
Submitted:
16 June 2023
Posted:
20 June 2023
You are already at the latest version
Abstract
The effect of legumes adapted to a temperate climate on gas production kinetics, enteric methane, and rumen fermentation in vitro was evaluated. The species studied were Acacia dealbatha, Acacia melanoxylon, Albizzia lopantha, Lupinus pubescens, Inga insignes, Senna multiglandulosa and Tecoma stans. The effective degradation of the dry matter was estimated, the in vitro gas production was measured up to 120 hours of incubation, and the production of enteric methane, VFAs, and ammonia were measured at 24 hours. Dry matter, organic matter, ash, crude protein, neutral detergent fiber, acid detergent fiber, and acid detergent lignin were evaluated. The content of total phenols and condensed tannins was quantified. To determine digestibility, the samples were incubated for up to 120 hours. I. insignes presented a higher dry matter content and a high crude protein content. L. pubescens presented low dry matter digestibility. The neutral detergent fiber content was higher in I. insignes and with a higher value in dry matter digestibility and lower content of total phenols and condensed tannins. While S. multiglandulosa showed low detergent acid fiber content. The acid detergent lignin value was higher in I. insignes and lower in L. pubescens. In vitro gas and enteric methane production was higher in L. pubescens. propionic acid and ammonia they were higher in S. multiglandulosa. It is concluded that the amount of secondary compounds affects methane production, compromises digestibility and rumen kinetics of the legumes studied.
Keywords:
Acacia dealbatha
; Acacia melanoxylon
; Albizzia lopantha
; Lupinus pubescens
; Inga insignes
; Senna multiglandulosa
INTRODUCTION
Greenhouse gas (GHG) emissions from ruminants contribute to global warming and climate change [1], and represent between 7 and 18 percent of total GHG emissions. worldwide of anthropogenic origin [2,3]. Among the main GHGs identified by international organizations [4] are methane (CH4), carbon dioxide (CO2), nitrous oxide (N2O), Hydrofluorocarbons (HFC), Perfluorocarbons (PFC), and Sulfur Hexafluoride (SF6).
Methane (CH4) is one of the main gases that is released as the end product of rumen methane genesis of species linked to animal production. Daily methane production values in B. taurus x B. indicus cross heifers of 88.08 g/heifer/day (18.9 min –150.1 max) [5]. In sheep, methane production was 13.7 g/sheep/day [6].
In this sense, several works have been carried out in search of reducing greenhouse gas emissions, mainly enteric methane [7,8,9], however it should be considered that in ruminants there are several factors that affect enteric methane production, such as the type of microbial population in the rumen, rumen pH, acetate: propionate ratio, animal breed, dry matter intake, diet composition, management practices, stress environmental, among others [6].
Therefore, strategies such as population modification of ruminant animals, biotechnological processes to modify methanogenic microorganisms, and nutritional manipulation have been used to mitigate enteric methane production and alter the different causes related to enteric fermentation [1].
With regard to nutritional manipulation, work carried out with plant species rich in secondary metabolites such as Acacia. most anguished, Sesbania. sesban, Leucaena leucocephala, Pennisetum.purpureum and Acacia. mearnsii have been used for their potential to reduce the production of enteric methane [10,11,12], which due to its tannin and saponin content have demonstrated their effect on the methanogenesis, the defaunation of methanogenic microorganisms and the non-inhibition of dietary protein synthesis.
The availability of tree legumes in each country and their use associated with traditional grazing [13], would prove to be a model to follow as an alternative system in ruminant feeding to contribute to the reduction of enteric methane emissions into the environment. atmosphere.
These findings support the objective of studying the rumen modulation capacity by tree legumes, considering their species, climate, content of secondary metabolites and their effect on rumen function.
MATERIALS AND METHODS
Animals
Four rambouillet sheep were used (66.6± 0.75 kg body weight; 4 years of age) with permanent rumen cannula were used as donors of rumen contents for in vitro incubations. The animals were cared for and handled by trained personnel in accordance with the Spanish guidelines for animal protection in accordance with European regulations, and the experimental procedures were approved by the Animal Experimentation Ethics Committee of the Autonomous Community of Madrid (Authorization number PROEX 035/17). The animals were fed a mixed diet consisting of 600 g of oat hay and 400 g of a commercial concentrate per kg (fresh matter basis). The concentrate consisted (g/kg, fresh matter basis) of 329 g barley grain, 328 g corn grain, 244 g wheat, 62 g soybean meal, 28 g CO3Ca, 6 g salt and 2 g of mineral-vitamin mixture. The mixed diet contained 112, 350, and 166 g crude protein (CP), neutral detergent fiber (NDF), and acid detergent fiber (ADF) per kg DM, respectively, and was offered at 50 g DM/kg metabolic weight. (PV 0.75) in two equal portions at 09:00 and 17:00. The consumption of water was at will.
