Submitted:
05 June 2023
Posted:
05 June 2023
You are already at the latest version
Abstract
The western flower thrips, Frankliniella occidentalis, is invasive and polyphagous. Its visual signal plays a crucial role in host location, suggesting that thrips has diurnal behavior. However, any diel rhythmicity of the thrips behavior is not yet well understood. This study analyzed the diurnal rhythmicity of the feeding and mating behaviors of thrips, and the patterns were explained in terms of diel rhythmicity by assessing the expression of genes associated with the circadian clock. Most feeding behaviors were observed during photophase in both larvae and adults. In mating behavior, which is mainly exhibited in 2~3 days-old young adults after emergence, most adults copulated at photophase. This diurnal pattern of the mating behavior was further supported by the analysis of a temporal calling behavior for mating, which was biased toward photophase. Four genes associated with the circadian clock were predicted in the F. occidentalis genome: period (PER), Timeless (TIM), Doubletime (DBT), and clock (CLK). All these genes exhibited a characteristic diel pattern in their gene expressions in a 24 h-period. The high expression peaks of these genes except for DBT were observed at photophase. To confirm the functional association of these genes with the feeding and mating behaviors, individual RNA interference specific to each gene was executed by feeding the gene-specific double-stranded RNA. Aside from DBT, suppression of these gene expressions resulted in significant deviations of the diel rhythms of the behaviors. These results suggest that the genetic components of the diel rhythmicity mediate the diurnal feeding and mating behaviors of F. occidentalis.
Keywords:
Circadian clock
; Frankliniella occidentalis
; Feeding
; Mating
; Behavior
1. Introduction
The western flower thrips, Frankliniella occidentalis, has been seriously considered as an insect pest in Korea since its invasion in 1993 and its subsequent dispersion to the entire country [1,2]. Its polyphagous feeding behavior underscores its pest severity, causing serious economic damage to high-value crops such as hot pepper [3]. In particular, F. occidentalis is the major thrips in hot pepper that is cultivated in greenhouses, and it transmits a tospovirus called TSWV (tomato spotted wilt virus) [4]. Compared to its related native species (= Frankliniella intonsa), F. occidentalis exhibits a high tolerance in survival and longevity under a high CO2 level (800 ppm), which can sometimes be the case in greenhouses [5]. This suggests that F. occidentalis would be a major thrips infesting various crops that are cultivated in greenhouses. Moreover, it has become more difficult to effectively control the insect pest due to its development of resistance to chemical insecticides [6].
Visual cues and olfactory signals are part of a major sensing system to locate hosts in F. occidentalis, wherein the visual recognition includes specific colors and shapes of objects [7]. Using an LED light source, its color discrimination was further analyzed, and the results suggested that it achieved color detection of three types of photoreceptors specific to blue, green, and UV [8]. In addition, short daylength reduces immature development and adult fecundity of F. occidentalis [9]. These suggest that F. occidentalis exhibits host-searching behavior in the daytime. However, its diel rhythmicity of the behaviors was still not well understood in genetic component(s).
The circadian clock represents a 24-h cyclic rhythmicity of various physiological processes in biological organisms including insects [10]. Moreover, entrainment to the daily environmental cycle such as the light-dark cycle is a fundamental property of the circadian clock. To adjust the rhythmicity to the light cycles, cryptochrome (CRY), which is a blue light receptor whose sequence is similar to that of DNA photolyases [11], plays a crucial role in the perception of light signal and transmitting the light signal to the other circadian clock components [12]. The circadian clock consists of two oscillatory loops: Period (PER)/Timeless (TIM) oscillatory loop and the CRY loop [13,14]. In each of these loops, Clock (CLK)/Cycle (CYC) acts as a transcriptional activator that promotes PER and TIM, or CRY transcription. The product proteins PER and TIM, or CRY are thought to provide negative feedback to inhibit the transcriptional activator. In the fruit fly, Drosophila melanogaster, CRY is expressed in specific clock neurons in the brain [15]. CRY is then activated by blue light and catalyzes TIM degradation through its protease activity, at which point the circadian clock is reset [16,17]. This resetting mechanism has been also demonstrated in monarch butterflies, which are known to navigate long distances during migration via time-compensated sun compass orientation [18]. Despite the well-known genetic components controlling circadian rhythm, little was known in the genetic factors modulating the behaviors such as feeding and mating in F. occidentalis.
The genome of F. occidentalis was first found in Thysanoptera, and it has 415.8 Mb encoding 16,859 genes [19]. The genome information allows us to investigate genes associated with circadian rhythm. This study demonstrated the diel rhythmicity of feeding and mating behaviors of F. occidentalis. To clarify their diel rhythmicity, circadian clock-associated genetic components were obtained from the annotated genome of the species and assessed in terms of their expression patterns. Moreover, the RNA interference (RNAi) of the circadian components followed by behavioral analysis supported the circadian rhythmicity of the feeding and mating behaviors.
2. Materials and Methods
2.1. Insect rearing
Both larvae and adults of F. occidentalis were obtained from the Department of Crop Protection, National Institute of Agricultural Sciences (Jeonju, Korea), and maintained at conditions of 25 ± 1°C temperature, 60 ± 5 % relative humidity, and a 14:10 h (L:D) light cycle without dawn and dusk transitions. Newly germinated beans (Phaseolus coccineus L.) were supplied ad libitum for immature growth and adult reproduction. Eggs that were newly laid on the beans in adult colonies were transferred to the breeding dish (10 cm diameter × 10 cm height, SPL Life Science, Seoul, Korea). After 3 days, at which point most larvae hatched, new beans were supplied every day. Under the laboratory conditions, larvae underwent two instars (L1 - L2) and were distinct from prepupae or pupae that developed wing pads.
2.2. Feeding behavior assay
To determine the feeding behavior of larvae and adults, beans were supplied as a diet in the breeding dish covered with a wet filter paper (5 cm diameter, Whatman, Maidstone, UK). After the test thrips were introduced, they exhibited feeding behavior by inserting the stylets into the diet and keeping stand-still position which was observed under a stereomicroscope (EZ4, Leica, Wetzlar, Germany). This behavior was regarded as feeding and counted at 1-hour intervals. Each experimental unit consisted of 20 test insects. During each time interval, the positive insects exhibiting feeding behavior were counted and used to calculate the feeding frequency in percentage. Each experimental unit was independently replicated three times.
2.3. Mating behavior assay
Before measuring the mating behavior, pupae were separated from the colony and individually transferred to a 1.5 mL microtube (Axygen, Union City, CA, USA) to avoid mating. After measuring the emergence time, a diet of bean (0.2 × 0.2 cm) was provided in the tube. A pair of male and female unmated adults at 48-72 h after emergence was placed on a cap of 1.5 mL microtube, which served as an arena, and covered with cover glass (18 × 18 mm, Marienfeld, Königshofen, Germany) for observation under a stereomicroscope. Each mating behavior was examined for 10 min after the introduction of both males and females under the conditions of 25 ± 1°C temperature and 60 ± 5% relative humidity in different light-dark regimes. Mating behavior under the dark regime was observed using an infrared light (Infrared light 150W, Couyor, China). Copulation behavior within 10 min in the arena was recorded as a successful mating. Each observation was replicated 10 times with every fresh adult.
2.4. Y-tube olfactometer assay
This experiment used a Y-tube olfactometer containing the main length (10 cm) with two arms (9 cm each) separated by a 45° angle. The Y-tube was placed in a dark room with infrared light (SE601, Qingdao Hanil, Pingdu, China) at room temperature. During this experiment, the air was filtered by a charcoal filter at a rate of 0.6 L/min (Power Air Pump, Seoul, Korea) and passed through the two glass vessels, then entered each of the arms. A test chamber was connected to a vessel containing the test thrips (N = 50, 2-3 day-old adults). The other control had an empty chamber. A run (= an experimental unit) used 50 thrips (the same age of adults) and was replicated three times. The positive responses of thrips were recorded by having them pass through the arms and reach over 5 cm from the crossroad. The scoring was made at 10 min. After each run, the Y-tube was cleaned with 70% ethanol.
2.5. Bioinformatic analysis
DNA and amino acid sequences were obtained from NCBI (National Center for Biotechnology Information: https://blast.ncbi.nlm.nih.gov) with accession numbers. MEGA6.0 was used to construct a phylogenetic tree through clustering using Maximum likelihood, where the evolutionary distances were computed using the Poisson correction method. Bootstrapping values were obtained with 1,000 replications to support branching and clustering. Protein domains were predicted using a program searching the conserved domain (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi), and InterPro (https://www.ebi.ac.uk/interpro/). The N-terminal signal peptide was determined using SignalP 5.0 server (https://services.healthtech.dtu.dk/ service.SignalP-5.0/). The resulting domains were drawn by Biorender (https://biorender.com/).
2.6. RNA extraction and cDNA preparation
Total RNAs were extracted using Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. For each RNA extraction, 25 individuals were macerated using 500 μL of Trizol reagent. Following the RNA extraction, RNA was resuspended in 30 μL of diethyl pyrocarbonate (DEPC) and quantified using a spectrophotometer (Nanodrop, Thermo Fisher Scientific, Wilmington, DE, USA). For cDNA synthesis, 400 ng of RNA was used in each sample with RT oligo dT premix (Intron Biotechnology, Seoul, Korea) containing oligo dT primer. A reaction mixture consisted of 2 μL of RNA extract and 18 μL of DEPC-treated water and was run according to the manufacturer’s instructions. The resulting cDNA samples were kept at -20°C before being used for experimentation.
