
Submitted:
25 May 2023
Posted:
29 May 2023
You are already at the latest version
Abstract
The widely conserved RACK1 protein is a WD-40 type scaffold protein that regulates diverse environmental stress signal transduction pathways. Arabidopsis RACK1A has been reported to interact with various proteins in salt stress and Light-Harvesting Complex (LHC) pathways. However, the mechanism of how RACK1 contributes to the photosystem and chlorophyll metabolism in stress conditions remains elusive. Using T-DNA-mediated activation tagging transgenic rice (Oryza sativa L.) lines, we show that leaves from rice RACK1B gene (OsRACK1B) gain-of-function (RACK1B-OX) plants exhibit the stay-green phenotype under salinity stress. In contrast, leaves from down-regulated OsRACK1B (RACK1B-UX) plants display an accelerated yellowing. qRT-PCR analysis revealed that several genes encoding chlorophyll catabolic enzymes (CCEs) are differentially expressed in both RACK1B-OX and RACK1B-UX rice plants. In addition to CCEs, STAY-GREEN (SGR) is a key component that forms the SGR-CCE complex in senescing chloroplasts, which causes LHCII complex instability. Transcript and protein profiling revealed a significant upregulation of OsSGR in RACK1B-UX plants than that in RACK1B-OX rice plants during salt treatment. The results imply that senescence-associated transcription factors (TFs) are altered in accordance with altered OsRACK1B expression, indicating a transcriptional reprogramming by OsRACK1B and a novel regulatory mechanism involving the OsRACK1B-OsSGR-TFs complex. Our findings suggest that ectopic expression of OsRACK1B negatively regulates chlorophyll degradation, leads to the steady level of LHC-II isoform Lhcb1, an essential prerequisite for the state transition of photosynthesis for adaptation, and delays salinity-induced senescence. Taken together, these results provide important insights into the molecular mechanisms of salinity-induced senescence, which can be useful in circumventing the effect of salt on photosynthesis and in reducing the yield penalty of important cereal crops, like rice, in global climate change conditions.
Keywords:
RACK1B
; Rice
; Salt stress
; Stay-green
; Light Harvesting Complex
; Chlorophyll degradation
; Senescence
1. Introduction
The Receptor for Activated C Kinase1 (RACK1) is a conserved multifunctional WD-40 type scaffold protein that functions as an assembly platform for diverse signaling pathways involved in plant development, stress responses, and immunity[1]. While mammalian RACK1 is well known for its role as a hub for cellular signaling cascades ranging from transcriptional regulation, protein translation, ribosome machinery biogenesis to cell proliferation, tumorigenesis, immune response, apoptosis, and cell senescence etc. [2,3,4,5,6], the function of plant RACK1 is still emerging. Despite a fundamental difference in phosphorylation sites, plant RACK1s appear to be functionally similar to mammalian RACK1 [7,8]. In plants, RACK1 has been implicated in seed germination, root development, flowering, pollen development and fruit ripening [9,10,11]. Mutant analysis in Arabidopsis (Arabidopsis thaliana) and rice (Oryza sativa L.) plants revealed that RACK1 plays a crucial role in response to environmental stress conditions such as drought and salinity [12]. Intriguingly, regulation of many of these pathways appears to be accomplished through the reported interaction of RACK1 with over 100 different proteins [1,13,14]. However, the physiological role of these interactions remains poorly understood.
The rice genome contains two homologs of RACK1 gene- OsRACK1A and OsRACK1B with 82% amino acid similarity. Nakashima et al., (2008) first reported that OsRACK1A serves as an adapter protein for a complex involving Rac1, RAR1, SGT1, and RbohB at the plasma membrane to provide resistance against blast pathogen [15]. Zhang and colleagues found that OsRACK1A is regulated by the circadian rhythm and interacts with many salt-responsive proteins [12]. Most recently, we have reported that overexpression of OsRACK1B induces ROS burst as a form of H2O2 through direct interaction with the N-terminal region of RbohD and affects the timing of anther dehiscence, pollen viability, and pollen cell wall integrity [11].
Rice feeds nearly half of the world’s population and is considered to be the single most important source of calories. However, adverse climate change coupled with environmental stressors such as drought, salinity, high and low temperatures, and pathogen attacks significantly constrain rice production. Compared to other cereal crops such as wheat and maize, rice is the most salinity-sensitive crop, and salinity of the soil causes an annual 30-40% yield loss in rice-growing countries [16]. Salinity affects plants in many ways, including disrupting cellular ionic homeostasis, dismantling photosynthetic apparatus, breaking down chlorophyll (Chl) pigments, and decreasing the efficiency of photosystem II (PS-II) and PS-I, leading to premature senescence [17,18,19].
Although higher plants undergo senescence processes during the reproductive stage as part of nutrient remobilization for grain and fruit ripening, the process can occur in vegetative stages in harsh environmental conditions. Abiotic and biotic stressors such as salinity, heat, drought, UV radiation, and pathogen attack can induce premature senescence [20,21]. Upon induction by stressors, the highly coordinated process of senescence is set through the activation of transcription factors (TFs) that modulate the expression of senescence-associated genes (SAGs), breakdown of chloroplast, and destabilization of the light harvesting complex which ultimately leads to cell death [17,22,23,24].
Abiotic stressors such as high salinity promote premature senescence that accompany yellowing of leaves due to the degradation of chlorophyll pigments and unmasking of preexisting carotenoids [25,26,27,28]. Chlorophyll retention and delayed senescence, however, lead to the stay-green phenotype, in which leaves remain green in color [29]. Recent research in crop and model plants has demonstrated that a regulatory protein, named Stay-Green (SGR) or NYE1 (also known as the protein for Mendel’s cotyledon trait), is involved in thylakoid photosystem disassembly and chlorophyll degradation during senescence. Mutant sgr plants exhibited delayed loss of chlorophyll during natural and dark-induced senescence in higher plants [30,31,32,33]. In contrast, SGR- overexpressing plants exhibited reductions in chlorophyll content and accelerated cell death in both rice and Arabidopsis seedlings. In Arabidopsis, SGR forms a complex with Chlorophyll catabolic enzymes (CCEs) for chlorophyll degradation and associates with chlorophyll binding proteins to aid chloroplast proteases to remove the damaged molecules [28,31,32,34,35]. Agronomically, the stay-green trait and delayed leaf senescence is of great interest for research due to its link with high yield [36]. In conjunction, several recent findings suggest the potential of delayed leaf senescence in grain filling and abiotic stress resilience in cereal crops like rice and wheat [33,37].
Though RACK1 has been reported to interact with several photosystem complex proteins, to date, there is no report on the precise role RACK1 plays within the photosystem complex under diverse environmental stimuli. As both RACK1 and photosystem complex plays central role in plant growth and development, understanding their crosstalk can provide insights into optimizing plant growth and yield potential. With this aim in mind, we sought to investigate RACK1’s role in photosystem complex under abiotic stress condition. We used a T-DNA insertional approach, an effective genetic tool that has the potential for activation (activation-tagging) or suppression of a native transcript depending on the exact position and direction of the insertion site within the genomic sequence [38]. We found that leaf discs from OsRACK1B overexpressed transgenic rice plants exhibit strong stay-green phenotypes by retaining more chlorophylls than that of the wild-type (WT) plants during salinity-induced senescence. By contrast, leaf discs from down-regulated OsRACK1B plants display chlorotic phenotypes and premature senescence. Gene expression analysis revealed that several genes involved in senescence and chlorophyll catabolism are differentially expressed in the RACK1B transgenic rice plants. Our study also indicates that OsRACK1B plays a key role in protecting the light harvest complex during stress-induced senescence.
2. Results
2.1. Identification of T-DNA insertion Activation Tagged rice plants overexpressing and down-regulating OsRACK1B
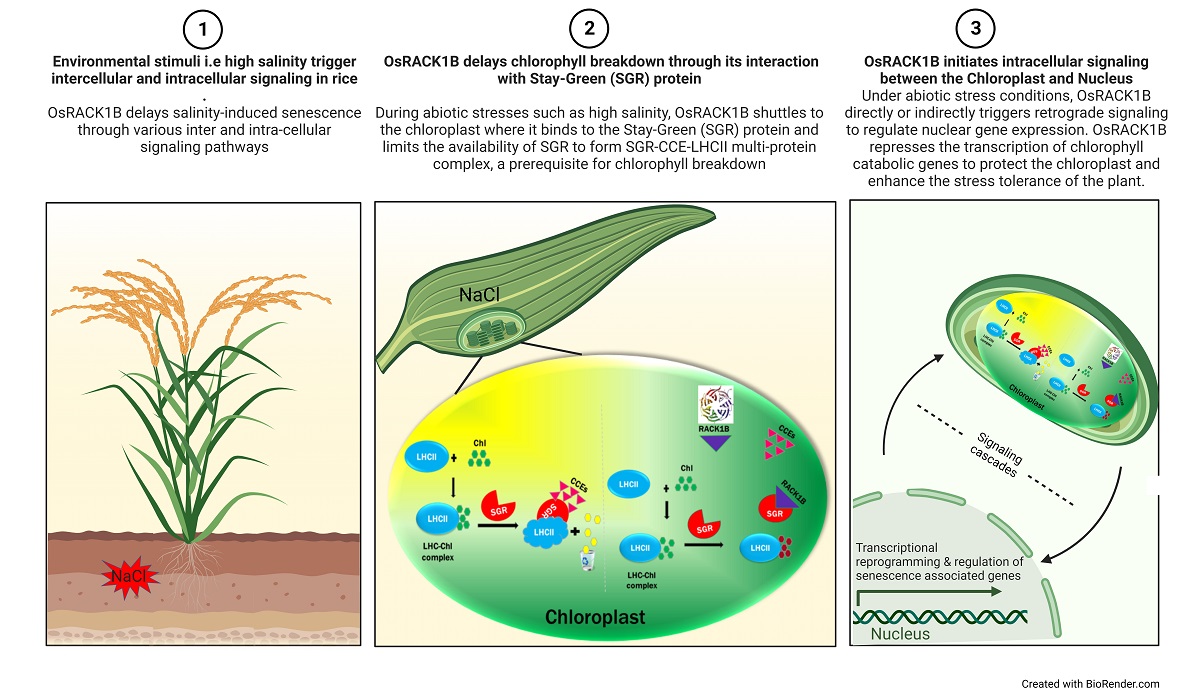
To elucidate the physiological function of OsRACK1B (Loc_Os05g47890) in salt stress, we screened both overexpression and loss of function lines from RICEGE database (http://signal.salk.edu/cgi-bin/RiceGE). We identified two putative OsRACK1B gain of function lines as described previously in Rahman et al., 2022 [11]. For this study, we identified PFG_3D-02734.L line as the putative loss-of-function line where T-DNA is located in the intronic region, between the two exons of the same locus. Expression analysis by qRT-PCR and western blot revealed that two plants from putative gain-of-function lines accumulated 4-7 folds more OsRACK1B transcripts and higher protein levels than that in Dongjin (hereafter termed WT) plants (Figure 1B, D). We refer to these two lines as the OsRACK1B overexpressed lines and designated them OX-1 (PFG_3A-07870.R) and OX-2 (PFG_3A-60871.L) (Rahman et. al., 2022). Similarly, OsRACK1B transcript and protein levels from two plants of the PFG_3D-02734.L line showed a substantial decrease (~50%) compared to the Hwayoung (WT) plant Figure 1C, E), hereafter referred to as the OsRACK1B knock-down line and designated as UX-1 and UX-2. TAIL-PCR and Sequence analysis revealed that the T-DNA insertion site of UX-1 and UX-2 is located in the intronic region between the two exons of OsRACK1B, 1018 bp downstream of the start codon (ATG) at Loc_Os05g47890. (Figure 1 A). Within the nearly 10 kb insertion site, except for several transposons, no other gene coding sequences could be seen, implying a low possibility of interference by ectopic activation of other nearby genes.