Collection of samples.
Seven tree legumes from the temperate climate of Ecuador located between 2500 - 2800 m.a.s.l were used. The samples collected were from Acacia dealbatha, Acacia melanoxylon, Albizzia lopantha, Lupinus pubescens, Inga insignes, Senna multiglandulosa and Tecoma stans. Each collected sample corresponded to the green leaves of plant, approximately 5 kg of green leaves were collected, the harvest was carried out during the months of February and March corresponding to the rainy season. The relative humidity of the sector is 55% - 60% and the average annual precipitation is approximately 1390 mm. Subsequently, the leaves were dried in an oven at 60 °C for 72 h. The particle size was reduced using a mill with a 1 mm sieve. Finally, the samples were stored for their subsequent chemical analysis and in vitro tests.
Chemical analysis
The content of dry matter (ID 934.01), ash (ID 942.05) and nitrogen (ID 990.03) were analyzed according to [14] AOAC (1999). Neutral Detergent Fiber (NDF) and Acid Detergent Fiber (ADF) were measured sequentially using the same sample in filter bags and expressed without including residual ash according to [15] Van Soest et al. (1991), using an ANKOM220 fiber analyzer (ANKOM Technology Corporation, Fairport, NY, USA). The NDF was determined using a thermostable amylase. Acid detergent lignin (ADL) was determined by solubilization of cellulose with sulfuric acid (72%) [15]. Total phenols (FT) were determined with the Folin-Ciocalteau method [16], and tannic acid as standard and condensed tannins (TC) were measured by the hydrochloric acid vanillin method [17], using catechin as standard, using a spectrophotometer Hitachi U2000 UV VIS
N-NH3 concentrations were determined using an Epoch microplate spectrophotometer (BioTek Instruments Inc., Winooski, VT, USA) by the phenol-hypochlorite method [18]. The production of volatile fatty acids (VFA) was determined by gas chromatography [19]. The methane analysis was carried out using a gas chromatograph [20], (Shimadzu GC 14B; Shimadzu Europa GmbH, Duisburg, Germany) equipped with a flame ionization detector and a column loaded with Carboxen 1000 (Supelco, Madrid, Spain).
In vitro incubations
in vitro incubations was obtained from four ewes immediately before morning feeding. The incubations were carried out using the inoculum from each sheep separately, to obtain four replicates per treatment. Glass vials (60 mL) were used in which 200 mg of dry matter from each of the samples were weighed. The rumen contents extracted from each animal were filtered through four layers of gauze and immediately transferred to the laboratory. The ruminal fluid was mixed with a culture medium [21] in a 1:4 (vol/vol) ratio at 39°C under continuous gassing with CO2, dosing 20 mL of the mixture in each vial by a peristaltic pump (Watson-Marlow 520UIP31). The vials were sealed and incubated at 39°C for 120 hours. Additionally, vials without substrate (two per inoculum) were included to correct the gas production values of endogenous substrates. Gas production was measured in each vial at 3, 6, 9, 12, 14, 22, 26, 30, 34, 48, 58, 72, 96, and 120 hours using a pressure transducer (Delta Ohm DTP704-2BGI; Herter Instruments SL, Barcelona, Spain) the produced gas was released after each measurement. The gas production data measured in each vial at each sampling time were adjusted to the model: mL gas= PG(1+(B/t) c)-1 [22], where PG: total gas production; B: gas asymptote and c: gas production rate.
On the other hand, in the vials incubated at 24 h, the gas produced was measured and a sample (10 mL) was taken in a vacuum tube (Terumo Europe NV, Leuven, Belgium) for methane analysis. The bottles were uncapped and the pH was immediately measured with a pH meter (Crison Basic 20; Crison Instruments, Barcelona, Spain). For AGV analysis, three milliliters of the contents of the vials were taken at the same time and 3 mL of deproteinizing solution (20 g of metaphosphoric acid and 0.6 g of crotonic acid per liter) was added and to determine N-NH3 used 2 mL of content plus 2 mL of HCl 0.5 mol L -1. The apparent in vitro digestibility of dry matter (IVDMS) was measured using a DAYSI II equipment (ANKOM Technology Corporation, Fairport, NY, USA) with samples that were incubated for up to 120 hours.
Statistical calculations and analysis
To estimate the kinetic parameters of fermentation (unpublished data except effective dry matter degradation) the gas production data were fitted to the exponential model: gas = A {1-exp [- c ( t -lag)]}, where A is the gas production asymptote, c is the fractional rate of gas production, lag is the initial delay in the start of gas production, and t is the gas measurement time. Parameters A , c and lag were estimated by an iterative least squares procedure using a SAS NLIN process (version 9.2, SAS Institute, Cary, NC, USA) [23]. The mean gas production time (T 1/2 ) was the time (h) when half the volume of asymptotic gas (A; mL) was produced and was calculated as T 1/2 = [(ln2 / c) + lag]. The average gas production rate (AGPR; mL gas h -1 ) was defined as the average gas production rate between the start of incubation and T 1/2 , and was calculated as AGPR = Ac/[2 (ln2 + c × lag)]. Finally, the effective degradability of dry matter (DEMS) was estimated assuming a passage rate ( Kp ) of 0.04 per h according to the equation: DEMS = [(IVDMS 120 × c ) / (c + Kp )] e ( -c × lag) .
