Submitted:
27 May 2023
Posted:
30 May 2023
You are already at the latest version
Abstract
Production of silkworm silk is the most economically important traits in the silk industry. Silk protein synthesis is regulated by juvenile hormone (JH) and 20-Hydroxyecdysone (20E). Therefore, it is important to understand the genetic regulation of silk production is thus a priority. JH binding protein (JHBP) transports JH from the hemolymph to target organs and cells and protects JH. In a previous study, we identified 41 genes containing a JHBP domain in the Bombyx mori genome. Only one JHBP gene, that is, BmJHBPd2, is highly expressed in the posterior silk gland (PSG) and its function remains unknown. In the present study, we investigated the expression levels of BmJHBPd2 and the major silk protein genes in the high silk-producing practical strain 872 (S872) and the low silk-producing local strain Dazao. Our results have shown that BmJHBPd2 was more highly expressed in S872, than in the Dazao strain, which is consistent with the expression pattern of fibroin genes. A subcellular localization assay indicated that BmJHBPd2 is located in the cytoplasm. In vitro hormone induction experiments showed that BmJHBPd2 was upregulated by treatment with juvenile hormone analogue (JHA). BmKr-h1 upregulation was significantly inhibited by overexpression of BmJHBPd2 at the cell level when induced by JHA. However, overexpression of BmJHBPd2 in the posterior silk gland by transgenic methods led to the inhibition of the expression of the silk fibroin gene, resulting in a reduction in silk yield. Further investigation has shown that in the BmJHBPd2OE individual, the key transcription factor Krüppel homolog 1 (Kr-h1) of the JH signaling pathway was inhibited, and 20E signaling pathway genes, such as broad complex (Brc), E74A, and ultraspiracle protein(USP), were upregulated. Our results have indicated that BmJHBPd2 plays an important role in the JH signaling pathway and was important for silk protein synthesis. Furthermore, our findings have helped to elucidate the mechanisms by which JH regulates silk protein synthesis.
Keywords:
Juvenile hormone
; Juvenile hormone binding protein
; Silk protein
; Bombyx mori
1. Introduction
Juvenile hormone (JH) plays a key role in insect development, metamorphosis, and reproduction of insects [1,2]. Juvenile hormone binding protein (JHBP) is the first key factor in the JH signaling pathway. The JHBP complex or JH-JHBP complex is a vital member of the intricate JH signaling transmission chain that binds to membrane receptors [3,4]. It broadens understanding of the crucial factors in the JH signaling pathway and the mode of action in target organs, and has far-reaching importance in pest control and other practical applications.
JH is a hemicterpenoid compound secreted by the corpora allata [5,6]. JHBP is a carrier protein that occours in hemolymph and cells. This protein transports hormone molecules from the JH synthesis site in the corpora allata to the peripheral target cells and assists JH molecules in entering the circulatory system. It can reduce the non-specific binding of JH [7], and protecting it from enzymatic degradation [8,9]. Intracellular JHBP can be divided into cytoplasmic and nuclear JHBP, which are considered juvenile hormone receptors or are involved in the effect of juvenile hormones on target genes. However, to date, the progress of related research progress has been relatively slow [10,11]. The expression level of JHBP was predominantly expressed in the fat body of the bamboo borer was at the medium level from the 3rd to 5th. The expression level was the highest at the early diapause stage, which continued until the middle diapause stage then decreased until the pupal stage [12]. In melon aphids, RNA interference has been used to silence the expression of JHBP and block the transmission of JH signals, resulting in mortality. This then results in effective pest management [13]. Proteomic analysis of male accessory gland secretions have shown that JHBPs affect female reproduction in oriental fruit flies [14]. These studies have shown that JHBP is involved in metabolism, growth, and reproduction. The JHBPs from this gene family are found in many lepidopteran species and form a separate group from other genes [15,16]. To date, studies on the JHBP have focused on many lepidopteran insects, including the tobacco hornworm (Manduca sexta) [17,18], tobacco budworm (Heliothis virescen) [8,19], greater wax moth (Galleria mellonella) [20], and silkworm (Bombyx mori) [21,22].
The domesticated silkworm is a typical lepidopteran holometabolous insect model organism. As a natural protein fiber, the use of silk has been applied in many fields [23]. Silkworm silk glands primarily synthesize and secrete two types of silk proteins, that is, fibroin and sericin. Fibroin is composed of the heavy chain (Fib-H) and light chain (Fib-L), and glycoprotein protein 25 kDa (P25), is synthesized in the posterior silk gland Silk fibroin [24,25]. In the study of increasing the amount of silk protein, injection, smearing, and feeding on exogenous JH or JH analogs (JHA) can prolong the larval stage of silkworms and increase the RNA and DNA contents of the silk protein gene [26,27,28,29]. The mechanism of action was not clear until recent results had shown that Krüppel homolog 1 (Kr-h1) is involved in the repression of metamorphosis. The silk glands of silkworms were significantly enlarged in transgenic silkworms with Kr-h1 overexpression [30], JH induces extended expression of the Bmfib-H gene, and BmKr-h1 may suppress larval–pupal metamorphosis by activating the expression of B. mori-derived dimmed (Bmdimm). Bmdimm is a transcription factors involved in the regulation of silk gene transcription that activates the transcription of the Bmfib-H gene in B. mori posterior silk gland (PSG) cells [31].
In previous studies, 41 JHBP genes were identified in the silkworms, which contained conserved structures of the binding proteins of juvenile hormone. Microarray data has shown different JHBP gene expression trends in silkworms. The expression of BmJHBP genes was generally higher in the head, integument, midgut, fat body, testes, and ovarise. Expression levels also differed in different tissues. The PSG had a specific and highly expression of JHBPd2 [22]. Our study aimed to investigate the role of JHBPd2 in silk protein synthesis.
2. Results
2.1. Expression of BmJHBPd2 in different silk producing strains
After many years of natural selection and artificial domestication, there have been significant differences in silk production among many silkworm strains. To analyze the mRNA levels of BmJHBPd2 among strains with different silk production, the high silk producing practical S872 and low silk-producing local strain Dazao were selected as materials to further analyze the difference between these two strains. The pupau and cocoons of S872 were substantially larger than those of strain Dazao (Figure S1A). This study investigated the indicators of four main economic characteristics of S872 and Dazao. The cocoon, pupae, and cocoon weight, particularly the shell weight percentage of S872 were significantly higher than those of Dazao (Figure S1A and B). Considering the high efficiency of silk gland synthesis of silk proteins in fifth instar larvae, the expression levels of genes in the PSG were investigated. In this study, we determined the expression levels of major silk protein genes using real-time RT-PCR. Bmfib-H, Bmfib-L and BmP25 were specifically expressed in PSG. The transcript levels of Bmfib-H and Bmfib-L in the S872 PSG was higher than those of Dazao at the 3rd and 5th day of the fifth instar (Figure 1A and B). The BmP25 expression of S872 and Dazao was at the same level (Figure 1C). Differences in the BmJHBPd2 expression were also analyzed. The transcript level of BmJHBPd2 in the S872 PSG was higher than that in Dazao on the 3rd and 5th day of the fifth instar. The expression level of S872 on the 5th day of the fifth instar was significantly higher than that in Dazao (Figure 1D). These results have suggested that BmJHBPd2 may be involved in silk protein synthesis.
2.2. Overexpression of BmJHBPd2 at the cellular level
JHBP plays an important role in JH regulation as a vital response factor in the juvenile hormone pathway and JH regulates silk protein synthesis. To further explore how BmJHBPd2 is involved in silk protein synthesis, the expression of the genes associated with the silk protein synthesis and the genes related to the JH signaling pathway in the exogenous JH induction were analyzed at the cellular level firstly. First of all, the BmJHBPd2 subcellular localization vector was constructed (Figure 2A), and immunofluorescence experiments showed that FLAG-tagged BmJHBPd2 was localized in the cytoplasm (Figure 2B). Those results are consistent with those reported in Li Wei’s research [22], and imply that BmJHBPd2 cannot be secreted from the cells, and plays a physiological role in cells. The qRT-PCR at the nucleic acid levels and western blotting at the protein levels have shown that the intracellular overexpression of BmJHBPd2 was successful (Figure 3A). The BmE cells have the signal transduction pathway for JH [31]. When adding JHA to BmE cells, the expression of BmJHBPd2 was significantly upregulated. JHA significantly upregulated the expression of the early response factor BmKr-h1, suggesting that JHA significantly activated the JH downstream signaling pathway (Figure 3B). When only overexpressing BmJHBPd2, Bmkr-h1 was not expressed nearly, and there was no change in the expression of the gene B. mori methoprene-tolerant 1 (BmMet1) (Figure 3C). However, the upregulated expression level of BmKr-h1 in the control group was higher than that in the experimental group (Figure 3D). This suggested that the overexpressed BmJHBPd2 protein reduced the response of BmE cells to JHA. These results have indicated that overexpression BmJHBPd2 inhibited BmKr-h1 expression and affects signal transmission in the JH signaling pathway. In addition, the expression of Bmfib-H was not detected in all our experiments. These findings have suggested that this may have been caused by the lack of transcription factors specific to silk proteins in the cells.
