Submitted:
25 May 2023
Posted:
26 May 2023
You are already at the latest version
Abstract
Endophytic microbes are a ubiquitous group of plant-associated communities that colonize the intercellular or intracellular host tissues while giving numerous beneficial effects to the plants. All the plant species are thought to be associated with endophytes, majorly constituted with bacteria and fungi. During the last two decades, there has been a considerable movement toward the study of endophytes associated with coffee plants. In this review, the main consideration is given to address the coffee-associated endophytic bacteria and fungi, particularly their action on plant growth promotion and the biocontrol of pests. In addition, we sought to identify and analyze the gaps in available research. Additionally, the potential of endophytes to improve the quality of coffee seeds is briefly discussed. Even though there are limited studies on the subject, it has been well recognized the potentiality of coffee endophytes in plant growth promotion through enhancing nitrogen fixation, availability of minerals, nutrient absorption, secretion of phytohormones, and other bioactive metabolites. Further, the antagonistic effect against various coffee pathogenic bacteria, fungi, nematodes, and also insect pests lead to the protection of the crop. Furthermore, it is recognized that endophytes enhance the sensory characteristics of coffee as a new field of study.
Keywords:
Ascomycetes
; bio decaffeination
; endophytic bacteria
; endophytic fungi
; entomopathogenic endophytes
; nitrogen fixing
; phosphate solubilizing
; phytohormones
; plant defense system
1. Introduction
1.1. Coffee as One of the Most Appealing Beverages
The genus coffee belongs to the Rubiaceae family. There are about 103 coffee species available, though, Arabica (Coffea arabica) and Robusta (Coffea canephora) are the two most dominant coffee species that grow worldwide and also provide about 99% global market [1,2]. Coffee is also recognized as the second most traded commodity worldwide after oil. According to the International Coffee Organization [3] world coffee exports amounted to 10.88 million (60-kilo) bags in December 2022, while exports in the first 3 months of the coffee year 2022/23 (October/22 to December/22) was 30.27 million (60-kilo) bags. The market of coffee is also rapidly expanding worldwide, for example, in China, coffee consumption has grown steadily by 20% each year, particularly in urban areas [4].
Moreover, as of today, the coffee plant is grown in over 60 countries by more than 25 million farmers, particularly small-scale growers in Africa, Asia, and the Americas [1]. In fact, those small-scale farmers dominate the market, 95% of coffee farmlands are 5 ha or smaller, and 84% are under 2 ha. The remaining 5% are produced by larger estates, and coffee plantations larger than 50 ha are rare in outside of Central and South America [5]. For instance, in Ethiopia, coffee is the top export, making between 20–25% of all foreign exchange earnings. Moreover, around 15 million people are thought to depend on coffee for their livelihood; and more than 80% of coffee growers are peasant farmers [6].
1.2. Concerns of Coffee Harvest Reduction Factors and Preventive Strategies
Despite the fact that coffee has a significant economic impact on low-income countries throughout the world, the sector has been faced with numerous persistent stresses and unexpected shocks over the decades challenging productivity. These common issues include climatic changes (e,g., temperature, and precipitation), diseases, and pests [7,8]. Addressing the issues of diseases, coffee rust caused by the fungus Hemileia vastatrix is among the leading troublesome organism, and the first known outbreak was documented in Sri Lanka in 1869. The thunderdome of the epidemic leads Sri Lanka to replace their coffee lands with tea and other alternative crops. Subsequently, the disease has affected nearly all regions of the world where coffee is grown [9]. In extreme outbreaks, foliage can be lost up to 100% and berries can be lost up to 70%, and the disease can result in yield losses of more than 75% [10]. Yet, the disease is not eradicated and is responsible for causing losses of one to two billion US dollars annually [11]. In addition, a lack of awareness and surveillance about the rust disease (also others) could be a future alarming threat to emerging coffee-grown countries like China [12]. Other prominent diseases include coffee berry disease caused by Colletotrichum kahawae [13], coffee wilt disease caused by Fusarium xylarioides [14], brown eye spot caused by Cercospora coffeicola [15], and Armillaria Root Rot caused by Armillaria mellea [16]. In turn, coffee berry borer Hypothenemus hampei [17], coffee leaf miner Leucoptera coffeella [18], and black coffee twig borer Xylosandrus compactus [19] are among the major insect pest of coffee. Despite those, pant-parasitic nematodes also severely hamper coffee plantations in many parts of the world [20]. The management of those disease and pest issues is generally achieved by synthetic pesticides, however, such practices are deemed to be negligible by many communities. At the same time, other available traditional techniques such as shade management, bark scrubbing, stem wrapping to prevent oviposition, mass trapping with the aid of pheromone-baited traps, and uprooting and burning of affected plants are not supposed to control the pest trouble fully [21,22].
1.3. Endophytes as a Prime Source for Improving Plant Health
Recently, endophytes have been recognized as prime candidates for protecting crops from both abiotic and biotic stresses, thus becoming a hot-topic [23,24]. Simply, endophytes can be described as any organism that lives within plant tissues. Interestingly, throughout their life cycles, endophytic communities are closely related to their hosts or spend portions of their lives in host tissues also without causing disease or negatively affecting them [25]. Thus, in general, endophytic association shows a mutualistic relationship, with some exceptions [26].
Theoretically, it is accepted that almost all plants are infested with endophytes, at least one or more of them [27]. A plethora of studies have been conducted on many different forms of endophytic populations, including bacteria and fungi regarding their various potential benefits to the plants. Those include producing growth-promoting metabolites, insect and pest repellents, antimicrobials to fight plant pathogens, and protect plants in stressful environments [27–29]. The majority of the studies showed that endophytes isolated from the same plants may have the ability to promote plant growth. However, several research studies have documented how endophytic bacteria can encourage the growth of non-host plants [30].
Other than coffee, studies have found that these endophyte microorganisms harbor many other crop plants including chickpea, chili, citrus, cotton, cowpea, maize, mustard, pearl millet, rice, soybean, strawberry, sugarcane, sunflower, tomato, and wheat [31–33]. Endophytes can also be found in below or above-ground parts of these plants, e.g., roots, tubers, fruits, leaves, barks, twigs, petioles, flowers, nodules ovules, seeds, and stems [34,35].
1.4. Aim of the Current Article
In the above context, we reviewed the studies of endophytic bacteria and fungi associated with coffee plants. The main purpose of this paper is three-fold; 1). emphasize the endophytes that possibly influence the coffee plant growth promotions, 2). report the potentiality of coffee endophytes in protecting coffee plants against various plant pathogens and insect pests, and 3). understand the scenario on the available studies, gaps, and future directions. In addition to those major considerations, we briefly discussed the ability of endophytes in improving the quality and flavor of coffee seeds.
2. Data Collection and Analysis
A thorough literature search was conducted by using three platforms, namely National Center for Biotechnology Information (NCBI), Google Scholar databases, and ProQuest. The search terminology was, ‘Coffee Endophytes’, and the initial search resulted in a total of 6,456 findings. However, the majority of results were not directly relevant to our objectives, thus eliminated after the primary screening. Thereafter, extensive secondary screening was done to filter the relevant publication. The final analysis was based on 107 studies published in 50 research papers. The data extraction pipeline is shown in Figure 1.
3. General Aspects of Endophytes
3.1. Endophytic Association with Plants
Endophytes are said to be universally distributed within the plant kingdom, and their presence in land, freshwater, and marine plants has been recorded [36,37]. Nonetheless, they can be latent pathogens, mutualists, commensals, temporary residents, or latent saprotrophs [38]. The majority of plant endophytes are found to be bacteria and fungi, while archaebacteria, algae, protozoa, viruses, and nematodes are rare endophytes [39]. According to modern molecular-based techniques and fossil records, the co-evaluation of endophytes has been reported to be from about 460 million years along with the plants [40,41]. This initial association is probably with the arbuscular mycorrhizal fungi, based on the sequence analyses it was further revealed that the development of those endophytes was along with the land plant colonization [42]. According to the preservations in the Rhynie chert, this association runs to the period of Early Devonian, where the plant lacks a ‘true’ root system [43]. The absence of well-preserved leaf endophyte fossils prompts the hypothesis that they likely originated later, during the Early Cretaceous period, concurrent with the advent and diversification of angiosperms. In contrast, studies show that fungus leaf endophytes existed at least during the Carboniferous period [44]. A study of leaf endophytes in the coffee family (Rubiaceae) done by Verstraete et al. [45] reviled that, plant lineages with endophytes show a higher level of speciation, which is also significant compared with the plant lineages without endophytes, however, there is a small disparity in the extinction rate. Furthermore, plants evolve twice as quickly toward endophytes than they do away from them, proving that the interaction may be advantageous to the plant [45].
It is relatively unusual for a plant to be colonized by just one kind of endophyte, instead, a number of endophytes habited with the plant also probable with directly or indirectly interacting with each other, e.g., bacteria-bacteria, fungi-fungi, and bacteria-fungi [46]. Thus, many millions of species of endophytes are thought to be interacting with the plants [47]. Despite the fact that bacteria and fungi are completely different forms of life as they are prokaryotes and eukaryotes respectively, they share similar characteristics as endophytes, e.g., the lifestyles of bacteria and fungi include intra- or intercellular colonization of plant tissues [48,49]. Furthermore, based on their life strategies, endophytic communities that may be present in the host plant, have either obligate or facultative associations. The obligatory endophytes can grow and survive only on the host plant; transmission through vertically (seeds) or vectors (pollinators, arthropods, or sap-feeders), rather than emanating from the rhizosphere. Unlike obligate endophytes, facultative endophytes do not show the absolute necessity of host plant, they have a stage in their life cycles where they exist independently of their hosts and they are generally associated with soil environment and atmosphere that is in close proximity with plant and colonize in the plant once the opportunity creates via coordinated infection [50,51]. In order to create an association with the plant, for example, soil-dwelling endophytes should first colonize the rhizosphere. Plant roots secrete exudates that contain various components, mainly including sugars, amino acids, organic acids, vitamins, and high molecular weight polymers thus providing various nutritional resources to the microbes. Those exudates are concentrated in the plant rhizosphere, thereby many microbes including endophytes attract towards (chemotaxis) and colonize [52]. Nevertheless, it is not an easy task to colonize in the rhizosphere as endophytes need to fight for space and nutrients along with other wealthy microbiota, which perhaps seems even to be antagonistic against endophytic colonization [30]. In turn, endophytes may outcompete the other microbes by growing aggressively and taking space more rapidly, however, little is understood about how they interact with other microorganisms [53,54]. The endophytes which thrive in this microenvironment could colonize further and reach the root system of the plants. In the case of bacteria, their traits including motility and the synthesis of polysaccharides are crucial for the colonization of plant rhizospheres [30,55]. Further, bacterial populations use long strings of closely associated cells to get the contact with the root system [56]. More precisely, endophytes get into the plant by using two different mechanisms namely, active and passive. In active colonization, endophytes get into plant tissue by searching and through cracks (e.g., open sites at lateral roots emergence, areas of elongation and differentiation of the root), wounds (e.g., insect bites), or hydathodes. Moreover, in active mechanism, they penetrate by degrading plant tissues with the help of hydrolytic enzymes such as cellulase and pectinase. The colonization of the plant by passive endophytes occurs as a result of random circumstances, rather than on purpose. Due to the absence of the cellular machinery needed for plant colonization, the passive life strategy may make them less competitive [51,57]. Note that endophytes that are transmitted via seeds ensure their presence in young plants. Similarly, the procedure described above is not necessary for plants that reproduce vegetatively [34].
It is possible for bacterial endophytes to colonize a plant host systematically even after a single entry. Further, like in rhizosphere colonization, bacterial endophytes use specific traits known as colonization traits to be done the entire plant colonization including communication between the bacterial endophytes and the plant [58]. In one study, Huang et al. [59] observed that endophytic colonization also leads the physiological changes in the plant, and according to their study, Bacillus subtilis invasion started from the root hair by swelling and bending of root hair tips also caused dichotomous branching of the root hair without bending. Turn back to the fungi, when the spores are attached to the host surface they start to germinate, by slight swelling, followed by germ tube emergence, development of aspersoria, penetration peg emergence, hyphae development, and invasion of plant tissues [60]. Noteworthy that, a comparatively small fraction of endophytes is colonizing the above-ground part of the plant due to the restriction for nutrients acquisition, ultraviolet light (UV), and desiccation, thus the greatest number of endophytes associated with the roots [61,62]. Unlike fungi, bacterial endophytes favor the pathway to colonize via the xylem vascular system and therefore can find systemic colonization of internal plant compartments. It has been demonstrated that bacteria can colonize xylem vessels, and the perforation plates' holes are big enough to let bacteria pass through [63]. Nevertheless, all the endophytes must endure the plant defense mechanism prior to colonization. Plant detects microbial invaders through microbe-associated molecular patterns (MAMPs), and through a series of defense responses, they attempt to overcome the threat [63]. However, endophytes have the ability to counteract the stimulation of the plant defense response, either synthesizing MAMPs with poor elicit capacity or downregulating their expression. Further, they can even synthesize the required genes to minimize the stimulation of plant defense reactions [64]. Production of reactive oxygen species (superoxide, hydroperoxyl radical, hydrogen peroxide, hydroxyl radical species), nitric oxide, and phytoalexins are one of such examples of plant defense reactions. In order to overcome this oxidative stress, for instance, Enterobacter sp. encodes superoxide dismutases, catalases, hydroperoxide reductases, chloroperoxidase, and thiol peroxidases [65]. It is also recognized that pre-communication between the endophytes and the plant is needed, which depends on the mutual signal exchange between them and also leads to balanced antagonisms [66]. Moreover, in general owing to the selective pressure driven by the plant, the colonization of endophytes is organ- and tissue-specific [67].
3.2. Brief Classification and Important Genera/Phylogenetic Placement
Endophytic bacteria fall within 16 phyla with more than 200 genera, nevertheless, the majority of them belong to the three phyla namely, Actinobacteria, Firmicutes, and Proteobacteria [68]. Endophytic bacteria are considered a subclass of rhizospheric bacteria, which is also known as plant growth-promoting rhizobacteria [30]. They also could be either Gram-negative or Gram-positive [68]. While, endophytic fungi predominantly categorize under the Ascomycetes, appearing sporadically in Basidiomycetes and Zygomycetes [29]. More precisely, endophytic fungi are classified into two main groups, 1). clavicipitaceous, which infects some grasses thus having a narrow host range; and 2). nonclavicipitaceous which infects vascular and non-vascular plants. All over, those species belonging to those two groups are classified into classes 1–4. Class 1 is categorized under the clavicipitaceous and other classes are placed under nonclavicipitaceous. Class 1 fungi (Ascomycota) colonize in plant’s shoots and rhizomes and class 2 (in Dikaryomycota-Ascomycota in major and minor in Basidiomycota). Most belong to the Ascomycota, with a minority of Basidiomycota. Endophytes colonize in shoots, roots, and rhizomes. Class 3 (Major in Dikaryomycota- Ascomycota where especially concentrated or, Basidiomycota) and class 4 (primarily, Ascomycota) colonize in shoot and root, respectively. Except for class 3, fungi in other classes show extensive colonization in plants [69]. In a recent literature review by Lugtenberg et al. [70] gave suggestions to update this classification as it hides the group of entomopathogenic fungi, which are symptomless endophytes of plants.
