
Submitted:
07 May 2023
Posted:
08 May 2023
You are already at the latest version
Abstract
Nutrigenomics is the study of the interaction of nutrition and genes, focusing on the influence of nutrients on the genome, proteome, and metabolome, and how nutrition affects human health. In the context of nutrigenomics, bioactive components are dietary ingredients that may transmit information from the external environment and alter gene expression in the cell, and hence the overall function of the organism. It is critical to consider food not only as a source of energy and essential nutrients necessary for life and growth, but also as a factor impacting health/disease, biochemical processes, biochemical pathway activation and affecting the diversity of the gut microbiome. Antimicrobial resistance in pathogenic and commensal microorganisms has emerged as a major public health concern due to emerging antimicrobial resistance genes in E. coli isolates from pig, cattle, chicken, and turkey meat, against tetracycline, streptomycin, and sulfonamides. Also, Salmonella spp. and Campylobacter spp. have shown antibiotic resistance at farms and slaughterhouses, and in animal-based food products. A correlation has been proven between a critical nutrient-responsive signaling system and catabolite control of gene expression, and a two-component signaling system that drives antibiotic resistance in E. faecalis, revealing a previously unknown integration between the nutritional status of the cell and intrinsic antibiotic resistance. Moreover, different nutrigenomic approaches can be applied to mitigate possible emergence of antimicrobial resistance against novel antibiotics. However, little progress has been achieved in converting nutrigenomics information into clinical advice, so far.
Keywords:
Nutrigenomics
; Antimicrobial resistance
; Novel Antibiotics
; gut microbiome
Introduction
Nutrigenomics is the study of the interactions between nutrition and genes. Nutrition plays an essential role in life since macro- and micronutrients are essential building blocks of sustaining life [1]. The pathways for nutrient-gene communication routes have existed through the evolution of life [2].
Furthermore, because some nutrients directly affect the genome or the epigenome through interaction with transcription factors or chromatin modifiers, they have a health impact [3]. An association between nutrients and genes has been in population-based studies. Each nutrient has its own set of target ligands, in contrast to distinguish the pharmacological properties of drug product we use drug molecule at low concentrations to discriminate its ability to target a limited number of biological targets. [4]
Overall, a nutrigenomics approach can provide a snapshot on genes, which are turned on/off at any given time (the genetic potential), a view of how gene/protein networks may interact to produce the observed response, and a method to determine the influence of nutrients on gene/protein expression. Increasing research in this field should lead to a better understanding of how nutrition affects metabolic pathways and homeostasis regulation, which may then be used for the prevention of chronic diet-related disorders [5].
One of the first studies to use transcriptomics technology for human dietary interventions assessed the effects of a high-carbohydrate breakfast compared to a high-protein meal on the gene expression profile in blood leukocytes of healthy males [6]. Breakfasts high in carbohydrates resulted in differential expression of those genes mostly engaged in glycogen metabolism, whereas breakfasts high in protein content led to differential expression of genes primarily involved in protein production [6].
A study was conducted to determine the function of parental folate pathway Single Nucleotide Polymorphisms (SNPs) in changing the susceptibility to neural tube defects (NTDs) in 50 couples with neural tube defect children and 80 couples with normal offspring. The investigators found a high frequency of NTDs in South India, where consanguinity and vitamin deficiencies are frequent, indicating the importance of genetic and dietary variables as likely etiological factors. Particularly, SNPs in folate pathways have been associated with NTDs It was confirmed that the maternal methylene tetrahydrofolate reductase (MTHFR) C77T SNP and parental glutamate carboxypeptidase (GCP) II C1561T were linked to an increased risk of NTDs [7].
The effects of chronic feeding of various diets comprising vanaspati (rich in trans fatty acids (TFAs)), palm oil (rich in saturated fatty acids (SFAs)), and sunflower oil (rich in polyunsaturated fatty acids (PUFAs)) at 10% level of all dietary composition on 11β-hydroxysteroid dehydrogenase type 1 enzyme (11β-HSD1) gene expression in rat retroperitoneal adipose tissue have been examined. When compared to a PUFA-enriched diet, meals high in TFAs and SFAs showed superior 11β-HSD1 gene expression in rat Retroperitoneal White Adipose Tissue (RPWAT). The risk of developing obesity and insulin resistance increased as a result of the enhanced local conversion of inactive to active glucocorticoids in adipose tissue [8].
Colistin (polymyxin E) is a polymyxin antibiotic extensively used in animal health for oral treatment of enterobacterial digestive infections in pigs, poultry, and cattle [9]. Previous studies utilizing culture-dependent techniques on farm animals in Tunisia revealed the prevalence of colistin-resistant Gram-negative bacteria. In a recent investigational study, DNA analysis for ten known mcr genes was conducted on cloacal swabs from 195 broiler chickens from six farms in Tunisia [10]. Eighty-one (41.5%) of the 195 animals tested positive for mcr-1 with positive cases on all farms. These findings corroborate the emergence of colistin resistance in farm animals in Tunisia and suggest that investigating antibiotic resistance genes (ARGs) might contribute to epidemiological research on antimicrobial resistance dissemination [10].
A nutrigenomics study showed the positive effects of phytobiotics and organic acids on ghrelin gene expression levels, gut microbiota composition, performance metrics, and intestinal histomorphological alterations in broiler chicks compared to antibiotics [11]. The addition of phytobiotics enhanced (p < 0.05) villus height and the ratio of villus height/crypt depth in the ileum, jejunum, and duodenum, while decreasing ghrelin gene expression levels. Total coliform and E. coli levels in cecal and ileal digesta were considerably lower (p < 0.05) compared to antibiotic treatment. Lactobacillus spp. were shown to be favourably associated with villus height/crypt depth ratio in the duodenum by correlation analysis. Based on the nutrigenomics methodology, the findings highlighted the significance of gene-nutrient-microbiota relationships. As a result, phytobiotics and organic acids may be viable alternatives to antibiotics for increased performance and immunity in chicken, as well as healthier meat production. [11] Moreover, phytobiotics will contribute to decreased antibiotics use and prevention of the potential emergence of microorganisms resistant to antibiotics.
Metagenomics and Illumina sequencing approaches have been employed to investigate the content and dynamic distribution of ARGs, as well as the microbial population, in three types of factory-processed Chinese garlic powder (GP) products [12]. The findings revealed that 126 ARG genes from 11 different ARG species were detected. With the processing of GP, the expression of ARGs increased at first and later decreased [12]. Since garlic is added to food, this finding raises concern of a possible emergence of antibiotic resistant strains from dietary supplies.
2. Materials and Methods
In the present review, the following sources were included: randomized controlled trials (RCTs), controlled non-randomized clinical trials (CCTs), retrospective and prospective comparative cohort studies, case control or nested case-control studies, reviews, systematic reviews, and thematic books.
A search strategy was designed using medical subject headings (MeSH). The MeSH terms of supplements, nutrition, nutrigenomics, gut microbiome, and antimicrobial resistance were used to systemically search PubMed, and MEDLINE databases. Only studies in the English language were included. All relevant publications up to April 2023, were included. No limits regarding study design or date were set for the search. Duplicate studies were removed from our study pool. All included studies were scanned against inclusion and exclusion criteria. Our inclusion criteria primarily focused on published literature that assessed the effect on influence of nutrients on genome and its correlation with emerging antimicrobial resistance for new antibacterial agents.
3. Results and Discussion
3.1. The effect of nutrition on human gut microbiome
Trillions of bacteria occupy the human gut, forming a dynamic ecological system involved in both health and disease. The gut microbiota composition is unique to each individual and tends to be generally constant throughout life, but daily transitory changes are noted. Diet is a fundamental modifiable element determining the makeup of the gut microbiota, implying that therapeutic dietary techniques to control microbial diversity, composition, and stability are possible. While food can cause changes in the gut microbiota, these changes appear to be transient. It is uncertain if long-term dietary changes may cause lasting changes to the gut microbiota, owing to a dearth of long-term human nutritional interventions or long-term follow-ups of short-term dietary treatments [13].
Aside from nutrition, the gut microbiota is influenced by a mix of extrinsic factors such as lifestyle and medicine, and intrinsic factors such as host genetics, immunological and metabolic control (Figure 1). It is commonly known that external variables show the greatest influence, with nutrition being the most extensively studied variable [14]. On the other hand, according to a 2016 data study of 1126 twins, genetics play a modest 8.8% average effect in determining the gut flora [15].
The human diet normally comprises many food types such as protein, fat, and carbohydrates, with a proper required balance. The community and diversity of the gut microbiome show a large impact on the kind and quantity of nutrition. The digestion of food elements results in variation in the end products, which play an important role in the prevention, management, and treatment of diseases like cancer and diabetes [16,17].
Moreover, the flourishing or fading of the beneficial microbiome may result in increased metabolites in a healthy direction or the appearance of opportunistic genera. Production of certain metabolites may result in a pathogenic appearance that can influence host physiology and gene expression, resulting in the development of various diseases. [18]
A fat-containing diet alters the gut microbial composition, usually resulting in a drop in Bacteroidetes and an increase in both Firmicutes and Proteobacteria [19]. Furthermore, certain genera of the class Gammaproteobacteria increased their abundance in comparison to other particular genera, resulting in a change in the microbial population and diversity. [20]
These alterations reduce the microbial synthesis of short-chain fatty acids and antioxidants. Changes in metabolites have consequences such as increased disease risk. Carbohydrates have been proven to boost cell survival in cancer by increased expression of genes related with fatty mass and obesity. [21]
Diets high in complex carbohydrates generated from plant tissue are slowly digested by the gut bacteria, particularly those found in the distal intestine. Plant fiber digestion enhances the symbiotic microbiota, which leads to an increase in short-chain fatty acids, which have a role in energy supply and hence in human health. Complex carbohydrates improve body weight, food intake, glucose homeostasis, and insulin sensitivity.[22].