2.7. RT-PCR and RT-qPCR
RT-PCR used the cDNA and amplified the target genes with a Taq polymerase (GeneALL, Seoul, Kore). A reaction mixture (25 μL) for PCR consisted of 2.5 μL of dNTP (each 10 pmol), 2.5 μL of 10× Taq buffer, 2 μL of forward and reverse primers (10 pmol/μL, Table S1), 0.5 μL of Taq polymerase, 1 μL of cDNA, and 16.5 μL of distilled deionized water. The PCR conditions began with an initial denaturation at 95°C for 5 min, which was followed by 35 amplification cycles consisting of 95°C for 1 min, 50 ~ 55°C for 30 sec, and 72°C for 1 min. At the end of the amplification cycle, an additional extension was performed at 72°C for 5 min. The PCR product was confirmed by 1% agarose gel electrophoresis. The qPCR used a Step One Plus Real-Time PCR System (Applied Biosystem) under the guidelines of Bustin et al. [20]. A sample of qPCR reaction (20 µL) contained 10 µL of Power SYBR Green PCR Master Mix (Toyobo, Osaka, Japan), 3 µL of cDNA template (100 ng), and 1 µL (10 pmol) each of the forward and reverse primers (Table S1). After an initial heat treatment at 95°C for 2 min, qPCR was performed with 40 cycles of denaturation at 95°C for 30 s, annealing at 50−55°C for 30 s, and extension at 72°C for 30 s. The transcript levels of elongation factor 1 (EF1) were used as a reference for the normalization of each test sample. Quantitative analysis was conducted using the comparative CT (2-ΔΔCT) method [21]. All experiments were independently replicated three times.
2.8. Measuring expression levels of circadian genes
For test thrips, L2 larvae and three-day-old adults were used. Each RNA extraction at every 2-h interval used 25 individuals and followed the cDNA synthesis and qPCR as described above. Each treatment was independently replicated three times.
2.9. RNA interference (RNAi) treatment and subsequent behavior assays
Template DNA was amplified with gene-specific primers (Table S1) containing T7 promoter sequence (5’-TAATACGACTCACTATAGGGAGA-3’) at the 5’ end. The PCR conditions were as described above. After confirming the PCR product, the resulting PCR product was used to in vitro synthesize double-stranded RNA (dsRNA) encoding F. occidentalis genes using T7 RNA polymerase with NTP mixture at 37°C for 3 h (MEGA script RNAi kit, Ambion, Austin, TX, USA). dsRNA (2,000 ng/10 μL) was mixed with a transfection reagent Metafectene PRO (Biontex, Plannegg, Germany) at a 1:1 (v/v) ratio and incubated at 25°C for 30 min to form liposomes, and it was then supplied with bean to F. occidentalis. Bean was coated with a mixture of 10 μL per 20 thrips and fed for 12 h. A control dsRNA (‘dsCON’) was prepared according to the method outlined by Vatanparast et al. [22]. Feeding and mating behaviors were assessed using the method described above.
2.10. Statistical analysis
Percent data were arcsine-transformed and the subsequent transformed data were confirmed to follow a normal distribution using PROC UNIVARIATE of SAS program [23]. Data obtained from the feeding or mating test were subjected to a one-way analysis of variance (ANOVA) using PROC GLM of the SAS program. The means were compared using the least significant difference (LSD) test at a Type I error of 0.05. Frequency data were analyzed by the mean difference test using PROC FREQ of the SAS program. Circadian rhythmicity of the gene expression was assessed using JTK_CYCLE [24].
3. Results
3.1. Diurnal feeding behavior of F. occidentalis
The feeding behavior of F. occidentalis was characterized by the insertion of their stylets to diet bean in both larvae and adults. Using this characteristic behavior, their diel rhythms were assessed in both developmental stages (Figure 1). The feeding behaviors were more active at photophase than they were at scotophase. This diurnal behavior was significant in larvae (F = 9. 45; df = 1, 22; P = 0.0050) and adults (F = 42.00; df = 1, 22; P < 0.0001).
3.2. Diurnal mating behavior of F. occidentalis
Mating behavior was analyzed in a small arena (Figure 2a) at different times of day. Mating rates of the unmated adults were varied with the adult ages, in which the maximal rates were observed at 2-3 days after adult emergence. Using this active age in mating behavior, the mating rates were monitored at every hour each day (Figure 2b). Most of the mating behaviors significantly (F = 3,114.92; df = 1, 47; P < 0.0001) occurred after light-on in a statistically finding.
3.3. Diurnal calling behavior of F. occidentalis
The diurnal mating behavior was further supported by the calling pattern in a day (Figure 3). Both males and females at 3 days after adult emergence were attractive to both sexes. However, their calling time varied within a day. This calling behavior followed the diel rhythm. Most calls occurred at photophase and decreased at scotophase: female to female (F = 26.34; df = 1,16; P < 0.0001), female to male (F = 28.34; df = 1,16; P < 0.0001), male to male (F = 55.33; df = 1,16; P < 0.0001), male to female (F = 25.94; df = 1,16; P < 0.0001).
3.4. Prediction of four genes associated with circadian clock and their expression profiles
Timeless (TIM), Period (PER), Doubletime (DBT), and Clock (CLK) are known to be some components of the circadian clock for all organisms [15]. To predict their orthologs from F. occidentalis, its genome was interrogated. Four corresponding genes were obtained and compared with other known orthologs (Figure 4a). This phylogenetic analysis of the orthologs indicates that insect orthologs are clustered and distinct from those of other animal systems. All these circadian genes were expressed in different stages of F. occidentalis (Figure 4b).
The expression levels of these circadian genes were assessed at intervals of every 2 h in a day in larvae and adults (Figure 5). All four genes showed diel patterns in their expression levels within a day. The diel patterns did not differ between the larval and adult stages (F = 0.12; df = 1, 239; P = 0.7200). However, the diel patterns differed between the four genes (F = 23.11; df = 3, 239; P < 0.0001). For TIM gene, the expression levels were higher at photophase compared to scotophase in larvae (F = 37.29; df = 1, 24; P < 0.0001) and adults (F = 48.87; df = 1, 24; P < 0.0001). This pattern was also detected in the expression of PER gene in both developmental stages: larvae (F = 22.76; df = 1, 24; P < 0.0001) and adults (F = 54.27; df = 1, 24; P < 0.0001). However, there was no significant different in the expression of DBT gene between photophase and scotophase in both developmental stages: larvae (F = 1.40; df = 1, 24; P = 0.2485) and adults (F = 9.40; df = 1, 24; P = 0.053). For CLK gene, the gene expression was also higher at photophase than at scotophase in both developmental stages: larvae (F = 31.93; df = 1, 24; P < 0.0001) and adults (F = 6.27; df = 1, 24; P = 0.0195). However, the oscillation patterns of the gene expression followed a circadian rhythmicity only in PER gene (P = 0.043 in larvae; P = 0.030 in adults) when they were assessed by a statistical algorithm to time-series data using JTK_CYCLE (Table S2).
