
Submitted:
16 June 2023
Posted:
19 June 2023
You are already at the latest version
Abstract
The genetic diversity of local coffee populations is crucial to breed new varieties better adapted to the increasingly stressful environment due to climate change and the evolving consumer preferences. Unfortunately, local coffee germplasm conservation and genetic assessment have not received much attention. Molecular tools offer substantial benefits in identifying and selecting new cultivars or clones suitable for sustainable commercial utilization. New annotation methods, such as chloroplast barcoding, are necessary to produce accurate and high-quality phylogenetic analyses. This study used DNA barcoding techniques to examine the genetic relationships among fifty-six accessions collected from the southern part of Saudi Arabia. PCR amplification and sequence characterization were used to investigate the effectiveness of four barcoding loci, namely atpB-rbcl, TrnL-TrnF, TrnT-TrnL, and TrnL. The maximum nucleotide sites, nucleotide diversity, and an average number of nucleotide differences were recorded for atpB-rbcl, while TrnT-TrnL had the highest variable polymorphic sites, segregating sites, and haploid diversity. Among the four barcode loci, TrnT-TrnL recorded the highest singleton variable sites, while TrnL recorded the highest parsimony information sites. Furthermore, the phylogenetic analysis clustered the Coffea arabica genotypes into four different groups, with three genotypes (KSA31, KSA38, and KSA46) found to be the most divergent genotypes standing alone in the cluster and remained apart during the analysis. The study demonstrates the presence of considerable diversity among coffee populations in Saudi Arabia. Furthermore, it also shows that DNA barcoding is an effective technique for identifying local coffee genotypes, with potential applications in coffee conservation and breeding efforts.
Keywords:
Coffea arabica
; chloroplastic DNA
; barcode
; genetic diversity
; population structure
; Saudi Arabia
1. Introduction:
Coffee is one of the most commercially significant crops, and the second most traded commodity after oil [1]. In addition to its high export value, coffee has also gained in cultural significance over the past few decades. Despite there being more than 125 reported species in the genus Coffea, only two species, Coffea arabica L. (also known as Arabica coffee) and C. canephora Pierre ex A. Froehner (known as Robusta coffee) are grown commercially [2]. The total annual global coffee production in 2020 was over 105.21 million tons, with 63.15 million tons of Arabica coffee accounting for 60.02% of the total, and 42.06 million tons of Robusta coffee comprising 39.97%. Coffee's genetic development is progressing at a sluggish pace despite its enormous economic relevance. The collection, characterization, and wise use of accessible germplasm material for any crop plant species contribute to its genetic development and long-term viability [3]. Therefore, enhancing diversity from both local and foreign sources is critical for the improvement of crops [4]. For historic reasons, the main issue with Arabica coffee has been its narrow genetic base that limits its adaptation to changing environments [2]. To get around this problem, breeders made use of wild coffee diploid species to introduce new genes into Arabica genotypes [2]. For instance, the leaf rust resistant Arabica cultivar Timor Hybrid got its resistance from its C. canephora parent; it was later used as a parent to develop several new rust resistant cultivars such as Catimor and Ruiru 11 [5]. For bean and liquor quality traits, the wild tetraploid Arabica genotypes from the species’ center of origin and the little-known ancient varieties from the Arabian Peninsula offer a wide gene pool to explore [6]. Despite the potential importance of coffee heirlooms in the Arabian Peninsula as a source of genetic diversity, there is limited information available on these genotypes. This information is essential for the development of new coffee varieties that can better adapt to changing environmental conditions, such as pest and disease pressure, abiotic stress, and changes in consumer preferences [7]. Furthermore, since over 60% of wild coffee species are in danger of extinction due to accelerated environmental change, gathering complete information, and characterizing this germplasm is of utmost importance [8]
Another issue facing the coffee industry as it struggles to cope with an over-supplied market, is adulteration. It has long been known that coffee is often adulterated with less expensive and readily available plant material [9]. Coffee adulteration has become a more serious issue for the industry in recent years due to the significant expansion in the variety of coffee recipes, stores, and ultimately consumers [10]. Therefore, developing molecular means like genetic barcodes to identify and authenticate the varieties can help mitigate the problem.
In Saudi Arabia and Yemen, C. arabica has been cultivated for at least four centuries on the terraced slopes and narrow valleys of the western mountains at different altitudes ranging mostly from 1200 to 2000m above sea level [11,12]. Most of what is grown now in southwestern Saudi Arabia are old cultivars that have been around for hundreds of years [12]. It is likely that these diverse populations are a result of successive introductions of genetic material from Eastern Ethiopia by Arab traders over centuries of uninterrupted exchange across the narrow strait of Bab El-Mandeb [13]. Therefore, it is safe to assume that the southwestern corner of the Arabian Peninsula contains the most genetic diversity of C. arabica outside the species' center of origin in the Ethiopian highlands [6]. Unfortunately, these genetic resources have not attracted enough attention from the scientific community, except for the 1989 FAO expedition to southern Yemen led by A. Eskes [14] and two recent studies by Tounekti et al. [12] and Montagnon et al. [6]. These three studies reported the existence of considerable diversity among coffee populations in the Arabian Peninsula. It is worth noticing that the present coffee populations have evolved over hundreds of years in a semi-arid environment [15] marked by recurring droughts, uneven distribution of rainfall, heat stress and high irradiance. Therefore, it is expected that these genotypes could be the source of interesting genes that confer stress tolerance [16].
The molecular phylogenies of coffee species have been established using variations in intergenic spacer sequences [17-19] and introns [20] of plastid DNA, as well as internal transcribed spacer (ITS) sequences of rDNA [21] and a combination of four plastid regions and ITS [22]. Chloroplast DNA (cpDNA) sequence variation is widely used in systematics and for making phylogenetic inferences at different taxonomic levels. Introns and intergenic spacers are known to exhibit high rates of mutation [23]. The trnT-trnL and trnL-trnF intergenic spacers, the trnL intron, and the atpB-rbcL intergenic region are useful for evolutionary studies at low taxonomic levels, and these regions have been extensively used in phylogenetic studies to analyze cytoplasmic polymorphism and to study the demographic history of several species [23,24,25,26,27].
Overall, further research is necessary to fully comprehend the diversity and potential of diploid and tetraploid coffee species and to utilize this information to develop new coffee varieties that can better meet the needs of farmers and consumers in the future. The present study aims to estimate the genetic diversity of local coffee populations in Saudi Arabia and to examine their genetic relatedness using chloroplast intergenic spacer markers.
2. Results
The successful amplification of all four intergenic spacer barcode sequences (atpB-rbcl, TrnT-TrnL, TrnL-trnF, TrnL) was achieved, resulting in a single band of the expected size (see supplementary Figure). The respective sequences for each barcode were submitted to the National Center for Biotechnology Information (NCBI) via Bankit submission, and accessions numbers for each barcode sequence are presented in Table 2. This study utilized the Coffea arabica (MN894552.1) chloroplast genome sequence, available on NCBI, as a reference genome sequence to detect polymorphisms among DNA sequences in 56 Coffea arabica accessions across all four barcoding primers.
The percentage of polymorphic sites for each sequence was determined by dividing the number of variable nucleotides by the length of the entire region and multiplying the result by 100. Table 3 presents the number of nucleotide sites (NNS), variable polymorphic sites (VPS), number of segregating sites (NSS), number of haplotypes (NH), nucleotide diversity (ND), and average number of nucleotide differences (ANND) for each barcode primer and the cumulative results for all four primers. The combined sequences showed the highest NSS, followed by the atpB-rbcl primer, while the TrnL primer had the lowest NNS. The TrnT-TrnL primer had the highest VPS (341), followed by atpB-rbcl, while the lowest (154) was recorded for TrnL-TrnF. The atpB-rbcl had the highest ND, followed by TrnT-TrnL (0.051), with TrnL-TrnF showing the lowest ND. Additionally, the atpB-rbcl had the highest ANND (185.54), while TrnL-TrnF exhibited the lowest value (25.23) for ANND.
NNS=Number of nucleotide sites, VPS= variable polymorphic sites, NSS= number of segregating sites, NH= number of haplotypes, ND= Nucleotide diversity, ANND= average number of nucleotide difference.
The nucleotide base composition of each barcode primer was determined and is presented in Table 4. The average nucleotide base composition of atpB-rbcl was recorded as 33.15% T(U), 16.60% C, 34.49% A, and 15.76% G. For TrnC, the composition was 26.7% T(U), 15.9% C, 37.6% A, and 19.8% G. TrnT-TrnL had a composition of 39.18% T(U), 13.88% C, 33.84% A, and 13.10% G (Table 4). The singleton variable sites (STVS) and parsimony information sites (PIS) for each chloroplast barcode are presented in Table 5. The TrnT-TrnL barcode recorded the highest number of STVS (338), followed by TrnL-TrnF (133), TrnL (52), and atpB-rbcl (1) had the lowest. Similarly, for PIS, TrnL had the highest number (155), followed by atpB-rbcl (137), while the minimum was recorded for TrnL-TrnF (45) across all sequences (Table 5). A phylogenetic analysis was constructed based on concatenated sequences of all four barcodes using the maximum likelihood method and Kimura 2-parameters model. The percentage of trees in which the associated taxa clustered together is shown next to the branches. This analysis involved 56 nucleotide sequences, and the final dataset comprised a total of 4114 positions. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The final phylogenetic tree divided the 56 accessions into five different groups.