Several studies have found a link between a higher fiber diet and a lower risk of irritable bowel syndrome, inflammatory bowel disease, cardiovascular disease, diabetes, and colon cancer [23].
3.2. The effect of nutrition on human genome expression
In the context of nutrigenetics and nutrigenomics, bioactive components are dietary ingredients that may transmit information from the external environment and alter gene expression in the cell, and hence affect the overall function of the organism. It is critical to consider nutrition not only as a source of energy and essential nutrients necessary for life and organism growth, but also as a factor impacting biochemical processes, biochemical pathway activation, and health/disease. Dietary bioactive components influence gene expression by chromatin structural modifications, activation of transcription factors via signalling cascades, or direct ligand binding to nuclear receptors [24].
Individual phenotypes can be altered by nutrient-induced gene expression. SNPs in a variety of genes involved in inflammation and lipid metabolism, on the other hand, can modify the bioactivity of essential metabolic pathways and mediators, as well as the capacity of nutrients to interact with these mediators and metabolic pathways [25].
Gene expression in human blood and other tissues has been demonstrated to be dependent on gender, age, and time of the day. However, additional variables that may influence gene expression have not been well investigated. For example, it is unclear whether the fasting or fed condition will provide a clear answer related to the association between gene expression in the blood and obesity [26].
Several recent studies have demonstrated the adaptability of biological networks by revealing rapid network rewiring in response to various environmental challenges. Fasting and feeding responses in rats have been studied with some success [27]. Other studies have demonstrated that circadian clock genes in mouse heart tissue are controlled differentially during food intake and fasting [28].
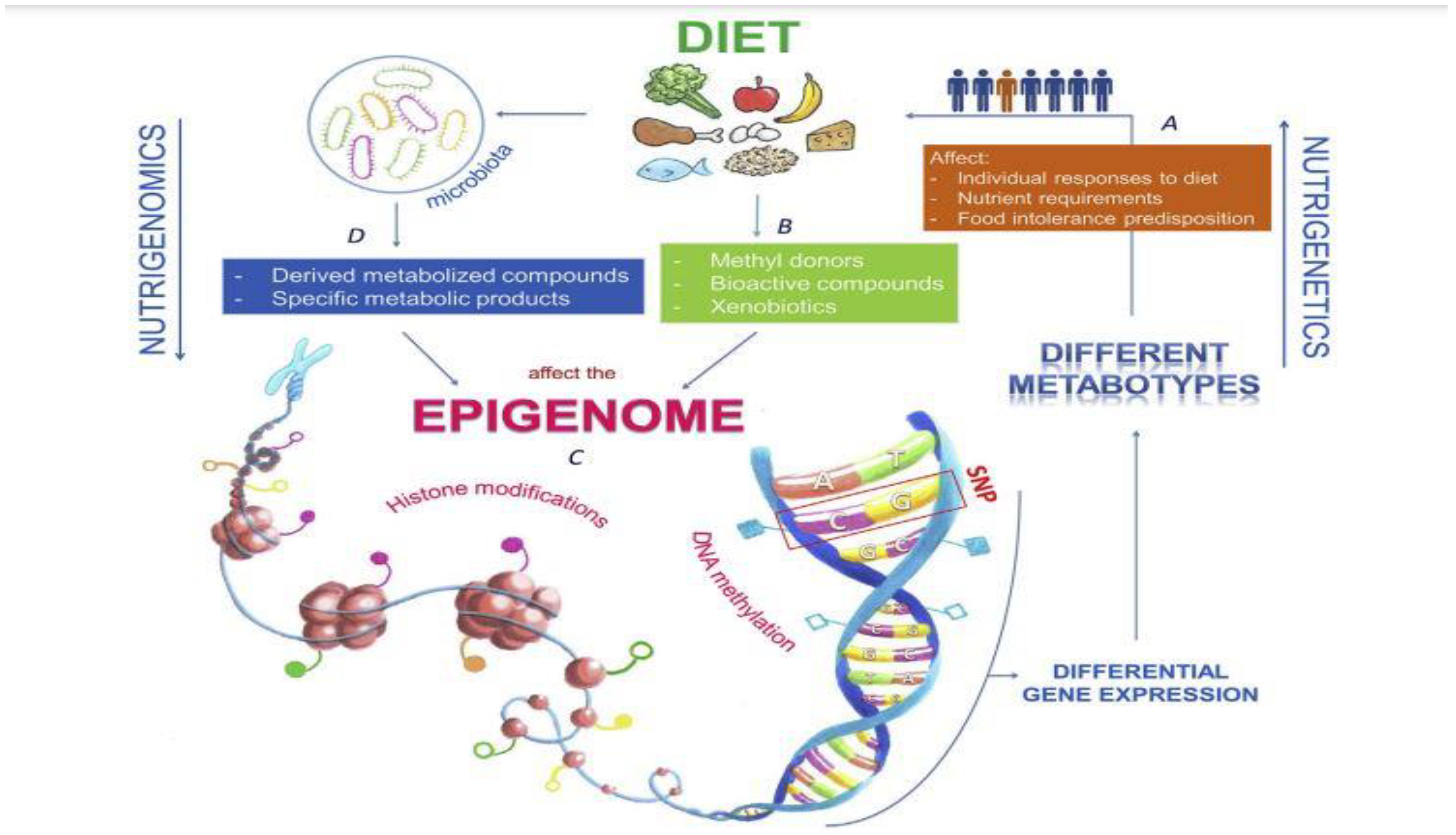
Figure 1.
Illustration of the relationships between nutrition and the genome [29]. (A)Nutrigenetics: genetic polymorphisms can induce differential gene expression. As a result, different metabotypes exist, which show different responses to nutrition, different nutrient requirements and potential food intolerance. Of note, the location of SNPs can also affect epigenetic modifications. (B-C-D) Nutrigenomics: methyl donor availability, bioactivity of dietary compound and xenobiotics (B) can affect the one-carbon cycle and other pathways, thus consequentially, affect DNA methylation and histone modifications (C). Not just parental molecules (B) but also derived compounds and metabolic products of microbial activity (D) can affect these pathways (C).
Figure 1.
Illustration of the relationships between nutrition and the genome [29]. (A)Nutrigenetics: genetic polymorphisms can induce differential gene expression. As a result, different metabotypes exist, which show different responses to nutrition, different nutrient requirements and potential food intolerance. Of note, the location of SNPs can also affect epigenetic modifications. (B-C-D) Nutrigenomics: methyl donor availability, bioactivity of dietary compound and xenobiotics (B) can affect the one-carbon cycle and other pathways, thus consequentially, affect DNA methylation and histone modifications (C). Not just parental molecules (B) but also derived compounds and metabolic products of microbial activity (D) can affect these pathways (C).
3.3. Antimicrobial resistance mechanisms
Antimicrobial resistance (AMR) is defined as the ability of bacteria to live and thrive in the presence of antimicrobial substances. Antimicrobial agents, such as antibiotics, disinfectants, and food preservatives, are available and can be employed against microorganisms to restrict their ability to grow, prevent their reproduction, or even kill them. Natural, semi-synthetic, and synthetic substances with distinct mechanisms capable of causing major changes on the metabolic and physiological levels of bacteria include cell wall synthesis modifications such as β-lactams and glycopeptides, protein synthesis inhibitors such as macrolides and tetracyclines, metabolic pathway inhibitors such as sulfonamides, and interference of DNA replication and translation such as fluoroquinolones [30].
Antimicrobial resistance is a dramatic natural phenomenon that arises spontaneously over time through genetic change inside bacteria. However, variables like drug usage and misuse speed up this transformation. For example, administering incorrect antibiotics for viral infections such as flu might contribute to the resistance process. It poses a comparable hazard to animals and food production sustainability. [30]
Antimicrobial resistance mechanisms are classified into four types: restriction of drug uptake; alteration of drug targets; inactivation of drugs; and active drug efflux (Figure 2). Intrinsic resistance mechanisms include drug target alterations, drug inactivation, and drug efflux; acquired resistance mechanisms include drug target modifications, drug inactivation, and drug efflux. Because of structural changes, etc., the processes utilized by gram-negative bacteria differ from those used by gram-positive bacteria [31].
3.4. Antimicrobial effect on human gut microbiome
The gut microbiota is vital for host health and is influenced by a variety of variables, including antibiotics. Antibiotic-induced changes in gut microbial composition can have a negative impact on health by reducing microbial diversity, changing functional attributes of the microbiota, and forming and selecting antibiotic resistant strains, and increasing the susceptibility to infection with pathogens like Clostridioides difficile [32].
Recently, scientists discovered that broad spectrum antibiotics had a negative influence on the gut microbiome [33]. The gut microbiota, which is home to bacteria, archaea, microeukaryotes, and viruses, plays an important role in human health. It inhibits pathogen colonization, modulates gut immunity, supplies necessary nutrients and bioactive metabolites, and aids in energy balance [33].
A recent study found that taking antibiotics orally is linked to an increased risk of colon cancer [34]. For example, extremely preterm children subjected to continuous antibiotic therapy had fewer varied bacterial communities, lower species richness, and higher ARGs in their gut [35,36].
Antibiotics can cause antibiotic associated diarrhea (AAD), and studies have shown that clindamycin might modify the microbial population, promoting the colonization of potential pathogens such as C. difficile, resulting in diarrhea and colitis [37]. The use of antibiotics has also been linked to changes in protein expression and energy consumption in the microbiota, with a minor increase following antibiotic therapy, possibly as a coping strategy to antibiotic stress, but decreasing at later stages and after exposure to antibiotics [38].
Antibiotics can affect the transcription of numerous important functional genes, including those encoding transport proteins, glucose metabolism genes, and protein synthesis genes. Exposure of Pseudomonas aeruginosa to sub inhibitory antibiotic doses elevated the expression of virulence associated genes, resulting in increased production of rhamnolipids and phenazines. Numerous investigations have indicated that aminoglycosides, lactams, vancomycin, and oxacillin may all promote biofilm development at sublethal doses [32]. These biofilms then serve as antibiotic resistance reservoirs. It provides further resistance to bacteria against various antibiotics and host defense, making treatment difficult in people and causing a variety of complications such as pipe/equipment obstruction in healthcare settings and food sectors [39].
