa was associated with more plants, this butterfly has been reported as a crop pest of Brassica oleracea (Brassicaceae) in central Mexico [28,54], but in our study site it was associated with other plant families. The migratory butterfly D. plexippus was associated with Alternanthera sp Forssk (Amaranthaceae), but in literature [22,31], it was associated with the plant Asclepias curassavica (Apocynaceae), were it lays its eggs. We didn’t find specimens of A. curassavica in our study site. The explanation may be that D. plexippus is a migratory butterfly the uses our agroecosystem as a feeding site, not as a reproduction site. The migratory butterflies E. proterpia and S. blomfildia visited more plant species; both have been reported in areas with crop fields and little remaining wooded sites [44], similar to our study site. Therefore the endemic butterfly Chlosyne ehrenbergii was positively associated with the plants Leonotis nepetifolia L.R.Br. (Lamiaceae) and Alternanthera sp. (Amaranthaceae).
5. Conclusions
The particular structure and high diversity of plants that exists in the agroecosystem under study played an important role in sustaining a high diversity of butterflies. However, we must consider that a larger-scale mosaic of different crops surrounds our study site, and that could be responsible for the diversity of the butterfly community, as mentioned elsewhere in the literature [6,10,21,55]. The high diversity of the native vegetation (including endemic species) and the positive correlation between butterflies and plant species, despite intensive crop management system, are probably the factors that generate most of the resources needed for a rich butterfly community that includes migratory and endemic species. These factors also create an effective network that allows for movement between habitats, serving probably as biological corridors [7,13,19]. Considering that many plant species found in crop field edges have ethnobotanical use, the results of this study suggest crop fields can provide refuge for butterflies as well as other benefits derived from the use of edge plants.
Author Contributions
PF-Z, SPR-A and ARA-H planed, designed and executed experimental work. PF-Z identified butterflies species and ARA-H identified plant species. SPR-A, NS O-S and DM conducted data analyses. SPR-A wrote the manuscript.
Funding
This research received no external funding
Acknowledgments
We thank the agroecosystem owners for allowing us to conduct this study. P Fascinetto-Zago and SP Rivas-Arancibia were financially supported by Vicerrectoría de Investigación y Estudios de Posgrado (VIEP) of Benemérita Universidad Autónoma de Puebla, México (BUAP) to conduct this research.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abós, F. El pluricultivo y la presencia de márgenes mantienen la diversidad biológica en los agrosistemas. Ecología 2002, 16, 273–285. [Google Scholar]
- Albino, C.; Cervantes, H.; López, M.; Ríos-Casanova, L.; Lira, R. Patrones de diversidad y aspectos etnobotánicos de las plantas arvenses del valle de Tehuacán-Cuicatlán: el caso de San Rafael, municipio de Coxcatlán, Puebla. Revista Mexicana de Biodiversidad 2011, 82, 1005–1019. [Google Scholar]
- Alignier, A.; Bretagnolle, V.; Petit, S. Spatial patterns of weeds along a gradient of landscape complexity. Basic Appl. Ecol. 2012, 13, 328–337. [Google Scholar] [CrossRef]
- Baquero, E.; Moraza, M.L.; Ariño, H.A.; Jordana, R. Mariposas Diurnas de Pamplona. Ayuntamiento de Pamplona: España; 2011.
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clausen, H.; Holbeck, H.; Reddersen, J. Factors influencing abundance of butterflies and burnet moths in the uncultivated habitats of an organic farm in Denmark. Biol. Conserv. 2001, 98, 167–178. [Google Scholar] [CrossRef]
- Collinge, S.K.; Prudic, K.L.; Oliver, J.C. Effects of Local Habitat Characteristics and Landscape Context on Grassland Butterfly Diversity. Conserv. Biol. 2003, 17, 178–187. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: statistical estimation of species richness and shared species from simples, version 8.0. 2006. Available from: http://purl. oclc. org/estimates.
- Davis, J.D.; Hendrix, S.D.; Debinski, D.M.; Hemsley, C.J. Butterfly, bee and forb community composition and cross-taxon incongruence in tallgrass prairie fragments. J. Insect Conserv. 2007, 12, 69–79. [Google Scholar] [CrossRef]
- Davros, N.M.; Debinski, D.M.; Reeder, K.F.; Hohman, W.L. Butterflies and Continuous Conservation Reserve Program Filter Strips: Landscape Considerations. Wildl. Soc. Bull. 2006, 34, 936–943. [Google Scholar] [CrossRef]
- Delgado-Castillo, J.C. Especies de Malezas Asociadas a Cultivos del Bajío de Guanajuato, México. Celaya, Guanajuato: Programa de Sanidad Vegetal, SAGARPA. 2010. Available from: http://www.agricolaunam.org.mx/colecciones%20virtuales/Malezas%20del%20Bajio%20JUAN%20CARLOS%20DELGADO.pdf.
- Erhardt, A. Diurnal Lepidoptera: Sensitive Indicators of Cultivated and Abandoned Grassland. J. Appl. Ecol. 1985, 22, 849. [Google Scholar] [CrossRef]
- Fahrig, L.; Girard, J.; Duro, D.; Pasher, J.; Smith, A.; Javorek, S.; King, D.; Lindsay, K.F.; Mitchell, S.; Tischendorf, L. Farmlands with smaller crop fields have higher within-field biodiversity. Agric. Ecosyst. Environ. 2014, 200, 219–234. [Google Scholar] [CrossRef]
- Fascinetto, Z.P. Dinámica e Identificación de la Comunidad de Mariposas Diurnas (Lepidoptera: Rhopalocera) Asociadas a un Agroecosistema en Atlixco, Puebla. Unpublished thesis, Benemérita Universidad Autónoma de Puebla, México. 2015.