Figure 1.
Evolutionary analysis by Maximum Likelihood method using four barcodes with 1000 bootstraps constructed in MEGA 10.0 using the concatenated sequence of atpB-rbcl, TrnLc, TrnT-TrnL and TrnL-TrnF.
Figure 1.
Evolutionary analysis by Maximum Likelihood method using four barcodes with 1000 bootstraps constructed in MEGA 10.0 using the concatenated sequence of atpB-rbcl, TrnLc, TrnT-TrnL and TrnL-TrnF.
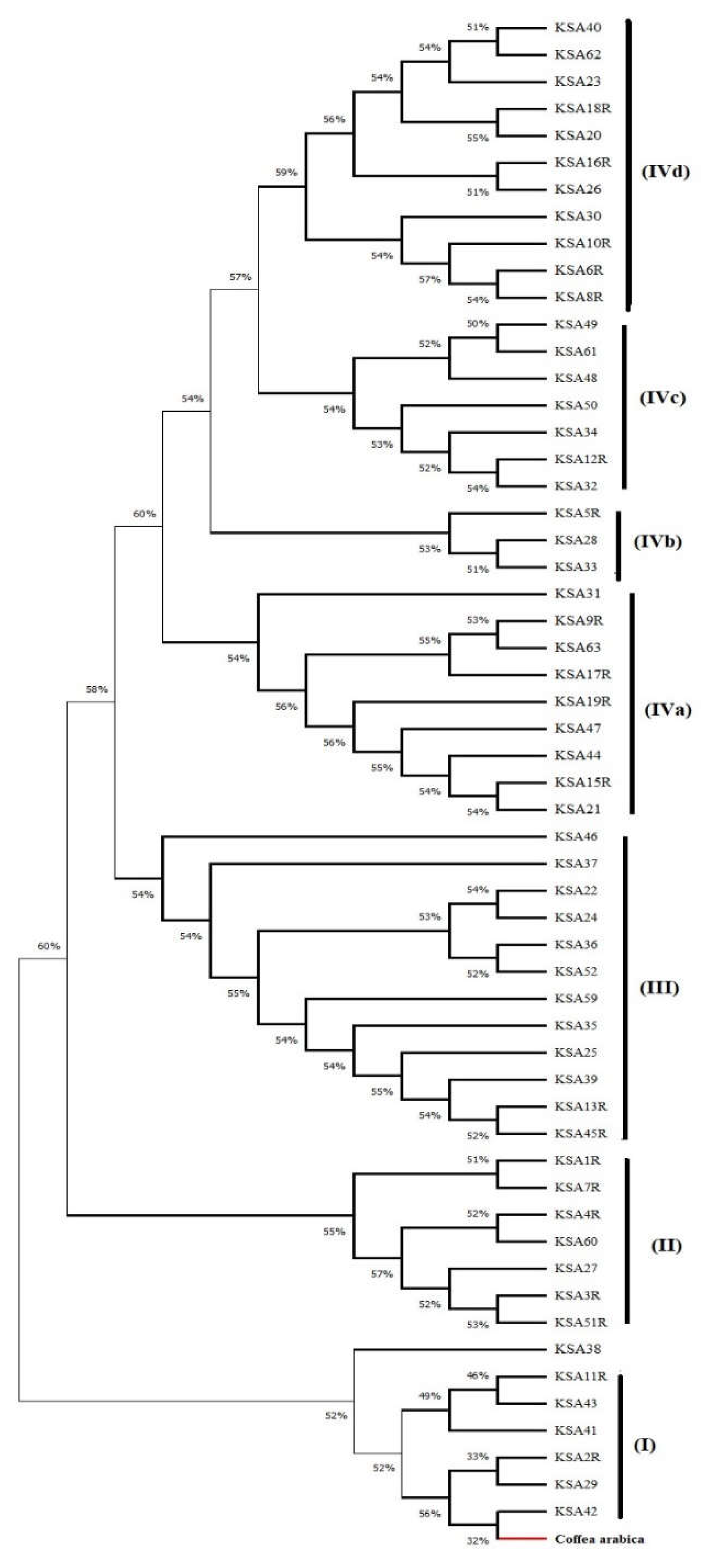
The phylogenetic analysis grouped the coffee accessions into four different groups. The first group contained six accessions (KSA42, KSA29, KSA2R, KSA41, KSA43, KSA11R, and KSA38) that were mostly from the Rayda district of Assir region. The second group contained seven accessions (KSA51R, KSA3R, KSA27, KSA60, KSA4R, KSA7R, and KSA1R), all from Jazan Region except KSA60 was from Assir. The third group contained 12 accessions (KSA45R, KSA13R, KSA39, KSA25, KSA35, KSA59, KSA52, KSA36, KSA24, KSA22, KSA37 and KSA46), all collected from Jazan Region except KSA59 from the north of Assir Region. The fourth and largest group contained 43 accessions that can be further subdivided into four subgroups. The first subgroup (IVa) was a diverse one and contained 12 accessions originating from the three regions. Subgroup IVb contained three accessions (KSA33, KSA28 and KSA5R) all from Jazan Region. Subgroup IVc contained seven accessions, six from Jazan and one from Al-Baha. Subgroup IVd contained 11 accessions, eight from Jazan region, two from Assir and one from Al-Baha.
3. Discussion:
The genetic diversity present in crop wild or primitive relatives (CWR) plays a crucial role in the effectiveness of crop improvement programs. These wild or unknown genotypes exit in diverse habitats, many of which are currently facing significant threats due to habitat degradation and climate change [8,14]. It is estimated that approximately 60% of wild coffee species are at risk of extinction worldwide; similarly, underutilized old varieties are disappearing from the orchards. This emphasizes the urgent need to conserve these species through both in situ and ex situ measures, which can help preserve their genetic diversity for future utilization.
While morphological descriptors are commonly used to characterize different coffee species, molecular markers are considered more efficient in distinguishing closely related species and cultivars [23], and more precise and reliable than morphological and biochemical markers [24]. Furthermore, several studies have demonstrated that specific regions of the chloroplast genome can serve as DNA barcodes for a wide variety of plant species [25,26]. Selection of suitable plastid genomes offer sufficient genetic information for distinguishing between genotypes. Additionally, when choosing suitable DNA barcoding loci, the variable regions should be given a primary consideration [27]. Therefore, the objective of this study was to evaluate the evolutionary and phylogenetic relationships among fifty-six Arabica coffee species by utilizing four DNA barcoding markers (atpB-rbcl, TrnL-TrnF, TrnL, and TrnT-TrnL). This research aimed to investigate the potential of four DNA barcode loci (specifically, atpB-rbcL, TrnL, TrnL-TrnF, and trnL-trnL from the chloroplast region) for the identification and provision of phylogenetic information on local Arabica coffee species. All four regions were successfully amplified using universal primers, yielding clear and reliable results. However, earlier studies have indicated that there were cases of partial amplification from the respective barcode loci's universal primers [28,29].
Similarly, other studies [30,31] have indicated that additional barcode primers, including matK, rbcL, and trnL-trnF, have demonstrated successful amplification within coffee species, which aligns with the findings of the present study. Despite the abundance of available data on DNA barcoding of angiosperms, there is currently limited information regarding specific barcodes that can guarantee an accurate species identification in all cases [32]. Often, a barcode that performs effectively for one group of plants may prove inadequate for another group, especially in the case of recently diverged species [33]. The current study successfully identified all fifty-six accessions as Coffea arabica, showcasing the effectiveness of the universal DNA barcode primers. Likewise, multiple studies have extensively documented the reliability of matK and rbcL, either individually or in combination, as DNA barcodes that can be used with confidence across various plant species [34]. Additional reports have recommended the utilization of rbcL as a valuable DNA barcode locus, primarily due to its relatively compact length of 500 bp, robust universal primer, high success rate in PCR amplification, and excellent sequencing quality [35]. However, other DNA barcodes, such as TrnL-TrnF and the TrnL spacer, have also been suggested as reliable alternative barcodes for identification of species [36]. The extent of sequence variation among the species or terminals under analysis is a crucial factor in determining the effectiveness of any barcoding locus [34]. Other reports have recommended the utilization of rbcL as a valuable DNA barcode locus due to its relatively compact length of 500 bp, robust universal primer, high success rate in PCR amplification, and excellent sequencing quality [25], along with other DNA barcodes such as TrnL-TrnF and the TrnL spacer [36]. The degree of sequence variation present among the species or terminals being analyzed is a determining factor in the efficacy of any barcoding loci [34].
The number of singleton variable sites was found to be higher in TrnLc, TrnL, and TrnL compared to atpB-rbcl. Similarly, TrnL and atpB-rbcl had more parsimony information sites than the rbcL barcode spacer region. These findings are consistent with a previous study conducted by [37], which reported that TrnL-trnF and matK barcodes exhibited greater variability than rbcl in Indian coffee arabica genotypes. The present study also found similar results for PIC among the four barcodes analyzed. Similarly, previous research has indicated that TrnL-TrnF and matK loci exhibit greater sequence polymorphism than rbcL, as suggested by [38,39], and [40]. The current study's results support these findings. Hence, the present study found that all four barcode sequences, which were evaluated as candidate barcode markers, met the DNA barcoding criteria outlined by [33]. Specifically, these markers exhibited sufficient sequence variability to enable effective discrimination among Saudi coffee genotypes. Similarly, [41] previously suggested that indels involving multiple residues may serve as useful diagnostic markers for species discrimination and phylogenetic analyses [42].