2.2. Transgenic rice plants overexpressing OsRACK1B exhibit stay-green phenotype under salinity stress
Despite having molecular evidence of RACK1 as an important regulator in response to salt stress, there has been a significant gap in understanding of the physiological role of RACK1 in stress-induced metabolic pathways, such as premature senescence. It was of interest to test whether RACK1 also contributes to this process concomitant with salt- stress signaling pathways. To this end, we investigated the physiological function of RACK1B in response to high salinity-induced senescence. The leaf discs from eight-week-old OX-1, OX-2 and WT plants were subjected to salt stress for 3 days under continuous light conditions. After 3 days of salt treatment (DST), phenotyping analysis revealed that leaf discs from OX-1 and OX-2 plants exhibited staygreen phenotypes while those of the WT turned yellow suggesting a delayed chlorophyll degradation in OX plants (Figure 2A, C ). ImageJ analysis of from similar area size revealed that chlorophyll pigmentation was higher in OX leaves than WT leaf discs. Consistent with the visible phenotype, OX leaf discs retained more total chlorophyll than WT leaf discs after 3 days of salt treatment (Supplimental Fig. S-3 A).
2.3. OsRACK1B down-regulated plant leaves display premature senescence in salinity stress
To further confirm the effect of RACK1 in chlorophyll catabolism under abiotic stress conditions, we subjected leaf discs from down-regulated RACK1B plants (UX-1 and UX-2) to salinity stress for 3 days under continuous light conditions. After 3 days, UX leaf discs exhibited severe yellowing compared to the WT leaf discs, which remained partially green (Figure 2B). In accordance with visible phenotypes, ImageJ analysis of the simialr area revealed that chlorophyll pigmentation was also lower than WT leaf discs (Figure 2D). The yellow phenotype in UX rice leaf discs indicated expedited chlorophyll degradation and early senescence compared to that in the WT leaves during salt stress. This phenotype contrasts with that of the OsRACK1B-OX leaves treated with the same concentration of salt for 3 days. Please note that the WT controls are different for the OX and UX lines. The total chlorphyll quantification also reflected the visible phenotype as UX rice leaves contained significantly low amount of chlorophyll compared the WT after 3 days of salt treatment (supplimental fig. S-3 B) To exclude the possibility of external factors influencing this phenotype, plants were grown and leaf discs were treated under the same conditions. The phenotypic differences in OsRACK1B-OX and UX leaf discs demonstrated that RACK1 is closely associated with salinity-induced senescence. To this end, we propose that RACK1B may act as a negative regulator of stress-induced chlorophyll degradation in rice plants. To rule out other possibilities influencing the phenotype, we screened the selected OsRACK1B transgenic lines for additional insertions in different chromosomal locations using Thermal Asymmetric Interlaced PCR (TAIL-PCR) as described in Rahman et al., 2022 [11]. While OX-1, OX-2, and UX-1 plants were confirmed to be a single copy of T-DNA insertion, a second insertion was found (Supp. Fig. S-1 A & B) nearby an EF-hand domain-containing protein (Loc_Os08g0558100) at Chr 8 in UX-2 plant (Supp. Fig. S-2 A & B)) and consequently was discarded from further investigation.
2.4. OsRACK1B negatively regulates the expression of chlorophyll degradation and senescence-associated genes
Leaf yellowing is the primary symptom of senescence and can be initiated by aging and environmental elicitors such as high salinity. In higher plants, senescence induction is tightly associated with chlorophyll metabolism [26,47]. Several genes involved in the chlorophyll synthesis or degradation are differentially expressed during senescence [47,48]. To elucidate the molecular mechanism of how overexpression or down-regulation of RACK1 perturbs the chlorophyll metabolism, we used quantitative PCR (qRT-PCR) to measure the transcript levels of some key genes involved in chlorophyll metabolism at baseline (0 hr) and after 24 hours of salt treatment. First, we measured the expression of a chlorophyll biosynthesis gene: chlorophyllide a oxygenase (CAO), which encodes Chl a oxygenase that catalyzes the transformation of Chl a into Chl b [49]. Gene expression analysis revealed that OsCAO levels are significantly up-regulated in OX plants and down-regulated in UX plants in comparison to WT levels following salt treatment (Figure 3A & B). Previously, it was found that plants overexpressing CAO exhibit the stay-green phenotype and a number of transcription factors were differentially expressed when artificial senescence was induced [50]. Therefore, we hypothesized that OsCAO upregulation in OX plants after salt treatment is one of the reasons for its stay-green phenotype in leaves upon salt exposure. Likewise, consistent with chlorotic phenotypes, low CAO expression in the UX plant suggests rapid chlorophyll breakdown in UX plants under high salinity. Next, we measured the mRNA expression of key genes encoding chlorophyll catabolic enzymes (CCE). Chlorophyll degradation is a multi-step process. The cascade starts with the conversion of Chl b to Chl a, catalyzed by Chl b reductase (NYC1). In the final step, the ring structure of the intermediate breakdown products is oxygenolytically opened by pheophorbide a oxygenase (PAO) to generate red chl-catabolite (RCC), which is degraded further by RCC reductase (RCCR). Several studies in plants found that mutations of NYC1 and PAO lead to chlorophyll retention, and their transcript levels increase during natural or salinity-induced senescence [26,29,51,52]. We found that transcript levels of OsNYC and OsRCCR were up-regulated in UX plants after salt exposure in comparison to that in the WT plants (Figure 3D and H). By contrast, their expression pattern in OX plants remains the same or even lower after 24 hours of salt treatment (Figure 3C and G ). Therefore, we speculated that the constitutive expression of OsRACK1B negatively influences the expression of CCEs, in consistance with their stay-green phenotype in OX plants. Likewise, accelerated leaf yellowing in UX plants indicate that functional RACK1 plays an important role in limiting the expression of CCE genes. Senescence is a complex process tightly controlled by a number of senescence-related transcriptional factors (TFs). We questioned whether RACK1B overexpression and down-regulation have any effect on transcriptional reprogramming during salt stress-induced senescence. In plants, NAC (NAM, ATAF1, -2, and CUC2) domain transcription factors represent the largest family members in transcriptional regulation and are well-known for their critical roles in stress-induced senescence [47]. One of the TFs -OsNAC092 (also called ORE1) has recently been identified as a key transcriptional activator during abiotic stress-dependent senescence. OsNAC092 gene expression is triggered by salt stress and activates a large number of senescence-associated genes [23,53]. As anticipated, we found a significant (up to 7 fold) up-regulation of OsNAC092 in UX plants compared to that in WT plants, indicating a possible activation of a large number of SAGs (Figure 3 J). By contrast, its expression was significantly lower in the OX plants (Figure 3 I) after salt treatment and this level was in good accordance with their stay-green phenotype.
2.5. OsRACK1B regulates the expression of OsSGR
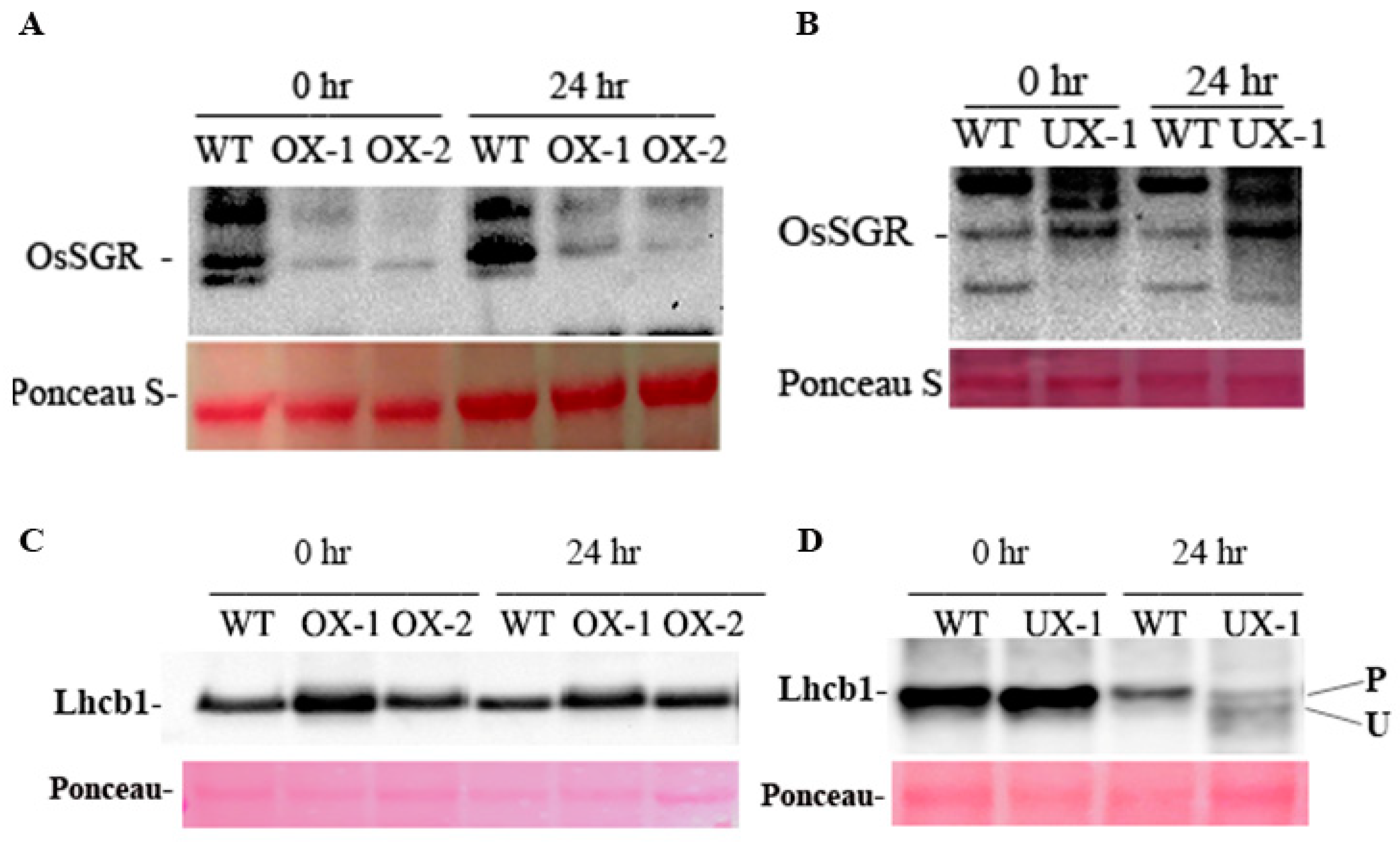
SGR has recently been recognized as a central regulator of natural and stress-induced chlorophyll breakdown. [30,33,54]. SGR interacts with all known CCEs, forms a multiprotein complex with CCEs and LHCII proteins for chlorophyll degradation, proteolytic cleavage of chloroplast, and channeling of phototoxic intermediate materials into vacuoles [31,32,55,56]. We speculated that RACK1B may regulate SGR, and this regulation might be associated with the chlorophyll catabolism in transgenic rice plants. In agreement with the phenotype observed, transcript and protein profiling revealed that SGR level is significantly altered in transgenic plants, especially after salt stress. qRT-PCR analysis revealed that OsSGR mRNA was much more abundant in UX plants before and after salt treatment than that in the WT plants; conversely, expression remains lower in OX plants (Figure 3E and F). In order to confirm that OsSGR is indeed a target of translational regulation, we analyzed OsSGR protein expression with western blot analysis. When protein extracts before and after salt treatment from leaves of OX, UX and respective WT plants were gel electrophoresed and stained with an anti-SGR antibody, we found differences in SGR protein levels between the OX and the UX plants compared with that in the WT. Consistent with their mRNA level, OX/SGR protein level was lower compared to that in the WT/SGR, indicating a down-regulation of SGR protein (Figure 4A) compared to their respective untreated WT cells, possibly due to constitutive expression of RACK1B. Even after 24 hours of salt treatment, SGR protein level was significantly high in the WT cells but the level remained steady in the OX cells. Similarly, down-regulation of RACK1B increased the SGR level as the UX/SGR band showed high intensity compared to the WT/SGR band (Figure 4 B). High salinity further induced the SGR expression in UX plants compared with that in the WT plants (Figure 4B). Collectively, these findings provided compelling evidence that SGR is regulated at both the transcriptional and translational stage by RACK1B during natural and high salinity conditions.