4. Coffee Endophytes: Insight
4.1. History of Studies, Current Trends, and Gaps
The understanding of endophytes goes back to more than a century, where in 1866 de Bary introduced the term endophytes for the first time [71]. Later on, discussions were made by further isolation of endophytes from various plant species. In the middle 1930s, Sampson’s [72] and early 1940s, Neill’s [73] studies showed the coexistence of endophyte fungi with, Lolium temulentum and L. perenne. Until the 1960s only a few studies were conducted, thereafter multiple studies have been conducted by addressing various dimensions of the endophytes [69]. Understanding the endophytes associated with different plant species also widens [74,75], while the most studied fungal endophytes are the grass associated with the genus Neotyphodium (Clavicipitaceae) [76]. However, according to the literature available, studies on endophytes associated with coffee plants emerged in the later 1990s, where Jimenez-Salgado et al. [77] demonstrated the bacterial endophyte Acetobacter diazotrophicus associated with coffee. Later on, the next study conducted by Sakiyama et al. [57] isolated several species of Paenibacillus. Initial studies focused more on bacterial endophytes, however, fungi endophytes are the most abundant group in the coffee plant rather than the bacteria, and vice versa in coffee-associated soil [78]. According to the literature survey, it is consumable that studies on coffee fungi have sprouted since 2005, and continuously gain more attention than bacterial studies. As shown in Figure 2, several studies only focused on both bacterial and fungi endophytes.
Furthermore, just above the half of studies (54.21%) concentrated on isolating and identifying the local coffee endophytes. The next most studies (13.08%) have been carried out to recognize the antagonistic activity of endophytes towards controlling phytopathogens and other pests. A handful of studies (10.28%) are also conducted to understand the in-depth diversity/population of local endophytic microflora. Despite that, several studies revealed the ability of endophytes in improving the quality of coffee seeds (4.67%), coffee plant growth promotion (3.74%), inoculating/introduction to the coffee cultivations (3.74%), and colonization (2.80%). Figure 3 shows the overview of the studies carried out.
It is worthwhile to bring some of the study findings on the isolation and identification of coffee endophytes. From 32 isolations of leaf endophytes, Bongiorno et al. [79] observed Colletotrichum (71.9%) as the major group, followed by Trichoderma (9.4%), Cercospora (6.3%) and rest (3.1% each) under the Ophiognomonia, Cladosporium, Mycosphaerella, and Schizophyllum. In a comprehensive study, Vega et al. [80] isolated 843 fungal endophytes associated with different coffee plant parts including leaves, roots, stems, and berries, abundant taxa with Colletotrichum, Fusarium, Penicillium, and Xylariaceae. Saucedo-García et al. [81] also found similar results, as Colletotrichum and Xylaria were the most associated endophytes (from 31 morphospecies), nevertheless, even with the coffee leaves. In a similar line, Oliveira et al. [82] identified 17 endophytic fungi species in 16 genera associated with coffee leaves. However, in a most recent molecular-based study, Gagliardi et al. [83] brought up the fact that the fungi endophytes alone are far more than we think, of course, it is obvious, nevertheless, as a proven study they have come up with 1,664 fungal amplicon sequence variants (ASVs) in coffee root tips alone. Table 1 shows some of the endophytes identified associated with coffee.
4.2. Endophytes in Coffee Plant Growth Promotion
Plant growth promotion due to the endophytes has been widely recognized for various plants including maize [97], potato [98], rice [99], sweet potato [100], tea [101], tomato [102], and so on [103]. It is interesting to know how those coffee endophytes do improve plant growth. The most frequent ways that endophytic associations benefit the plants' growth include direct effects; nutrient absorption promotion, enhancing the availability of minerals, nitrogen fixation, secretion of phytohormones, and other bioactive metabolites, indirectly via stress reduction [34,104,105]. The possible pathway of growth promotion in coffee plants is shown in the Figure 4.
4.2.1. Nutrient Absorption Promotion and Enhancing the Availability of Minerals
It has been recognized that endophytic fungi have considerable capacity to improve the nutrient up-taking ability of plants from soil both macro- (e.g., calcium, magnesium, nitrogen, phosphorus, potassium) and micro- elements (e.g., boron, zinc) [106,107].
One of the foremost studies conducted by Jimenez-Salgado et al. [77] identified the nitrogen (N) fixing Acetobacter diazotrophicus bacteria associated, particularly from the coffee root systems. It has been suggested that increasing populations of endophytic N-fixing bacteria in plants may boost nitrogen fixation, as reported by Jimenez-Salgado and co-authors [77] those recorded bacteria have the ability to colonize extensively, thus increasing values for the plant. In addition, the bacteria found to be in coffee root tissues, Jimenez-Salgado et al. [77] found that they also largely habitat in the rhizosphere. This could be due to the falling of coffee leaves and berries collected in soil lines, degraded by the microbiome, converted further to organic matter, and enriched the rhizosphere with carbon (sugar) that allows utilization by the A. diazotrophicus for their development. By taking into account this phenomenon, it is clear that most probability N fixing bacteria are associated with coffee with mutualistic symbiosis, however, no evidence was found during our literature survey. Looking back, Pratiwi et al. [108] isolated several more N-fixing bacteria from coffee roots, among them Rahnella aquatilis and Pseudomonas tolaasii as the most prominent in fixing N, thus in the ammonium producing. Note that endophytic bacteria modify the molecular nitrogen, then convert into ammonium, which is then used by host plants for their growth [109]. In turn, Further studies done by Pratiwi and colleagues [108] recognized that the phosphate (P) solubilizing potentiality of several bacterial species, Rahnella aquatilis, and Kluyvera intermedia had the highest and most significant P solubilizing capacity. The study findings of them are very competitive with the results obtained by Muleta et al. [110], where they worked on coffee root-associated soil rhizobacterial species, which have P solubilizing capacity. In a recent supportive study, Teshome et al. [111] recognized that Pseudomonas spp. (14.5%) are the most prominent P solubilizing in their isolations, followed by Citrobacter spp. (3.6%)., Rhodococcus spp. (9.1%)., Stenotrophomonas spp. (7.3%)., Gordonia and Bacillus spp. (3.6%). Added to that, Pratiwi et al. [108] recognized that the P solubilizing potentiality of several bacterial species, Rahnella aquatilis, and Kluyvera intermedia had the highest phosphate solubilizing capacity.
4.2.2. Secretion of Phytohormones and Other Bioactive Metabolites
It has also been recognized that endophytes synthesize phytohormones (or maybe like compounds), particularly abscisic acid, cytokinin, gibberellins (GAs), and indole-3-acetic acid (IAA), which can promote plant growth development and also aid plant hosts in fending off the negative effects of abiotic stressors [68,112]. In a study, Silva et al. [113] found that bacterial strains of Acinetobacter calcoaceticus, Escherichia fergusonii, and Salmonella enterica have the ability to synthesize aforesaid IAA, cytokinins, gibberellins, and also siderophores, thus directly enhance the coffee plant growth. Highlighting one such phytohormonal effect, endophytic changes the plant toot architecture by producing plant growth regulators such as IAA, where it affects the roots physiology and function in a positive direction via enhancing its development (also increasing root surface area), leading to better absorption of nutrients and water [106]. While, small molecules known as siderophores are capable of chelating iron, and can be used by plants while depriving pathogens of iron. Further, siderophores aid in fixing nitrogen since diazotrophic organisms need ferrous ion (Fe2+) and molybdenum (Mo) factors for their functioning and synthesis of nitrogenase, eventually beneficial to the iron-deficient plant [32]. Recent improvements with technologies, it has been discovered many secondary metabolites secreted by endophytes and their functions [70]. Those molecules not only resistant plants against pests but also protect plants from abiotic stresses to some extent such as drought, flooding, and heat. However, this was not on the agenda relevant to the coffee endophytes particularly how those protect the plants from abiotic challenges; thus we did not consider those in this paper.
4.2.3. Successful Field and Laboratory Experiments on Coffee Plant Growth Promotion.
There are only limited studies conducted on coffee plant growth improvement through endophytes. For instance, Asyiah et al. [114] found that Bacillus sp. has the ability to promote plant growth, of this, they have found that Coffea canephora grown in pots along with the bacterial consortium increased plant height by an average of 5.2 cm compared with the untreated control. Furthermore, it was observed that the Bacillus sp. treatment alone increases the number and area of coffee leaves, e.g., 10.7 ± 2.66 (control: 4.4 ± 2.30) and 416.3 ± 224.9 cm2 (control: 109.9± 81.5 cm2) respectively. Added to this, Mamani-Huayhua et al. [95] evident that Trichoderma sp. has a great ability to promote plant growth. They sprayed conidial suspension of 1x107 cfu mL-1 on soil and foliage at the transplanting of the seedlings, also 30 days intervals after the transplant. They tested the growth parameters of coffee plants after three months of transplanting and, as shown in Figure 5 received considerable evidence for growth enhanced by the means of increases in plant height, stem diameter, length of the main root, and number of leaves compared with untreated control. In an in vitro assay, Silva et al. [112] found that out of 119 bacterial strains, six of them namely, 85G (Escherichia fergusonii), 161G, 163G, 160G, 150G (Acinetobacter calcoaceticus), and 109G (Salmonella enterica) shown significant impact on promoting the growth of coffee seedlings. Further, they have shown that the growth-promoting index for those species is higher as 574, 560, 549, 545, 524, and 522, respectively. In contrast, this study is vital, as they observed, that not all bacterial strains could promote plant growth. Also, some reduce the growth by giving a deleterious effect. However, overall, those findings are clear enough to understand endophytic capacity as growth promoters may be competitive as rhizobacteria.
4.3. Endophytes in Coffee Plant Protection
Endophyte shows the potential of protecting plants from a wide range of pathogens and chewing, boring, and sucking insects and nematodes, thus helping them to survive threats [115]. It is intriguing to know first how the endophytes play a role in controlling the phytopathogens.
4.3.1. Protection against Fungi Pathogens
It is also interesting to understand the biocontrol mechanism of pathogenic diseases by endophytic bacteria. Secondary metabolites (e.g., harmine), extracellular lytic enzymes (e.g., chitinase, β-1, 3-glucanase, protease, lipase, gelatinases), volatile compounds (e.g., Hydrogen cyanide), and siderophores were discovered to have synergistic effects in inhibiting pathogenic fungus [84,116,117].
Accounting for the endophytic fungi, they inhibit the phytopathogenic fungi using different mechanisms. Those strategies include mycoparasitism, competition, antibiosis, and induction of the plant defense system [118]. In mycoparasitism, the endophytic fungi directly prey on the phytopathogenic fungi. Here, first, the endophytic fungi recognize the praying fungi through the host-based signals and grow towards them, attached, coiling around the prey hyphae, penetrating inside the host, and acquiring the nutrients. This led to the deaths of praying fungi and ultimately controlled the phytopathogenic fungi population [119]. The competition basically takes place among the fungi species for ecological niches and nutrients. Endophytic fungi could grow faster than pathogenic fungi and take advantage of space and nutrients, thus minimizing the colonization of the opponent [54,120]. In antibiosis, to control the pathogens, endophytic fungi use different types of antimicrobial compounds such as aliphatic compounds, alkaloids, peptides, phenylpropanoids, polyketides, and terpenoids [121]. In another way, indirect control of pathogens takes place by inducing the plant defense mechanism. In this regard, along with the association with the plant host, endophytic trigger the induced defense mechanism, by altering the phytohormone biosynthesis, and production of defense-related enzymes and secondary metabolites [122,123,124].
Silva et al. [112] found that Bacillus megaterium, Brevibacillus choshinensis, Cedecea davisae, Microbacterium testaceum, Pectobacterium carotovorum, and Salmonella enterica significantly reduces the coffee leaf rust, Hemileia vastatrix. Of this, Silva and co-authors [112] recognized that particularly Brevibacillus choshinensis shows higher disease-controlling ability where the disease severity index is reduced by 100. Further, it also shows that endophytic bacteria affect on germination of urediniospores. In a similar study, Shiomi et al. [85] did a comprehensive study on controlling Hemileia vastatrix. First, they isolated the endophytic bacteria from both Coffea arabica and Coffea robusta. They have found that out of 40 selected bacteria isolations, 23 isolations (e.g., Acinetobacter calcoaceticus, Bacillus cereus, Bacillus lentimorbus and Clavibacter michiganensis) have the capacity to inhibit H. vastatrix urediniospore germination in more than 40%. A further compulsive observation was that germination tubes had deformations that impeded normal growth. Furthermore, through the leaf disc assay, Shiomi and co-workers [85] recognized that the endophytic bacteria also have the ability to reduce the disease severity of the coffee leaves caused by Hemileia vastatrix, tremendously. In this aspect, Bacillus lentimorbus was found to be the most effective, control levels were above 63%. In contrast, Cacefo et al. [125] observed that H. vastatrix could also be controlled with Bacillus subtilis, however, not as effective as fungicide application where once fungicide showed 53% of disease control, the bacterial treatment could control by 24%. According to the Kejela et al [117] study, it is clear that Bacillus sp. has the ability to control other coffee pathogens such as Colletotrichum gloeosporioides and Fusarium oxysporum. Even though their study tested under laboratory conditions, the findings are significant, colonies of C. gloeosporioides and F. oxysporum are inhibited by 78% and 86% respectively by Bacillus sp. Looking at controlling Fusarium oxysporum, Duong et al. [84] recognized that Burkholderia sp. (closely related to B. cenocepacia) shows a higher effect than Bacillus sp. (closely related to B. subtilis), where fungi growth is inhibited by 49.77 ± 0.08% and 40.77 ± 0.06% respectively. In addition, they found that Streptomyces sp. (closely related to S. mobaraensis) exhibits pathogen controlling ability similar (40.76 ± 0.04%) to Bacillus sp. However, a comparatively lower level of Fusarium oxysporum inhibition was reported when tested with other genes including Brachybacterium sp. (10.26 ± 0.09), Burkholderia sp. (26.99 ± 0.01%), Curtobacterium sp. (17.68 ± 0.06%), Kitasatospora sp. (9.00 ± 0.04%), Luteibacter sp. (8.62 ± 0.02%), and Pseudomonas sp. (22.18 ± 0.04%).