The phylogenetic analysis grouped the Saudi Coffea arabica genotypes into four groups with a clear influence of geographic origin suggesting the genotypes of each region share one or more common ancestor (Figure 2). For instance, accessions KSA11R, KSA41, KSA42 and KSA43 from the isolated Rayda district of Assir region were grouped in clusters I and II. The accessions representing very old trees (KSA36, KSA44, KSA46, KSA47) segregated in the middle of the phylogenic tree in groups III and IVa. Similar results were reported by [37] where the grouping using single and multilocus barcode primers was strongly influenced by the geographic origin of the genotypes. A molecular analysis of coffee arabica genotypes from Saudi Arabia using SRAP markers grouped them into five distinct groups based mostly on their origin [23]. The accessions collected from Jazan region primarily clustered in groups II and IV, whereas those from Al-Baha and Assir regions constituted a different group. Similar surveys of genetic diversity among coffee populations in northern Yemen [6] and southern Yemen [14] found that each district (valley) have its own cultivars. Another study using genotyping by sequencing (GBS) showed that genetic closeness correlated with geographic proximity [43].The current study provides further evidence to support this finding. It was also suggested chloroplast sequences provide high level effective and accurate information for plant evolution and high level preservation [44]. For future studies on this economically significant crop, we recommend using resequencing and genome-wide association studies (GWAS) to discover additional polymorphic markers associated with important agro-morphological traits. These markers would be beneficial for a range of investigations in Coffea crop. Ultimately, the polymorphic markers established and confirmed in this research hold potential as a valuable genomic asset for molecular breeding, identification, and biogeography studies of Arabica coffee.
4. Materials and Methods
4.1. Plant Material
A survey was carried out at several sites in the Sarawat mountain range, running parallel to the Red Sea from the southeast to the northwest through the three administrative regions of Jazan, Assir, and Al-Baha. The survey covered a strip of terraced mountains located between latitudes 17˚N and 20˚N, the most northern location where coffee is commercially grown in the world. The coffee gardens included in the survey were found at altitudes ranging from 900 to 2000 m a.s.l. In total, we collected young leaves from 56 accessions, from Jebel Fayfa (Fayfa district), Eddayer, Maadi (Haroub district), Jebel Al-Gahr (Al-Rayth district), Rayda valley (Assouda district in Assir region), Mahayel Assir district, Al-Majarda district and Jebel Shada (Al-Mekhwah district of Al-Baha region) (Table 1). We tagged and sampled 1-3 trees representing each tree population. Each accession was given a code starting with the acronym “KSA” (e.g., KSA-1), but, for the sake of simplicity, we dropped the acronym in the figures. The letter “R” was added to the code of accessions 1-19, 45, and 51 to indicate that they were sourced from a small, local coffee germplasm collection established in the Fayfa district.
4.2. DNA extraction
Plant material, consisting of young leaves from various C. arabica accessions, was collected from representative trees in each population and transported to the lab in a cooler. The leaves were sanitized by immersing them in a 5% sodium hypochlorite solution for 1-2 minutes and then rinsing them with sterile distilled water. The material was then ground in liquid nitrogen and stored in an -80ºC freezer. DNA was extracted from 100 mg of mixed powder using an innuPREP Plant DNA Kit (Analytik Jena), following the manufacturer's protocol. DNA quality and concentration were determined using a Nanodrop ND-1000 spectrophotometer (Saveen Werner, Sweden).
4.3. Chloroplastic DNA amplification and sequencing
Four chloroplast DNA regions were considered (Table 2). PCR was performed in a 25 μl volume containing 2 μl of template DNA, 10 μl of 1X innuMix Standard PCR, and 1 μM of each primer (Table 2). The Gene Amp PCR System 9700 was used with the following program: initial denaturation at 94°C for 5 min, 35 cycles of denaturation at 94°C for 1 min, annealing at 49-52°C for 60-75s, and elongation at 72°C for 60-75s, followed by a final polymerization at 72°C for 10 min (Table 2). To check the effectiveness of PCR, positive control using sterile water was included in all amplifications. The PCR products were checked by electrophoresis on 1% agarose gel in TAE buffer, and DNA was visualized under UV light after staining with ethidium bromide.

Table 2.
General information about the PCR primers used in this study.
| Sr# | Sequence 5′-3′ | Target | PCR condition | Source |
|---|---|---|---|---|
| 1 | CATTACAAATGCGATGCTCT | trnT-trnL | Hybridation : 50°C/1 min Elongation : 72°C/1min | [45] |
| TCTACCGATTTCGCCATATC | ||||
| 2 | CGAAATCGGTAGACGCTACG | TrnL | Hybridation : 49°C/1.15 min Elongation : 72°C/1.15 min | [45] |
| GGGGATAGAGGGACTTGAAC | ||||
| 3 | GGTTCAAGTCCCTCTATCCC | TrnL-trnF | Hybridation :52°C/1 min Elongation : 72°C/1min | [45] |
| ATTTGAACTGGTGACACGAG | ||||
| 4 | GAAGTAGTAGGATTGATTCTC | atpB-rbcL | Hybridation : 50°C/1 min Elongation : 72°C/1min | [46] |
| TACAGTTGTCCATGTACCAG |
The amplified products were purified using the GFX PCR kit (GE Healthcare). Sequencing reactions were carried out by Congenic using Sanger technology, separately for each strand to obtain independent forward and reverse sequences. The forward and reverse fragments were aligned, and additional reactions were conducted in case of any discrepancies.
4.4. Sequence Analysis:
The scanner software-2 was utilized to determine the quality of the sequences. The four barcode samples of each coffee arabica genotype were manually curated and aligned using the contig assembly program in Bio Edit 7.0 software to ensure high-quality sequences. Nucleotide sequences obtained from the 57 accessions were initially aligned using CLUSTAL W [47] and analyzed with MEGA program version X. The number of individuals, number of nucleotide sites, variable polymorphic sites, number of segregating sites, number of haplotypes, nucleotide diversity, and average number of nucleotide differences of each barcode marker and consensus sequence were measured using DNAsp (v6) [48]. The quantification of insertion events in the sequence was determined by the number of variable sites where the addition of one or more nucleotides signals polymorphism. Likewise, the number of deletions was determined by the variable sites where polymorphism arises due to the removal of one or more nucleotides. The identification of the number of transitions in the sequences was based on the number of variable sites where polymorphism occurred due to the exchange between two purines (A and G) or two pyrimidines (C and T). On the other hand, the number of transversions was determined by the variable sites where polymorphism resulted from the replacement of a purine with a pyrimidine (Table 4). To determine the number of mutation events that have occurred in a sequence, the sum of variable sites and the number of distinct mutations observed at the same nucleotide site across different samples are combined. This quantification considers both different types of polymorphisms and multiple occurrences of mutations within the sequence. Various parameters were estimated for each sequence region to differentiate them, based on the number of monomorphic or polymorphic sites, the number of parsimony informative sites (PIC), nucleotide diversity (π), haplotype diversity (Hd), and the total number of mutations [49], singleton variable site (STVC) [50] (Table 5).
4.5. Evolutionary analysis by Maximum Likelihood method:
The evolutionary history was inferred by using the Maximum Likelihood method and Kimura 2-parameter model [51]. The tree with the highest log likelihood (-22360.57) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The proportion of sites where at least 1 unambiguous base is present in at least 1 sequence for each descendent clade is shown next to each internal node in the tree. This analysis involved 57 nucleotide sequences. There was a total of 5381 positions in the final dataset. Evolutionary analyses were conducted in MEGA X [52].
5. Conclusion:
To summarize, this study utilized a DNA barcoding approach to investigate the molecular relationships among fifty-six Arabica coffee samples collected from the southern region of Saudi Arabia. The three-barcode region, namely TrnT-TrnL, TrnL-TrnF, and TrnL, exhibited higher sequence variability compared to the atpB-rbcl barcode region and effectively differentiated the local coffee genotypes by the presence of unique variable sites (singletons and parsimony). Moreover, the combination of DNA sequences from these barcode loci, analyzed through phylogenetic analysis using the maximum likelihood method, grouped similar coffee genotypes, providing improved resolution, and understanding of the studied genotypes. These findings will contribute to future research programs in the identification and conservation of Arabica coffee using DNA barcoding markers.
Acknowledgments
The authors extend their appreciation to the Deputyship for Research & Innovation, Ministry of Education in Saudi Arabia for funding this research work through project number 1.092.
References
- Mussatto, S.I.; Machado, E.M.; Martins, S.; Teixeira, J.A. Production, composition, and application of coffee and its industrial residues. Food and Bioprocess Technology 2011, 4, 661–672. [Google Scholar] [CrossRef]
- Mishra, M.K. Genetic resources and breeding of coffee (Coffea spp.). Advances in Plant Breeding Strategies: Nut and Beverage Crops, 2019, 4, 475-515.
- Nguyen, G.N.; Norton, S.L. Genebank phenomics: A strategic approach to enhance value and utilization of crop germplasm. Plants 2020, 9, 817. [Google Scholar] [CrossRef] [PubMed]
- Migicovsky, Z.; Warschefsky, E.; Klein, L.L.; Miller, A.J. Using living germplasm collections to characterize, improve, and conserve woody perennials. Crop Science 2019, 59, 2365–2380. [Google Scholar] [CrossRef]
- Word Coffee Research. High-yield ing, dwarf/compact hybrid tolerant to coffee leaf rust and resistant to coffee berry disease (CBD) released in Kenya. https://varieties.worldcoffeeresearch.org/varieties/ruiru-11, 2023, (accessed on 6 June 2023).