Metabolomics profiles were examined in antibiotic-treated pigs from postnatal days 7 to 42, subjected to a corn-soy baseline diet with or without infeed antibiotics [40]. The antibiotic-treated group exhibited increased amounts of metabolites linked with amino acid metabolism, resulting in lower amino acid concentrations. Short-chain fatty acid production was also reduced as butyrate and propionate levels declined [40].
3.5. Phylogenetic Groups and Antimicrobial Resistance Genes from poultry
There is little information on the occurrence of metal and antibiotic resistance in potentially pathogenic E. coli entering the food chain from pigs, which might endanger human health. The phenotypic and genotypic resistances of E. coli to 18 antibiotics and three metals (mercury, silver, and copper) at pig slaughterhouses in the United Kingdom has been investigated, which revealed resistance to streptomycin, sulphonamide, oxytetracycline, ampicillin, chloramphenicol, ceftiofur, amoxicillin-clavulanic acid, trimethoprim-sulfamethoxazole, aztreonam, and nitrofurantoin [41].
E. coli isolated from meat originating from different animal species may have ARGs and therefore pose a risk to human health. To identify antimicrobial resistance genes in E. coli isolates from pig, cattle, chicken, and turkey meat, and to determine if resistance genotypes are related to phylogenetic groupings or meat species, a normal culture procedure was applied including 313 isolated E. coli samples. Resistance genes could be detected by PCR in 98% of resistant isolates. The tetracycline resistance genes tet(A) and tet(B), the streptomycin resistance genes strA and aadA1, the sulphonamide resistance genes sulI and sulII, the kanamycin resistance genes dfr and aphA, and the ampicillin resistance blaTEM have been identified. One stx1 positive E. coli isolate recovered from pig carried the tet(A) gene and belonged to the phylogenetic group B2, whereas another stx1 positive strain isolated from cattle was multi-resistant and tested positive for blaTEM, aphA, strA-B, sulII, and tet(A) and belonged to the phylogenetic group A. Most E. coli populations with diverse resistance genes to a single antibiotic showed statistically significant differences in MIC values [42].
The genomic backbone and plasmid correlations with antimicrobial resistance were investigated [43]. A total of 72 pathogenic avian E. coli (APEC) strains were studied. Isolates that were resistant to tetracycline and trimethoprim-sulfamethoxazole (87.5% each) and harbored blaTEM (61.1%) dominated. Furthermore, phylogroup D was the most common in total 23.6% and among multidrug-resistant (MDR) isolates (14/63). The results indicated that group D strains have a high capacity to host a wide range of plasmids (Inc-types) harboring various AMR genes. This means that phylogroup D might provide a problem in dealing with antimicrobial resistance in poultry [43].
3.6. New antibiotics against microbial resistance
Antibiotic resistance and growth of multidrug-resistant bacterial strains have now become extremely widespread in hospitals and pose a threat to worldwide infectious disease control. Possible antibiotic resistance approaches are now examined and different mechanisms for antibiotic resistance of certain previously beneficial antibiotics are under investigation [44].
If an active substance does not exhibit cross-resistance to current antibiotics, it is considered novel. Cross-resistance is defined in this context as resistance to the same class of antibiotics that may be assessed by systematic, in vitro sensitivity testing to genetically determined bacteria. If sufficient information on cross-resistance is lacking or unavailable, an active substance is considered innovative if it belongs to a new class of antibiotics, has a novel target or binding site, or demonstrates a novel mechanism of action [44].
Since 2017, eight novel antibacterial active compounds have been authorized, including one for the treatment of tuberculosis (TB). The non-profit organization TB Alliance created pretomanid, a therapeutic compound for the treatment of multidrug-resistant TB. Approximately half of the new antibiotics licensed target carbapenem-resistant Enterobacteriaceae (CRE), oxacillinase-48-producing Enterobacteriaceae (OXA-48), and β-lactamase-producing Enterobacteriaceae (ESBL). Despite significant advances in TB and C. difficile research, antibiotics are still ineffective to treat carbapenem-resistant Acinetobacter baumannii and Pseudomonas aeruginosa [45].
Since 2019, the new chemical compound zoliflodacin is in Phase III for the treatment of multidrug-resistant Neisseria gonorrhoeae [46]. It is the first antibiotic of the spiropyrimidinetrion class to be synthesized. Its novel action comprises inhibition of type II bacterial topoisomerase targeting a different location than fluoroquinolones [47].
Zoliflodacin has a very low resistance frequency and is efficacious not only against multidrug-resistant N. gonorrhoeae with MICs ranging from 0.002 to 0.25 μg/mL but also against a variety of Gram-positive and Gram-negative bacteria [42]. Even when bacteria are subjected to combinations of zoliflodacin and antibiotics currently in use, such as ceftriaxone, doxycycline, and gentamycin, no resistant mutations of N. gonorrhoeae have been detected. Zoliflodacin does not exhibit cross-resistance with currently available fluoroquinolones. [46]
Ridinilazole is a synthetic fast acting antibiotic from the bis-benzimidazole family for the treatment of C. difficile [48]. It is poorly absorbed by the gastrointestinal lumen after oral administration. Ridinilazole is virtually perfect for the treatment of CDI as it showed in vitro selective efficacy against C. difficile, minimal systemic absorption, and decreased impact on gut microbiota. Ridinilazole-resistant strains have not been identified. The findings of the action on the microbiota are quite encouraging, as the microflora remained essentially intact following ridinilazole treatment. In contrast, the most recent fidaxomycin therapy showed altered gut microbiota, and vancomycin therapy resulted in a significant drop in Bifidobacteria [44].
Delafloxacin, a new antibacterial DNA and topoisomerase IV inhibitor fluoroquinolone hows increased activity in acidic medium [49]. It possesses comparable affinity to both DNA gyrase and topoisomerase IV in Gram-positive (Staphylococcus aureus) and Gram-negative (E. coli) bacteria limits the the potential antibiotic resistance, requiring the accumulation of numerous mutations in both enzyme genes [44].
Eravacycline is a completely synthetic fluorocycline of the tetracycline family that was engineered to treat complicated intra-abdominal infections (cIAI) and to primarily combat acquired resistance of regular tetracyclines [50]. The acquisition of genes encoding certain efflux pumps and the presence of ribosomal protection proteins (RPPs) are the two key mechanisms causing pathogen resistance to tetracyclines. [51].
Various efflux pumps have been found in Gram-positive and Gram-negative bacteria. The most common efflux pumps are encoded by the tet(A) and tet(B) genes in Gram-negative bacteria and the tet(K) and tet(L) genes in Gram-positive bacteria [45]. Eravacycline has the same pharmacophore as tetracyclines, but it has two distinct modifications in ring D at positions C7 (addition of a fluorine atom) and C9 (addition of a pyrrolidine acetamide group) [52].
The modifications at positions C7 and C9 render eravacycline efficacy against Gram-positive and Gram-negative bacterial strains otherwise resistant to first- and second-generation tetracyclines. Eravacycline, like other tetracyclines, inhibits the entrance of molecules from the aminoacyl-tRNA complex by reversibly binding to the ribosomal 30S subunit. However, compared to typical tetracyclines, the interaction between eravacycline and ribosomes is significantly stronger due to the recognition of several target sites. The first- and second-generation compounds are bacteriostatic; however, eravacycline possesses in vitro bactericidal activity against select strains of A. baumannii, E. coli, and K. pneumoniae [44].
Plazomicin is a novel aminoglycoside derived from a modified sisomicin (a particular antibiotic against Gram-negative infections for which gentamicin, the first-choice molecule, was ineffective) [53]. Plazomicin inhibits most of the Aminoglycoside Modifying Enzymes (AME) that inactivate aminoglycosidic drugs in Enterobacteria spp. due to its novel chemical structure in comparison to other aminoglycosides. It differs significantly from the structures of gentamycin and tobramycin but is similar to amikacin [44].
3.7. How genomics mitigates the public health impact of antimicrobial resistance
Whole-genome sequencing (WGS) and, more recently, metagenomic investigations have considerably improved our understanding of the antimicrobial resistance (AMR) process, and these technologies are guiding mitigation measures for better understanding and controlling of AMR (Table 1) [54].
Culture-based antimicrobial susceptibility testing (AST), which is still used in clinical microbiology and patient care, has historically been used to identify AMR. While phenotyping gives clear visual evidence of how bacteria will interact with an antibiotic, it typically provides little or no information about resistance mechanisms, with divergent genetic clones frequently exhibiting similar resistance profiles [55].
Multi-locus sequence typing (MLST), a genetic typing method, provides a better level of pathogen resolution than AST but is very limited since it only describes a tiny part of a genome. WGS, on the other hand, gives genome-wide information at the single nucleotide level that may be utilized to determine the existence and mechanisms of AMR, as well as pathogen identification, virulence, and origin [56,57]. With the advent of next-generation sequencing (NGS), which uses high-throughput, parallel sequencing of DNA fragments, pathogen genomes may now be identified quickly and at a cheap cost [58,59].
Comparative phylogenetic analysis can be used to determine the degree of relatedness between different isolates based on the extent of genome similarity and, when combined with epidemiological and clinical data, can help us understand the specific temporal patterns of AMR and transmission [55].
Furthermore, recent advancements in metagenomic sequencing methods have eliminated completely the need for establishing of bacterial cultures. As a result of combining all accessible genetic information in a sample, metagenomic analysis allows for a shift in focus from an individual pathogen to the community, microbiome landscape, resulting in a highly comprehensive model of how pathogens interact, mobilize and access AMR genes. [60]

Table 1.
Case studies on using whole-genome sequencing (WGS) to reduce the public health burden of antibiotic resistance (AMR).
Table 1.