3.5. RNAi treatments of circadian genes disrupt the circadian rhythmicity of the feeding and mating behaviors
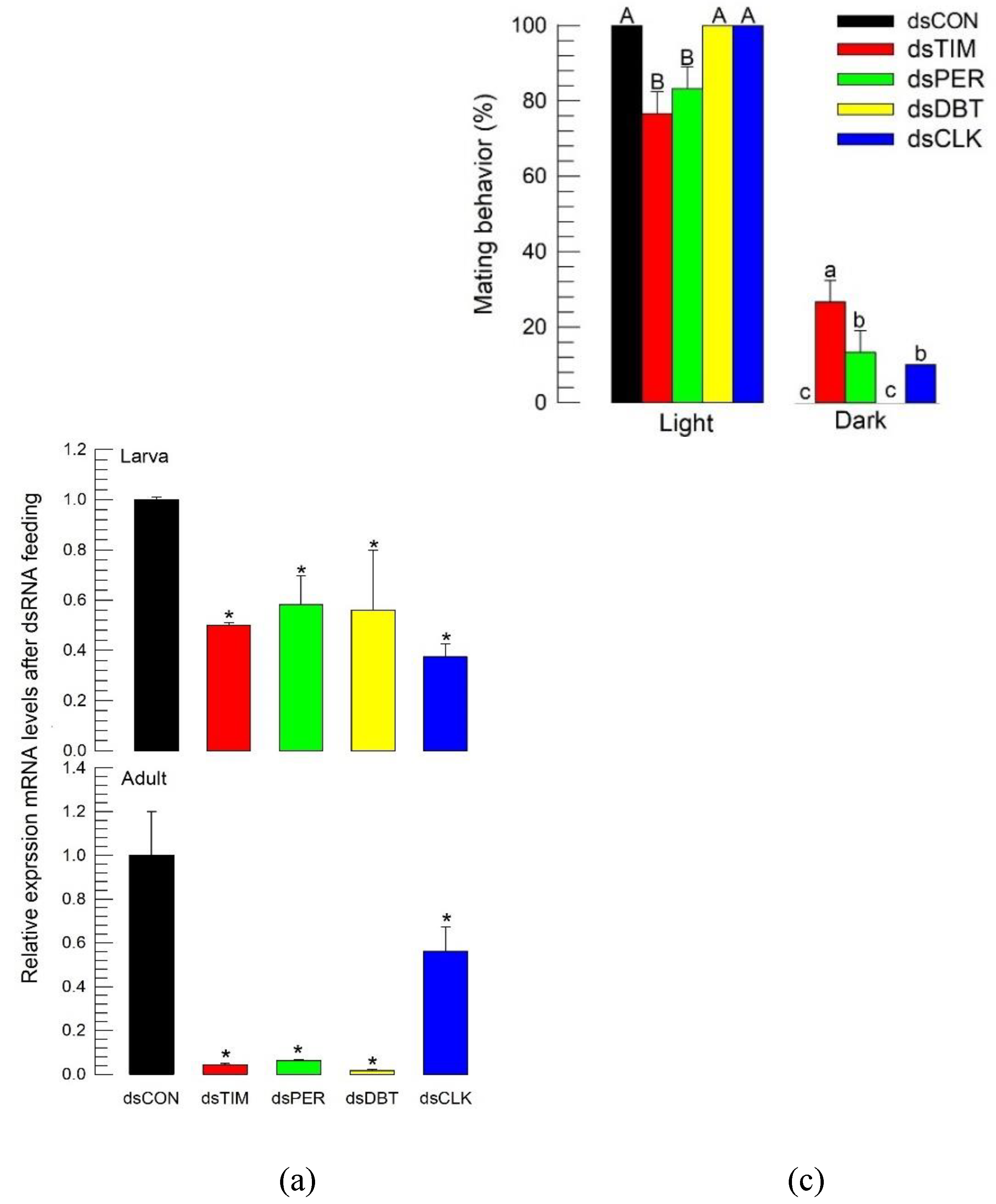
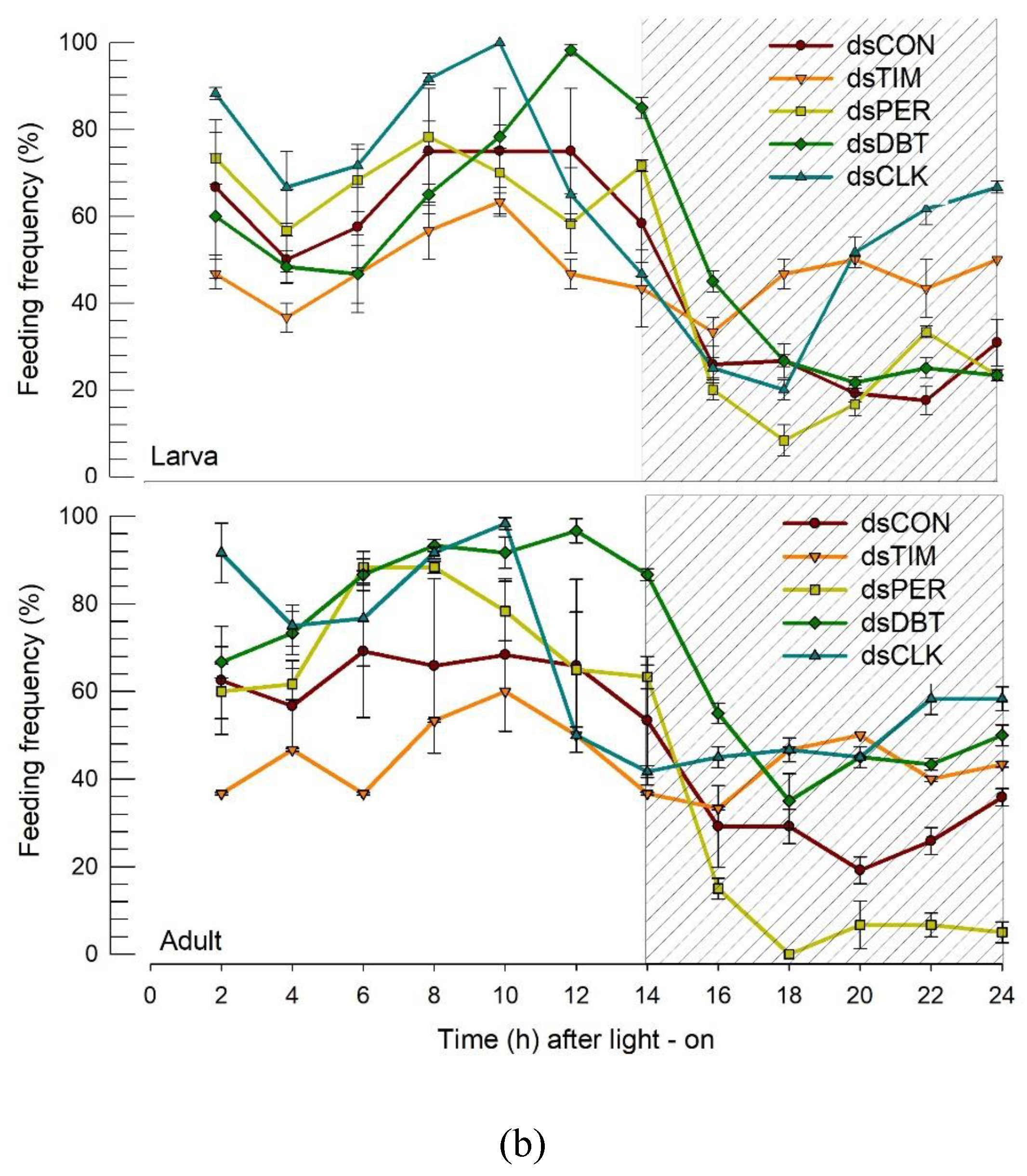
The expression of each circadian gene was suppressed by feeding specific dsRNA (Figure 6a). The RNAi of circadian gene treatment significantly suppressed the gene expression compared to control RNAi: in TIM RNAi, larvae (F = 18.45; df = 1, 4; P = 0.0255) and adults (F = 13.36; df = 1, 4; P = 0.0200), in PER RNAi, larvae (F = 13.22; df = 1, 4; P = 0.0221) and adults (F = 13.86; df = 1, 4; P = 0.0204), in DBT RNAi, larvae (F = 19.89; df = 1, 4; P = 0.0112) and adults (F = 13.47; df = 1, 4; P = 0.00214), and in CLK RNAi, larvae (F = 18.22; df = 1, 4; P = 0.0130) and adults (F = 11.50; df = 1, 4; P = 0.0275). Under some RNAi conditions, the feeding behaviors were altered by decreasing the frequency at photophase while increasing it at scotophase (Figure 6b). For example, RNAi treatments specific to TIM significantly altered the feeding behavior in both developmental stages: larvae (F = 27.07; df = 1, 106; P < 0.0001) and adults (F = 26.45; df = 1, 107; P < 0.0001). In PER RNAi, the altered behavior was observed in adults (F = 13.66; df = 1, 106; P = 0.0003), but not in larvae (F = 0.46; df = 1, 106; P = 0.5007). In DBT RNAi, neither of the stages exhibited behavioral changes: larvae (F = 3.02; df = 1, 106; P = 0.9902) and adults (F = 0.31; df = 1, 106; P = 0.5776). In CLK RNAi, both stages showed significant changes: larvae (F = 7.16; df = 1, 106; P = 0.0086) and adults (F = 9.20; df = 1, 106; P = 0.0030).
The alteration in behavior induced by RNAi treatment was also observed in adult mating behavior (Figure 6c). RNAi treatments against TIM and PER expressions significantly altered the behavior by decreasing mating behavior in photophase and increasing it in scotophase compared to control. In RNAi treatment against DBT expression, there was no significant difference in the behavior compared to control. In RNAi treatment against CLK expression, there was a significant difference in photophase, but not in scotophase.
4. Discussion
The feeding and mating behaviors of F. occidentalis followed a typical diurnal rhythmicity. Both larvae and adults exhibit similar diurnal feeding behavior. Diurnal behavior has been reported in other thrips such as Thrips hawaiiensis, Scirtothrips dorsalis, Frankliniella schultzei, and Megalurothrips usitatus infesting Mango, in which a sequential sampling at an interval of every two hours indicates the most abundant collection at midday [25]. These suggest that the feeding and mating behaviors are under the control of circadian rhythm.