2.3. Transgenic overexpression of BmJHBPd2 in the silk gland
To further explore the biological function of BmJHBPd2 in the silk gland, a piggyBac transgenic vector was constructed containing a combination of BmJHBPd2 and a Myc foreign label with the Fib-L promoter, Fib-L terminator, and pBac [3xP3 EGFP] (Figure 4A). Transgenic and helper plasmids were injected into 271 pre-blastoderm eggs, of which136 hatched and developed to the adult stage. An EGFP-positive brood was obtained and used to establish the transgenic overexpression line (Figure 4B). The mRNA levels of BmJHBPd2 in the PSG of 3rd day of fifth instar larvae (L53D) of the transgenic and wild-type lines were detected using qRT-PCR. The PSG showed that BmJHBPd2 levels were substantially higher in the transgenic line than in the wild-type line (Figure 4C). To confirm whether BmJHBPd2 with a Myc-tag was synthesized in the transgenic line, proteins were extracted from the PSG of L53D for western blotting. The results have shown that signals were only detected in the BmJHBPd2OE strain with the Myc antibody, but not in the wild-type line (Figure 4D). These results have indicated that BmJHBPd2 was overexpressed in the PSG.
2.4. Overexpression of BmJHBPd2 affects silk synthesis and silk yield
This study investigated the strain overexpressing the BmJHBPd2 gene obtained above, predominantly focusing on the silk gland and silk yield of 5th instar larvae. The BmJHBPd2OE line was raised to the L53D stage, and its silk glands were dissected and observed. There was no pronounced difference in the biological characteristics of the silk glands (Figure 5A). However, we found that the overexpression of BmJHBPd2 resulted in thinner and smaller cocoon shells than those in the WT lines (Figure 5B). Further observation of the cocoon shells of the two lines showed that the whole cocoon weight, cocoon weight, pupau weight, and cocoon shell rate were significantly reduced in the BmJHBPd2OE line (Figure 5C–F). Based on the differences in cocoon shells between the BmJHBPd2OE line and WT line, we chose the L5D3 stage to determine the mRNA levels of silk protein-related transcription factors and silk fibroin genes in both lines. Among the silk fibroins tested, Bmfib-H and Bmfib-L were significantly downregulated at the L5D3 in the BmJHBPd2OE line (Figure 5G and H), except for BmP25, which was not significantly different between the two lines (Figure 5I). Among the silk-related transcription factors, the expression of Bmsage and Bmdimm were significantly downregulated in the BmJHBPd2OE line (Figure S2A and B). However, the expression of Bmsgf-1 was significantly upregulated (Figure S2C). These results have indicated that this study successfully overexpressed the BmJHBPd2 gene, the expression of the silk fibroin gene and silk fibroin-related transcription factors in the BmJHBPd2OE line was significantly reduced, which affected the silk yield.
2.5. Overexpression of BmJHBPd2 led to repressed the silk synthesis by inhibiting Bmkr-h1 expression in the silk glands
JHBPd2 affected the expression of Kr-h1 in the JH pathway at the cellular level. Therefore, the major JH regulatory pathways genes were investigated in the silk glands of the BmJHBPd2OE and WT lines. The relative expression level of the early response factor BmKr-h1 in the JH pathway was significantly reduced in the BmJHBPd2OE line by RT-qPCR (Figure 6A). JH receptors, such as Met1, methoprene-tolerant 2 (Met2), and steroid receptor co-activator (SRC), were also downregulated (Figure 6C–D). The key enzymes of the JH metabolic pathway, juvenile hormone esterase (JHE), and juvenile hormone epoxide hydrolase (JHEH), were significantly downregulated (Figure S3A and B). Therefore, both the major genes in the JH pathway and the JH-degrading enzymes were downregulated, indicating that the JH pathway was affected by overexpression of BmJHBPd2.
Kr-h1 can directly inhibit the biosynthesis of 20-Hydroxyecdysone (20E) and the expression of some early transcript factors in 20E [32]. Therefore, the expression of some early transcription factors in 20E was investigated. In the present study, the early transcription factor B. mori broad complex (BmBrc) of 20E was significantly upregulated (Figure 7A). The relative expression levels of E74A and ultraspiracle protein (USP) were significantly upregulated in the BmJHBPd2OE line (Figure 7B and C). Meanwhile, those of the ecdysteroid receptor (EcR), hormone receptor 3 (HR3), and E75A were not significantly different between the two lines (Figure 7D–F). These results have suggested that overexpression of BmJHBPd2 increases the expression of early transcription factors in the 20E signaling pathway.
3. Discussion
The most valuable aspect of silkworm studies is the silk production [23]. It is important to understand the process of silk protein synthesis to increase silk production[33]. In our study, we found that there is a certain correlation between the expression levels of BmJHBPd2 and silk protein genes and silk yield. At the cellular level, BmJHBPd2 was induced by JHA and suppressed JH signaling by inhibiting the expression of Bmkr-h1. The individual experiments showed that overexpression of BmJHBPd2 promoted the expression of 20E related transcription factors by inhibiting the expression of Bmkr-h1, thereby decreasing the expression of silk protein genes and silk production. Our research results have indicated that BmJHBPd2 plays an important role in regulating the JH signaling in silk glands. Simply increasing the expression level of JHPBd2 cannot increase silk yield, but rather inhibited silk protein synthesis. Our research has provided a reference for future genetic modification to improve silk yield.
As a specific carrier of the endocrine hormone JH in silkworms, JHBP protects and transports synthesized and secreted JH from the corpora allatum [7]. Given that the organs are used for silk protein synthesis and secretion, silk glands grow rapidly during the fifth instar. Although JH was largely absent from the blood of the 5th instar silkworm [34], to date there have been no reports in the literature on whether the 5th instar silk gland of the silkworm contains JH. Based on the functional studies, the findings of this study have suggested that the 5th instar silk gland of the silkworm is likely to contain JH, and that JHBPd2 can play a role in regulating the concentration gradient of JH in the silk gland functioning. The rationale is that the overexpression of JHBPd2 alone in the PSG increases its protein product, and JH entering the silk gland without reaching the JH concentration in the blood binds to the overexpressed JHBPd2. This can result in a decrease in free JH functioning in the silk gland, which, in turn, reduces the expression of the JH signaling pathway Kr-h1. As a key transcription factor connecting the JH and the 20E pathways, Kr-h1 can directly inhibit the biosynthesis of 20E [32], thereby inhibiting insect growth. Kr-h1 also directly binds to the Kr-h1 binding site (KBS) elements of E93, Brc, and E75 promoters to inhibit their expression [35,36,37,38]. During crosstalk between the JH and 20E pathways, Kr-h1 is located upstream of the 20E pathway genes and inhibits their expression. 20E transcription factors, such as Brc, strongly repress silk protein synthesis. Consequently, the expression of silk proteins is reduced, which then leads to decreased silk yield.
The expression pattern of JHBPd2 gene is highly similar to that of the silk fibroin gene, both of which are highly expressed at the 5th instar stage and are mainly expressed in the PSG. This indicates that a close relationship with silk fibroin synthesis [39]. The larval stage of the silkworms was positively correlated with the silk yield [40,41]. The expression of JHBPd2 was significantly higher in high silk yield varieties than in low silk yield varieties, and there was a positive correlation between its expression and silk yield. However, the overexpression of JHBPd2 was found to inhibit silk yield. This study also conducted an in-depth analysis of this issue. This contradictory result provides further evidence that JHBPd2 plays a role in the regulation of JH concentration. The larval stage of high silk yield S872, especially at the end of the 5th instar, is 2–3 days longer than that of the low silk yield variety Dazao. This suggested that high silk yield S872 has more JH in vivo than Dazao, a which can be inferred from applying JH to the silkworm body surface prolonging the developmental time of the silkworm [31]. With more JH in the high silk yield S872, there is a corresponding increase in JH content in the silk gland, which requires more JHBPd2 protein to bind and protect JH. Therefore, the silk glands of the high silk yield S872 had more time to synthesize more silk protein and thus produce more silk. After the overexpression of JHBPd2, the balance of JH concentration in the silk gland was disrupted because the amount of JH in the silkworm was not affected, which inhibited silk protein synthesis. Our results have also indicated that simply changing a gene that is positively associated with silk production may not improve silk yield. with silk yield being a quantitative trait, it is controlled by multiple genes. Varieties with high silk yields can result from artificial selection, which is the result of the synergistic regulation of multiple genes. Altering only one gene, such as JHBPd2 in this study, may disrupt the homeostasis in vivo, which, in turn, inhibits silk protein synthesis. Therefore, further research is required to determine how to improve silk production through genetic manipulation. Further research on the regulatory mechanism of silk protein synthesis is expected to identify the most critical factors affecting silk protein synthesis.