- Hernández, D.M.F. Butterflies of the agricultural experiment station of tropical roots and tubers, and Santa Ana, Camagüey, Cuba: an annotated list. Acta ZoolÓGica Mex. 2007, 23, 43–75. [Google Scholar] [CrossRef]
- Fernandez, A.L.; Sheaffer, C.C. Tautges NE, Putnam DH, Hunter M. Alfalfa, Wildlife and the Environment. National Alfalfa and Forage Alliance, St. Paul: Minnesota; 2019.
- Gámez, A.; De Gouveia, M.; Álvarez, W.; Pérez, H.; Mondragón, A.; Alvarado, H.; Vásquez, C. Flora arvense asociada a un agroecosistema tipo conuco en la comunidad de Santa Rosa de Ceiba Mocha en el estado Guárico. Bioagro 2014, 26, 177–182. [Google Scholar]
- Gerell, R. Management of roadside vegetation: effects on density and species diversity of butterflies in Scania, south Sweden. Entomologisk Tidskrift 1997, 118, 171–176. [Google Scholar]
- Giuliano, D.; Cardarelli, E.; Bogliani, G. Grass management intensity affects butterfly and orthopteran diversity on rice field banks. Agric. Ecosyst. Environ. 2018, 267, 147–155. [Google Scholar] [CrossRef]
- Glassberg, J. A Swift Guide to the Butterflies of Mexico and Central America. Sunstreak Books: Princeton University Press; 2018.
- González-Estébanez, F.J.; García-Tejero, S.; Mateo-Tomás, P.; Olea, P.P. Effects of irrigation and landscape heterogeneity on butterfly diversity in Mediterranean farmlands. Agric. Ecosyst. Environ. 2011, 144, 262–270. [Google Scholar] [CrossRef]
- Graves, S.D.; Shapiro, A.M. Exotics as host plants of the California butterfly fauna. Biol. Conserv. 2003, 110, 413–433. [Google Scholar] [CrossRef]
- Grill, A.; Cleary, D.F. Diversity patterns in butterfly communities of the Greek nature reserve Dadia. Biol. Conserv. 2003, 114, 427–436. [Google Scholar] [CrossRef]
- Gurevitch, J.; Chester Jr, S.T. Analysis of repeated measures experiments. Ecology 1986, 67, 251–255. [Google Scholar] [CrossRef]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. Past Palaeontological, Software version 1.18. Copyright Hammer and Harper; 2003.
- Hassannejad, S.; Ghafarbi, S.P. Weed flora survey in alfalfa (Medicago sativa L.) fields of Shabestar (northwest of Iran). Arch. Agron. Soil Sci. 2014, 60, 971–991. [Google Scholar] [CrossRef]
- Hernández-Mejía, C.; Llorente-Bousquets, J.; Vargas-Fernández, I.; Luis-Martínez, A. 2008: Las mariposas (Hesperioidea y Papilionoidea) de Malinalco, Estado de México. Rev. Mex. De Biodivers. 2014, 79, 117–130. [Google Scholar]
- Hernández-Mejía, C.; Flores-Gallardo, A.; Llorente-Bousquets, J. Morfología del corion en Leptophobia (Lepidoptera: Pieridae) e importancia Taxonómica. Southwest. Entomol. 2015, 40, 351–368. [Google Scholar] [CrossRef]
- Hernández, P.E.; Bastida, F.; Pujadas, S.A.; Pallavicini, Y.; Izquierdo, F.J.; González, A.J.L. Efecto de la intensificación agrícola en la diversidad taxonómica y funcional de las especies arvenses del banco de semillas en cultivos cerealistas. Poster session presented at: XV Congreso de la Sociedad Española de Malherbología. 2015 Octubre 19-22, Sevilla, España.
- Hintze, J. Software NCSS, PASS and GESS, Kaisville Utah: NCSS; 2008.
- Iwashina, T. Flavonoid Function and Activity to Plants and Other Organisms. Biol. Sci. Space 2003, 17, 24–44. [Google Scholar] [CrossRef]
- Iwema, D.; Janes, E.; Rosenberger, D. Late Summer Butterfly Species Richness and Abundance in Bourbonnais Township, Northeastern Illinois. Department of Biological Sciences, Olivet Nazarene University; 2016.
- Jaccard, P. Nouvelles recherches sur la distribution florale. Bull. De La Société Vaudoise Des Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Jiménez-Valverde, A.; Hortal, J. Las curvas de acumulación de especies y la necesidad de evaluar los inventarios biológicos. Rev. IbÉRica De Aracnol. íA 2003, 8, 151–161. [Google Scholar]
- Johnson, D.E. Métodos Multivariados Aplicados al Análisis de Datos, International Thomson, México; 1998.
- Kitahara, M.; Sei, K.; Fujii, K. Patterns in the structure of grassland butterfly communities along a gradient of human disturbance: further analysis based on the generalist/specialist concept. Popul. Ecol. 2000, 42, 135–144. [Google Scholar] [CrossRef]
- Kivinen, S.; Luoto, M.; Kuussaari, M.; Saarinen, K. Effects of land cover and climate on species richness of butterflies in boreal agricultural landscapes. Agric. Ecosyst. Environ. 2007, 122, 453–460. [Google Scholar] [CrossRef]
- Neff, P.K.; Locke, C.; Lee-Mӓder, E. Assessing a farmland set-aside conservation program for an endangered butterfly: USDA State Acres for Wildlife Enhancement (SAFE) for the Karner blue butterfly. J. Insect Conserv. 2017, 21, 929–941. [Google Scholar] [CrossRef]
- Kovach, W.L. MVSP-A MultiVariate Statistical Package for Windows. Verion 3.13. Pentraeth, Wales: Kovach Computing Services; 2004.