Coming back to the disease control of coffee leaf rust, endophytic fungi have also proven to be effective as endophytic bacteria. Of this, Mamani-Huayhua et al. [95] presented their findings on controlling Hemileia vastatrix using five endophytic Trichoderma strains (TE1, TE2, TE3, TE4, TE5). They have sprayed spore suspension, as a coffee foliage application at the laboratory. Nevertheless, they have resulted in TE1 and TE2 being significant, whereas TE-1 indicated the best bio-controlling ability as most efficiently reduces disease incidence by 35.8% and severity by 8.95%. Mulaw et al. [89] tested the Trichoderma spp. isolated from healthy coffee roots against the coffee tracheomycosis. They have observed that T. flagellatum strongly inhibits the Fusarium spp. In addition, further, they have found that the endophytes control the other pathogens such as Alternaria alternata, Botrytis cinerea, and Sclerotinia sclerotiorum, with inhibition percentages of 39.9 ± 2.2, 38.55 ± 9.56, and 10.67 ± 2.78, respectively.
In turn, Monteiro et al. [93] demonstrated the effect of volatile compounds produced by coffee endophytic fungi on various phytopathogens. They have found that endophytic Muscodor coffeanum (COAD 1842), and M. yucatanensis (HZM64) completely inhibit the growth of Botrytis cinerea, Cercospora coffeicola, Phoma sp., and Rizoctonia solani, while partially inhibit Aspergillus ochraceus and Pestalotia longisetula. Nevertheless, Muscodor vitigenus (HZM39) and Simplicillium sp. (C12) also showed complete inhibition of C. coffeicola, Phoma sp., and R. solani, in addition to P. longisetula, while partially controlling the B. cinerea. In another laboratory experiment with Induratia coffeana, Gomes et al. [126] showed the antimicrobial activity owing to the volatile compounds produced, against Aspergillus caespitosus, A. elegans, A. expansum, A. flavus, A. foetidus, A. niger, A. ochraceus A. sclerotiorum, A. sydowii, A. tamari, A. tubingensis, and A. versicolor either by decreasing the growth rate or, mainly, by completely inhibiting colony growth.
Other than the fungal pathogens, bacteria also could cause considerable disease prevalence in the coffee such as bacterial halo blight, bacterial leaf blight, bacterial leaf spot, and coffee leaf scorch; however, the antagonistic effect towards them by endophytes has not yet been tested, which shows another area to cover [127].
4.3.2. Protection against Parasitic Nematodes
It was found that a number of nematodes also could be associated with the coffee plantations and threaten, e.g., Apratylenchus spp., Criconemella spp., Pratylenchus spp., Radopholus spp., Rotylenchulus spp., and Xiphinema spp. [20]. It is also recorded that endophytic bacteria could control the nematodes, and also use various direct and indirect mechanisms to affect nematodes. For instance, parasitizing, (e.g., producing toxins, antibiotics, extracellular enzymes, and volatile compounds), interfering with nematode–plant-host recognition, competing for nutrients, triggering plant systemic defense mechanisms, and promoting plant health [128]. Enthrallingly, endophytic fungi also control the parasitic nematode by preying on them. In this case, endophytic fungi make constricting rings and trap the nematodes, and absorb the nutrients to let the host die. Those endophytes also have the ability to produce antimicrobial and nematicidal compounds like linoleic acid or pleurotin [129]. In a greenhouse study, Asyiah et al. [114] planted the Robusta coffee plants with a different bacterial consortium of Bacillus sp., and Pseudomonas dimunita. They found that the number of Pratylenchus coffeae in root or soil on a plant treated with bacterial consortium was significantly different from the control plant. Particularly, the application of Bacillus sp. alone indicated that 48.2%. suppression of the number of Pratylenchus coffeae in roots compared to the control. Further, Bacillus sp. only also minimizes the nematode population present in the rhizosphere by 54.6%. Contrary to the above findings, Asyiah and co-workers [114] recognized that the combination of endophytic bacteria with other rhizobacteria (e.g., Pseudomonas dimunita and Bacillus subtilis) showed more effectiveness than the single species isolations. For example, a combination of different endophyte isolates with Pseudomonas dimunita showed suppression of root-inhibiting Pratylenchus coffeae by 71.9% and soil dwellings by 76.1%. However, note that the beneficial effects of endophytic bacteria on host plants are typically greater than those of rhizospheric bacteria; thus the importance of endophytic bacteria in the consortium is highly recognized [30]. However, it is quite argumentative to conclude that a consortium of endophytes could work better than single isolation or vice versa. There are several factors to get varied or for inconsistent results in controlling probably due to the following of various study protocols, selected microbial strains, working conditions, geographical variation, and so on [130,131,132].
In one study, Asyiah et al. [87] showed that Bacillus subtilis controls and minimizes the penetration of Pratylenchus coffeae to coffee plants by up to 85%. A comprehensive study conducted by Hoang et al. [20] found that a coffee root endophytic bacterium, Streptomyces sp. has a great ability on controlling coffee nematode Meloidogyne incognita. From an in vitro assay, they observed that nematode eggs and larvae (L2) were destroyed by 85.8% and 85% respectively. In addition, negative effects on hatched juveniles were also recorded. Intriguingly, Hoang and co-authors [20] reported the further nematicidal activity of Streptomyces sp. on nematode cadaver, as the body shrunk after five days and morphologically visible the lysed body just after a week suggesting the toxic mechanism on the host. In a similar study, Mekete et al. [133] recognized several more root endophytic bacterial effects against the Meloidogyne incognita (L2 larvae). Further, they have recorded Agrobacterium radiobacter, Bacillus brevis, B. licheniformis, B. megaterium, B. mycoides, B. pumilus, Cedecea davisae, Chryseobacterium balustinum, Cytophaga johnsonae, Lactobacillus paracasei, Micrococcus halobius, M. luteus, Pseudomonas syringae, and Stenotrophomonas maltophilia sown significant controlling ability of the aforesaid nematode from 38 to 98%. Moreover, Mekete and colleagues [133] observed that Bacillus pumilus and B. mycoides were the most effective in minimizing the number of galls by 33% and egg masses by 39% causing the M. incognita. In a similar study, Duong and co-authors [84] reported the lethal effect of Bacillus sp. (closely related to B. cereus and B. mycoides) on Pratylenchus coffeae and Radopholus duriophilus.
4.3.3. Protection against Insect Pests
The application of entomopathogenic fungi in controlling pests has become a catchy topic for many researchers [134,135,136]. It is a gripping mechanism to evaluate how entomopathogenic fungi control insect pests. Of this, first fungi spores attach to the cuticle of the insect body, and start to germinate and penetrate through the cuticle (also via the digestive tract) and reach hemocoel. Inside the hemocoel, fungi start to replicate by utilizing nutrients, and insect hosts become weak due to the mycotoxins produced, as well as owing to the tissue invasion and nutrition deprivation, eventually die [137,138,139].
It is possible to find many entomopathogenic fungi naturally habited with coffee plants as endophytes, one such study, for example, Vega et al. [76] identified 16 species of entomopathogenic fungi from five genera, associated with coffee plants, namely Acremonium spp., Beauveria spp., Cladosporium spp., Clonostachys sp., and Paecilomyces spp. By far, endophytic Beauveria bassiana has been the most experimented species, also commercialized worldwide as mycoinsecticides [140]. Owing to their broad range of infecting on arthropods, many studies targeted the use of them in different crop fields [141,142]. Posada et al. [143] highlighted the different application methods of Beauveria bassiana to coffee plants such as foliar spraying, stem injection, and soil drench, which are helpful to introduce to the field when the absence of fungi or improve the field population. In a recent study, Bayman et al. [144] tested 29 local isolations of B. Bassiana and also compared their effectiveness with commercially available formulations-Mycotrol in controlling Hypothenemus hampei. From the laboratory experiment, they found that mortality of H. hampei varied according to the different strains of fungi, however, nearly half of the isolations showed over 80% controlling ability, moreover, several strains were headed the Mycotrol. Further, Bayman and colleagues [144] get successful results in their field experiment, where it was reported to reduce infestation and the number of pests per fruit. Nevertheless, the researchers further highlighted that the effect of B. bassiana on the pest depends on the microclimates, or with the changes in the abiotic and biotic environment. According to the study conducted by, Samuels et al. [145], other than B. bassiana, Metarhizium anisopliae also has the potential to control H. hampei significantly, however, may not be greater than B. bassiana does. In a subsequent paper, Vega et al. [76] also discussed their findings on the management of H. hampei using B. bassiana and Clonostachys rosea. According to their findings, B. bassiana_infection leads to the death of insects by just 4.8 ± 0.2 days and 14.7 ± 0.75 days for the insect treated with C. rosea.
Jia-ning et al. [146] demonstrated the control ability of coffee stem borers, Xylotrechus quardripes and Acalolepta cervinus using Beauveria bassiana in laboratory and field experiments. They have found effective control of aforesaid pests by B. bassiana in both environmental conditions. The control ability of different life stages of both pests is recognized, mortality percentage of young larvae, elder larvae, pupae, and adult of X. quardripes after 15 days of fungi application are 98.4 ± 3.2, 96.2 ± 0.5, 94.1 ± 0.8, and 89.1 ± 0.7 respectively. While the mortality percentage of young larvae and elder larvae of A. cervinus were 92.4 ± 6.6 and 89.1 ± 5.1 respectively. Further, in the field experiment, the results indicated that coffee stem borer damage was reduced (in 120 days) after the application of conidia spray of B. Bassiana by 44.4 ± 5.0. and 43.9 ± 4.4 when dusting fungus powder products by hand.
4.4. Role of Endophytes in the Improvement of Quality and Flavor of Coffee Seeds
Microbial communities associated with coffee play a crucial role in determining the quality of the final product. There could be negative and positive effects offered by the microflora, for example, Batista et al. [147] showed that Aspergillus flavus and Penicillium spp. associated with coffee seeds produce toxic compounds such as aflatoxin (B1 and B2) and ochratoxin A, respectively. However, our perception is of the beneficial effects on endophytes. The importance of endophytes has been shown particularly during the fermentation of coffee beans; however, their role during coffee fruit maturation in natural settings remains unknown [148]. It is believed that the prevalence of certain microbial groups in coffee cherries and the biochemical ramifications of their presence has significantly benefited the quality of coffee by means of improving both aroma and flavor. Pathways of inevitable colonization happening in coffee cherries by endophytes remain uncertain, nevertheless, previously discussed pathways to enter into the host could be applicable here, as once endophytes invaded systematic spreading is possible [148].
Coming to the point of benefiting effect, the importance of endophytes is recognized during the processing of coffee. First, knowing about coffee fermentation is worthwhile, at this stage; it removes mucilage from parchment coffee. Microorganisms, particularly bacteria, fungi, and yeast play a major role in degrading mucilage by producing various enzymes, alcohols, and acids during the fermentation process. In addition, this step is important for the development of aroma and flavor precursors [149]. In an early study, Sakiyama et al. [57] conducted a study with the aim of finding the effect of endophytic bacterial activity on the quality of coffee beverages. They have isolated the Paenibacillus amylolyticus associated with coffee beans. Further, Sakiyama and co-authors [57] found that the isolated P. amylolyticus have the great potential to produce extracellular pectinase, which plays a vital role during fermentation, ultimately enhancing the quality of the beverage. In a recent study, Liu et al. [150] isolated four naturally occurring strains of endophytes from the parchment and endosperm of green coffee beans and tested them to understand their ability to improve coffee fermentation. Among them, they have found that Bacillus cereus, and B. subtilis increase the anti-inflammatory chlorogenic acid by 38.5 and 51.5% respectively, compared to the unfermented beans.
Today, different levels of caffeine, a stimulant that is the key component in coffee beverages, are favored by consumers, in this regard the importance of coffee endophytes recognized, above the other traditional methods [151]. Liu and co-workers [150], fermented the coffee with Pichia guilliermondii and observed the biodecaffeination ability, dropping caffeine levels by about 38% compared to the unfermented beans. In addition, the study resulted that the aforesaid two Bacillus sp. improves the sensory characteristics of the coffee such as sweetness, after-taste, and clean cup. However, over the Liu et al. [150] findings on the lower level of biodecaffeination, Baker et al. [87] recognized that Pseudomonas sp., exhibiting higher caffeine degradation (98.61%), where cells induce is enough for degrading caffeine as the bacterium uses high concentrated caffeine as a sole source of carbon and nitrogen. Nunes and de Melo. [152] got similar results, testing with Pseudomonas putida as it completely degraded the alkaloid. In contrast, Chaves et al. [90] recognized that Aspergillus oryzae found in coffee leaves have the potential to increase the caffeine level. Moreover, a study by Li and colleagues [124] revealed the presence of health-benefiting genes in B. cereus and Priestia aryabhattai, which opened a new window for the research communities.
5. Conclusions
The studies on coffee endophytes are relatively new compared to many other plant-associated microbial studies. In 25 years of coffee endophytes’ history of research, only 107 studies have been conducted, basically identifying new species. However, the majority of studies are based on one-time sampling of the plant tissues (mainly, the leaf), and periodical sampling seems to be missing. It is also understood that the dynamic of endophytic microflora due to the climate and other environmental effects, is therefore vulnerable to change in the composition of the population, which may be hindering the real scenario. Additionally, many studies conducted in several geographical locations, making it impossible to comprehend the universal distribution of coffee endophytes. Though there are only limited studies available on both endophytic ability on coffee plant growth promotion and their availability as a biocontrol agent, available findings are striking. Further, as most of the studies are limited to lab experiments, there is a necessity to bring them into the field and test them for commercial applicability. Further, no studies conducted in relation to the endophytic effect on mitigating the abiotic stress of plants, thus future studies could be focused on this as well since climatic change is one of the burning issues in today's world that mount in agricultural productivity. The use of endophytes in those areas is undoubtedly advantageous, also by means of avoiding unfriendly pesticides and fertilizers driven for nature-friendly organic farming towards sustainable agriculture. In addition, one fascinating finding would be the use of endophytes in improving the sensory attributes of coffee, thus showing enhanced advantages in the coffee beverage industry.
Author Contributions
“Conceptualization, A.K.H.P., S.A. and S.C.K.; methodology, A.K.H.P.; software, A.K.H.P.; validation, S.A., S.T. and S.C.K.; formal analysis, Y.L.L., J.Q.Z., Z.Q.F., L.K.Z. and K.S.; investigation, C.N., I.P. and L.L.; data curation, S.T.; writing—original draft preparation, A.K.H.P.; writing—review and editing, A.K.H.P., S.A., S.C.K., Y.L.L., J.Q.Z., Z.Q.F., L.K.Z., K.S., C.N., I.P. and L.L.; visualization, S.C.K., and S.A.; supervision, S.C.K, S.T. and S.A.; project administration, S.A.; funding acquisition, S.A. All authors have read and agreed to the published version of the manuscript.
Funding
National Natural Science Foundation of China grant number 32260004 is thanked for the financial support.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available on request.