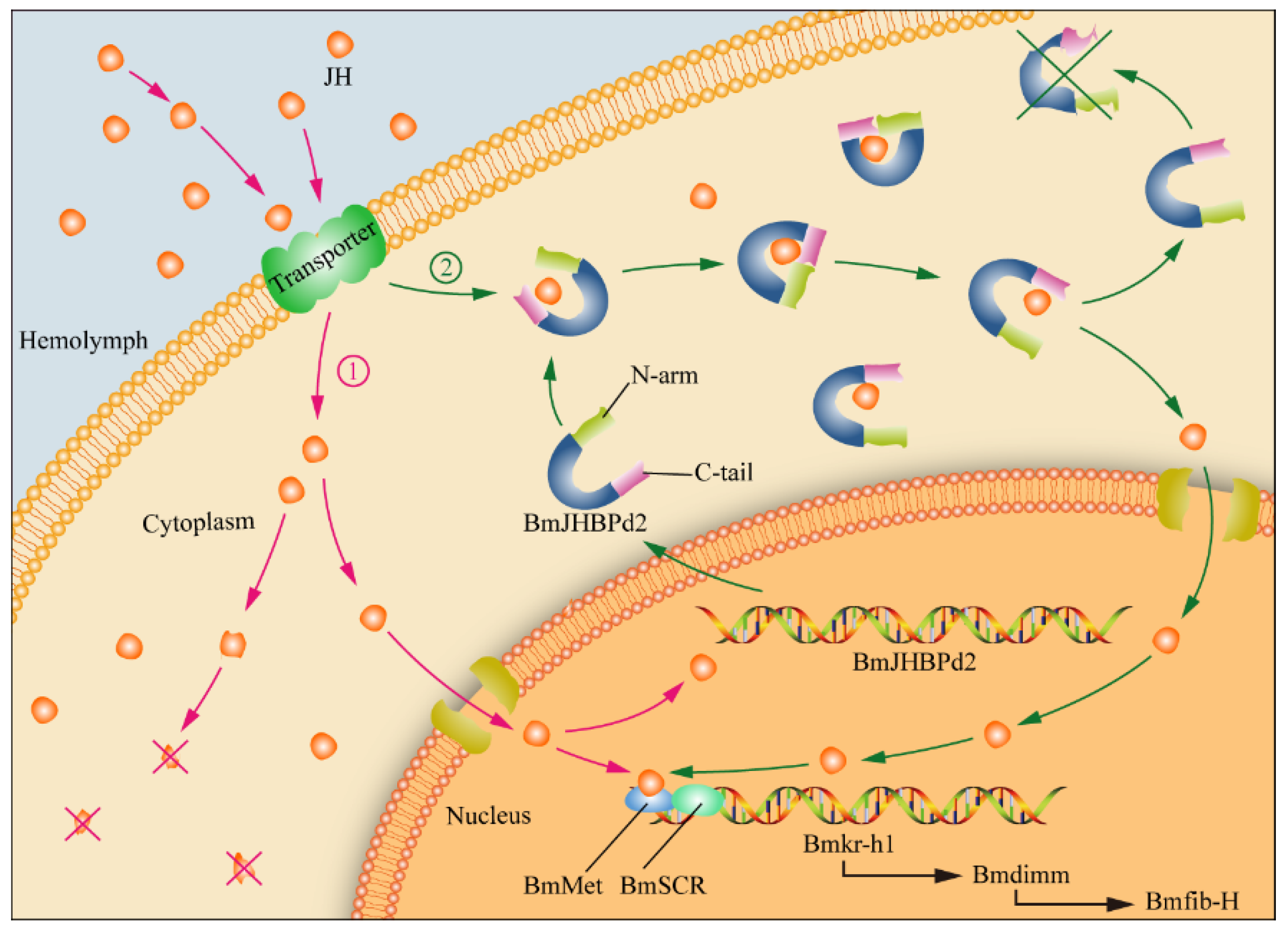
The JH in JHBPd2OE line silk glands was not measured in this study, because there was too little material available. We performed JH assays on normal silk gland tissues and found that the silk glands contained JH (data not published). We have proposed the following regarding the expression of BmJHBPd2 in the PSG (Figure 8). JH is transported into PSG cells from the hemolymph early in the fifth instar stage. Free JH then binds to the nuclear receptor BmMet, and forms a complex with BmSRC [42]. This complex activates the expression of BmKr-h1, and subsequently, the expression of the transcription factor Bmdimm to regulate Bmfib-H [31]. Simultaneously, JH induces the upregulated expression of BmJHBPd2 (Figure 3B). Cytoplasmic BmJHBPd2 can bind to redundant JH and slowly release it to maintain the JH level, which continuously regulates gene expression for silk synthesis. Therefore, there are two potential sources of JH in the silk glands, one of which could originate from the blood. There is a consensus that JH is released into the blood after synthesis by the pharyngeal lateral body, and that JH in the blood is bound by JHBP in the blood and transported to various tissues and organs. However, it is difficult to understand that at 5th instar, JH is essentially undetectable in the blood. Therefore, it is unlikely that other tissues and organs contain JH. However, at the early age of the 5th instar, JH is likely synthesized by the corpora allatum and then transported to other tissues and organs after being bound to JHBP in each organ. Here, JHBP functions as a sponge, which slowly releases JH and regulates the growth and development of each tissue and organ. However, the silk gland may synthesize JH independently. Although the corpora allatum is the main JH-synthesizing organ, the potential for other tissues and organs not synthesizing JH cannot be excluded. In addition, a substantial number of JH synthesis enzymes have been detected in the silk glands. Further confirmation is required to confirm this inference in the later stages.
4. Materials and Methods
4.1. Silkworm strain and cell culture
Bombyx mori, the low silk-producing strain Dazao, and the high silk-producing strain 872 (S872), were provided by the State Key Laboratory of Silkworm Genome Biology, Southwest University in Chongqing, China. The silkworm eggs were cultured at standard temperature of 25℃ under 12 h light and 12 h dark cycle conditions. The larvae were reared with fresh mulberry leaves with 75% relative humidity. The B. mori cell line BmE [43], originally derived from embryo cells, and was maintained at 27℃ in Grace’s medium supplemented with 10% FBS (HyClone).
4.2. RNA preparation and quantitative real time-PCR (qRT-PCR)
Total RNA from the eggs and silk glands was extracted using TRIzol™ reagent (Invitrogen, USA). Reverse transcription was performed using the PrimeScript™ RT reagent Kit with gDNA Eraser (Takara, Japan). Quantitative PCR was performed using SYBR® Premix Ex Taq™ II (Takara) and qPCR reaction under the following conditions:95℃ for 10 s, then 40 cycles of treatment at 95℃ 5 s, 60℃ 31 s. The silkworm ribosomal protein L3 (BmRpl3) was used as the internal marker gene. Three independent replicates were used for each performed.
4.3. Subcellular localization
For the subcellular localization assay of silkworm BmJHBPd2, BmE cells were seeded onto coverslips in 24-well plates. After 12 h of culture, pSLfa1180-Basic, pSLfa1180-GFP, and pSLfa1180-GFP-BmJHBPd2 including the promoter region of the BmActin4 gene from silkworm genomic DNA, fusion protein with FLAG-labeled protein and BmJHBPd2, SV40 polyA (SV40) cloned into the pSLaf1180 vector were separately transfected into BmE cells at 1 µg per well. Cells were transfected with expression plasmids using X-tremeGENE HP DNA Transfection Reagent (Roche Applied Science). After transfection for 48 h, cells were fixed for 10 min at room temperature with 4% (v/v) formaldehyde in PBS. They were then blocked for 30 min in PBS containing 0.1% (w/v) BSA and 5% (v/v) goat serum. The samples were treated with a primary antibody (anti-FLAG monoclonal M2 mouse (Sigma),), for 1 h before being incubated with a secondary antibody (anti-mouse Alexa 488) for 30 min at room temperature. The samples were then mounted using a mounting medium containing 4,6-diamidino-2-phenylindole (DAPI) and photographed using a confocal microscope (Olympus FV1000, Japan).
4.4. Western blot
Radio-immunoprecipitation assay (RIPA) lysis buffer (Beyotime, China) was used to extract proteins from the cells. Protease inhibitors were then added to the supernatants. Protein concentrations were measured using a bicinchoninic acid (BCA) protein assay kit (Beyotime, China). The proteins were separated using10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto a polyvinylidene difluoride(PVDF) membrane (Roche, Switzerland). The PVDF membrane was blocked using 5% skimmed milk overnight at 4℃ and incubated with a primary antibody against BmJHBPd2 for 2 h at 37℃. After washing the PVDF membrane six times at 5 min intervals, the membranes were incubated with goat anti-rabbit IgG labeled with horseradish peroxidase (HRP) (Sigma, USA) secondary antibody and visualized with SuperSignal™ West Femto Maximum Sensitivity Substrate (Thermo Scientific, USA) using the automatic exposing pattern of Genome XRQ (Gene Company, Hong Kong).
4.5. Statistical method for cocoon layer rate
The whole cocoon was weighed and then gently peeled, the pupa was removed, the epidermis was shed, and the remaining cocoon was then weighed. The ratio of this weight to the whole cocoon weight was determined as the cocoon layer ratio, which was measured in 15 transgenic lines and wild-type individuals.
4.6. Plasmid Construction
To construct the transgenic overexpression lines, we used the pBac [3xp3-EGFP] system. JHBPd2 was driven by the posterior silk gland-specific Fib-L promoter to obtain the pBac [3xp3-EGFP-Fib-L-JHBPd2-myc]. The target gene, JHBPd2, was amplified by PCR using cDNA from the silk gland tissue of the Dazao cultivar after the 3rd day of the fifth instar as the template. The 5′ end of BmJHBPd2-F was selected to add the BamH I restriction endonuclease site. The 5′ end of BmJHBPd2-R was selected to add the Not I restriction endonuclease site and Myc foreign label sequence for amplification. Full-length PCR products were digested with BamH I and Not I and connected to the 1180 vector skeleton with the Fib-L promoter to construct the psl1180 [FibL-BmJHBPd2-myc] expression vector. The Asc I enzyme was used to digest psl1180 [FibL-BmJHBPd2-myc] carrier and pBac [3xp3-EGFP] vector. Solution I was used to construct an overexpression vector. The primers used to construct the plasmids are listed in Table S1 in the supplementary materials.