The results obtained were analyzed under a completely randomized design (DCA) using the SAS statistical package (version 9.2, SAS Institute, Cary, NC, USA). In vitro gas production curves were obtained using Prism 4 software, Graphpad Software, Inc. of San Diego, CA, USA. The comparison of means was carried out using Tukey's test at 5%. The relationship between the content of secondary compounds and the gas and methane production parameters were evaluated by means of a simple correlation analysis using the PROC CORR procedure of the SAS statistical package (version 9.2, SAS Institute, Cary, NC, USA).
RESULTS
Chemical composition
Table 1 reports the chemical composition of the legumes analyzed, where it is observed that I. insignis presented the highest dry matter content (497.00 g/kg -1) with respect to the other species. The range of crude protein content in all species was variable and was between 134.66 - 238.45 g/kg -1 DM, however, in T. stans the 53.28% of its crude protein was bound to the insoluble cell wall (detergent acid fiber). The neutral detergent fiber content was much higher in I. insignes with 647.65 g/kg -1 DM, while S. multiglandulosa and L. pubescens had a lower content of acid detergent fiber (198.25 g/kg -1 DM) and lignin. detergent acid (72.29 g/kg -1 DM) respectively.
On the other hand, the lowest content of total phenols was found in T. stans (1.19%), finally L. pubescens presented 0.42% of condensed tannins, a lower value compared to the other species.
In vitro gas production and enteric methane
I. insignes presented a statistically different lower gas production (p=0.0001) when compared with the other species subjected to in vitro fermentation (Table 2). The gas production asymptote and the fermentation rate were the same or higher in all species evaluated. In Figure 1 distinguishes differentiated groups with higher and lower gas production.
While the species that presented the greatest potential in the reduction of enteric methane produced were I. insignes and A. melanoxylon (1.09 and 1.35 mL CH4/g DM -1 respectively). Additionally, a significant variation (p=0.0001) was evidenced in the pH of the inocula at 24 hours of fermentation in a range of 6.82 - 7.25
Volatile fatty acid profile and rumen function
Table 3 showed the molar proportion of volatile fatty acids (%), where S. multiglandulosa presented 61.69, 29.51 and 4.72 for acetate, propionate and butyrate respectively, percentages that differed significantly (p=0.0001) compared to the other species. Similarly, the highest concentration of N-NH3 was for S. multiglandulosa. However, the highest DEMS and DIVMS was for I. insignes with 381.28 and 791.46 g/kg respectively.
The correlation analysis (Table 4) indicates a strong associativity between the content of total phenols, condensed tannins and the production of total gas (r= -0.664, r= -0.813) and enteric methane (r= -0.602, r= -0.796). A direct relationship was observed between the content of secondary compounds and effective degradation of dry matter.
DISCUSSION
The dry matter content is related to the availability of nutrients in the plant [24], in this sense I. insignis, A. dealbatha, A. melanoxylon and A. lophanta presented amounts close to 500 g/kg MS -1 which would apparently reflect a higher biomass available for the animal compared to the other species studied.
However, when analyzing the nutritional components, it was observed that in species such as T. stans and I. insignes, despite their acceptable levels of crude protein, 53.28 and 40.43% of that protein, respectively, were bound to detergent acid fiber, thus reducing the availability of protein for its degradation at the ruminal level.
In the other hand, if we consider the fibrous components of plants as predictors of dry matter intake [25], indicates that species with high lignin content (LAD) such as I. insignes, A. dealbatha and A. melanoxylon would reduce consumption levels by animals and, on the contrary, it has been shown that there is a relationship between consumption preference in animals and NDF content (r = 0.737; P < 0.0001).
The content of secondary compounds (total phenols and condensed tannins) was lower than that reported in other studies in tropical legumes [26,27,28] due to their different metabolic pathways (C 3 and C 4) used for photorespiration [29].
Regarding the amount of phenols found in this research, they would not represent any limitation on fermentation parameters, because at the rumen level non-tannic phenols do not have the affinity to bind with proteins and other nutrients [16]. Meanwhile, tannins have a different mechanism of action, being the most effective hydrolysables in reducing methanogenesis by having a direct effect on rumen microorganisms, inhibiting their growth. In this investigation, the amount of condensed tannins (between 0.42 - 3.45%) was determined. These low toxicity compounds [30], have an indirect effect, but more evident at the rumen level, exerting their action on nutrients, decreasing digestibility, gas and methane production [31]. In addition, those shrubs that have a higher content of condensed tannins and protein would be related to a greater availability for the formation of tannin-protein complexes [25], as was the case of I. insignes in this research.