Acknowledgments
Samantha C. Karunarathna thanks Qujing Normal University, and National Institute of Fundamental Studies (NIFS), Sri Lanka. Suhail Asad thanks Pu’er University for the support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pham, Y.; Reardon-Smith, K.; Mushtaq, S.; Cockfield, G. The Impact of Climate Change and Variability on Coffee Production: A Systematic Review. Clim. Change 2019, 156, 609–630. [CrossRef]
- Bastian, F.; Hutabarat, O.S.; Dirpan, A.; Nainu, F.; Harapan, H.; Emran, T.B.; Simal-Gandara, J. From Plantation to Cup: Changes in Bioactive Compounds during Coffee Processing. Foods 2021, 10, 2827. [CrossRef]
- International Coffee Organization - What’s New Available online: https://www.ico.org/ (accessed on 16 April 2023).
- Ma, J.; Li, J.; He, H.; Jin, X.; Cesarino, I.; Zeng, W.; Li, Z. Characterization of Sensory Properties of Yunnan Coffee. Curr. Res. Food Sci. 2022, 5, 1205–1215. [CrossRef]
- Bermudez, S.; Voora, V.; Larrea, C. (the year is 2022) Coffee Prices and Sustainability Available online: https://www.iisd.org/system/files/2022-09/2022-global-market-report-coffee.pdf (accessed on 9 April 2023).
- Yebasse, M.; Shimelis, B.; Warku, H.; Ko, J.; Cheoi, K.J. Coffee Disease Visualization and Classification. Plants 2021, 10, 1257. [CrossRef]
- Noponen, M.R.A.; Haggar, J.P.; Edwards-Jones, G.; Healey, J.R. Intensification of Coffee Systems Can Increase the Effectiveness of REDD Mechanisms. Agric. Syst. 2013, 119, 1–9. [CrossRef]
- Rhiney, K.; Guido, Z.; Knudson, C.; Avelino, J.; Bacon, C.M.; Leclerc, G.; Aime, M.C.; Bebber, D.P. Epidemics and the Future of Coffee Production. Proc. Natl. Acad. Sci. U. S. A. 2021, 118, e2023212118. [CrossRef]
- McCook, S.; Vandermeer, J. The Big Rust and the Red Queen: Long-Term Perspectives on Coffee Rust Research. Phytopathology 2015, 105, 1164–1173. [CrossRef]
- Gichuru, E.; Alwora, G.; Gimase, J.; Kathurima, C. Coffee Leaf Rust (Hemileia vastatrix) in Kenya—A Review. Agronomy 2021, 11, 2590. [CrossRef]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L.; et al. The Coffee Leaf Rust Pathogen Hemileia vastatrix : One and a Half Centuries around the Tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [CrossRef]
- Li, L.; Várzea, V.M.P.; Xia, Q.; Xiang, W.; Tang, T.; Zhu, M.; He, C.; Pereira, A.P.; Lavado da Silva, M. do C.M.; Wu, W.; et al. First Report of Hemileia vastatrix (Coffee Leaf Rust) Physiological Races Emergent in Coffee Germplasm Collections in the Coffee-Cropping Regions of China. Plant Dis. 2021, 105, 4162. [CrossRef]
- Motisi, N.; Ribeyre, F.; Poggi, S. Coffee Tree Architecture and Its Interactions with Microclimates Drive the Dynamics of Coffee Berry Disease in Coffee Trees. Sci. Rep. 2019, 9, 2544. [CrossRef]
- Mulatu, A.; Megersa, N.; Teferi, D.; Alemu, T.; Vetukuri, R.R. Biological Management of Coffee Wilt Disease (Fusarium xylarioides) Using Antagonistic Trichoderma Isolates. Front. Plant Sci. 2023, 14, 1113949. [CrossRef]
- Ramos, J.B.; de Resende, M.L.V.; Botelho, D.M.d.S.; Pereira, R.C.M.; Reichel, T.; Balieiro, A.A.F.; Botega, G.P.; Abrahão, J.C. d. R. Screening Coffee Genotypes for Brown Eye Spot Resistance in Brazil. PLoS One 2022, 17, e0258822. [CrossRef]
- Nair, K.P.P. Coffee. In The Agronomy and Economy of Important Tree Crops of the Developing World; Elsevier: Amsterdam, Netherlands, 2010; pp. 181–208.
- Johnson, M.A.; Ruiz-Diaz, C.P.; Manoukis, N.C.; Verle Rodrigues, J.C. Coffee Berry Borer (Hypothenemus hampei), a Global Pest of Coffee: Perspectives from Historical and Recent Invasions, and Future Priorities. Insects 2020, 11, 882. [CrossRef]
- Dantas, J.; Motta, I.O.; Vidal, L.A.; Nascimento, E.F.M.B.; Bilio, J.; Pupe, J.M.; Veiga, A.; Carvalho, C.; Lopes, R.B.; Rocha, T.L.; et al. A Comprehensive Review of the Coffee Leaf Miner Leucoptera coffeella (Lepidoptera: Lyonetiidae)-A Major Pest for the Coffee Crop in Brazil and Others Neotropical Countries. Insects 2021, 12, 1130. [CrossRef]
- Gugliuzzo, A.; Criscione, G.; Biondi, A.; Aiello, D.; Vitale, A.; Polizzi, G.; Tropea Garzia, G. Seasonal Changes in Population Structure of the Ambrosia Beetle Xylosandrus compactus and Its Associated Fungi in a Southern Mediterranean Environment. PLoS One 2020, 15, e0239011. [CrossRef]
- Hoang, H.; Tran, L.H.; Nguyen, T.H.; Nguyen, D.A.T.; Nguyen, H.H.T.; Pham, N.B.; Trinh, P.Q.; de Boer, T.; Brouwer, A.; Chu, H.H. Occurrence of Endophytic Bacteria in Vietnamese Robusta Coffee Roots and Their Effects on Plant Parasitic Nematodes. Symbiosis 2020, 80, 75–84. [CrossRef]
- Infante, F. Pest Management Strategies against the Coffee Berry Borer (Coleoptera: Curculionidae: Scolytinae). J. Agric. Food Chem. 2018, 66, 5275–5280. [CrossRef]
- Pandey, M.; Kayastha, P.; Khanal, S.; Shrestha, S.; Thakur, G.; Adhikari, K.; Shah, K.K.; Pant, D.; Khanal, D. An Overview on Possible Management Strategies for Coffee White Stem Borer Xylotrechus quadripes Chevrolat (Coleoptera: Cerambycidae) in Nepal. Heliyon 2022, 8, e10445. [CrossRef]
- Chaudhary, P.; Agri, U.; Chaudhary, A.; Kumar, A.; Kumar, G. Endophytes and Their Potential in Biotic Stress Management and Crop Production. Front. Microbiol. 2022, 13, 933017. [CrossRef]
- Grabka, R.; d’Entremont, T.W.; Adams, S.J.; Walker, A.K.; Tanney, J.B.; Abbasi, P.A.; Ali, S. Fungal Endophytes and Their Role in Agricultural Plant Protection against Pests and Pathogens. Plants 2022, 11, 384. [CrossRef]
- Aamir, M.; Rai, K.K.; Zehra, A.; Kumar, S.; Yadav, M.; Shukla, V.; Upadhyay, R.S. Fungal Endophytes: Classification, Diversity, Ecological Role, and Their Relevance in Sustainable Agriculture. In Microbial Endophytes; Kumar, A., Singh, V.K., Eds.; Elsevier: Sawston, UK, 2020; pp. 291–323.
- Jia, M.; Chen, L.; Xin, H.-L.; Zheng, C.-J.; Rahman, K.; Han, T.; Qin, L.-P. A Friendly Relationship between Endophytic Fungi and Medicinal Plants: A Systematic Review. Front. Microbiol. 2016, 7, 906. [CrossRef]
- Golinska, P.; Wypij, M.; Agarkar, G.; Rathod, D.; Dahm, H.; Rai, M. Endophytic Actinobacteria of Medicinal Plants: Diversity and Bioactivity. Antonie Van Leeuwenhoek 2015, 108, 267–289. [CrossRef]
- Mishra, V.K.; Passari, A.K.; Leo, V.V.; Singh, B.P. Molecular Diversity and Detection of Endophytic Fungi Based on Their Antimicrobial Biosynthetic Genes. In Fungal Biology, Singh, B.P., Gupta, V.K., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–35.
- Gakuubi, M.M.; Munusamy, M.; Liang, Z.-X.; Ng, S.B. Fungal Endophytes: A Promising Frontier for Discovery of Novel Bioactive Compounds. J. Fungi 2021, 7, 786. [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant Beneficial Endophytic Bacteria: Mechanisms, Diversity, Host Range and Genetic Determinants. Microbiol. Res. 2019, 221, 36–49. [CrossRef]
- Fávaro, L.C. de L.; Sebastianes, F.L. de S.; Araújo, W.L. Epicoccum nigrum P16, a Sugarcane Endophyte, Produces Antifungal Compounds and Induces Root Growth. PLoS One 2012, 7, e36826. [CrossRef]
- Fadiji, A.E.; Babalola, O.O. Elucidating Mechanisms of Endophytes Used in Plant Protection and Other Bioactivities with Multifunctional Prospects. Front. Bioeng. Biotechnol. 2020, 8, 467. [CrossRef]
- Ahlawat, O.P.; Yadav, D.; Kashyap, P.L.; Khippal, A.; Singh, G. Wheat Endophytes and Their Potential Role in Managing Abiotic Stress under Changing Climate. J. Appl. Microbiol. 2022, 132, 2501–2520. [CrossRef]
- Rosenblueth, M.; Martínez-Romero, E. Bacterial Endophytes and Their Interactions with Hosts. Mol. Plant. Microbe. Interact. 2006, 19, 827–837. [CrossRef]
- Yadav, G.; Meena, M. Bioprospecting of Endophytes in Medicinal Plants of Thar Desert: An Attractive Resource for Biopharmaceuticals. Biotechnol. Rep. 2021, 30, e00629. [CrossRef]
- Flewelling, A.J.; Currie, J.; Gray, C.A.; and Johnson. J.A. Endophytes from Marine Macroalgae: Promising Sources of Novel Natural Products. Curr. Sci. 2015, 109, 88–111.
- Deutsch, Y.; Gur, L.; Frank, I.B.; Ezra, D. Endophytes from Algae, a Potential Source for New Biologically Active Metabolites for Disease Management in Aquaculture. Front. Mar. Sci. 2021, 8. [CrossRef]
- Suryanarayanan, T.S. Endophyte Research: Going beyond Isolation and Metabolite Documentation. Fungal Ecol. 2013, 6, 561–568. [CrossRef]
- Aleynova, O.A.; Kiselev, K.V. Interaction of Plants and Endophytic Microorganisms: Molecular Aspects, Biological Functions, Community Composition, and Practical Applications. Plants 2023, 12, 714. [CrossRef]
- Redecker, D.; Kodner, R.; Graham, L.E. Glomalean Fungi from the Ordovician. Science 2000, 289, 1920–1921. [CrossRef]
- Krings, M.; Taylor, T.N.; Hass, H.; Kerp, H.; Dotzler, N.; Hermsen, E.J. Fungal Endophytes in a 400-Million-Yr-Old Land Plant: Infection Pathways, Spatial Distribution, and Host Responses. New Phytol. 2007, 174, 648–657. [CrossRef]
- Schüßler, A. Molecular phylogeny, taxonomy, and evolution of Geosiphon pyriformis and arbuscular mycorrhizal fungi. Plant Soil 2002, 244, 75–83. [CrossRef]
- Remy, W.; Taylor, T.N.; Hass, H.; Kerp, H. Four Hundred-Million-Year-Old Vesicular Arbuscular Mycorrhizae. Proc. Natl. Acad. Sci. 1994, 91, 11841–11843.
- Krings, M.; Taylor, T.N.; Dotzler, N. Fungal Endophytes as a Driving Force in Land Plant Evolution: Evidence from the Fossil Record. In Biocomplexity of Plant-Fungal Interactions; Southworth, D., Eds.; John Wiley & Sons: Oxford, UK, 2012; pp. 5–27.
- Verstraete, B.; Janssens, S.; Rønsted, N. Non-Nodulated Bacterial Leaf Symbiosis Promotes the Evolutionary Success of Its Host Plants in the Coffee Family (Rubiaceae). Mol. Phylogenet. Evol. 2017, 113, 161–168. [CrossRef]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical Ecology of Endophytic Fungi: Origins of Secondary Metabolites. Chem. Biol. 2012, 19, 792–798. [CrossRef]
- El-Bondkly, E.A.M.; El-Bondkly, A.A.M.; El-Bondkly, A.A.M. Marine Endophytic Fungal Metabolites: A Whole New World of Pharmaceutical Therapy Exploration. Heliyon 2021, 7, e06362. [CrossRef]
- Schulz, B.; Boyle, C. What Are Endophytes? In Soil Biology, Schulz, B.J.E., Boyle, C.J.C., Sieber, T.N., Eds.; Springer: Berlin, Heidelberg: Germany, 2007; Volume 9, pp. 1–13.
- Tiwari, P.; Kang, S.; Bae, H. Plant-Endophyte Associations: Rich yet under-Explored Sources of Novel Bioactive Molecules and Applications. Microbiol. Res. 2023, 266, 127241. [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of Bacterial Endophytes and Their Proposed Role in Plant Growth. Trends Microbiol. 2008, 16, 463–471. [CrossRef]
- Rochín-Hernández, L.S.; Rochín-Hernández, L.J.; Flores-Cotera, L.B. Endophytes, a Potential Source of Bioactive Compounds to Curtail the Formation–Accumulation of Advanced Glycation End Products: A Review. Molecules 2022, 27, 4469. [CrossRef]
- Yarzábal, L.A.; Chica, E.J. Role of Rhizobacterial Secondary Metabolites in Crop Protection against Agricultural Pests and Diseases. In New and Future Developments in Microbial Biotechnology and Bioengineering; Gupta, V.K., Pandey, A., Eds; Elsevier: Amsterdam, Netherlands, 2019; pp. 31–53.
- Oszust, K.; Cybulska, J.; Frąc, M. How Do Trichoderma Genus Fungi Win a Nutritional Competition Battle against Soft Fruit Pathogens? A Report on Niche Overlap Nutritional Potentiates. Int. J. Mol. Sci. 2020, 21, 4235. [CrossRef]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. Int. J. Mol. Sci. 2022, 23, 2329. [CrossRef]
- Santoyo, G.; Urtis-Flores, C.A.; Loeza-Lara, P.D.; Orozco-Mosqueda, M.d.C.; Glick, B.R. Rhizosphere Colonization Determinants by Plant Growth-Promoting Rhizobacteria (PGPR). Biology 2021, 10, 475. [CrossRef]
- Hansen, M.; Kragelund, L.; Nybroe, O.; Sørensen, J. Early Colonization of Barley Roots by Pseudomonas fluorescens Studied by Immunofluorescence Technique and Confocal Laser Scanning Microscopy. FEMS Microbiol. Ecol. 1997, 23, 353–360. [CrossRef]
- Sakiyama, C.C.H; Paula, E.M.; Pereira, P.C.; Borges, A.C.; Silva, D.O. Characterization of Pectin Lyase Produced by an Endophytic Strain Isolated from Coffee Cherries. Lett. Appl. Microbiol. 2001, 33, 117–121. [CrossRef]
- Maela, P.M.; Serepa-Dlamini, M.H. Current Understanding of Bacterial Endophytes, Their Diversity, Colonization and Their Roles in Promoting Plant Growth. Appl. Microbiol. Open Access 2019, 5, 1–13.