Case studies on using whole-genome sequencing (WGS) to reduce the public health burden of antibiotic resistance (AMR).
| Case 1: International surveillance— determination the population structure and epidemiology of carbapenem-resistant K. pneumoniae (CR-Kp) across Europe [61] | |||
| Justification | WGS/workflow | Main findings | Advantages of WGS |
| The primary reservoirs and transmission dynamics of CR-Kp in Europe are still poorly understood. | For sequencing, hospital European laboratories have submitted consecutive clinical isolates of CR-Kp, along with a susceptible strain for comparison. | Carbapenemase acquisition was the primary cause of CR-Kp dissemination; nosocomial acquisition was the other main source of CR-Kp spread. | Provided a baseline for continuous CR-Kp monitoring. The importance of nosocomial spread was emphasized. |
| Case 2: Enhancing the national surveillance of antimicrobial resistance in the Philippines [55] | |||
| Justification | WGS/workflow | Main findings | Advantages of WGS |
| National laboratory-based surveillance has revealed an increase in AMR incidences over the preceding ten years, but the understanding of the epidemiology and causes of AMR have remained limited. | The capacity of WGS was added to the current surveillance project. To provide baseline data and guide control strategies, retrospective sequencing of MDR GNB collected before the introduction was performed and examined with phenotypic and epidemiological data. | Through the discovery of the introduction and country-wide dissemination of a high-risk epidemic clone, E. coli ST410, drivers of carbapenem resistance at several healthcare system levels were found, including a localized outbreak of plasmid-driven CR-Kp impacting a single healthcare facility. | The implementation of efficient infection control methods was made possible by a thorough understanding of the epidemiology and causes of AMR. Data was provided to worldwide AMR surveillance initiatives, which improved global coverage. |
| Case3: Investigating an MRSA outbreak in a neonatal unit in the UK [62] | |||
| Justification | WGS/workflow | Main findings | Advantages of WGS |
| Over a 6-month period, phenotypically comparable MRSA isolates were found in patients on a special baby care unit but could not be connected chronologically or geographically, implying that the entire breadth of the epidemic had not been recognized. | WGS was performed on all MRSA isolates received from a special baby unit patients over a 6-month period, independent of phenotypic features. MRSA isolates from the community with antibiograms comparable to the epidemic strain, as well as screening samples from elsewhere in the hospital, were also sequenced. | Phylogenetic research revealed that two previously excluded isolates were part of the epidemic, allowing temporal linkages between patients to be established. Beyond the newborn unit, a large transmission network was discovered. | WGS enabled the testing of a large number of isolates and the precise identification of related strains, allowing for comprehensive epidemic reconstruction. Combining WGS data with clinical and epidemiological data allowed for the identification of the source of the epidemic and the successful implementation of infection control measures. |
| Case 4: Investigating the direction of transmission in an A. baumannii outbreak in a UK hospital [63] | |||
| Justification | WGS/workflow | Main findings | Advantages of WGS |
| The molecular typing of a cluster of A. baumannii isolates acquired at a UK hospital suggested a clonal epidemic, but the route of transmission between cases could not be established based on the available laboratory, clinical, and epidemiological data. | WGS analysis was performed on a group of isolates acquired from patients with similar molecular typing profiles and antibiograms in order to better understand direct transmission between patients. | The index case was identified using phylogenetic analysis, and the subsequent chain of transmission was determined. One patient/isolate was found to be unconnected, and the outbreak investigation was abandoned. | The directionality of transmission may be identified by WGS, allowing for a precise reconstruction of the outbreak. |
| Case 5: Contact tracing and detection of secondary cases of TB in the Netherlands [64] | |||
| Justification | WGS/workflow | Main findings | Advantages of WGS |
| Secondary TB detection and screening are critical for TB control. The poor precision of molecular typing makes the accurate identification of case clusters and transmission networks difficult. | In 2016, clinical TB isolates in the Netherlands were examined using both molecular typing and WGS. The two techniques were evaluated in terms of discrimination and accuracy in identifying possibly related situations. | WGS proved more capable of determining the relatedness of isolates than molecular typing, grouping a lesser proportion of isolates as related (25% vs. 14%) and boosting the proportion verified as epidemiologically connected (57% vs. 31%). | WGS aided in the identification of transmission episodes, allowing for contact tracing and generating a broader knowledge of TB control. |
| Case 6: Identifying the drivers of AMR in atypical enteropathogenic E. coli (aEPEC) strains isolated from children < 5 years in four sub-Saharan African countries and three South Asian countries [65] | |||
| Justification | WGS/workflow | Main findings | Advantages of WGS |
| The incidence, causes, and drivers of AMR in E. coli intestinal isolates from children in the community in many places throughout the world were unclear. | The phenotypic susceptibility of isolates and WGS were investigated and linked with antibiotic usage, disease state (symptomatic/asymptomatic), phylogenetic lineage, and geographic location. | AMR was shown to be prevalent, with 65% of isolates resistant to at least three antimicrobial medication classes. A wide spectrum of genetic pathways of AMR was discovered, with geographic location and antimicrobial usage pattern being the best predictors of AMR. | WGS was utilized to conduct a thorough examination of AMR across a vast geographical area, revealing information about AMR epidemiology, distribution, and causes. |
| Case 7: Investigation of colistin resistance detected in commensal E. coli in food stock animals in China [66] | |||
| Justification | WGS/workflow | Main findings | Advantages of WGS |
| Routine surveillance had revealed a significant increase in the rates of colistin resistance in bacteria colonizing pigs in China, but the cause of this resistance remained unknown. | To validate the presence of plasmid-associated, transmissible colistin resistance, conjugation tests were performed. The WGS on plasmids was utilized to identify the relevant gene. | The plasmid-associated colistin resistance gene sequence was identified and named mcr-1. | The genetic foundation of a novel AMR mechanism has been identified and characterized, allowing for continuing surveillance and guiding inquiry and identification of this growing danger. |
| Case 8: Understanding of the epidemiology of MDR and XDR pathogens amenable to control by vaccination [67,68] | |||
| Justification | WGS/workflow | Main findings | Advantages of WGS |
| AMR is affecting the effectiveness of typhoid fever therapy. Resistance to azithromycin, the final effective oral drug, was discovered in Bangladesh and later in Pakistan, but the genetic basis and likelihood of spread remained unclear. | WGS was used to examine clinical isolates of azithromycin-resistant S. Typhi. The phylogenetic analysis allowed the strains to be contextualized among contemporaneous S. Typhi isolates in both contexts. | Phylogenetic analysis revealed that resistant isolates in Bangladesh and Pakistan arose from the separate acquisition of mutations in the same gene, showing the breadth of azithromycin selection pressure and the critical need for disease management by vaccination. | Two independent epidemics of azithromycin-resistant S. Typhi were identified and investigated using WGS. These findings aided in the development of innovative typhoid conjugate vaccines for infection control. |
3.8. Potential effect of nutrigenomics on increased antimicrobial resistance against new antibiotics
According to metagenomic research, commensal bacteria in healthy people help keeping pathogenic bacteria at a low density, implying that carriage is not often of concern. When patients have invasive surgery, however, there is a loss of microbial diversity, which is followed by the colonization of harmful bacteria. The use of antimicrobials, which commonly leads to the selection of drug-resistant pathogens and enables horizontal gene transfer (HGT) of AMR genes between bacterial lineages and species, might increase this impact. WGS investigations are currently being utilized to uncover the colonization variables that allow specific infections to grow quickly and survive in such conditions, with the hope that treatments targeting persistent organisms will be developed to reduce pathogen colonization [69,70,71,72,73,74].
ARGs from heat-treated bacteria might be transmitted to other bacteria via a variety of horizontal gene transfer (HGT) pathways (Table 2). HGT is aided by mobile genetic elements (MGEs) like plasmids, integrons, and transposons, which allow genes to travel more freely. The frequency of HGT is heavily influenced by the qualities of MGEs, the characteristics of the donor and recipient populations, and the environment. Three primary conventional HGT pathways have been indicated (1) conjugation, (2) transformation, or (3) transduction. Other, less well-known processes of DNA transfer may occur. [75]
Conjugation takes place between living bacterial cells and is not possible if the cells are heat inactivated [76]. DNA fragments, including ARGs, may be released from lysed heat inactivated bacteria and transmitted through transformation. Natural transformation has been seen in over 60 bacterial species and is likely to occur in many more [77,78].
Few studies have specifically examined exogenous DNA absorption by bacteria in food. There is evidence that DNA stability is inversely proportional to DNA length and that while heat treatments destroy lysed exposed ARGs, fragments may still be long enough to be transformed by other bacteria [79].
Transduction is a kind of HGT that is mediated by bacteriophages (phages) and related particles known as gene transfer agents (GTAs). Phages can package some genetic material (including ARGs) of their hosts by replicating within the host cell before lysing it (lytic) or by integration into the host cell genome (lysogenic) [76].
Table 2.