- Montagnon, C.; Mahyoub, A.; Solano, W.; Sheibani, F. Unveiling a unique genetic diversity of cultivated Coffea arabica L. in its main domestication center: Yemen. Genetic Resources and Crop Evolution 2021, 68, 2411–2422. [Google Scholar] [CrossRef]
- Herrera, J.C.; Lambot, C. The coffee tree—Genetic diversity and origin. In The craft and science of coffee; Elsevier: 2017, pp. 1-16.
- Davis, A.P.; Chadburn, H.; Moat, J.; O’Sullivan, R.; Hargreaves, S.; Nic Lughadha, E. High extinction risk for wild coffee species and implications for coffee sector sustainability. Science advances 2019, 5, eaav3473. [Google Scholar] [CrossRef]
- Oliveira, L.S.; Franca, A.S. An overview of the potential uses for coffee husks. Coffee in health and disease prevention 2015, 283–291. [Google Scholar]
- Choudhary, A.; Gupta, N.; Hameed, F.; Choton, S. An overview of food adulteration: Concept, sources, impact, challenges and detection. International Journal of Chemical Studies 2020, 8, 2564–2573. [Google Scholar] [CrossRef]
- Al-Asmari, K.M.; Zeid, I.M.A.; Al-Attar, A.M. Coffee Arabica in Saudi Arabia: An Overview. International Journal of Pharmaceutical and Phytopharmacological Research (eIJPPR) 2020, 10, 71–78. [Google Scholar]
- Tounekti, T.; Mahdhi, M.; Al-Turki, T.A.; Khemira, H. Genetic diversity analysis of coffee (Coffee arabica L.) Germplasm accessions growing in the Southwestern Saudi Arabia using quantitative traits. Natural Resources 2017, 8, 321–336. [Google Scholar] [CrossRef]
- Montagnon, C.; Sheibani, F.; Benti, T.; Daniel, D.; Bote, A.D. Deciphering Early Movements and Domestication of Coffea arabica through a Comprehensive Genetic Diversity Study Covering Ethiopia and Yemen. Agronomy 2022, 12, 3203. [Google Scholar] [CrossRef]
- Eskes, A.B. Identification, description and collection of coffee types in P. D. R. Yemen. IPGRI, Rome, 1989. pp. 70.
- De Pauw, E. An agroecological exploration of the Arabian Peninsula. ICARDA, Aleppo, Syria, 2022, pp. 77.
- Tounekti T., M. Mahdhi, T. A. Al-Turki, H. Khemira. Water relations and photo-protection mechanisms during drought stress in four coffee (Coffea arabica) cultivars from southwestern Saudi Arabia. South African Journal of Botany, 2018, 117, 17–25. [Google Scholar] [CrossRef]
- Lashermes, P.; Trouslot, P.; Anthony, F.; Combes, M.-C.; Charrier, A. Genetic diversity for RAPD markers between cultivated and wild accessions of Coffea arabica. Euphytica 1996, 87, 59–64. [Google Scholar] [CrossRef]
- Cros, J.; Combes, M.-C.; Trouslot, P.; Anthony, F.; Hamon, S.; Charrier, A.; Lashermes, P. Phylogenetic analysis of chloroplast DNA variation in Coffea L. Molecular phylogenetics and evolution 1998, 9, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, K.; Govers, K.; Bekele, E.; Borsch, T. ISSR fingerprinting of Coffea arabica throughout Ethiopia reveals high variability in wild populations and distinguishes them from landraces. Plant systematics and evolution 2014, 300, 881–897. [Google Scholar] [CrossRef]
- Tesfaye, K.; Borsch, T.; Govers, K.; Bekele, E. Characterization of Coffea chloroplast microsatellites and evidence for the recent divergence of C. arabica and C. eugenioides chloroplast genomes. Genome 2007, 50, 1112–1129. [Google Scholar] [CrossRef]
- Lashermes, P.; Combes, M.-C.; Trouslot, P.; Charrier, A. Phylogenetic relationships of coffee-tree species (Coffea L.) as inferred from ITS sequences of nuclear ribosomal DNA. Theoretical and Applied Genetics 1997, 94, 947–955. [Google Scholar] [CrossRef]
- Maurin, O.; Davis, A.P.; Chester, M.; Mvungi, E.F.; Jaufeerally-Fakim, Y.; Fay, M.F. Towards a phylogeny for Coffea (Rubiaceae): identifying well-supported lineages based on nuclear and plastid DNA sequences. Annals of Botany 2007, 100, 1565–1583. [Google Scholar] [CrossRef]
- Al-Ghamedi, K.; Alaraidh, I.; Afzal, M.; Mahdhi, M.; Al-Faifi, Z.; Oteef, M.D.; Tounekti, T.; Alghamdi, S.S.; Khemira, H. Assessment of genetic diversity of local coffee populations in Southwestern Saudi Arabia using SRAP markers. Agronomy 2023, 13, 302. [Google Scholar] [CrossRef]
- Jingade, P.; Huded, A.K.; Kosaraju, B.; Mishra, M.K. Diversity genotyping of Indian coffee (Coffea arabica L.) germplasm accessions by using SRAP markers. Journal of Crop Improvement 2019, 33, 327–345. [Google Scholar] [CrossRef]
- Mashaly, A.; Alajmi, R.; Mustafa, A.E.-Z.; Rady, A.; Alkhedir, H. Species abundance and identification of forensically important flies of Saudi Arabia by DNA barcoding. Journal of medical entomology 2017, 54, 837–843. [Google Scholar] [CrossRef]
- Al-Dakhil, M.; Alghamdi, S.; Migdadi, H.; Afzal, M.; Ali, A.A. Morphological characterization and DNA barcoding of Duckweed species in Saudi Arabia. Plants 2021, 10, 2438. [Google Scholar] [CrossRef] [PubMed]
- Mekbib, Y.; Saina, J.K.; Tesfaye, K.; Eshetu, G.; Hu, G.; Chen, J.; Li, Z. Chloroplast genome sequence variations and development of polymorphic markers in Coffea arabica. Plant Molecular Biology Reporter 2020, 38, 491–502. [Google Scholar] [CrossRef]
- Cuénoud, P.; Savolainen, V.; Chatrou, L.W.; Powell, M.; Grayer, R.J.; Chase, M.W. Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences. American Journal of Botany 2002, 89, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.J.; Liu, J.; Möller, M.; Zhang, L.; Zhang, X.M.; Li, D.Z.; Gao, L.M. DNA barcoding of Rhododendron (Ericaceae), the largest Chinese plant genus in biodiversity hotspots of the Himalaya–Hengduan Mountains. Molecular ecology resources 2015, 15, 932–944. [Google Scholar] [CrossRef]
- Wu, F.; Li, M.; Liao, B.; Shi, X.; Xu, Y. DNA barcoding analysis and phylogenetic relation of mangroves in Guangdong Province, China. Forests 2019, 10, 56. [Google Scholar] [CrossRef]
- Vickers, N.J. Animal communication: when i’m calling you, will you answer too? Current biology 2017, 27, R713–R715. [Google Scholar] [CrossRef] [PubMed]
- Weigand, H.; Beermann, A.J.; Čiampor, F.; Costa, F.O.; Csabai, Z.; Duarte, S.; Geiger, M.F.; Grabowski, M.; Rimet, F.; Rulik, B. DNA barcode reference libraries for the monitoring of aquatic biota in Europe: Gap-analysis and recommendations for future work. Science of the Total Environment 2019, 678, 499–524. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: from gene to genome. Biological Reviews 2015, 90, 157–166. [Google Scholar] [CrossRef]
- Carneiro de Melo Moura, C.; Brambach, F.; Jair Hernandez Bado, K.; Krutovsky, K.V.; Kreft, H.; Tjitrosoedirdjo, S.S.; Siregar, I.Z.; Gailing, O. Integrating DNA barcoding and traditional taxonomy for the identification of dipterocarps in remnant lowland forests of Sumatra. Plants 2019, 8, 461. [Google Scholar] [CrossRef]
- Hong, Z.; He, W.; Liu, X.; Tembrock, L.R.; Wu, Z.; Xu, D.; Liao, X. Comparative Analyses of 35 Complete Chloroplast Genomes from the Genus Dalbergia (Fabaceae) and the Identification of DNA Barcodes for Tracking Illegal Logging and Counterfeit Rosewood. Forests 2022, 13, 626. [Google Scholar] [CrossRef]
- Kang, Y. Molecular identification of Aquilaria species with distribution records in China using DNA barcode technology. Mitochondrial DNA Part B 2021, 6, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.; Jingade, P.; Huded, A.K.C. DNA barcoding analysis and phylogenetic relationships of Indian wild coffee species. Turkish Journal of Botany 2022, 46, 109–122. [Google Scholar] [CrossRef]
- Hao, D.C.; Huang, B.L.; Chen, S.L.; Mu, J. Evolution of the chloroplast trnL-trnF region in the gymnosperm lineages Taxaceae and Cephalotaxaceae. Biochemical Genetics 2009, 47, 351–369. [Google Scholar] [CrossRef] [PubMed]
- Skuza, L.; SzuĆKo, I.; Filip, E.; Adamczyk, A. DNA barcoding in selected species and subspecies of Rye (Secale) using three chloroplast loci (matK, rbcL, trnH-psbA). Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2019, 47, 54–62. [Google Scholar] [CrossRef]
- Meena, R.K.; Negi, N.; Uniyal, N.; Shamoon, A.; Bhandari, M.S.; Pandey, S.; Negi, R.K.; Sharma, R.; Ginwal, H.S. Chloroplast-based DNA barcode analysis indicates high discriminatory potential of matK locus in Himalayan temperate bamboos. 3 Biotech 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Mahadani, P.; Ghosh, S.K. Utility of indels for species-level identification of a biologically complex plant group: a study with intergenic spacer in Citrus. Molecular biology reports 2014, 41, 7217–7222. [Google Scholar] [CrossRef]
- Wu, L.; Wu, M.; Cui, N.; Xiang, L.; Li, Y.; Li, X.; Chen, S. Plant super-barcode: a case study on genome-based identification for closely related species of Fritillaria. Chinese medicine 2021, 16, 1–11. [Google Scholar] [CrossRef]
- Hamon, P.; Grover, C.E.; Davis, A.P.; Rakotomalala, J.-J.; Raharimalala, N.E.; Albert, V.A.; Sreenath, H.L.; Stoffelen, P.; Mitchell, S.E.; Couturon, E. Genotyping-by-sequencing provides the first well-resolved phylogeny for coffee (Coffea) and insights into the evolution of caffeine content in its species: GBS coffee phylogeny and the evolution of caffeine content. Molecular phylogenetics and evolution 2017, 109, 351–361. [Google Scholar] [CrossRef]
- Guyeux, C.; Charr, J.-C.; Tran, H.T.; Furtado, A.; Henry, R.J.; Crouzillat, D.; Guyot, R.; Hamon, P. Evaluation of chloroplast genome annotation tools and application to analysis of the evolution of coffee species. PLoS One 2019, 14, e0216347. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of. Plant molecular biology 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Savolainen, V.; Corbaz, R.; Moncousin, C.; Spichiger, R.; Manen, J.-F. Chloroplast DNA variation and parentage analysis in 55 apples. Theoretical and Applied Genetics 1995, 90, 1138–1141. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic acids research 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.; Sánchez-Gracia, A. DnaSP version 6 for 32-bit and 64-bit environments (ZIP file of 11.7 Mb).