However, it has also been reported [32,33] the action of condensed tannins on ruminal microorganisms as a selective process, acting especially on the population of ruminal methanogens, cellulitic bacteria and protozoa associated with the methanogenesis, causing a decrease in the availability of hydrogen in the rumen, which may affect hydrolysis, nutrient degradation and microbial protein synthesis. Studies relates these effects based on the concentration and type of tannins in the forage and adaptation of the rumen population, qualifying the use of forages rich in tannins as beneficial [34].
The content of condensed tannins found in the tree legumes was significantly related (p<0.05) with the production of gas and enteric methane, expected results as detailed by studies [35], in this context, it was evidenced that as the concentration of condensed tannins increased, gas and methane production decreased. However, the methane reduction was more marked when it was calculated per unit of nutrient (mL/CH4/g DM) compared to the total percentage of methane produced at 24 hours of incubation, behavior perhaps related to the type of tannins (condensed or hydrolyzable) as mentioned by studies [31], which also indicates that the greater the number of hydroxyl groups in the condensed tannins, the greater their potential to reduce methane production. In this sense, lower gas production may also be related to higher protein utilization and microbial protein synthesis [36].
Fermentation kinetics patterns (asymptote, fermentation rate) were also affected by condensed tannin levels, perhaps due to variability in carbohydrate (FAD) and lignin content present in plants [37]. On the other hand, in others studies [24] it was observed that the higher molar proportion of propionate was not related to a decrease in methane production, perhaps due more to the amount of condensed tannins present in the plants and their antimethanogenic activity rather than decrease fiber digestibility.
The effective degradation of dry matter was higher in those forages with higher content of condensed tannins, behavior similar to that reported in similar studies [38]. On the other hand, the condensed tannins decreased (not significantly) the concentration of N-NH3, a desirable characteristic up to a certain point according to others authors [39], in this sense, one investigation [38] mention that N-NH 3 is part of the nutrient supply for proteolytic and cellulolytic bacteria. Furthermore, [39] indicates that the presence of tannins could decrease the ammonia concentration, exhibiting the efficient use of volatile fatty acids (VFA) for the synthesis of microbial proteins.
The digestibility values were low in those legumes with a lower content of condensed tannins (S. multiglandulosa) due to a greater extent to the lignin content that could be related to low rates of particle size reduction and slower outflow rates affecting at the end to consumption [25].
CONCLUSION
The effects caused by secondary metabolites present in the legumes studied on enteric methane production and ruminal kinetics were different, these results being conditioned to the type and amount of secondary compounds present. On the other hand, it seems that the detrimental effects on digestibility would be related to the content of indigestible fibers rather than the content of condensed tannins.
Funding
The authors thank the financial support from the Research and Development Direction (DIDE, acronym in Spanish) of the Technical University of Ambato, Ecuador, project (PFCAGP19)
Acknowledgments
The authors thank Department of Agricultural Production, Higher Technical School of Agricultural, Food and Biosystems Engineering, Polytechnic University of Madrid for the infrastructure and laboratory facilities.
Conflicts of Interest
The authors declare we have no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bonilla-Cárdenas J and Lemus-Flores C (2012) Enteric methane emission by ruminants and its contribution to global climate change Review. Rev Mex Cienc Pecu 3(2):215-246.
- IPCC (Intergovernmental Panel on Climate Change) (2006). Chapter 10: Emissions resulting from livestock and manure management. In 2006 IPCC Guidelines for National Greenhouse Gas Inventories. Volume 4: Agriculture, forestry and other land uses. pp 10.1–10.
- FAO (Food and Agriculture Organization of the United Nations). (2013) Mitigation of greenhouse gas emissions in livestock production – A review of technical options for the reduction of non-CO2 gas emissions. no.177.
- Kyoto Protocol (1998) United Nations Framework Convention on climate change. United Nations.
- Ku-Vera JC, et al. (2018) Determination of methane yield in cattle fed tropical grasses as measured in open-circuit respiration chambers. Agricultural and Forest Meteorology. [CrossRef]
- Veerasamy S, Rattan L, Jeffrey L, Thaddeus E (2011) Measurement and prediction of enteric methane emission. Agricultural and Forest Meteorology.
- Animut G, Puchala R, Goetsch AL, Patra AK, Sahlu T, Varel VH, Wells J (2008) Methane emission by goats consuming diets with different levels of condensed tannins from lespedeza. Anim Feed Sci Technol 144:212–22.