Studies on the fate of antibiotic resistance genes (ARGs) after exposure to heat.
| Procedure | Medium | Evaluation temperature (°C) |
Species | Antimicrobial Resistance Genes (ARGs) present | Stated antimicrobial resistance profiles | Recipient species | ARGs detected post-treatment from non-culturable samples |
Transformation demonstrated | Reference | |
|---|---|---|---|---|---|---|---|---|---|---|
| Cooking—boiled (20 min), grilled (10 min), microwaved (5 min, 900 W), or autoclaved (20 min, 121 °C) | Chicken, beef, pork ?? |
Not Stated | E. faecalis | aac(6′)-Ie-aph(2″)-Ia | Aminoglycosides, except to streptomycin(predicted profile, not tested) | E. faecalis | YES | NO | [80] | |
| General heat treatments | Saline | 40, 50,60, 70, 80, 90, 100 | E. coli |
blaCTX-M-1, blaCMY-2, tetA, strA |
Cephalosporins, tetracycline, streptomycin | E. coli | YES | YES | 70 °C for 30 min | [81] |
| Milk pasteurizatio-n (sterilization) | Milk and elution buffer | 63.5, 121 |
S. aureus, S. sciuri |
blaZ, mecC, tetK | Penicillin, methicillin, tetracycline | S. aureus | YES | YES | 63.5 °C for 30 min | [82] |
| Non-food autoclaving | Distilled water and in presence of salt | 121, 135 | Plasmid (pUC18) | NS | Ampicillin | E. coli | - | YES | 121 °C for 15 min | [83] |
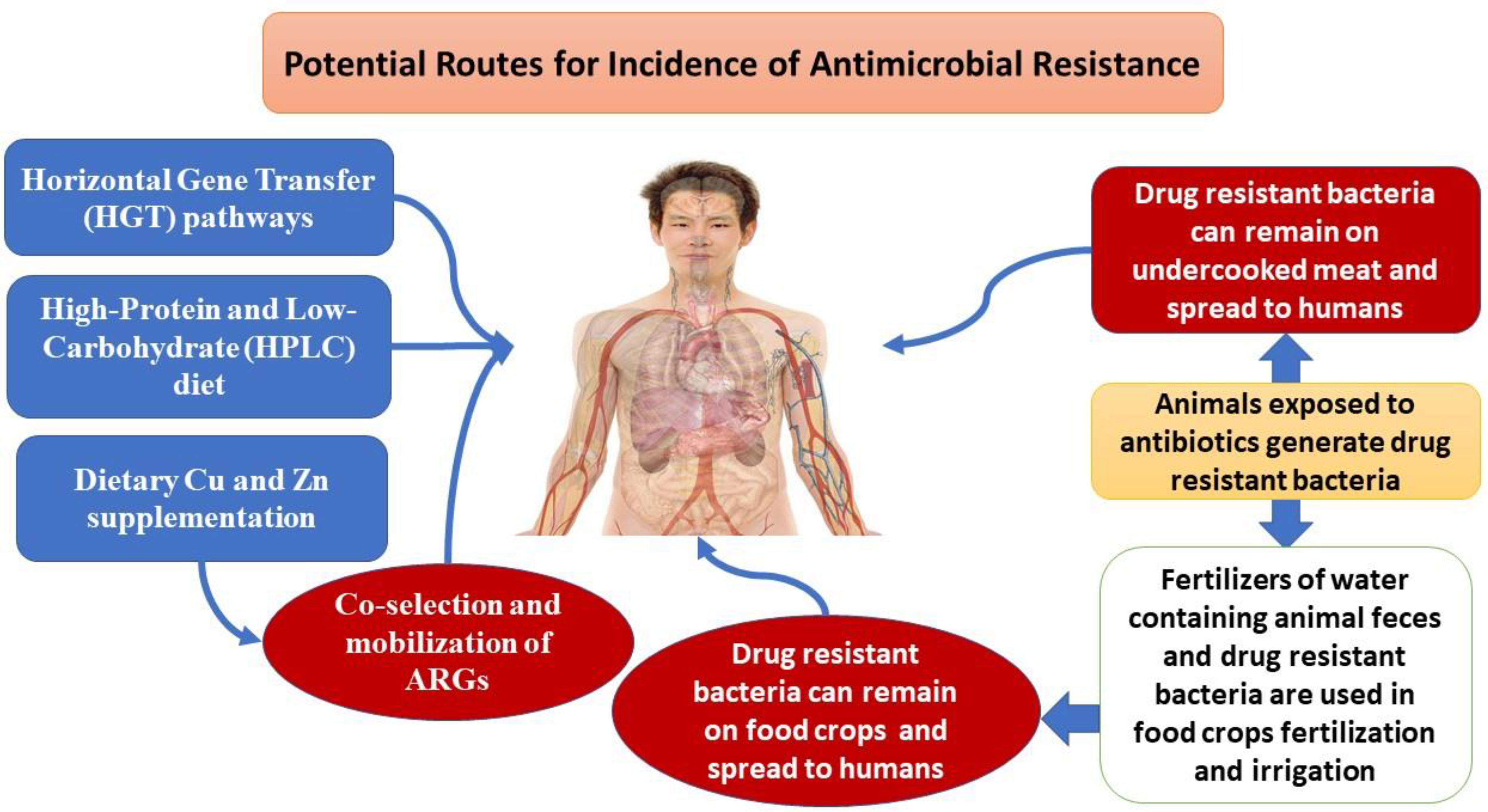
Individual patient risk factors for getting infected by extended-spectrum β-lactamase-producing bacteria include extended hospital stay prior to infection, antibiotic exposure, and recent travel abroad (Figure 3) [68]. Male sex, older age, and co-morbidities are all risk factors for multi-drug resistant (MDR) Gram-negative bacteria infections [69]. Changes in the human microbiota occur in response to disease, particularly when antibiotic exposure is frequent and/or chronic. Enterobacterales are common gastrointestinal colonizers that can serve as key reservoirs for mobile AMR genes [60].
Enterococci are innately resistant to cephalosporins, allowing them to develop to unusually high densities in patients subjected to cephalosporin treatment, encouraging diffusion to other areas where infection occurs. The cognate response sensor (CroS) kinase and its cognate response regulator (CroR) are essential for cephalosporin resistance in Enterococcus faecalis, but little is known about the variables that govern this signaling system to modify resistance. To detect protein-protein interactions in E. faecalis cells, a protein fragment complementation assay was used, which revealed a previously unknown association of CroR with the HPr protein of the phosphotransferase system (PTS), which is responsible for carbohydrate uptake and catabolite control of gene expression. The potential of CroS to increase cephalosporin resistance and gene expression in a nutrient-dependent manner is limited by its connection with HPr, according to genetic and physiological investigations. Mutational studies revealed that the interface via which HPr associates with CroR differs from the interface via which it associates with other cellular partners. [84]
A study in United States showed that aminoglycosides were discovered to be the most common and widespread cause of AMR in healthy people, and aminoglycoside-O-phosphotransferases (aph3-dprime) associated negatively with total calories and consumption of soluble fibers. Data revealed that individuals with low ARGs ingested much more fiber in their diets than medium and high ARG individuals, which was possessed greater abundances of obligate anaerobes in their gut microbiota, particularly from the family Clostridiaceae. Moreover, machine learning was used to look for connections between 387 nutritional, physiological, and lifestyle characteristics and antimicrobial resistance, and discovered that enhanced phylogenetic diversity of food was related with individuals with low ARG levels [85].
Data from animal studies showed that in both canines and felines 23 antibiotic resistant genes (ARGs) were found in 50% of the samples, with tetracycline and aminoglycoside resistance genes being the most common. The abundance of a particular ARG tended to respond similarly to nutritional intervention. When compared to dogs subjected to a baseline diet, dogs on the High-Protein and Low-Carbohydrate (HPLC) diet had a higher abundance of tetracycline resistance genes tet(W), tet(O), tet (44), and macrolide resistance genes mefA and mel, but a lower abundance of the β-lactam ARG CfxA6. The quantity of these ARGs was similar in HPLC-fed kittens and Moderate-Protein/Moderate-Carbohydrate (MPMC)-fed kittens [86].
The tetracycline resistance gene tet(W) was found in the greatest number of taxa, mostly in Firmicutes. Bifidobacterium, a genus widely utilized in dairy product fermentation and as probiotics, shared tet(W) with a wide range of other species. [86]
Dietary Cu and Zn supplementation in swine production may further increase the likelihood of antibiotic resistance spreading through co-selection and mobilization of ARGs and subsequent transmission to humans. [87,88]
Co-selection can occur when ARGs and metal resistance genes (MRGs) are genetically linked (co-resistance), when the same resistance mechanism confers resistance to both metals and antibiotics (cross-resistance), or when a common regulator controls the expression of both metal and antibiotic resistance systems [89]. Indeed, increased dietary Cu and Zn dosages used to promote swine development have been demonstrated to select for Cu or Zn resistance and to co-select for antibiotic resistance in certain groups of swine gut bacteria containing harmful strains [90].
The previously mentioned new antibiotics in section 3.6 showed no emerging resistance against their antibacterial activity that make them more adventitious over other existing antibiotics in different clinical settings. However, the aforementioned evidence regarding nutrigenomic effects on antimicrobial resistance suggest potential emergence of resistance one day against these new antibiotics.
4. Conclusions
It is apparent that significant effort is being dedicated to the development of antibiotic alternatives to promote better clinical outcomes with a low resistance incidence. As we move forward, we must keep resistance mechanisms in mind so that they can be perpetuated. Nutrigenomic pathways seem to carry a lot of unknowns that may result in emergence of antibiotic resistance. Till the moment, new antibiotics seem to overcome the occurrence of resistance, but one-day resistance against those agents may occur.