Circadian behavioral rhythms are conserved in virtually all organisms, including plants [26], photosynthetic bacteria [27], Neurospora [28], Drosophila [29], and mice [30]. In particular, at a molecular level, Drosophila demonstrates the negative feedback regulation of transcription as a central theme of circadian rhythms [31]. The first Drosophila clock component that was identified was the PER gene [32]. The second essential pacemaker component that was identified was TIM, and both PER and TIM genes reciprocally autoregulate their transcriptions, where TIM and PER dimerization is crucial for the posttranscriptional regulation and nuclear entry of both proteins [33,34]. By contrast, both CLK and CYC belong to the basic helix-loop-helix (bHLH)–PAS (Per-Arnt-Sim) transcription factor family, the members of which are involved in a wide range of other life processes [35]. Drosophila mutants in CLK and CYC genes exhibit a retardation in the rate of transcription of the two major clock components, PER and TIM [36]. Further, CLK and CYC were found to bind the promoter containing the 21-bp E-box-containing sequence of PER gene [37]. CLK-CYC dimer was also found to activate transcription from promoters containing four copies of the 18-bp E-box-containing sequence from both the PER and TIM promoters in Drosophila [38]. Moreover, two phosphorylation sites located in a putative nuclear localization signal on Drosophila PER protein are phosphorylated by DBT, and these regulate the daily changes in PER stability [39]. In our current study, these four circadian components were identified from the F. occidentalis genome and expressed in different development stages.
The light signal triggered the feeding and mating behaviors and induced the expression of TIM and PER genes in F. occidentalis. For mating analysis, we used unmated male and female adults to avoid any biased behavior due to previous mating [40]. Both male and female adults were attracted to the virgin males or females. Even though males release attractants to males and females for mating [41], a recent study demonstrated that females also produce and release the aggregation pheromone in F. occidentalis [42]. The light cycle is the most powerful Zeitgeber entraining the circadian clock in most organisms. Insects use CRY and/or the compound eye for the light perception necessary for photic entrainment [43]. The molecular mechanism underlying CRY-dependent entrainment is currently well understood. In the cricket Gryllus bimaculatus, RNAi of TIM gene expression in the photic entrainment mechanism disrupted the entrainment to light-dark cycles, as TIM protein levels showed daily cycling characterized by an increase during the night and reduction with exposure to light [44]. In our current study, the increase of TIM and PER expression of F. occidentalis during photophase is explained by their autoregulation through the induction of their gene expressions by the derepressed CYC-CLK dimer in the nucleus based on the circadian clock of Drosophila model [45]. Whittaker and Kirk [46] also reported the increase of the thrips’ behavior with increase of daylength, but they observed the significant decrease in 24-h constant light condition. This suggests the light-dark Zeitgeber entraining in the diel rhythmicity of the thrips.
Through oral administration via diet, specific dsRNAs against the four genes associated with circadian rhythm effectively suppressed their specific target genes. This led to alterations of feeding behavior when TIM or PER genes were suppressed by their specific RNAi treatments. Specific suppression of target genes through RNAi offers an efficient strategy with which to address the target gene function associated with circadian rhythm. Indeed, the dietary RNAi has been reported to be effective against F. occidentalis [47]. In our current study, the time shifting of the mating behavior by RNAi treatments may lead to a mismatch in locating the opposite sex and thus preventing the reproduction of diploid female progeny. Thus, the RNAi technique via diet feeding, especially of bean leaf, allows for novel tactics of F. occidentalis for sustainable agriculture, because it allows for greater specificity than chemical insecticides [48]. The ingestion of dsRNA can be costly because effective RNAi to kill the thrips requires continuous and repeated exposures with large amounts of dsRNA in this kind of environmental RNAi [49].
Altogether, the feeding and mating behaviors of F. occidentalis exhibit a typical diel rhythm, which is active at photophase. This diel rhythm is associated with PER and TIM gene expressions because their RNAi treatments altered the diel pattern. These results support the circadian control of the feeding and mating behaviors in F. occidentalis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Y.K.; methodology, C.K. and S.A.; software, C.K.; validation, C.K. and S.A.; formal analysis, C.K.; investigation, C.K. and Y. K.; resources, Y.K.; data curation, C.K.; writing—original draft preparation, C.K., S.A. and Y.K.; writing—review and editing, Y.K.; visualization, C.K.; supervision, Y.K.; project administration, Y.K.; funding acquisition, Y.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Cooperative Research Program for Agriculture Science & Technology Development funded by Rural Development Administration, Republic of Korea, grant number PJ01578901.
Data Availability Statement
All data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Woo, K.S.; Ahn, S.B.; Lee, S.H.; Kwon, H.M. First record of Frankliniella occidentalis and its distribution and host plants in Korea. Korean J. Appl. Entomol. 1994, 33, 127. [Google Scholar]
- Lee, G.S.; Lee, J.H.; Kang, S.H.; Woo, K.S. Thrips species (Thysanoptera: Thripidae) in winter season and their vernal activities on Jeju island, Korea. J. Asia Pac. Entomol. 2001, 4, 115–122. [Google Scholar] [CrossRef]
- Lim, U.T.; Kim, E.; Mainali, B.P. Flower model traps reduced thrips infestations on a pepper crop in field. J. Asia Pac. Entomol. 2013, 16, 143–145. [Google Scholar] [CrossRef]
- Kim, C.Y.; Choi, D.Y.; Kang, J.H.; Ahmed, S.; Kil, E.J.; Kwon, G.M.; Lee, G.S.; Kim, Y. Thrips infesting hot pepper cultured in greenhouses and variation in gene sequences encoded in TSWV. Korean J. Appl. Entomol. 2021, 60, 381–401. [Google Scholar]
- Shuqi, H.; Ying, L.; Lei, Q.; Zhihua, L.; Chao, X.; Lu, Y.; Furong, G. The influence of elevated CO2 concentration on the fitness traits of Frankliniella occidentalis and Frankliniella intonsa (Thysanoptera: Thripidae). Environ. Entomol. 2017, 46, 722–728. [Google Scholar] [CrossRef]
- Bhuyain, M.M.H.; Lim, U.T. Relative susceptibility to pesticides and environmental conditions of Frankliniella intonsa and F. occidentalis (Thysanoptera: Thripidae), an underlying reason for their asymmetrical occurrence. PLoS ONE 2020, 15, e0237876. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Wu, S.; Xing, Z.; Xu, R.; Cai, W.; Lei, Z. Behavioral responses of western flower thrips (Frankliniella occidentalis) to visual and olfactory cues at short distances. Insects 2020, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Stukenberg, N.; Pietruska, M.; Waldherr, A.; Meyhöfer, R. Wavelength-specific behavior of the western flower thrips (Frankliniella occidentalis): evidence for a blue-green chromatic mechanism. Insects 2020, 11, 423. [Google Scholar] [CrossRef] [PubMed]
- Brødsgaard, H.F. Effect of photoperiod on the bionomics of Frankliniella occidentalis (Pergande) (Thysanoptera, Thripidae). J. Appl. Entomol. 1994, 117, 498–507. [Google Scholar] [CrossRef]
- Crespo-Flores, S.L.; Barber, A.F. The Drosophila circadian clock circuit is a nonhierarchical network of peptidergic oscillators. Curr. Opin. Insect Sci. 2022, 52, 100944. [Google Scholar] [CrossRef]
- Lin, C.; Todo, T. The cryptochromes. Genome Biol. 2005, 6, 220. [Google Scholar] [CrossRef]
- Emery, P.; So, W.V.; Kaneko, M.; Hall, J.C.; Rosbash, M. CRY, a Drosophila clock and light-regulated cryptochrome, is a major contributor to circadian rhythm resetting and photosensitivity. Cell 1998, 95, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Tokuoka, A.; Itoh, T.Q.; Hori, S.; Uryu, O.; Danbara, Y.; Nose, M.; Bando, T.; Tanimura, T.; Tomioka, K. cryptochrome genes form an oscillatory loop independent of the per/tim loop in the circadian clockwork of the cricket Gryllus bimaculatus. Zool. Lett. 2017, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, K.; Matsumoto, A. The circadian system in insects: cellular, molecular, and functional organization. Adv. Insect Physiol. 2019, 56, 73–115. [Google Scholar]
- Emery, P.; Stanewsky, R.; Hall, J.C.; Rosbash, M. Drosophila cryptochromes: a unique circadian-rhythm photoreceptor. Nature 2000, 404, 456–457. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Parikh, V.; Itsukaichi, T.; Bae, K.; Edery, I. Resetting the Drosophila clock by photic regulation of PER and a PER-TIM complex. Science 1996, 271, 1740–1744. [Google Scholar] [CrossRef] [PubMed]
- Ceriani, M.F.; Darlington, T.K.; Staknis, D.; Mas, P.; Petti, A.A.; Weitz, C.J.; Kay, S.A. Light-dependent sequentation of TIMELESS by CRYPTOCHROME. Science 1999, 285, 553–556. [Google Scholar] [CrossRef]
- Zhu, H.; Sauman, I.; Yuan, Q.; Casselman, A.; Emery-Le, M.; Emery, P.; Reppert, S.M. Cryptochromes define a novel circadian clock mechanism in monarch butterflies that may underlie sun compass navigation. PLoS Biol. 2008, 6, e4. [Google Scholar] [CrossRef]
- Rotenberg, D.; Baumann, A.A.; Ben-Mahmoud, S.; Christiaens, O.; Dermauw, W.; Ioannidis, P.; Jacobs, C.G.C.; Vargas Jentzsch, I.M.; Oliver, J.E.; Poelchau, M.F.; et al. Genome-enabled insights into the biology of thrips as crop pests. BMC Biol. 2020, 18, 142. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data analysis using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vatanparast, M.; Ahmed, S.; Herrero, S.; Kim, Y. A non-venomous sPLA2 of a lepidopteran insect: its physiological functions in development and immunity. Dev. Comp. Immunol. 2018, 89, 83–92. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute Inc. SAS/STAT user’s guide, Release 6.03, Ed. Cary, NC. 1989.