- Hosein, F.N.; Austin, N.; Maharaj, S.; Johnson, W.; Rostant, L.; Ramdass, A.C.; Rampersad, S.N. Utility of DNA barcoding to identify rare endemic vascular plant species in Trinidad. Ecology and Evolution 2017, 7, 7311–7333. [Google Scholar] [CrossRef] [PubMed]
- Pettengill, J.B.; Neel, M.C. An evaluation of candidate plant DNA barcodes and assignment methods in diagnosing 29 species in the genus Agalinis (Orobanchaceae). American journal of botany 2010, 97, 1391–1406. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of molecular evolution 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Molecular biology and evolution 2018, 35, 1547. [Google Scholar] [CrossRef]
Table 2.
Accession numbers of four barcode primers of 56 Coffea arabica genotypes.
| Genotype ID | atpB-rbcl | TrnL-TrnF | TrnT-TrnL | TrnL |
|---|---|---|---|---|
| KSA1R | OQ718327 | OQ914867 | OQ914923 | OQ953999 |
| KSA2R | -------- | OQ914868 | OQ914924 | OQ954000 |
| KSA3R | OQ844066 | OQ914869 | OQ914925 | OQ954001 |
| KSA4R | OQ914863 | OQ914870 | OQ914926 | OQ954002 |
| KSA5R | OQ914864 | OQ914871 | OQ914927 | OQ954003 |
| KSA6R | OQ914865 | OQ914872 | OQ914928 | OQ954004 |
| KSA7R | OQ914866 | OQ914873 | OQ914929 | OQ954005 |
| KSA8R | OQ850301 | OQ914874 | OQ914930 | OQ954006 |
| KSA9R | OQ850302 | OQ914875 | OQ914931 | OQ954007 |
| KSA10R | OQ850303 | OQ914876 | OQ914932 | OQ954008 |
| KSA11R | OQ850304 | OQ914877 | OQ914933 | OQ954009 |
| KSA12R | OQ850305 | OQ914878 | OQ914934 | OQ954010 |
| KSA13R | OQ850306 | OQ914879 | OQ914935 | OQ954011 |
| KSA15R | OQ851715 | OQ914880 | OQ914936 | OQ954012 |
| KSA16R | OQ851716 | OQ914881 | OQ914937 | OQ954013 |
| KSA17R | OQ851717 | OQ914882 | OQ914938 | OQ954014 |
| KSA18R | OQ851718 | OQ914883 | OQ914939 | OQ954015 |
| KSA19R | OQ851719 | OQ914884 | OQ914940 | OQ954016 |
| KSA20 | OQ851720 | OQ914885 | OQ914941 | OQ954017 |
| KSA21 | OQ872544 | OQ914886 | OQ914942 | OQ954018 |
| KSA22 | OQ872545 | OQ914887 | OQ914943 | OQ954019 |
| KSA23 | OQ872546 | OQ914888 | OQ914944 | OQ954020 |
| KSA24 | OQ872547 | OQ914889 | OQ914945 | OQ954021 |
| KSA25 | OQ872548 | OQ914890 | OQ914946 | OQ954022 |
| KSA26 | OQ872549 | OQ914891 | OQ914947 | OQ954023 |
| KSA27 | OQ872550 | OQ914892 | OQ914948 | OQ954024 |
| KSA28 | OQ872551 | OQ914893 | OQ914949 | OQ954025 |
| KSA29 | OQ872552 | OQ914894 | OQ914950 | OQ954026 |
| KSA30 | OQ872553 | OQ914895 | OQ914951 | OQ954027 |
| KSA31 | OQ872554 | OQ914896 | OQ914952 | OQ954028 |
| KSA32 | OQ872555 | OQ914897 | OQ914953 | OQ954029 |
| KSA33 | OQ872556 | OQ914898 | OQ914954 | OQ954030 |
| KSA34 | OQ872557 | OQ914899 | OQ914955 | OQ954031 |
| KSA35 | OQ872558 | OQ914900 | OQ914956 | OQ954032 |
| KSA36 | OQ872559 | OQ914901 | OQ914957 | OQ954033 |
| KSA37 | OQ872560 | OQ914902 | OQ914958 | OQ954034 |
| KSA38 | OQ872561 | OQ914903 | OQ914959 | OQ954035 |
| KSA39 | OQ872562 | OQ914904 | OQ914960 | OQ954036 |
| KSA40 | OQ872563 | OQ914905 | OQ914961 | OQ954037 |
| KSA41 | ------------- | OQ914906 | OQ914962 | OQ954038 |
| KSA42 | ------------- | OQ914907 | OQ914963 | OQ954039 |
| KSA43 | ------------- | OQ914908 | OQ914964 | OQ954040 |
| KSA44 | OQ852764 | OQ914909 | OQ914965 | OQ954041 |
| KSA45R | OQ852765 | OQ914910 | OQ914966 | OQ954042 |
| KSA46 | OQ852766 | OQ914911 | OQ914967 | OQ954043 |
| KSA47 | OQ852767 | OQ914912 | OQ914968 | OQ954044 |
| KSA48 | OQ852768 | OQ914913 | OQ914969 | OQ954045 |
| KSA49 | OQ852769 | OQ914914 | OQ914970 | OQ954046 |
| KSA50 | OQ852770 | OQ914915 | OQ914971 | OQ954047 |
| KSA51R | OQ852771 | OQ914916 | OQ914972 | OQ954048 |
| KSA52 | OQ852772 | OQ914917 | OQ914973 | OQ954049 |
| KSA59 | OQ852773 | OQ914918 | OQ914974 | OQ954050 |
| KSA60 | OQ852774 | OQ914919 | OQ914975 | OQ954051 |
| KSA61 | OQ852775 | OQ914920 | OQ914976 | OQ954052 |
| KSA62 | OQ852776 | OQ914921 | OQ914977 | OQ954053 |
| KSA63 | OQ852777 | OQ914922 | OQ914978 | OQ954054 |
Note: KSA2; KSA41; KSA42; KSA43 were not identified in the data base for atpB-rbcl barcode.
Table 3.
Summary of nucleotide sites, variable polymorphic sites, number of segregating sites, haploid diversity, nucleotide diversity, and average number of nucleotide difference.
Table 3.
Summary of nucleotide sites, variable polymorphic sites, number of segregating sites, haploid diversity, nucleotide diversity, and average number of nucleotide difference.
| Barcode Name | Individual | NNS | VPS | NSS | NH | ND | ANND |
|---|---|---|---|---|---|---|---|
| atpB-rbcl | 56 | 1139 | 341 | 341 | 17 | 0.54 | 185.54 |
| TrnL | 55 | 551 | 237 | 237 | 31 | 0.056 | 18.93 |
| TrnL-TrnF | 56 | 1055 | 154 | 154 | 17 | 0.046 | 25.23 |
| TrnT-TrnL | 56 | 988 | 421 | 421 | 50 | 0.051 | 40.50 |
| atpB-rbcl + TrnL+TrnL-TrnF + TrnT-TrnL | 223 | 4114 | 651 | 651 | 37 | 0.11 | 295 |
Table 4.