To test whether OsSGR is regulated by OsRACK1B through direct interaction in planta, we performed bimolecular fluorescence complementation (BiFC) assays (Supplimental Fig. S-4 A). Microscopic examination revealed strong reconstituted YFP fluorescence when nYFP-RACK1B is co-expressed with OsSGR-cYFP (Supp. Fig S-4 B). Yellow fluorescent signal co-localized with the blue fluorescence of DAPI, thus confirming an interaction between OsRACK1B and OsSGR in the nucleus (Supp Fig. S-4 B, top panel). Similarly, the fluorescence could be seen in both nucleus and cytoplasm when onion peels were treated with 200 mM NaCl for 30 min. (Supp Fig. S-4 B; second panel). This result implies that OsRACK1B negatively regulates chlorophyll degradation during salt-induced senescence at least in part by directly interacting with OsSGR.
2.6. A functional OsRACK1 is required for LHCII state transition
Previously, it was shown that RACK1 interacts with chlorophyll a/b binding proteins in the light-harvesting complex II (LHCII) [13] . As SGR interacts with all known CCEs [54,57,58] in the LHCII during chlorophyll breakdown, we investigated whether the physical interaction between RACK1B and SGR would have an effect on the PSII-LHCII super-complex and on PSII stability, especially under stress conditions. To gain further insight into the effect on PSII-LHCII, western blot analysis of light-harvesting chlorophyll a/b binding protein 1 (Lhcb1), one of the two major LHCII proteins, was performed. Lhcb1 is the most abundant in the LHCII super-complex among the trimers Lhcb1, Lhcb2, and Lhcb3 [59] . It undergoes rapid phosphorylation catalyzed by a protein kinase-STN7 and dephosphorylation during the state transition from PSII to PSI [59,60,61]. The process of state transition is one of the mechanisms adopted by plants to tune the performances of PSI and PSII, especially during stress adaptation [62,63]. To investigate whether Lhcb1 level is perturbed in RACK1B transgenic lines, we performaed a westenblot ananlysis. We found that the expression of Lhcb1 in OX and UX plants remained similar as compared to their respective WT plants in normal condition. However, after 24 hours of salt treatment, the level remained stable in OX plants but slightly decreased in WT plant (Figure 4C). By contrast, the Lhcb1 protein level in UX plants had been reduced significantly than that in the WT plants after 24 hours of salt treatment (Figure 4D). Furthermore, we observed an accumulation of an apparent non-phosphorylated form of Lhcb1 in UX plants after salt treatment whereas it was completely absent in the OX plants and their respective WT plants. (Figure 4C-D). These observations suggest that loss of fucntion of RACK1B led to loss of LHC integrity, which is an essential prerequisite for the state I-to-state II transition and triggers the dissociation of LHCII from PSII to PSI. It also suggests the possibility of an activated STN7 in OX plants duirng salt treatment. Indeed, the qPCR analysis revealed an increased mRNA level of STN7 in OX plants after salt treatment (Figure 4K). Similarly, a reduction of phosphorylated Lhcb1 in UX plant indicates that the LHC failed to maintain a smooth transition between PSI and PSII in the absence of a functional RACK1. This is also evident by the appearance of an aparent non-phosphorylated Lhcb1 in UX plants. It is therefore likely that the degradation of LHC is due to increased activity of SGR and/or low activity of STN7, particularly under salt stress conditions. Notably, SGR was found to be coimmunoprecipitated with Lhcb1 [28,64]. Interestingly, the low mRNA expression level of STN7 (Figure 4L) also agrees with the findings of the increased non-phosphorylated band in UX plants.
3. Discussion
In plants, the regulatory network of RACK1 has been well studied in the model plant Arabidopsis but little is known for monocot species, such as rice, as there have been only a few reports available to date. Kundu et al., (2013) [13] used AtRACK1A as bait to screen a split-ubiquitin based inflorescence cDNA library and demonstrated that AtRACK1A interacts with a significant number of proteins found in photosynthesis and light-regulated physiological processes in normal and stress conditions. OsRACK1 was reported to be regulated by circadian rhythms and to be involved in the regulation of salt stress responses and was found to be colocalized with PsbP, an extrinsic subunit of photosystem II (PSII) [12]. However, the regulatory network of RACK1’s role in the photosynthesis-related processes remain unclear. To further dissect the role of RACK1 in photosystem complex and salt response, we investigated the physiological and molecular response of a monocot model and economically important crop, the rice plant, to salinity stress. Here, we have provided several lines of evidence showing a novel function of OsRACK1B in regulating chlorophyll metabolism and salinity-induced senescence. We also show that RACK1B retains chlorophyll by inhibiting the functional expression of key chlorophyll catabolic genes (CCGs), namely SGR, NYC1, RCCR, and PAO under senescence-promoting high salinity conditions.
Our study revealed that leaves from OsRACK1B-OX plants exhibit persistent greenness or the “stay-green” phenotype much longer than WT leaves during high salt stress conditions (200 mM NaCl). By contrast, under the same conditions, OsRACK1B-UX leaves display leaf yellowing, an early senescence phenotype, then the WT leaves. Therefore, we speculated that the stay-green phenotype in OsRACK1B-OX plants or premature senescence in OsRACK1B-UX plants under stress conditions is due to the initiation of the cellular programs for chlorophyll catabolism. Indeed, this assumption is supported by the findings that in the OsRACK1B-OX plants, chlorophyll catabolic genes were down-regulated. Conversely, enhanced levels of chlorophyll catabolic gene expression was observed in the RACK1B-UX plant than in the WT plant. These results indicate that OsRACK1B plays a critical role in stress-induced leaf senescence. Notably, using RNA-interfered transgenic rice plants, Zhang et al., (2018) found that suppression of OsRACK1A increased chlorophyll content under 150 mM NaCl stress for up to 72 hours [12]. This disagreement may be attributed to the differences in amino acid sequence and expression pattern depending on the tissue-specific location of the two homologs. Though OsRACK1A and OsRACK1B share 82% amino acid identity, a distinct function for each of the genes is evident from discernable phenotypes from single gene manipulations. In fact, using double and triple mutants of RACK1 genes in Arabidopsis, it was shown that the genes act with unequal redundancy [10]. Previously, our lab reported that in addition to key residue phosphorylation-based interactions with more than 100 different proteins; the RACK1 proteins homo- and hetero-dimerize [44]. In this context, an epistatic relationship between these two RACK1 proteins in the chlorophyll catabolism pathway is quite possible.
The Chlorophyll catabolic pathway in plants has been intensively studied and most, if not all the genes involved are isolated and known as CCEs. In rice, some key genes such as OsSGR, OsNYC1, and OsRCCR have been recognized as hallmark genes for leaf sense cence [29,31,65,66,67]. Our results show that under strong abiotic stress conditions, RACK1B-OX leaves maintain more chlorophylls and RACK1B-UX leaves turn yellow much faster than WT leaves. Thus, regulation of the expression of CCE genes in these transgenic plants was not surprising. Indeed, when eight-week-old plants are treated with 200mM NaCl for 24 hours, we found low expression of OsSGR, OsNYC1, and OsRCCR genes in RACK1B-OX plants. In contrast, expression of these genes are significantly induced in RACK1B-UX plant. This implies a substrate delimited catabolic feedback pathway where limited availability of substrate molecules would impede the downstream catabolic activities - hence a down-regulation of the catabolic enzymes in a sequential pathway.
Under abiotic stress conditions, chloroplast homeostasis is maintained through fine coordination with nuclear gene expression for coping with stress. Intriguingly, although all CCEs are located in the chloroplast, a majority of the chloroplast proteome is nucleus-encoded and must be translocated to plastids after their synthesis in the cytosol [28,29,68,69,70]. Therefore, regulation of nuclear gene expression in response to the functional or metabolic state of the plastids, driven by retrograde signals from the plastids, is essential[71]. In rice, RACK1 was found in the nucleus, cytosol, and in microsomal fractions [15,72]. RACK1 was also found to shuttle proteins around the cell [11,73,74]. Evidence suggests that the ability of RACK1’s temporal and spatial regulation of diverse signal transduction can be attributed to its translocation among various cellular compartments, which is influenced by the particular cohort of proteins interacting with RACK1 at any given time[2,9]. Moreover, the lack of specific localization motifs in RACK1’s sequence also contributes to this ability. Thus, it is assumed that RACK1B itself regulates transcription factors in the nucleus or facilitates translocation of components for a transcriptional re-programming as an adaptor protein. For instance, RACK1A interacts with the Osrap2.6 transcription factor in the cytoplasm and nucleus and contributes to rice innate immunity [72]. Conversely, RACK1A also interacts with Rac1 at the periphery of the plasma membrane to exert the same function[15]. Whether RACK1 has any role in the chloroplast ribosome biogenesis and function is still unknown. Remarkably, chloroplast ribosome-associated proteins are found to support translation during stress conditions[75].
Over the last decade, several functional studies in Arabidopsis and rice have implicated SGR as one of the master regulators of Chl degradation, and sgr mutant plants displayed delayed Chl-degradation phenotype during natural and stress-induced senescence[65,76,77,78]. Rice has two SGR homologs, SGR and SGR-like (SGRL), with overlapping biochemical functions [56]. The Arabidopsis genome contains three homologs - SGR1, SGR2, and SGRL [32]. In rice, overexpression of OsSGR and OsSGRL both cause leaf yellowing during stress-induced and natural senescence processes [30,35,56]. Intriguingly, in Arabidopsis, SGR1 and SGRL positively regulate Chl degradation, while SGR2 is a negative regulator of Chl catabolism during natural and stress-induced senescence, despite 76% amino acid sequence similarity between SGR1 and SGR2[33,51,64,79].
Our experiment revealed that SGR expression is significantly induced in RACK1-UX leaves, while its expression is considerably reduced in RACK1B-OX leaves even with 200 mM NaCl treatment for 24 hours. These findings indicated that altered transcript and proteomic expression of OsSGR directly correlates with the altered expression of OsRACK1B in both normal and high salinity conditions. SGR interacts with six chlorophyll degradation enzymes (CCEs) including NYC1 and RCCR[28]. Taken together, based on the expression patterns of OsSGR and CCEs in our experiment, we assumed that OsRACK1B most likely inhibits the function of OsSGR by physically interacting with it, especially under stress conditions. We tested this hypothesis by using BiFC analysis in onion epidermal cells. Our analysis revealed that OsRACK1B interacts with OsSGR in the nucleus and outside of the nucleus under normal and high salinity stress conditions, suggesting a transcriptional and post-transcriptional regulation of OsSGR by OsRACK1B in chlorophyll catabolism. Although, use of green tissues for such activities would have helped evaluate whether the interaction in the chloroplast is a pre-requisite for inhibiting SGR activities, which functionally take place within the chloroplast.