4.7. Silkworm germline transformation
For silkworm germline transformation, the ultrapure plasmid of BmJHBPd2 overexpression transgene vector and helper plasmid were mixed in a 1:1 volume ratio, and the final concentration for embryo injection was 300–500 ng/ μL. After being sealed, they were moved into an artificial culture room at 25℃ and incubated it for 10–12 days until the larvae hatched. After being hatched by injecting the silkworm eggs, the G0 generation could not be screened for transgenic individuals. Males and females were randomly mated and laid eggs for the G1 generation. The G1 generation eggs were fluorescently screened around the sixth day of development. Egg circles with eyes emitting green fluorescence were screened as positive individual egg circles and raised to the 5th instar for molecular-level detection in subsequent experiments.
4.8. Statistical Analysis
All the data were statistically analyzed using an independent-sample t-test. Asterisks indicate significant differences (* P < 0.05, ** P <0.01, *** P < 0.001).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1. Primer sets used in the transgenic overexpression. Table S2. Primer sets used in qRT-PCR. Figure S1. Main economic characters of different silk-producing strain. Figure S2. Expression of silk protein transcription factor in wild type and transgenic lines. Figure S3. Overexpression of BmJHBPd2 results in inhibition of JH catabolic enzyme in silk gland assayed.
Author Contributions
Conceptualization, C.L.; methodology, X.Z, H.Z and W.L.; validation, J.Z and J.L.; formal analysis, J.Z.; visualization, J.Z.; writing—original draft preparation, J.Z, X.Z and C.L.; writing—review and editing, J.Z and C.L.; supervision, C.L.; project administration, C.L.; funding acquisition, C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31972618), the Talent Plan Project of Chongqing (cstc2021ycjh-bgzxm0048), and project of graduate innovation of Chongqing (CYB22159).
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Not applicable
Conflicts of Interest
The authors declare no conflict of interest.
References
- Truman, J. W.; Riddiford, L. M., The origins of insect metamorphosis. Nature 1999, 401, 447-52. [CrossRef]
- Shin, S. W.; Zou, Z.; Saha, T. T.; Raikhel, A. S., bHLH-PAS heterodimer of methoprene-tolerant and Cycle mediates circadian expression of juvenile hormone-induced mosquito genes. Proceedings of the National Academy of Sciences of the United States of America 2012, 109, 16576-81. [CrossRef]
- Wiśniewski, J. R.; Muszyńska-Pytel, M.; Grzelak, K.; Kochman, M., Biosynthesis and degradation of juvenile hormone in corpora allata and imaginal wing discs of Galleria mellonella (L.). Insect Biochemistry 1987, 17, 249-254. [CrossRef]
- Sok, A. J.; Andruszewska, G.; Niewiadomska-Cimicka, A.; Grad, I.; Rymarczyk, G.; Pajdzik, D.; Orłowski, M.; Schmidt, M. T.; Grajek, W.; Ozyhar, A.; Kochman, M., Regulatory elements in the juvenile hormone binding protein gene from Galleria mellonella--topography of binding sites for Usp and EcRDBD. Biochimica et biophysica acta 2008, 1779, 390-401. [CrossRef]
- Röller, D. H.; Dahm, K. H.; Sweeley, C. C.; Trost, B. M., The structure of the juvenile hormone. Angewandte Chemie 2010, 79, 190-191. [CrossRef]
- Gilbert, L. I.; Granger, N. A.; Roe, R. M., The juvenile hormones: Historical facts and speculations on future research directions. Insect biochemistry and molecular biology 2000, 30, 617-44. [CrossRef]
- Hammock, B.; Nowock, J.; Goodman, W.; Stamoudis, V.; Gilbert, L. I., The influence of hemolymph-binding protein on juvenile hormone stability and distribution in Manduca sexta fat body and imaginal discs in vitro. Molecular and cellular endocrinology 1975, 3, 167-84. [CrossRef]
- Sanburg, L. L.; Kramer, K. J.; Kezdy, F. J.; Law, J. H.; Oberlander, H., Role of juvenile hormone esterases and carrier proteins in insect development. Nature 1975, 253, 266-7. [CrossRef]
- Tauchman, S. J.; Lorch, J. M.; Orth, A. P.; Goodman, W. G., Effects of stress on the hemolymph juvenile hormone binding protein titers of Manduca sexta. Insect biochemistry and molecular biology 2007, 37, 847-54. [CrossRef]
- Chang, E. S.; Bruce, M. J.; Prestwich, G. D., Further characterization of the juvenile hormone binding protein from the cytosol of a Drosophila cell line: Use of a photoaffinity label. Insect Biochemistry 1985, 15, 197-204. [CrossRef]
- Shemshedini, L.; Lanoue, M.; Wilson, T. G., Evidence for a juvenile hormone receptor involved in protein synthesis in Drosophila melanogaster. The Journal of biological chemistry 1990, 265, 1913-8. [CrossRef]
- Ritdachyeng, E.; Manaboon, M.; Tobe, S. S.; Singtripop, T., Molecular characterization and gene expression of juvenile hormone binding protein in the bamboo borer, Omphisa fuscidentalis. Journal of insect physiology 2012, 58, 1493-501. [CrossRef]
- Rebijith, K. B.; Asokan, R.; Ranjitha, H. H.; Rajendra Prasad, B. S.; Krishna, V.; Krishna Kumar, N. K., Diet-Delivered dsRNAs for Juvenile Hormone-Binding Protein and Vacuolar ATPase-H Implied Their Potential in the Management of the Melon Aphid (Hemiptera: Aphididae). Environmental entomology 2016, 45, 268-75. [CrossRef]
- Wei, D.; Li, H. M.; Tian, C. B.; Smagghe, G.; Jia, F. X.; Jiang, H. B.; Dou, W.; Wang, J. J., Proteome analysis of male accessory gland secretions in oriental fruit flies reveals juvenile hormone-binding protein, suggesting impact on female reproduction. Scientific reports 2015, 5, 16845. [CrossRef]
- Saito, K.; Su, Z. H.; Emi, A.; Mita, K.; Takeda, M.; Fujiwara, Y., Cloning and expression analysis of takeout/JHBP family genes of silkworm, Bombyx mori. Insect molecular biology 2006, 15, 245-51. [CrossRef]
- Whitmore, E.; Gilbert, L. I., Haemolymph lipoprotein transport of juvenile hormone. Journal of insect physiology 1972, 18, 1153-67. [CrossRef]
- Kramer, K. J.; Sanburg, L. L.; Kézdy, F. J.; Law, J. H., The Juvenile Hormone Binding Protein in the Hemolymph of Manduca sexta Johannson (Lepidoptera: Sphingidae). Proceedings of the National Academy of Sciences of the United States of America 1974, 71, 493-7. [CrossRef]
- Lerro, K. A.; Prestwich, G. D., Cloning and sequencing of a cDNA for the hemolymph juvenile hormone binding protein of larval Manduca sexta. The Journal of biological chemistry 1990, 265, 19800-6. [CrossRef]
- Wojtasek, H.; Prestwich, G. D., Key disulfide bonds in an insect hormone binding protein: cDNA cloning of a juvenile hormone binding protein of Heliothis virescens and ligand binding by native and mutant forms. Biochemistry 1995, 34, 5234-41. [CrossRef]
- Rodriguez Parkitna, J. M.; Ozyhar, A.; Wiśniewski, J. R.; Kochman, M., Cloning and sequence analysis of Galleria mellonella juvenile hormone binding protein--a search for ancestors and relatives. Biological chemistry 2002, 383, 1343-55. [CrossRef]
- Vermunt, A. M.; Kamimura, M.; Hirai, M.; Kiuchi, M.; Shiotsuki, T., The juvenile hormone binding protein of silkworm haemolymph: Gene and functional analysis. Insect molecular biology 2001, 10, 147-54. [CrossRef]
- Li, W.; Cheng, T.; Hu, W.; Peng, Z.; Liu, C.; Xia, Q., Genome-wide identification and analysis of JHBP-domain family members in the silkworm Bombyx mori. Molecular genetics and genomics : MGG 2016, 291, 2159-2171. [CrossRef]
- Xia, Q.; Li, S.; Feng, Q., Advances in silkworm studies accelerated by the genome sequencing of Bombyx mori. Annu Rev Entomol 2014, 59, 513-36. [CrossRef]
- Gamo, T.; Inokuchi, T.; Laufer, H., Polypeptides of fibroin and sericin secreted from the different sections of the silk gland in Bombyx mori. Insect Biochemistry 1977, 7, 285-295. [CrossRef]
- Inoue, S.; Tanaka, K.; Arisaka, F.; Kimura, S.; Ohtomo, K.; Mizuno, S., Silk fibroin of Bombyx mori is secreted, assembling a high molecular mass elementary unit consisting of H-chain, L-chain, and P25, with a 6:6:1 molar ratio. The Journal of biological chemistry 2000, 275, 40517-28. [CrossRef]
- Akai, H.; Kiguchi, K.; Mori, K., Increased Accumulation of Silk Protein Accompanying JH-Induced Prolongation of Larval Life in Bombyx mori L.(Lepidoptera : Bombycidae). Applied Entomology & Zoology 1971, 6, 218-220. [CrossRef]
- Akai, H.; Kobayashi, M., Induction of Prolonged Larval Instar by the Juvenile Hormone in Bombyx mori L. : Lepidoptera : Bombycidae. Applied Entomology & Zoology 1971, 6, 138-139. [CrossRef]
- Kurata, S.; Koga, K.; Sakaguchi, B., Nucleolar size in parallel with ribosomal RNA synthesis at diapause termination in the eggs of Bombyx mori. Chromosoma 1978, 68, 313-7. [CrossRef]
- Kurata, K., Effect of a juvenile hormon analogue given at various ages of 5th instar larvae on RNA synthesis in the posterior silk gland of the silkworm, Bombyx mori. Journal of Insect Biotechnology & Sericology 2010, 53, 421-426.