Nucleotide base substitution matrix of four barcoding markers in Arabica coffee.
| Atpb-rbcl | TrnL | TrnC | TrnT-TrnL | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | T(U) | C | A | G | Total | T(U) | C | A | G | Total | T(U) | C | A | G | Total | T(U) | C | A | G | Total |
| KSA1R | 36.36 | 16.01 | 30.48 | 17.15 | 968 | 32.94 | 20.38 | 32.46 | 14.22 | 422 | 26.9 | 15.7 | 37.9 | 19.6 | 562 | 39.48 | 13.49 | 33.79 | 13.24 | 808 |
| KSA2R | 33.25 | 19.21 | 33.50 | 14.04 | 406 | 28.5 | 15.5 | 35.8 | 20.2 | 1017 | 39.58 | 14.02 | 33.75 | 12.66 | 806 | |||||
| KSA3R | 35.60 | 15.89 | 31.48 | 17.03 | 969 | 33.25 | 19.76 | 32.53 | 14.46 | 415 | 27.3 | 15.5 | 38.0 | 19.3 | 561 | 39.14 | 13.83 | 34.20 | 12.84 | 810 |
| KSA4R | 35.62 | 15.86 | 31.12 | 17.40 | 977 | 33.74 | 19.56 | 32.03 | 14.67 | 409 | 26.9 | 15.6 | 38.0 | 19.5 | 558 | 36.78 | 14.23 | 33.72 | 15.28 | 949 |
| KSA5R | 35.34 | 15.89 | 31.57 | 17.21 | 982 | 33.58 | 20.34 | 31.86 | 14.22 | 408 | 27.6 | 15.7 | 37.7 | 19.0 | 562 | 37.93 | 14.81 | 33.60 | 13.67 | 878 |
| KSA6R | 34.38 | 15.57 | 31.90 | 18.15 | 1047 | 33.66 | 19.85 | 32.20 | 14.29 | 413 | 27.9 | 16.1 | 36.9 | 19.1 | 559 | 39.63 | 13.46 | 34.69 | 12.22 | 810 |
| KSA7R | 34.34 | 16.49 | 31.90 | 17.27 | 1025 | 32.94 | 19.81 | 32.94 | 14.32 | 419 | 26.4 | 16.3 | 38.0 | 19.3 | 569 | 39.15 | 13.66 | 34.88 | 12.32 | 820 |
| KSA8R | 35.20 | 16.02 | 30.92 | 17.86 | 980 | 33.33 | 20.29 | 31.88 | 14.49 | 414 | 26.4 | 15.5 | 38.4 | 19.7 | 549 | 38.00 | 13.65 | 34.47 | 13.88 | 850 |
| KSA9R | 35.79 | 15.79 | 30.97 | 17.44 | 975 | 27.97 | 17.72 | 34.27 | 20.05 | 429 | 26.4 | 15.6 | 38.3 | 19.6 | 556 | 38.63 | 13.92 | 33.18 | 14.27 | 862 |
| KSA10R | 35.35 | 16.14 | 31.14 | 17.37 | 973 | 30.37 | 22.73 | 32.64 | 14.26 | 484 | 26.3 | 15.6 | 38.1 | 20.0 | 551 | 38.31 | 14.24 | 32.54 | 14.92 | 885 |
| KSA11R | 31.89 | 17.34 | 35.36 | 15.41 | 1038 | 32.39 | 22.98 | 29.76 | 14.88 | 457 | 25.9 | 16.5 | 37.3 | 20.3 | 557 | 36.42 | 13.22 | 31.97 | 18.39 | 832 |
| KSA12R | 32.69 | 16.04 | 36.33 | 14.93 | 991 | 34.00 | 18.60 | 32.00 | 15.40 | 500 | 27.0 | 15.7 | 38.0 | 19.3 | 548 | 39.53 | 13.84 | 33.54 | 13.09 | 802 |
| KSA13R | 33.13 | 16.36 | 35.60 | 14.92 | 972 | 33.25 | 20.15 | 31.80 | 14.81 | 412 | 26.7 | 16.1 | 37.5 | 19.7 | 554 | 40.08 | 13.94 | 34.17 | 11.81 | 796 |
| KSA15R | 32.80 | 16.10 | 36.22 | 14.89 | 994 | 33.66 | 20.34 | 31.72 | 14.29 | 413 | 27.0 | 15.4 | 37.9 | 19.7 | 544 | 39.85 | 14.25 | 33.92 | 11.98 | 793 |
| KSA16R | 33.13 | 16.26 | 35.38 | 15.24 | 978 | 32.89 | 20.00 | 32.00 | 15.11 | 450 | 26.7 | 16.4 | 37.5 | 19.4 | 566 | 39.43 | 14.18 | 33.58 | 12.81 | 804 |
| KSA17R | 32.66 | 16.13 | 36.19 | 15.02 | 992 | 33.98 | 19.28 | 32.05 | 14.70 | 415 | 26.8 | 15.5 | 37.6 | 20.1 | 548 | 39.87 | 13.96 | 33.71 | 12.45 | 795 |
| KSA18R | 32.49 | 16.05 | 35.81 | 15.66 | 1022 | 33.09 | 20.19 | 31.63 | 15.09 | 411 | 26.9 | 15.8 | 37.7 | 19.6 | 551 | 40.30 | 14.23 | 33.88 | 11.59 | 794 |
| KSA19R | 32.79 | 15.94 | 36.14 | 15.13 | 985 | 31.80 | 19.80 | 32.00 | 16.40 | 500 | 26.8 | 15.6 | 37.9 | 19.7 | 557 | 39.53 | 14.00 | 34.20 | 12.27 | 807 |
| KSA20 | 32.40 | 15.90 | 36.40 | 15.30 | 1000 | 33.41 | 19.95 | 31.73 | 14.90 | 416 | 26.1 | 15.4 | 38.3 | 20.1 | 566 | 38.99 | 13.74 | 33.79 | 13.49 | 808 |
| KSA21 | 33.16 | 15.76 | 36.23 | 14.84 | 977 | 30.93 | 19.77 | 34.19 | 15.12 | 430 | 26.1 | 15.3 | 38.2 | 20.4 | 555 | 39.88 | 14.00 | 33.50 | 12.63 | 800 |
| KSA22 | 32.99 | 16.55 | 35.65 | 14.81 | 979 | 32.49 | 19.22 | 33.18 | 15.10 | 437 | 26.6 | 15.0 | 38.4 | 20.0 | 515 | 39.88 | 14.00 | 33.75 | 12.38 | 800 |
| KSA23 | 33.07 | 15.95 | 35.80 | 15.18 | 1028 | 32.85 | 20.05 | 31.64 | 15.46 | 414 | 27.1 | 15.4 | 37.9 | 19.6 | 565 | 39.88 | 14.00 | 33.38 | 12.75 | 800 |
| KSA24 | 32.40 | 16.15 | 35.95 | 15.50 | 929 | 33.50 | 20.39 | 31.55 | 14.56 | 412 | 26.4 | 14.9 | 38.2 | 20.5 | 523 | 39.48 | 13.82 | 33.62 | 13.08 | 803 |
| KSA25 | 32.97 | 15.55 | 36.22 | 15.26 | 1016 | 28.67 | 14.00 | 34.89 | 22.44 | 450 | 26.5 | 16.6 | 37.0 | 19.9 | 548 | 39.88 | 14.00 | 33.63 | 12.50 | 800 |
| KSA26 | 32.90 | 15.33 | 36.45 | 15.33 | 1070 | 34.12 | 19.43 | 31.75 | 14.69 | 422 | 26.6 | 15.8 | 37.9 | 19.7 | 549 | 40.03 | 13.80 | 33.75 | 12.42 | 797 |
| KSA27 | 31.87 | 15.99 | 36.05 | 16.09 | 957 | 33.65 | 19.47 | 32.21 | 14.66 | 416 | 27.2 | 15.7 | 37.4 | 19.7 | 548 | 39.50 | 14.13 | 33.75 | 12.63 | 800 |
| KSA28 | 32.47 | 16.80 | 35.57 | 15.15 | 970 | 32.44 | 19.11 | 32.00 | 16.44 | 450 | 26.2 | 16.3 | 36.4 | 21.1 | 583 | 39.63 | 14.00 | 33.75 | 12.63 | 800 |
| KSA29 | 35.42 | 25.33 | 21.87 | 17.38 | 1070 | 33.57 | 19.08 | 31.40 | 15.94 | 414 | 25.7 | 16.8 | 36.0 | 21.5 | 600 | 39.15 | 14.09 | 33.29 | 13.47 | 802 |