- Kuussaari, M.; Heliölä, J.; Luoto, M.; Pöyry, J. Determinants of local species richness of diurnal Lepidoptera in boreal agricultural landscapes. Agric. Ecosyst. Environ. 2007, 122, 366–376. [Google Scholar] [CrossRef]
- Loos, J.; Dorresteijn, I.; Hanspach, J.; Fust, P.; Rakosy, L.; Fischer, J. Low-Intensity Agricultural Landscapes in Transylvania Support High Butterfly Diversity: Implications for Conservation. PLOS ONE 2014, 9, e103256. [Google Scholar] [CrossRef]
- Magurran, A.E. Diversity Indices and Species Abundance Models. Ecological Diversity and Its Measurement: Springer, Dordrecht; 1988.
- Mas, M.T.; Verdú, A.M.C. Biodiversidad de la flora arvense en cultivos de mandarino según el manejo del suelo en las interfilas. Boletín De Sanid. Veg. Plagas 2005, 31, 231–242. [Google Scholar]
- Marín, L.; León-Cortés, J.L.; Stefanescu, C. The effect of an agro-pasture landscape on diversity and migration patterns of frugivorous butterflies in Chiapas, Mexico. Biodivers. Conserv. 2008, 18, 919–934. [Google Scholar] [CrossRef]
- Martínez-Noble, J.I.; Meléndez-Ramírez, V.; Delfín-González, H.; Pozo, C. Mariposas de la selva mediana subcaducifolia de Tzucacab, con nuevos registros para Yucatán, México. Rev. Mex. de Biodivers. 2015, 86, 348–357. [Google Scholar] [CrossRef]
- Munguira, M.L.; Thomas, J.A. Use of Road Verges by Butterfly and Burnet Populations, and the Effect of Roads on Adult Dispersal and Mortality. J. Appl. Ecol. 1992, 29, 316. [Google Scholar] [CrossRef]
- Oostermeijer, J.; van Swaay, C. The relationship between butterflies and environmental indicator values: a tool for conservation in a changing landscape. Biol. Conserv. 1998, 86, 271–280. [Google Scholar] [CrossRef]
- Pedroza, S.A.; Ríos, F.J.L.; Torres, M.M.; Cantú BJE, Piceno SC, Yáñez CLG. Eficiencia del agua de riego en la producción de maíz forrajero (Zea mays L.) y alfalfa (Medigaco sativa): impacto social y económico. Terra Latinoamericana 2014, 32, 231–239. [Google Scholar]
- Pozo de la Tijera, M.C.; Salas-Suárez, N.; Maya-Martínez, A.; Prado-Cuellar, B. Uso y Monitoreo de los Recursos Naturales en el Corredor Biológico Mesoamericano (Áreas Focales Xpujil-Zoh Laguna y Carrillo Puerto). Colegio de la Frontera Sur, Unidad Chetumal, CONABIO; 2005.
- Proctor, M.; Yeo, P.; Lack, A. The Natural History of Pollination. HarperCollins Publishers, Britain; 1996. [CrossRef]
- Ramírez-Hernández, A.; Micó, E.; Marcos-García, M.d.L. .; Brustel, H.; Galante, E. The “dehesa”, a key ecosystem in maintaining the diversity of Mediterranean saproxylic insects (Coleoptera and Diptera: Syrphidae). Biodivers. Conserv. 2014, 23, 2069–2086. [Google Scholar] [CrossRef]
- Saarinen, K. A comparison of butterfly communities along field margins under traditional and intensive management in SE Finland. Agric. Ecosyst. Environ. 2002, 90, 59–65. [Google Scholar] [CrossRef]
- Sans, F.X.; Armengot, L.; Bassa, M.; Blanco-Moreno, J.M.; Caballero-López, B.; Chamorro, L.; José-María, L. La intensificación agrícola y la diversidad vegetal en los sistemas cerealistas de secano mediterráneos: implicaciones para la conservación. Rev. Ecosistemas 2013, 22, 30–35. [Google Scholar]
- Santiago, L.J.A.; García, B.L.E.; Rojas, J.C.; Perales, R.H. Host selection behavior of Leptophobia aripa (Lepidoptera: Pieridae). Fla. Entomol. 2006, 89, 127–134. [Google Scholar] [CrossRef]
- Sawchik, J.; Dufrêne, M.; Lebrun, P. Estimation of habitat quality based on plant community, and effects of isolation in a network of butterfly habitat patches. Acta Oecologica 2003, 24, 25–33. [Google Scholar] [CrossRef]
- Sparks, T.; Parish, T. Factors affecting the abundance of butterflies in field boundaries in Swavesey fens, Cambridgeshire, UK. Biol. Conserv. 1995, 73, 221–227. [Google Scholar] [CrossRef]
- Tabashnik, B.E. Host range evolution: the shift from native legume hosts to alfalfa by the butterfly, Colias philodice eriphyle. Evolution 1983, 37, 150–162. [Google Scholar] [CrossRef]
- Tórrez, M.; Arendt, W.; Maes, J.M. Comunidades de aves y lepidópteros diurnos y las relaciones entre ellas en bosque nuboso y cafetal de Finca Santa Maura, Jinotega. Encuentro 2013, 60–79. [Google Scholar] [CrossRef]
- Valtonen, A.; Saarinen, K.; Jantunen, J. Intersection reservations as habitats for meadow butterflies and diurnal moths: Guidelines for planning and management. Landsc. Urban Plan. 2007, 79, 201–209. [Google Scholar] [CrossRef]
- Vibrans, H.; Tenorio, L.P. Malezas de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, CONABIO, México; 2012.