- Hughes, M.E.; Hogenesch, J.B. Kornacker, K. JTK_CYCLE: an efficient nonparametric algorithm for detecting rhythmic components in genome-scale data sets. J. Biol. Rhythms 2010, 25, 372–380. [Google Scholar] [CrossRef]
- Aliakbarpour, H.; Rawi, C.S. Diurnal activity of four species of thrips (Thysanoptera: Thripidae) and efficiencies of three nondestructive sampling techniques for thrips in mango inflorescences. J. Econ. Entomol. 2010, 103, 631–640. [Google Scholar] [CrossRef]
- Millar, A.J.; Kay, S.A. The genetics of phototransduction and circadian rhythms in Arabidopsis. BioEssays 1997, 19, 209–214. [Google Scholar] [CrossRef]
- Ishiura, M.; Kutsuna, S.; Aoki, S.; Iwasaki, H.; Andersson, C.R.; Tanabe, A.; Golden, S.S.; Johnson, C.H.; Kondo, T. Expression of a gene cluster kaiABC as a circadian feedback process in cyanobacteria. Science 1998, 281, 1519–1523. [Google Scholar] [CrossRef]
- Dunlap, J.C. Genetics and molecular analysis of circadian rhythms. Annu. Rev. Genet. 1996, 30, 579–601. [Google Scholar] [CrossRef] [PubMed]
- Rosbash, M.; Allada, R.; Dembinska, M.E.; Guo, W.Q.; Le, M.; Marrus, S.B.; Qian, Z.; Rutila, J.E.; Yaglom, J.; Zeng, H. A Drosophila circadian clock. Cold Spring Harbor, Cold Spring Harbor Laboratory Press, NY, pp. 265–278. 1996.
- Vitaterna, M.H.; King, D.P.; Chang, A.M.; Kornhauser, J.M.; Lowrey, P.L.; McDonald, J.D.; Dove, W.F.; Pinto, L.H.; Turek, F.W.; Takahashi, J.S. Mutagenesis and mapping of a mouse gene, Clock, essential for circadian behavior. Science 1994, 264, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Hardin, P.E.; Hall, J.C.; Rosbash, M. Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature 1990, 343, 536–540. [Google Scholar] [CrossRef]
- Reddy, P.; Zehring, W.A.; Wheeler, D.A.; Pirrotta, V.; Hadfield, C.; Hall, J.C.; Rosbash, M. Molecular analysis of the period locus in Drosophila melanogaster and identification of a transcript involved in biological rhythms. Cell 1984, 38, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Vosshall, L.B.; Price, J.L.; Sehgal, A.; Saez, L.; Young, M.W. Specific block in nuclear localization of period protein by a second clock mutation, timeless. Science 1994, 263, 1606–1609. [Google Scholar] [CrossRef]
- Saez, L.; Young, M.W. Regulation of nuclear entry of the Drosophila clock proteins PERIOD and TIMELESS. Neuron. 1996, 17, 911–920. [Google Scholar] [CrossRef]
- Rutila, J.E.; Suri, V.; Le, M.; So, W.V.; Rosbash, M.; Hall, J.C. CYCLE is a second bHLH-PAS protein essential for circadian transcription of Drosophila period and timeless. Cell 1998, 93, 805–814. [Google Scholar] [CrossRef]
- Allada, R.; White, N.E.; So, W.V.; Hall, J.C.; Rosbash, M. A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless. Cell 1998, 93, 791–804. [Google Scholar] [CrossRef]
- Gekakis, N.; Staknis, D.; Nguyen, H.B.; Davis, C.F.; Wilsbacher, L.D.; King, D.P.; Takahashi, J.S.; Weitz, C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science 1998, 280, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Darlington, T.K.; Wager-Smith, K.; Ceriani, M.F.; Staknis, D.; Gekakis. N.; Steeves, T.D.L.; Weitz, C.J.; Takahashi, J.S.; Kay, S.A. Closing the circadian loop: CLOCK-induced transcription of its own inhibitors per and tim. Science 1998, 280, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Price, J.L.; Blau, J.; Rothenfluh, A.; Abodeely, M.; Kloss, B.; Young, M.W. double-time is a novel Drosophila clock gene that regulates PERIOD protein accumulation. Cell 1998, 94, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Akinyemi, A.O.; Kirk, W.D.J. Experienced males recognise and avoid mating with non-virgin females in the western flower thrips. PLoS ONE 2019, 14, e0224115. [Google Scholar] [CrossRef] [PubMed]
- de Kogel, W.J.; van Deventer, P. Intraspecific attraction in the western flower thrips, Frankliniella occidentalis; indications for a male sex pheromone. Entomol. Exp. Appl. 2003, 107, 87–89. [Google Scholar] [CrossRef]
- Khan, F.; Kim, K.; Sung, J.; Lim, H.; Kim, S.G.; Choi, M.Y.; Kim, Y. A novel physiological function of pheromone biosynthesis-activating neuropeptide in production of aggregation pheromone. Sci. Rep. 2023, 13, 5551. [Google Scholar] [CrossRef]
- Foley, L.E.; Emery, P. Drosophila cryptochrome: variations in blue. J. Biol. Rhythms 2020, 35, 16–27. [Google Scholar] [CrossRef]
- Moriyama,Y. ; Takeuchi, K.; Shinohara, T.; Miyagawa, K.; Matsuka, M.; Yoshii, T.; Tomioka, K. Timeless plays an important role in compound eye-dependent photic entrainment of the circadian rhythm in the cricket Gryllus bimaculatus. Zool. Sci. 2022, 39, 397–405. [Google Scholar]
- Liu, T.; Mahesh, G.; Yu, W.; Hardin, P.E. CLOCK stabilizes CYCLE to initiate clock function in Drosophila. Proc. Natl. Acad. Sci. USA, 2017, 114, 10972–10977. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, M.S.; Kirk, W.D.J. The effect of photoperiod on walking, feeding, and oviposition in the western flower thrips. Entomol. Exp. Appl. 2004, 2004. 111, 209–214. [Google Scholar] [CrossRef]
- Andongma, A.A.; Greig, C.; Dyson, P.J.; Flynn, N.; Whitten, M.M.A. Optimization of dietary RNA interference delivery to western flower thrips Frankliniella occidentalis and onion thrips Thrips tabaci. Arch. Insect Biochem. Physiol. 2020, 103, e21645. [Google Scholar] [CrossRef] [PubMed]
- Whitten, M.; Dyson, P. Gene silencing in non-model insects: overcoming hurdles using symbiotic bacteria for trauma-free sustainable delivery of RNA interference. BioEssays 2017, 39, 1600247. [Google Scholar] [CrossRef]
- Zotti, M.; Smagghe, G. RNAi technology for insect management and protection of beneficial insects from diseases: lessons, challenges, and risk assessments. Neotrop. Entomol. 2015, 44, 197–213. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Diel rhythm of feeding behaviors in larvae (upper) and adults (lower) of F. occidentalis. Feeding behavior was determined by the stylet insertion to diet bean at an interval of every hour. Ten individuals were observed at each time point and replicated three times. Light:Dark cycle was 14:10 h. Temperature was 25 ± 2°C.
Figure 1.
Diel rhythm of feeding behaviors in larvae (upper) and adults (lower) of F. occidentalis. Feeding behavior was determined by the stylet insertion to diet bean at an interval of every hour. Ten individuals were observed at each time point and replicated three times. Light:Dark cycle was 14:10 h. Temperature was 25 ± 2°C.