- Huang, B.; Lv, C.; Zhuang, P.; Zhang, H.; Fan, L. Endophytic Colonisation of Bacillus subtilis in the Roots of Robinia pseudoacacia L. Plant Biol. 2011, 13, 925–931. [CrossRef]
- Patel, P.; Kumar, S.; Modi, A.; Kumar, A. Deciphering Fungal Endophytes Combating Abiotic Stresses in Crop Plants (Cereals and Vegetables). In Microbial Management of Plant Stresses; Ajay Kumar, A., Droby, S., Eds.; Elsevier: Sawston, UK, 2021; pp. 131–147.
- Malfanova, N.; Lugtenberg, B.J.J.; Berg, G. Bacterial Endophytes: Who and Where, and What Are They Doing There? In Molecular Microbial Ecology of the Rhizosphere; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 391–403.
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The Hidden World within Plants: Ecological and Evolutionary Considerations for Defining Functioning of Microbial Endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [CrossRef]
- Ding, L.-N.; Li, Y.-T.; Wu, Y.-Z.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X.-L. Plant Disease Resistance-Related Signaling Pathways: Recent Progress and Future Prospects. Int. J. Mol. Sci. 2022, 23, 16200. [CrossRef]
- Deng, Y.; Chen, H.; Li, C.; Xu, J.; Qi, Q.; Xu, Y.; Zhu, Y.; Zheng, J.; Peng, D.; Ruan, L.; et al. Endophyte Bacillus subtilis Evade Plant Defense by Producing Lantibiotic Subtilomycin to Mask Self-Produced Flagellin. Commun. Biol. 2019, 2, 368. [CrossRef]
- Taghavi, S.; van der Lelie, D.; Hoffman, A.; Zhang, Y.-B.; Walla, M.D.; Vangronsveld, J.; Newman, L.; Monchy, S. Genome Sequence of the Plant Growth Promoting Endophytic Bacterium Enterobacter Sp. 638. PLoS Genet. 2010, 6, e1000943. [CrossRef]
- Ji, L.; Yang, X.; Qi, F. Distinct Responses to Pathogenic and Symbionic Microorganisms: The Role of Plant Immunity. Int. J. Mol. Sci. 2022, 23, 10427. [CrossRef]
- Mengistu, A. A. Endophytes: Colonization, Behaviour, and Their Role in Defense Mechanism. Int. J. Microbiol. 2020, 2020, 6927219. [CrossRef]
- Eid, A.M.; Fouda, A.; Abdel-Rahman, M.A.; Salem, S.S.; Elsaied, A.; Oelmüller, R.; Hijri, M.; Bhowmik, A.; Elkelish, A.; Hassan, S.E.-D. Harnessing Bacterial Endophytes for Promotion of Plant Growth and Biotechnological Applications: An Overview. Plants 2021, 10, 935. [CrossRef]
- Rodriguez, R.J.; White Jr, J.F.; Arnold, A.E.; Redman, R.S. Fungal Endophytes: Diversity and Functional Roles. New Phytol. 2009, 182, 314–330. [CrossRef]
- Lugtenberg, B.J.J.; Caradus, J.R.; Johnson, L.J. Fungal Endophytes for Sustainable Crop Production. FEMS Microbiol. Ecol. 2016, 92, fiw194. [CrossRef]
- Petrini, O. Fungal Endophytes of Tree Leaves. In: Andrews, J.H., Hirano, S.S. (eds) Microbial Ecology of Leaves. Brock/Springer Series in Contemporary Bioscience. Springer, New York, NY., 1991; pp. 179–197 ISBN 9781461278221.
- Sampson, K. The Presence and Absence of an Endophytic Fungus in Lolium temulentum and L. perenne. Trans. Br. Mycol. Soc. 1935, 19, 337–343. [CrossRef]
- Neill, J.C. The endophyte of ryegrass (Lolium perenne). N. Z. J. Sci. Technol., 1940, 21, 280–291.
- Carroll, G. Forest Endophytes: Pattern and Process. Can. J. Bot. 1995, 73, 1316–1324. [CrossRef]
- Petrini, O.; Fisher, P.J. Fungal Endophytes in Salicornia perennis. Trans. Br. Mycol. Soc. 1986, 87, 647–651. [CrossRef]
- Vega, F.E.; Posada, F.; Catherine Aime, M.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic Fungal Endophytes. Biol. Control 2008, 46, 72–82. [CrossRef]
- Jimenez-Salgado, T.; Fuentes-Ramirez, L.E.; Tapia-Hernandez, A.; Mascarua-Esparza, M.A.; Martinez-Romero, E.; Caballero-Mellado, J. Coffea arabica L., a New Host Plant for Acetobacter diazotrophicus, and Isolation of Other Nitrogen-Fixing Acetobacteria. Appl. Environ. Microbiol. 1997, 63, 3676–3683. [CrossRef]
- Jurburg, S.D.; Shek, K.L.; McGuire, K. Soil Microbial Composition Varies in Response to Coffee Agroecosystem Management. FEMS Microbiol. Ecol. 2020, 96. [CrossRef]
- Bongiorno, V.A.; Rhoden, S.A.; Garcia, A.; Polonio, J.C.; Azevedo, J.L.; Pereira, J.O.; Pamphile, J.A. Genetic Diversity of Endophytic Fungi from Coffea arabica Cv. IAPAR-59 in Organic Crops. Ann. Microbiol. 2016, 66, 855–865. [CrossRef]
- Vega, F.E.; Simpkins, A.; Aime, M.C.; Posada, F.; Peterson, S.W.; Rehner, S.A.; Infante, F.; Castillo, A.; Arnold, A.E. Fungal Endophyte Diversity in Coffee Plants from Colombia, Hawai’i, Mexico and Puerto Rico. Fungal Ecol. 2010, 3, 122–138. [CrossRef]
- Saucedo-García, A.; Anaya, A.L.; Espinosa-García, F.J.; González, M.C. Diversity and Communities of Foliar Endophytic Fungi from Different Agroecosystems of Coffea arabica L. in Two Regions of Veracruz, Mexico. PLoS One 2014, 9, e98454. [CrossRef]
- Oliveira, R.J.V.; Souza, R.G.; Lima, T.E.F; Cavalcanti, M.A.Q. Endophytic Fungal Diversity in Coffee Leaves (Coffea arabica) Cultivated Using Organic and Conventional Crop Management Systems. Mycosphere 2014, 5, 523–530. [CrossRef]
- Gagliardi, S.; Avelino, J.; Fulthorpe, R.; Virginio Filho, E. de M.; Isaac, M.E. No Evidence of Foliar Disease Impact on Crop Root Functional Strategies and Soil Microbial Communities: What Does This Mean for Organic Coffee? Oikos 2022, 2022, e08987. [CrossRef]
- Duong, B.; Nguyen, H.X.; Phan, H.V.; Colella, S.; Trinh, P.Q.; Hoang, G.T.; Nguyen, T.T.; Marraccini, P.; Lebrun, M.; Duponnois, R. Identification and Characterization of Vietnamese Coffee Bacterial Endophytes Displaying in Vitro Antifungal and Nematicidal Activities. Microbiol. Res. 2021, 242, 126613. [CrossRef]
- Shiomi, H.F.; Silva, H.S.A.; de Melo, I.S. de; Nunes, F.V.; Bettiol, W. Bioprospecting Endophytic Bacteria for Biological Control of Coffee Leaf Rust. Sci. Agric. (Piracicaba, Braz.) 2006, 63, 32–39. [CrossRef]
- Vega, F.E.; Pava-Ripoll, M.; Posada, F.; Buyer, J.S. Endophytic Bacteria in Coffea arabica L. J. Basic Microbiol. 2005, 45, 371–380. [CrossRef]
- Asyiah, I.N.; Soekarto; Hoesain, M; Iqbal, M.;Hindersah, R.; Narulita, E; Mudakir,I. The Endophytic Bacteria Isolation as Biological Control Agent of Pratylenchus coffeae. Asian Jr. of Microbiol. Biotech. Env. Sc. 20, 2018: 159–165.
- Baker, S.; Sahana, S.; Rakshith, D.; Kavitha, H.U.; Kavitha, K.S.; Satish, S. Biodecaffeination by Endophytic Pseudomonas sp. Isolated from Coffee arabica L. J. Pharm. Res. 2012, 5, 3654–3657.
- Mulaw, T.B.; Druzhinina, I.S.; Kubicek, C.P.; Atanasova, L. Novel Endophytic Trichoderma spp. Isolated from Healthy Coffea arabica Roots are Capable of Controlling Coffee Tracheomycosis. Diversity 2013, 5, 750-766. [CrossRef]
- Chaves, F.C.; Gianfagna, T.J.; Aneja, M.; Posada, F.; Peterson, S.W.; Vega, F.E. Aspergillus oryzae NRRL 35191 from Coffee, a Non-Toxigenic Endophyte with the Ability to Synthesize Kojic Acid. Mycol. Prog. 2012, 11, 263–267. [CrossRef]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Hyde, K.D. Characterization of Colletotrichum Species Associated with Coffee Berries in Northern Thailand. Fungal Divers. 2009, 89–109.
- Monteiro, M.C.P.; Tavares, D.G.; Nery, E.M.; de Queiroz, M.V.; Pereira, O.L.; Cardoso, P.G. Enzyme Production by Induratia spp. Isolated from Coffee Plants in Brazil. Braz. Arch. Biol. Technol. 2020, 63. [CrossRef]
- Monteiro, M.C.; Alves, N.M.; Queiroz, M.V.; Pinho, D.B.; Pereira, O.L.; Souza, S.M.; Cardoso, P.G. Antimicrobial activity of endophytic fungi from coffee plants. Biosci. j. 2017, 381–389.
- Peterson, S.W.; Vega, F.E.; Posada, F.; Nagai, C. Penicillium Coffeae,a New Endophytic Species Isolated from a Coffee Plant and Its Phylogenetic Relationship to P. fellutanum, P. thiersiiand P. brocae Based on Parsimony Analysis of Multilocus DNA Sequences. Mycologia 2005, 97, 659–666. [CrossRef]
- Mamani-Huayhua, G.; Leon-Ttacca, B; Palao-Iturregui, L.A.; Borja-Loza, Y.R. Biocontrol of coffee yellow rust (Hemileia vastatrix Berk. & Br.) with Trichoderma sp. endophyte strains. Cultivos Tropicales. 2021, 42, e01.