- Hammond KJ, Crompton LA, Bannink A, Dijkstra J, Yáñez-Ruiz DR, O'Kiely P, Kebreab E, Eugène MA, Yu Z, Shingfield KJ, Schwarm A, Hristov AN, Reynolds CK (2016) Review of current in vivo Measurement techniques for quantifying enteric methane emissions from ruminants. Anim Feed Sci Technol 219:13–30.
- Vanegas JL, Carro MD, Alvir MR and González J (2016) Protection of sunflower seed and sunflower meal protein with malic acid and heat: effects on in vitro ruminal fermentation and methane production. J Sci Food Agriculture. [CrossRef]
- Perna Junior F, Cassiano ECO, Martins MF, Romero LAS, Zapata DCV, Pinedo LA, Marino CT, and Rodrigues PHM (2017) Effect of tannins-rich extract from Acacia mearnsii or monensin as feed additives on ruminal fermentation efficiency in cattle. Livestock Science. [CrossRef]
- Barros-Rodríguez MA, Solorio-Sánchez FJ, Sandoval-Castro CA, Ahmed AMM, Rojas-Herrera R, Briceño-Poot EG, Ku-Vera JC (2014) Effect of intake of diets containing tannins and saponins on in vitro gas production and sheep performance. Animal Production Science 54:1486-1489. https:// doi.org/10. [CrossRef]
- Soliva CR, Zeleke AB, Clément C, Hess HD, Fievez V and Kreuzer M (2008). In vitro screening of various tropical foliages, seeds, fruits and medicinal plants for low methane and high ammonia generating potentials in the rumen. Anim Feed Sci Technol.
- Muir JP, Pitman WD, Dubeux JC Jr, and Foster JL (2014) The future of warm season, tropical and subtropical forage legumes in sustainable pastures and rangelands. African Journal of Range & Forage Science, 31:3, 187-198. [CrossRef]
- AOAC (1999) Official Methods of Analysis, 16th edition, 5th revision.
- Van Soest PJ, Robertson JB and Lewis BA (1991) Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci 74:3583–3597.
- Makkar HPS (2003) Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Ruminant Res 49:241-256.
- Broadhurst RB and Jones WT (1978) Analysis of condensed tannins using acidified vanillin. J. Sci. J. Sci. Food Agric 29:788-794.
- Weatherburn MW (1967) Phenol–hypochlorite reaction for determination of ammonia. Anal Chem 39:971–974.
- Carro MD, Lebzien P and Rohr K (1992) Influence of yeast culture on the in vitro fermentation (Rusitec) of diets containing variable portions of concentrates. Anim Feed Sci Technol 37:209–220.
- Martínez ME, Ranilla MJ, Tejido ML, Ramos S and Carro MD (2010) The effect of the diet fed to donor sheep on in vitro methane production and ruminal fermentation of diets of variable composition. Anim Feed Sci Technol 158:126–135.
- Goering MK and Van Soest PJ (1970) Forage Fiber Analysis (Apparatus, Reagents, Procedures and Some Applications), Agricultural Handbook, n ∘ 379.
- Groot JCJ, Cone JW, Williams BA, Debersaques FMA and Lantinga EA (1996) Multiphasic analysis of gas production kinetics for in vitro fermentation of ruminant feeds. Anim Feed Sci Technol 64(1):77–89. [CrossRef]
- Molina-Alcaide E, Carro MD, Rodela MY, Weisbjerg MR, Lind V, Novoa-Garrido M (2017) In vitro rumen fermentation and methane production of different seaweed species. Anim Feed Sci Technol 228(1):1-12.
- McDonald P, Edwards RA, Greenhalgh JFD and Morgan CA (2002) Animal Nutrition. 6th ed. (Prentice Hall: Harlow, England, UK).
- Hernández-Orduño G, Torres-Acosta JFJ, Sandoval-Castro CA, Capetillo-Leal CM, Aguilar-Caballero AJ, Alonso-Díaz MA (2015) A tannin-blocking agent does not modify the preference of sheep towards tannin-containing plants.
- Bhatta R, Saravanan, M, Baruah L and Sampath KT (2012) Nutrient content, in vitro ruminal fermentation characteristics and methane reduction potential of tropical tannin-containing leaves. J Sci Food Agric. [CrossRef]
- Soltan YA, Morsy AS, Sallam SMA, Louvandini H, and Abdalla AL (2012) Comparative in vitro evaluation of forage legumes (prosopis, acacia, atriplex, and leucaena) on rumen fermentation and methanogenesis. Journal of Animal and Feed Sciences, 21:759–772.
- Oduguwa B, Olusoji A, Arigbede OM, & Adesunbola JO, & Sudekum KH (2013) Feeding potential of cassava (Manihot esculenta Crantz) peles ensiled with Leucaena leucocephala and Gliricidia sepium assessed with West African dwarf goats. Trop Anim Health Prod 45:1363–368.