- Villarreal, H.; Álvarez, M.; Córdoba, S.; Escobar, F.; Fagua, G.; Gast, F.; Mendoza, H.; Ospina, M.; Umaña, A. Manual de Métodos para el Desarrollo de Inventarios de Biodiversidad. Instituto de Investigaciones de Recursos Biológicos Alexander Von Humboldt. Colombia; 2006.
- Villaseñor, J.L.; Espinosa-Garcia, F.J. The alien flowering plants of Mexico. Divers. Distrib. 2004, 10, 113–123. [Google Scholar] [CrossRef]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. de Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef]
- Wettstein, W.; Schmid, B. Conservation of arthropod diversity in montane wetlands: effect of altitude, habitat quality and habitat fragmentation on butterflies and grasshoppers. J. Appl. Ecol. 1999, 36, 363–373. [Google Scholar] [CrossRef]
- Zambrano-González, G.; Ortiz-Ordóñez, G. Diversidad de Lepidópteros diurnos en tres localidades del corredor biológico y multicultural Munchique-Pinche, Cauca, Colombia. Bol. Cient. Mus. Hist. Nat. 2009, 13, 241–224. [Google Scholar]
Figure 1.
Canonical Correspondence Analysis between Lepidoptera (Rhopalocera) families and environmental variables (Temperature, Humidity, Speed of wind) along two years in the edges of the Medicago sativa crop (38.6% in the first ordination axis and 10.2% in the second).
Figure 1.
Canonical Correspondence Analysis between Lepidoptera (Rhopalocera) families and environmental variables (Temperature, Humidity, Speed of wind) along two years in the edges of the Medicago sativa crop (38.6% in the first ordination axis and 10.2% in the second).
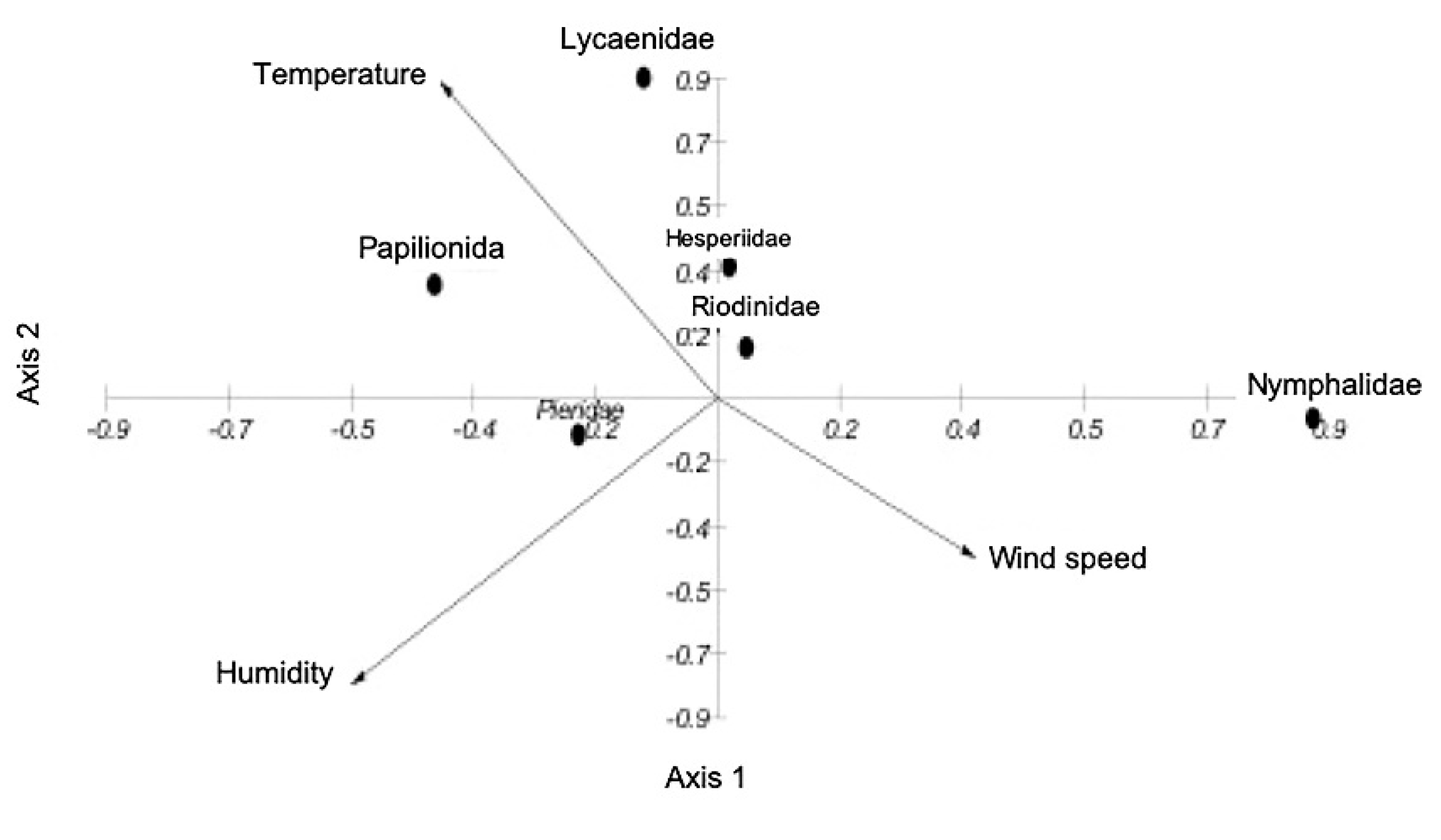
Figure 2.