- Kayukawa, T.; Murata, M.; Kobayashi, I.; Muramatsu, D.; Okada, C.; Uchino, K.; Sezutsu, H.; Kiuchi, M.; Tamura, T.; Hiruma, K.; Ishikawa, Y.; Shinoda, T., Hormonal regulation and developmental role of Krüppel homolog 1, a repressor of metamorphosis, in the silkworm Bombyx mori. Developmental biology 2014, 388, 48-56. [CrossRef]
- Zhao, X. M.; Liu, C.; Jiang, L. J.; Li, Q. Y.; Zhou, M. T.; Cheng, T. C.; Mita, K.; Xia, Q. Y., A juvenile hormone transcription factor Bmdimm-fibroin H chain pathway is involved in the synthesis of silk protein in silkworm, Bombyx mori. The Journal of biological chemistry 2015, 290, 972-86. [CrossRef]
- Zhang, T.; Song, W.; Li, Z.; Qian, W.; Wei, L.; Yang, Y.; Wang, W.; Zhou, X.; Meng, M.; Peng, J.; Xia, Q.; Perrimon, N.; Cheng, D., Krüppel homolog 1 represses insect ecdysone biosynthesis by directly inhibiting the transcription of steroidogenic enzymes. Proceedings of the National Academy of Sciences of the United States of America 2018, 115, 3960-3965. [CrossRef]
- Ma, L.; Xu, H.; Zhu, J.; Ma, S.; Liu, Y.; Jiang, R. J.; Xia, Q.; Li, S., Ras1(CA) overexpression in the posterior silk gland improves silk yield. Cell research 2011, 21, 934-43. [CrossRef]
- Furuta, K.; Ichikawa, A.; Murata, M.; Kuwano, E.; Shinoda, T.; Shiotsuki, T., Determination by LC-MS of juvenile hormone titers in hemolymph of the silkworm, Bombyx mori. Bioscience, biotechnology, and biochemistry 2013, 77, 988-91. [CrossRef]
- Kayukawa, T.; Nagamine, K.; Ito, Y.; Nishita, Y.; Ishikawa, Y.; Shinoda, T., Krüppel Homolog 1 Inhibits Insect Metamorphosis via Direct Transcriptional Repression of Broad-Complex, a Pupal Specifier Gene. The Journal of biological chemistry 2016, 291, 1751-1762. [CrossRef]
- Kayukawa, T.; Jouraku, A.; Ito, Y.; Shinoda, T., Molecular mechanism underlying juvenile hormone-mediated repression of precocious larval-adult metamorphosis. Proceedings of the National Academy of Sciences of the United States of America 2017, 114, 1057-1062. [CrossRef]
- Li, K.; Tian, L.; Guo, Z.; Guo, S.; Zhang, J.; Gu, S. H.; Palli, S. R.; Cao, Y.; Li, S., 20-Hydroxyecdysone (20E) Primary Response Gene E75 Isoforms Mediate Steroidogenesis Autoregulation and Regulate Developmental Timing in Bombyx. The Journal of biological chemistry 2016, 291, 18163-75. [CrossRef]
- Liu, X.; Dai, F.; Guo, E.; Li, K.; Ma, L.; Tian, L.; Cao, Y.; Zhang, G.; Palli, S. R.; Li, S., 20-Hydroxyecdysone (20E) Primary Response Gene E93 Modulates 20E Signaling to Promote Bombyx Larval-Pupal Metamorphosis. The Journal of biological chemistry 2015, 290, 27370-27383. [CrossRef]
- Li, J. Y.; Yang, H. J.; Lan, T. Y.; Wei, H.; Zhang, H. R.; Chen, M.; Fan, W.; Ma, Y. Y.; Zhong, B. X., Expression profiling and regulation of genes related to silkworm posterior silk gland development and fibroin synthesis. Journal of proteome research 2011, 10, 3551-64. [CrossRef]
- Zeng, B.; Huang, Y.; Xu, J.; Shiotsuki, T.; Bai, H.; Palli, S. R.; Huang, Y.; Tan, A., The FOXO transcription factor controls insect growth and development by regulating juvenile hormone degradation in the silkworm, Bombyx mori. The Journal of biological chemistry 2017, 292, 11659-11669. [CrossRef]
- Zhang, Z.; Liu, X.; Shiotsuki, T.; Wang, Z.; Xu, X.; Huang, Y.; Li, M.; Li, K.; Tan, A., Depletion of juvenile hormone esterase extends larval growth in Bombyx mori. Insect biochemistry and molecular biology 2017, 81, 72-79. [CrossRef]
- Kayukawa, T.; Minakuchi, C.; Namiki, T.; Togawa, T.; Yoshiyama, M.; Kamimura, M.; Mita, K.; Imanishi, S.; Kiuchi, M.; Ishikawa, Y., Transcriptional regulation of juvenile hormone-mediated induction of Krüppel homolog 1, a repressor of insect metamorphosis. Proceedings of the National Academy of Sciences of the United States of America 2012, 109, 11729-11734. [CrossRef]
- Pan, M. H.; Xiao, S. Q.; Chen, M.; Hong, X. J.; Lu, C., Establishment and characterization of two embryonic cell lines of Bombyx mori. In vitro cellular & developmental biology. Animal 2007, 43, 101-4. [CrossRef]
Figure 1.
The expression of BmJHBPd2 was different in the posterior silk gland of the different silk-producing strain Dazao and S872. A–D. Relative Bmfib-H, Bmfib-L, BmP25 and BmJHBPd2 mRNA levels in the PSG from Dazao and S872 larvae on days 3rd and 5th of the fifth instar as analyzed by quantitative RT-PCR. BmRpl3 expression is shown as a control The results are expressed as means ± SD of three independent experiments; * P < 0.05; ** P <0.01; *** P < 0.001.
Figure 1.
The expression of BmJHBPd2 was different in the posterior silk gland of the different silk-producing strain Dazao and S872. A–D. Relative Bmfib-H, Bmfib-L, BmP25 and BmJHBPd2 mRNA levels in the PSG from Dazao and S872 larvae on days 3rd and 5th of the fifth instar as analyzed by quantitative RT-PCR. BmRpl3 expression is shown as a control The results are expressed as means ± SD of three independent experiments; * P < 0.05; ** P <0.01; *** P < 0.001.
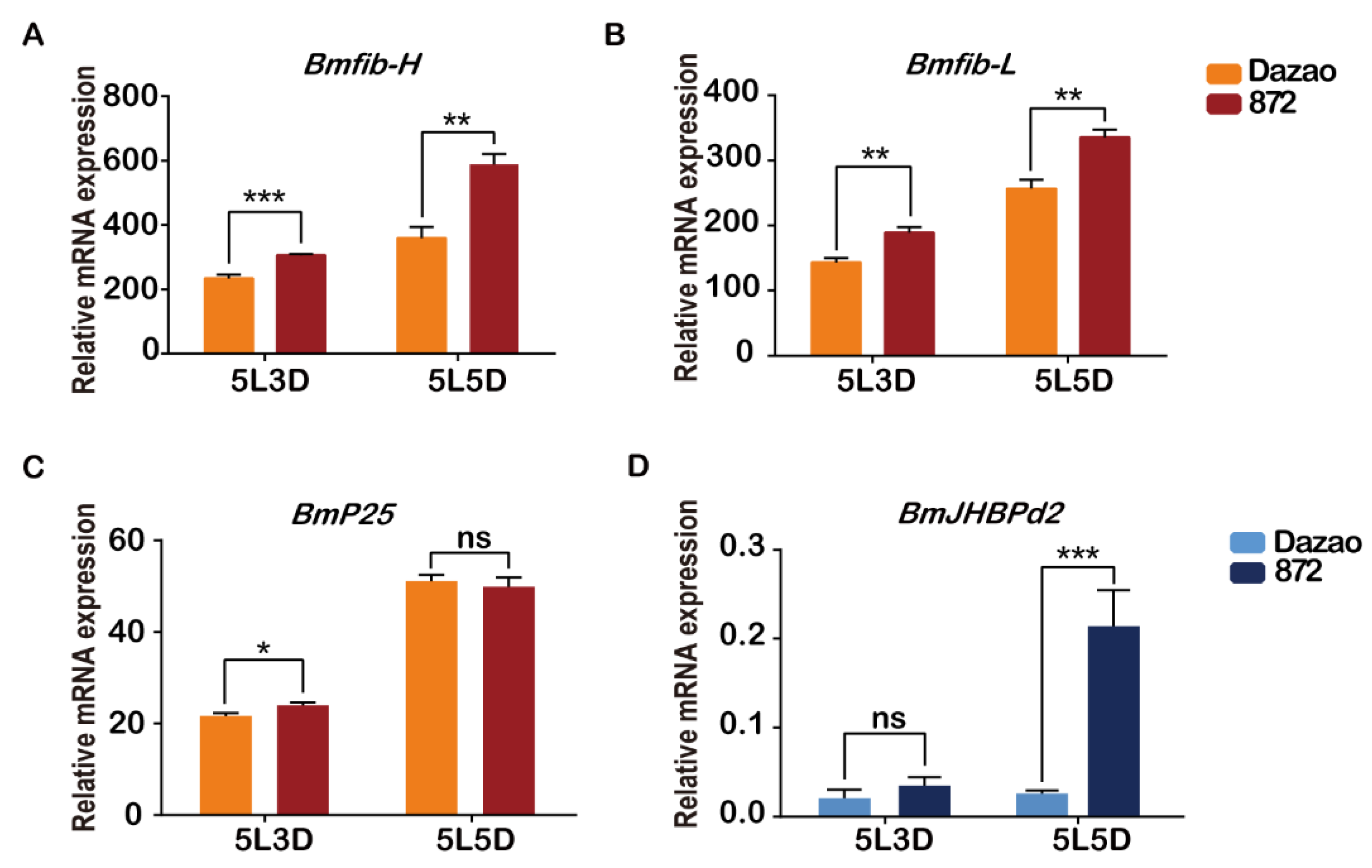
Figure 2.