Although SGR localizes to the thylakoid membrane of chloroplasts[28] , reports suggest that SGRs do not only occur in chloroplasts. Its functional presence and interaction with other proteins were also observed in different locations [78,80]. Elucidating detailed mechanisms of how OsRACK1B bonding with OsSGR inactivates SGR activity or represses its expression needs further studies. Two mechanisms can be postulated for how RACK1B prevents chlorophyll degradation by regula ing OsSGR: the most likely possibility is that by forming a heterodimer with SGR, RACK1B limits the availability of SGR to form the SGR-CCE-LHCII multiprotein complex, a prerequisite for chlor phyll breakdown [33]. This hypothesis can be substantiated by recent findings showing that in Arabidopsis, SGR2 negatively regulates chlorophyll degradation by forming homo- or heterodimers with SGR1 [33,64,79] . Considering SGR2 itself can bind to LHCII, the authors proposed that heter dimerization of SGR1 with SGR2 interrupts the formation of the SGR1–CCE–LHCII protein complexes, and thereby limits Chl degradation under stress conditions.
Since there is no report of an orthologue of SGR2 in rice, the question of whether rice SGRL function in a similar fashion through the formation of a heterodimer with SGR needs further investigation. Another possibility is that tethering between RACK1B and SGR triggers a conformational change that makes the complex a better substrate for proteases. A wide range of chloroplast proteins undergo degradation and removal processes such as autophagy, senescence-associated vacuole (SAV) lysis, or intra-plastidial proteolysis by different forms of proteases (such as Clp, FtsH etc.) for the biogenesis and maintenance of chloroplasts, especially under stress conditions [24,81,82,83]. Of interest, a similar finding was observed in Arabidopsis: Chloroplast Vesiculation (CV) protein interacts with photosystem II subunit PsbO1 via a highly conserved C-terminal domain and appears to alter the structure and stability of the PSII complex, consequently facilitating the degradation of core proteins such as D1 by thylakoid proteases under salt stress conditions[84]. We do not know yet whether OsSGR is subject to such recognition of different N or C termini, modified or not, by the proteases in the chloroplast. Further studies are required to address this potentially important link.
Previous studies have demonstrated that in rice and Arabidopsis, SGR forms a complex together with CCEs and LHCII proteins such as Lhcb1, but not with LHCI or photosystem core proteins. Phosphorylation of LHCII is required for the transition from state-I to state-II for adaptation in response to environmental changes [62,85]. A serine-threonine protein kinase named as STN7 is necessary for phosphorylation of the LHCII to regulate the reversible association between PSII and LHCII during PSII repair [60,86,87]. Our expression analysis revealed a steady level of light-harvesting chlorophyll a/b binding 1 (Lhcb1) protein RACK1B-OX plants, suggesting maintained integrity of LHCII complex in response to salinity stress. Conversely, high activity of SGR prompted the degradation of LHCII complex, as reflected in the low amount of Lhcb1 protein and reduced expression of STN7 coinciding the amount of non-phosphorylated Lhcb1. Since state transitions depend on STN7 kinase[60], our data can be linked to the functional stability of the chloroplast and a redox balance between PSII and PSI for photosynthetic efficiency in response to stress conditions. Being localized mostly in the nucleus and cytoplasm it is unclear whether RACK1 shuttles photosynthetic proteins in the chloroplast to form a large complex in the photosynthesis process or works as a molecular chaperone for structural remodeling of the light-harvesting complex during state transitions. Given that RACK1 interacts with chlorophyll a/b binding protein, we cannot rule out the possibility that OsRACK1B might interact with other CCEs or LHCII subunits. Taken together, we conclude that RACK1B is required for the dynamics of PSII–LHCII supramolecular complexes under stress conditions.
There are limitations to our study. First, we could not confirm whether the stay-green phenotype is functional or cosmetic stay-green and leaves of OsRACKB-OX plants were active in photosynthesis during salinity stress condition. This is important since functional stay-greens retain both Chl and photosynthetic capacity longer than WT, and cosmetic stay-greens retain Chl but undergo other aspects of senescence as WT[36] . This can be assessed in the future using Fv/Fm measurements. Second, we could not test that if the SGR and RACK1B protein coexist in the similar multiporotein protein complex. A pull-down assay would confirm the hypothesis. Also, our present experimental approach of BiFC in onion peel cells did not allow us to evaluate whether the cellular localization of RACK1B-SGR interaction has any influence on chlorophyll catabolism. Since, SGR localizes to the thylakoid membrane of chloroplasts and functions for chlorophyll breakdown by forming the SGR-CCE-LHCII multiprotein complex, further studies are needed to prove the interaction between RACK1B and SGR in plant cells with chloroplasts such as Arabidopsis mesophyll cell protoplasts.
4. Materials and Methods
4.1. Plant materials, growth condition, and stress treatment
We identified OsRACK1 T-DNA insertion lines from Rice functional genomic express database (http://signal.salk.edu/cgi-bin/RiceGE). T-DNA tagged lines PFG_3A-60871.L and PFG_3A-07870.R both in the Dongjin (Oryza sativa ssp. Japonica cv. Dongjin) background and PFG_3D-02734.L in the Hwayoung (Oryza sativa ssp. Japonica cv. Hwayoung) background seeds were purchased from Crop Biotech Inst., Korea [39,40] Seeds were surface sterilized and germinated on full strength Murashige and Skoog (MS) [41] culture media (Caisson Laboratories, Inc.) at room temperature. Leaves from two-week-old germinated seedlings were used for DNA extraction and genotyping. Wild-type (WT) and selected transgenic lines were then transferred to hydroponic nutrient solution prepared following the protocol as described by Lakshmanan et al., (2015). Plants were grown at 28º C during a 14 h light (300 μmol m-2 s-1) period and 24 °C during 10 h of darkness with 60% relative humidity.
For salt stress treatment, eight-week-old plants were transferred to hydroponic solution in which NaCl (200 mM) was added gradually. After 24 hours, plants were washed with tap water and placed in the fresh hydroponic solution for recovery. Leaf tissues from plants were sampled before and after salt stress, flash-frozen in liquid nitrogen and stored at –80 °C for protein and RNA extraction.
4.2. Genotyping of the T-DNA flanking region of OsRACK1B transgenic lines
Transgenic lines were screened for possible T-DNA insertions positioned near RACK1B (Loc_Os05g47890) by PCR. Genotyping of RACK1B overexpression lines were performed according to the protocol described in Rahman et al., 2022 [11] . For putative loss of function lines, similar genotyping PCR was performed for Salk line PFG_3D-027334 using T-DNA and gene-specific primers revealing T-DNA insertion in two rice plants from PFG_3D-02734 lines.
4.3. RNA extraction, Complementary DNA (cDNA) Synthesis and quantitative Reverse Transcriptase PCR (qRT-PCR) analysis
Total RNA was extracted before and after salt stress from eight-week-old rice leaves. Freshly harvested leaves were ground in fine powder using liquid nitrogen and pre-chiled mortar and pestle. 100 mg of powdered tissue sample was used for RNA extraction using the RNeasy Plant Mini Kit (Qiagen, USA). Finally, 500 ng of total RNA for each sample was reverse transcribed to make cDNA using SuperScript IV VILO Master Mix kit (Thermo Fisher Scientific, USA) following manufacturers instruction.
Quantitative Reverse-Transcriptase PCR (qRT-PCR) was performed using cDNA and PowerUP SYBR Green master mix (Thermo Fisher Scientific,USA) with a CFX96 real-time PCR detection system (Bio-Rad,USA). Normalized expression (DDC(t) method) was calculated using the Bio-Rad CFX manager software employing the housekeeping gene OsActin1 (Os03g50885) as a reference gene. All experiments were performed in triplicate for technical repeats. The results were plotted as relative values ±SEM and graphically displayed using GraphPad Prism version 8.2.0 (GraphPad Software Inc., USA). The primer sequences used are listed in Table S2.
4.4. Protein Extraction and Western blot analysis
Total protein from leaves was extracted from 100 mg of finely powdered tissue using lysis buffer (CelLytic P, Sigma-Aldrich) containing protease and phosphatase inhibitor cocktail (Sigma-Aldrich, USA). Total protein content for each sample was quantified by Bradford assay using Quick Start Bradford dye reagent (Bio-Rad,USA).
Western blot analysis was performed using an equal amount of total proteins per sample and resolved in 4-12% pre-cast XT-MES gel (Bio-Rad). The proteins were transferred to polyvinylidene difluoride (PVDF) membrane, incubated with primary polyclonal antibodies and secondary antibodies. Membranes were developed using the ECL detection kit (Bio-Rad, USA). The signals of the bands were visualized and captured using the Chemi-DocXRS system (Bio-Rad, USA). The OsRACK1B peptide specific mAb antibodies were raised using two peptides- AGVLRGHNDM and QDLKPEVQAF corresponding to amino acids 10-19 at the N-terminus and 286-295 at C-terminus end OsRACK1B protein sequences, respectively (Abmart, China). Commercially purchased Anti-Lhcb1 (cat# AS01004 Agrisera, Sweden) antibody used in study demonstrated reactivity as stated in the manufacturer’s website and as shown previously by Liu et al., 2019. Similarly, a polyclonal Anti-SGR (Cat# PHY1024S, PhytoAB Inc., USA) antibody was used. Because of the high conservation and specificity of the peptides, anti-SGR antibody derived from synthetic peptide using Arabidopsis SGR (AT4G22920) was used to detect OsSGR. Antibody binding was validated against Arabidopsis and rice protein samples to confirm reactivity and specificity against SGR protein (data not shown). Ponceau-S (40% methanol (v/v), 15% acetic acid (v/v), 0.25% Ponceau-S) staining of Rubisco was used as loading control.
4.5. Bimolecular Fluorescence Complementation (BiFC) Assay
For BiFC assay, total RNA was extracted from eight-week-old wild-type rice leaves using Trizol reagent (Thermo Fisher, USA) according to the manufacturer's instructions. 1 µg of purified total RNA per sample was reverse transcribed using SuperScript™ IV VILO Master Mix kit (Thermo Fisher, USA) following manufacturers instruction. Full-length coding sequences of OsRACK1B and OsSGR with or without stop codons were amplified by PCR using Q5® High-Fidelity DNA Polymerase (NEB, MA, USA). Amplicons were purified (QIAquick PCR Purification Kit, Qiagen, USA), sanger sequenced (Genewiz, USA) and cloned into the Gateway entry vector pCR8/GW/TOPO (Invitrogen, CA) according to the manufacturer’s instruction. Orientation was confirmed by sequencing the plasmids from selected colonies. Entry clones were sub-cloned into the BiFC plasmid sets pSAT5-DEST-cEYFP(175-end)-C1(pE3130), pSAT5(A)-DEST-cEYFP(175-end)-N1 (pE3132), pSAT4(A)-DESTnEYFP(1-174)-N1 (pE3134), and pSAT4-DEST-nEYFP(1-174)-C1 (pE3136) destination vectors (https://www.bio.purdue.edu/people/faculty/gelvin/nsf/protocols_vectors.htm) using Gateway LR clonase-II enzyme mix (Invitrogen, USA). Orientation was further confirmed by sanger sequencing of the fusion plasmids isolated (PureLink® HiPure Plasmid Midiprep Kit, Invitrogen) from selected colonies using primers listed in SI Table 2. Each pair of recombinant plasmids encoding nEYFP and cEYFP fusions was mixed 1:1 (w/w), co-bombarded with gold particles (1 um, Au, Bio-Rad) into onion epidermal layers. Each pair of recombinant plasmids encoding nEYFP or cEYFP fusion proteins was co-bombarded into onion epidermal cells using Helios DNA particle delivery system (Biolistic PDS-1000/He, BioRad) as described by Hollender and Liu (2010) [43]. Bombarded epidermal cells were incubated in MS liquid media for 16–24 hr at 22°C under dark incubation, followed by observation for YFP fluorescence with an inverted spinning-disk confocal microscope (Eclipse Ti-E-PFS, Nikon, USA). For salt treatment, onion cells were incubated in 200 mM NaCl for 30 min before imaging. Confocal fluorescent images and DIC images were acquired and processed using the Nikon NIS-Elements software. AtRACK1A was used as a positive control as described by Sabila et al., 2016 [44]. Negative controls for interaction were provided by empty YFPC and YFPN vectors in combination with the OsRACK1 and OsSGR1 pSAT expression plasmids (Methods S2). Primers used for BiFC plasmids are listed in Table S3.