5. Recommendations
Extending the application of different nutrigenomic approaches in healthcare settings
6. Limitations
The possible chances of integration of nutrigenomics in clinical practice
Author Contributions
Conceptualization, N.A.S.; writing—original draft preparation, E.M.S, M.A.E and S.A.R; writing—review and editing, M.A.E and A.S.M; supervision, N.A.S and A.S.M; project administration, N.A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Drug Research Center (DRC), Cairo-Egypt for the great effort and continuous support during development of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, Y.; Michalak, M.; Agellon, L.B. Importance of nutrients and nutrient metabolism on human health. Yale J. Biol. Med. 2018, 91, 95–103. [Google Scholar]
- Carlberg, C. Nutrigenomics in the context of evolution. Redox Biol. 2023, 62, 102656. [Google Scholar] [CrossRef]
- Ordovas, J.M.; Ferguson, L.R.; Tai, E.S.; Mathers, J.C. Personalised nutrition and health. BMJ 2018, 361, bmj.k2173. [Google Scholar] [CrossRef]
- Mishra, U.N.; Jena, D.; Sahu, C.; Devi, R.; Kumar, R.; Jena, R.; Irondi, E.A.; Rout, S.; Tiwari, R.K.; Lal, M.K.; et al. Nutrigenomics: An Inimitable Interaction amid Genomics, Nutrition and Health. Innovative Food Science & Emerging Technologies 2022, 82, 103196. [Google Scholar]
- Neeha, V.S.; Kinth, P. Nutrigenomics Research: A Review. Journal of Food Science and Technology 2012, 50, 415–428. [Google Scholar] [CrossRef]
- Brennan, L.; de Roos, B. Nutrigenomics: Lessons Learned and Future Perspectives. The American Journal of Clinical Nutrition 2021, 113, 503–516. [Google Scholar] [CrossRef]
- Naushad, S.M.; Rama Devi, A.R. Role of Parental Folate Pathway Single Nucleotide Polymorphisms in Altering the Susceptibility to Neural Tube Defects in South India. Journal of Perinatal Medicine 2010, 38. [Google Scholar] [CrossRef]
- Vara Prasad, S.S.; Jeya Kumar, S.S.; Kumar, P.U.; Qadri, S.S.; Vajreswari, A. Dietary Fatty Acid Composition Alters 11β-Hydroxysteroid Dehydrogenase Type 1 Gene Expression in Rat Retroperitoneal White Adipose Tissue. Lipids in Health and Disease 2010, 9. [Google Scholar] [CrossRef]
- Valiakos, G.; Kapna, I. Colistin Resistant Mcr Genes Prevalence in Livestock Animals (Swine, Bovine, Poultry) from a Multinational Perspective. A Systematic Review. Veterinary Sciences 2021, 8, 265. [Google Scholar] [CrossRef]
- Di Francesco, A.; Salvatore, D.; Sakhria, S.; Bertelloni, F.; Catelli, E.; Ben Yahia, S.; Tlatli, A. Colistin Resistance Genes in Broiler Chickens in Tunisia. Animals 2023, 13, 1409. [Google Scholar] [CrossRef]
- Gilani, S.M.H.; Rashid, Z.; Galani, S.; Ilyas, S.; Sahar, S.; Zahoor-ul-Hassan; Al-Ghanim, K.; Zehra, S.; Azhar, A.; Al-Misned, F.; et al. Growth Performance, Intestinal Histomorphology, Gut Microflora and Ghrelin Gene Expression Analysis of Broiler by Supplementing Natural Growth Promoters: A Nutrigenomics Approach. Saudi Journal of Biological Sciences 2021, 28, 3438–3447. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, P.; Wu, Y.; Wang, X.; Lu, X.; Liu, C.; Li, N.; Sun, J.; Xiao, J.; Jesus, S.-G. The Formation of Antibiotic Resistance Genes in Bacterial Communities During Garlic Powder Processing. Frontiers in Nutrition 2021, 8. [Google Scholar] [CrossRef]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Davenport, E.R.; Beaumont, M.; Jackson, M.A.; Knight, R.; Ober, C.; Spector, T.D.; Bell, J.T.; Clark, A.G.; Ley, R.E. Genetic Determinants of the Gut Microbiome in UK Twins. Cell Host & Microbe 2016, 19, 731–743. [Google Scholar]
- Martín-Peláez, S.; Fito, M.; Castaner, O. Mediterranean Diet Effects on Type 2 Diabetes Prevention, Disease Progression, and Related Mechanisms. A Review. Nutrients 2020, 12, 2236. [Google Scholar] [CrossRef]
- Soldati, L.; Di Renzo, L.; Jirillo, E.; Ascierto, P.A.; Marincola, F.M.; De Lorenzo, A. The Influence of Diet on Anti-Cancer Immune Responsiveness. Journal of Translational Medicine 2018, 16. [Google Scholar] [CrossRef]
- Mansour, S.R.; Moustafa, M.A.A.; Saad, B.M.; Hamed, R.; Moustafa, A.-R.A. Impact of Diet on Human Gut Microbiome and Disease Risk. New Microbes and New Infections 2021, 41, 100845. [Google Scholar] [CrossRef]
- Hildebrandt, M.A.; Hoffmann, C.; Sherrill–Mix, S.A.; Keilbaugh, S.A.; Hamady, M.; Chen, Y.; Knight, R.; Ahima, R.S.; Bushman, F.; Wu, G.D. High-Fat Diet Determines the Composition of the Murine Gut Microbiome Independently of Obesity. Gastroenterology 2009, 137, 1716–1724.e2. [Google Scholar] [CrossRef]
- Agans, R.; Gordon, A.; Kramer, D.L.; Perez-Burillo, S.; Rufián-Henares, J.A.; Paliy, O. Dietary Fatty Acids Sustain the Growth of the Human Gut Microbiota. Applied and Environmental Microbiology 2018, 84. [Google Scholar] [CrossRef]
- Doaei, S.; Gholamalizadeh, M.; Akbari, M.E.; Akbari, S.; Feradova, H.; Rahimzadeh, G.; Mosavi Jarrahi, A. Dietary Carbohydrate Promotes Cell Survival in Cancer Via the Up-Regulation of Fat Mass and Obesity-Associated Gene Expression Level. Malaysian Journal of Medical Sciences 2019, 26, 8–17. [Google Scholar] [CrossRef]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. Journal of Lipid Research 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut Microbiome Development along the Colorectal Adenoma–Carcinoma Sequence. Nature Communications 2015, 6. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Boba, A.; Czemplik, M.; Kulma, A.; Wojtasik, W. Influence of the Bioactive Diet Components on the Gene Expression Regulation. Nutrients 2021, 13, 3673. [Google Scholar] [CrossRef]
- Paoloni-Giacobino, A.; Grimble, R.; Pichard, C. Genetics and Nutrition. Clinical Nutrition 2003, 22, 429–435. [Google Scholar] [CrossRef]
- Leonardson, A.S.; Zhu, J.; Chen, Y.; Wang, K.; Lamb, J.R.; Reitman, M.; Emilsson, V.; Schadt, E.E. The Effect of Food Intake on Gene Expression in Human Peripheral Blood. Human Molecular Genetics 2009, 19, 159–169. [Google Scholar] [CrossRef]
- Wolfrum, C.; Asilmaz, E.; Luca, E.; Friedman, J.M.; Stoffel, M. Foxa2 Regulates Lipid Metabolism and Ketogenesis in the Liver during Fasting and in Diabetes. Nature 2004, 432, 1027–1032. [Google Scholar] [CrossRef]
- Kobayashi, H.; Oishi, K.; Hanai, S.; Ishida, N. Effect of Feeding on Peripheral Circadian Rhythms and Behaviour in Mammals. Genes to Cells 2004, 9, 857–864. [Google Scholar] [CrossRef]
- Bordoni, L.; Gabbianelli, R. Primers on Nutrigenetics and Nutri(Epi)Genomics: Origins and Development of Precision Nutrition. Biochimie 2019, 160, 156–171. [Google Scholar] [CrossRef]
- Abushaheen, M.A.; Muzaheed; Fatani, A.J.; Alosaimi, M.; Mansy, W.; George, M.; Acharya, S.; Rathod, S.; Divakar, D.D.; Jhugroo, C.; et al. Antimicrobial Resistance, Mechanisms and Its Clinical Significance. Disease-a-Month 2020, 66, 100971. [Google Scholar] [CrossRef]
- C Reygaert, W. An Overview of the Antimicrobial Resistance Mechanisms of Bacteria. AIMS Microbiology 2018, 4, 482–501. [Google Scholar] [CrossRef] [PubMed]
- Patangia, D.V.; Anthony Ryan, C.; Dempsey, E.; Paul Ross, R.; Stanton, C. Impact of Antibiotics on the Human Microbiome and Consequences for Host Health. MicrobiologyOpen 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.; Stanton, C.; Lane, J.; Smith, G.; Ross, R. Precision Nutrition and the Microbiome, Part I: Current State of the Science. Nutrients 2019, 11, 923. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, D.-C. Facing a New Challenge. Chinese Medical Journal 2019, 132, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, A.J.; Wang, B.; Sun, X.; Kennedy, E.A.; Hernandez-Leyva, A.; Ndao, I.M.; Tarr, P.I.; Warner, B.B.; Dantas, G. Persistent Metagenomic Signatures of Early-Life Hospitalization and Antibiotic Treatment in the Infant Gut Microbiota and Resistome. Nature Microbiology 2019, 4, 2285–2297. [Google Scholar] [CrossRef]
- Gibson, M.K.; Wang, B.; Ahmadi, S.; Burnham, C.-A.D.; Tarr, P.I.; Warner, B.B.; Dantas, G. Developmental Dynamics of the Preterm Infant Gut Microbiota and Antibiotic Resistome. Nature Microbiology 2016, 1. [Google Scholar] [CrossRef]
- McDonald, L.C. Effects of Short- and Long-Course Antibiotics on the Lower Intestinal Microbiome as They Relate to Traveller’s Diarrhea. Journal of Travel Medicine 2017, 24, S35–S38. [Google Scholar] [CrossRef]
- Pérez-Cobas, A.E.; Artacho, A.; Knecht, H.; Ferrús, M.L.; Friedrichs, A.; Ott, S.J.; Moya, A.; Latorre, A.; Gosalbes, M.J. Differential Effects of Antibiotic Therapy on the Structure and Function of Human Gut Microbiota. PLoS ONE 2013, 8, e80201. [Google Scholar] [CrossRef]
- Muhammad, M.H.; Idris, A.L.; Fan, X.; Guo, Y.; Yu, Y.; Jin, X.; Qiu, J.; Guan, X.; Huang, T. Beyond Risk: Bacterial Biofilms and Their Regulating Approaches. Frontiers in Microbiology 2020, 11. [Google Scholar] [CrossRef]
- Mu, C.; Yang, Y.; Yu, K.; Yu, M.; Zhang, C.; Su, Y.; Zhu, W. Alteration of Metabolomic Markers of Amino-Acid Metabolism in Piglets with in-Feed Antibiotics. Amino Acids 2017, 49, 771–781. [Google Scholar] [CrossRef]
- Yang, H.; Wei, S.-H.; Hobman, J.L.; Dodd, C.E.R. Antibiotic and Metal Resistance in Escherichia Coli Isolated from Pig Slaughterhouses in the United Kingdom. Antibiotics 2020, 9, 746. [Google Scholar] [CrossRef] [PubMed]
- Sacher-Pirklbauer, A.; Klein-Jöbstl, D.; Sofka, D.; Blanc-Potard, A.-B.; Hilbert, F. Phylogenetic Groups and Antimicrobial Resistance Genes in Escherichia Coli from Different Meat Species. Antibiotics 2021, 10, 1543. [Google Scholar] [CrossRef] [PubMed]
- Tohmaz, M.; Askari Badouei, M.; Kalateh Rahmani, H.; Hashemi Tabar, G. Antimicrobial Resistance, Virulence Associated Genes and Phylogenetic Background versus Plasmid Replicon Types: The Possible Associations in Avian Pathogenic Escherichia Coli (APEC). BMC Veterinary Research 2022, 18. [Google Scholar] [CrossRef] [PubMed]
- Terreni, M.; Taccani, M.; Pregnolato, M. New Antibiotics for Multidrug-Resistant Bacterial Strains: Latest Research Developments and Future Perspectives. Molecules 2021, 26, 2671. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Antibacterial Agents in Clinical Development: An Analysis of the Antibacterial Clinical Development Pipeline. [(accessed on 1 May 2021)];2019 Available online: http://apps.who.int/iris/bitstream/handle/10665/258965/WHO-EMP-IAU-2017.11-eng.pdf;jsessionid=9955EDC07F0D8D09ACAA2CAC34DB921F?sequence=1. (accessed on 1 May 2021).