Overexpressing BmJHBPd2 in BmE cells. A. Structural map of subcellular localization overexpression of BmJHBPd2 vector. B. Immunofluorescence experiment of BmJHBPd2 in BmE cells.
Figure 2.
Overexpressing BmJHBPd2 in BmE cells. A. Structural map of subcellular localization overexpression of BmJHBPd2 vector. B. Immunofluorescence experiment of BmJHBPd2 in BmE cells.
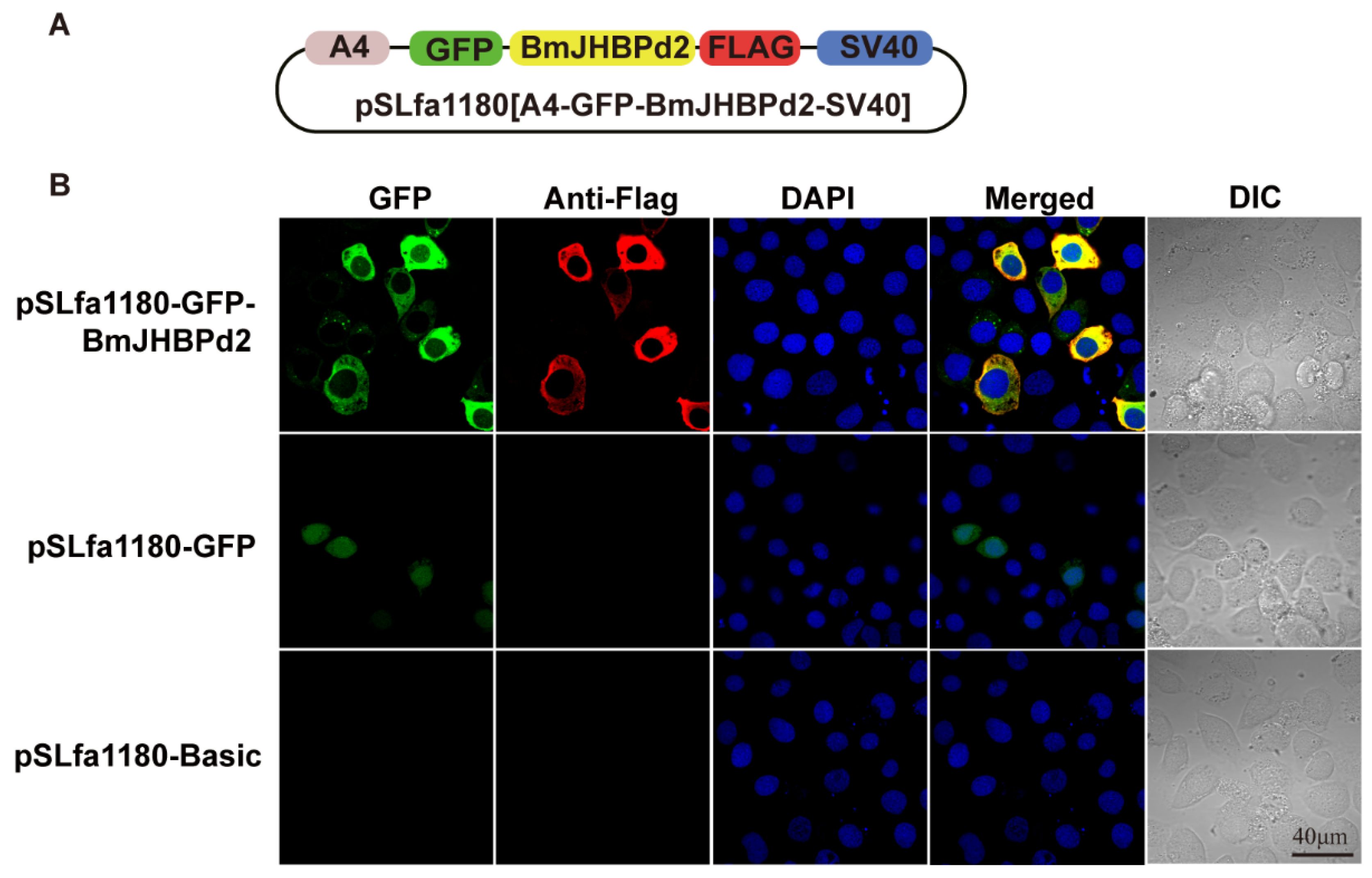
Figure 3.
BmJHBPd2 responds to JHA in BmE cells. A. Overexpression of BmJHBPd2 assayed by qRT-PCR and protein level of in BmE cells overexpressed BmJHBPd2 using the flag tag antibody, and Tubulin was shown as a control. B. Expression of BmJHBPd2, Bmfib-H, BmKr-h1 and BmMet1 after adding JHA or DMSO to normal BmE cells. C. Expression of Bmfib-H, BmKr-h1 and BmMet1 after overexpression of BmJHBPd2 assayed by qRT-PCR. D. Bmfib-H, BmKr-h1 expression level of BmE cells overexpressed BmJHBPd2 after adding JHA. BmRpl3 expression is shown as a control. Results are expressed as the means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
Figure 3.
BmJHBPd2 responds to JHA in BmE cells. A. Overexpression of BmJHBPd2 assayed by qRT-PCR and protein level of in BmE cells overexpressed BmJHBPd2 using the flag tag antibody, and Tubulin was shown as a control. B. Expression of BmJHBPd2, Bmfib-H, BmKr-h1 and BmMet1 after adding JHA or DMSO to normal BmE cells. C. Expression of Bmfib-H, BmKr-h1 and BmMet1 after overexpression of BmJHBPd2 assayed by qRT-PCR. D. Bmfib-H, BmKr-h1 expression level of BmE cells overexpressed BmJHBPd2 after adding JHA. BmRpl3 expression is shown as a control. Results are expressed as the means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
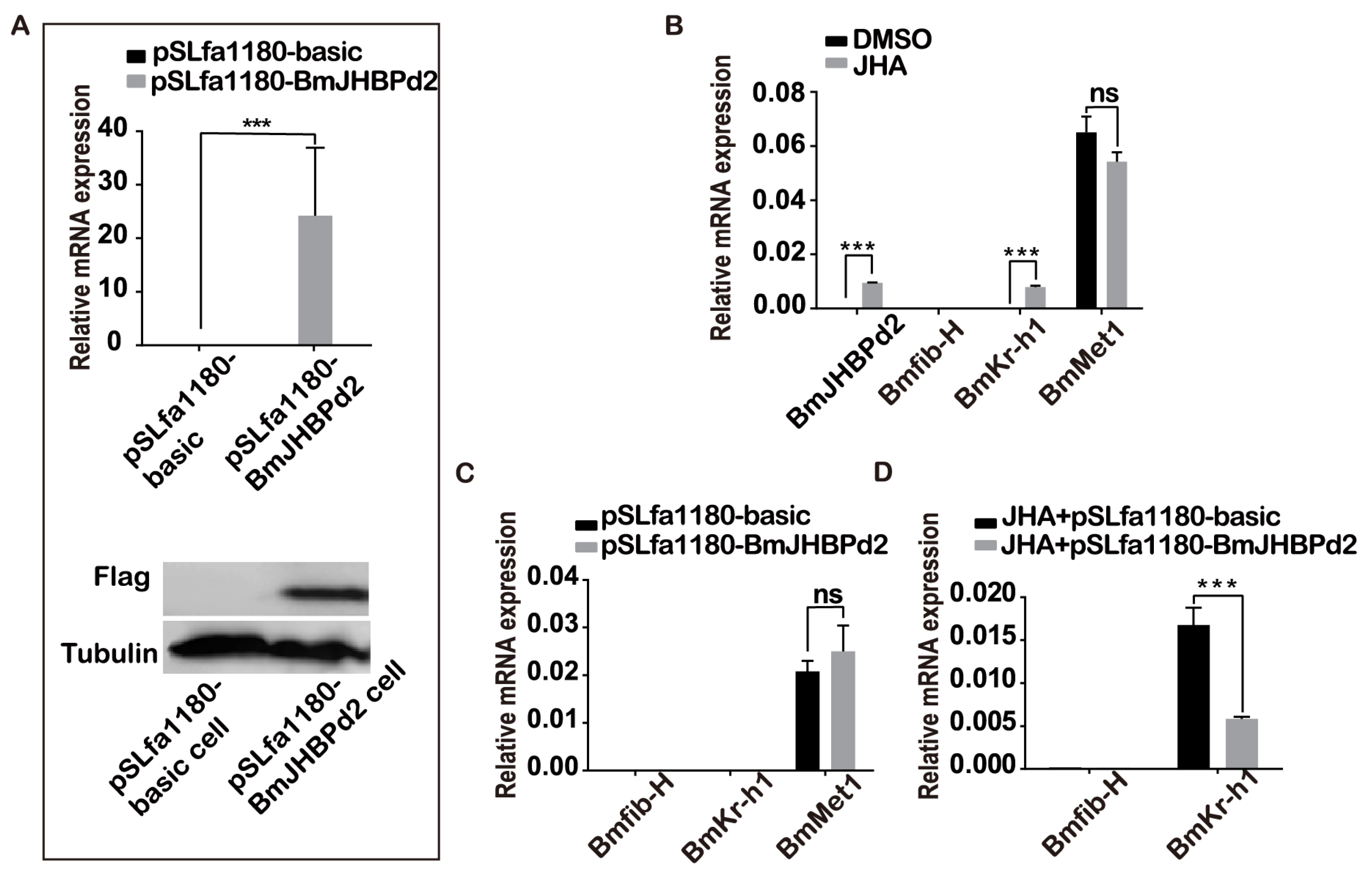
Figure 4.