4.6. Leaf disc assay and chlorophyll pigment analysis
Leaf discs were excised from fully expanded leaves of eight-week-old WT and transgenic plants. Then leaf strips were floated on water (as experimental control) or 200 mM NaCl (for salinity stress) in 6 well plates. Plates were kept under continuous light for 72 hours at room temperature. After 72 hours, leaf discs were photographed (Leica EZ4 stereo microscope) and used for spectrophotometric measurement of chlorophyll contents using protocols described previously [45]. Briefly, equal size leaf discs were incubated with 7.5 ml DMSO at 650 C for 30 min. The extracted chlorophyll solution was made up to 10 ml with DMSO. 3 ml of the total extracted solution for each sample was transferred to a cuvette, and the OD values were measured at 645 and 663 nm using GENESYS 20 UV/Vis spectrophotometer (Thermo Scientific, USA). The Chlorophyll concentration was calculated as described by Arnon (1949) [46] using the following equation:
Total Chlorophyll (ug/mL) = 0.0202 A663 + 0.00802 A645, where: A645 = absorbance at a wavelength of 645 nm; A663 = absorbance at a wavelength of 663 nm.
For pigment analysis, the acquired images were processed using the free ImageJ software (NIH, USA). Color thresholding was applied, in RGB color space, adjusting the parameters in order to select the green pigment and separate them from the background
5. Conclusions
In this work, we reveal the physiological function of RACK1 in chlorophyll catabolism which was enigmatic since, to date, reports have only indicated its interaction with photosynthesis-related proteins. Delaying senescence or increasing chlorophyll stability during stress conditions is linked to increased photosynthetic efficiency and improved grain yield [88,89]. Grain filling and increased yields are agronomic traits of interest. Therefore, our study provides important insights into the molecular mechanisms of salt-tolerance and could be used to ameliorate the effect of salt on photosynthesis to increase the yield potential of rice, an important cereal crop, under current global climate change.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
M.A.R & H.U planned and designed the experiments. M.A.R performed the experiments, analyzed the data, and wrote the manuscript; H.U conceived the project and contributed to analyze the data and critical revision of the article.
Funding
This work was partly supported by the National Science Foundation grant to HU (MCB 0542312). M.A.R thanks the Dept. of Biology and Graduate School of the HU for the Teaching Assistantship and E. Just-Percy L. Julian Graduate Research Assistantship. Any opinions, findings, conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation, USA.
Data Availability Statement
Acknowledgments
Fluorescence images were obtained with a spinning disk confocal fluorescent microscope acquired through a Department of Defense HBCU/MI Equipment/Instrumentation Grant (#64684-RT-REP) to Anna K. Allen of the Biology Department, Howard University. We thank Clarence M Lee of the Biology department; Karl Thompson, and Qiyi Tang of the Medical School, HU, for granting access to their laboratory facilities. Plasmids for BiFC analysis were kindly provided by Stanton Gelvin of Purdue University. Rice mutant seeds were obtained from G. An, Republic of Korea.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Islas-Flores, T.; Rahman, A.; Ullah, H.; Villanueva, M.A. The Receptor for Activated C Kinase in Plant Signaling: Tale of a Promiscuous Little Molecule. Front Plant Sci 2015, 6, 1090. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.R.; Ron, D.; Kiely, P.A. RACK1, A multifaceted scaffolding protein: Structure and function. Cell Commun Signal 2011, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- McCahill, A.; Warwicker, J.; Bolger, G.B.; Houslay, M.D.; Yarwood, S.J. The RACK1 scaffold protein: a dynamic cog in cell response mechanisms. Mol Pharmacol 2002, 62, 1261–1273. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.H.; Flygaard, R.K.; Jenner, L.B. Structural analysis of ribosomal RACK1 and its role in translational control. Cell Signal 2017, 35, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Ron, D.; Adams, D.R.; Baillie, G.S.; Long, A.; O'Connor, R.; Kiely, P.A. RACK1 to the future--a historical perspective. Cell Commun Signal 2013, 11, 53. [Google Scholar] [CrossRef]
- Xiao, T.; Zhu, W.; Huang, W.; Lu, S.S.; Li, X.H.; Xiao, Z.Q.; Yi, H. RACK1 promotes tumorigenicity of colon cancer by inducing cell autophagy. Cell Death Dis 2018, 9, 1148. [Google Scholar] [CrossRef]
- Chen, J.G. Phosphorylation of RACK1 in plants. Plant Signal Behav 2015, 10, e1022013. [Google Scholar] [CrossRef]
- Ullah, H.; Scappini, E.L.; Moon, A.F.; Williams, L.V.; Armstrong, D.L.; Pedersen, L.C. Structure of a signal transduction regulator, RACK1, from Arabidopsis thaliana. Protein Sci 2008, 17, 1771–1780. [Google Scholar] [CrossRef]
- Chen, J.G.; Ullah, H.; Temple, B.; Liang, J.; Guo, J.; Alonso, J.M.; Ecker, J.R.; Jones, A.M. RACK1 mediates multiple hormone responsiveness and developmental processes in Arabidopsis. J Exp Bot 2006, 57, 2697–2708. [Google Scholar] [CrossRef]
- Guo, J.; Chen, J.G. RACK1 genes regulate plant development with unequal genetic redundancy in Arabidopsis. BMC Plant Biol 2008, 8, 108. [Google Scholar] [CrossRef]
- Rahman, M.A.; Fennell, H.; Ullah, H. Receptor for Activated C Kinase1B (OsRACK1B) Impairs Fertility in Rice through NADPH-Dependent H2O2 Signaling Pathway. International Journal of Molecular Sciences 2022, 23, 8455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, Y.; Shen, J.; Yin, J.; Li, D.; Gao, Y.; Xu, W.; Liang, J. OsRACK1A, encodes a circadian clock-regulated WD40 protein, negatively affect salt tolerance in rice. Rice (N Y) 2018, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Kundu, N.; Dozier, U.; Deslandes, L.; Somssich, I.E.; Ullah, H. Arabidopsis scaffold protein RACK1A interacts with diverse environmental stress and photosynthesis related proteins. Plant Signal Behav 2013, 8, e24012. [Google Scholar] [CrossRef] [PubMed]
- Urano, D.; Czarnecki, O.; Wang, X.; Jones, A.M.; Chen, J.G. Arabidopsis receptor of activated C kinase1 phosphorylation by WITH NO LYSINE8 KINASE. Plant Physiol 2015, 167, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Chen, L.; Thao, N.P.; Fujiwara, M.; Wong, H.L.; Kuwano, M.; Umemura, K.; Shirasu, K.; Kawasaki, T.; Shimamoto, K. RACK1 functions in rice innate immunity by interacting with the Rac1 immune complex. Plant Cell 2008, 20, 2265–2279. [Google Scholar] [CrossRef]
- Hoang, T.M.; Moghaddam, L.; Williams, B.; Khanna, H.; Dale, J.; Mundree, S.G. Development of salinity tolerance in rice by constitutive-overexpression of genes involved in the regulation of programmed cell death. Front Plant Sci 2015, 6, 175. [Google Scholar] [CrossRef]
- Allu, A.D.; Soja, A.M.; Wu, A.; Szymanski, J.; Balazadeh, S. Salt stress and senescence: identification of cross-talk regulatory components. J Exp Bot 2014, 65, 3993–4008. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P. Photosynthesis under stressful environments: an overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Suo, J.; Zhao, Q.; David, L.; Chen, S.; Dai, S. Salinity Response in Chloroplasts: Insights from Gene Characterization. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front Plant Sci 2015, 6, 69. [Google Scholar] [CrossRef]
- Van Hautegem, T.; Waters, A.J.; Goodrich, J.; Nowack, M.K. Only in dying, life: programmed cell death during plant development. Trends Plant Sci 2015, 20, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Balazadeh, S.; Riano-Pachon, D.M.; Mueller-Roeber, B. Transcription factors regulating leaf senescence in Arabidopsis thaliana. Plant Biol (Stuttg) 2008, 10 Suppl 1, 63–75. [Google Scholar] [CrossRef]
- Balazadeh, S.; Siddiqui, H.; Allu, A.D.; Matallana-Ramirez, L.P.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Kohler, B.; Mueller-Roeber, B. A gene regulatory network controlled by the NAC transcription factor ANAC092/AtNAC2/ORE1 during salt-promoted senescence. Plant J 2010, 62, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Mendoza, M.; Velasco-Arroyo, B.; Santamaria, M.E.; Gonzalez-Melendi, P.; Martinez, M.; Diaz, I. Plant senescence and proteolysis: two processes with one destiny. Genet Mol Biol 2016, 39, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Hortensteiner, S. Chlorophyll degradation during senescence. Annu Rev Plant Biol 2006, 57, 55–77. [Google Scholar] [CrossRef] [PubMed]
- Hortensteiner, S.; Krautler, B. Chlorophyll breakdown in higher plants. Biochim Biophys Acta 2011, 1807, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Sade, N.; Del Mar Rubio-Wilhelmi, M.; Umnajkitikorn, K.; Blumwald, E. Stress-induced senescence and plant tolerance to abiotic stress. J Exp Bot 2018, 69, 845–853. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Schelbert, S.; Park, S.Y.; Han, S.H.; Lee, B.D.; Andres, C.B.; Kessler, F.; Hortensteiner, S.; Paek, N.C. STAY-GREEN and chlorophyll catabolic enzymes interact at light-harvesting complex II for chlorophyll detoxification during leaf senescence in Arabidopsis. Plant Cell 2012, 24, 507–518. [Google Scholar] [CrossRef]
- Kusaba, M.; Ito, H.; Morita, R.; Iida, S.; Sato, Y.; Fujimoto, M.; Kawasaki, S.; Tanaka, R.; Hirochika, H.; Nishimura, M.; et al. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell 2007, 19, 1362–1375. [Google Scholar] [CrossRef]
- Jiang, H.; Li, M.; Liang, N.; Yan, H.; Wei, Y.; Xu, X.; Liu, J.; Xu, Z.; Chen, F.; Wu, G. Molecular cloning and function analysis of the stay green gene in rice. Plant J 2007, 52, 197–209. [Google Scholar] [CrossRef]
- Park, S.Y.; Yu, J.W.; Park, J.S.; Li, J.; Yoo, S.C.; Lee, N.Y.; Lee, S.K.; Jeong, S.W.; Seo, H.S.; Koh, H.J.; et al. The senescence-induced staygreen protein regulates chlorophyll degradation. Plant Cell 2007, 19, 1649–1664. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; An, K.; Liao, Y.; Zhou, X.; Cao, Y.; Zhao, H.; Ge, X.; Kuai, B. Identification of a novel chloroplast protein AtNYE1 regulating chlorophyll degradation during leaf senescence in Arabidopsis. Plant Physiol 2007, 144, 1429–1441. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Park, S.Y.; Paek, N.C. The Divergent Roles of STAYGREEN (SGR) Homologs in Chlorophyll Degradation. Mol Cells 2015, 38, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Christ, B.; Hörtensteiner, S. Mechanism and significance of chlorophyll breakdown. Journal of plant growth regulation 2014, 33, 4–20. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, Y.; Li, M.; Xu, X.; Wu, G. Overexpression of SGR results in oxidative stress and lesion-mimic cell death in rice seedlings. J Integr Plant Biol 2011, 53, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.; Howarth, C.J. Five ways to stay green. Journal of experimental botany 2000, 51, 329–337. [Google Scholar] [CrossRef]
- Pinto, R.S.; Lopes, M.S.; Collins, N.C.; Reynolds, M.P. Modelling and genetic dissection of staygreen under heat stress. Theoretical and Applied Genetics 2016, 129, 2055–2074. [Google Scholar] [CrossRef]
- Ülker, B.; Peiter, E.; Dixon, D.P.; Moffat, C.; Capper, R.; Bouché, N.; Edwards, R.; Sanders, D.; Knight, H.; Knight, M.R. Getting the most out of publicly available T-DNA insertion lines. The Plant Journal 2008, 56, 665–677. [Google Scholar] [CrossRef]
- An, S.; Park, S.; Jeong, D.-H.; Lee, D.-Y.; Kang, H.-G.; Yu, J.-H.; Hur, J.; Kim, S.-R.; Kim, Y.-H.; Lee, M. Generation and analysis of end sequence database for T-DNA tagging lines in rice. Plant physiology 2003, 133, 2040–2047. [Google Scholar] [CrossRef]
- Jeong, D.H.; An, S.; Park, S.; Kang, H.G.; Park, G.G.; Kim, S.R.; Sim, J.; Kim, Y.O.; Kim, M.K.; Kim, S.R.; et al. Generation of a flanking sequence-tag database for activation-tagging lines in japonica rice. Plant J 2006, 45, 123–132. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia plantarum 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Citovsky, V.; Lee, L.Y.; Vyas, S.; Glick, E.; Chen, M.H.; Vainstein, A.; Gafni, Y.; Gelvin, S.B.; Tzfira, T. Subcellular localization of interacting proteins by bimolecular fluorescence complementation in planta. J Mol Biol 2006, 362, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Hollender, C.A.; Liu, Z. Bimolecular fluorescence complementation (BiFC) assay for protein-protein interaction in onion cells using the helios gene gun. Journal of visualized experiments: JoVE 2010.