- de Lima, T.E.F.; de Oliveira, R.J.V.; Neves, R.P.; Bezerra, J.L.; de Queiroz Cavalcanti, M.A. Endophytic Yeasts of Coffea arabica and Vitis labrusca Cv. Isabel from Pernambuco, Brazil. Nova Hedwigia 2013, 96, 463–469. [CrossRef]
- Cun, H.; Munir, S.; He, P.; Wu, Y.; He, P.; Ahmed, A.; Che, H.; Li, J.; He, Y. Diversity of Root Endophytic Bacteria from Maize Seedling Involved in Biocontrol and Plant Growth Promotion. Egypt. J. Biol. Pest Contr. 2022, 32. [CrossRef]
- Sessitsch, A.; Reiter, B.; Berg, G. Endophytic Bacterial Communities of Field-Grown Potato Plants and Their Plant-Growth-Promoting and Antagonistic Abilities. Can. J. Microbiol. 2004, 50, 239–249. [CrossRef]
- Walitang, D.I.; Kim, K.; Madhaiyan, M.; Kim, Y.K.; Kang, Y.; Sa, T. Characterizing Endophytic Competence and Plant Growth Promotion of Bacterial Endophytes Inhabiting the Seed Endosphere of Rice. BMC Microbiol. 2017, 17, 209. [CrossRef]
- Dhungana, S.A; Adachi, F.; Hayashi, S.; Puri, R.R; Itoh, K. Plant Growth Promoting Effects of Nepalese Sweet Potato Endophytes. Horticulturae 2018, 4, 53. [CrossRef]
- Yan, X.; Wang, Z.; Mei, Y.; Wang, L.; Wang, X.; Xu, Q.; Peng, S.; Zhou, Y.; Wei, C. Isolation, Diversity, and Growth-Promoting Activities of Endophytic Bacteria from Tea Cultivars of Zijuan and Yunkang-10. Front. Microbiol. 2018, 9, 1848. [CrossRef]
- Morsy, M.; Cleckler, B.; Armuelles-Millican, H. Fungal Endophytes Promote Tomato Growth and Enhance Drought and Salt Tolerance. Plants 2020, 9, 877. [CrossRef]
- Fouda, A.; Eid, A.M.; Elsaied, A.; El-Belely, E.F.; Barghoth, M.G.; Azab, E.; Gobouri, A.A.; Hassan, S.E.-D. Plant Growth-Promoting Endophytic Bacterial Community Inhabiting the Leaves of Pulicaria incisa (Lam.) DC Inherent to Arid Regions. Plants 2021, 10, 76. [CrossRef]
- Hassan, S.E.-D. Plant Growth-Promoting Activities for Bacterial and Fungal Endophytes Isolated from Medicinal Plant of Teucrium polium L. J. Adv. Res. 2017, 8, 687–695. [CrossRef]
- Lin, H.; Liu, C.; Peng, Z.; Tan, B.; Wang, K.; Liu, Z. Distribution Pattern of Endophytic Bacteria and Fungi in Tea Plants. Front. Microbiol. 2022, 13, 872034. [CrossRef]
- Verma, S.K.; Sahu, P.K.; Kumar, K.; Pal, G.; Gond, S.K.; Kharwar, R.N.; White, J.F. Endophyte Roles in Nutrient Acquisition, Root System Architecture Development and Oxidative Stress Tolerance. J. Appl. Microbiol. 2021, 131, 2161–2177. [CrossRef]
- Chaudhary, R.; Kumar, V.; Gupta, S.; Naik, B.; Prasad, R.; Mishra, S.; Saris, P.E.J.; Kumar, V. Finger Millet (Eleusine coracana) Plant–Endophyte Dynamics: Plant Growth, Nutrient Uptake, and Zinc Biofortification. Microorganisms 2023, 11, 973. [CrossRef]
- Pratiwi, E.R.; Ardyati, T.; Suharjono, S. Plant Growth Promoting Endophytic Bacteria of Coffea canephora and Coffea arabica L. in UB Forest. J. Exp. Life Sci. 2020, 10, 119–126. [CrossRef]
- Jha, C.K.; Patel, B.; Saraf, M. Stimulation of the Growth of Jatropha curcas by the Plant Growth Promoting Bacterium Enterobacter cancerogenus MSA2. World J. Microbiol. Biotechnol. 2012, 28, 891–899. [CrossRef]
- Muleta, D.; Assefa, F.; Börjesson, E.; Granhall, U. Phosphate-Solubilising Rhizobacteria Associated with Coffea arabica L. in Natural Coffee Forests of Southwestern Ethiopia. J. Saudi Soc. Agric. Sci. 2013, 12, 73–84. [CrossRef]
- Teshome, B.; Wassie, M.; Abatneh, E. Isolation, Screening and Biochemical Characterization of Phosphate-Solubilizing Rhizobacteria Associated with Coffea arabica L. J Fertil Pestic 2017, 8. [CrossRef]
- García-Latorre, C.; Rodrigo, S.; Santamaria, O. Effect of Fungal Endophytes on Plant Growth and Nutrient Uptake in Trifolium subterraneum and Poa pratensis as Affected by Plant Host Specificity. Mycol. Prog. 2021, 20, 1217–1231. [CrossRef]
- Silva, H.S.A.; Tozzi, J.P.L.; Terrasan, C.R.F.; Bettiol, W. Endophytic Microorganisms from Coffee Tissues as Plant Growth Promoters and Biocontrol Agents of Coffee Leaf Rust. Biol. Control 2012, 63, 62–67. [CrossRef]
- Asyiah, I.N.; Mudakir, I.; Hoesain, M.; Pradana, A.P.; Djunaidy, A.; Sari, R.F. Consortium of Endophytic Bacteria and Rhizobacteria Effectively Suppresses the Population of Pratylenchus coffeae and Promotes the Growth of Robusta Coffee. Biodiversitas 2020, 21, 4702–4708. [CrossRef]
- Clay, K. The Ecology and Evolution of Endophytes. Agric. Ecosyst. Environ. 1993, 44, 39–64. [CrossRef]
- Ali, S.; Hameed, S.; Imran, A.; Iqbal, M.; Lazarovits, G. Genetic, Physiological and Biochemical Characterization of Bacillus sp. Strain RMB7 Exhibiting Plant Growth Promoting and Broad Spectrum Antifungal Activities. Microb. Cell Fact. 2014, 13, 144. [CrossRef]
- Kejela, T.; Thakkar, V.R.; Thakor, P. Bacillus Species (BT42) Isolated from Coffea arabica L. Rhizosphere Antagonizes Colletotrichum gloeosporioides and Fusarium oxysporum and Also Exhibits Multiple Plant Growth Promoting Activity. BMC Microbiol. 2016, 16, 277. [CrossRef]
- Palmieri, D.; Ianiri, G.; Del Grosso, C.; Barone, G.; De Curtis, F.; Castoria, R.; Lima, G. Advances and Perspectives in the Use of Biocontrol Agents against Fungal Plant Diseases. Horticulturae 2022, 8, 577. [CrossRef]
- Trutmann, P.; Keane, P.J. Trichoderma koningii as a Biological Control Agent for Sclerotinia sclerotiorum in Southern Australia. Soil Biol. Biochem. 1990, 22, 43–50. [CrossRef]
- Morandi, M.A.B.; Sutton, J.C.; Maffia, L.A. Effects of Host and Microbial Factors on Development of Clonostachys rosea and Control of Botrytis cinerea in Rose. Eur. J. Plant Pathol. 2000, 106, 439–448. [CrossRef]
- Mousa, W.K.; Raizada, M.N. The Diversity of Anti-Microbial Secondary Metabolites Produced by Fungal Endophytes: An Interdisciplinary Perspective. Front. Microbiol. 2013, 4, 65. [CrossRef]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal Effectors and Plant Susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [CrossRef]
- Baiyee, B.; Ito, S.-I.; Sunpapao, A. Trichoderma asperellum T1 Mediated Antifungal Activity and Induced Defense Response against Leaf Spot Fungi in Lettuce (Lactuca sativa L.). Physiol. Mol. Plant Pathol. 2019, 106, 96–101. [CrossRef]
- Li, Z.; Wen, W.; Qin, M.; He, Y.; Xu, D.; Li, L. Biosynthetic Mechanisms of Secondary Metabolites Promoted by the Interaction between Endophytes and Plant Hosts. Front. Microbiol. 2022, 13, 928967. [CrossRef]
- Cacefo, V.; De Araújo, F. F.; Pacheco, A. C. Biological control of Hemileia vastatrix berk. & broome with bacillus subtilis cohn and biochemical changes in the coffee. Coffee Sci. 2017, 11, 567–574.
- Gomes, A.A.M.; Paes, S.A.; Ferreira, A.P.S.; Pinho, D.B.; de Lourdes Cardeal, Z.; Menezes, H.C.; Cardoso, P.G.; Pereira, O.L. Endophytic Species of Induratia from Coffee and Carqueja Plants from Brazil and Its Potential for the Biological Control of Toxicogenic Fungi on Coffee Beans by Means of Antimicrobial Volatiles. Braz. J. Microbiol. 2023, 54, 349–360. [CrossRef]
- Badel, J.L.; Zambolim, L. Coffee Bacterial Diseases: A Plethora of Scientific Opportunities. Plant Pathol. 2019, 68, 411–425. [CrossRef]
- Tian, B.; Yang, J.; Zhang, K.-Q. Bacteria Used in the Biological Control of Plant-Parasitic Nematodes: Populations, Mechanisms of Action, and Future Prospects: Nematophagous Bacteria. FEMS Microbiol. Ecol. 2007, 61, 197–213. [CrossRef]
- Moosavi, M.R.; Zare, R. Fungi as Biological Control Agents of Plant-Parasitic Nematodes. In Plant Defence: Biological Control; Mérillon, J., Ramawat, K., Eds.; Springer: Dordrecht, Netherlands, 2012; Volume 12, pp. 67–107.
- Usta, C. Microorganisms in Biological Pest Control—A Review (Bacterial Toxin Application and Effect of Environmental Factors). In Current Progress in Biological Research; Silva-Opps, M., Eds.; InTech: London, UK, 2013, pp. 287–317.
- Chattopadhyay, P.; Banerjee, G.; Mukherjee, S. Recent Trends of Modern Bacterial Insecticides for Pest Control Practice in Integrated Crop Management System. 3 Biotech 2017, 7. [CrossRef]
- Kumar, K.K.; Dara, S.K. Fungal and Bacterial Endophytes as Microbial Control Agents for Plant-Parasitic Nematodes. Int. J. Environ. Res. Public Health 2021, 18, 4269. [CrossRef]
- Mekete, T.; Hallmann, J.; Kiewnick, S.; Sikora, R. Endophytic Bacteria from Ethiopian Coffee Plants and Their Potential to Antagonise Meloidogyne incognita. Nematology 2009, 11, 117–127. [CrossRef]
- Ambele, C.F.; Ekesi, S.; Bisseleua, H.D.B.; Babalola, O.O.; Khamis, F.M.; Djuideu, C.T.L.; Akutse, K.S. Entomopathogenic Fungi as Endophytes for Biological Control of Subterranean Termite Pests Attacking Cocoa Seedlings. J. Fungi 2020, 6, 126. [CrossRef]
- Deka, B.; Babu, A.; Peter, A.J.; Kumhar, K.C.; Sarkar, S.; Rajbongshi, H.; Dey, P.;Amalraj, E.L.D.; Talluri, V.R. Potential of the Entomopathogenic Fungus, Metarhizium anisopliae s.l. in Controlling Live-Wood Eating Termite, Microtermes obesi (Holmgren) (Blattodea: Termitidae) Infesting Tea Crop. Egypt. J. Biol. Pest Contr. 2021, 31. [CrossRef]
- Viswakethu, V.; Balakrishanan, P.; Murugan, L.; Narayana swamy, B.; Subbaraya, U. Entomopathogenic Fungi as a Promising Biological Control Agent against Banana Fruit Scarring Beetle, Basilepta subcostata (Jac.) (Chrysomelidae: Coleoptera). Egypt. J. Biol. Pest Contr. 2021, 31. [CrossRef]
- Donzelli, B. G.G.; Krasnoff, S. B. Molecular Genetics of Secondary Chemistry in Metarhizium Fungi. Adv. Genet., 2016, 94, 365–436. [CrossRef]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; et al. Insect-Fungal-Interactions: A Detailed Review on Entomopathogenic Fungi Pathogenicity to Combat Insect Pests. Microb. Pathog. 2021, 159, 105122. [CrossRef]
- Zhou, Q.; Yu, L.; Ying, S.-H., Feng, M.-G. Comparative Roles of Three Adhesin Genes (adh1-3) in Insect-Pathogenic Lifecycle of Beauveria bassiana. Appl. Micro;iol. Biotechnol. 2021, 105, 5491–5502. [CrossRef]
- Rehner, S.A.; Posada, F.; Buckley, E.P.; Infante, F.; Castillo, A.; Vega, F.E. Phylogenetic Origins of African and Neotropical Beauveria Bassiana s.l. Pathogens of the Coffee Berry Borer, Hypothenemus hampei. J. Invertebr. Pathol. 2006, 93, 11–21. [CrossRef]
- Posada, F.; Vega, F.E. Inoculation and Colonization of Coffee Seedlings (Coffea arabica L.) with the Fungal Entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycoscience 2006, 47, 284–289. [CrossRef]
- Gasmi, L.; Baek, S.; Kim, J.C.; Kim, S.; Lee, M.R.; Park, S.E.; Shin, T.Y.; Lee, S.J.; Parker, B.L.; Kim, J.S. Gene Diversity Explains Variation in Biological Features of Insect Killing Fungus, Beauveria bassiana. Sci. Rep. 2021, 11, 91. [CrossRef]
- Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A.; Vega, F.E. Inoculation of Coffee Plants with the Fungal Entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol. Res. 2007, 111, 748–757. [CrossRef]
- Bayman, P.; Mariño, Y.A.; García-Rodríguez, N.M.; Oduardo-Sierra, O.F.; Rehner, S.A. Local Isolates of Beauveria bassiana for Control of the Coffee Berry Borer Hypothenemus hampei in Puerto Rico: Virulence, Efficacy and Persistence. Biol. Control 2021, 155, 104533. [CrossRef]
- Samuels, R.I.; Pereira, R.C.; Gava, C.A.T. Infection of the Coffee Berry Borer Hypothenemus hampei (Coleoptera: Scolytidae) by Brazilian Isolates of the Entomopathogenic Fungi Beauveria bassiana and Metarhizium anisopliae (Deuteromycotina: Hyphomycetes). Biocontrol Sci. Technol. 2002, 12, 631–635. [CrossRef]
- Jia-ning, W.; Rong-ping, K. Biological Control of Coffee Stem Borers, Xylotrechus quardrlpes and Acalolepla cervznus, By Beaweha bassiana Preparation. Insect Sci. 2002, 9, 43–50. [CrossRef]
- Batista, L.R.; Chalfoun, S.M.; Prado, G.; Schwan, R.F.; Wheals, A.E. Toxigenic Fungi Associated with Processed (Green) Coffee Beans (Coffea arabica L.). Int. J. Food Microbiol. 2003, 85, 293–300. [CrossRef]
- Oliveira, M.N.V.; Santos, T.M.A.; Vale, H.M.M.; Delvaux, J.C.; Cordero, A.P.; Ferreira, A.B.; Miguel, P.S.B.; Tótola, M.R.; Costa, M.D.; Moraes, C.A.; et al. Endophytic Microbial Diversity in Coffee Cherries of Coffea arabica from Southeastern Brazil. Can. J. Microbiol. 2013, 59, 221–230. [CrossRef]
- Haile, M.; Kang, W.H. The Role of Microbes in Coffee Fermentation and Their Impact on Coffee Quality. J. Food Qual. 2019, 4836709. [CrossRef]
- Liu, F.; Song, Z.; Zhang, T.; Tong, X.; Chen, M.Y.; Gao, D.; Chen, J.; Ho, C.L. Characterization of the Therapeutic Properties and Flavor Profile of Coffee via Monoculture Fermentation with Endophytic Microbial Isolates. ACS Food Sci. Technol. 2022, 2, 1039–1049. [CrossRef]
- Abrahão, M.R.E.; Molina, G.; Pastore, G.M. Endophytes: Recent Developments in Biotechnology and the Potential for Flavor Production. Food Res. Int. 2013, 52, 367–372. [CrossRef]
- Nunes, F.V.; de Melo, I.S. Isolation and Characterization of Endophytic Bacteria of Coffee Plants and Their Potential in Caffeine Degradation. In Proceedings of the Environmental Toxicology, Kungolos, A.G., Brebbia, C.A., Samaras, C.P., Eds.; WIT Press: Southampton, UK, 2006, pp. 293–297.
Figure 1.
Procedure for the literature selection.
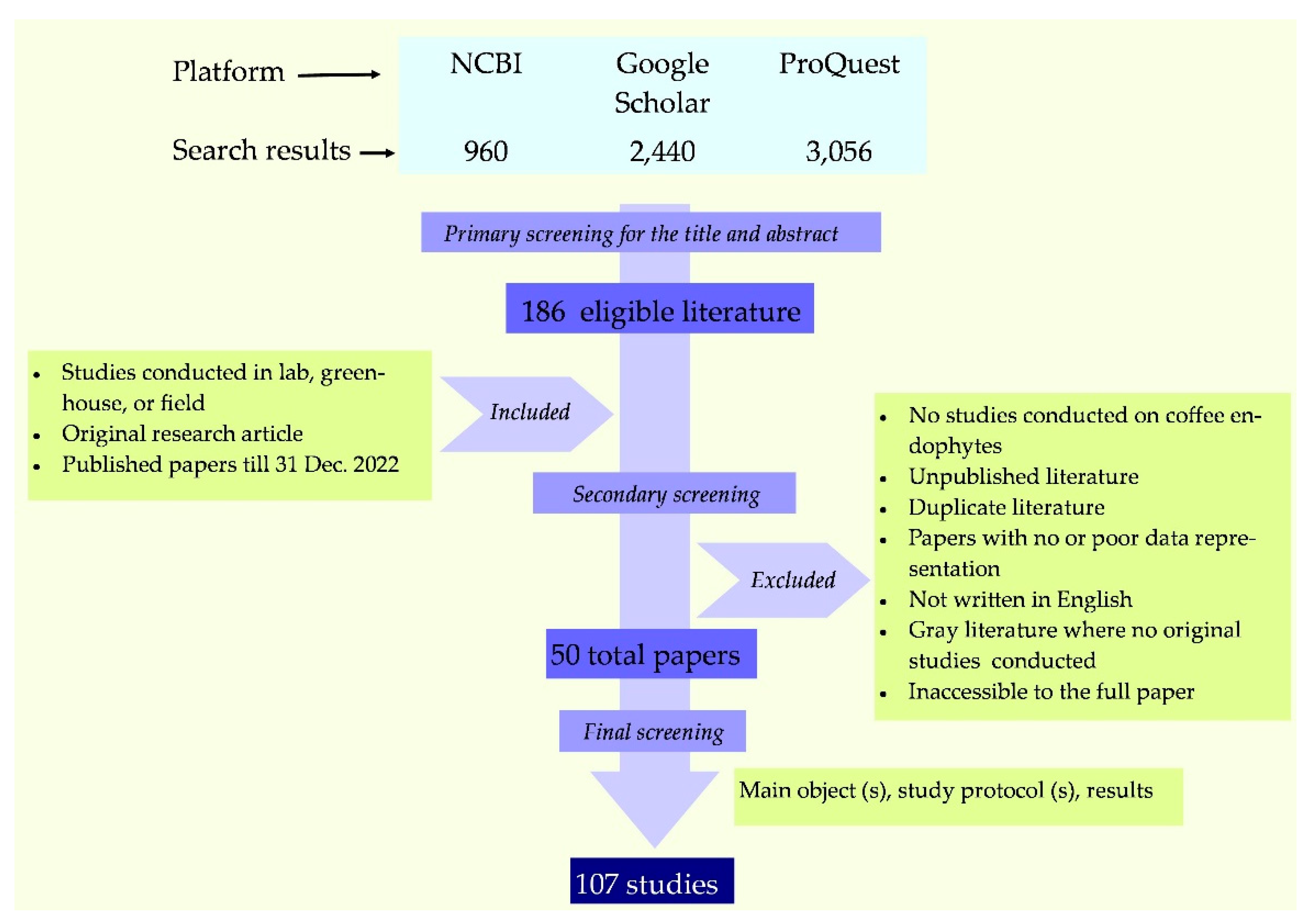
Figure 2.