Canonical Correspondence Analysis between butterfly and plant families (in the first ordination axis 58.4%; second ordination axis 42.6%). Butterfly species: Ag.v: Agraulis vanillae, An.a: Anaea aidea, A.a: Ancyloxypha arene, As.m: Ascia monuste, At.i: Asterocampa idyja, Ba.p: Battus philenor, Bi.h: Biblis hyperia, Cl.sp: Calephelis sp., Ca.n: Catasticta nimbice, Ch.e: Chlosyne ehrenbergii, Ch.l: Chlosyne lacinia, Ci.s: Cissia similis, Cg.h: Cogia hippalus, Co.c: Colias cesonia, Co.e: Colias eurytheme, Da.g: Danaus gilippus, Da.p: Danaus plexippus, Di.m: Dione moneta, E.d: Eurema daira, E.p: Eurema proterpia, E.s: Eurema salome, H.c: Hemiargus ceraunus, H.i: Hemiargus isola, J.c: Junonia coenia, Lp.a: Leptophobia aripa, Le.c: Leptotes cassius, Le.m: Leptotes marina, L.sp: Lerema sp., N.i: Nathalis iole, N.a: Nymphalis antiopa, Pa.p: Papilio polyxenes, Pr.p: Parides photinus, Pb.b: Phoebis boisdusvalii, Pl.m: Pholisora mejicanus, Pc.p: Phyciodes pallescens, Ah.s: Anthanassa sitalces, Ah.t: Anthanassa texana, Po.z: Poanes zabulon, Py.c: Pyrgus communis, Sm.b: Smyrna blomfildia, St.a: Strymon astiocha, St.m: Strymon melinus, U.d: Urbanus dorantes, U.p: Urbanus procnes, V.an:Vanessa anabella, V.at: Vanessa atalanta y Z.c: Ziegleria ceromia.
Figure 2.
Canonical Correspondence Analysis between butterfly and plant families (in the first ordination axis 58.4%; second ordination axis 42.6%). Butterfly species: Ag.v: Agraulis vanillae, An.a: Anaea aidea, A.a: Ancyloxypha arene, As.m: Ascia monuste, At.i: Asterocampa idyja, Ba.p: Battus philenor, Bi.h: Biblis hyperia, Cl.sp: Calephelis sp., Ca.n: Catasticta nimbice, Ch.e: Chlosyne ehrenbergii, Ch.l: Chlosyne lacinia, Ci.s: Cissia similis, Cg.h: Cogia hippalus, Co.c: Colias cesonia, Co.e: Colias eurytheme, Da.g: Danaus gilippus, Da.p: Danaus plexippus, Di.m: Dione moneta, E.d: Eurema daira, E.p: Eurema proterpia, E.s: Eurema salome, H.c: Hemiargus ceraunus, H.i: Hemiargus isola, J.c: Junonia coenia, Lp.a: Leptophobia aripa, Le.c: Leptotes cassius, Le.m: Leptotes marina, L.sp: Lerema sp., N.i: Nathalis iole, N.a: Nymphalis antiopa, Pa.p: Papilio polyxenes, Pr.p: Parides photinus, Pb.b: Phoebis boisdusvalii, Pl.m: Pholisora mejicanus, Pc.p: Phyciodes pallescens, Ah.s: Anthanassa sitalces, Ah.t: Anthanassa texana, Po.z: Poanes zabulon, Py.c: Pyrgus communis, Sm.b: Smyrna blomfildia, St.a: Strymon astiocha, St.m: Strymon melinus, U.d: Urbanus dorantes, U.p: Urbanus procnes, V.an:Vanessa anabella, V.at: Vanessa atalanta y Z.c: Ziegleria ceromia.