- Sabila, M.; Kundu, N.; Smalls, D.; Ullah, H. Tyrosine Phosphorylation Based Homo-dimerization of Arabidopsis RACK1A Proteins Regulates Oxidative Stress Signaling Pathways in Yeast. Front Plant Sci 2016, 7, 176. [Google Scholar] [CrossRef] [PubMed]
- Hiscox, J.; Israelstam, G. A method for the extraction of chlorophyll from leaf tissue without maceration. Canadian journal of botany 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Kim, J.; Kim, J.H.; Lyu, J.I.; Woo, H.R.; Lim, P.O. New insights into the regulation of leaf senescence in Arabidopsis. J Exp Bot 2018, 69, 787–799. [Google Scholar] [CrossRef]
- Leng, Y.; Ye, G.; Zeng, D. Genetic Dissection of Leaf Senescence in Rice. Int J Mol Sci 2017, 18. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.H.; Yoo, E.S.; Lee, C.H.; Hirochika, H.; An, G. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol Biol 2005, 57, 805–818. [Google Scholar] [CrossRef]
- Tanaka, R.; Koshino, Y.; Sawa, S.; Ishiguro, S.; Okada, K.; Tanaka, A. Overexpression of chlorophyllide a oxygenase (CAO) enlarges the antenna size of photosystem II in Arabidopsis thaliana. The Plant Journal 2001, 26, 365–373. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Jeong, J.; Kang, M.Y.; Kim, J.; Paek, N.C.; Choi, G. Phytochrome-interacting transcription factors PIF4 and PIF5 induce leaf senescence in Arabidopsis. Nat Commun 2014, 5, 4636. [Google Scholar] [CrossRef]
- Xiao, H.J.; Liu, K.K.; Li, D.W.; Arisha, M.H.; Chai, W.G.; Gong, Z.H. Cloning and characterization of the pepper CaPAO gene for defense responses to salt-induced leaf senescence. BMC Biotechnol 2015, 15, 100. [Google Scholar] [CrossRef] [PubMed]
- Rauf, M.; Arif, M.; Dortay, H.; Matallana-Ramirez, L.P.; Waters, M.T.; Gil Nam, H.; Lim, P.O.; Mueller-Roeber, B.; Balazadeh, S. ORE1 balances leaf senescence against maintenance by antagonizing G2-like-mediated transcription. EMBO Rep 2013, 14, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, Y.; Ito, H.; Tanaka, A. Arabidopsis STAY-GREEN, Mendel's Green Cotyledon Gene, Encodes Magnesium-Dechelatase. Plant Cell 2016, 28, 2147–2160. [Google Scholar] [CrossRef] [PubMed]
- Hortensteiner, S. Stay-green regulates chlorophyll and chlorophyll-binding protein degradation during senescence. Trends Plant Sci 2009, 14, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Rong, H.; Tang, Y.; Zhang, H.; Wu, P.; Chen, Y.; Li, M.; Wu, G.; Jiang, H. The Stay-Green Rice like (SGRL) gene regulates chlorophyll degradation in rice. J Plant Physiol 2013, 170, 1367–1373. [Google Scholar] [CrossRef]
- Kuai, B.; Chen, J.; Hortensteiner, S. The biochemistry and molecular biology of chlorophyll breakdown. J Exp Bot 2018, 69, 751–767. [Google Scholar] [CrossRef]
- Matsuda, K.; Shimoda, Y.; Tanaka, A.; Ito, H. Chlorophyll a is a favorable substrate for Chlamydomonas Mg-dechelatase encoded by STAY-GREEN. Plant Physiol Biochem 2016, 109, 365–373. [Google Scholar] [CrossRef]
- Pietrzykowska, M.; Suorsa, M.; Semchonok, D.A.; Tikkanen, M.; Boekema, E.J.; Aro, E.M.; Jansson, S. The light-harvesting chlorophyll a/b binding proteins Lhcb1 and Lhcb2 play complementary roles during state transitions in Arabidopsis. Plant Cell 2014, 26, 3646–3660. [Google Scholar] [CrossRef]
- Bellafiore, S.; Barneche, F.; Peltier, G.; Rochaix, J.D. State transitions and light adaptation require chloroplast thylakoid protein kinase STN7. Nature 2005, 433, 892–895. [Google Scholar] [CrossRef]
- Leoni, C.; Pietrzykowska, M.; Kiss, A.Z.; Suorsa, M.; Ceci, L.R.; Aro, E.M.; Jansson, S. Very rapid phosphorylation kinetics suggest a unique role for Lhcb2 during state transitions in Arabidopsis. Plant J 2013, 76, 236–246. [Google Scholar] [CrossRef]
- Chen, Y.; Hoehenwarter, W. Changes in the phosphoproteome and metabolome link early signaling events to rearrangement of photosynthesis and central metabolism in salinity and oxidative stress response in Arabidopsis. Plant Physiology 2015, 169, 3021–3033. [Google Scholar] [CrossRef] [PubMed]
- Minagawa, J. State transitions--the molecular remodeling of photosynthetic supercomplexes that controls energy flow in the chloroplast. Biochim Biophys Acta 2011, 1807, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Kim, D.; Kim, Y.S.; Hortensteiner, S.; Paek, N.C. Arabidopsis STAYGREEN-LIKE (SGRL) promotes abiotic stress-induced leaf yellowing during vegetative growth. FEBS Lett 2014, 588, 3830–3837. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.W.; Lee, Y.J.; Koh, H.J.; Lee, B.M.; Nam, Y.W.; Paek, N.C. Isolation, characterization, and mapping of the stay green mutant in rice. Theor Appl Genet 2002, 104, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.; Sato, Y.; Masuda, Y.; Nishimura, M.; Kusaba, M. Defect in non-yellow coloring 3, an alpha/beta hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J 2009, 59, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, M.; Chen, Y.; Wu, P.; Wu, G.; Jiang, H. Knockdown of OsPAO and OsRCCR1 cause different plant death phenotypes in rice. J Plant Physiol 2011, 168, 1952–1959. [Google Scholar] [CrossRef] [PubMed]
- Pruzinska, A.; Anders, I.; Aubry, S.; Schenk, N.; Tapernoux-Luthi, E.; Muller, T.; Krautler, B.; Hortensteiner, S. In vivo participation of red chlorophyll catabolite reductase in chlorophyll breakdown. Plant Cell 2007, 19, 369–387. [Google Scholar] [CrossRef]
- Pruzinska, A.; Tanner, G.; Anders, I.; Roca, M.; Hortensteiner, S. Chlorophyll breakdown: pheophorbide a oxygenase is a Rieske-type iron-sulfur protein, encoded by the accelerated cell death 1 gene. Proc Natl Acad Sci U S A 2003, 100, 15259–15264. [Google Scholar] [CrossRef]
- Sato, Y.; Morita, R.; Katsuma, S.; Nishimura, M.; Tanaka, A.; Kusaba, M. Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J 2009, 57, 120–131. [Google Scholar] [CrossRef]
- Kakizaki, T.; Inaba, T. New insights into the retrograde signaling pathway between the plastids and the nucleus. Plant Signal Behav 2010, 5, 196–199. [Google Scholar] [CrossRef]
- Wamaitha, M.J.; Yamamoto, R.; Wong, H.L.; Kawasaki, T.; Kawano, Y.; Shimamoto, K. OsRap2.6 transcription factor contributes to rice innate immunity through its interaction with Receptor for Activated Kinase-C 1 (RACK1). Rice (N Y) 2012, 5, 35. [Google Scholar] [CrossRef] [PubMed]
- Dongping, Z.; Li, C.; Bing, L.; Jiansheng, L. The scaffolding protein RACK1: a platform for diverse functions in the plant kingdom. Journal of Plant Biology & Soil Health 2013, 1. [Google Scholar]
- Li, J.J.; Xie, D. RACK1, a versatile hub in cancer. Oncogene 2015, 34, 1890–1898. [Google Scholar] [CrossRef] [PubMed]
- Pulido, P.; Zagari, N.; Manavski, N.; Gawronski, P.; Matthes, A.; Scharff, L.B.; Meurer, J.; Leister, D. CHLOROPLAST RIBOSOME ASSOCIATED Supports Translation under Stress and Interacts with the Ribosomal 30S Subunit. Plant Physiol 2018, 177, 1539–1554. [Google Scholar] [CrossRef] [PubMed]
- Armstead, I.; Donnison, I.; Aubry, S.; Harper, J.; Hortensteiner, S.; James, C.; Mani, J.; Moffet, M.; Ougham, H.; Roberts, L.; et al. Cross-species identification of Mendel's I locus. Science 2007, 315, 73. [Google Scholar] [CrossRef] [PubMed]
- Barry, C.S.; McQuinn, R.P.; Chung, M.Y.; Besuden, A.; Giovannoni, J.J. Amino acid substitutions in homologs of the STAY-GREEN protein are responsible for the green-flesh and chlorophyll retainer mutations of tomato and pepper. Plant Physiol 2008, 147, 179–187. [Google Scholar] [CrossRef]
- Zhou, C.; Han, L.; Pislariu, C.; Nakashima, J.; Fu, C.; Jiang, Q.; Quan, L.; Blancaflor, E.B.; Tang, Y.; Bouton, J.H.; et al. From model to crop: functional analysis of a STAY-GREEN gene in the model legume Medicago truncatula and effective use of the gene for alfalfa improvement. Plant Physiol 2011, 157, 1483–1496. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Park, S.Y.; Kim, Y.S.; Wang, S.H.; Yoo, S.C.; Hortensteiner, S.; Paek, N.C. Arabidopsis STAY-GREEN2 is a negative regulator of chlorophyll degradation during leaf senescence. Mol Plant 2014, 7, 1288–1302. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, J.; Li, J.; Yang, C.; Wang, T.; Ouyang, B.; Li, H.; Giovannoni, J.; Ye, Z. A STAY-GREEN protein SlSGR1 regulates lycopene and beta-carotene accumulation by interacting directly with SlPSY1 during ripening processes in tomato. New Phytol 2013, 198, 442–452. [Google Scholar] [CrossRef]
- Ishida, H.; Yoshimoto, K. Chloroplasts are partially mobilized to the vacuole by autophagy. Autophagy 2008, 4, 961–962. [Google Scholar] [CrossRef]
- Izumi, M.; Nakamura, S. Chloroplast Protein Turnover: The Influence of Extraplastidic Processes, Including Autophagy. Int J Mol Sci 2018, 19. [Google Scholar] [CrossRef]
- Wada, S.; Ishida, H.; Izumi, M.; Yoshimoto, K.; Ohsumi, Y.; Mae, T.; Makino, A. Autophagy plays a role in chloroplast degradation during senescence in individually darkened leaves. Plant Physiol 2009, 149, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Blumwald, E. Stress-induced chloroplast degradation in Arabidopsis is regulated via a process independent of autophagy and senescence-associated vacuoles. Plant Cell 2014, 26, 4875–4888. [Google Scholar] [CrossRef] [PubMed]
- Fristedt, R.; Willig, A.; Granath, P.; Crevecoeur, M.; Rochaix, J.-D.; Vener, A.V. Phosphorylation of photosystem II controls functional macroscopic folding of photosynthetic membranes in Arabidopsis. The Plant Cell 2009, 21, 3950–3964. [Google Scholar] [CrossRef] [PubMed]
- Bonardi, V.; Pesaresi, P.; Becker, T.; Schleiff, E.; Wagner, R.; Pfannschmidt, T.; Jahns, P.; Leister, D. Photosystem II core phosphorylation and photosynthetic acclimation require two different protein kinases. Nature 2005, 437, 1179–1182. [Google Scholar] [CrossRef]
- Pesaresi, P.; Hertle, A.; Pribil, M.; Kleine, T.; Wagner, R.; Strissel, H.; Ihnatowicz, A.; Bonardi, V.; Scharfenberg, M.; Schneider, A.; et al. Arabidopsis STN7 kinase provides a link between short- and long-term photosynthetic acclimation. Plant Cell 2009, 21, 2402–2423. [Google Scholar] [CrossRef]
- Gregersen, P.L.; Culetic, A.; Boschian, L.; Krupinska, K. Plant senescence and crop productivity. Plant molecular biology 2013, 82, 603–622. [Google Scholar] [CrossRef]
- Sade, N.; Umnajkitikorn, K.; Rubio Wilhelmi, M.d.M.; Wright, M.; Wang, S.; Blumwald, E. Delaying chloroplast turnover increases water-deficit stress tolerance through the enhancement of nitrogen assimilation in rice. Journal of experimental botany 2017, 69, 867–878. [Google Scholar] [CrossRef]
Figure 1.