Overexpression of BmJHBPd2 in the posterior silk gland. A. Schematic diagram of carrier construction. B. Screening of transgenic moths, the right shows a positive individual and the green fluorescence appears in the eyes under the fluorescence microscope. C. Overexpression of BmJHBPd2 assayed by qRT-PCR, with BmRpl3 expression being shown as a control. D. Detection of overexpression of BmJHBPd2 at the protein level using Myc tag antibody, Tubulin was shown as a control. Results are expressed as means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
Figure 4.
Overexpression of BmJHBPd2 in the posterior silk gland. A. Schematic diagram of carrier construction. B. Screening of transgenic moths, the right shows a positive individual and the green fluorescence appears in the eyes under the fluorescence microscope. C. Overexpression of BmJHBPd2 assayed by qRT-PCR, with BmRpl3 expression being shown as a control. D. Detection of overexpression of BmJHBPd2 at the protein level using Myc tag antibody, Tubulin was shown as a control. Results are expressed as means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
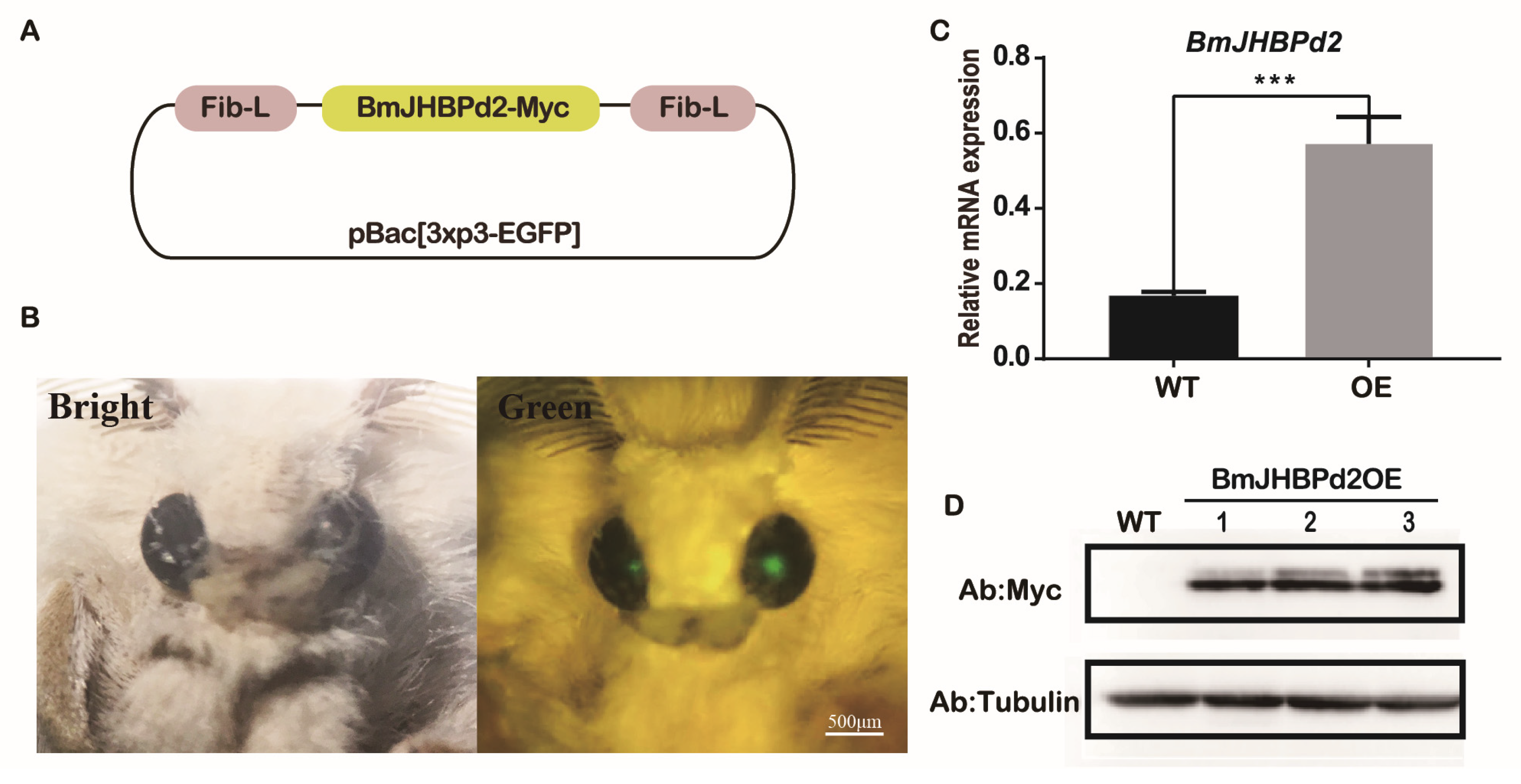
Figure 5.
Analysis of fibroin gene expression and cocoon traits. A-F. Phenotype of silk gland at the fifth instar and cocoon of BmJHBPd2OE and WT lines. G-I. Expression of silk fibroin protein gene in the wild type and transgenic lines. BmRpl3 expression was used as a control. Results are expressed as means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
Figure 5.
Analysis of fibroin gene expression and cocoon traits. A-F. Phenotype of silk gland at the fifth instar and cocoon of BmJHBPd2OE and WT lines. G-I. Expression of silk fibroin protein gene in the wild type and transgenic lines. BmRpl3 expression was used as a control. Results are expressed as means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
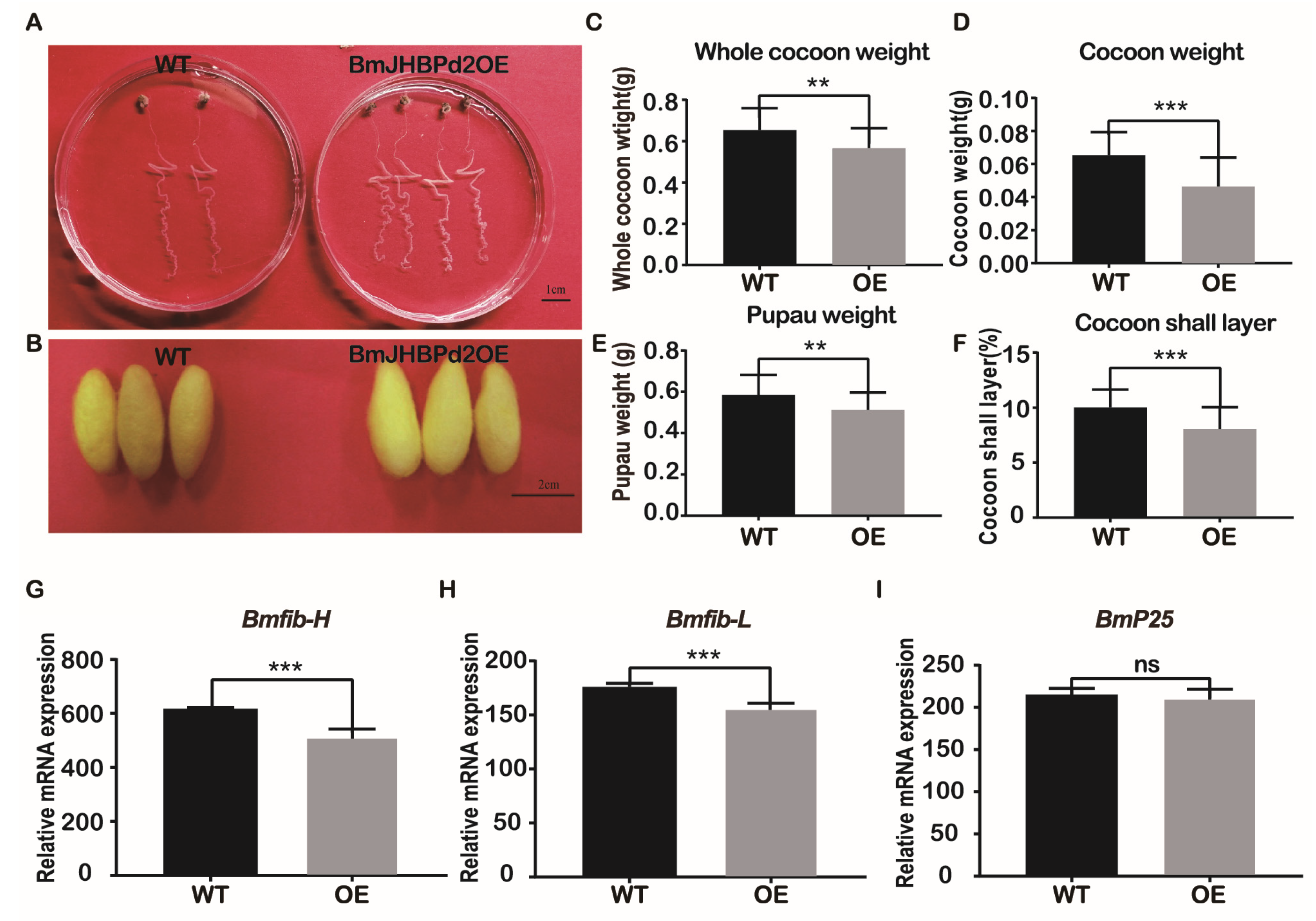
Figure 6.