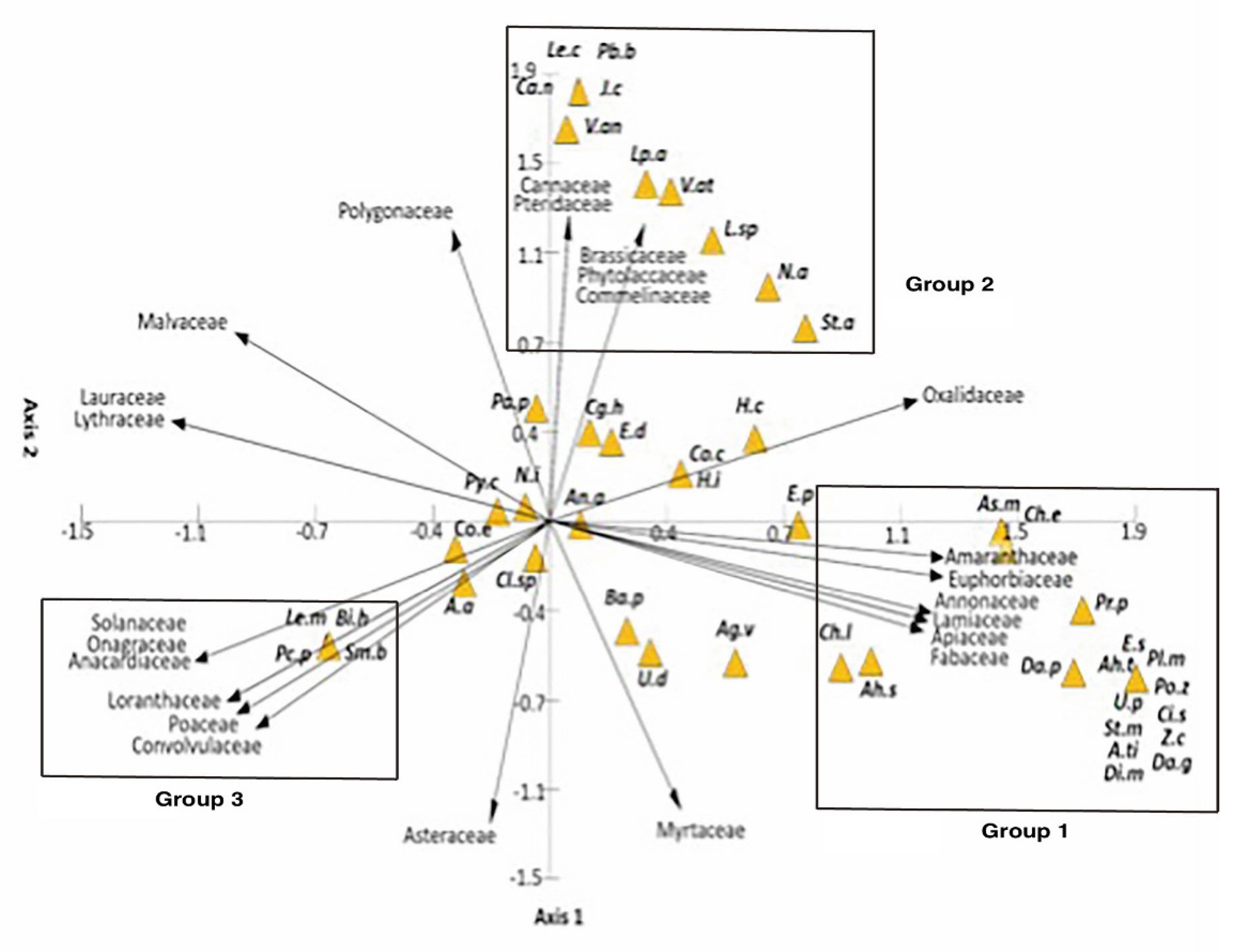

Table 1.
Abundance and ethnobotanical use [60] of plant taxa found in the edges of a Medicago sativa field in central Mexico. *Species native to Mexico [62,63]. (-)No ethnibotanical use registered.
| Families | Taxa | Abundance | Ethnobotanical use |
|---|---|---|---|
| Apiaceae | Foeniculum vulgare, Mill., 1768 | 3 | curative and comestible |
| Asteraceae | *Aldama dentata, La Llave, 1824 | 19 | forage |
| *Bidens odorata, L., 1753 | 32 | curative, forage and comestible | |
| *Galinsoga parviflora, Cav., 1796 | 4 | forage | |
| *Sanvitalia procumbens, Lam., 1792 | 13 | curative and ornament | |
| Taraxacum officinale, F.H.Wigg., 1780 | 56 | curative, forage, comestible, and melliferous | |
| Brassicaceae | *Lepidium virginicum, L., 1753 | 6 | curative, forage and comestible |
| Nasturtium officinale, W.T. Aiton, 1812 | 4 | curative and comestible | |
| Amaranthaceae | Alternanthera sp., Forssk, 1775 | 40 | -- |
| Chenopodium album, L., 1753 | 4 | Curative | |
| Phytolaccaceae | *Phytolacca americana, L., 1753 | 3 | curative, comestible, ornament and to colour |
| Polygonaceae | *Persicaria hydropiperoides, (Michx.) Small, 1903 | 2 | curative, forage and to colour |
| Rumex conglomeratus, Murray, 1770 | 8 | curative and comestible | |
| Commelinaceae | *Commelina diffusa, Burm.f., 1768 | 60 | curative, forage and ornament |
| Fabaceae | *Erythrina coralloides, Moc. y Sessé ex DC., 1825 | 1 | curative, comestible, ornament and artisan |
| Melilotus albus, Medik, 1786 | 2 | forage and melliferous | |
| Medicago lupulina, L., 1753 | 31 | forage and melliferous | |
| *Vigna luteola, (Jacq.) Benth., 1859 | 3 | curative and comestible | |
| Trifolium repens, L., 1753 | 586 | forage and comestible | |
| Lamiaceae | Leonotis nepetifolia, (L.) R.Br.,1811 | 56 | curative, ornament and melliferous |
| *Salvia mexicana, L., 1753 | 4 | forage, comestible, ornament and melliferous | |
| *Salvia longistyla, Benth, 1833 | 93 | Curative | |
| Lauraceae | *Persea americana, Mill., 1768 | 5 | curative and comestible |
| Annonaceae | Annona cherimola, Mill., 1768 | 2 | comestible and combustible |
| Euphorbiaceae | *Euphorbia heterophylla, L., 1753 | 2 | curative |
| Ricinus communis, L., 1753 | 74 | curative | |
| Malvaceae | *Anoda cristata, (L.) Schltdl., 1837 | 6 | curative, forage, comestible and ornament |
| *Kearnemalvastrum lacteum, (Ait.) D.M.Bates, 1967 | 2 | curative, and forage | |
| Malva parviflora, L., 1753 | 9 | curative, forage and comestible | |
| *Sida haenkeana, C.Presl, 1835 | 32 | -- | |
| Lythraceae | *Cuphea angustifolia, Jacq. ex Koehne, 1877 | 8 | curative |
| Myrtaceae | *Psidium guajava, L., 1753 | 5 | curative, forage, comestible, artisan, to colour and combustible |
| Onagraceae | *Oenothera rosea, L´Hér. ex Ait., 1789 | 70 | Curative and ornament |
| Oxalidaceae | *Oxalis corniculata, L., 1753 | 98 | curative, forage, comestible and ornament |
| Poaceae | Arundo donax, L., 1753 | 37 | curative, forage, artisan and construction |
| Bromus carinatus, Hook. & Arn., 1840 | 6 | forage and comestible | |
| Chloris gayana, Kunth., 1829 | 1096 | forage | |
| *Ixophorus unisetus, (J.Presl) Schltdl.,1861 | 44 | forage | |
| *Setaria parviflora, (Poir.) Kerguélen, 1987 | 28 | forage | |
| Pteridaceae | Adiantum sp., L., 1753 | 10 | - |
| Loranthaceae | *Psittacanthus calyculatus, G.Don, 1834 | 1 | curative and artisan |