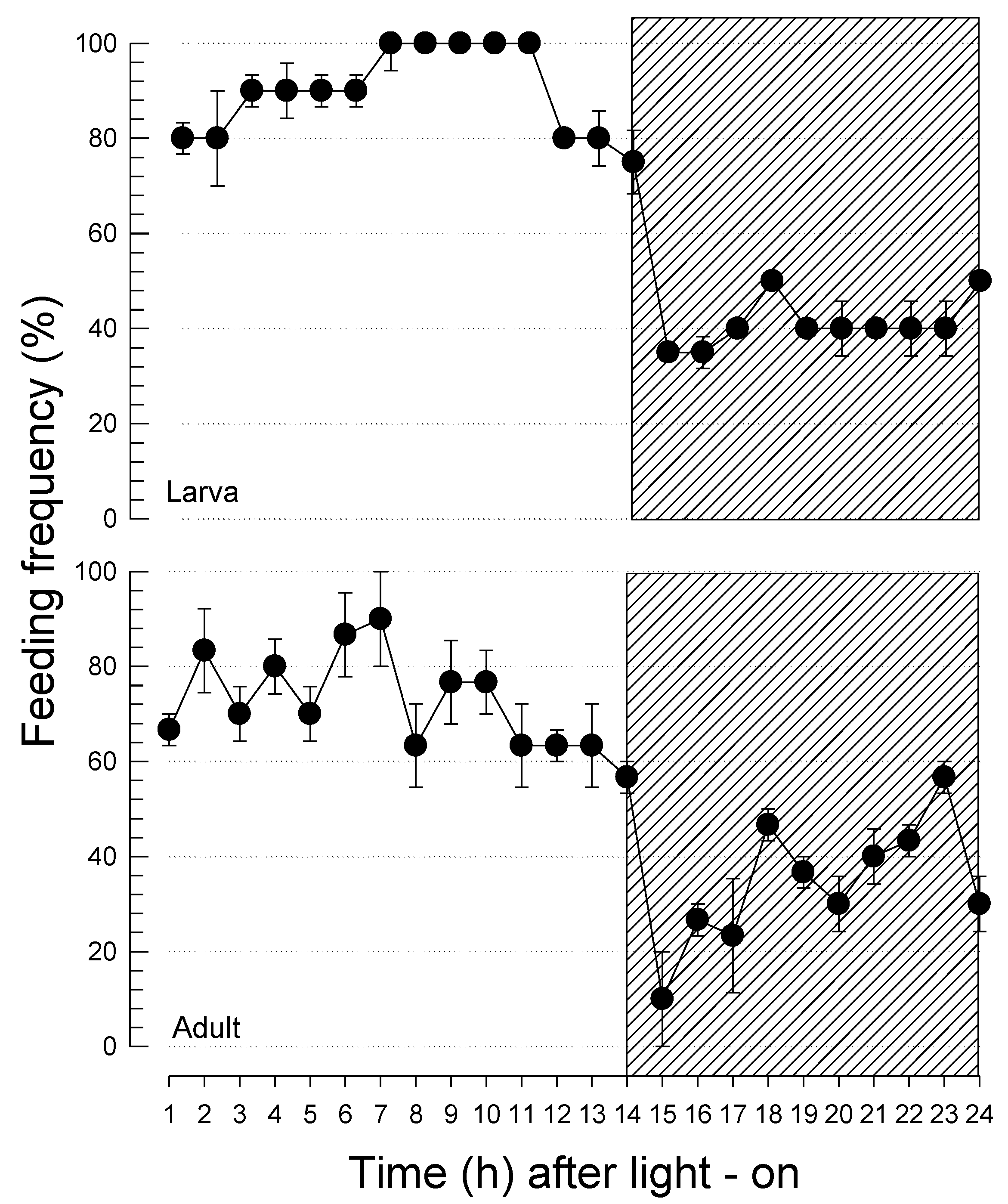
Figure 2.
Diel rhythm of mating behavior in F. occidentalis adults. Mating behavior was determined by copulation between a male and a female in a dish arena (1 cm diameter). Ten pairs were observed at each time point and replicated three times. (A) Variation of mating frequency at different adult ages. Behavioral assessments were performed at photophase. (B) Diurnal mating behavior of three-day-old adults. The assessments were performed at an interval of every hour during photophase and an interval of every three hours at scotophase. Light:Dark cycle was 14:10 h. Temperature was 25 ± 2°C.
Figure 2.
Diel rhythm of mating behavior in F. occidentalis adults. Mating behavior was determined by copulation between a male and a female in a dish arena (1 cm diameter). Ten pairs were observed at each time point and replicated three times. (A) Variation of mating frequency at different adult ages. Behavioral assessments were performed at photophase. (B) Diurnal mating behavior of three-day-old adults. The assessments were performed at an interval of every hour during photophase and an interval of every three hours at scotophase. Light:Dark cycle was 14:10 h. Temperature was 25 ± 2°C.
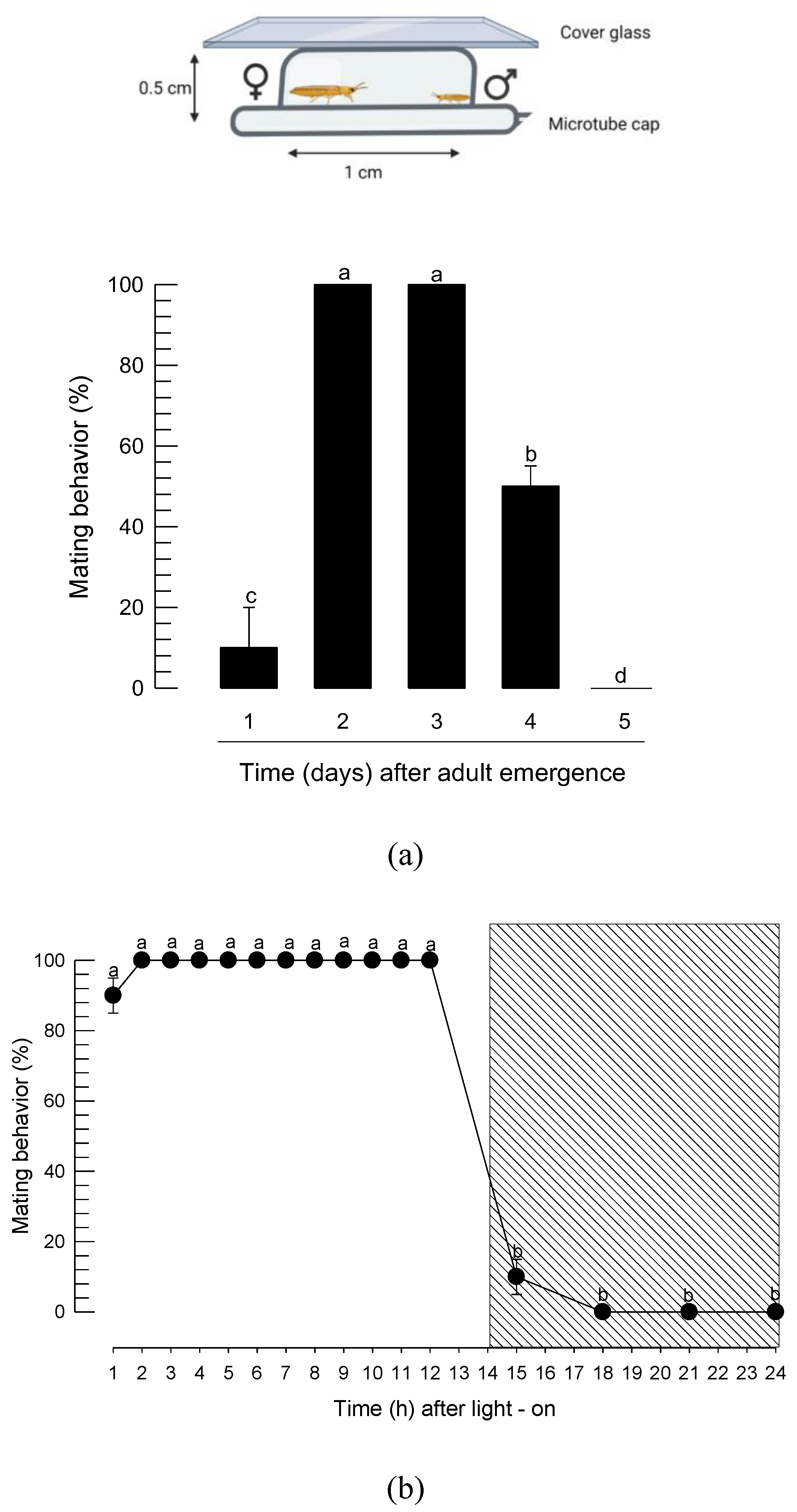
Figure 3.
Diel rhythm of calling for mating. (A) Diagram of Y-tube olfactometer experiment. Test insects (‘T1’) for sending calling signals were inserted into the A or B chamber. Responding insects (‘T2’) (N = 50) were added to the base of the Y tube. The experiment was run at dark condition under infrared light. Each treatment was replicated three times. (B) Calling pattern in a day. Different letters above the standard deviation bars indicate significant differences among means at Type I error = 0.05 (LSD test).
Figure 3.
Diel rhythm of calling for mating. (A) Diagram of Y-tube olfactometer experiment. Test insects (‘T1’) for sending calling signals were inserted into the A or B chamber. Responding insects (‘T2’) (N = 50) were added to the base of the Y tube. The experiment was run at dark condition under infrared light. Each treatment was replicated three times. (B) Calling pattern in a day. Different letters above the standard deviation bars indicate significant differences among means at Type I error = 0.05 (LSD test).