- Bradford, P.A.; Miller, A.A.; O’Donnell, J.; Mueller, J.P. Zoliflodacin: An Oral Spiropyrimidinetrione Antibiotic for the Treatment of Neisseria Gonorrheae, Including Multi-Drug-Resistant Isolates. ACS Infectious Diseases 2020, 6, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Kowalska-Krochmal, B.; Dudek-Wicher, R. The Minimum Inhibitory Concentration of Antibiotics: Methods, Interpretation, Clinical Relevance. Pathogens 2021, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Cho, J. Ridinilazole: A Novel Antimicrobial for Clostridium Difficile Infection. Annals of Gastroenterology 2018. [Google Scholar] [CrossRef] [PubMed]
- Tulkens, P.M.; Van Bambeke, F.; Zinner, S.H. Profile of a Novel Anionic Fluoroquinolone—Delafloxacin. Clinical Infectious Diseases 2019, 68, S213–S222. [Google Scholar] [CrossRef]
- Sartelli, M.; Chichom-Mefire, A.; Labricciosa, F.M.; Hardcastle, T.; Abu-Zidan, F.M.; Adesunkanmi, A.K.; Ansaloni, L.; Bala, M.; Balogh, Z.J.; Beltrán, M.A.; et al. The Management of Intra-Abdominal Infections from a Global Perspective: 2017 WSES Guidelines for Management of Intra-Abdominal Infections. World Journal of Emergency Surgery 2017, 12. [Google Scholar]
- Nguyen, F.; Starosta, A.L.; Arenz, S.; Sohmen, D.; Dönhöfer, A.; Wilson, D.N. Tetracycline Antibiotics and Resistance Mechanisms. Biological Chemistry 2014, 395, 559–575. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Cheung, D.; Adam, H.; Zelenitsky, S.; Golden, A.; Schweizer, F.; Gorityala, B.; Lagacé-Wiens, P.R.S.; Walkty, A.; Gin, A.S.; et al. Review of Eravacycline, a Novel Fluorocycline Antibacterial Agent. Drugs 2016, 76, 567–588. [Google Scholar] [CrossRef] [PubMed]
- Endimiani, A.; Hujer, K.M.; Hujer, A.M.; Armstrong, E.S.; Choudhary, Y.; Aggen, J.B.; Bonomo, R.A. ACHN-490, a Neoglycoside with Potent In Vitro Activity against Multidrug-Resistant Klebsiella Pneumoniae Isolates. Antimicrobial Agents and Chemotherapy 2009, 53, 4504–4507. [Google Scholar] [CrossRef] [PubMed]
- Waddington, C.; Carey, M.E.; Boinett, C.J.; Higginson, E.; Veeraraghavan, B.; Baker, S. Exploiting Genomics to Mitigate the Public Health Impact of Antimicrobial Resistance. Genome Medicine 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Argimón, S.; Masim, M.A.L.; Gayeta, J.M.; Lagrada, M.L.; Macaranas, P.K.V.; Cohen, V.; Limas, M.T.; Espiritu, H.O.; Palarca, J.C.; Chilam, J.; et al. Integrating Whole-Genome Sequencing within the National Antimicrobial Resistance Surveillance Program in the Philippines. Nature Communications 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Whole-Genome Sequencing as Part of National and International Surveillance Programmes for Antimicrobial Resistance: A Roadmap. BMJ Global Health 2020, 5, e002244. [CrossRef] [PubMed]
- Didelot, X.; Gardy, J.; Colijn, C. Bayesian Inference of Infectious Disease Transmission from Whole-Genome Sequence Data. Molecular Biology and Evolution 2014, 31, 1869–1879. [Google Scholar] [CrossRef] [PubMed]
- Ashton, P.M.; Nair, S.; Peters, T.M.; Bale, J.A.; Powell, D.G.; Painset, A.; Tewolde, R.; Schaefer, U.; Jenkins, C.; Dallman, T.J.; et al. Identification of Salmonellafor Public Health Surveillance Using Whole Genome Sequencing. PeerJ 2016, 4, e1752. [Google Scholar] [CrossRef]
- Wainwright, C.; Altamirano, L.; Cheney, M.; Cheney, J.; Barber, S.; Price, D.; Moloney, S.; Kimberley, A.; Woolfield, N.; Cadzow, S.; et al. A Multicenter, Randomized, Double-Blind, Controlled Trial of Nebulized Epinephrine in Infants with Acute Bronchiolitis. New England Journal of Medicine 2003, 349, 27–35. [Google Scholar] [CrossRef]
- Blake, K.S.; Choi, J.; Dantas, G. Approaches for Characterizing and Tracking Hospital-Associated Multidrug-Resistant Bacteria. Cellular and Molecular Life Sciences 2021, 78, 2585–2606. [Google Scholar] [CrossRef]
- David, S.; Reuter, S.; Harris, S.R.; Glasner, C.; Feltwell, T.; Argimon, S.; Abudahab, K.; Goater, R.; Giani, T.; Errico, G.; et al. Epidemic of Carbapenem-Resistant Klebsiella Pneumoniae in Europe Is Driven by Nosocomial Spread. Nature Microbiology 2019, 4, 1919–1929. [Google Scholar] [CrossRef]
- Harris, S.R.; Cartwright, E.J.; Török, M.E.; Holden, M.T.; Brown, N.M.; Ogilvy-Stuart, A.L.; Ellington, M.J.; Quail, M.A.; Bentley, S.D.; Parkhill, J.; et al. Whole-Genome Sequencing for Analysis of an Outbreak of Meticillin-Resistant Staphylococcus Aureus: A Descriptive Study. The Lancet Infectious Diseases 2013, 13, 130–136. [Google Scholar] [CrossRef]
- Lewis, T.; Loman, N.J.; Bingle, L.; Jumaa, P.; Weinstock, G.M.; Mortiboy, D.; Pallen, M.J. High-Throughput Whole-Genome Sequencing to Dissect the Epidemiology of Acinetobacter Baumannii Isolates from a Hospital Outbreak. Journal of Hospital Infection 2010, 75, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Jajou, R.; Neeling, A. de; Hunen, R. van; Vries, G. de; Schimmel, H.; Mulder, A.; Anthony, R.; Hoek, W. van der; Soolingen, D. van Epidemiological Links between Tuberculosis Cases Identified Twice as Efficiently by Whole Genome Sequencing than Conventional Molecular Typing: A Population-Based Study. PLoS ONE 2018, 13, e0195413. [Google Scholar]
- Ingle, D.J.; Levine, M.M.; Kotloff, K.L.; Holt, K.E.; Robins-Browne, R.M. Dynamics of Antimicrobial Resistance in Intestinal Escherichia Coli from Children in Community Settings in South Asia and Sub-Saharan Africa. Nature Microbiology 2018, 3, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. The Lancet Infectious Diseases 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Hooda, Y.; Sajib, M.S.I.; Rahman, H.; Luby, S.P.; Bondy-Denomy, J.; Santosham, M.; Andrews, J.R.; Saha, S.K.; Saha, S. Molecular Mechanism of Azithromycin Resistance among Typhoidal Salmonella Strains in Bangladesh Identified through Passive Pediatric Surveillance. PLOS Neglected Tropical Diseases 2019, 13, e0007868. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Dehraj, I.F.; Carey, M.E.; Dyson, Z.A.; Garrett, D.; Seidman, J.C.; Kabir, F.; Saha, S.; Baker, S.; Qamar, F.N. A Race against Time: Reduced Azithromycin Susceptibility in Salmonella Enterica Serovar Typhi in Pakistan. mSphere 2020, 5. [Google Scholar] [CrossRef]