| Anacardiaceae | Schinus molle, L., 1753 | 1 | curative, forage, comestible, to colour, combustible and construction |
| Convolvulaceae | *Ipomoea purpurea, (L.) Roth., 1787 | 28 | curative and ornament |
| Solanaceae | *Physalis philadelphica, Lam., 1786 | 2 | curative, forage and comestible |
| *Solanum americanum, Mill., 1768 | 4 | curative, comestible and melliferous | |
| *Solanum lanceolatum, Cav., 1795 | 17 | curative, forage, comestible, and melliferous | |
| Solanum sp., L., 1753 | 3 | - | |
| Cannaceae | Canna indica, L., 1753 | 90 | comestible, ornament and artisan |
| 24 | 48 | 2710 |
Table 2.
Abundance and feeding guilds of the butterfly taxa (Lepidoptera: Rhopalocera) found in the edges of the Medicago sativa field. *Endemic species of Mexico [27,45]. ** Migratory species [20], (x) presence (-) absence.
| Families | Species | Abundance | Nectarfeeding | Yeastfeeding | Mud-puddling |
|---|---|---|---|---|---|
| Pieridae | Colias eurytheme Boisduval, 1852 | 1035 | x | - | - |
| Colias cesonia (Stoll,1790) | 5 | x | - | - | |
| **Eurema mexicana (Boisduval, 1836) | 2 | x | - | - | |
| Eurema salome (Reakirt, 1866) | 2 | x | - | - | |
| **Eurema daira (Godart, 1819) | 23 | x | - | x | |
| **Eurema proterpia (Fabricius, 1775) | 11 | x | x | x | |
| Eurema boisduvaliana (C. Felder & R. Felder, 1865) | 3 | x | - | - | |
| Leptophobia aripa (Boisduval, 1836) | 99 | x | - | - | |
| Catasticta nimibice (Boisduval, 1836) | 1 | x | - | - | |
| **Ascia monuste (Linnaeus, 1764) | 11 | x | - | - | |
| Nathalis iole Boisduval, 1836 | 22 | x | - | - | |
| Phoebis boisdusvalii (C. Felder & R. Felder, 1861) | 1 | x | - | - | |
| Phoebis agarithe (Boisduval, 1836) | 2 | x | x | x | |
| Nymphalidae | Chlosyne lacinia (Geyer, 1837) | 20 | x | - | - |
| *Chlosyne ehrenbergii (Geyer, [1833]) | 64 | x | - | - | |
| Vanessa atalanta (Frühstorfer, 1909) | 16 | x | x | - | |
| Vanessa annabella (Field, 1971) | 8 | x | - | - | |
| Anthanassa texana (W.H. Edwards, 1863) | 16 | x | - | - | |
| * Phyciodes pallescens (R. Felder, 1869) | 3 | x | - | - | |
| Anthanassa sitalces (A. Hall, 1917) | 6 | x | - | - | |
| Agraulis vanillae (Riley, 1926) | 2 | x | - | - | |
| Anaea aidea (Guérin-Méneville,[1844]) | 17 | - | x | - | |
| Dione moneta Butler, 1873 | 1 | x | - | - | |
| Danaus gilippus (H.W. Bates, 1863) | 2 | x | - | - | |
| **Danaus plexippus (Linnaeus, 1758) | 15 | x | - | - | |
| Junonia coenia Hübner, [1822] | 1 | x | - | - | |
| Asterocampa idyja (Geyer, [1828]) | 1 | x | x | - | |
| Cissia similis (Butler, 1867) | 1 | - | x | - | |
| Nymphalis antiopa (Linnaeus, 1758) | 3 | x | x | ||
| Biblis hyperia (Cramer, 1779) | 1 | - | x | - | |
| **Smyrna blomfidia (Fabricius, 1781) | 1 | - | x | - | |
| Morpho polyphemus Westwood, 1851 | 5 | - | x | - | |
| *Hamadryas atlantis (H. Bates, 1864) | 1 | - | x | - | |
| Cyllopsis sp. R. Felder, 1869 | 14 | x | x | - | |
| Papilionidae | Battus philenor (Linnaeus, 1771) | 38 | x | - | - |
| Parides photinus (Doubleday, 1844) | 13 | x | - | x | |
| Papilio polyxenes Cramer, 1782 | 8 | x | - | - | |
| Lycaenidae | Hemiargus isola (Reakirt, [1867]) | 3 | x | x | x |
| Hemiargus ceraunus (Butler & H. Druce, 1872) | 8 | x | x | x | |
| Strymon melinus Hübner, 1818 | 1 | x | - | - | |
| Strymon astiocha (Prittwitz, 1865) | 5 | x | - | - | |
| Ziegleria ceromia (Hewitson, 1877) | 1 | x | - | x | |
| Leptotes marina (Reakirt, 1868) | 9 | x | x | x | |
| Leptotes cassius (Cramer, 1775) | 3 | x | x | x | |