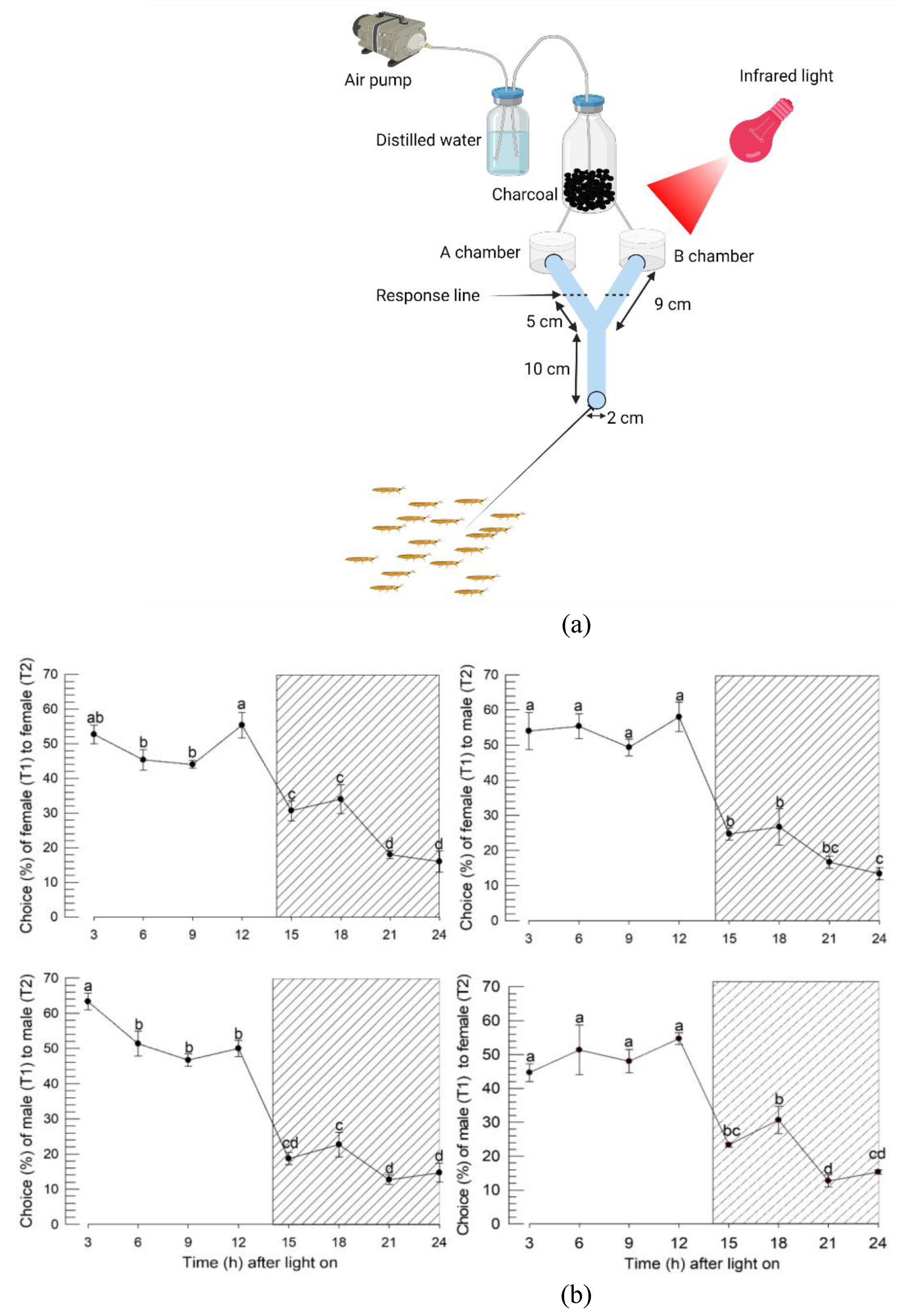
Figure 4.
Phylogenetic analysis of four genes associated with circadian rhythm in F. occidentalis: (A) Timeless (TIM), (B) Period (PER), (C) Doubletime (DBT), and (D) Clock (CLK). The tree was generated by the Neighbor-joining method using MEGA 6.0. Bootstrapping values were obtained with 1,000 repetitions to support branch and clustering. Amino acid sequences were retrieved from GenBank, along with accession numbers. Protein domains were predicted using NCBI conserved Domain and InterPro. The N-terminal signal peptide was determined using the SignalP 5.0 server.
Figure 4.
Phylogenetic analysis of four genes associated with circadian rhythm in F. occidentalis: (A) Timeless (TIM), (B) Period (PER), (C) Doubletime (DBT), and (D) Clock (CLK). The tree was generated by the Neighbor-joining method using MEGA 6.0. Bootstrapping values were obtained with 1,000 repetitions to support branch and clustering. Amino acid sequences were retrieved from GenBank, along with accession numbers. Protein domains were predicted using NCBI conserved Domain and InterPro. The N-terminal signal peptide was determined using the SignalP 5.0 server.
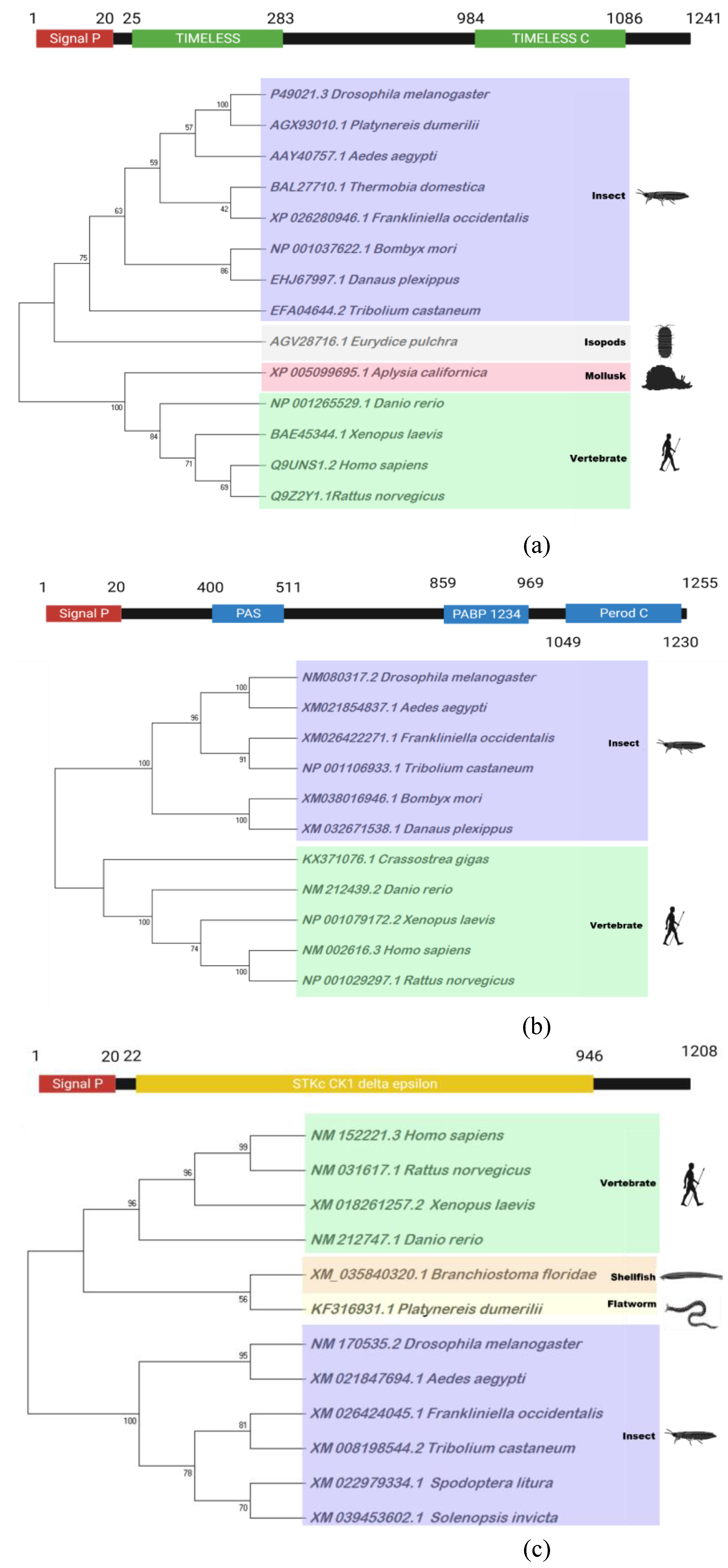
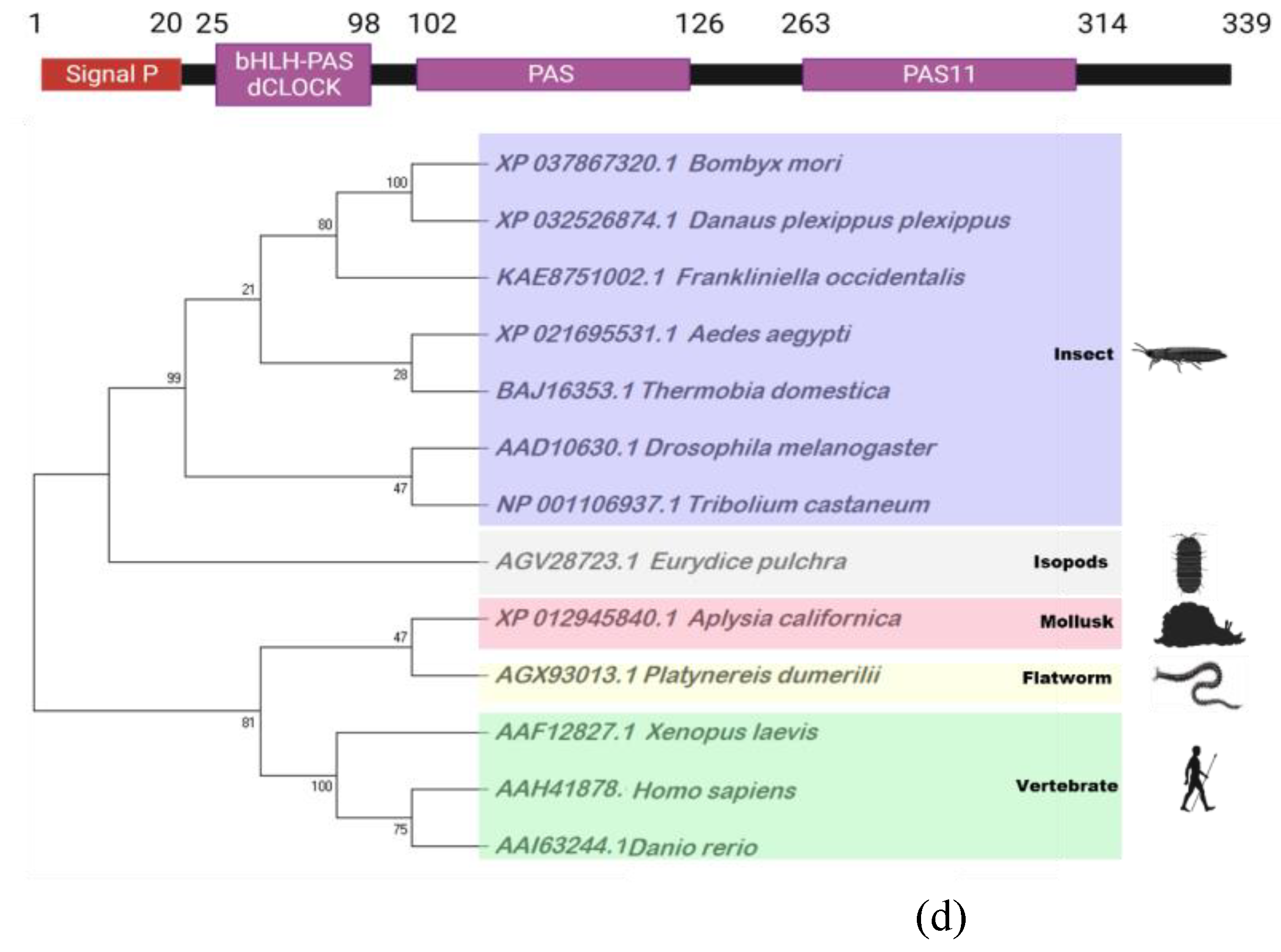
Figure 5.