| KSA30 | 32.48 | 16.45 | 36.16 | 14.91 | 979 | 32.77 | 19.76 | 32.77 | 14.70 | 415 | 26.0 | 15.6 | 38.3 | 20.0 | 569 | 39.60 | 13.70 | 33.62 | 13.08 | 803 |
| KSA31 | 33.23 | 16.16 | 35.69 | 14.93 | 978 | 32.53 | 20.24 | 32.53 | 14.70 | 415 | 25.1 | 16.0 | 39.2 | 19.7 | 589 | 33.37 | 11.21 | 40.97 | 14.45 | 803 |
| KSA32 | 32.72 | 16.62 | 35.49 | 15.18 | 975 | 33.41 | 20.58 | 32.20 | 13.80 | 413 | 27.1 | 15.9 | 37.7 | 19.3 | 554 | 39.88 | 13.88 | 33.63 | 12.63 | 800 |
| KSA33 | 32.93 | 16.21 | 36.05 | 14.80 | 993 | 33.57 | 20.05 | 31.88 | 14.49 | 414 | 26.5 | 16.2 | 37.5 | 19.8 | 550 | 38.83 | 14.02 | 34.12 | 13.03 | 806 |
| KSA34 | 32.28 | 16.65 | 36.06 | 15.02 | 979 | 33.50 | 19.90 | 31.80 | 14.81 | 412 | 26.0 | 16.2 | 38.4 | 19.4 | 573 | 38.64 | 13.73 | 34.26 | 13.37 | 823 |
| KSA35 | 32.67 | 16.04 | 36.16 | 15.14 | 1004 | 32.60 | 19.95 | 32.85 | 14.60 | 411 | 26.7 | 15.7 | 38.1 | 19.5 | 554 | 39.29 | 13.92 | 33.62 | 13.18 | 812 |
| KSA36 | 33.54 | 15.90 | 35.28 | 15.28 | 975 | 31.66 | 19.36 | 33.94 | 15.03 | 439 | 26.5 | 16.0 | 37.9 | 19.6 | 551 | 39.41 | 13.63 | 34.32 | 12.64 | 807 |
| KSA37 | 32.99 | 15.73 | 35.96 | 15.32 | 979 | 32.50 | 19.09 | 32.73 | 15.68 | 440 | 27.1 | 15.5 | 38.7 | 18.8 | 595 | 39.70 | 13.52 | 34.37 | 12.41 | 806 |
| KSA38 | 32.96 | 15.61 | 36.14 | 15.30 | 974 | 33.49 | 20.48 | 31.33 | 14.70 | 415 | 27.1 | 15.9 | 37.7 | 19.3 | 584 | 40.08 | 14.03 | 34.13 | 11.76 | 791 |
| KSA39 | 33.03 | 15.64 | 36.40 | 14.93 | 978 | 32.94 | 20.56 | 31.31 | 15.19 | 428 | 27.3 | 15.7 | 37.8 | 19.3 | 535 | 39.41 | 13.88 | 33.46 | 13.26 | 807 |
| KSA40 | 32.39 | 16.47 | 35.21 | 15.93 | 923 | 33.01 | 19.86 | 32.54 | 14.59 | 418 | 26.4 | 15.7 | 38.1 | 19.9 | 554 | 39.18 | 13.84 | 33.37 | 13.60 | 809 |
| KSA41 | 33.49 | 19.95 | 32.80 | 13.76 | 436 | 26.6 | 14.9 | 38.2 | 20.3 | 523 | 39.67 | 14.11 | 33.75 | 12.47 | 794 | |||||
| KSA42 | 34.49 | 20.14 | 30.79 | 14.58 | 432 | 26.4 | 15.5 | 37.3 | 20.8 | 576 | 40.05 | 14.30 | 33.58 | 12.06 | 804 | |||||
| KSA43 | 33.82 | 20.19 | 31.63 | 14.36 | 411 | 27.0 | 14.7 | 37.9 | 20.4 | 530 | 39.88 | 13.75 | 33.75 | 12.63 | 800 | |||||
| KSA44 | 31.89 | 16.76 | 35.57 | 15.78 | 925 | 31.65 | 19.50 | 31.19 | 17.66 | 436 | 26.8 | 15.9 | 37.7 | 19.6 | 560 | 39.30 | 13.81 | 33.21 | 13.68 | 804 |
| KSA45R | 32.86 | 15.84 | 35.68 | 15.62 | 922 | 32.06 | 19.38 | 33.01 | 15.55 | 418 | 27.2 | 15.7 | 37.2 | 20.0 | 541 | 39.60 | 13.82 | 33.50 | 13.08 | 803 |
| KSA46 | 32.86 | 16.43 | 35.61 | 15.10 | 980 | 32.64 | 19.44 | 32.18 | 15.74 | 432 | 26.7 | 15.8 | 37.3 | 20.2 | 544 | 39.35 | 14.16 | 33.58 | 12.91 | 798 |
| KSA47 | 33.10 | 15.68 | 35.95 | 15.27 | 982 | 33.17 | 19.61 | 32.93 | 14.29 | 413 | 26.1 | 16.0 | 38.2 | 19.7 | 563 | 39.63 | 14.13 | 33.50 | 12.75 | 800 |
| KSA48 | 32.39 | 16.30 | 35.65 | 15.65 | 920 | 33.63 | 19.41 | 32.05 | 14.90 | 443 | 28.6 | 21.6 | 32.4 | 17.4 | 574 | 39.63 | 14.16 | 33.66 | 12.55 | 805 |
| KSA49 | 32.65 | 16.05 | 35.79 | 15.51 | 922 | 32.61 | 20.86 | 31.89 | 14.63 | 417 | 26.9 | 17.7 | 35.5 | 19.9 | 583 | 39.30 | 13.93 | 33.58 | 13.18 | 804 |
| KSA50 | 32.68 | 16.02 | 35.82 | 15.48 | 924 | 32.27 | 21.59 | 30.00 | 16.14 | 440 | 26.9 | 15.4 | 37.9 | 19.8 | 551 | 39.78 | 13.97 | 33.29 | 12.97 | 802 |
| KSA51R | 32.39 | 15.71 | 36.51 | 15.38 | 923 | 33.58 | 19.95 | 32.12 | 14.36 | 411 | 26.5 | 16.5 | 37.9 | 19.1 | 570 | 39.46 | 13.93 | 33.91 | 12.70 | 811 |
| KSA52 | 32.68 | 16.07 | 35.40 | 15.85 | 921 | 32.85 | 20.68 | 31.39 | 15.09 | 411 | 26.7 | 15.7 | 37.1 | 20.5 | 536 | 38.75 | 14.02 | 33.58 | 13.65 | 813 |
| KSA59 | 32.36 | 16.13 | 35.71 | 15.80 | 924 | 33.50 | 19.75 | 31.50 | 15.25 | 400 | 26.3 | 15.6 | 38.5 | 19.6 | 556 | 38.27 | 13.49 | 33.54 | 14.70 | 823 |
| KSA60 | 32.50 | 16.36 | 35.75 | 15.38 | 923 | 33.64 | 20.23 | 30.45 | 15.68 | 440 | 26.7 | 16.2 | 37.6 | 19.6 | 551 | 38.56 | 14.20 | 33.78 | 13.46 | 817 |
| KSA61 | 32.29 | 15.87 | 35.64 | 16.20 | 926 | 32.70 | 20.14 | 32.46 | 14.69 | 422 | 25.9 | 16.6 | 37.0 | 20.5 | 595 | 39.23 | 14.20 | 33.50 | 13.08 | 803 |
| KSA62 | 32.86 | 16.02 | 36.22 | 14.90 | 980 | 32.00 | 18.00 | 35.50 | 14.50 | 400 | 27.4 | 15.6 | 36.9 | 20.0 | 544 | 39.70 | 13.86 | 33.58 | 12.86 | 801 |
| KSA63 | 33.09 | 15.93 | 35.96 | 15.02 | 979 | 32.81 | 19.82 | 32.21 | 15.16 | 426 | 26.4 | 15.9 | 37.7 | 19.9 | 552 | 39.57 | 13.43 | 34.05 | 12.95 | 834 |
| Avg. | 33.15 | 16.60 | 34.49 | 15.76 | 963.8 | 26.7 | 15.9 | 37.6 | 19.8 | 566 | 39.18 | 13.88 | 33.84 | 13.10 | 812 | |||||
Table 5.
Singleton variable sites and parsimony information sites of the four chloroplasts barcode.