- Archiméde H, Eugène M, Magdeleinea CM, Boval M, Martin C, Morgavib DP, Lecomtec P, Doreaub M (2011) Comparison of methane production between C3 and C4 grasses and legumes. Anim Feed Sci Technol 166–167: 59–64.
- Beauchemin KA, Kreuzer M, O'Mara F and McAllister TA (2008) Nutritional management for enteric methane abatement: a review, Australian Journal of Experimental Agriculture 48:21–27.
- Gunjan G and Makkar HPS (2012) Methane mitigation from ruminants using tannins and saponins Trop Anim Health Prod 44:729–739.
- Tavendale MH, Meagher LP, Pacheco D, Walker N, Attwood GT and Sivakumaran S (2005) Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis, Anim Feed Sci Technol, 123 –124:403–419.
- Jayanegara A, Leiber F and Kreuzer M (2011) Meta-analysis of the relationship between dietary tannin level and methane formation in ruminants from in vivo and in vitro experiments, Journal of Animal physiology and Animal nutrition. [CrossRef]
- McSweeney CS, Palmer B, McNeil DM, et al., 2001. Microbial interactions with tannins: nutritional consequences for ruminants, Anim Feed Sci Technol, 91, 83–93.
- Pellikaan WF, Stringano E, Leenaars J, Bongers D, Van Laar-van Schuppen S, Plant J, Mueller-Harvey I (2011) Evaluating effects of tannins on extent and rate of in vitro gas and CH4 production using an automated pressure evaluation system (APES). Anim Feed Sci Technol 166–167: 377–390.
- Blümmel, M, Makkar HPS, Becker K (1997) In vitro gas production: A technique revisited Journal of Animal Physiology and Animal Nutrition 77 (1):24-34.
- Galindo J, Marrero Y, Ruiz TE, González N, Díaz A, Aldama AI, Moreira O, Hernández JL, Torres V and Sarduy L (2009) Effect of a multiple mixture of herbaceous legumes and Leucaena leucocephala on the microbial population and fermentative products in the rumen of Zebu upgraded yearling steers. Cuban Journal of Agricultural Science. 43:251-257.
- Barros-Rodríguez MA, Solorio-Sánchez FJ, Sandoval-Castro CA, Klieve A, Rojas-Herrera RA, Briceño-Poot EG and Ku-Vera JC (2015) Rumen function in vivo and in vitro in sheep fed Leucaena leucocephala Trop Anim Health Prod. [CrossRef]