Expression profile of four genes associated with circadian rhythm in F. occidentalis: Timeless (TIM), Period (PER), Doubletime (DBT), and Clock (CLK). The experimental unit at each time point was 25 thrips to extract RNA, and each unit was independently replicated three times. The assessments were performed at every two-hour interval. The black hatch area indicates scotophase. Light:Dark cycle was 14:10 h. Temperature was 25 ± 2°C. Different letters above the standard deviation bars indicate significant differences among means at Type I error = 0.05 (LSD test).
Figure 5.
Expression profile of four genes associated with circadian rhythm in F. occidentalis: Timeless (TIM), Period (PER), Doubletime (DBT), and Clock (CLK). The experimental unit at each time point was 25 thrips to extract RNA, and each unit was independently replicated three times. The assessments were performed at every two-hour interval. The black hatch area indicates scotophase. Light:Dark cycle was 14:10 h. Temperature was 25 ± 2°C. Different letters above the standard deviation bars indicate significant differences among means at Type I error = 0.05 (LSD test).
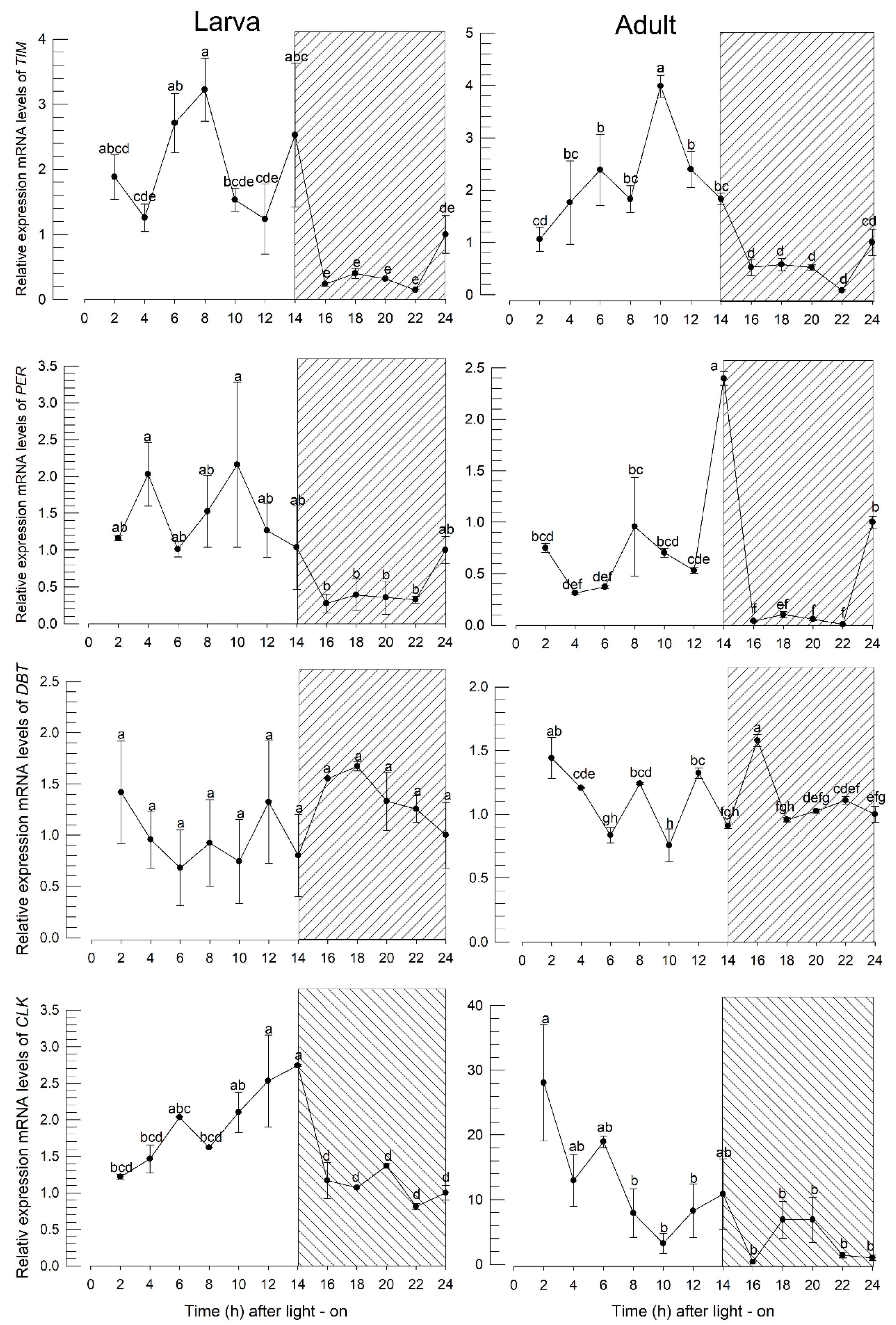
Figure 6.
Alterations of feeding and mating behavior by RNA interference (RNAi) of four circadian clock genes in F. occidentalis: Timeless (TIM), Period (PER), Doubletime (DBT), and Clock (CLK). RNAi was performed by feeding dsRNA (dsTIM, dsPER, dsDBT, or dsCLK) specific to each circadian gene. (a) RNAi efficiency by measuring transcript levels using RT-qPCR at 24 h after dsRNA feeding. Each measurement used 25 thrips to extract RNA for RT-qPCR and was independently replicated three times. An asterisk indicates a significant difference of means compared to control (‘dsCON’) at Type I error = 0.05 (LSD test). (b) Alteration in feeding rhythm after feeding dsRNA. In each time point, 20 thrips as an experimental unit were used to assess feeding behavior. Each measurement was replicated three times. (c) Alteration in mating behavior after feeding dsRNA. Mating behavior was observed at 2 pm (Light) and 2 am (Dark). Ten pairs were observed at each time point and replicated three times. Different letters above the standard deviation bars indicate significant differences among means at Type I error = 0.05 (LSD test) in each light and dark condition.
Figure 6.
Alterations of feeding and mating behavior by RNA interference (RNAi) of four circadian clock genes in F. occidentalis: Timeless (TIM), Period (PER), Doubletime (DBT), and Clock (CLK). RNAi was performed by feeding dsRNA (dsTIM, dsPER, dsDBT, or dsCLK) specific to each circadian gene. (a) RNAi efficiency by measuring transcript levels using RT-qPCR at 24 h after dsRNA feeding. Each measurement used 25 thrips to extract RNA for RT-qPCR and was independently replicated three times. An asterisk indicates a significant difference of means compared to control (‘dsCON’) at Type I error = 0.05 (LSD test). (b) Alteration in feeding rhythm after feeding dsRNA. In each time point, 20 thrips as an experimental unit were used to assess feeding behavior. Each measurement was replicated three times. (c) Alteration in mating behavior after feeding dsRNA. Mating behavior was observed at 2 pm (Light) and 2 am (Dark). Ten pairs were observed at each time point and replicated three times. Different letters above the standard deviation bars indicate significant differences among means at Type I error = 0.05 (LSD test) in each light and dark condition.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