| Gene | Parameter | Sites | Position | Total | Grand Total |
| atpB-rbcl. | STVS | Two variants’ sites | 448 | 1 | 1 |
| Three variants’ sites | 0 | ||||
| PIC | Two variants’ sites | 338 362 408 442 443 444 447 457 469 878 913 | 11 | 137 | |
| Three variants’ sites | 335 339 340 350 356 360 361 363 364 366 369 370 383 387 388 394 395 396 398 399 400 403 404 405 410 414 416 417 430 432 433 434 435 437 438 439 441 445 446 451 452 453 454 455 459 460 464 465 466 467 470 471 488 489 490 494 495 496 497 498 499 500 505 507 510 512 513 515 520 522 523 524 530 531 534 536 539 540 543 545 546 549 552 553 554 555 562 563 564 567 568 569 570 595 602 766 768 771 776 779 780 784 789 794 795 826 828 829 834 844 845 855 866 867 869 874 882 896 897 907 908 911 914 915 921 924 | 126 | |||
| TrnL | STVS | Two variants’ sites | 40 60 103 134 149 155 165 173 178 182 186 192 215 216 218 219 237 242 246 335 343 344 369 378 386 390 391 397 398 400 402 405 407 408 411 413 417 418 419 427 430 431 | 42 | 52 |
| Three variants’ sites | 124 157 217 379 406 410 414 428 429 439 | 10 | |||
| PIC | Two variants’ sites | 37 42 49 50 58 63 64 71 72 73 74 76 77 79 84 85 86 87 88 92 93 99 100 102 105 106 108 109 112 113 114 116 117 123 126 129 130 131 132 140 141 142 146 148 154 156 161 162 163 167 168 170 172 180 184 190 191 195 196 198 199 200 201 203 204 221 223 226 227 228 232 233 239 244 245 248 250 251 252 253 255 256 258 260 261 263 266 267 270 273 274 275 276 278 280 281 282 284 288 289 294 295 297 302 303 304 305 309 312 313 314 315 318 320 321 322 323 324 327 328 329 330 332 333 334 337 341 342 346 347 348 349 351 352 353 354 356 357 358 360 361 362 363 365 367 373 375 382 387 392 393 401 426 437 440 | 155 | 182 | |
| Three variants’ sites | 21 30 43 44 45 53 78 89 90 118 181 185 291 345 350 355 366 368 372 374 376 377 381 383 384 403 432 | 27 | |||
| TrnL-TrnF | STVS | Two variants’ sites | 32 33 50 51 64 91 224 256 268 271 273 281 282 284 285 286 287 290 291 293 297 300 301 307 308 310 312 314 315 318 320 321 324 325 327 328 330 332 336 337 338 340 346 347 348 349 350 351 353 354 356 357 362 363 364 365 366 367 369 370 371 373 374 375 376 385 388 389 392 393 396 403 406 407 410 411 416 419 426 427 429 430 432 443 444 445 447 448 452 455 456 457 459 460 467 470 471 472 475 476 484 487 490 494 495 497 501 504 505 506 508 510 520 521 535 543 548 549 551 | 119 | 133 |
| Three variants’ sites | 37 368 394 399 404 420 446 454 464 482 488 489 530 552 | 14 | |||
| PIC | Two variants’ sites | 31 46 47 86 88 382 386 400 414 415 422 423 431 442 449 450 453 458 463 473 474 477 478 479 485 486 492 493 502 518 525 533 538 541 547 | 35 | 45 | |
| Three variants’ sites | 23 441 499 500 522 531 532 539 545 546 | 10 | |||
| TrnT-TrnL | STVS | Two variants’ sites | 33 35 36 37 39 40 41 42 43 45 46 49 51 52 53 60 62 63 65 69 73 76 77 78 80 81 82 84 86 87 88 91 92 96 97 98 99 101 106 107 108109 114 115 122 124 125 126 127 129 130 131 132 136 137 138 141 142 143 150 151 152 154 155 161 162 163 166 167 169 174 175 177 178 180 184 189 190 191 192 195 196 197 202 204 211 213 217 221 222 223 224 227 229 230 233 236 237 249 253 254 256 257 258 259 260 261 262 265 267 268 270 271 273 275 276 283 284 287 288 294 299 301 303 304 305 306 310 311 312 313 321 322 326 327 328 330 332 335 339 346 347 351 352 354 356 357 358 359 366 368 371 375 377 379 381 382 385 386 391 394 398 400 403 405 411 414 415 416 417 420 422 428 432 435 437 445 446 447 449 451 456 457 464 468 471 473 475 476 477 481 482 490 491 492 493 497 498 499 500 502 504 509 513 514 515 516 519 520 527 528 529 532 533 534 535 538 542 543 544 545 546 548 551 554 565 568 576 579 580 582 595 598 599 600 601 604 605 618 626 630 631 633 638 639 640 642 643 645 646 647649 652 654 655 656 661 664 668 670 674 676 678 679 680 682 684 685 688 691 695 696 697 698 701 703 706 709 711 714 715 716 718 719 722 723 725 727 749 752 756 760 762 766 768 771 774 777 778 782 791 800 814 | 303 | 338 |
| Three variants’ sites | 61 72 550 570 581 606 607 613 622 625 627 628 653 659 675 707 710 713 721 724 728 732 734 741 754 763 769 770 775 776 780 792 795 796 797 | 35 | |||
| PIC | Two variants’ sites | 28 240 393 421 444 452 470 530 541 547 549 573 574 586 590 592 594 608 614 619 620 621 624 636 650 660 665 666 671 677 683 705 708 712 731 748 751 753 755 798 799 802 805 812 819 820 821 | 47 | 74 | |
| Four variants’ sites | 22 24 27 48 635 641 648 672 673 689 720 726 739 740 764 765 767 779 783 794 801 806 807 809 810 816 822 | 27 |
Table 1.
Altitude and latitude of the sites where the coffee accessions were sourced. The sites are located between longitudes 42˚22' and 43˚07'E.
Table 1.
Altitude and latitude of the sites where the coffee accessions were sourced. The sites are located between longitudes 42˚22' and 43˚07'E.
| # | Accession no. | Region | District | Altitude (m a.s.l.) | Latitude |
|---|---|---|---|---|---|
| 1 | KSA1R | Jazan | Khacher/Al-Zoughli | 1254 | 17°18'03"N |
| 2 | KSA2R | Jazan | Khacher/Al-Guatil | 1484 | 17°19'01"N |
| 3 | KSA3R | Jazan | Khacher/Al-Guatil | 1484 | 17°19'01"N |
| 4 | KSA4R | Jazan | Jebel Fayfa | 1541 | 17°15'21"N |
| 5 | KSA5R | Jazan | Wadi Dafa | 1254 | 17°25'41"N |
| 6 | KSA6R | Jazan | Tallan | 1672 | 17°23'12"N |
| 7 | KSA7R | Jazan | Tallan | 1672 | 17°23'12"N |
| 8 | KSA8R | Jazan | Tallan | 1546 | 17°23'01"N |
| 9 | KSA9R | Jazan | Tallan | 1672 | 17°23'12"N |
| 10 | KSA10R | Jazan | Khacher/Al-Zoughli | 1254 | 17°18'03"N |
| 11 | KSA11R | Assir | Rayda | 1594 | 18º11'37"N |
| 12 | KSA12R | Jazan | Maaddi | 1287 | 17º29'29"N |
| 13 | KSA13R | Jazan | Maaddi | 1344 | 17º29'29"N |
| 14 | KSA15R | Al-Baha | Shada Al-ala | 1548 | 19º50'54"N9 |
| 15 | KSA16R | Assir | Rayda | 1594 | 18º11'37"N |
| 16 | KSA17R | Assir | Rayda | 1519 | 18º11'37"N |
| 17 | KSA18R | Assir | Al-Majarda | 1329 | 19º09'35"N |
| 18 | KSA19R | Assir | Al-Majarda | 1300 | 19º09'35"N |
| 19 | KSA20 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 20 | KSA21 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 21 | KSA22 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 22 | KSA23 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 23 | KSA24 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 24 | KSA25 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 25 | KSA26 | Jazan | Jebel Fayfa | 1550 | 17º15'24"N |
| 26 | KSA27 | Jazan | Jebel Fayfa | 1550 | 17º15'24"N |
| 27 | KSA28 | Jazan | Jebel Fayfa | 1550 | 17º15'24"N |
| 28 | KSA29 | Jazan | Al-Gahr | 1846 | 17º38'08"N |
| 29 | KSA30 | Jazan | Al-Gahr | 1846 | 17º38'08"N |
| 30 | KSA31 | Jazan | Al-Gahr | 1846 | 17º38'08"N |
| 31 | KSA32 | Jazan | Al-Gahr | 1846 | 17º38'08"N |
| 32 | KSA33 | Jazan | Al-Gahr | 1846 | 17º38'08"N |
| 33 | KSA34 | Jazan | Jebel Fayfa | 1660 | 17º15'55"N |
| 34 | KSA35 | Jazan | Jebel Fayfa | 1660 | 17º15'55"N |
| 35 | KSA36 | Jazan | Jebel Fayfa | 1450 | 17º15'59"N |
| 36 | KSA37 | Jazan | Eddayer | 1100 | 17º22'10"N |
| 37 | KSA38 | Jazan | Eddayer | 1228 | 17º22'10"N |
| 38 | KSA39 | Jazan | Eddayer | 1228 | 17º22'10"N |
| 39 | KSA40 | Jazan | Haroub | 1100 | 17º29'29"N |
| 40 | KSA41 | Assir | Rayda | 1450 | 18º11'37"N |
| 41 | KSA42 | Assir | Rayda | 1450 | 18º11'37"N |
| 42 | KSA43 | Assir | Rayda | 1400 | 18º11'37"N |
| 43 | KSA44 | Jazan | Jebel Fayfa | 1524 | 17º15'48"N |
| 44 | KSA45R | Jazan | Jebel Fayfa | 1524 | 17º15'48"N |
| 45 | KSA46 | Jazan | Al-Gahr | 1750 | 17º39'01"N |
| 46 | KSA47 | Jazan | Al-Gahr | 1750 | 17º39'01"N |
| 47 | KSA48 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 48 | KSA49 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 49 | KSA50 | Jazan | Jebel Fayfa | 1260 | 17º15'20"N |
| 50 | KSA51R | Jazan | Jebel Fayfa | 1524 | 17º17'13"N |
| 51 | KSA52 | Jazan | Jebel Fayfa | 1550 | 17º15'24"N |
| 52 | KSA59 | Assir | Al-Majarda | 1329 | 19º09'35"N |
| 53 | KSA60 | Assir | Al-Majarda | 1300 | 19º09'35"N |
| 54 | KSA61 | Al-Baha | Shada Al-ala | 1548 | 19º50'54"N |
| 55 | KSA62 | Al-Baha | Shada Al-ala | 1548 | 19º50'54"N |
| 56 | KSA63 | Al-Baha | Shada Al-ala | 1548 | 19º50'54"N |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