Percentage (above) and number of studies (below) carried out on coffee endophytes until 2022.
Figure 2.
Percentage (above) and number of studies (below) carried out on coffee endophytes until 2022.
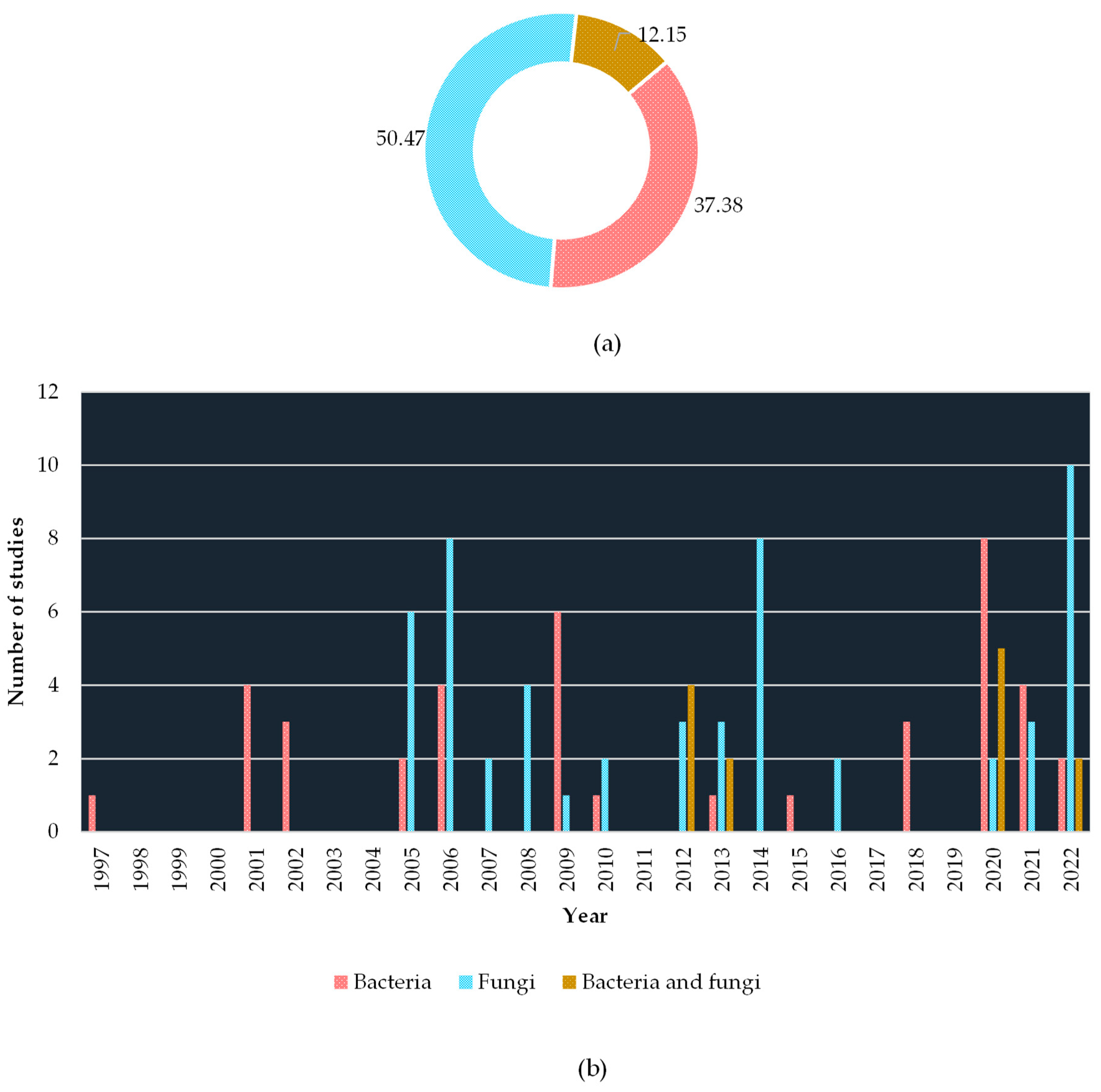
Figure 3.
Studies on coffee endophytic fungi. Different dimensions of studies were carried out throughout two and half decades. Most studies focused on isolation and identifying the endophytes associated with coffee plants, followed by biocontrol ability and diversity/population studies. While “other” studies concentrated on endophytic variation, phylogenetic origin, synthesis of mycotoxin, and organic compounds. .
Figure 3.
Studies on coffee endophytic fungi. Different dimensions of studies were carried out throughout two and half decades. Most studies focused on isolation and identifying the endophytes associated with coffee plants, followed by biocontrol ability and diversity/population studies. While “other” studies concentrated on endophytic variation, phylogenetic origin, synthesis of mycotoxin, and organic compounds. .
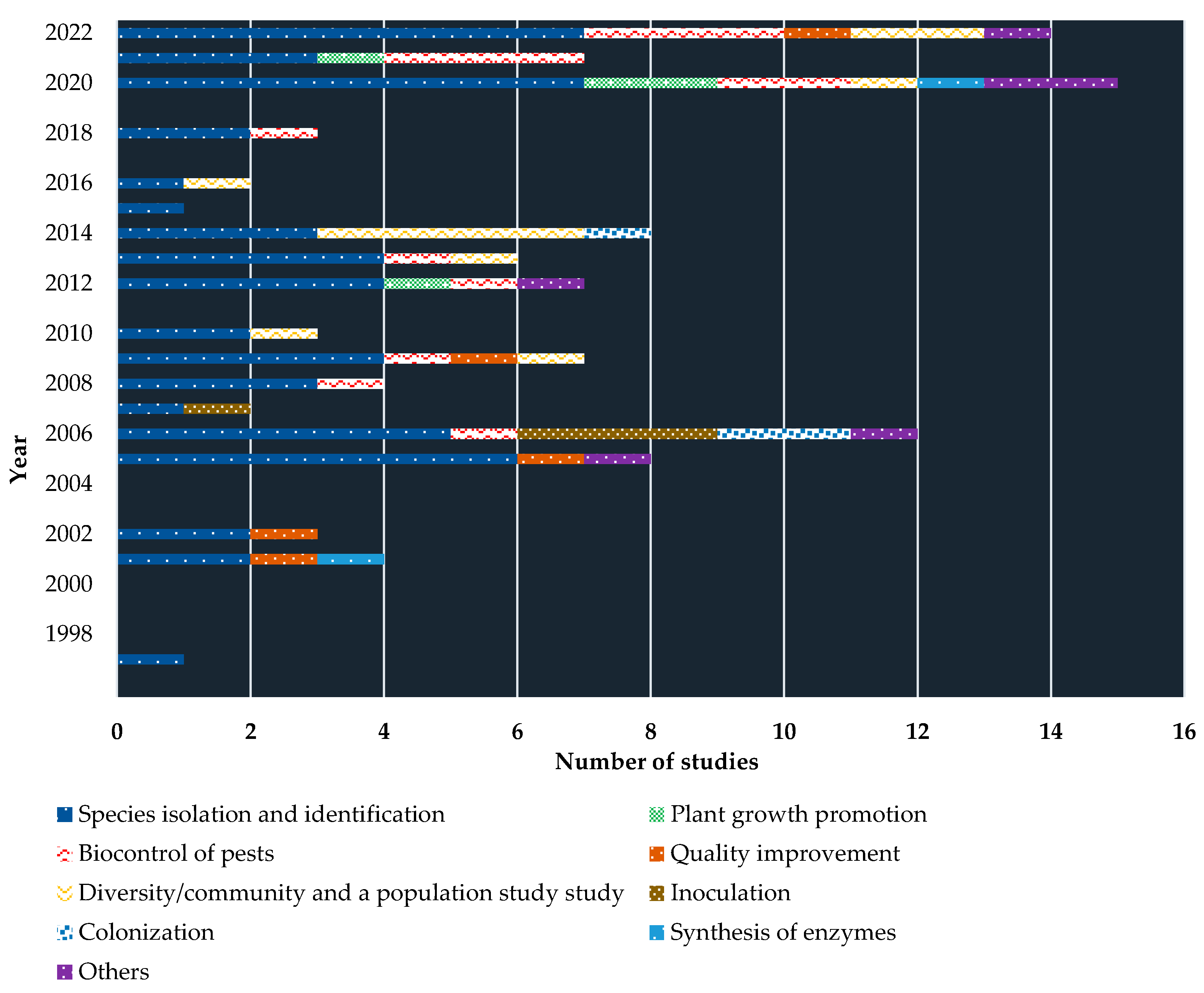
Figure 4.
Coffee plant growth improvement and protection mechanism by endophytes. (a). Highlight the enhancement of nutrition acquisition by the plant. Hyphae of fungi absorb the nutrients from the rhizosphere and through the invasion/colonization in the plant root system, they supply the carried nutrients to the plant and distribute throughout the plant. Small arrows show the nutrient movement pathway. (b). Endophytic fungi protect plants from other pathogenic fungi through mycoparasitism. (c). Some endophytic fungi trap the parasitic nematodes using constricting rings and destroy them. (d). Entomopathogenic fungi infect insect pests and spend a parasitic phase inside the living host and infection progress leads the insect to die. The fungi further colonize inside the corpse and further grow up on the insect body and spend a saprophytic lifestyle.
Figure 4.
Coffee plant growth improvement and protection mechanism by endophytes. (a). Highlight the enhancement of nutrition acquisition by the plant. Hyphae of fungi absorb the nutrients from the rhizosphere and through the invasion/colonization in the plant root system, they supply the carried nutrients to the plant and distribute throughout the plant. Small arrows show the nutrient movement pathway. (b). Endophytic fungi protect plants from other pathogenic fungi through mycoparasitism. (c). Some endophytic fungi trap the parasitic nematodes using constricting rings and destroy them. (d). Entomopathogenic fungi infect insect pests and spend a parasitic phase inside the living host and infection progress leads the insect to die. The fungi further colonize inside the corpse and further grow up on the insect body and spend a saprophytic lifestyle.
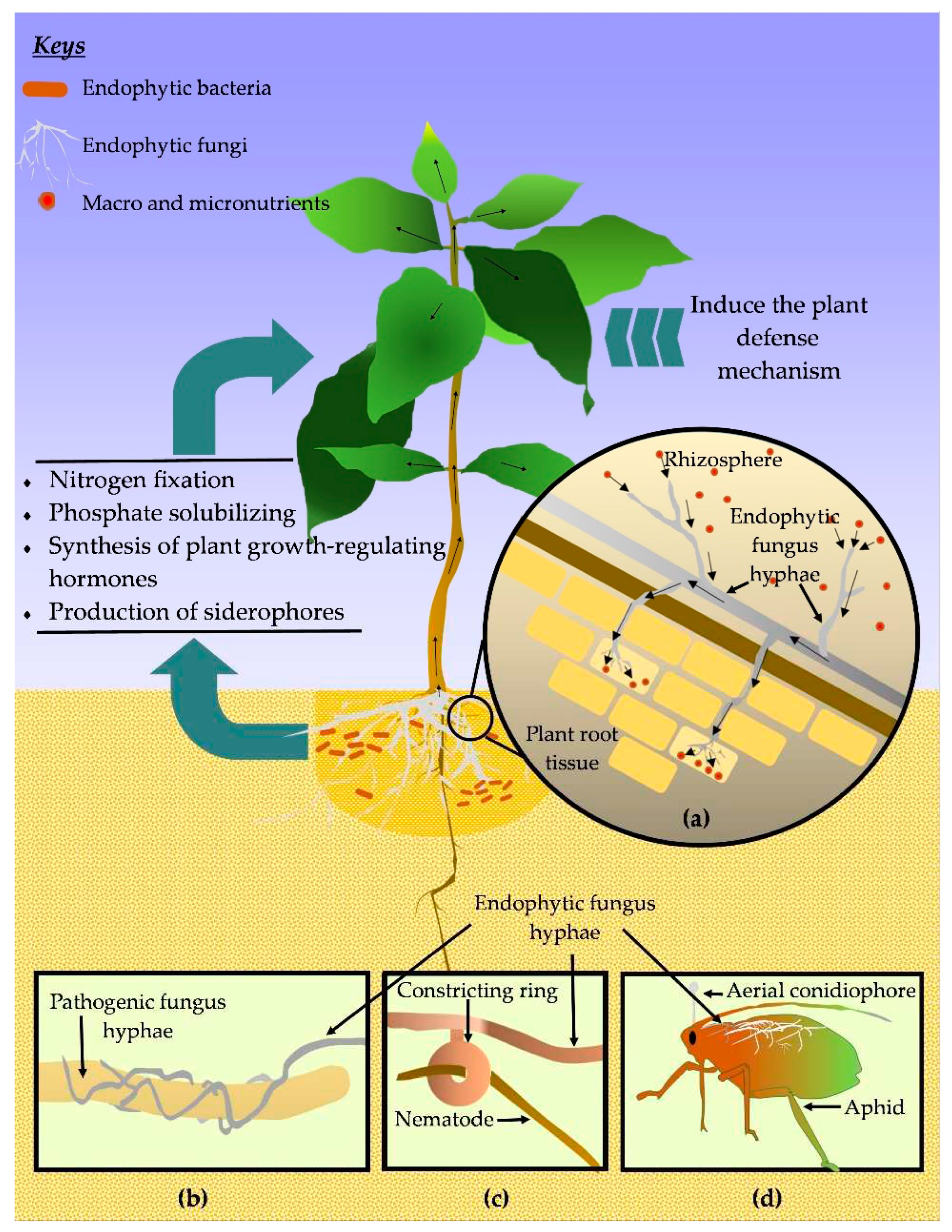
Figure 5.