Characterization of T-DNA insertion lines with analysis of gene expression and protein abundance. A. Schematic diagram depicting the positions of T-DNA insertions in OsRACK1B. Gray, orange and white bars represent the exons, 5´-UTR, 3´-UTR region respectively. ATG & TGA are start and stop codons. The gray line represents the intron. The triangles indicate OsRack1b mutant alleles (OX-1, PFG_3A-07870.R; OX-2, PFG_3A-60871.L; UX-1 and UX-2, PFG_3D-02734.L). T-DNA insertion in PFG_3A-60871.L and PFG_3A-07870.R lines (green inverted triangle) resulted in RACK1 overexpression shown in B and D. Insertion in PFG_3D-02734.L (orange inverted triangle) is down-regulated allele revealed by expression analysis shown in Western blot (C) and qRT-PCR (E). (B, C) Western blot analysis of the OsRACK1B in Four-week-old wild-type (WT) and OsRack1b mutant rice plants. Equal amount of leaf proteins extracted from the indicated genotypes was subjected to immunoblot analysis stained with anti-OsRACK1B antibody. Ponceau-stained membrane is shown as the loading control. Molecular weight markers are indicated in kDa. (D, E) qRT-PCR analysis of normalized expression level of OsRACK1B in transgenic rice lines in compared to their respective wild-type plants. Total RNA was extracted from leaf tissues sampled from detached leaves of four-week-old wild-type and transgenic plants as described in Methods. OsActin-1(LOC4333919) was used to standardize transcript levels in each sample. The data are shown as the means ± SE of three technical repeats. Single (*) and double asterisks (**) indicate significant differences compared with the wild-type values (Student’s t-test; * P < 0.05; ** P<0.01). Primers are listed in Table S1.
Figure 1.
Characterization of T-DNA insertion lines with analysis of gene expression and protein abundance. A. Schematic diagram depicting the positions of T-DNA insertions in OsRACK1B. Gray, orange and white bars represent the exons, 5´-UTR, 3´-UTR region respectively. ATG & TGA are start and stop codons. The gray line represents the intron. The triangles indicate OsRack1b mutant alleles (OX-1, PFG_3A-07870.R; OX-2, PFG_3A-60871.L; UX-1 and UX-2, PFG_3D-02734.L). T-DNA insertion in PFG_3A-60871.L and PFG_3A-07870.R lines (green inverted triangle) resulted in RACK1 overexpression shown in B and D. Insertion in PFG_3D-02734.L (orange inverted triangle) is down-regulated allele revealed by expression analysis shown in Western blot (C) and qRT-PCR (E). (B, C) Western blot analysis of the OsRACK1B in Four-week-old wild-type (WT) and OsRack1b mutant rice plants. Equal amount of leaf proteins extracted from the indicated genotypes was subjected to immunoblot analysis stained with anti-OsRACK1B antibody. Ponceau-stained membrane is shown as the loading control. Molecular weight markers are indicated in kDa. (D, E) qRT-PCR analysis of normalized expression level of OsRACK1B in transgenic rice lines in compared to their respective wild-type plants. Total RNA was extracted from leaf tissues sampled from detached leaves of four-week-old wild-type and transgenic plants as described in Methods. OsActin-1(LOC4333919) was used to standardize transcript levels in each sample. The data are shown as the means ± SE of three technical repeats. Single (*) and double asterisks (**) indicate significant differences compared with the wild-type values (Student’s t-test; * P < 0.05; ** P<0.01). Primers are listed in Table S1.
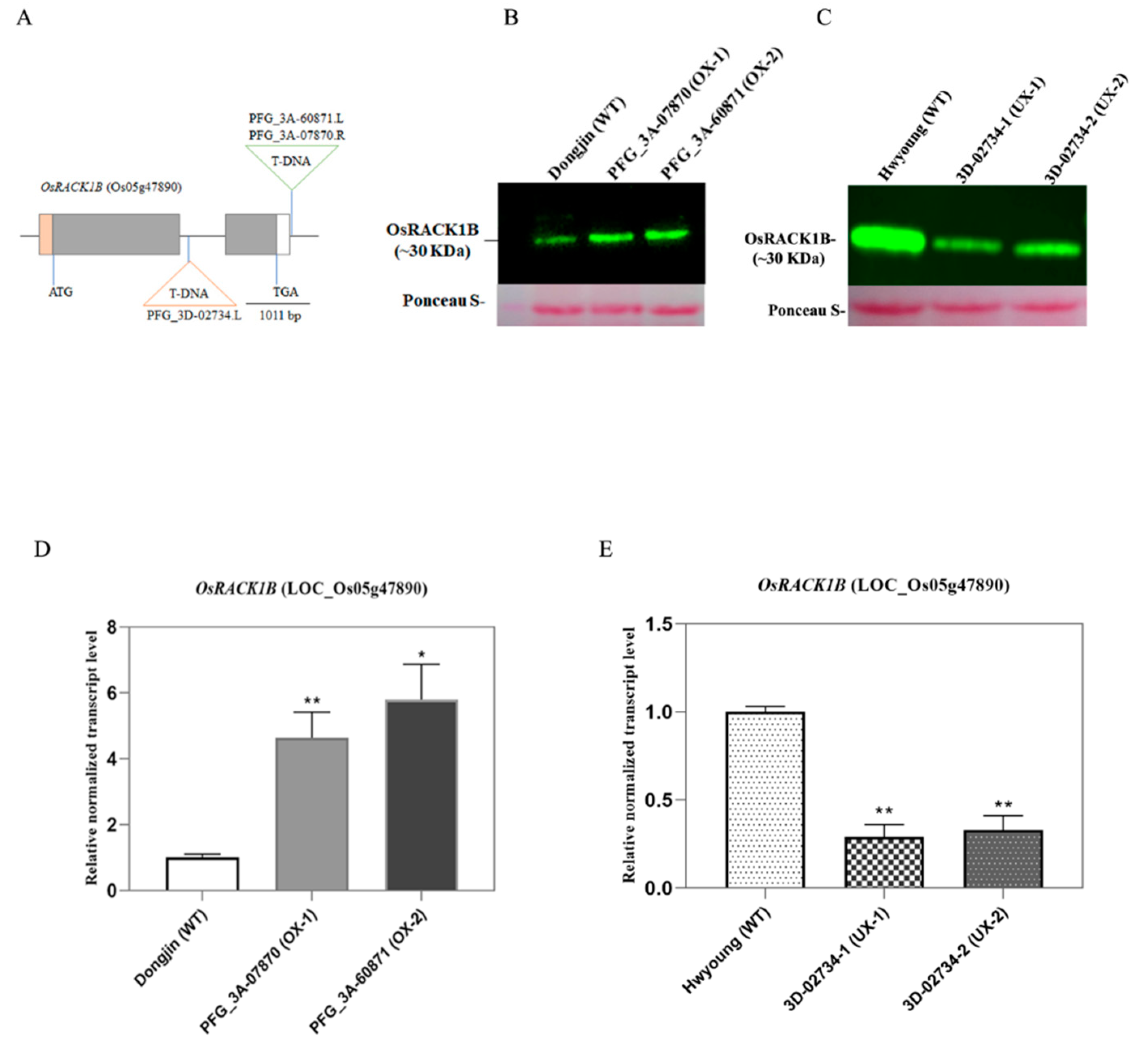
Figure 2.
The OsRACK1B overexpressed rice leaves retains more chlorophyll than wild-type during salinity stress. A. The stay-green phenotype was observed in OX-1, and OX-2 leaf discs after 3 days of salt DST (200mM NaCl) treatment in comparison to the WT leaf discs. (A and C) Changes in green pigment (measured as pixel density of same area by the ImageJ) in leaf discs from eight-week-old OsRACK1B overexpressed rice plants after 3 days of salt treatment (3 DST) and 3 days of water treatment (Mock) as control. B and D, The OsRACK1B down-regulated rice leaf discs exhibit yellowing (premature senescence) phenotype than wild-type during salinity stress. B. Expedite chlorotic phenotype was observed in leaf discs from eight-week-old OsRACK1B down-regulated UX-1 and UX-2 leaf discs after 3 days of salt (200mM NaCl) treatment in comparison to the WT. D, Green pigment changes (measured as pixel density of the same area by the ImageJ) in leaves of UX-1 and UX-2 plants after 3 days of salt treatment (3 DST) and 3 days of water treatment (Mock) as control.
Figure 2.
The OsRACK1B overexpressed rice leaves retains more chlorophyll than wild-type during salinity stress. A. The stay-green phenotype was observed in OX-1, and OX-2 leaf discs after 3 days of salt DST (200mM NaCl) treatment in comparison to the WT leaf discs. (A and C) Changes in green pigment (measured as pixel density of same area by the ImageJ) in leaf discs from eight-week-old OsRACK1B overexpressed rice plants after 3 days of salt treatment (3 DST) and 3 days of water treatment (Mock) as control. B and D, The OsRACK1B down-regulated rice leaf discs exhibit yellowing (premature senescence) phenotype than wild-type during salinity stress. B. Expedite chlorotic phenotype was observed in leaf discs from eight-week-old OsRACK1B down-regulated UX-1 and UX-2 leaf discs after 3 days of salt (200mM NaCl) treatment in comparison to the WT. D, Green pigment changes (measured as pixel density of the same area by the ImageJ) in leaves of UX-1 and UX-2 plants after 3 days of salt treatment (3 DST) and 3 days of water treatment (Mock) as control.