- Bhatta R, Saravanan M, Baruah L and Prasad CS (2015) Effects of graded levels of tannin-containing tropical tree leaves on in vitro rumen fermentation, total protozoa and methane production..Journal of applied microbiology 118:557-564. [CrossRef]
Figure 1.
Gas production curves of tree legumes subjected to different incubation times 1: A. dealbatha; 2: A. melanoxylon; 3: A. lopantha; 4: L. pubescens; 5: I. insignes; 6: S. multiglandulosa; 7: T. stans.
Figure 1.
Gas production curves of tree legumes subjected to different incubation times 1: A. dealbatha; 2: A. melanoxylon; 3: A. lopantha; 4: L. pubescens; 5: I. insignes; 6: S. multiglandulosa; 7: T. stans.
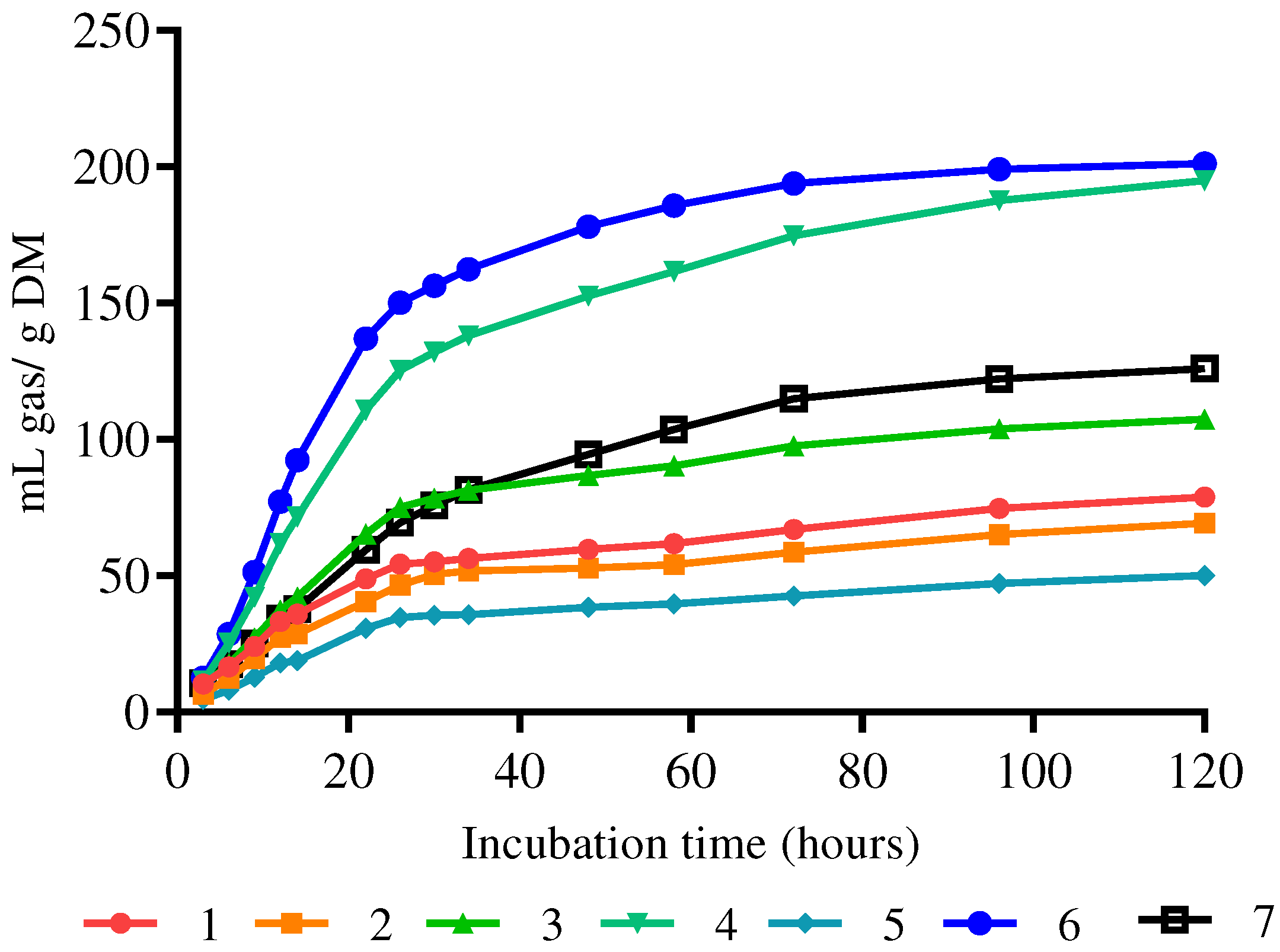

Table 1.
Chemical composition of different tree legumes from temperate climates expressed as g/kg -1 of dry matter (except where otherwise indicated).
Table 1.
Chemical composition of different tree legumes from temperate climates expressed as g/kg -1 of dry matter (except where otherwise indicated).
| Item | Acacia dealbatha | Acacia melanoxylon | Albizzia lophanta | Lupinus pubescens | Inga insignis |
Senna multi glandular |
Tecoma stans |
|---|---|---|---|---|---|---|---|
| MS 1 | 484.00 | 484.74 | 457.23 | 245.64 | 497.00 | 336.64 | 346.66 |
| MO | 933.12 | 934.92 | 934.94 | 910.56 | 908.00 | 846.75 | 926.63 |
| ASHES | 66.93 | 65.18 | 65.13 | 89.58 | 92.00 | 153.38 | 73.46 |
| CP | 162.32 | 134.66 | 141.87 | 238.45 | 176.85 | 162.95 | 165.37 |
| PC-FAD(%) | 34.86 | 32.79 | 21.98 | 4.13 | 40.43 | 4.64 | 53.28 |
| NDF | 519.88 | 573.45 | 329 | 402.94 | 647.65 | 275.92 | 492.68 |
| FAD | 408.64 | 406.90 | 242.33 | 221.15 | 518.86 | 198.25 | 368.00 |
| LAD | 285.00 | 252.25 | 145.54 | 72.29 | 311.29 | 117.12 | 229.47 |
| LAD/FAD(%) | 69.74 | 61.99 | 60.05 | 32.77 | 59.99 | 59.19 | 62.43 |
| TF2 ( %) | 2.32 | 1.95 | 2.94 | 1.73 | 3.76 | 1.69 | 1.19 |
| CT 3 (%) | 1.24 | 1.48 | 2.18 | 0.42 | 3.45 | 0.35 | 1.15 |
DM: dry matter; 1 g/kg of fresh matter; OM: organic matter; PC: crude protein; PC-FAD: crude protein bound to detergent acid fiber.; FND: ash-free neutral detergent fiber; FAD: ash-free acid detergent fiber; LAD: lignin acid free detergent ash; LAD/FAD: lignin in relation to acid detergent fiber; 2 TF: total phenols using tannic acid as standard; 3 CT: condensed tannins using catechin as standard.
Table 2.