- Jung, H.-J.; Littmann, E.R.; Seok, R.; Leiner, I.M.; Taur, Y.; Peled, J.; van den Brink, M.; Ling, L.; Chen, L.; Kreiswirth, B.N.; et al. Genome-Wide Screening for Enteric Colonization Factors in Carbapenem-Resistant ST258 Klebsiella Pneumoniae. mBio 2019, 10. [Google Scholar] [CrossRef]
- Taur, Y.; Xavier, J.B.; Lipuma, L.; Ubeda, C.; Goldberg, J.; Gobourne, A.; Lee, Y.J.; Dubin, K.A.; Socci, N.D.; Viale, A.; et al. Intestinal Domination and the Risk of Bacteremia in Patients Undergoing Allogeneic Hematopoietic Stem Cell Transplantation. Clinical Infectious Diseases 2012, 55, 905–914. [Google Scholar] [CrossRef]
- Bell, B.G.; Schellevis, F.; Stobberingh, E.; Goossens, H.; Pringle, M. A Systematic Review and Meta-Analysis of the Effects of Antibiotic Consumption on Antibiotic Resistance. BMC Infectious Diseases 2014, 14. [Google Scholar] [CrossRef]
- van Schaik, W. The Human Gut Resistome. Philosophical Transactions of the Royal Society B: Biological Sciences 2015, 370, 20140087. [Google Scholar] [CrossRef] [PubMed]
- Osthoff, M.; McGuinness, S.L.; Wagen, A.Z.; Eisen, D.P. Urinary Tract Infections Due to Extended-Spectrum Beta-Lactamase-Producing Gram-Negative Bacteria: Identification of Risk Factors and Outcome Predictors in an Australian Tertiary Referral Hospital. International Journal of Infectious Diseases 2015, 34, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Leal, H.F.; Azevedo, J.; Silva, G.E.O.; Amorim, A.M.L.; de Roma, L.R.C.; Arraes, A.C.P.; Gouveia, E.L.; Reis, M.G.; Mendes, A.V.; de Oliveira Silva, M.; et al. Bloodstream Infections Caused by Multidrug-Resistant Gram-Negative Bacteria: Epidemiological, Clinical and Microbiological Features. BMC Infectious Diseases 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- James, C.; Dixon, R.; Talbot, L.; James, S.J.; Williams, N.; Onarinde, B.A. Assessing the Impact of Heat Treatment of Food on Antimicrobial Resistance Genes and Their Potential Uptake by Other Bacteria—A Critical Review. Antibiotics 2021, 10, 1440. [Google Scholar] [CrossRef] [PubMed]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.-A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial Resistance in the Food Chain: A Review. International Journal of Environmental Research and Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rodríguez, F.; Mercanoglu Taban, B. A State-of-Art Review on Multi-Drug Resistant Pathogens in Foods of Animal Origin: Risk Factors and Mitigation Strategies. Frontiers in Microbiology 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Rizzotti, L.; Felis, G.E.; Torriani, S. Horizontal Gene Transfer among Microorganisms in Food: Current Knowledge and Future Perspectives. Food Microbiology 2014, 42, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Kharazmi, M.; Bauer, T.; Hammes, W.P.; Hertel, C. Effect of Food Processing on the Fate of DNA with Regard to Degradation and Transformation Capability in Bacillus Subtilis. Systematic and Applied Microbiology 2003, 26, 495–501. [Google Scholar] [CrossRef]
- Koncan, R.; García-Albiach, R.; Bravo, D.; Cantón, R.; Baquero, F.; Cornaglia, G.; del Campo, R. P571 The Fate of Antibiotic Resistance Genes in Cooked Meat. International Journal of Antimicrobial Agents 2007, 29, S130. [Google Scholar] [CrossRef]
- Le Devendec, L.; Jouy, E.; Kempf, I. Evaluation of Resistance Gene Transfer from Heat-Treated Escherichia Coli. International Journal of Food Microbiology 2018, 270, 39–43. [Google Scholar] [CrossRef]
- Taher, E.M.; Hemmatzadeh, F.; Aly, S.A.; Elesswy, H.A.; Petrovski, K.R. Survival of Staphylococci and Transmissibility of Their Antimicrobial Resistance Genes in Milk after Heat Treatments. LWT 2020, 129, 109584. [Google Scholar] [CrossRef]
- Masters, C.I.; Miles, C.A.; Mackey, B.M. Survival and Biological Activity of Heat Damaged DNA. Letters in Applied Microbiology 1998, 27, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Snyder, H.; Kellogg, S.L.; Skarda, L.M.; Little, J.L.; Kristich, C.J. Nutritional Control of Antibiotic Resistance via an Interface between the Phosphotransferase System and a Two-Component Signaling System. Antimicrobial Agents and Chemotherapy 2014, 58, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Xue, Z.; Villanueva, Y.T.; Durbin-Johnson, B.; Alkan, Z.; Taft, D.H.; Liu, J.; Korf, I.; Laugero, K.D.; Stephensen, C.B.; et al. Association of Diet and Antimicrobial Resistance in Healthy U.S. Adults. mBio 2022, 13. [Google Scholar] [CrossRef]
- Kim, Y.; Leung, M.H.Y.; Kwok, W.; Fournié, G.; Li, J.; Lee, P.K.H.; Pfeiffer, D.U. Antibiotic Resistance Gene Sharing Networks and the Effect of Dietary Nutritional Content on the Canine and Feline Gut Resistome. Animal Microbiome 2020, 2. [Google Scholar] [CrossRef] [PubMed]
- Muurinen, J.; Richert, J.; Wickware, C.L.; Richert, B.; Johnson, T.A. Swine Growth Promotion with Antibiotics or Alternatives Can Increase Antibiotic Resistance Gene Mobility Potential. Scientific Reports 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Su, J.-Q.; An, X.-L.; Huang, F.-Y.; Rensing, C.; Brandt, K.K.; Zhu, Y.-G. Feed Additives Shift Gut Microbiota and Enrich Antibiotic Resistance in Swine Gut. Science of The Total Environment 2018, 621, 1224–1232. [Google Scholar] [CrossRef]
- Poole, K. At the Nexus of Antibiotics and Metals: The Impact of Cu and Zn on Antibiotic Activity and Resistance. Trends in Microbiology 2017, 25, 820–832. [Google Scholar] [CrossRef]
- Yazdankhah, S.; Rudi, K.; Bernhoft, A. Zinc and Copper in Animal Feed – Development of Resistance and Co-Resistance to Antimicrobial Agents in Bacteria of Animal Origin. Microbial Ecology in Health & Disease 2014, 25. [Google Scholar]
Figure 2.
General antimicrobial resistance mechanisms [31]. Reproduced from Kosmidis 2015.
Figure 2.
General antimicrobial resistance mechanisms [31]. Reproduced from Kosmidis 2015.
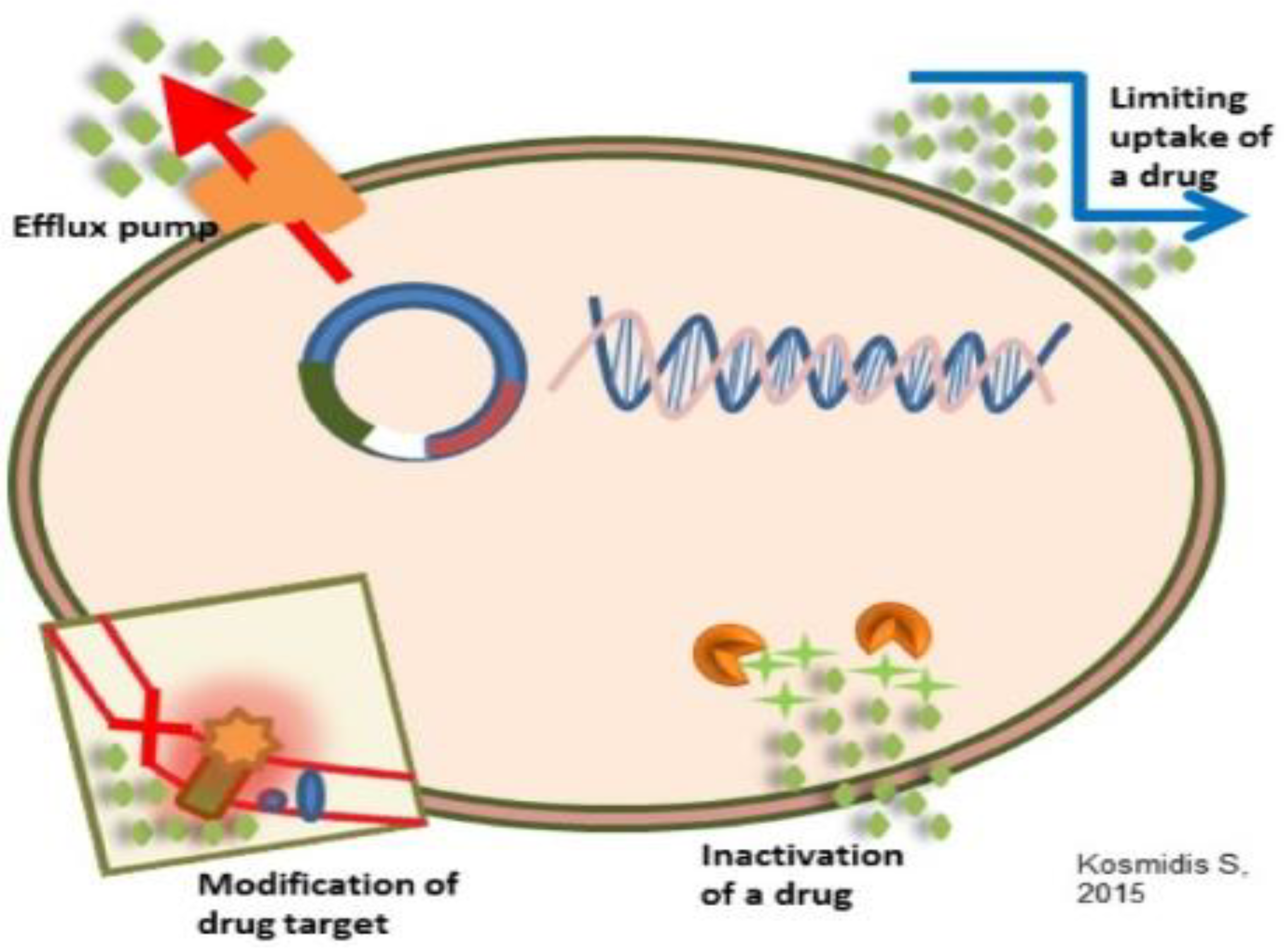
Figure 3.
Potential Routes for Antimicrobial Resistance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