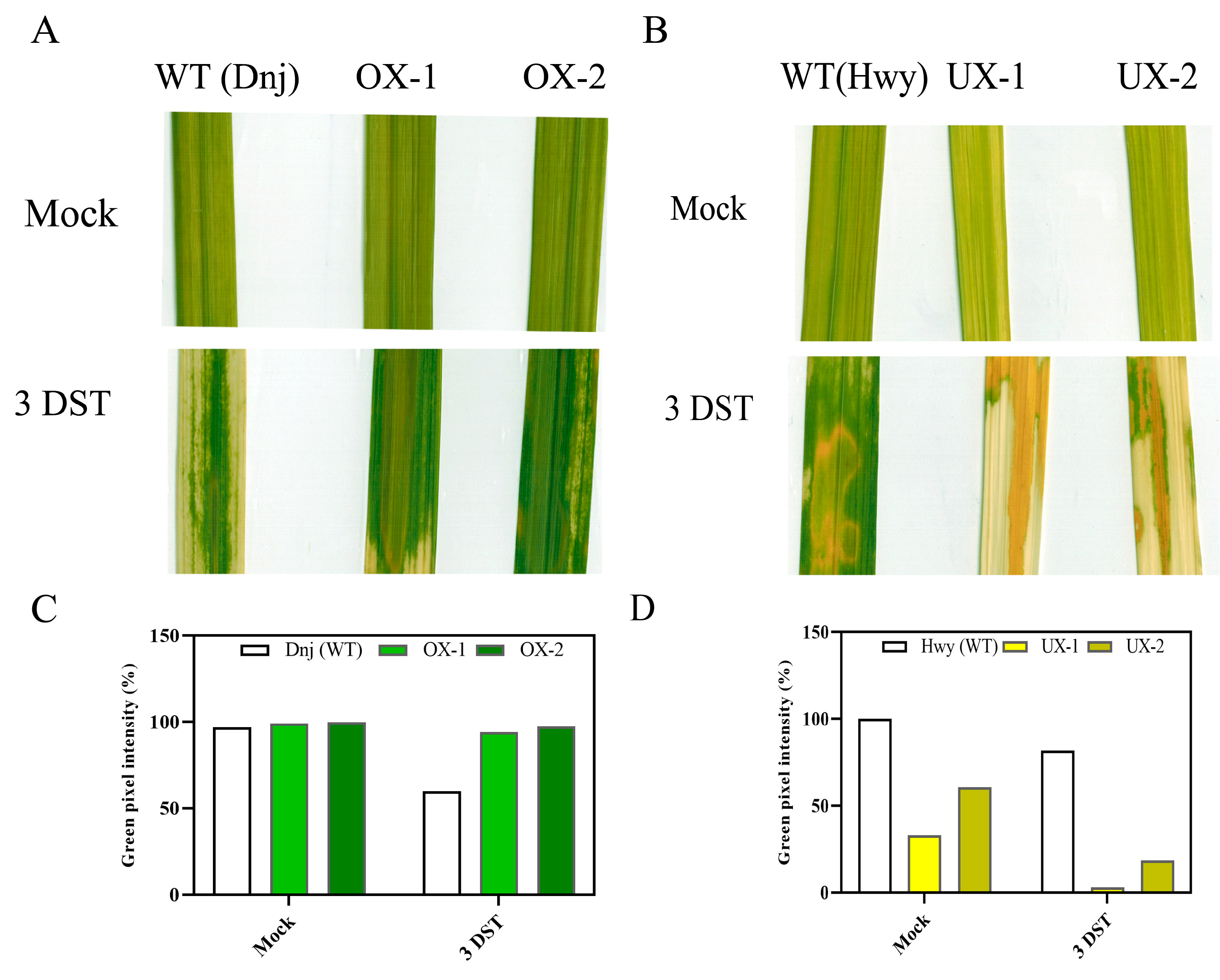
Figure 3.
Altered expression of chlorophyll metabolic genes under salinity stress condition. Total mRNA was extracted from leaf tissue from eight-week-old OsRACK1B overexpressed (OX-1 and OX-2), OsRACK1B down-regulated (UX-1) plant and their respective wild-type (WT) plants ( Dongjin and Hwayoung) at 0 hr and after 24 hr of salt (200 mM NaCl) treatment. The transcript levels of chlorophyll biosynthetic gene chlorophyllide a oxygenase (CAO) and catabolic enzymes (CCEs) encoding genes: red chlorophyll catabolite reductase (RCCR) and NONYELLOW COLORING 1 (NYC1); senescence-associated transcription factor (TF) NAC092, chlorophyll degradation related gene STAY-GREEN (SGR); and LHCII phosphorylation and state transitions related gene Serine/threonine-protein kinase (STN7) were analyzed using qRT-PCR. The transcript levels of all genes were normalized using OsActin-1 (LOC4333919). Error bars indicate the standard errors (SE) of the means (n = 3) and values are means ±SE. Single and double asterisks indicate statistical significance: *P < 0.05 and **P< 0.01 compared to the wild-type (Student’s t-test). Primers are listed in Table S2.
Figure 3.
Altered expression of chlorophyll metabolic genes under salinity stress condition. Total mRNA was extracted from leaf tissue from eight-week-old OsRACK1B overexpressed (OX-1 and OX-2), OsRACK1B down-regulated (UX-1) plant and their respective wild-type (WT) plants ( Dongjin and Hwayoung) at 0 hr and after 24 hr of salt (200 mM NaCl) treatment. The transcript levels of chlorophyll biosynthetic gene chlorophyllide a oxygenase (CAO) and catabolic enzymes (CCEs) encoding genes: red chlorophyll catabolite reductase (RCCR) and NONYELLOW COLORING 1 (NYC1); senescence-associated transcription factor (TF) NAC092, chlorophyll degradation related gene STAY-GREEN (SGR); and LHCII phosphorylation and state transitions related gene Serine/threonine-protein kinase (STN7) were analyzed using qRT-PCR. The transcript levels of all genes were normalized using OsActin-1 (LOC4333919). Error bars indicate the standard errors (SE) of the means (n = 3) and values are means ±SE. Single and double asterisks indicate statistical significance: *P < 0.05 and **P< 0.01 compared to the wild-type (Student’s t-test). Primers are listed in Table S2.
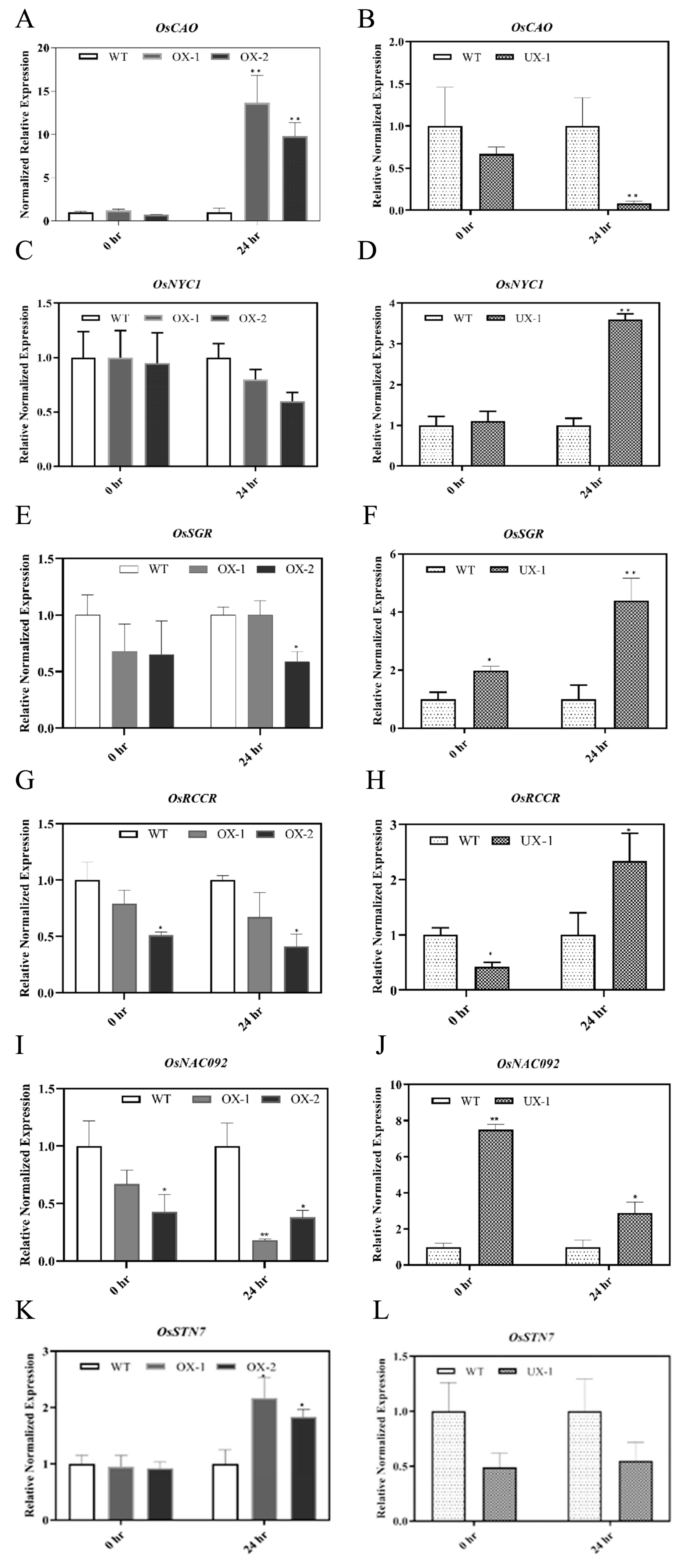
Figure 4.
Immunoblot analysis of Photosynthetic Proteins. The altered level of OsRACK1B protein causes a change in the abundance of Lhcb1 and Stay-Green protein (SGR1) during salinity induced leaf senescence. Total protein was extracted from harvested leaves from eight-week-old transgenic rice plants and their respective WT plants at 0 hr and 24 hr of salt (200mM NaCl) treatment. Anti-Lhcb1 and anti-Sgr antibodies were used for immunoblot analysis. A and B, Immunodetection of SGR1. C and D, Immunodetection of Lhcb1. Antisera against Lhcb1 recognize two bands in UX sample. Upper band (slower migrating band) represents the phosphorylated form. P and U indicate the phosphorylated and unphosphorylated forms, respectively. Each lane contained equal amount of total protein from leaf tissue. .
Figure 4.
Immunoblot analysis of Photosynthetic Proteins. The altered level of OsRACK1B protein causes a change in the abundance of Lhcb1 and Stay-Green protein (SGR1) during salinity induced leaf senescence. Total protein was extracted from harvested leaves from eight-week-old transgenic rice plants and their respective WT plants at 0 hr and 24 hr of salt (200mM NaCl) treatment. Anti-Lhcb1 and anti-Sgr antibodies were used for immunoblot analysis. A and B, Immunodetection of SGR1. C and D, Immunodetection of Lhcb1. Antisera against Lhcb1 recognize two bands in UX sample. Upper band (slower migrating band) represents the phosphorylated form. P and U indicate the phosphorylated and unphosphorylated forms, respectively. Each lane contained equal amount of total protein from leaf tissue. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