Gas production parameters and enteric methane of temperate climate tree legumes.
| Gas production | Methane production | ||||||
|---|---|---|---|---|---|---|---|
| Species | GP | B. | c | pH | mL CH 4 /g DM 24h |
% CH 4 total 24h | |
| A .dealbatha | 84.3 d | 19.5 ab | 1.220 b | 7.23 a | 1.76 bc | 21.06 | |
| A. melanoxylon | 70.4 ed | 17.3b | 1.390 b | 7.25 a | 1.35 c | 15.56 | |
| A. lopantha | 110.4 c | 17.9 ab | 1.540ab | 7.07 b | 1.86 bc | 15.03 | |
| L. pubescens | 201.9 a | 20.3 ab | 1.540ab | 6.87 c | 4.54 a | 21.54 | |
| I. insignes | 50.7 e | 18.0 ab | 1.467ab | 7.29 a | 1.09 c | 15.98 | |
| S. multiglandulosa | 204.4 a | 15.6 b | 1.856 a | 6.82 c | 3.61 ba | 15.61 | |
| T. stans | 149.3 b | 30.5 a | 1.322 b | 7.02 b | 2.56 abc | 18.37 | |
| EEM | 5.51 | 2.79 | 0.0949 | 0.0246 | 0.450 | 2.178 | |
| Value P | 0.0001 | 0.0243 | 0.0039 | 0.0001 | 0.0002 | 0.1982 | |
GP: total gas production (mL/g DM); B: gas asymptote (mL/g DM); and c: gas production rate (%/hour). abc Values followed by different letter between rows differ significantly (p <0.005)
Table 3.
Volatile fatty acids (mol/100mol), ammoniacal nitrogen (N-NH3 mg/L), effective degradation of dry matter (EDDM g/kg -1) and in vitro digestibility of dry matter (IVDDM g/kg - 1) of temperate climate tree legumes.
Table 3.
Volatile fatty acids (mol/100mol), ammoniacal nitrogen (N-NH3 mg/L), effective degradation of dry matter (EDDM g/kg -1) and in vitro digestibility of dry matter (IVDDM g/kg - 1) of temperate climate tree legumes.
|
A. dealbatha |
A. melanoxylon |
A. lopantha |
L. pubescens |
I. insignes |
S. multi glandulosa |
T. stans |
EEM | ValueP | |
|---|---|---|---|---|---|---|---|---|---|
| Acetate | 69.77 b | 71.74 b | 70.99 b | 70.20 b | 71.36 b | 61.69 c | 75.65 a | 0.771 | 0.0001 |
| Propionate | 20.63 b | 21.53b | 22.77 b | 21.87 b | 17.45 c | 29.51 a | 16.36 c | 0.491 | 0.0001 |
| butyrate | 6.25 ba | 4.31c | 4.37 c | 5.37 bc | 7.40 a | 4.72 bc | 5.73 bc | 0.353 | 0.0001 |
| isobutyrate | 0.82 a | 0.64 a | 0.46 a | 0.60 a | 0.93 a | 0.80 a | 0.66 a | 0.116 | 0.1296 |
| isovalerate | 1.32 ba | 0.98 ba | 0.69 b | 1.11 ba | 1.65 a | 1.32 ba | 0.90 b | 0.147 | 0.0036 |
| valerate | 1.18 b | 0.80 cb | 0.70 c | 0.84 cb | 1.19 b | 1.65 a | 0.67 c | 0.098 | 0.0001 |
| A/P | 3.38 b | 3.33 b | 3.11 b | 3.20 b | 4.12 a | 2.11 c | 4.64 a | 0.117 | 0.0001 |
| N-NH3 | 133.65 | 125.12 | 115.70 | 128.53 | 161.15 | 211.53 | 109.88 | 26.199 | 0.1459 |
| EDDM4% | 339.24 ab | 363.61 a | 309.58 ab | 173.18 c | 381.28a | 232.16 bc | 272.63abc | 2.637 | 0.0001 |
| IVDDM | 748.57 ab | 686.58 ab | 661.44 b | 477.05 c | 791.46a | 469.91 c | 698.82 ab | 2.806 | 0.0001 |
abc Values followed by a different letter between columns are significantly different (p <0.05).
Table 4.
Parameters of Pearson correlation between the content of secondary compounds and the production of gas, methane and ruminal function of tree legumes from temperate climates.
Table 4.
Parameters of Pearson correlation between the content of secondary compounds and the production of gas, methane and ruminal function of tree legumes from temperate climates.
| Gas production |
methane production |
ruminal function | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| pH | N-NH3 | EDDM 4% | ||||||||
| r | P | r | P | r | P | r | P | r | P | |
| TF | -0.664 | 0.104 | -0.602 | 0.153 | 0.051 | 0.913 | 0.097 | 0.836 | 0.616 | 0.141 |
| TC | -0.813 | 0.026 | -0.796 | 0.032 | -0.093 | 0.843 | -0.143 | 0.760 | 0.781 | 0.038 |
FT: total phenols; TC: condensed tannins.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