Graphical representation of the data obtained from Mamani-Huayhua et al. [95], which shows the comparison of coffee plant growth parameters along with the Trichoderma sp. application. C: Untreated control, T1: Treatment 1; T2–T5: Treatment 2–5. Each treatment represents different endophytic strains. .
Figure 5.
Graphical representation of the data obtained from Mamani-Huayhua et al. [95], which shows the comparison of coffee plant growth parameters along with the Trichoderma sp. application. C: Untreated control, T1: Treatment 1; T2–T5: Treatment 2–5. Each treatment represents different endophytic strains. .
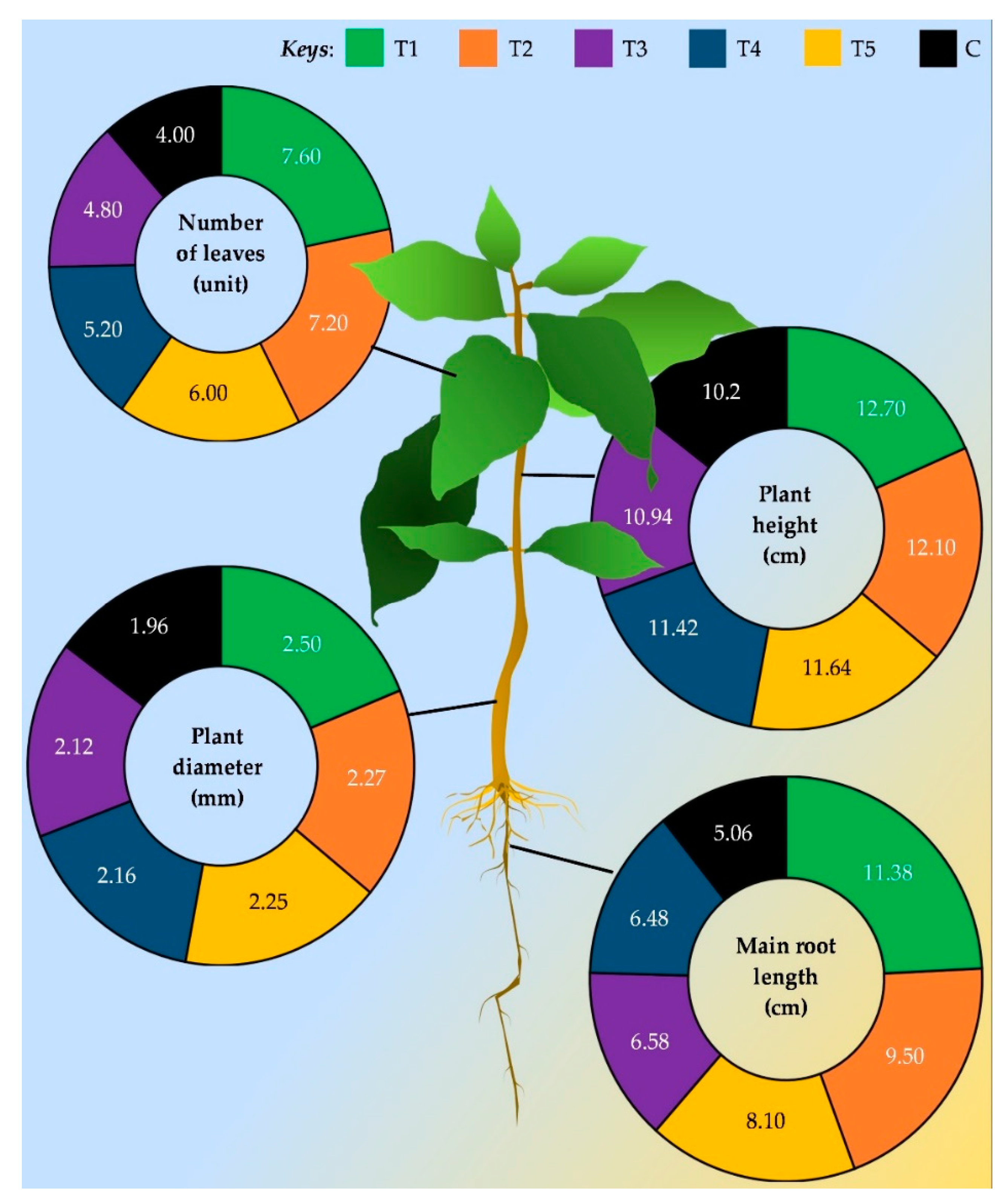

Table 1.
Some of the identified bacterial and fungal coffee endophytes.
| Phylum/Division | Species | Associated coffee species | Isolated part of the coffee plant | Locality | Reference |
|---|---|---|---|---|---|
| Bacteria | |||||
|
Actinobacteria |
Arthrobacter sp. | C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam |
[84] |
| Clavibacter michiganensis |
C.c |
LE | State of São Paulo, Brazil |
[85] | |
| SE | Cacahoatán, Chiapas, Mexico | [86] | |||
| Curtobacterium sp. | C.a | ST | Centro Nacional de Investigaciones de café, Cenicafé, Chinchin, Colombia |
[86] | |
| Curtobacterium sp. | C.c | RO | Buon Ma Thuot, Bao Loc | [84] | |
| Curtobacterium flaccumfaciens | C.a | SE and LE | Centro Nacional deInvestigaciones de café, Cenicafé, Chinchin, Colombia | [86] | |
| Gordona sp. | C.a | CH | Hawaii | [86] | |
| Kitasatospora sp. | C.c | RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Kocuria sp. | C.c | RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Kocuria kristinae | C.a | LE | State of São Paulo, Brazil |
[85] | |
| Lechevalieria sp. | C.a and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Sinomonas sp. | C.c | RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Streptomyces sp. | C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Firmicutes |
Bacillus sp. |
C.a | LE | State of São Paulo, Brazil |
[85] |
| C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | ||
| Bacillus cereus | C.c | LE | State of São Paulo, Brazil |
[85] | |
| C.a | LE | Hawaii | [86] | ||
| Bacillus lentimorbus | C.c | ST | State of São Paulo, Brazil |
[85] | |
| Bacillus megaterium | C.a | LE | Hawaii | [86] | |
| Bacillus subtilis | C.a | CR | Hawaii | [86] | |
| C.a | RO | East Java Province, Indonesia |
[87] | ||
| Lysinibacillus sp. | C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc | [84] | |
| Paenibacillus sp. | C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Paenibacillus amylolyticus. | C.a | CH | Minas Gerais, Brazil. | [57] | |
| Staphylococcus sp. | C.C and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
|
Proteobacteria |
Acinetobacter calcoaceticus | C.a | ST | State of São Paulo, Brazil |
[85] |
| Acetobacter diazotrophicus | C.a | ST and RO | Guerrero and Puebla, Mexico | [77] | |
| Burkholderia sp. | C.c | SE | Buon Ma Thuot, Vietnam | [84] | |
| Cedecea davisae | C.a | LE | State of São Paulo, Brazil |
[85] | |
| Enterobacter sp. | C.c | RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Herbaspirillum sp. | C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Klebsiella pneumonia | Cc | LE | State of São Paulo, Brazil |
[85] | |
| Luteibacter sp. | C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc | [84] | |
| Methylobacterium sp. | C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Methylobacterium radiotolerans | C.a | CH | Centro Nacional deInvestigaciones de café, Cenicafé, Chinchin, Colombia | [86] | |
| Pandoraea pnomenusa | C.a | LE | State of São Paulo, Brazil |
[85] | |
| Paracoccus sp. | C.c and C.l | SE and RO | Buon Ma Thuot, Bao Loc, Vietnam | [84] | |
| Pseudomonas sp. | C.a | ST, RO, and LE (NS) | Southern parts of India. | [88] | |
| Fungi | |||||
| Ascomycota | Acremonium alternatum | C.a | EP | Caldas, Colombia | [76] |
| Aspergillus sp. | C.a | RO | Ethiopia | [89] | |
| Aspergillus oryzae | C.a | LE | Caldas, Colombia | [89] | |
| Aureobasidium pullulans | C.a | LE | Pernambuco, Brazil | [82] | |
| Beauveria bassiana | C.a | SE, EP, EP, and CR | Caldas, Colombia | [76] | |
| Cercospora iranica | C.a | LE | Paraná, Brazil. | [79] | |
| Cercospora tezpurensis | C.a | LE | Paraná, Brazil. | [79] | |
| Cladosporium sp. | C.a | RO | Ethiopia | [89] | |
| Cladosporium cladosporioides | C.a | LE | Adjuntas, Puerto Rico and Kona, Hawaii, USA | [76] | |
|
Cladosporium pini-ponderosae |
C.a | LE | Paraná, Brazil. | [79] | |
| Cladosporium sphaerospermum | C.a | LE | Beltsville, Maryland, USA | [76] | |
| Cladosporium tenuissimum | C.a | LE | Pernambuco, Brazil | [82] | |
| Clonostachys rosea | C.a | LE | Chinchina, Caldas, Colombia | [76] | |
| Cladosporium cladosporioides | C.a | LE | Pernambuco, Brazil | [82] | |
| Colletotrichum asianum | C.a | CH | Chiang Mai, Thailand | [91] | |
| Colletotrichum brassicicola | C.a | LE | Veracruz, Mexico | [81] | |
| Colletotrichum coffeanum | C.a | LE | Pernambuco, Brazil | [82] | |
| Colletotrichum endophytica | C.a | LE | Paraná, Brazil. | [79] | |
| Colletotrichum fructicola | C.a | CH | Chiang Mai, Thailand | [91] | |
| Colletotrichum gloeosporioides | C.a | LE | Veracruz, Mexico | [81] | |
| Colletotrichum gloeosporioides | C.a | LE | Pernambuco, Brazil | [82] | |
| Colletotrichum musae | C.a | LE | Veracruz, Mexico | [81] | |
| Colletotrichum queenslandicum | C.a | LE | Paraná, Brazil. | [79] | |
| Colletotrichum siamense | C.a | CH | Chiang Mai, Thailand | [91] | |
|
Cryptosporiopsis corticola |
C.a | LE | Veracruz, Mexico | [81] | |
| Diaporthe liquidambaris | C.a | LE | Pernambuco, Brazil | [82] | |
|
Dipodascaceae sp. |
C.a | RO | Ethiopia | [89] | |
| Drechslera biseptata | C.a | LE | Pernambuco, Brazil | [82] | |
| Fusarium equiseti | C.a | RO | Ethiopia | [89] | |
| Fusarium oxysporum | C.a | RO | Ethiopia | [89] | |
| Fusarium solani | C.a | RO | Ethiopia | [89] | |
| Glomerella cingulata | C.a | LE | Veracruz, Mexico | [81] | |
| Guignardia mangiferae | C.a | LE | Veracruz, Mexico | [81] | |
| Induratia coffeana | C.a | NS | Minas Gerais, Brazil | [92] | |
| Induratia yucatanensis | C.a | NS | Minas Gerais, Brazil | [92] | |
| Khuskia oryzae | C.a | LE | Pernambuco, Brazil | [82] | |
| Lasiodiplodia pseudotheobromae | C.a | LE | Pernambuco, Brazil | [82] | |
| Macrophomina sp. | C.a | RO | Ethiopia | [89] | |
| Muscodor coffeanum* | C.a | LE | Minas Gerais, Brazil | [93] | |
| Muscodor vitigenus* | C.a | ST | Minas Gerais, Brazil | [93] | |
| Muscodor yucatanensis* | C.a | LE | Minas Gerais, Brazil | [93] | |
| Mycosphaerella pseudovespa | C.a | LE | Paraná, Brazil. | [79] | |
| Nodulisporium gregarium | C.a | LE | Pernambuco, Brazil | [82] | |
|
Ophioceras leptosporum |
C.a | LE | Paraná, Brazil. | [79] | |
| Ophiognomonia sp. | C.a | LE | Paraná, Brazil. | [79] | |
| Paecilomyces fumosoroseus | C.a | CR | Adjuntas, Puerto Rico | [76] | |
| Paecilomyces javanicus | C.a | PE | Chinchina, caldas, Colombia | [76] | |
| Paenibacillus amylolyticus | C.a | CH | Minas Gerais, Brazil. | [57] | |
| Penicillium sp. | C.a | RO | Ethiopia | [89] | |
| Penicillium coffeae | C.a | NS | Kunia, Hawaii |
[94] | |
| Pestalotiopsis maculans | C.a | LE | Pernambuco, Brazil | [82] | |
| Phoma eupyrena | C.a | LE | Pernambuco, Brazil | [82] | |
| Phomopsis sp. | C.a | RO | Ethiopia | [89] | |
| Phomopsis arnoldiae | C.a | LE | Veracruz, Mexico | [81] | |
| Phyllosticta sp. | C.a | LE | Pernambuco, Brazil | [82] | |
|
Phyllosticta capitalensis (=Guignardia mangiferae) |
C.a | LE | Pernambuco, Brazil | [82] | |
|
Pleosporales sp. |
C.a | RO | Ethiopia | [89] | |
| Sarocladium bacillisporum | C.a | LE | Pernambuco, Brazil | [82] | |
|
Simplicillium sp. |
C.a | ST | Minas Gerais, Brazil |
[93] | |
| Trichoderma sp. | C.a | LE and ST | San Juan del Oro, Peru |
[95] | |
| Trichoderma flagellatum | C.a | RO | Ethiopia | [89] | |
| Trichoderma hamatum | C.a | RO | Ethiopia | [89] | |
| Trichoderma neokoningii | C.a | LE | Paraná, Brazil. | [79] | |
| Xylaria spp. | C.a | LE | Veracruz, Mexico | [81] | |
| Xylaria sp. | C.a | LE | Pernambuco, Brazil | [82] | |
| Basidiomycota | |||||
| Rhodotorula acheniorum | C.a | LE | Pernambuco, Brazil | [96] | |
| Rhodotorula aurantiaca | C.a | LE | Pernambuco, Brazil | [96] | |
| Rhodotorula aurantiaca | C.a | LE | Pernambuco, Brazil | [82] | |
| Rhodotorula glutinis | C.a | LE | Pernambuco, Brazil | [96] | |
| Rhodotorula mucilaginosa | C.a | LE | Pernambuco, Brazil | [96] | |
| Schizophyllum commune | C.a | LE | Paraná, Brazil. | [79] | |
| Trametes sp. | C.a | RO | Ethiopia | [89] | |
C.a: Coffea arabica; C.c: Coffea canephora; C.l: Coffea liberica; CH: Cherrie; CR: Crown; EP: Epicarp; LE: Leaves; NS: Not specified; PE: Peduncle; RO: Roots; SE: Seeds; ST: Stems, * Muscodor now transferred to Induratia genus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