Overexpression of BmJHBPd2 results in inhibition of JH signaling pathway in silk gland assayed by qRT-PCR. The following JH signaling pathway related genes were selected: Kr-h1 (A), Met1 (B), Met2 (C), SRC (D). BmRpl3 expression is shown as a control. Results are expressed as means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
Figure 6.
Overexpression of BmJHBPd2 results in inhibition of JH signaling pathway in silk gland assayed by qRT-PCR. The following JH signaling pathway related genes were selected: Kr-h1 (A), Met1 (B), Met2 (C), SRC (D). BmRpl3 expression is shown as a control. Results are expressed as means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
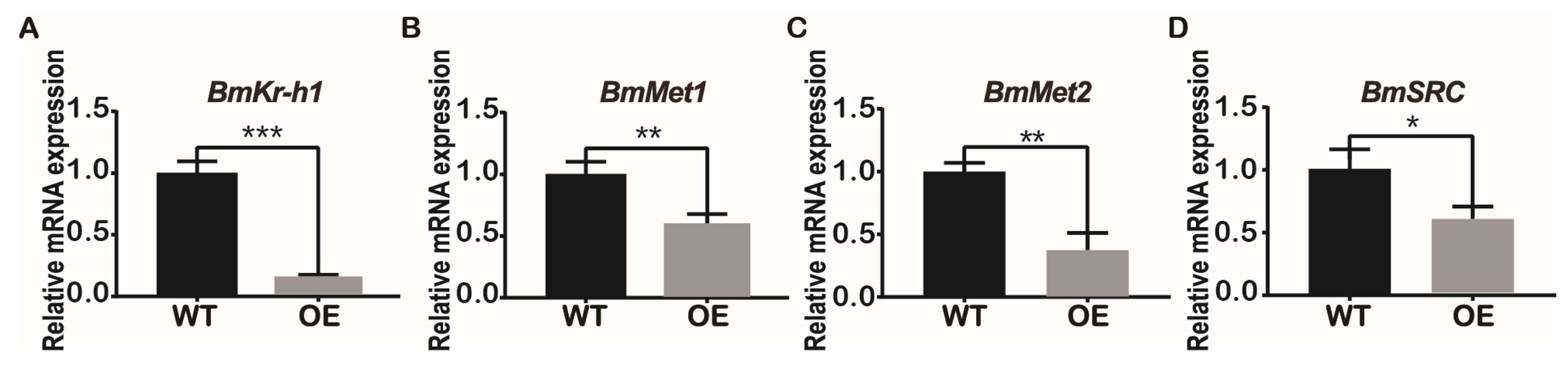
Figure 7.
Overexpression of BmJHBPd2 results in upregulation of 20E signaling pathway in silk gland assayed by qRT-PCR. The following 20E signaling pathway related genes were selected: Brc (A), E74A (B), USP (C), EcR (D), HR3 (E), and E75A (F). BmRpl3 expression is shown as a control. Results are expressed as means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
Figure 7.
Overexpression of BmJHBPd2 results in upregulation of 20E signaling pathway in silk gland assayed by qRT-PCR. The following 20E signaling pathway related genes were selected: Brc (A), E74A (B), USP (C), EcR (D), HR3 (E), and E75A (F). BmRpl3 expression is shown as a control. Results are expressed as means ± S.D. of three independent experiments. * P < 0.05; ** P <0.01; *** P < 0.001.
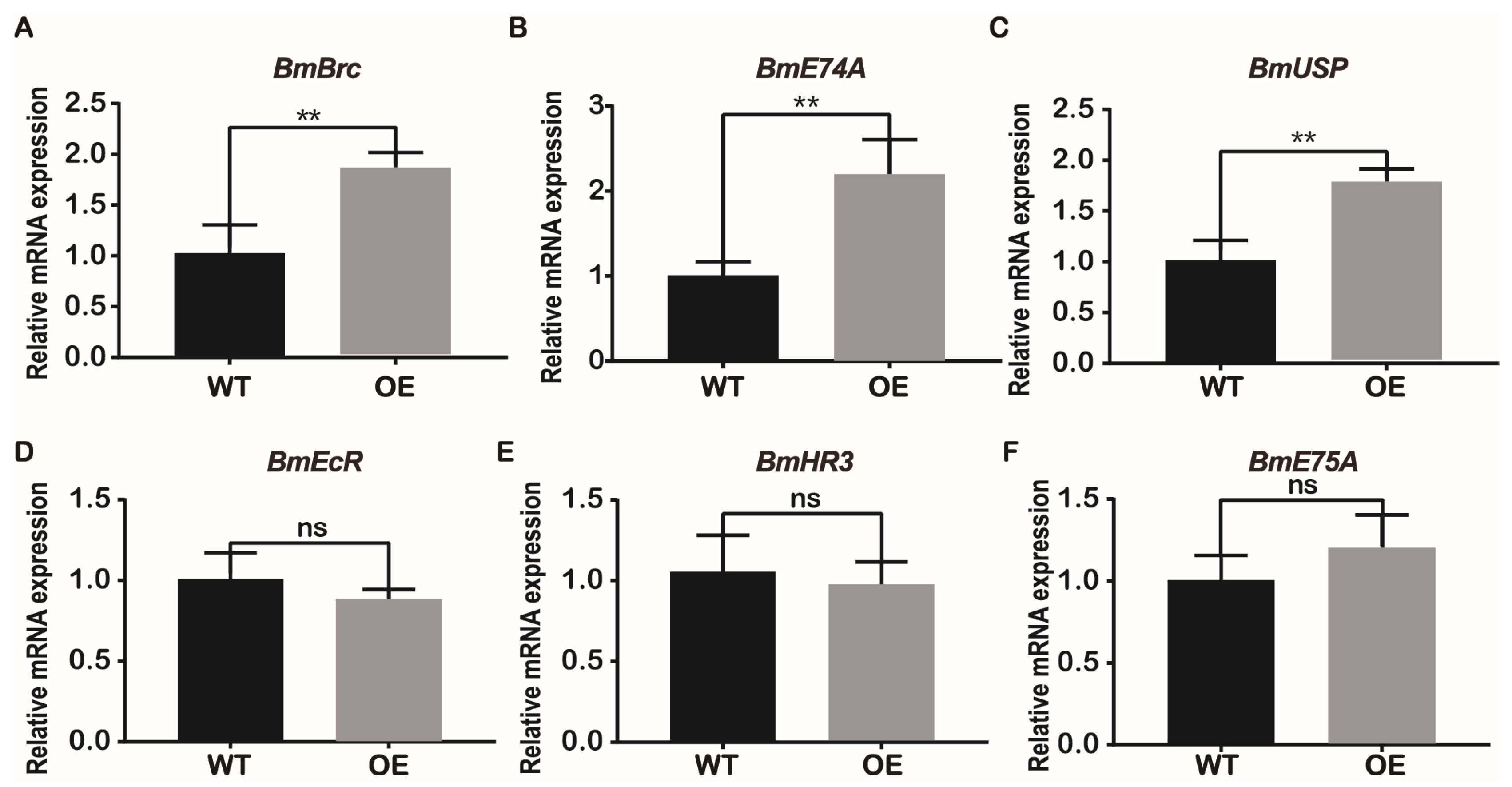
Figure 8.
Expression pattern of BmJHBPd2 in the PSG.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