| Riodinidae | Calephelis spp. Grote & Robinson, 1869 | 35 | x | - | - |
| Hesperiidae | Pyrgus communis (Grote, 1872) | 109 | x | x | x |
| Ancyloxypha arene (W.H. Edwards, 1871) | 60 | x | - | - | |
| Lerema spp. Scudder, 1872 | 4 | x | - | - | |
| Urbanus dorantes (Stoll, 1790) | 17 | x | - | - | |
| Urbanus procne (Plötz, 1881) | 2 | x | - | - | |
| Pholisora mexicana (Reakirt, 1867) | 1 | x | - | - | |
| Cogia hippalus (W.H. Edwards, 1882) | 6 | x | - | - | |
| Poanes zabulon (Boisduval & Le Conte, [1837]) | 2 | x | - | - | |
| Pyrrhopyge chalybea Scudder, 1872 | 1 | x | - | - | |
| Cymaenes fraus (Godman, 1900) | 1 | x | - | - | |
| Pompeius pompeius (Latreille, [1824]) | 3 | x | - | - | |
| Staphylus spp. Godman & Salvin, 1896 | 4 | x | - | - | |
| 6 | 57 | 1749 |
Table 3.
Correlation analysis between plant and butterfly species in the edges of the Medicago sativa crop. Groups are defined in Figure 2.
Table 3.
Correlation analysis between plant and butterfly species in the edges of the Medicago sativa crop. Groups are defined in Figure 2.
| Group | Plant species | Butterfly species | p | r2 |
|---|---|---|---|---|
| 1 | Erythrina coralloides | Ascia monuste | 0.0001 | 0.42 |
| Anthanassa texana | 0.004 | 0.164 | ||
| Anthanassa sitalces | 0.0001 | 0.609 | ||
| 1 | Salvia longistyla | Ascia monuste | 0.0001 | 0.503 |
| Anthanassa texana | 0.004 | 0.067 | ||
| Anthanassa sitalces | 0.0001 | 0.718 | ||
| Chlosyne lacinia | 0.006 | 0.208 | ||
| 1 | Melilotus albus | Anthanassa sitalces | 0.014 | 0.066 |
| 1 | Leonotis nepetifolia | Chlosyne ehrenbergii | 0.019 | 0.032 |
| 1 | Alternanthera sp. | Chlosyne ehrenbergii | 0.0001 | 0.378 |
| Ziegleria ceromia | 0.0001 | 0.385 | ||
| 1 | Melilotus albus | Anthanassa sitalces | 0.014 | 0.066 |
| 1 | Vigna luteola | Chlosyne ehrenbergii | 0.0002 | 0.104 |
| 1 | Euphorbia heterophylla | Chlosyne ehrenbergii | 0.0009 | 0.078 |
| Ziegleria ceromia | 0.0003 | 0.104 | ||
| 1 | Ricinus communis | Ziegleria ceromia | 0.0001 | 0.88 |
| 2 | Lepidium virginicum | Leptophobia aripa | 0.0001 | 0.909 |
| 2 | Nasturtium officinale | Leptophobia aripa | 0.0001 | 0.039 |
| 2 | Commelina diffusa | Leptophobia aripa | 0.007 | 0.01 |
| 2 | Canna indica | Leptophobia aripa | 0.0001 | 0.049 |
| Junonia coenia | 0.0020 | 0.286 | ||
| Vanessa anabella | 0.0010 | 0.303 | ||
| 3 | Chloris gayana | Biblis hyperia | 0.001 | 0.561 |
| 3 | Bromus carinatus | Smyrna blomfildia | 0.003 | 0.201 |
| 3 | Ixophorus unisetus | Smyrna blomfildia | 0.003 | 0.194 |
Table 4.
Correlation analysis between plant species and migratory butterfly species in the Medicago sativa crop.
Table 4.
Correlation analysis between plant species and migratory butterfly species in the Medicago sativa crop.
| Plant species | Migratory butterfly species | p | r2 |
|---|---|---|---|
| Erythrina coralloides | Eurema daira | 0.0009 | 0.231 |
| Ascia monuste | 0.0001 | 0.420 | |
| Aldama dentata | Eurema proterpia | 0.0275 | 0.070 |
| Medicago lupulina | Eurema proterpia | 0.0001 | 0.570 |
| Sanvitalia procumbens | Eurema proterpia | 0.0018 | 0.362 |
| Salvia longistyla | Ascia monuste | 0.0001 | 0.503 |
| Leonotis nepetifolia | Ascia monuste | 0.007 | 0.087 |
| Alternanthera sp. | Danaus plexippus | 0.027 | 0.194 |
| Chloris gayana | Smyrna blomfildia | 0.001 | 0.598 |
| Bromus carinatus | Smyrna blomfildia | 0.003 | 0.201 |
| Ixophorus unisetus | Smyrna blomfildia | 0.003 | 0.194 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



